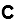zhao, B., li, T., li, y., Admirador, Z., xiong, D., Wang, X., gao, METRO., Herrero, SM, & Zhu, h. (2023). Un
atlas of trait associations with resting-state and task-evoked human brain functional organizations
in the UK Biobank. Neurociencia de la imagen. Publicación anticipada.
https://doi.org/10.1162/imag_a_00015
An atlas of trait associations with resting-state and task-evoked human
brain functional organizations in the UK Biobank
Bingxin Zhao1,2,10, Tengfei Li3,4,10, Yujue Li2, Zirui Fan1,2, Di Xiong5, Xifeng Wang5, Mufeng
Gao5, Stephen M. Smith6, and Hongtu Zhu4,5,7,8,9*
1Department of Statistics and Data Science, Universidad de Pennsylvania, Filadelfia, Pensilvania
19104, EE.UU.
2Department of Statistics, Purdue University, West Lafayette, EN 47907, EE.UU.
3Department of Radiology, University of North Carolina at Chapel Hill, Chapel Hill, CAROLINA DEL NORTE
27599, EE.UU.
4Biomedical Research Imaging Center, School of Medicine, University of North Carolina
at Chapel Hill, Chapel Hill, CAROLINA DEL NORTE 27599, EE.UU.
5Department of Biostatistics, University of North Carolina at Chapel Hill, Chapel Hill, CAROLINA DEL NORTE
27599, EE.UU.
6Wellcome Centre for Integrative Neuroimaging, FMRIB, Nuffield Department of Clinical
Neurosciences, Universidad de Oxford, Oxford, Reino Unido.
7Department of Genetics, University of North Carolina at Chapel Hill, Chapel Hill, CAROLINA DEL NORTE
27599, EE.UU.
8Department of Computer Science, University of North Carolina at Chapel Hill, Capilla
Colina, CAROLINA DEL NORTE 27599, EE.UU.
9Department of Statistics and Operations Research, University of North Carolina at
Chapel Hill, Chapel Hill, CAROLINA DEL NORTE 27599, EE.UU.
10These authors contributed equally to this work.
*Autor correspondiente: Hongtu Zhu, 3105C McGavran-Greenberg Hall, 135 Dauer Drive,
Chapel Hill, CAROLINA DEL NORTE 27599. Dirección de correo electrónico: htzhu@email.unc.edu Phone: (919) 966-7250
© 2023 Instituto de Tecnología de Massachusetts. Publicado bajo una atribución Creative Commons 4.0
Internacional (CC POR 4.0) licencia. 1
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
Abstracto
Functional magnetic resonance imaging (resonancia magnética funcional) has been widely used to identify brain
regions linked to critical functions, such as language and vision, and to detect tumors,
strokes, brain injuries, and diseases. It is now known that large sample sizes are
necessary for fMRI studies to detect small effect sizes and produce reproducible results.
Here we report a systematic association analysis of 647 traits with imaging features
extracted from resting-state and task-evoked fMRI data of more than 40,000 Reino Unido
Biobank participants. We used a parcellation-based approach to generate 64,620
functional connectivity measures to reveal fine-grained details about cerebral cortex
functional organizations. The difference between functional organizations at rest and
during task was examined, and we have prioritized important brain regions and
networks associated with a variety of human traits and clinical outcomes. Por ejemplo,
depression was most strongly associated with decreased connectivity
in the
somatomotor network. We have made our results publicly available and developed a
browser framework to facilitate the exploration of brain function-trait association
resultados (http://fmriatlas.org/).
Palabras clave: Brain function; Conectividad funcional; Human traits; Mental Health; descansando
resonancia magnética funcional; Task fMRI; UK Biobank.
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
2
1. Introducción
Functional magnetic resonance imaging (resonancia magnética funcional) is a noninvasive and comprehensive
method of assessing functional organizations of the human brain. By measuring blood
oxygen level dependent (BOLD) signal changes, fMRI can map complex brain functions
and estimate neural correlations between different brain regions (Power et al., 2011).
When the subject is performing a specific task, fMRI can detect brain signals and regions
that link to the task (Ogawa et al., 1990), which is known as task-evoked fMRI. As an
alternativa, resting-state fMRI can observe brain signals during rest and measure
intrinsic functional organization without performing any tasks (Biswal et al., 1995). Ambos
task-evoked and resting-state
fMRIs have been widely used
in clinical and
epidemiological neuroscience
investigación
to explore
el
relationship between
inter-individual variations in brain function and human traits. Por ejemplo, resting-state
functional abnormalities are frequently observed in neurological and psychiatric
disorders,
semejante
como
Alzheimer’s
enfermedad
(Agosta
et
Alabama.,
2012),
attention-deficit/hyperactivity disorder (TDAH) (Posner et al., 2014), schizophrenia (Hu
et al., 2017), and major depressive disorder (MDD) (Mulders et al., 2015). fMRI has also
been used to identify the influence of multi-system diseases and complex traits, como
diabetes (Macpherson et al., 2017), alcohol consumption (Ewing et al., 2014), y
dietary behaviors (Zhao et al., 2017), on brain functions.
A major limitation of most fMRI association studies has been their small sample
tamaño, which is usually less than one hundred or a few hundred. As functional connectivity
measures may be noisy and have large intra-subject variations (Elliott et al., 2020), él
may be difficult to replicate fMRI-trait associations found in small studies (Marek et al.,
2022). This problem can be resolved statistically by increasing the sample size of fMRI
estudios, which can detect weaker signals and reduce the uncertainty of the results. Para
ejemplo, Marek et al. (2022) showed that when the sample size is larger than 2,000,
brain-behavioral phenotype associations can become more reproducible. Sin embargo, el
high assessment costs of fMRI may make it difficult to increase sample sizes sufficiently
to collect the necessary data in every study. In the last few years, several large-scale
fMRI datasets involving over 10,000 subjects have become publicly available, incluido
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
3
the Adolescent Brain Cognitive Development (Chaarani et al., 2021) (ABCD), the Chinese
Imaging Genetics (CHIMGEN) (Xu et al., 2020), and the UK Biobank (Miller et al., 2016)
(UKB). Particularly, the UKB study collected a rich variety of human traits and disease
variables (Bycroft et al., 2018), providing the opportunity to discover and validate
fMRI-trait associations in a large-scale cohort.
Based on fMRI data from more than 40,000 subjects in the UKB study, nosotros
investigated resting-state and
task-evoked
functional organizations and
su
associations with human traits and health outcomes. By processing raw fMRI images
from the UKB study, we represented the brain as a functional network containing 360
brain areas in a parcellation (Glasser et al., 2016) developed using the Human
Connectome Project (Van Essen et al., 2013) (HCP) datos (referred to as the Glasser360
atlas, Higo. 1, Higo. S1, and Table S1). The Glasser360 atlas contained 64,620 (360 × 359/2)
full correlation measures to represent the functional connections among 360 cerebro
areas in 12 functional networks (Ji et al., 2019): the primary visual, secondary visual,
auditory,
somatomotor,
cingulo-opercular, modo por defecto, dorsal attention,
frontoparietal, idioma, posterior multimodal, ventral multimodal, and orbito-affective
redes. Compared to the functional connectome data provided by the UKB study,
which were generated from whole brain spatial independent component analysis (ICA)
(Alfaro-Almagro et al., 2018; beckman & Herrero, 2004; Hyvarinen, 1999), el
parcellation-based approach (like Glasser360) can provide more fine-grained details of
brain functional organizations.
We explored brain-trait associations by performing a systematic analysis with 647
traits and diseases (selected to represent a wide range of traits and health conditions)
using a discovery-validation design. Functional brain regions and networks were found
to be strongly associated with a range of disorders and complex traits. In order to
evaluate how the choice of parcellation may impact our results, we additionally applied
another parcellation (Schaefer et al., 2018) on the same datasets, which divided the
brain into 200 regiones, referred to as the Schaefer200 atlas (Higo. S2 and Table S2). Nosotros
found that the two parcellations can yield similar conclusions and patterns, mientras que el
Glasser360 atlas can provide more biological insights due to its finer partitioning. Nosotros
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
4
also explored the differences between resting-state and task-evoked functional
organizaciones, as well as age and sex-related effects. Numerous studies have
investigated the impacts of age and sex disparities on brain structures and functions.
Sin embargo, the specific locations and patterns of these identified differences can vary
across studies
(Ritchie et al., 2018; Scheinost et al., 2015). Por
leveraging
parcellation-based data from the comprehensive UKB study, our aim is to provide a
more in-depth exploration of differences in resting-state functional connectivity and
their correlations with age and sex.
In order to facilitate the exploration of our extensive results obtained from large-scale
fMRI data, we have developed an
interactive browser
tool, accessible at
http://fmriatlas.org/. This tool acts as a gateway for users to navigate and delve deeper
into our research findings. While we will highlight several pivotal discoveries in the
forthcoming sections of the main body, we urge readers to consult the supplementary
materials and utilize our online tool for a more comprehensive understanding and
discovery of additional patterns. It is worth noting that our bioinformatics resource will
be regularly updated and broadened to include new findings and data. Future updates
will encompass integration with new brain parcellations, alternative data processing
pipelines, and the addition of future large-scale fMRI datasets. These improvements will
further augment the tool’s functionality, keeping it current and providing the research
community with a continually updated platform for the exploration of fMRI data.
2. Material and methods
2.1 Brain imaging data
We generated functional connectivity measures from the raw resting and task fMRI
data downloaded from the UKB data categories 111 y 106, respectivamente. Details of
image acquisition and preprocessing procedures were
summarized
en
el
Supplementary Note. We mapped the preprocessed images onto the Glasser360 atlas
(Glasser et al., 2016), which projected the fMRI data onto a brain parcellation with 360
areas, resulting in a 360 × 360 functional full correlation matrix for each subject (full
correlation). The Glasser360 atlas was originally a surface-based parcellation (Dickie et
Alabama., 2019), and has been converted into a volumetric atlas that is compatible with UKB
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
5
datos. El 360 brain functional areas were grouped into 12 functional networks (Ji et al.,
2019),
including the primary visual, secondary visual, auditory, somatomotor,
cingulo-opercular, modo por defecto, dorsal attention, frontoparietal, idioma, posterior
multimodal, ventral multimodal, and orbito-affective (Table S1). El 64,620 (360 ×
359/2) functional connectivity measures were studied in our main analyses. Estos
high-resolution fMRI traits provided fine details on cerebral cortex functional
organization and allowed us to compare the resting and task-evoked functional
organizaciones.
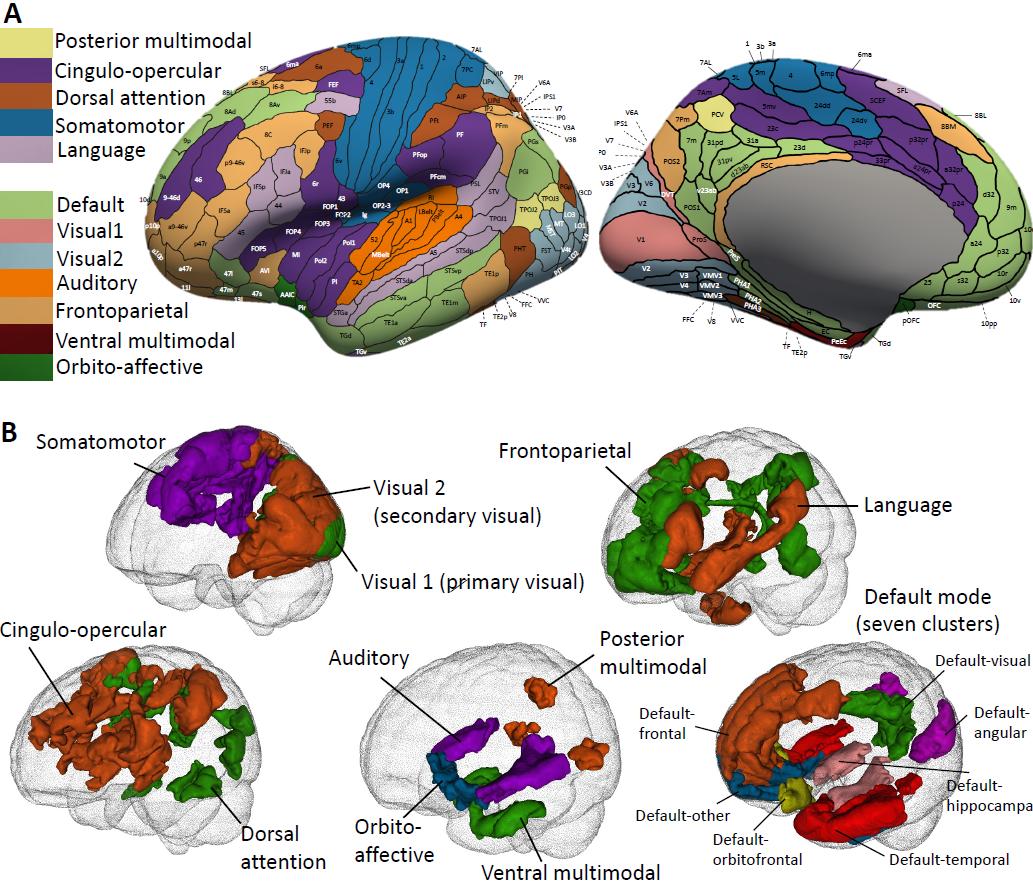
Higo. 1 Illustration of functional areas and networks in the Glasser360 atlas.
