Tracking the Emergence of Location-based Spatial
Representations in Human Scene-Selective Cortex
Sam C. Berens1,2 , Bárður H. Joensen1,3,4, and Aidan J. Horner1,5
抽象的
■ Scene-selective regions of the human brain form allocentric
representations of locations in our environment. These represen-
tations are independent of heading direction and allow us to know
where we are regardless of our direction of travel. 然而, 我们
know little about how these location-based representations are
形成的. Using fMRI representational similarity analysis and linear
mixed models, we tracked the emergence of location-based repre-
sentations in scene-selective brain regions. We estimated patterns
of activity for two distinct scenes, taken before and after partici-
pants learnt they were from the same location. During a learning
phase, we presented participants with two types of panoramic
视频: (1) an overlap video condition displaying two distinct
场景 (0° and 180°) from the same location and (2) a no-overlap
video displaying two distinct scenes from different locations (哪个
served as a control condition). In the parahippocampal cortex
(PHC) and retrosplenial cortex (RSC), representations of scenes
from the same location became more similar to each other only
after they had been shown in the overlap condition, suggesting
the emergence of viewpoint-independent location-based repre-
句子. Whereas these representations emerged in the PHC
regardless of task performance, RSC representations only emerged
for locations where participants could behaviorally identify the two
scenes as belonging to the same location. The results suggest that
we can track the emergence of location-based representations in
the PHC and RSC in a single fMRI experiment. 更远, 他们
support computational models that propose the RSC plays a key
role in transforming viewpoint-independent representations into
behaviorally relevant representations of specific viewpoints. ■
介绍
Rapidly learning the spatial layout of a new environment
is a critical function that supports flexible navigation.
This ability is thought to depend on the emergence of
location-based representations in scene-selective brain
regions that signal where we are irrespective of our cur-
rent heading direction. As we are unable to sample all
possible viewpoints from a given location simultaneously,
the formation of location-based representations requires
the integration of scenes from differing viewpoints.
Despite evidence for the existence of location-based rep-
resentations in scene-selective regions (例如, Marchette,
Vass, Ryan, & 爱泼斯坦, 2015; Vass & 爱泼斯坦, 2013), 我们
know little about how such representations emerge.
Models of spatial navigation suggest that distinct brain
regions are responsible for supporting allocentric (viewpoint-
独立的) and egocentric (viewpoint-dependent) 代表-
sentations of our environment (Julian, Keinath, Marchette,
& 爱泼斯坦, 2018; Byrne, Becker, & 伯吉斯, 2007). 具体来说,
the parahippocampal cortex (PHC) and hippocampus
are thought to encode allocentric spatial representations
related to navigational landmarks/boundaries (爱泼斯坦,
Patai, Julian, & Spiers, 2017; 伯吉斯, Becker, 国王, &
奥基夫, 2001) and spatial context more broadly (爱泼斯坦
1约克大学, 2University of Sussex, 3UCL Institute of
Cognitive Neuroscience, 4UCL Institute of Neurology, 5约克
Biomedical Research Institute
& Vass, 2014). The hippocampus also supports a wider
variety of spatial and nonspatial associative/configural
functions in the service of memory and navigation (例如,
Henson & Gagnepain, 2010; Hannula & Ranganath, 2009;
Kumaran et al., 2007; 奥基夫 & 伯吉斯, 2005; Eichenbaum,
2004). 这里, we focus on the PHC given its more specific
role in spatial allocentric processing relative to the hippo-
campus. 相比之下, the parietal lobe is thought to support
egocentric representations of specific viewpoints that
underpin route planning (Calton & Taube, 2009; Byrne
等人。, 2007). To enable efficient route planning, a transfor-
mation between allocentric and egocentric representations
is thought to occur in the retrosplenial cortex (RSC), cueing
allocentric representations from egocentric inputs and vice
versa (Bicanski & 伯吉斯, 2018; 伯恩等人。, 2007).
In support of these models, human fMRI studies using
representational similarity analyses (RSA) have found
evidence for viewpoint-independent representations of
specific locations (henceforth referred to as “location-based
representations”) in a network of brain regions including the
PHC and RSC (Marchette, Vass, Ryan, & 爱泼斯坦, 2014;
Vass & 爱泼斯坦, 2013). 最近, panoramic videos
have been used to experimentally induce the formation
of location-based representations (罗伯逊, Hermann,
Mynick, Kravitz, & Kanwisher, 2016). Assessing pattern
similarity for distinct scenes taken from the same location,
Robertson et al. provided evidence for greater pattern sim-
ilarity in the RSC and occipital place area (OPA) 后
© 2020 麻省理工学院. Published under a
Creative Commons Attribution 4.0 国际的 (抄送 4.0) 执照.