(A) Functional areas defined in the Glasser360 atlas (left hemisphere). See Table S1 for
information on these areas and Figure S1 for maps of the whole brain (ambos
hemispheres). Visual1, the primary visual network; Visual2, the secondary visual
6
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
network. (B) Annotation of the 12 functional networks in the human brain. The default
mode network (bottom right) is further divided into seven clusters, mainly based on
their physical locations.
To investigate the potential cross-parcellation variability, we also projected the
fMRI data onto the Schaefer200 atlas (Schaefer et al., 2018) and obtained the 200 × 200
functional connectivity matrices (full correlation, Table S2). The resting and task fMRI
data from the HCP study were also used in our analysis. In addition to functional
connectivity measures, we generated amplitude measures for the brain functional areas
in the Glasser360 atlas, which quantified the brain functional activity (Alfaro-Almagro et
Alabama., 2018; Bijsterbosch et al., 2017; Zou et al., 2008). Precise mathematical definitions
and previous examples of amplitude applications in UKB and HCP studies can be found
in Bijsterbosch et al. (2017).
2.2 Consistency, fiabilidad, and comparison of resting and task fMRI
Following the previous Glasser360 paper (Glasser et al., 2016), we first checked the
group mean maps of two independent sets of UKB subjects (UKB phases 1 & 2 data and
UKB phase 3 datos). In the UKB phase 3 datos, we removed the relatives of early phase
subjects. We obtained the group means for each functional connectivity measure
separately in the two datasets. To measure the similarity/consistency of the two sets of
group means, we calculated their Pearson correlation. For both the resting and task
resonancia magnética funcional, the same analysis was conducted, and we also compared the group mean maps
between resting and task fMRI by using Pearson correlation. Próximo, we evaluated the
intra-subject reliability by using repeated images. We generated and compared the
group mean maps for the original visit and repeated visit separately as we did in the
above two-phase analysis. For each functional connectivity measure, we also checked
the individual-level differences by taking the Pearson correlation across all subjects with
two visits. Finalmente, we repeated the group mean and intra-subject reliability analyses by
using repeated scans in HCP study.
2.3 Age effects and sex differences analysis
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
7
Between 2006 y 2010, approximately half a million participants aged 40 a 69
were recruited for the UKB study. The UKB imaging study is an ongoing project to
re-invite 100,000 UKB participants to collect multi-modal brain and body imaging
datos(Littlejohns et al., 2020). We used the UKB phases 1 a 4 datos (released up through
early 2021, norte = 40,880 for resting fMRI and 34,671 for task fMRI) in our analysis. The age
(at imaging) range of subjects was 44 a 82 (edad media = 64.15, standard error = 7.74)
and the proportion of females was 51.6%. In the age and sex analysis, we fitted the
following model for each fMRI trait: where is the
standardized fMRI trait, is the standardized age, is the sex factor (0 for female and
1 for male), is the set of adjusted covariates, is the main effect of on ,
is the main effect of on , is the effect of age-sex interaction term on ,
represents effects of covariates, and is random error variable. We adjusted the
following covariates: imaging site, head motion, head motion-squared, brain position,
brain position-squared, volumetric scaling, height, weight, body mass index, heel bone
mineral density, and the top 10 genetic principal components. For each continuous trait
or covariate variable, we removed values greater than five times the median absolute
deviation from the median. These removed values will be treated as missing entries in
the dataset. We performed the analysis in a discovery-validation design and only
reported the results that were significant in both discovery and validation datasets (en
different significance levels). Específicamente, as in previous studies (Zhao et al., 2022), nosotros
used the UKB white British subjects in phases 1 a 3 datos (norte = 33,795 for resting and 28,
907 for task) as our discovery sample. The assignment of ancestry in UKB was based on
self-reported ethnicity and has been verified in Bycroft et al. (2018). The UKB non-British
subjects in phases 1 a 3 data and the individuals in newly released UKB phase 4 datos (norte
= 5,961 for resting and 4,884 for task, removed relatives of the discovery sample) eran
treated as the validation sample. We reported P values from the two-sided t test and
focused on the results that were significant at the Bonferroni significance level (7.73 ×
10-7, 0.05/64,620 for the Glasser360 atlas; y 2.51 × 10-6, 0.05/19,900 for the
Schaefer200 atlas) in the discovery dataset and were also significant at nominal
significance level (0.05) in the validation dataset.
2.4 Trait-fMRI association analysis
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
8
For each fMRI trait, we performed linear regression with 647 fenotipos, cual
were selected to reflect a variety of traits and diseases across different domains (Mesa
S3). Específicamente, there were 24 mental health traits (Category 100060), 10 cognitivo
traits (Category 100026), 12 physical activity traits (Category 100054), 6 electronic
device use traits (Category 100053), 8 sun exposure traits (Category 100055), 3 sexual
factor traits (Category 100056), 3 social support traits (Category 100061), 12 familia
history of diseases (Category 100034), 21 diet traits (Category 100052), 9 alcohol
drinking traits (Category 100051), 6 smoking traits (Category 100058), 34 sangre
biochemistry biomarkers (Category 17518), 3 blood pressure traits (Category 100011), 3
spirometry traits (Category 100020), 20 early life factors (Categories 135, 100033,
100034, y 100072), 9 greenspace and coastal proximity (Category 151), 2 hand grip
strength (Category 100019), 13 residential air pollution traits (Category 114), 5
residential noise pollution traits (Category 115), 2 body composition traits by impedance
(Category 100009), 4 health and medical history traits (Category 100036), 3 femenino
specific factors (Category 100069), 1 education trait (Category 100063), 48 curated
disease phenotypes based on Dey et al. (2020), y 386 disease diagnosis coded
according to International Classification of Diseases (ICD-10, Category 2002). Nosotros
selected all diseases in Category 2002 that had at least 100 patients in our resting fMRI
imaging cohort.
For all traits, we adjusted for the effects of age (at imaging), age-squared, sexo,
age-sex interaction, age-squared-sex interaction, imaging site, head motion, cabeza
motion-squared, brain position, brain position-squared, volumetric scaling, height,
weight, body mass index, heel bone mineral density, and the top 10 genetic principal
componentes. Similar to the age and sex analysis, we used the UKB white British subjects
in phases 1 a 3 datos (n = 33,795 for resting and 28, 907 for task) as our discovery
sample and validated our results in the hold-out independent validation dataset (n =
5,961 for resting and 4,884 for task, removed relatives of the discovery sample). Nosotros
reported P values from the two-sided t test and prioritized the results that were
significant at the false discovery rate (FDR) level of 5% in the discovery dataset and were
also significant at the nominal significance level (0.05) in the validation dataset. En
comparison to the conservative Bonferroni correction, the popular FDR multiple testing
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
9
procedimiento (Benjamini & Hochberg, 1995) was more powerful and was consistent with
the exploratory nature of our fMRI-trait analysis. De este modo, we mainly used FDR multiple
testing control in this paper and the subset of associations further passed the stringent
Bonferroni significance level were also provided in our website.
2.5 Prediction models with multiple data types
We built prediction models for fluid intelligence using multi-modality neuroimaging
traits, incluido 64,620 resting fMRI traits, 64,620 task fMRI traits, 215 DTI parameters
from dMRI (Zhao et al., 2021), y 101 regional brain volumes from sMRI (Zhao et al.,
2019). After removing relatives according to Bycroft et al. (2018), we randomly
partitioned the white British imaging subjects into three independent datasets: training
(norte = 20,270), validation (norte = 6,764), and testing (norte = 6,761). The effect sizes of imaging
predictors were estimated from the training data (norte = 20,270). We removed the effects
of age, age-squared, sexo, age-sex interaction, age-squared-sex interaction, imaging site,
head motion, head motion-squared, brain position, brain position-squared, volumetric
scaling, height, weight, body mass index, heel bone mineral density, and the top 10
genetic principal components.
We also integrated other data types into our prediction model, including genetic
variants and several categories of traits studied in our trait-fMRI association analysis
(Table S4). For non-neuroimaging traits, the effect sizes were estimated from all UKB
white British subjects except for the ones in validation and testing data (after removing
parientes). We adjusted for all the covariates listed above for neuroimaging traits, excepto
for the imaging-specific variables including imaging site, head motion, volumetric
scaling, and brain position. The genetic effects were estimated by fastGWA (Jiang et al.,
2019) and were aggregated using polygenic risk scores via lassosum (Mak et al., 2017).
We downloaded imputed genotyping data (Category 100319) and performed the
following quality controls (Zhao et al., 2019): 1) excluded subjects with more than 10%
missing genotypes; 2) excluded variants with minor allele frequency less than 0.01; 3)
excluded variants with missing genotype rate larger than 10%; 4) excluded variants that
failed the Hardy-Weinberg test at 1 × 10-7 nivel; y 5) removed variants with
imputation INFO score less than 0.8. All non-genetic predictors (including neuroimaging
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
10
traits) were modeled using ridge regression via glmnet (Friedman et al., 2010) (R version
3.6.0). All model parameters were tuned in the validation dataset, and we evaluated the
prediction performance on the testing data by calculating the correlation between the
predicted values and the observed ones.
3. Resultados
3.1 Consistency and reliability of the cerebral cortex functional organizations
We examined the consistency and reliability of functional connectivity using
annotations from the Glasser360 atlas in the UKB study. As in Glasser et al. (2016), nosotros
first compared the group means of two independent sets of UKB subjects: the UKB
phases 1 y 2 datos (imaging data released up through 2018 (Zhao et al., 2021), n =
17,374 for resting and 15,891 for task) and the UKB phase 3 datos (data released in early
2020, n = 16,852 for resting and 13,232 for task, removing the relatives of subjects in
early released data). Figure S3 illustrates the consistent spatial patterns of functional
connectivity across the two independent groups. Similar to previous studies of other
conjuntos de datos (Chaarani et al., 2021; Glasser et al., 2016; Herting et al., 2018), the group
mean maps in the two independent datasets of the UKB study were highly similar, con
the correlation (r) a través del 64,620 functional connectivity being 0.996 in resting fMRI
y 0.994 in task fMRI. These results may suggest that the HCP-trained parcellation can
provide a set of well-defined and biologically meaningful brain functional traits in the
UKB datasets.
Próximo, we evaluated the intra-subject reliability of the Glasser360 atlas using the
repeat scans from the UKB repeat imaging visit (norte = 2,771 for resting and 2,014 for task,
average time between visits = 2 años). We performed two analyses. The first analysis is
to compare the group mean maps of the original imaging visit to those of the repeat
visit. Group means were highly consistent between the two visits, with a correlation of
0.997 y 0.994 for resting and task fMRIs, respectivamente (ranges across different
networks were [0.995, 0.999] for resting and [0.987, 0.998] for task, Higo. S4). El segundo
analysis quantified individual-level differences between the two visits. Específicamente, nosotros
evaluated the reliability of each functional connectivity by calculating the correlation
between two observations from all revisited individuals. En general, the correlation was r =
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
11
0.37 (standard error = 0.11) for resting fMRI and r = 0.31 (standard error = 0.08) for task
resonancia magnética funcional (Higo. S5). The correlation of within-network connectivity was generally high in
resting fMRI (Higo. 2A, mean r = 0.46). During task fMRI, the overall correlation was
decreased (mean r = 0.32) and the secondary visual and posterior multimodal networks
exhibited higher functional connectivity on average than others. Además, el
connectivity within activated functional areas (defined by group-level Z-statistic maps,
Supplementary Note) showed a higher correlation than that within nonactivated areas
(Figs. 2B and S6A, mean r = 0.40 vs. 0.30, PAG < 2.2 × 10-16). The majority of the
above-defined activations occurred in the secondary visual, dorsal attention and
somatomotor networks. Furthermore, we examined the reliability of amplitude
measures of fMRI (Alfaro-Almagro et al., 2018; Bijsterbosch et al., 2017; Zou et al.,
2008), which quantified the functional activity within each of the 360 brain areas. The
average amplitude correlation was r = 0.60 (standard error = 0.08) for resting fMRI and r
= 0.45 (standard error = 0.07) for task fMRI (Fig. 2C). In accordance with the findings in
functional connectivity, the reliability of amplitude measurements of activated areas in
task fMRI was higher than that of nonactivated areas (Fig. 2D, mean r = 0.49 vs. 0.43, P =
1.1 × 10-12).
Finally, we compared the spatial patterns of UKB and HCP studies. The correlation
between UKB and HCP was r = 0.90 for resting fMRI and r = 0.78 for task fMRI in the
group mean analysis (Fig. S7). These results demonstrate a substantial level of overall
consistency between the typical subjects in a healthy young adult cohort and those of
middle age and older age. We also examined the reliability of functional connectivity in
the Glasser360 atlas using the repeated scans in the HCP study (n = 1075, average time
between two scans = 1 day). The average correlation was r = 0.40 (standard error = 0.09)
for resting fMRI and r = 0.22 (standard error = 0.11) for task fMRI (the emotion task)
(Fig. S6B). These results show that the two studies have similar reliability, suggesting
that the quality of fMRI traits in the biobank-scale UKB study is comparable to that of
the HCP project. Similar to the UKB study, the connectivity among activated functional
areas (defined by group-level Z-statistic maps, Supplementary Note) had higher
reliability than the nonactivated connectivity in HCP task fMRI (Fig. S6C, mean r = 0.382
vs. 0.225, P < 2.2 × 10-16). In general, the excellent group mean map consistency, as well
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
12
as the similar reliability between the UKB and the HCP studies, provides confidence that
the Glasser360 atlas will be able to consistently annotate the functional organization of
typical subjects in a healthy population. On the other hand, the relatively low
intra-subject reliability of fMRI matches previous findings (Elliott et al., 2020), which
may suggest that a large sample size is needed to produce reproducible association
results in downstream analyses (Marek et al., 2022).