认知神经科学杂志 33:3, PP. 445–462
https://doi.org/10.1162/jocn_a_01654
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
哦
C
n
_
A
_
0
1
6
5
4
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
participants had seen a panoramic video showing that two
scenes were from the same location. This effect was not ev-
ident when participants could not learn that two scenes were
from the same location. 有趣的是, they also provided ev-
idence for an effect in the PHC that occurred inboth video
conditions—that is, regardless of whether participants could
learn the scenes were from the same location—suggesting a
more general associative role for the PHC.
Despite these results, we still know little about (1) 如何
quickly such representations are formed, (2) what types
of spatial information they encode, 和 (3) under what
conditions they are evoked. 第一的, it remains unclear
whether location-based representations emerge rapidly
after short exposures to a new environment or whether
they only develop after prolonged experience. 罗伯逊
等人. had participants watch videos outside the scanner,
over the course of 2 天, before assessing pattern simi-
larity inside the scanner. To test whether location-based
representations can form rapidly, we developed a proto-
col that permitted us to scan participants before and after
a short learning phase, allowing us to estimate changes in
pattern similarity as a function of learning in a single fMRI
实验. 第二, without tracking the formation of
location-based representations, it is difficult to determine
exactly what type of information they are representing.
例如, shared representations across viewpoints
may relate to long-term semantic knowledge that is in-
voked when seeing different views of a well-known loca-
的 (see Marchette, Ryan, & 爱泼斯坦, 2017). 相比之下,
rapidly learning representations that are shared across
different viewpoints of a new environment implies that
the information being encoded is more likely to be spa-
tial rather than semantic in nature.
第三, we do not know whether location-based repre-
sentations are involuntary retrieved during visual pro-
cessing. Computational models of spatial navigation
predict that allocentric representations are automatically
activated and updated by egocentric viewpoints (Bicanski
& 伯吉斯, 2018; 伯恩等人。, 2007). 此外, elec-
trophysiological studies in rodents have shown that allo-
centric representations are automatically activated and
updated during exploration (例如, 摩纳哥, 饶, Roth, &
Knierim, 2014; 奥基夫 & Dostrovsky, 1971). 然而,
evidence in humans is lacking. Robertson et al. 必需的
participants to recall whether scenes were presented on
the left or right of the screen, introducing a task that
explicitly required them to recall the panorama, 和
the position of the specific scene within the panorama.
Suggesting some level of involuntary retrieval, one fMRI
study found that viewpoint-independent representations
of specific buildings may be activated when participants
judge whether the building is well known to them
(Marchette et al., 2014). In the current study, 参与者
performed an unrelated low-level attentional task as the
scenes were presented. The activation of location-based
representations under these conditions would suggest that
they can be retrieved in a relatively automatic manner.
这里, we test whether location-based representations of
novel environments can be learnt by integrating visual in-
formation across different scenes. Although location-based
representations are predicted by models of spatial navigation,
they may also be consistent with various other cognitive
型号 (see Discussion). 像这样, we define location-
based representations to be any type of information that
encodes the relationship between different, nonoverlap-
ping views of the same location. We recorded patterns of
BOLD activity as participants passively observed a number
of scenes depicting different views of novel locations.
随后, using an experimental manipulation intro-
duced by Robertson et al. (2016), participants watched
videos showing these scenes as part of a wider panorama.
Half of the videos allowed participants to learn the spatial
relationship between two scenes from the same location
(overlap condition). The remaining videos acted as a
control by presenting scenes from different locations
(no-overlap condition). After the videos, we again recorded
patterns of activity for each of the scenes. Whereas Robertson
等人. (2016) only assessed scene representations after
video presentation, we also scanned before and during
the videos; see Clarke, Pell, Ranganath, and Tyler (2016)
for a similar preexperimental versus postexperimental
design focused on changes in object representations. 这
allowed us to track the potential emergence of location-
based representations using RSAs as well as assess neural
activity when these representations were being formed.
Using generalized linear mixed models, we show that
patterns evoked by different scenes become more similar
in scene-selective regions of the PHC and RSC after the
presentation of the video panoramas. This increase in sim-
ilarity was specific to the “overlap” video condition, 在哪里
scenes from the same location were presented together,
and was not observed in the no-overlap condition. 这
suggests the emergence of location-based representations
in the PHC and RSC. 重要的, whereas this increase in
pattern similarity emerged in the PHC regardless of behav-
ioral performance, the same pattern was only present in
the RSC when participants could remember which scenes
came from the same location. This finding supports
computational models that propose the RSC is critical in
translating viewpoint-independent representations in the
medial temporal lobe into more behaviorally relevant
egocentric representations.
方法
参加者
Twenty-eight right-handed participants were recruited
from the University of York, 英国. These partic-
ipants had no prior familiarity with the locations used as
stimuli in the experiment (见下文). All participants gave
written informed consent and were reimbursed for their
时间. Participants had either normal or corrected-to-
normal vision and reported no history of neurological or
446
认知神经科学杂志
体积 33, 数字 3
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
哦
C
n
_
A
_
0
1
6
5
4
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
psychiatric illness. Data from five participants could not be
included in the final sample because of problems with fMRI
data acquisition (one participant), excess of motion-related
artifacts in the imaging data (three participants), and a
failure to respond during one of the in-scanner tasks
(one participant). 像这样, analyses included 23 partici-
pants (10 男人) with a mean age of 21.96 年 (标准差=
3.22 年). The study was approved by a local research
ethics committee at the University of York.