Fig. 2 Reliability across brain functional areas and networks.
(A) Comparison of reliability of functional connectivity across 12 brain functional
networks in resting (left panel) and task (right panel) fMRI. (B) Comparison of reliability
of functional connectivity between the activated areas (within activation) and the
nonactivated areas (out of activation) in task fMRI. (C) Comparison of reliability of
amplitude measures in resting (left panel) and task (right panel) fMRI. See Table S1 for
information of the labeled brain areas. (D) Comparison of reliability of amplitude
measures between the activated areas (within activation) and the nonactivated areas
(out of activation) in task fMRI.
13
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
3.2 Comparison of resting-state and task-evoked functional organizations
The correlation between resting fMRI and task fMRI group mean maps was 0.754 in
the UKB study and 0.782 in the HCP study, indicating the high degree of similarity
between
intrinsic and extrinsic functional organizations (Fig. S7). Resting-task
differences were observed across different networks. For example, in the auditory
network, task fMRI revealed stronger intra-hemispheric connections than resting fMRI
(mean = 0.482 vs. 0.314, P = 5.6 × 10-11), while the inter-hemispheric connections in task
fMRI generally weakened (mean = 0.214 vs. 0.280, P = 8.0 × 10-6). Task-related changes
were more complex in the default mode network. To summarize the patterns, we
grouped the 77 areas in the default mode network into seven clusters, mainly based on
their physical locations. We found that functional connectivity within the frontal, visual,
and hippocampal clusters was stronger in task fMRI than in resting fMRI (mean = 0.314
vs. 0.384, P = 1.7 × 10-9), while the connectivity between the frontal and the other two
clusters decreased (mean = 0.191 vs. 0.086, P < 2.2 × 10-16). Moreover, the frontal
cluster of the default mode network can be further divided into two subclusters, the
first subcluster consisted of left/right 9a, 9m, 9p, 8BL, 8Ad, and 8Av areas, mainly in the
dorsolateral superior frontal gyrus (referred to as the dorsolateral superior subcluster);
and the second one included left/right 10v, 10r, p32, a24, and 10d areas in the medial
orbital superior frontal gyrus and pregenual anterior cingulate cortex (referred to as the
medial orbital superior subcluster). The dorsolateral superior subcluster had decreased
connectivity with the areas in other clusters of the default mode network in task fMRI,
especially those in the temporal cluster. On the other hand, the medial orbital superior
subcluster had a greater level of connectivity with a few other areas of the default mode
network when performing the task, especially with the orbitofrontal complex (OFC)
cluster and the neighboring 10pp area. Furthermore, the visual cluster maintained
strong intra-cluster connectivity during the task, whereas its connectivity with the
angular, frontal, and temporal clusters decreased (mean = 0.271 vs. 0.177, P < 2.2 ×
10-16).
Several areas of the secondary visual network were less connected to other visual
areas when the task was performed, including the left/right V6A (in the superior
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
14
occipital), V6 (in the cuneus), VMV1 (in the lingual gyrus), and VMV2 (in the lingual and
fusiform gyrus). Interestingly, some of these visual areas, such as the left/right V6, had
increased functional connectivity with the default mode network. There was also an
increase in connections between the default mode network and other major cognitive
networks, such as the cingulo-opercular and frontoparietal. For the somatomotor
network, the insula-related areas (including left/right Ig, FOP2, OP2-3, and right RI) had
reduced connections with other somatomotor areas in task fMRI. Similar to the auditory
network, the inter-hemispheric connectivity in the cingulo-opercular network decreased
in task fMRI. Additionally, we found that the dorsal attention, frontoparietal, and
language networks had similar functional connectivity patterns in resting and task fMRI.
In summary, our results confirm the similarity of functional structures between resting
and task fMRI, while also
identifying specific patterns of differences. These
network-specific patterns can be explored on our website http://fmriatlas.org/.
3.3 Age effects and sex differences in functional organizations
By using the large-scale fMRI data, we quantified the age and sex effect patterns on
resting and task functional organizations. We used unrelated white British subjects in
UKB phases 1-3 data release (until early 2020) as our discovery sample (n = 33,795 for
resting and 28, 907 for task) and validated the results in an independent hold-out
dataset, which included non-British subjects in UKB phases 1-3 data release and all
subjects in UKB phase 4 data release (early 2021 release, removed the relatives of our
discovery sample, n = 5, 961 for resting and 4, 884 for task). The full list of the adjusted
covariates can be found in the Methods section. Below we highlighted the results
passing the stringent Bonferroni significance level (7.73 × 10-7 = 0.05/64,620) in the
discovery dataset and being significant at the nominal significance level (0.05) in the
validation dataset.
There were widespread age effects on functional connectivity of resting and task
fMRI, and network and area-specific details were revealed (Figs. S8A-B). For example, as
age increased, the connections within the auditory, secondary visual, somatomotor,
language, and cingulo-opercular networks were generally weaker. Some areas had
particularly large age-effects, such as the left/right PoI2 (the posterior insular area 2)
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
15
areas in the cingulo-opercular network. However, both positive and negative age effects
were observed in the frontoparietal and default mode networks (Fig. S9). For example,
the left/right POS2 (the parieto-occipital sulcus area 2) areas in the frontoparietal
network and left/right POS1 (the parieto-occipital sulcus area 1) areas in the default
mode network had strong aging effects. Negative age effects in the default mode
network were strongest in the hippocampal cluster, such as the left/right PHA1 (the
parahippocampal area 1) areas.
In task fMRI, age effects were different from those in resting fMRI. We highlighted
a few patterns. First, the age effects in the auditory network were mainly on the
inter-hemispheric connections, where the connectivity between the left and right
hemispheres decreased with aging. Similarly, the
inter-hemispheric connectivity
between the auditory and cingulo-opercular networks declined as we aged. The age
effects on intra-hemispheric connections were much weaker. Except for a few areas
(such as the right 8Ad and right PEF), most areas in the cingulo-opercular and default
mode networks had reduced functional connectivity with aging (Fig. S10). On the other
hand, most of the functional connectivity in the secondary visual network increased
with aging, especially the left/right V3A and V6A areas in the superior occipital gyrus.
There were both positive and negative effects of aging on other networks, such as
somatomotor, frontoparietal, and dorsal attention. Overall, these results describe the
detailed age effect pattern for functional organizations at rest and during task
performance.
We also examined the age effects on amplitude measures. In resting fMRI,
age-related decreases in brain activity were observed in most brain areas, with the
strongest effects in left and right PreS areas (the presubiculum, a subarea of the
parahippocampal region, < -0.222, P < 5.01 × 10-193, Fig. 3A). In task fMRI, however,
both strong positive and negative effects on brain activity were widely observed (Fig.
3B). Because widespread age effects were detected on both functional connectivity and
amplitude traits, we examined the conditional age effects on functional connectivity
traits after additionally including amplitude traits as covariates. After adjusting for
amplitude traits, most of the age effects on functional connectivity traits became much
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
16
smaller and were not significant at the Bonferroni significance level, especially in resting
fMRI (Fig. S11). For example, although a few of the strongest amplitude-adjusted age
effects remained significant, most of the other moderate amplitude-adjusted age effects
failed to pass the Bonferroni significance level in the default mode network. Overall,
these results for amplitude traits indicate that age has a significant effect on the
variation of amplitude traits across subjects, which may also be carried over to
functional connectivity traits (Bijsterbosch et al., 2017).
Functional connectivity patterns differed between males and females. We found
widespread sex differences across different resting fMRI networks, with the strongest
differences occurring in the somatomotor network (Fig. S8C). Males had stronger
functional connectivity in the somatomotor and auditory networks as well as a few
specific areas, including the left/right VIP (in the superior parietal gyrus), LIPv (in the
superior parietal gyrus), PH (in the inferior temporal gyrus), and V6A (in the superior
occipital gyrus) of the secondary visual network, the left/right PFcm (in the superior
temporal gyrus) and 43 (in the rolandic operculum) of the cingulo-opercular network,
the left/right a9-46v and p9-46v (both in the middle frontal gyrus) of the frontoparietal
network, and the left/right PGp (in the middle occipital gyrus) of the dorsal attention
network. In the default mode network, the sex difference had a complicated pattern.
Specifically, males had stronger connectivity in the hippocampal and OFC clusters,
especially in the left 47m area of the posterior orbital gyrus. On the other hand, females
had stronger connectivity in many other areas of the default mode network (Fig. S12).
We observed significant sex differences in task fMRI within several brain regions.
These include the right V6A (located in the superior occipital gyrus) and left VMV2
(found in the lingual and fusiform gyrus) within the secondary visual network, the
left/right PHA3 (situated in the fusiform gyrus) within the dorsal attention network, and
the left/right RSC (located in the middle cingulate cortex) of the frontoparietal network
(P < 7.73 × 10-7, refer to Figs. S13A-C). Within the language, auditory, and somatomotor
networks, males exhibited stronger functional connectivity than females in numerous
brain regions (see Figs. S13D-F). Additionally, males had stronger connectivity in the
hippocampal and frontal areas of the default mode network, whereas females had
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
17
stronger connectivity between the visual cluster and the frontal cluster (Fig. S14). As for
the amplitude measures, females had stronger brain activity in many areas of the
default mode network, whereas males had stronger brain activity in most other
networks in resting fMRI (Fig. 3C). Sex differences were generally reduced in task fMRI
amplitude measurements (Fig. 3D). Lastly, we estimated the amplitude-adjusted sex
effects on functional connectivity traits by additionally controlling for the amplitude
traits as covariates. Similar to the findings of the age effects, the majority of
amplitude-adjusted sex effects on functional connectivity traits can be explained by
amplitude traits, such as in the somatomotor and default mode networks (Fig. S15).
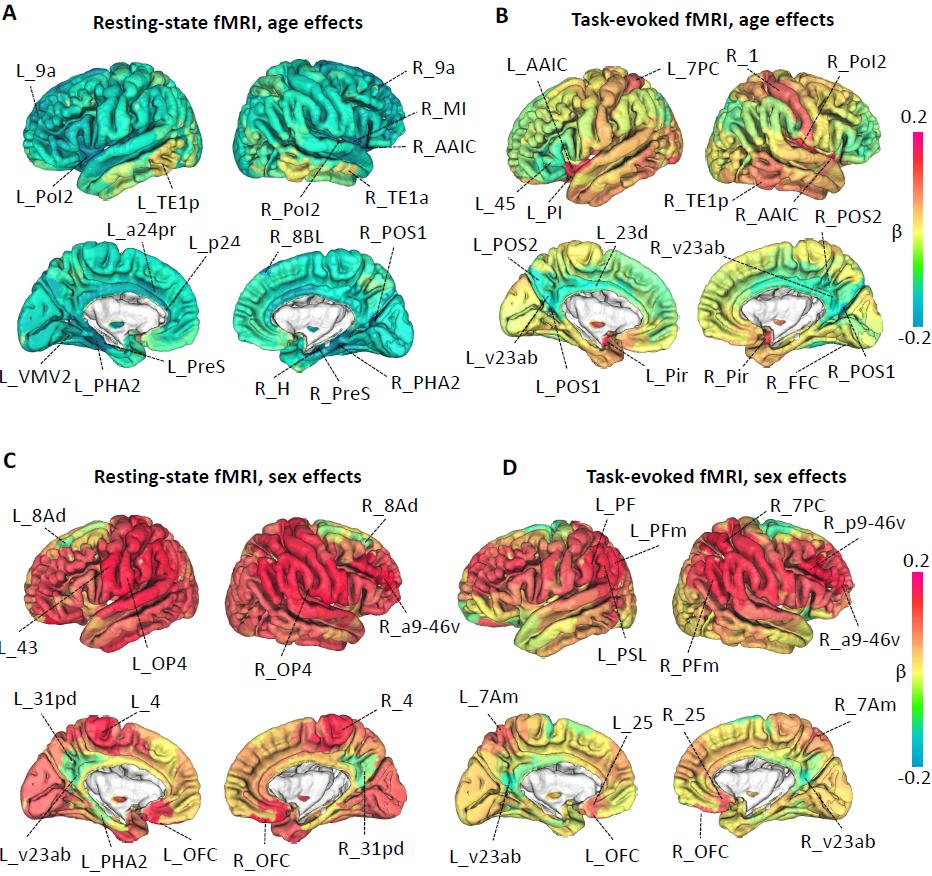
Fig. 3 Spatial pattern of age and sex effects on brain functional organizations.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
18
We illustrate the spatial pattern of age effects on amplitude measures in (A) for resting
fMRI and in (B) for task fMRI. See Table S1 for information on the labeled brain areas.
(C) and (D) display the spatial pattern of sex effects on amplitude measures of resting
and task fMRI, respectively. We labeled the brain areas with the strongest age and sex
effects in amplitude measures.
3.4 An atlas of trait associations with cerebral cortex functional areas
We aimed to explore the associations between resting and task functional
organizations and 647 phenotypes. Similar to the age and sex analyses, we used
unrelated white British subjects in UKB phases 1-3 data release as the discovery sample
(n = 33,795 for resting and 28, 907 for task) and validated the results in an independent
hold-out dataset (n = 5, 961 for resting and 4, 884 for task). We prioritized significant
associations that survived at the FDR 5% level in the discovery sample and remained
significant at the nominal significance level (0.05) in the validation sample. Among the
647 traits, 120 had at least one significant association with resting fMRI functional
connectivity measures, among which 82 further survived the Bonferroni significance
level (7.73 × 10-7, 0.05/64,620) (Table S3). We detail below the patterns of associations
relating to mental health, cognitive function, and disease status. For the complete set of
results, please visit http://165.227.92.206/traitList.html.
We observed strong associations between resting fMRI and multiple mental health
traits, including risk-taking, depression, MDD, and neuroticism. Enrichments in specific
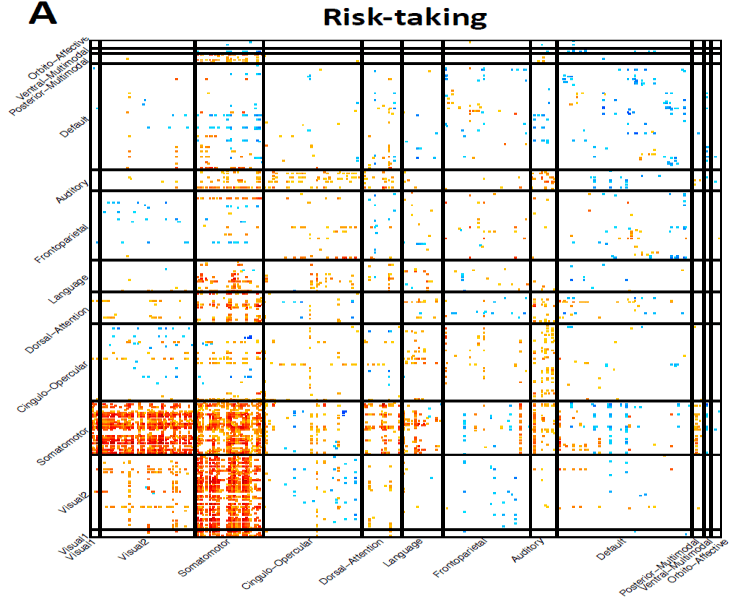
networks and brain areas were observed. For example, risk-taking (Data field 2040) was
strongly positively associated with the somatomotor network and the connections
between the somatomotor and visual networks (Fig. 4A). Risk-taking was also negatively
associated with the functional connections of the default mode network. Functional
connectivity of sensory/motor areas was recently found to be positively associated with
risk-taking
(Rolls et al., 2022) and our
findings were consistent with the
“sensory-motor-cognitive” mode of brain functional amplitude changes related to aging
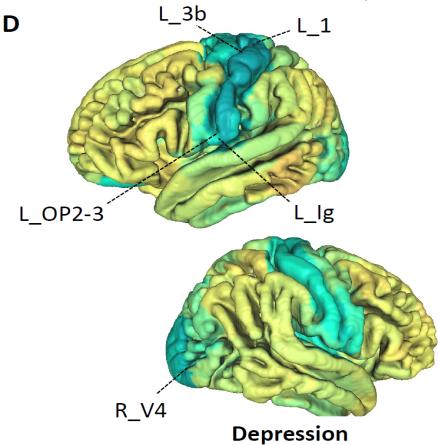
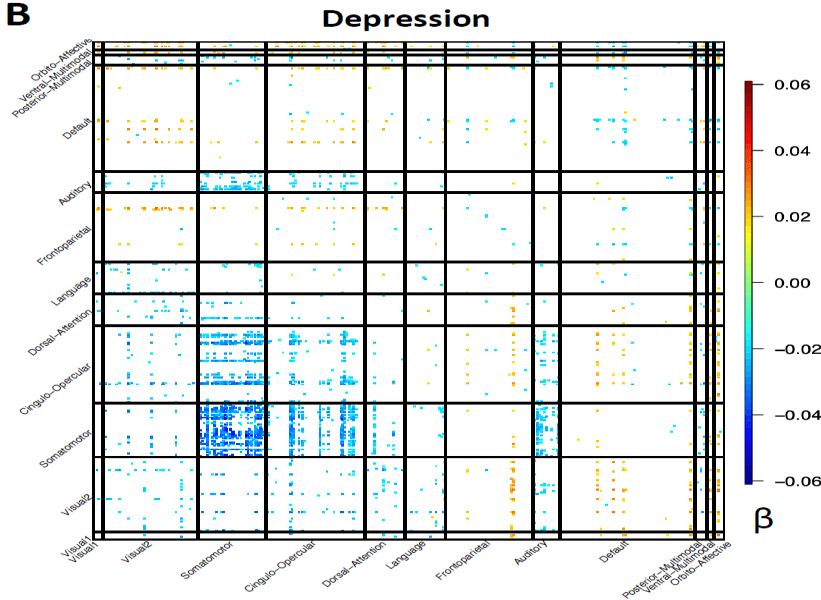
(Smith et al., 2020). In addition, depression was mostly associated with reduced
connectivity in the somatomotor and cingulo-opercular networks (curated disease
phenotype based on ICD-10 codes, Fig. 4B). Consistent patterns were also observed in
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
19
MDD (ICD-10 code F329), nervous feelings (Data field 1970), seen doctor for nerves
anxiety tension or depression (Data field 2090), neuroticism score (Data field 20127),
and suffer from nerves (Data field 2010).
Multiple cognitive traits were associated with functional connectivity in fMRI, such
as fluid intelligence (Data field 20016), the number of puzzles correctly solved (Data field
6373), duration to complete alphanumeric path (Data field 6350), and maximum digits
remembered correctly (Data field 4282). These cognitive traits showed different
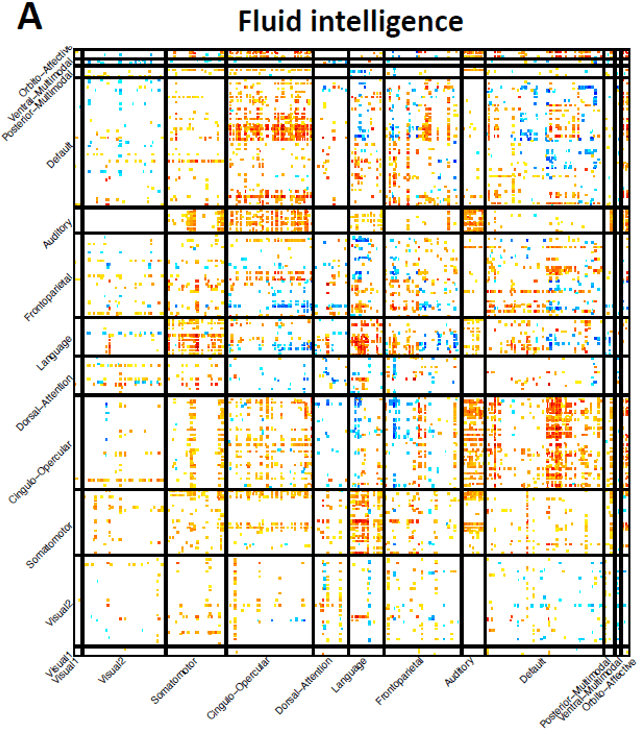
association patterns. Fluid intelligence, for example, was associated with functional
connectivity in the auditory, language, cingulo-opercular, dorsal attention, and default
mode networks, most of the associations were positive (Fig. 5A). The duration to
complete alphanumeric path was mainly negatively associated with functional
connectivity in the secondary visual network (Fig. S16A), the number of puzzles
correctly solved was mostly related to the functional connectivity within the default
mode, somatomotor, and secondary visual networks (Fig. S16B), and the maximum
digits remembered correctly was positively related to the auditory and language
networks (Fig. S16C). The links between brain function and several other brain-related
complex traits were detected, such as the strong connections between handedness
(Data field 1707) and the cingulo-opercular network (Fig. S16D). Resting functional
connectivity was also widely associated with lifestyle and environmental traits, including
physical activity, electronic device use, smoking, diet, alcohol, and sun exposure. For
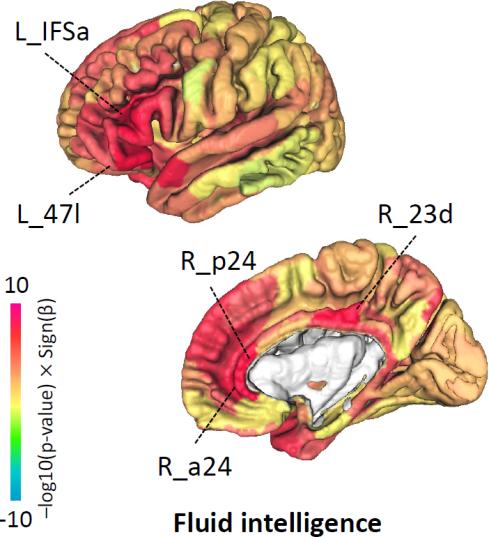
example, watching television (TV) for longer periods of time (Data field 1070) may
weaken functional connectivity in the somatomotor and visual networks as well as
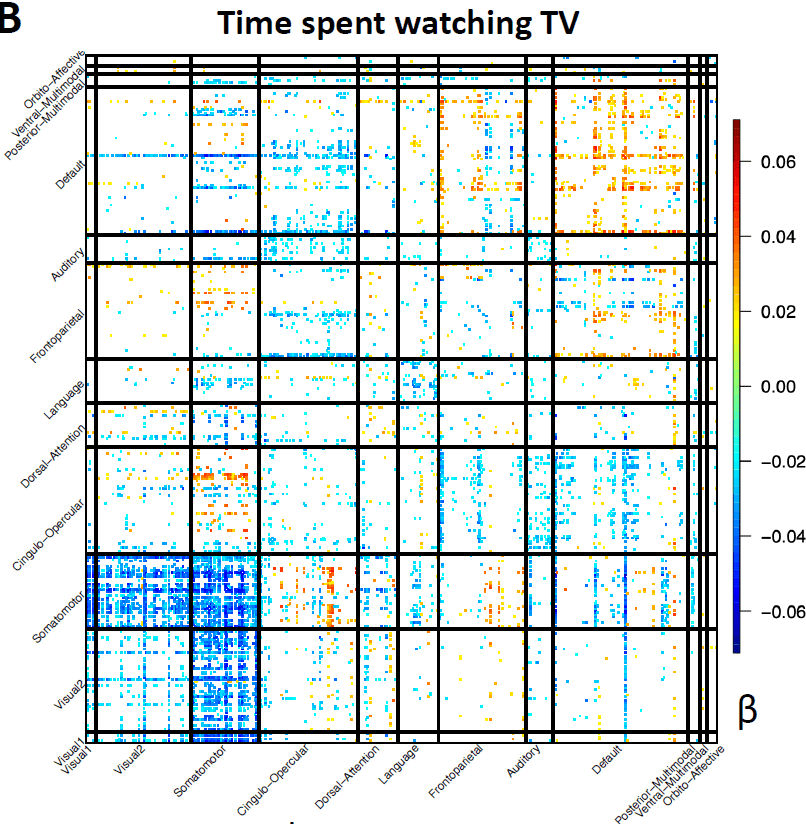
strengthen functional connectivity in the default mode network (Fig. 5B).
Strong associations between increased functional connectivity and cardiovascular
diseases were identified, including atrial fibrillation (curated disease phenotype and
ICD-10 code I48), vascular/heart problems diagnosed by doctor (Data field 6150), and
hypertension (curated disease phenotype and ICD-10 code I10). Atrial fibrillation is the
most common clinically significant arrhythmia, and increasing evidence suggests it is
associated with cognitive decline and dementia (Alonso & de Larriva, 2016). We found
that atrial fibrillation was widely associated with functional connectivity across different
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
20
networks (Figs. S17A-B). Hypertension and vascular/heart problems were associated
with reduced functional connectivity in the auditory, somatomotor, secondary visual,
and cingulo-opercular networks (Figs. S17C-D). Hypertension is a major risk factor for
vascular dementia and Alzheimer’s Disease and altered functional connections may
reflect the early effects of vascular risk factors on brain functions (Carnevale et al.,
2020).
In task fMRI, 96 traits had at least one significant association at the FDR 5% level
(and significant at the nominal level in the validation dataset), and 59 further survived
the Bonferroni significance level (7.73 × 10-7 = 0.05/64,620) (Table S3). Of the 96 traits,
69 were also significantly associated with resting fMRI at the 5% FDR level. The
association patterns in task and resting fMRI were very similar for a few traits, such as
atrial fibrillation (Fig. S18). For many traits, however, we observed different patterns in
resting and task fMRI, including fluid intelligence (Figs. S19A-B) and the number of
puzzles correctly solved (Figs. S19C-D) (P < 2.2 × 10-16). For example, both fluid
intelligence and the number of solved puzzles were positively associated with
intra-hemispheric connections of the auditory network in task fMRI, whereas no or
negative associations were observed with inter-hemispheric connections. There were
similar intra- and inter-hemispheric connection differences in the cingulo-opercular
network.