Stimuli
We generated 12 panoramic images of different urban
locations from the City of Sunderland, and Middlesbrough
town center, 英国 (数字 1; osf.io/cgy97).
These panoramas spanned a 210° field-of-view horizontally
but were restricted in the vertical direction to limit the
appearance of proximal features (<2 m from the camera).
Throughout experiment, 24 “endpoint images” display-
ing 30° scenes taken either end of each panorama
were shown (i.e., centered at 0° and 180°; Figure 1A).
These images were both inside outside the
scanner to assess participants’ spatial knowledge the
depicted locations for RSA (see below).
Endpoints also in a series videos (see
osf.io> 1° and translations > 1 毫米), and a
Fourier basis set implementing a 1/128-Hz high-pass
filter. For the analyses of univariate BOLD activations, EPI
data were warped to Montreal Neurological Institute space
with transformation parameters derived from structural
scans (using the DARTEL toolbox; Ashburner, 2007).
随后, the EPI data were spatially smoothed with
an isotropic 8-mm FWHM Gaussian kernel before GLM
分析 (regressors included the same nuisance effects
noted above).
ROIs
We generated four binary masks per participant to repre-
sent each ROI in native space. 要做到这一点, a first-level GLM
of the functional localizer data modeled BOLD responses to
scene and face stimuli presented during the localizer task.
Each ROI was then defined as the conjunction between a
“scene > face” contrast and an anatomical mask of each
region that had been warped to native space (left/right
PHC sourced from Tzourio-Mazoyer et al., 2002; left/right
RSC sourced from Julian, Fedorenko, 韦伯斯特, & Kanwisher,
2012). 因此, the ROIs were functionally defined but
Berens, Joensen, 和霍纳
449
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
哦
C
n
_
A
_
0
1
6
5
4
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
constrained to anatomical regions known to be spatially
可选择的. Normalized group averages of these ROIs are
available at osf.io/gbznp/ and neurovault.org/collections
/4819.
Recent evidence suggests that the RSC is composed of at
least two functionally distinct subregions, both of which
may be scene selective: (1) a retinotopically organized
medial place area in posterior sections of the RSC and (2)
a more anterior region corresponding to BAs 29 和 30
associated with more integrative mnemonic processes
(Silson, Steel, & 贝克, 2016). In the current study, 我们
focus on the functionally defined RSC as a whole and do
not differentiate between these subregions. 然而,
the functional ROIs that we identified for each participant
principally cover anterior sections of the RSC corre-
sponding to BAs 29 和 30 and show little overlap with
the retinotopic areas identified by Silson et al.
The OPA has also been implicated as a critical scene-
selective region (例如, Robertson et al., 2016; Marchette
等人。, 2015). Recent research suggests that this region
is principally involved in representing environmental
boundaries and navigable paths during visual perception
(Malcolm, Silson, 亨利, & 贝克, 2018; Bonner & 爱泼斯坦,
2017; Julian, Ryan, 汉密尔顿, & 爱泼斯坦, 2016). 然而,
computational models of spatial navigation do not predict
that the OPA maintains location-based representations that
are viewpoint invariant (Bicanski & 伯吉斯, 2018; Byrne
等人。, 2007). 此外, we were only able to reliably
delineate the OPA bilaterally in 6 的 23 participants in
our sample. 像这样, we did not focus on this region in
the current study; 反而, we restricted our main analyses
and family-wise error (FWE) corrections to the PHC and RSC
bilaterally. 尽管如此, for completeness, we generated an
OPA mask using a normalized group-level contrast and ran
the location-based RSA analyses reported below on this
region separately (statistical outputs available at osf.io
/d8ucj/). No effects of interest were identified in either
the left or right OPA.
RSAs
Our general approach to the RSA involved modeling the
observed similarity between different BOLD patterns as
a linear combination of effects of interest and nuisance
变量. 这里, the similarity between BOLD responses
was taken as the correlation of normalized voxel intensi-
领带 (t statistics) across all voxels in an ROI. The resulting
correlation coefficients were then Fisher-transformed
before being subjected to statistical analysis. This trans-
form ensures that the sampling distribution of similarity
scores is approximately normal to meet the assumption
of normality for statistical inference. We then entered all
the transformed similarity scores under test from each
participant and stimulus set into a general linear mixed-
effects regression model. Although underused in the
neurosciences (although see Motley et al., 2018), 这些
models are common in the psychological literature as
they offer a robust method of modeling nonindependent
observations with few statistical assumptions (Baayen,
戴维森, & Bates, 2008). 这里, we used mixed-effects
models to predict observed representational similarity
between endpoints with a set of fixed-effects and random-
effects predictors (下面讨论).