We also quantified the association patterns with amplitude traits and prioritized
brain areas whose functional activity was related to traits and diseases. We observed
similar patterns to the functional connectivity results. For example, risk-taking has the
strongest associations with the brain activity of the postcentral gyrus
in the
somatomotor network, especially the primary somatosensory cortex (Rolls et al., 2022)
(Fig. 4C, > 0.033, PAG < 8.14 × 10-6). The postcentral gyrus, insula, and Rolandic
operculum areas of the somatomotor network were most negatively related to
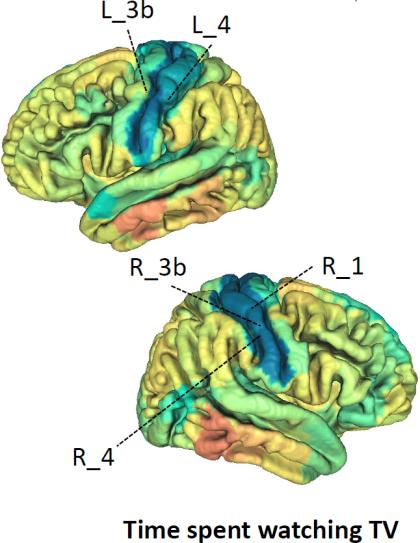
depression (Fig. 4D, < -0.036, P < 7.10 × 10-7). All significant associations with fluid
intelligence were positive, with the top three areas being the middle cingulate, anterior
cingulate, and orbital part of the inferior frontal gyrus (IFG pars orbitalis) in the default
mode network (Fig. 5C, > 0.053, PAG < 1.31 × 10-12). Time spent watching TV was strongly
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
21
negatively associated with the postcentral gyrus, precentral gyrus, paracentral lobule,
and the supplementary motor area in the somatomotor network (Fig. 5D, < -0.050, P <
2.03 × 10-12).
3.5 Alternative analyses using the Schaefer200 atlas
Fig. 4 Selected complex traits that were associated with brain functional organizations.
(A) Associations between risk-taking (Data field 2040) and functional connectivity of
resting fMRI. This figure and the top-ranked brain areas can be viewed in an interactive
version at http://165.227.92.206/trait/trait85.html.
(B) Associations between
depression (curated disease phenotype) and functional connectivity of resting fMRI. This
figure and the top-ranked brain areas can be viewed in an interactive version at
http://165.227.92.206/trait/trait230.html. We illustrated the estimated correlation
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
22
coefficients that were significant at FDR 5% level in the discovery sample (n = 33,795)
and were also significant at the nominal significance level (0.05) in the validation dataset
(n = 5, 961). (C) and (D) display the spatial pattern of associations with amplitude
measures of resting fMRI for risk-taking and depression, respectively. Brain areas with
the strongest associations were labeled. See Table S1 for information on these areas.
Fig. 5 Selected complex traits that were associated with brain functional organizations.
(A) Associations between fluid
intelligence (Data field 20016) and functional
connectivity of resting fMRI. This figure and the top-ranked brain areas can be viewed in
an interactive version at http://165.227.92.206/trait/trait158.html. (B) Associations
between time spent watching TV (Data field 1070) and functional connectivity of resting
fMRI. This figure and the top-ranked brain areas can be viewed in an interactive version
at http://165.227.92.206/trait/trait101.html. We illustrated the estimated correlation
23
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
coefficients that were significant at FDR 5% level in the discovery sample (n = 33,795)
and were also significant at the nominal significance level (0.05) in the validation dataset
(n = 5, 961). (C) and (D) display the spatial pattern of associations with amplitude
measures of resting fMRI for fluid intelligence and time spent watching TV, respectively.
Brain areas with the strongest associations were labeled. See Table S1 for information
on these areas.
The brain parcellation may play a crucial role in the definition of the brain
functional network and affect the results of downstream analysis (Popovych et al., 2021).
To explore the impact of parcellation choice on the large-scale UKB study, we
additionally applied another parcellation (the Schaefer200 atlas (Schaefer et al., 2018))
and repeated our analysis of the same set of subjects. Briefly, the Schaefer200 atlas
partitioned the brain into 200 regions, resulting in 19,900 pairwise functional full
correlation measures (200 × 199/2). We mapped the 200 regions onto the same 12
networks used in the Glasser360 atlas (Table S2).
The average reliability in the Schaefer200 atlas was r = 0.387 (standard error =
0.10) for resting fMRI and r = 0.312 (standard error = 0.07) for task fMRI, which was in
the same range as the Glasser360 atlas. Figure S20 compares the reliability of the two
parcellations. Glasser360 and Schaefer200 atlases showed similar patterns across a
variety of networks, with the largest differences being observed in the secondary visual
network, where the Glasser360 atlas was more reliable. In addition, consistent spatial
patterns of functional connectivity were observed in the two parcellations, although the
strength of connectivity was slightly higher in the Schaefer200 atlas, which may partly
be explained by the smaller number of brain areas (Fig. S21). These results demonstrate
the good generalizability of functional organizations modeled by the Glasser360 atlas.
We evaluated the age and sex effects in the Schaefer200 atlas. Figure S22
compares the age effect patterns in the Schaefer200 and Glasser360 atlases. In both
atlases, decreasing resting functional connectivity was consistently associated with
aging, especially in the auditory, cingulo-opercular, and somatomotor networks. The
main difference was in the secondary visual network, where the age effects in the
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
24
Glasser360 atlas were stronger than those in the Schaefer200 atlas. This finding may be
attributed to the lower reliability of the Schaefer200 atlas in the secondary visual
network, suggesting that the Glasser360 atlas may be more suitable for studying the
brain connectivity of
the visual cortex.
In addition, consistent
intra- and
inter-hemispheric association differences in task fMRI were observed. The Schaefer200
and Glasser360 atlases also showed similar sex effect patterns, in which the strongest
effects were both detected in the somatomotor and auditory networks (Fig. S23).
Next, we repeated the association analysis with the 647 traits. In resting fMRI, 131
traits had at least one significant association at the FDR 5% level and 83 further passed
the Bonferroni significance level (2.51 × 10-6 = 0.05/19,900, also passing the nominal
significance level (0.05) in the independent validation dataset, Table S3). Of the 120
traits with significant associations in the Glasser360 atlas analysis, 109 (90.83%) were
also significant in the Schaefer200 atlas analysis. Additionally, the association maps
were largely consistent in the two atlases. For example, time spent watching TV was
consistently associated with decreased functional connections of the somatomotor and
visual networks, as well as increased functional connectivity in the default mode
network (Figs. S24A-B). Moreover, fluid intelligence was consistently linked to increased
functional connectivity, particularly in the language and auditory networks (Figs.
S24C-D).
In both atlases, depression was associated with reduced functional
connectivity in the somatomotor and cingulo-opercular networks (Fig. S25). At the FDR
5% level, 90 traits showed significant associations with task fMRI, including 76 of the 96
(79.2%) traits that were significant in the Glasser360 atlas analysis. All these results are
available on our website. In summary, the Schaefer200 atlas results agree well with
those of the Glasser360 atlas, indicating that the patterns observed in our Glasser360
analysis are not parcellation-specific.
Finally, we examined the trait associations with 1,701 functional connectivity traits
based on the whole brain spatial ICA (Alfaro-Almagro et al., 2018; Beckmann & Smith,
2004; Hyvarinen, 1999) approach in resting fMRI. These ICA functional connectivity
traits
were
available
from
the
UK
Biobank
data
release
(https://www.fmrib.ox.ac.uk/ukbiobank/index.html, Data fields 25752 and 25753),
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
25
which were partial correlations and the timeseries were estimated from group ICA maps
via the dual-regression(Alfaro-Almagro et al., 2018). Of the 647 traits, 76 demonstrated
at least one significant association at the FDR 5% level and 58 remained significant at
the Bonferroni significance level (2.94 × 10-5 = 0.05/1,701, also passing the nominal
significance level in the independent validation dataset). Among the 76 ICA-significant
traits, 65 (85.53%) were also significant in the above Glasser360 atlas analysis.
Compared to the ICA-derived traits, parcellation-based traits from the Glasser360 atlas
(which identified significant associations with 120 complex traits at the FDR 5% level and
82 at the Bonferroni significance level) were able to detect associations with more traits.
In addition, we ranked the 58 ICA-significant complex traits (at the Bonferroni
significance level) by the number of their significant associations with ICA-derived traits.
Then we compared the association strengths of the top ten traits with ICA-derived traits
and those with Glasser360 traits. On these ten traits, ICA-derived traits and Glasser360
traits showed similar levels of association strength (Fig. S26). For example, many
ICA-derived and Glasser360 traits were found to be significantly associated with systolic
blood pressure (Data field 4080), and most of these associations were in a similar range
of effect size (Fig. S27). These results align with the results of a recent study on the
functional connectome signature of blood pressure (Jiang et al., 2023). The results of
Glasser360 traits indicate that the auditory and somatomotor networks may be more
strongly associated with systolic blood pressure than other networks. These networks
and areas may be targeted when studying hypertension-related cognitive dysfunction
and brain functional damages (Carnevale et al., 2020; Naumczyk et al., 2017). In
summary, parcellation-based traits may reveal more network and area-level details with
comparable association strength to ICA-derived traits.
3.6 Fluid intelligence prediction by integrating multiple data types.
Our association analyses demonstrate the potential value of large-scale fMRI data
for a variety of complex traits and disorders in clinical and epidemiological research. For
example, it is of great interest to construct prediction models by integrating fMRI data
and other data types (He et al., 2020; Pervaiz et al., 2020; Shen & Thompson, 2019).
Fluid intelligence is a key indicator of cognitive ability and is associated with multiple
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
26
neurological and neuropsychiatric disorders (Keyes et al., 2017). In this section, we
performed prediction for fluid intelligence using neuroimaging traits from multiple
modalities, including resting fMRI, task fMRI, diffusion MRI (dMRI) (Zhao et al., 2021),
and structural MRI (sMRI) (Zhao et al., 2019). We further integrated these neuroimaging
data with a wide range of other data types, including common genetic variants,
biomarkers, local environments, early life factors, diet, and behavioral traits. The
relative contributions and joint performance of these data types were assessed in a
training, validation, and testing design. All model parameters were tuned using the
validation data and we evaluated the prediction performance on the independent
testing data by calculating the correlation between the predicted values and the
observed intelligence, while adjusting for the covariates listed in the Methods section.
The prediction performance of multi-modality neuroimaging traits was summarized
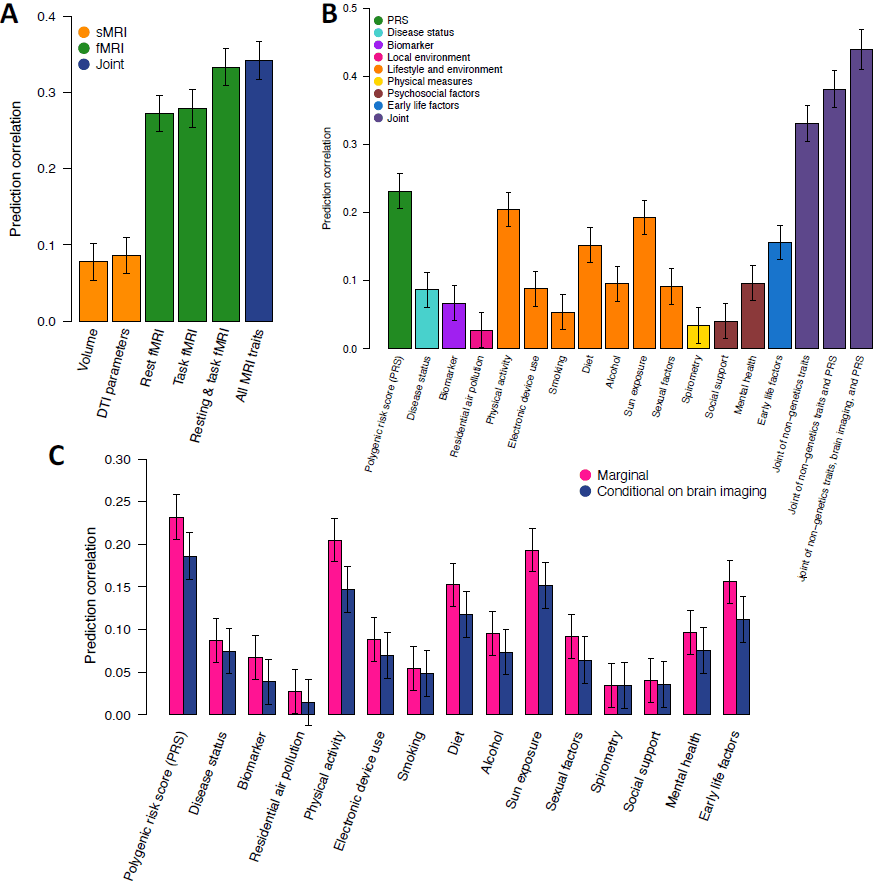
in Figure 6A. The prediction correlation of resting fMRI was 0.272 (standard error =
0.012), suggesting that about 7.4% variation in fluid intelligence can be predicted by
resting fMRI connectivity. The prediction correlation was similar
in task fMRI
(correlation = 0.279) and was improved to 0.333 by jointly using resting and task fMRI,
which suggests that resting and task fMRI had different contributions to intelligence
prediction. This improvement aligned with previous results reported in the HCP and
Philadelphia Neurodevelopmental Cohort (PNC) studies (Gao et al., 2019), and matched
our association results where both resting and task fMRI showed strong associations
with fluid intelligence with different spatial patterns. In addition, the dMRI and sMRI
traits had much lower prediction accuracy than fMRI traits. Specifically, the prediction
correlation was 0.09 for diffusion tensor imaging (DTI) parameters of dMRI and 0.08 for
regional brain volumes of sMRI. Moreover, adding these structural traits in addition to
fMRI traits did not substantially improve the prediction performance (correlation =
0.342), indicating the prediction power of brain structural traits for intelligence can be
largely captured by the functional traits.