重要的, mixed-effects models allow us to include
estimates of pattern similarity across individual items
(endpoints) and participants in the same statistical model.
The fixed-effects predictors in each model specified key
hypotheses of interest. The random effects accounted for
statistical dependencies between related observations at
both the item and participant levels. RSAs of fMRI data typ-
ically either assess patterns across all items (不管
状况) or average across items in the same condition,
meaning that important variation within conditions is
被忽略. Our modeling approach allows us to examine
changes in representational similarity at the level of both
items and conditions simultaneously while controlling for
statistical dependencies between related observations.
Raw similarity data and mean similarity matrices are
available on the Open Science Framework (osf.io/cgy97).
This page also includes MATLAB functions for estimating
each statistical model as well as the model outputs.
Visual Representations of Specific Endpoints
We first examined whether the passive viewing of end-
point images evoked stimulus-specific visual representa-
tions in each of our four ROIs (left and right PHC and
RSC). Multivariate BOLD responses to the endpoints
were estimated for Session 1 (pre-videos) and Session 2
(post-videos) separately. We then computed the similar-
ity of these responses across sessions by correlating
BOLD patterns in Session 1 with patterns in Session 2.
This resulted in a nonsymmetric, 24 × 24 correlation ma-
trix representing the similarity between all BOLD pat-
terns observed in Session 1 and those observed in
Session 2. The correlation coefficients (n = 576 per par-
ticipant) were then Fisher-transformed and entered as a
dependent variable into a mixed-effects regression model
with random effects for participants and endpoints. 这
main predictor of interest was a fixed effect that con-
trasted correlations between the same endpoints (例如,
A1–A1, B1–B1; n = 24 per participant) with correlations
between different endpoints (例如, A1–A2, A1–B1; n =
552 per participant) across the two sessions.
As well as running this analysis in each ROI, we per-
formed a complementary searchlight analysis to detect
endpoint-specific representations in other brain regions.
这里, local pattern similarity was computed for each brain
voxel using spherical searchlights with a 3-voxel radius
(the mean number of voxels per searchlight was 105.56;
searchlights were not masked by gray-/white-matter tissue
probability maps). Fisher-transformed correlations for
same versus different endpoints were contrasted at the first
level before running a group-level random-effects analysis.
450
认知神经科学杂志
体积 33, 数字 3
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
哦
C
n
_
A
_
0
1
6
5
4
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
Location-based Memory Representations
We next tested our principal hypothesis—whether repre-
sentations of Endpoints A1 and A2 became more similar to
one another as a result of watching the overlap videos—in
each ROI. Using the multivariate BOLD responses from
Sessions 1 和 2, we computed the neural similarity
between endpoints that were presented within the same
image set and the same session. This resulted in eight sym-
metric, 6 × 6 correlation matrices for each participant—one
per set in Session 1 and one per set in Session 2. 一切
correlation coefficients from the lower triangle of these
matrices (n = 15) were then Fisher-transformed and
entered as a dependent variable into a mixed-effects regres-
sion model (see Figure 1C). 像这样, the model included
120 correlation coefficients per participant (2 sessions ×
4 sets × 15 similarity scores).
One fixed-effects predictor modeled unspecific changes
in similarity between sessions (hereafter referred to the
session effect) by coding whether similarity scores were
recorded in Session 1 or Session 2. 相似地, a further three
fixed-effects predictors modeled similarity differences
attributable to (1) endpoints in the overlap condition
(IE。, A1–A2), (2) endpoints shown in the same video
(A1–A2, B1–C2), 和 (3) endpoints that were not shown
in any video (C1–B2)—shown in Figure 1C. 一起,
these predictors and their interactions constituted a 2 ×
3 factorial structure (Session [1 与. 2] × Condition [重叠
与. no-overlap vs. unseen]) and so were tested with a
Session × Condition F test. 尽管如此, our principal
hypothesis holds that there will be a specific interaction
between the Session and Overlap predictors (referred to
as the Session × Overlap effect), which we report along-
side the F test. The model also included a predictor indicat-
ing whether endpoints were from the same location
(A1–A2, B1–B2, C1–C2), thereby allowing us to estimate
changes in similarity between them. This ensured that
variance loading onto the Session × Overlap effect was
properly attributable to the learning of spatially coherent
representations rather than some combination of other
因素 (例如, same location + seen in the same video).
Note that this model term quantifies similarity differences
between overlap endpoints and all other endpoints that
“change” between Session 1 and Session 2. A positive effect
may indicate either an increase in similarity in the overlap
condition or a decrease across all other similarity scores
regardless of condition (或两者). 像这样, the model is
structured to account for any systematic change in the
baseline level of similarity across sessions (see Results).
此外, the Session × Overlap term is only sensitive
to a learning effect that causes relative shifts in similarity
scores specific to the overlap condition and cannot be
attributed to any other combination of effects.