Next, we examined the prediction performance of non-neuroimaging data types
(Fig. 6B). The prediction correlation of intelligence genetic polygenic risk score was
0.232 (standard error = 0.013), which was slightly lower than the performance of resting
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
27
fMRI. Several categories of lifestyle and environmental traits had strong predictive
power, including physical activity (correlation = 0.205), sun exposure (correlation =
0.193), and diet (correlation = 0.153). Moreover, biomarkers, disease records, and early
life factors all had significant predictive performance, with prediction correlations being
0.067, 0.087, and 0.156, respectively. By combining all these non-neuroimaging data
types, the prediction correlation increased to 0.381. The performance was further
improved to 0.440 by including neuroimaging data, which was much higher than when
using only one type of data.
To explore whether the predictive power of non-neuroimaging traits can be
explained by brain structural and functional variations, we evaluated their conditional
predictive performance on fluid intelligence after controlling for neuroimaging traits.
There was a reduction of performance on multiple categories of non-neuroimaging
predictors, suggesting their effects on intelligence may be indirect and partially
mediated by brain structure and function (Fig. 6C and Table S4). For example, the
prediction performance of the polygenic risk score decreased from 0.232 to 0.186,
indicating that 19.8% of the genetic predictive power on intelligence can be captured by
brain structural and functional variations measured by brain MRI. The proportion was
28.3% for physical activity, 23.1% for diet, and 28.6% for early life factors. Overall, these
results illustrate that neuroimaging traits, especially the ones from resting and task
fMRI, are powerful predictors of cognitive function. Future studies can integrate genetic,
biomarker, behavioral/environmental factors, and multi-modality MRI data for better
prediction of brain-related complex traits and disorders.
4. Discussion
Inter-individual variations in brain function and their relationship to human health
and behavior are of great interest. The intra-individual reliability of brain fMRI traits is
generally low, although the group-level consistency is high (Chaarani et al., 2021; Elliott
et al., 2020; Herting et al., 2018; Noble et al., 2021). Then it has been suggested that a
large sample size is needed for fMRI studies to detect trait associations with small effect
sizes (Kennedy et al., 2021; Smith & Nichols, 2018). The UKB study provided an
extensive biobank-scale data resource for quantifying fMRI associations with many
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
28
phenotypes. The present study conducted a systematic analysis of intrinsic and extrinsic
functional organizations with a parcellation-based approach using fMRI data collected
from over 40,000 individuals. We measured differences between resting and task fMRI,
investigated age and sex effects on brain function, and examined the cross-parcellation
variability of our findings. We explored the fMR’s association with 647 traits chosen
from
Fig. 6 Integrative prediction model for fluid intelligence.
(A) Prediction accuracy of neuroimaging traits for fluid intelligence. Volume, region
brain volumes from brain structural MRI (sMRI); DTI parameters, diffusion tensor
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
29
imaging parameters to measure brain white matter microstructures; All MRI traits,
including brain volume, DTI parameters, resting fMRI, and task fMRI. (B) Prediction
accuracy of non-neuroimaging traits from different trait categories and their joint
performance. PRS, polygenic risk scores of genetic variants. (C) Comparison of predictive
power of non-neuroimaging traits before (“marginal”) and after controlling for the
neuroimaging traits (“conditional on brain imaging”).
a variety of trait domains. In comparison to the prior literature (Miller et al., 2016),
which applied data-driven spatial ICA (Alfaro-Almagro et al., 2018; Beckmann & Smith,
2004; Hyvarinen, 1999) to about 5000 subjects, the parcellation-based approach and
much larger sample size allowed us to quantify functional organizations in fine-grained
details. We found distinct brain functional areas and networks that were strongly
related to traits from various categories, such as mental health, physical activity,
cognitive performance, and biomarkers. We developed integrative prediction models
for fluid intelligence, suggesting that integrating fMRI traits with multiple data types can
improve prediction performance for brain-related complex traits and diseases.
4.1 Resting-state and task-evoked functional organizations
The study of how the brain alters its functionality in response to tasks or stimuli is a
topic of significant interest and has broad clinical applications (Zheng et al., 2022). For
instance, fMRI studies involving an emotional task have consistently demonstrated
abnormalities
in the prefrontal cortex-limbic area among patients with anxiety
disorders, who typically exhibit exaggerated responses to emotional stimuli (Li et al.,
2020). Despite relatively small sample sizes, previous studies have found that intrinsic
and extrinsic functional architectures share substantial similarities, with minor but
consistent differences observed across various tasks (Cole et al., 2014; Cole et al., 2021;
Gonzalez-Castillo & Bandettini, 2018; Gratton et al., 2016; Gratton et al., 2018; Smith et
al., 2009; Tavor et al., 2016). Leveraging parcellation-based data from the extensive UKB
study, we corroborate that group-level intrinsic and extrinsic functional spatial patterns
are largely alike (correlation = 0.754), consistent with previous fMRI datasets with
smaller sample sizes (Cole et al., 2014; Cole et al., 2021; Gonzalez-Castillo & Bandettini,
2018; Gratton et al., 2016; Gratton et al., 2018; Tavor et al., 2016). Moreover, we
provide a more detailed analysis of resting-state functional connectivity differences. For
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
30
example, our results described the complicated task-positive and task-negative
functional connectivity change patterns in the default mode network. Although the
default mode network has been originally recognized as brain areas with greater
connectivity in resting fMRI than task fMRI (Raichle et al., 2001), recent studies have
found that the default mode network also had positive functional contributions to tasks,
which may result in increased activity in task fMRI (Elton & Gao, 2015).
Furthermore, our results demonstrate a remarkable spatial correlation between
the UKB and HCP studies in both resting and task fMRI. This high degree of consistency
across independent studies underscores the possibility of innovative joint analyses of
human connectome data. Through meta-analytic amalgamation of these fMRI datasets,
we have the potential to gain a more profound understanding of trait-fMRI associations'
replication and enhance fMRI's predictive power for a variety of phenotypes (He et al.,
2022). The integration of data from multiple sources may lead to more robust and
reliable outcomes in the field of fMRI research.
4.2 Sex difference in fMRI
Our area- and network-specific sex effect maps can be useful for understanding sex
differences in brain functional activity, as well as brain function-related cognitive
impairment and brain disorders. We found that the strongest sex difference in resting
fMRI was in the somatomotor network, where females had weaker functional
connectivity than males (Fig. 3C). Additionally, depression was strongly associated with
decreased connectivity in the somatomotor network (Fig. 4B). Considering the fact that
depression is two times more prevalent in females than in males (Salk et al., 2017), our
results may help understand the brain function-related sex differences in depression
(Labaka et al., 2018). In addition, we found that a wide variety of complex traits were
strongly associated with the functional connectivity between the visual and
somatomotor networks, such as risk-taking and time spent watching TV (Figs. 4A and
5B). Future studies could investigate the biological mechanisms underlying these
functional connectivity alterations as well as causal medication pathways among
lifestyle, brain function, and mental health (Zhao & Castellanos, 2016).
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
31
Additionally, our findings
indicate that males demonstrated stronger task
functional connectivity than females in numerous areas within the language network
(Refer to Fig. S13D). This could potentially be attributable to males' more frequent use
of language strategies, such as silent naming during the Hariri’s faces/shapes emotion
task. On the other hand, females might rely more heavily on visual or spatial strategies.
This observation calls for further investigation.
4.3 Trait-fMRI associations
We conducted an analysis of fMRI data alongside a range of complex traits using a
discovery-validation design, generating association maps that correspond to the
functional organization of the human brain during both resting and task states. These
results may contribute to the development of improved disease prediction models and
the
identification of clinically beneficial neuroimaging biomarkers. For
instance,
depression and depressive mood disorders have been associated with abnormal brain
connectivity across several intrinsic networks (Brakowski et al., 2017; Gudayol-Ferré et
al., 2015; Korgaonkar et al., 2019). Our findings spotlight specific patterns of decreased
resting functional connectivity, particularly within the somatomotor network. Extended
periods of TV viewing have been linked to structural variations in the visual cortex and
sensorimotor areas (Takeuchi et al., 2013). This activity has also been associated with
cognitive decline (Fancourt & Steptoe, 2019) and increased dementia risk (Raichlen et
al., 2022)—both closely connected with the default mode network (Grieder et al., 2018).
Moreover, visual impairment and diminished functional connectivity within the visual
network have been identified in Alzheimer’s disease (Huang et al., 2021; Littlejohns et
al., 2022). Our results suggest that resting fMRI traits of the default mode and visual
networks could serve as valuable endophenotypes for investigating the effects of
environmental and lifestyle factors on aging and dementia.
The large-scale UKB data also revealed that resting and task fMRI may have
different association patterns with complex traits, such as mental health and cognitive
abilities. For example, depression was strongly associated with resting fMRI, but not
with task fMRI. Moreover, in resting and task fMRI, the associations with fluid
intelligence had different spatial distributions. Our prediction analysis further suggests
that task fMRI has additional predictive power on intelligence on top of resting fMRI.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
32
These results demonstrate the differences between resting and task-evoked brain
functions in terms of their connections with brain health and cognition.
4.4 Online resource and future development
Using the large-scale fMRI data in the UKB study, we were able to study hundreds
of brain regions in a parcellation-based approach. We have utilized the rich phenotypic
data in the UKB database in our fMRI-trait association analysis, which was an
exploratory analysis designed to offer a publicly accessible web interface. The
bioinformatics resource we have developed offers significant potential for fMRI
researchers in various ways. Firstly, it allows for swift comparisons between our findings
and those of existing studies within the field. Researchers can easily evaluate the
congruencies or disparities in trait-fMRI associations when utilizing data from distinct
studies or when identical data are analyzed by different research groups and
methodologies (Botvinik-Nezer et al., 2020). Furthermore, our results can offer
corroborating evidence and preliminary data for future study designs and grant
proposals. Researchers can harness our findings to justify the necessity for additional
data collection and the development of advanced techniques. Additionally, our resource
has the potential to unearth further insights in subsequent studies through the
incorporation of other fMRI data resources. For instance, conducting joint analyses with
other large-scale neuroimaging studies, such as the ABCD (Chaarani et al., 2021) and
CHIMGEN (Xu et al., 2020) studies, could support the replication of association findings
and provide insights into age-related or cohort-related interactions throughout the
lifespan. In conclusion, the online resource we have developed offers a wealth of
opportunities for fMRI researchers to gain insights, compare results, support the design
of future studies, and integrate with other data sources. This integration fosters an
enhanced understanding and collaboration within the field.
The ongoing UKB imaging study, which aims to scan 100,000 subjects within a few
years (Littlejohns et al., 2020), presents an opportunity for us to continuously update
and augment our online resource. This will involve not only replicating our reported
findings based on the Glasser360 and Schaefer (Schaefer et al., 2018) atlases, but also
integrating additional common parcellation schemes such as the Gordon (Gordon et al.,
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
33
2016), Power (Power et al., 2011), DiFuMo (Dadi et al., 2020), and data-driven ICA
(Alfaro-Almagro et al., 2018; Beckmann & Smith, 2004; Hyvarinen, 1999) atlases.
Moreover, we plan to explore and incorporate different data preprocessing pipelines to
understand their effects on the results. For example, we will examine the effects of
topographical misalignments on trait-fMRI associations and sex differences. There has
been an observation in the HCP study that the cross-subject variability can be explained
by the misalignment in topography between individual subjects' true connectivity
topography and group-average ICA maps used by the ICA dual regression (Bijsterbosch
et al., 2019; Bijsterbosch et al., 2018). This residual functional misalignment can mean
that between-subject spatial variability appears as variability in network connectivity;
the extent of this problem of misinterpretation may vary across different analysis
methods (e.g., group-ICA with dual-regression vs hard parcellation). It would be
interesting to quantify the effects of spatial misalignment on both parcellation-based
and whole-brain ICA-based fMRI traits in the large-scale UKB dataset.
In addition, our main analyses were based on parcellation-based full correlations.
Although the FMRIB's ICA-based X-noiseifier (FIX) has been applied to the UKB dataset
to remove scanner artifacts and motion effects, full correlation measures can be more
sensitive to the remaining global artifacts and noises than partial correlations (Feis et al.,
2015; Griffanti et al., 2014). It is possible to further remove global artifacts by measuring
the partial functional connectivity between paired brain regions after removing the
dependency of other brain regions (Elliott et al., 2018). Future studies need to explore
parcellation-based partial correlation traits for a large number of parcels (such as the
360 regions in the Glasser360 atlas) with a limited number of time points in the UKB
study. Finally, we welcome user feedback and suggestions, which will help improve our
project and resources to better meet the needs of the fMRI research community.
Declaration of Competing Interest
The authors declare no competing financial interests.
Credit authorship contribution statement
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
34
B.Z., H.Z., and S.M.S designed the study. B.Z., T.L., Z.F., D.X., X.W., and M.G. processed
and analyzed the data. Y.L. and B.Z. designed the website and developed online
resources. B.Z. wrote the manuscript with feedback from all authors.