最后, the model included a behavioral predictor spec-
ifying whether participants were able to match Endpoints
A1–A2 in the postscanner task (mean centered with three
级别: 0, 1, 或者 2 correct responses per pair). This examined
whether changes in representational similarity were
dependent on participants’ ability to identify that end-
points from the overlap condition belonged to the same
location after scanning (IE。, a three-way interaction:
Session × Overlap × Behavior). Random effects in the
model accounted for statistical dependencies across
image sets, sessions, and participants.
To complement the ROI analyses, we ran a searchlight
analysis that tested for RSA effects across the whole brain
(searchlight radius: 3 voxels). 这里, first-level contrast esti-
mates compared the Fisher-transformed correlations
between overlap endpoints (IE。, A1–A2) and all other
endpoint correlations (例如, B1–B2, B1–C1). A group-level
analysis then compared these similarity contrasts between
sessions to test the Session × Overlap interaction. To test
for a Session × Overlap × Behavior interaction, the group-
level model also included a behavioral predictor specifying
a participant’s average performance in matching A1 to A2
during the postscanner task (mean centered). 注意
this searchlight analysis is not able to control for the poten-
tial contributions of other important factors (IE。, 相同的
地点, same video) that our mixed-effects approach
explicitly controls. It is complementary, but secondary, 到
the ROI analyses.
Statistical Validation and Inference
To ensure that each mixed-effects regression model was
not unduly influenced by outlying data points, we systemat-
ically excluded observations that produced unexpectedly
large residuals more than 2.5 SDs above or below model
估计. This was conducted regardless of condition
and so did not bias the analyses to finding an effect (if no
effect were present). 此外, a highly similar pattern
of results was seen when not excluding outliers, supporting
the robustness of our findings (see osf.io/dzy3p). 下列的
these exclusions, Kolmogorov–Smirnov tests indicated that
residuals were normally distributed across all the linear
mixed-effects models. 此外, visual inspection of scat-
terplots showing residual versus predicted scores indicated
no evidence of heteroscedasticity or nonlinearity. 在哪里
effects size estimates are contrasted across different
型号, we report the result as an unequal variance t test
with the degrees of freedom being approximated using
the Welch–Satterthwaite equation (Welch, 1947).
All p values are reported as two-tailed statistics. FWE cor-
rections related to the multiple comparisons across our
four ROIs are made for each a priori hypothesis (denoted
pFWE). 此外, we report whole-brain effects from
searchlight and mass univariate analyses when they survive
FWE-corrected thresholds ( pFWE < .05) at the cluster level
(cluster-defining threshold: p < .001 uncorrected). All
other p values are noted at uncorrected levels. As well as
reporting null hypothesis significance tests, we present
the results of complimentary Bayesian analyses. Unlike
the frequentist statistics, these indicate whether the null
is statistically preferred over the alternative hypothesis.
Berens, Joensen, and Horner
451
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
o
c
n
_
a
_
0
1
6
5
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
p
ffiffiffiffiffiffiffi
0:5
As such, we use the Bayesian analyses to determine
whether there is evidence for the null when frequentist
tests are nonsignificant. For each t test, a Bayes factor in
favor of the null hypothesis (BF01) was computed with a
Cauchy prior centered at zero (i.e., no effect) and a scale
(see Gelman, Jakulin, Pittau, & Su,
parameter (r) of
2008). Bayes factors greater than 3 are taken as evidence in
favor of the null hypothesis, whereas those less than 1/3 are
taken as evidence in favor of the alternative (Kass & Raftery,
1995). Finally, alongside the inferential statistics, we report
Cohen’s d effect sizes for each t test. When effects are
tested in the context of a mixed-effects model, estimates
of Cohen’s d are computed from the fixed effects only
and exclude variance attributed to random effects.