Data and code availability statement
Our results and summary-level data can be downloaded and browsed at
http://fmriatlas.org/. The individual-level UK Biobank data can be obtained from
https://www.ukbiobank.ac.uk/. Code used
in
this
study
is
available
at
https://zenodo.org/record/8235805.
Ethics statement
This research utilized data from the UKB study and the HCP study. The UKB has obtained
ethics approval from the North West Multi-Centre Research Ethics Committee (MREC,
approval number: 11/NW/0382), and obtained written informed consent from all
participants prior to the study. All experimental procedures in the HCP study were
approved by the institutional review boards at Washington University (approval number
201204036).
Acknowledgments
This research was partially supported by partially supported by MH116527 (T.L. and
H.Z.) and AG082938-01 (H.Z.). We thank the individuals represented in the UKB and HCP
studies for their participation and the research teams for their work in collecting,
processing and disseminating these datasets for analysis. We would like to thank the
University of North Carolina at Chapel Hill and Purdue University and their Research
Computing groups for providing computational resources and support that have
contributed to these research results. This research has been conducted using the UK
Biobank resource (application number 22783), subject to a data transfer agreement.
REFERENCES
Agosta, F., Pievani, M., Geroldi, C., Copetti, M., Frisoni, G. B., & Filippi, M. (2012).
Resting state fMRI in Alzheimer's disease: beyond the default mode network.
Neurobiology of aging, 33(8), 1564-1578.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
35
Alfaro-Almagro, F., Jenkinson, M., Bangerter, N. K., Andersson, J. L., Griffanti, L.,
Douaud, G., Sotiropoulos, S. N., Jbabdi, S., Hernandez-Fernandez, M., & Vallee, E.
(2018). Image processing and Quality Control for the first 10,000 brain imaging
datasets from UK Biobank. Neuroimage, 166, 400-424.
Alonso, A., & de Larriva, A. P. A. (2016). Atrial fibrillation, cognitive decline and
dementia. European Cardiology Review, 11(1), 49.
Beckmann, C. F., & Smith, S. M. (2004). Probabilistic independent component analysis
for functional magnetic resonance imaging. IEEE transactions on medical
imaging, 23(2), 137-152.
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery rate: a practical and
powerful approach to multiple testing. Journal of the royal statistical society.
Series B (Methodological), 289-300.
Bijsterbosch, J., Harrison, S., Duff, E., Alfaro-Almagro, F., Woolrich, M., & Smith, S.
(2017). Investigations into within-and between-subject resting-state amplitude
variations. Neuroimage, 159, 57-69.
Bijsterbosch, J. D., Beckmann, C. F., Woolrich, M. W., Smith, S. M., & Harrison, S. J.
(2019). The relationship between spatial configuration and functional
connectivity of brain regions revisited. Elife, 8, e44890.
Bijsterbosch, J. D., Woolrich, M. W., Glasser, M. F., Robinson, E. C., Beckmann, C. F., Van
Essen, D. C., Harrison, S. J., & Smith, S. M. (2018). The relationship between
spatial configuration and functional connectivity of brain regions. Elife, 7,
e32992.
Biswal, B., Zerrin Yetkin, F., Haughton, V. M., & Hyde, J. S. (1995). Functional
connectivity in the motor cortex of resting human brain using echo‐planar MRI.
Magnetic resonance in medicine, 34(4), 537-541.
Botvinik-Nezer, R., Holzmeister, F., Camerer, C. F., Dreber, A., Huber, J., Johannesson,
M., Kirchler, M., Iwanir, R., Mumford, J. A., Adcock, R. A., Avesani, P., Baczkowski,
B. M., Bajracharya, A., Bakst, L., Ball, S., Barilari, M., Bault, N., Beaton, D.,
Beitner, J., . . . Schonberg, T. (2020). Variability in the analysis of a single
neuroimaging dataset by many teams. Nature, 582(7810), 84-88.
https://doi.org/10.1038/s41586-020-2314-9
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
36
Brakowski, J., Spinelli, S., Dörig, N., Bosch, O. G., Manoliu, A., Holtforth, M. G., & Seifritz,
E. (2017). Resting state brain network function in major depression – Depression
symptomatology, antidepressant treatment effects, future research. Journal of
Psychiatric Research, 92, 147-159.
https://doi.org/10.1016/j.jpsychires.2017.04.007
Bycroft, C., Freeman, C., Petkova, D., Band, G., Elliott, L. T., Sharp, K., Motyer, A.,
Vukcevic, D., Delaneau, O., O’Connell, J., Cortes, A., Welsh, S., Young, A.,
Effingham, M., McVean, G., Leslie, S., Allen, N., Donnelly, P., & Marchini, J.
(2018). The UK Biobank resource with deep phenotyping and genomic data.
Nature, 562(7726), 203-209. https://doi.org/10.1038/s41586-018-0579-z
Carnevale, L., Maffei, A., Landolfi, A., Grillea, G., Carnevale, D., & Lembo, G. (2020). Brain
Functional Magnetic Resonance Imaging Highlights Altered Connections and
Functional Networks in Patients With Hypertension. Hypertension,
HYPERTENSIONAHA. 120.15296.
Chaarani, B., Hahn, S., Allgaier, N., Adise, S., Owens, M., Juliano, A., Yuan, D., Loso, H.,
Ivanciu, A., & Albaugh, M. (2021). Baseline brain function in the preadolescents
of the ABCD Study. Nature neuroscience, 1-11.
Cole, Michael W., Bassett, Danielle S., Power, Jonathan D., Braver, Todd S., & Petersen,
Steven E. (2014). Intrinsic and Task-Evoked Network Architectures of the Human
Brain. Neuron, 83(1), 238-251. https://doi.org/10.1016/j.neuron.2014.05.014
Cole, M. W., Ito, T., Cocuzza, C., & Sanchez-Romero, R. (2021). The Functional Relevance
of Task-State Functional Connectivity. The Journal of Neuroscience, 41(12),
2684-2702. https://doi.org/10.1523/jneurosci.1713-20.2021
Dadi, K., Varoquaux, G., Machlouzarides-Shalit, A., Gorgolewski, K. J., Wassermann, D.,
Thirion, B., & Mensch, A. (2020). Fine-grain atlases of functional modes for fMRI
analysis. NeuroImage, 221. https://doi.org/10.1016/j.neuroimage.2020.117126
Dey, R., Zhou, W., Kiiskinen, T., Havulinna, A., Elliott, A., Karjalainen, J., Kurki, M., Qin, A.,
Lee, S., & Palotie, A. (2020). An efficient and accurate frailty model approach for
genome-wide survival association analysis controlling for population structure
and relatedness in large-scale biobanks. bioRxiv.
Dickie, E. W., Anticevic, A., Smith, D. E., Coalson, T. S., Manogaran, M., Calarco, N.,
Viviano, J. D., Glasser, M. F., Van Essen, D. C., & Voineskos, A. N. (2019). Ciftify: A
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
37
framework for surface-based analysis of legacy MR acquisitions. Neuroimage,
197, 818-826.
Elliott, L. T., Sharp, K., Alfaro-Almagro, F., Shi, S., Miller, K. L., Douaud, G., Marchini, J., &
Smith, S. M. (2018). Genome-wide association studies of brain imaging
phenotypes in UK Biobank. Nature, 562(7726), 210-216.
Elliott, M. L., Knodt, A. R., Ireland, D., Morris, M. L., Poulton, R., Ramrakha, S., Sison, M.
L., Moffitt, T. E., Caspi, A., & Hariri, A. R. (2020). What is the test-retest reliability
of common task-functional MRI measures? New empirical evidence and a
meta-analysis. Psychological science, 31(7), 792-806.
Elton, A., & Gao, W. (2015). Task-positive functional connectivity of the default mode
network transcends task domain. Journal of cognitive neuroscience, 27(12),
2369-2381.
Ewing, S. W. F., Sakhardande, A., & Blakemore, S.-J. (2014). The effect of alcohol
consumption on the adolescent brain: A systematic review of MRI and fMRI
studies of alcohol-using youth. NeuroImage: clinical, 5, 420-437.
Fancourt, D., & Steptoe, A. (2019). Television viewing and cognitive decline in older age:
findings from the English Longitudinal Study of Ageing. Sci Rep, 9(1), 2851.
https://doi.org/10.1038/s41598-019-39354-4
Feis, R. A., Smith, S. M., Filippini, N., Douaud, G., Dopper, E. G., Heise, V., Trachtenberg,
A. J., van Swieten, J. C., van Buchem, M. A., & Rombouts, S. A. (2015). ICA-based
artifact removal diminishes scan site differences in multi-center resting-state
fMRI. Frontiers in neuroscience, 9, 395.
Friedman, J., Hastie, T., & Tibshirani, R. (2010). Regularization paths for generalized
linear models via coordinate descent. Journal of statistical software, 33(1), 1.
Gao, S., Greene, A. S., Constable, R. T., & Scheinost, D. (2019). Combining multiple
connectomes improves predictive modeling of phenotypic measures.
Neuroimage, 201, 116038.
Glasser, M. F., Coalson, T. S., Robinson, E. C., Hacker, C. D., Harwell, J., Yacoub, E.,
Ugurbil, K., Andersson, J., Beckmann, C. F., & Jenkinson, M. (2016). A
multi-modal parcellation of human cerebral cortex. Nature, 536(7615), 171-178.
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
38
Gonzalez-Castillo, J., & Bandettini, P. A. (2018). Task-based dynamic functional
connectivity: Recent findings and open questions. NeuroImage, 180, 526-533.
https://doi.org/10.1016/j.neuroimage.2017.08.006
Gordon, E. M., Laumann, T. O., Adeyemo, B., Huckins, J. F., Kelley, W. M., & Petersen, S.
E. (2016). Generation and Evaluation of a Cortical Area Parcellation from
Resting-State Correlations. Cerebral Cortex, 26(1), 288-303.
https://doi.org/10.1093/cercor/bhu239
Gratton, C., Laumann, Timothy O., Gordon, Evan M., Adeyemo, B., & Petersen, Steven E.
(2016). Evidence for Two Independent Factors that Modify Brain Networks to
Meet Task Goals. Cell Reports, 17(5), 1276-1288.
https://doi.org/10.1016/j.celrep.2016.10.002
Gratton, C., Laumann, T. O., Nielsen, A. N., Greene, D. J., Gordon, E. M., Gilmore, A. W.,
Nelson, S. M., Coalson, R. S., Snyder, A. Z., Schlaggar, B. L., Dosenbach, N. U. F., &
Petersen, S. E. (2018). Functional Brain Networks Are Dominated by Stable
Group and Individual Factors, Not Cognitive or Daily Variation. Neuron, 98(2),
439-452.e435. https://doi.org/10.1016/j.neuron.2018.03.035
Grieder, M., Wang, D. J. J., Dierks, T., Wahlund, L. O., & Jann, K. (2018). Default Mode
Network Complexity and Cognitive Decline in Mild Alzheimer's Disease. Front
Neurosci, 12, 770. https://doi.org/10.3389/fnins.2018.00770
Griffanti, L., Salimi-Khorshidi, G., Beckmann, C. F., Auerbach, E. J., Douaud, G., Sexton, C.
E., Zsoldos, E., Ebmeier, K. P., Filippini, N., & Mackay, C. E. (2014). ICA-based
artefact removal and accelerated fMRI acquisition for improved resting state
network imaging. Neuroimage, 95, 232-247.
Gudayol-Ferré, E., Peró-Cebollero, M., González-Garrido, A. A., & Guàrdia-Olmos, J.
(2015). Changes in brain connectivity related to the treatment of depression
measured through fMRI: a systematic review. Frontiers in Human Neuroscience,
9. https://doi.org/10.3389/fnhum.2015.00582
He, T., An, L., Chen, P., Chen, J., Feng, J., Bzdok, D., Holmes, A. J., Eickhoff, S. B., & Yeo, B.
T. T. (2022). Meta-matching as a simple framework to translate phenotypic
predictive models from big to small data. Nature Neuroscience, 25(6), 795-804.
https://doi.org/10.1038/s41593-022-01059-9
Downloaded from http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 September 2023
39
He, T., Kong, R., Holmes, A. J., Nguyen, M., Sabuncu, M. R., Eickhoff, S. B., Bzdok, D.,
Feng, J., & Yeo, B. T. (2020). Deep neural networks and kernel regression achieve
comparable accuracies for functional connectivity prediction of behavior and
demographics. Neuroimage, 206, 116276.
Herting, M. M., Gautam, P., Chen, Z., Mezher, A., & Vetter, N. C. (2018). Test-retest
reliability of longitudinal task-based fMRI: Implications for developmental
studies. Developmental cognitive neuroscience, 33, 17-26.
Hu, M.-L., Zong, X.-F., Mann, J. J., Zheng, J.-J., Liao, Y.-H., Li, Z.-C., He, Y., Chen, X.-G., &
Tang, J.-S. (2017). A review of the functional and anatomical default mode
network in schizophrenia. Neuroscience bulletin, 33(1), 73-84.
Huang, J., Beach, P., Bozoki, A., & Zhu, D. C. (2021). Alzheimer's Disease Progressively
Reduces Visual Functional Network Connectivity. J Alzheimers Dis Rep, 5(1),
549-562. https://doi.org/10.3233/ADR-210017
Hyvarinen, A. (1999). Fast and robust fixed-point algorithms for independent
component analysis. IEEE transactions on Neural Networks, 10(3), 626-634.
Ji, J. L., Spronk, M., Kulkarni, K., Repovš, G., Anticevic, A., & Cole, M. W. (2019). Mapping
the human brain's cortical-subcortical functional network organization.