RESULTS
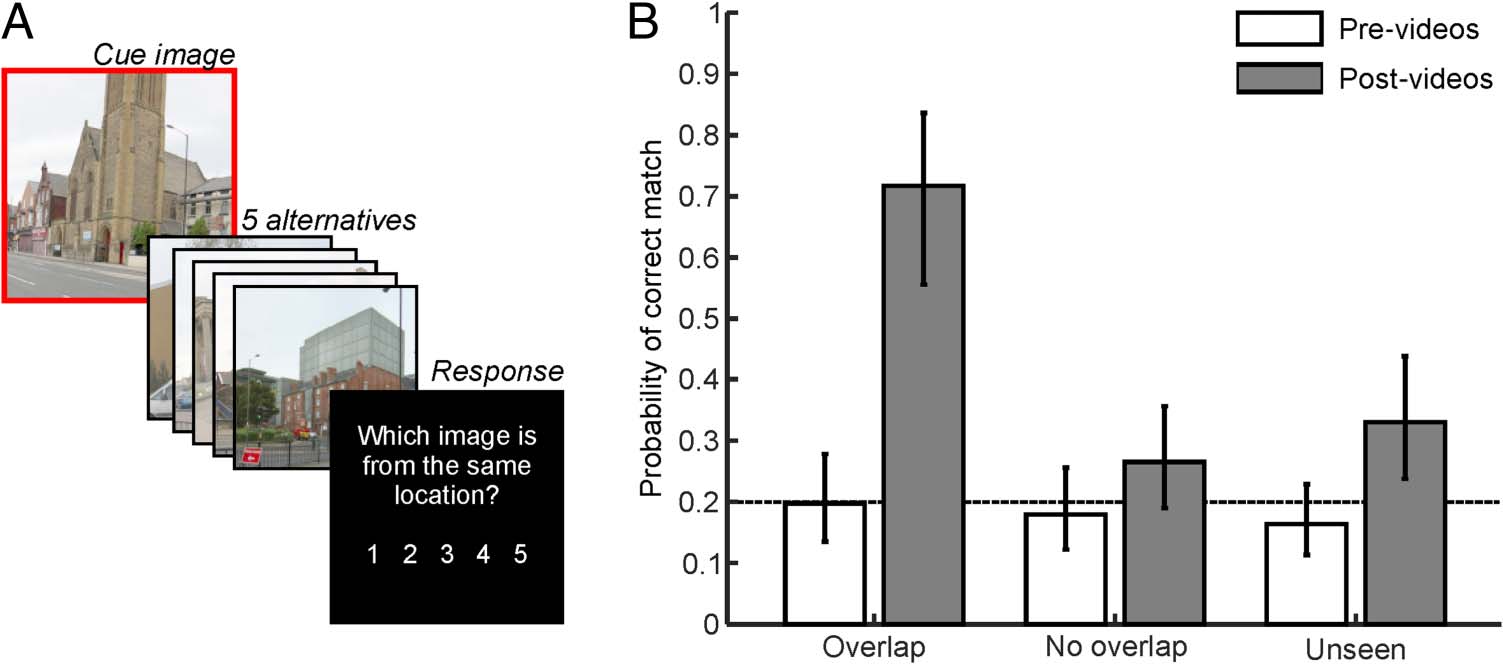
Behavioral Performance
We first analyzed behavioral responses to the prescanner
and postscanner tasks to determine (a) whether partici-
pants were able to identify which endpoints belonged to
the same location and (b) whether performance in-
creased as a result of watching the overlap videos. A gen-
eralized linear mixed-effects analysis modeled correct
versus incorrect matches between cue and target end-
points as a function of Session (pre-videos vs. post-
videos) and Experimental Condition (overlap, no overlap,
and unseen). As such, the model constituted a 2 × 3 fac-
torial design with random intercepts and slopes for both
participants and endpoints.
The results, displayed in Figure 2B, revealed significant
main effects of Session, F(1, 1098) = 47.302, p < .001, and
Condition, F(2, 1098) = 6.500, p = .002, as well as an in-
teraction between them, F(2, 1098) = 11.231, p < .001.
The interaction indicated that performance was at chance
level across all conditions before the videos (min: p =
.249, BF01 = 2.531, d = −0.241) but substantially in-
creased in the overlap video condition after the videos,
t(1098) = 6.867, p < .001, BF01 < 0.001, d = 1.432
(post-video > pre-video). This increase was not seen in
the no-overlap condition, t(1098) = 1.761, p = .079,
BF01 = 1.212, d = 0.3672; 然而, a significant increase
was seen in the unseen condition, t(1098) = 3.159, p =
.002, BF01 = 0.105, d = 0.659. The performance increases
in the control conditions (only significant in the unseen
状况) were likely the result of participants being able
to exclude overlap endpoints as nontarget alternatives in
the 5-AFC test (IE。, a recall-to-reject strategy, disregarding
A1 and A2 when cued with either B1, B2, C1, or C2).
Consistent with this, Session 2 performance in the no-
overlap and unseen conditions was not significantly different
from chance level in a 3-AFC test (0.33, as opposed to 0.2 在一个
5-AFC test; no-overlap: t(1098) = −1.494, p = .135, BF01 =
1.729, d = −0.312; unseen: t(1098) = −0.054, p = .957,
BF01 = 4.567, d = −0.011). 尽管如此, performance in
the overlap condition did significantly differ from this
adjusted chance level, t(1098) = 4.514, p < .001, BF01
= 0.006, d = 0.941.
Participants’ increased ability to match endpoints in
the overlap condition was not characteristic of a general
tendency to match endpoints that appeared in the same
video (i.e., selecting B1 when cued with C2). This was ev-
ident because matches between no-overlap endpoints
were not more likely in Session 2 compared with
Session 1, t(366) = 0.646, p = .519, BF01 = 3.785, d =
0.135. In contrast, performance increases in the overlap
condition (i.e., the post-video > pre-video effect report-
ed above) were significantly larger than this general effect
of matching all endpoints that appeared in the same vid-
eo, t(949.20) = 5.027, p < .001, BF01 = 0.002, d = 1.048.
In addition, participants were unable to explicitly match
no-overlap endpoints shown in the same video during
the final behavioral task (comparison to 0.2 chance level:
t(334) = −0.467, p = .641, BF01 = 4.141, d = −0.097).
In summary, participants rapidly learnt which scenes
were from the same location; however, this was only
seen in the overlap condition (and not in the no-overlap
condition).
Visual Representations of Specific Endpoints
First, we report the results of the mixed-effects model
testing for representations of specific endpoints that re-
mained relatively unchanged across sessions (i.e., pre-
videos to post-videos). This revealed that correlations
between the same endpoints (e.g., A1–A1, B1–B1) were
greater than correlations between different endpoints
(e.g., A1–A2, A1–B1) in both the right PHC, t(13224) =
5.229, pFWE < .001, BF01 = 0.001, d = 1.090, and the left
PHC, t(13200) = 6.351, pFWE < .001, BF01 < 0.001, d =
1.324. This effect was not significant in either the right
or left RSC, t(13210) = 1.185, pFWE = .945, BF01 = 2.454,
d = 0.247, and t(13202) = −0.231, pFWE = .999, BF01 =
4.463, d = −0.048, respectively.
The searchlight analysis that tested for consistent rep-
resentations of specific endpoints across the whole brain
revealed representations in one large cluster that peaked
in the right occipital lobe (area V1; t(22) = 11.50, pFWE <
.001, k = 5202, BF01 < 0.001, d = 2.398) and extended
into the areas V2, V3, and V4 and the fusiform gyri bilat-
erally. Three smaller clusters were also detected in the
right precuneus, t(22) = 4.64, pFWE = .011, k = 44,
BF01 = 0.005, d = 0.968, right inferior parietal lobule,
t(22) = 4.40, pFWE = .028, k = 37, BF01 = 0.008, d =
0.918, and right RSC, t(22) = 4.32, pFWE = .025, k =
38, BF01 = 0.008, d = 0.901. The latter effect overlapped
considerably with the right RSC ROI identified for each
participant. However, the effect size estimated in the
ROI analysis was weaker than the peak searchlight effect,
principally because it was variable across endpoints and
as such largely accounted for by random effects in the
model. Unthresholded statistical maps of these effects
are available at neurovault.org/collections/4819.
452
Journal of Cognitive Neuroscience
Volume 33, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
o
c
n
_
a
_
0
1
6
5
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
In summary, we find evidence that the PHC (bilaterally),
the right RSC, and a number of early visual areas main-
tained consistent representations of specific endpoints
across scanning sessions. Note that whether a region codes
such representations across scanning sessions is indepen-
dent of whether it may learn location-based memory rep-
resentations in the second session; these effects are, in
principle, dissociable.
As part of a supplementary analysis, we also tested for
visual representations of specific scenes that remained
stable within (but not necessarily across) scanning sessions
(see osf.io/exzba/). To quantify the BOLD similarity of spe-
cific scenes within each session, we required two indepen-
dent pattern representations per session. Thus, across
both sessions, we estimated voxel patters derived from
four distinct periods: (a) first half of Session 1 (pre-videos),
(b) second half of Session 1 (pre-videos), (c) first half of
Session 2 (post-videos), and (d) second half of Session 2
(post-videos). As a result, each of these voxel representa-
tions was only derived from four endpoint presentations.
Nonetheless, when similarity scores were modeled in a
mixed-effects regression, each ROI showed greater levels
of similarity between representations of the same endpoint
relative to the similarity between different endpoints
(weakest effect in the left PHC: t(26492) = 2.211, p =
.027, BF01 = 0.606, d = 0.461). Furthermore, this analysis
revealed that representations of the same endpoints be-
came more similar to one another after the videos in the
right RSC and left PHC (weakest effect: t(26492) = 2.598,
p = .009, BF01 = 0.308, d = 0.542). This latter effect was
insensitive in the right PHC and left RSC (weakest effect:
t(26492) = 1.671, p = .095, BF01 = 1.375, d = 0.348).
Location-based Memory Representations
Effects in the Right PHC
Next, we report the results of the mixed-effects model
examining whether pattern similarity between different
endpoints changed across sessions as a result of watching
the videos. This revealed a significant Session × Condition
interaction in the right PHC, F(2, 2739) = 6.827, pFWE = .004
(average similarity matrices shown in Figure 3; condition
estimates and confidence intervals plotted in Figure 4A).
Post hoc tests showed that this effect was driven by a
difference between pre-video to post-video sessions for
endpoints in the overlap condition, t(2739) = 2.923, p =
.004, BF01 = 0.167, d = 0.610. This difference was not ob-
served in any other condition (no overlap: t(2739) = 0.756,
p = .450, BF01 = 3.533, d = 0.156; unseen: t(2739) =
−0.970, p = .332, BF 0 1 = 3.001, d = −0.202).