Neuroimage, 185, 35-57.
Jiang, L., Zheng, Z., Qi, T., Kemper, K., Wray, N., Visscher, P., & Yang, J. (2019). A
resource-efficient tool for mixed model association analysis of large-scale data.
Nature genetics, 51(12), 1749.
Jiang, R., Calhoun, V. D., Noble, S., Sui, J., Liang, Q., Qi, S., & Scheinost, D. (2023). A
functional connectome signature of blood pressure in> 30 000 participants from
the UK biobank. Cardiovascular Research, 119(6), 1427-1440.
Kennedy, j. T., Harms, METRO. PAG., Korucuoglu, o., Astafiev, S. v., Respeto, D. METRO., Thompson, W..
K., Bjork, j. METRO., & Anokhin, A. PAG. (2021). Reliability and Stability Challenges in
ABCD Task fMRI Data. bioRxiv.
Keyes, k. METRO., Platón, J., Kaufman, A. S., & McLaughlin, k. A. (2017). Association of fluid
intelligence and psychiatric disorders in a population-representative sample of
US adolescents. JAMA psychiatry, 74(2), 179-188.
Korgaonkar, METRO. S., Goldstein-Piekarski, A. NORTE., Proporcionó, A., & williams, l. METRO. (2019).
Intrinsic connectomes are a predictive biomarker of remission in major
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
40
depressive disorder. Molecular Psychiatry, 25(7), 1537-1549.
https://doi.org/10.1038/s41380-019-0574-2
Labaka, A., Goñi-Balentziaga, o., Lebeña, A., & Pérez-Tejada, j. (2018). Biological sex
differences in depression: a systematic review. Biological research for nursing,
20(4), 383-392.
li, J., Zhong, y., Mamá, Z., Wu, y., Angustia, METRO., Wang, C., Liu, NORTE., Wang, C., & zhang, norte. (2020).
Emotion reactivity-related brain network analysis in generalized anxiety disorder:
a task fMRI study. BMC Psychiatry, 20(1).
https://doi.org/10.1186/s12888-020-02831-6
Littlejohns, t. J., Hayat, S., Luben, r., Brayne, C., Conroy, METRO., Foster, PAG. J., Khawaja, A. PAG.,
& Kuzma, mi. (2022). Visual Impairment and Risk of Dementia in 2
Population-Based Prospective Cohorts: UK Biobank and EPIC-Norfolk. J Gerontol
A Biol Sci Med Sci, 77(4), 697-704. https://doi.org/10.1093/gerona/glab325
Littlejohns, t. J., Holliday, J., Gibson, l. METRO., Garratt, S., Oesingmann, NORTE., Alfaro-Almagro,
F., Campana, j. D., Boultwood, C., collins, r., & Conroy, METRO. C. (2020). The UK Biobank
imaging enhancement of 100,000 Participantes: rationale, data collection,
management and future directions. Nature communications, 11(1), 1-12.
Macpherson, h., Formica, METRO., harris, MI., & Daly, R. METRO. (2017). Brain functional
alterations in Type 2 Diabetes–A systematic review of fMRI studies. Fronteras en
neuroendocrinology, 47, 34-46.
Mak, t. S. h., Porsch, R. METRO., Choi, S. w., zhou, X., & Sham, PAG. C. (2017). Polygenic scores
via penalized regression on summary statistics. Genetic epidemiology, 41(6),
469-480.
Marek, S., Tervo-Clemmens, B., Calabro, F. J., Montez, D. F., kay, B. PAG., Hatoum, A. S.,
Donohue, METRO. r., Foran, w., Molinero, R. l., & Hendrickson, t. j. (2022).
Reproducible brain-wide association studies require thousands of individuals.
Naturaleza, 603(7902), 654-660.
Molinero, k. l., Alfaro-Almagro, F., Bangerter, norte. K., tomás, D. l., Yacoub, MI., Xu, J.,
Bartsch, A. J., Jbabdi, S., Sotiropoulos, S. NORTE., & andersson, j. l. (2016).
Multimodal population brain imaging in the UK Biobank prospective
epidemiological study. Nature neuroscience, 19(11), 1523-1536.
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
41
Mulders, PAG. C., van Eijndhoven, PAG. F., Schene, A. h., beckman, C. F., & Tendolkar, I.
(2015). Resting-state functional connectivity in major depressive disorder: a
revisar. Neurociencia & Revisiones de biocomportamiento, 56, 330-344.
Naumczyk, PAG., Sabisz, A., Witkowska, METRO., Graff, B., Jodzio, K., Gąsecki, D., Szurowska, MI.,
& Narkiewicz, k. (2017). Compensatory functional reorganization may precede
hypertension-related brain damage and cognitive decline: a functional magnetic
resonance imaging study. Journal of hypertension, 35(6), 1252.
Noble, S., Scheinost, D., & Constable, R. t. (2021). A guide to the measurement and
interpretation of fMRI test-retest reliability. Opinión actual en comportamiento
Ciencias, 40, 27-32.
Ogawa, S., Sotavento, T.-M., kay, A. r., & Tank, D. W.. (1990). Brain magnetic resonance
imaging with contrast dependent on blood oxygenation. Actas de la
Academia Nacional de Ciencias, 87(24), 9868-9872.
Pervaiz, Ud., Vidaurre, D., lana rica, METRO. w., & Herrero, S. METRO. (2020). Optimising network
modelling methods for fMRI. Neuroimagen, 211, 116604.
Popovych, oh. v., Jung, K., Manos, T., Diaz-Pier, S., Hoffstaedter, F., Schreiber, J., yo, B.
T., & Eickhoff, S. B. (2021). Inter-subject and inter-parcellation variability of
resting-state whole-brain dynamical modeling. Neuroimagen, 118201.
posner, J., Parque, C., & Wang, z. (2014). Connecting the dots: a review of resting
connectivity MRI studies in attention-deficit/hyperactivity disorder.
Neuropsychology review, 24(1), 3-15.
Fuerza, j. D., cohen, A. l., nelson, S. METRO., Peluca, GRAMO. S., Barnes, k. A., Church, j. A., Vogel, A.
C., Laumann, t. o., Miezín, F. METRO., & Schlaggar, B. l. (2011). Functional network
organization of the human brain. Neurona, 72(4), 665-678.
Raichle, METRO. MI., MacLeod, A. METRO., Snyder, A. Z., Powers, W.. J., Gusnard, D. A., & Shulman,
GRAMO. l. (2001). A default mode of brain function. Actas del Nacional
Academia de Ciencias, 98(2), 676-682.
Raichlen, D. A., Klimentidis, Y. C., Sayre, METRO. K., Bharadwaj, PAG. K., Dejar, METRO. h. C., Wilcox, R.
r., & Alexander, GRAMO. mi. (2022). Leisure-time sedentary behaviors are differentially
associated with all-cause dementia regardless of engagement in physical activity.
Proc Natl Acad Sci U S A, 119(35), e2206931119.
https://doi.org/10.1073/pnas.2206931119
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
42
Ritchie, S. J., Cox, S. r., shen, X., Lombardo, METRO. v., Reus, l. METRO., Alloza, C., harris, METRO. A.,
Alderson, h. l., Cazador, S., Neilson, MI., Liewald, D. C. METRO., Auyeung, B., Whalley, h.
C., Lawrie, S. METRO., Gale, C. r., Bastin, METRO. MI., McIntosh, A. METRO., & Deary, I. j. (2018).
Sex Differences in the Adult Human Brain: Evidencia de 5216 UK Biobank
Participantes. Corteza cerebral, 28(8), 2959-2975.
https://doi.org/10.1093/cercor/bhy109
Rolls, mi. T., Wan, Z., cheng, w., & feng, j. (2022). Risk-taking in humans and the medial
orbitofrontal cortex reward system. Neuroimagen, 118893.
Salk, R. h., Hyde, j. S., & Abramson, l. Y. (2017). Gender differences in depression in
representative national samples: Meta-analyses of diagnoses and symptoms.
Psychological bulletin, 143(8), 783.
Schaefer, A., kong, r., gordon, mi. METRO., Laumann, t. o., Zuo, X.-N., holmes, A. J., Eickhoff,
S. B., & yo, B. t. (2018). Local-global parcellation of the human cerebral cortex
from intrinsic functional connectivity MRI. Corteza cerebral, 28(9), 3095-3114.
Scheinost, D., Finn, mi. S., Tokoglu, F., shen, X., Papademetris, X., Hampson, METRO., &
Constable, R. t. (2015). Sex differences in normal age trajectories of functional
redes cerebrales. Mapeo del cerebro humano, 36(4), 1524-1535.
https://doi.org/10.1002/hbm.22720
shen, l., & Thompson, PAG. METRO. (2019). Brain imaging genomics: integrated analysis and
aprendizaje automático. Actas del IEEE, 108(1), 125-162.
Herrero, S. METRO., eliot, l. T., Alfaro-Almagro, F., McCarthy, PAG., Nichols, t. MI., Douaud, GRAMO., &
Molinero, k. l. (2020). Brain aging comprises many modes of structural and
functional change with distinct genetic and biophysical associations. Elife, 9,
e52677.
Herrero, S. METRO., Fox, PAG. T., Molinero, k. l., Glahn, D. C., Fox, PAG. METRO., Mackay, C. MI., Filippini, NORTE.,
watkins, k. MI., Toro, r., Laird, A. r., & beckman, C. F. (2009). Correspondence of
the brain’s functional architecture during activation and rest. Actas de la
Academia Nacional de Ciencias, 106(31), 13040-13045.
https://doi.org/10.1073/pnas.0905267106
Herrero, S. METRO., & Nichols, t. mi. (2018). Statistical challenges in “big data” human
neuroimaging. Neurona, 97(2), 263-268.
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
43
Takeuchi, h., Taki, y., Hashizume, h., Asano, K., Asano, METRO., Sassa, y., Yokota, S., Kotozaki,
y., Nouchi, r., & Kawashima, R. (2013). The Impact of Television Viewing on Brain
Structures: Cross-Sectional and Longitudinal Analyses. Corteza cerebral, 25(5),
1188-1197. https://doi.org/10.1093/cercor/bht315
Tavor, I., jones, oh. PAG., Mars, R. B., Herrero, S. METRO., Behrens, t. MI., & Jbabdi, S. (2016).
Task-free MRI predicts individual differences in brain activity during task
actuación. Ciencia, 352(6282), 216-220.
https://doi.org/10.1126/science.aad8127
VanEssen, D. C., Herrero, S. METRO., Respeto, D. METRO., Behrens, t. MI., Yacoub, MI., Ugurbil, K., &
Consorcio, W.-M. h. (2013). The WU-Minn human connectome project: un
descripción general. Neuroimagen, 80, 62-79.
Xu, P., guo, l., cheng, J., Wang, METRO., Geng, Z., Zhu, w., zhang, B., Liao, w., Qiu, S., &
zhang, h. (2020). CHIMGEN: a Chinese imaging genetics cohort to enhance
cross-ethnic and cross-geographic brain research. Molecular psychiatry, 25(3),
517-529.
zhao, B., li, T., Herrero, S. METRO., xiong, D., Wang, X., Cual, y., luo, T., Zhu, Z., shan, y., &
Matoba, norte. (2022). Common variants contribute to intrinsic human brain
functional networks. Nature Genetics in press
zhao, B., li, T., Cual, y., Wang, X., luo, T., shan, y., Zhu, Z., xiong, D., Hauberg, METRO. MI., &
Bendl, j. (2021). Common genetic variation influencing human white matter
microstructure. Ciencia, 372(6548), eabf3736.
zhao, B., luo, T., li, T., li, y., zhang, J., shan, y., Wang, X., Cual, l., zhou, F., & Zhu, z.
(2019). Genome-wide association analysis of 19,629 individuals identifies
variants influencing regional brain volumes and refines their genetic
co-architecture with cognitive and mental health traits. Nature genetics, 51(11),
1637-1644.
zhao, J., li, METRO., zhang, y., Song, h., von Deneen, k. METRO., Shi, y., Liu, y., & Él, D. (2017).
Intrinsic brain subsystem associated with dietary restraint, disinhibition and
hunger: an fMRI study. Brain imaging and behavior, 11(1), 264.
zhao, y., & Castellanos, F. X. (2016). Annual research review: discovery science
strategies in studies of the pathophysiology of child and adolescent psychiatric
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
44
disorders‐promises and limitations. Journal of Child Psychology and Psychiatry,
57(3), 421-439.
Zheng, Y.-Q., Farahibozorg, S.-R., Gong, w., Rafipoor, h., Jbabdi, S., & Herrero, S. (2022).
Accurate predictions of individual differences in task-evoked brain activity from
resting-state fMRI using a sparse ensemble learner. NeuroImagen, 259.
https://doi.org/10.1016/j.neuroimage.2022.119418
Zou, Q.-H., Zhu, C.-Z., Cual, y., Zuo, X.-N., Largo, X.-Y., Cao, Q.-J., Wang, Y.-F., & Zang,
Y.-F. (2008). An improved approach to detection of amplitude of low-frequency
fluctuation (ALFF) for resting-state fMRI: fractional ALFF. Revista de neurociencia
Métodos, 172(1), 137-141.
Descargado de http://direct.mit.edu/imag/article-pdf/doi/10.1162/imag_a_00015/2154903/imag_a_00015.pdf by guest on 07 Septiembre 2023
45