Furthermore, a significant Session × Overlap interaction
highlighted that the similarity differences in the overlap
condition were attributable to the video manipulation
alone rather than some combination of other factors, t
(2739) = 2.549, pFWE = .043, BF01 = 0.337, d = 0.532.
Importantly, before the videos were shown, pairs of
endpoints from the same location (i.e., A1–A2, B1–B2,
and C1–C2) were found to evoke neural patterns that
were more similar to each other than pairs of endpoints
from different locations in the right PHC (e.g., A1–B2,
B1–C2), t(2739) = 2.498, pFWE = .050, BF01 = 0.369,
d = 0.521 (see osf.io/uxhs9 for a plot of this effect). This
“same-location” effect suggests that, even before the spa-
tial relationship between scenes were known, the right
PHC encoded visual properties of those scenes that gen-
eralized across different views. These data demonstrate
that, despite controlling for similarity across stimuli using
both the GIST descriptor and a pixel-wise correlation,
and despite participants being unable to infer which end-
points were from the same location before watching the
videos, we still found evidence for a “same-location” ef-
fect in the right PHC. This underlies the critical role of
estimating pattern similarity before learning to identify
significant increases in similarity post-video relative to
pre-video (cf. Robertson et al., 2016). Note that this
“same-location” effect is only seen when collapsing across
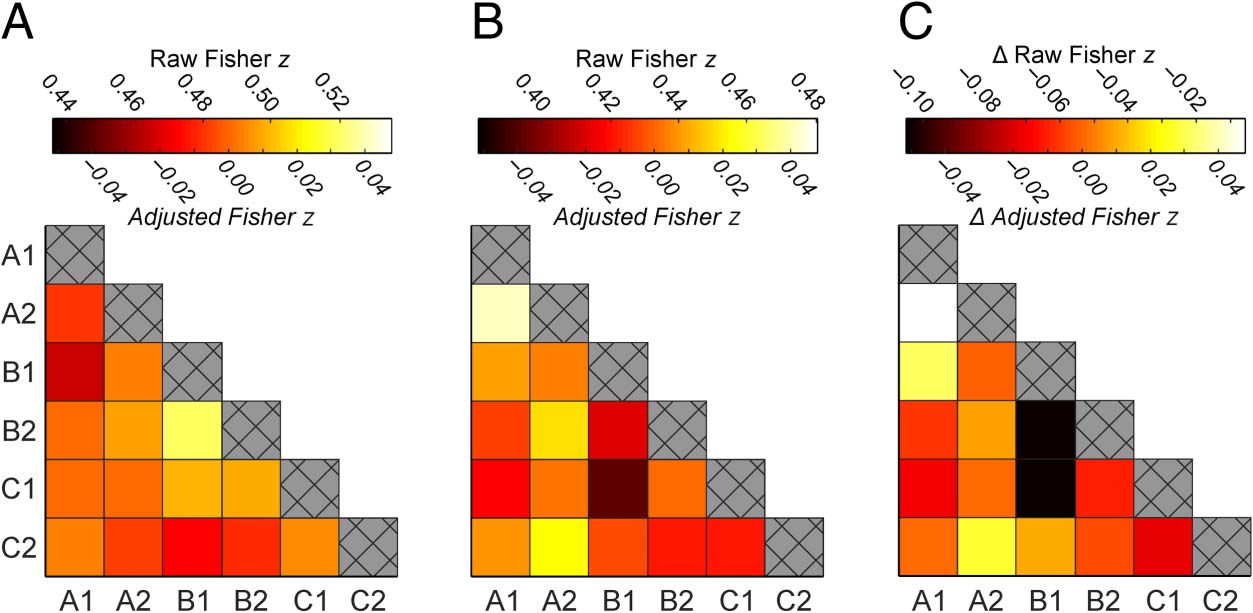
Figure 3. Mean representational
similarity between endpoints in
the right PHC, averaged across all
participants and image sets. (A)
Similarity between endpoints
before the panoramic videos
were shown (i.e., in Session 1).
(B) Similarity between endpoints
after the panoramic videos were
shown (i.e., in Session 2). (C)
Change in similarity that followed
the panoramic videos (i.e.,
Session 2 minus Session 1). Color
bars indicate both raw and
baseline-adjusted Fisher z
statistics (above and below the
color bar, respectively). Adjusted
statistics account for trivial differences in similarity across scanning sessions caused by motion and scanner drift. This is achieved by subtracting out a baseline
level of similarity between nonassociated endpoints (i.e., endpoints that were not from the same location, video, or experimental condition). Note that the
baseline-adjusted statistics are shown for illustrative purposes only; each RSA was conducted on the raw Fisher z statistics alone. Crosshatchings along the
diagonal elements represent perfect correlations between identical BOLD responses and so were not included in the analyses.
Berens, Joensen, and Horner
453
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
o
c
n
_
a
_
0
1
6
5
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
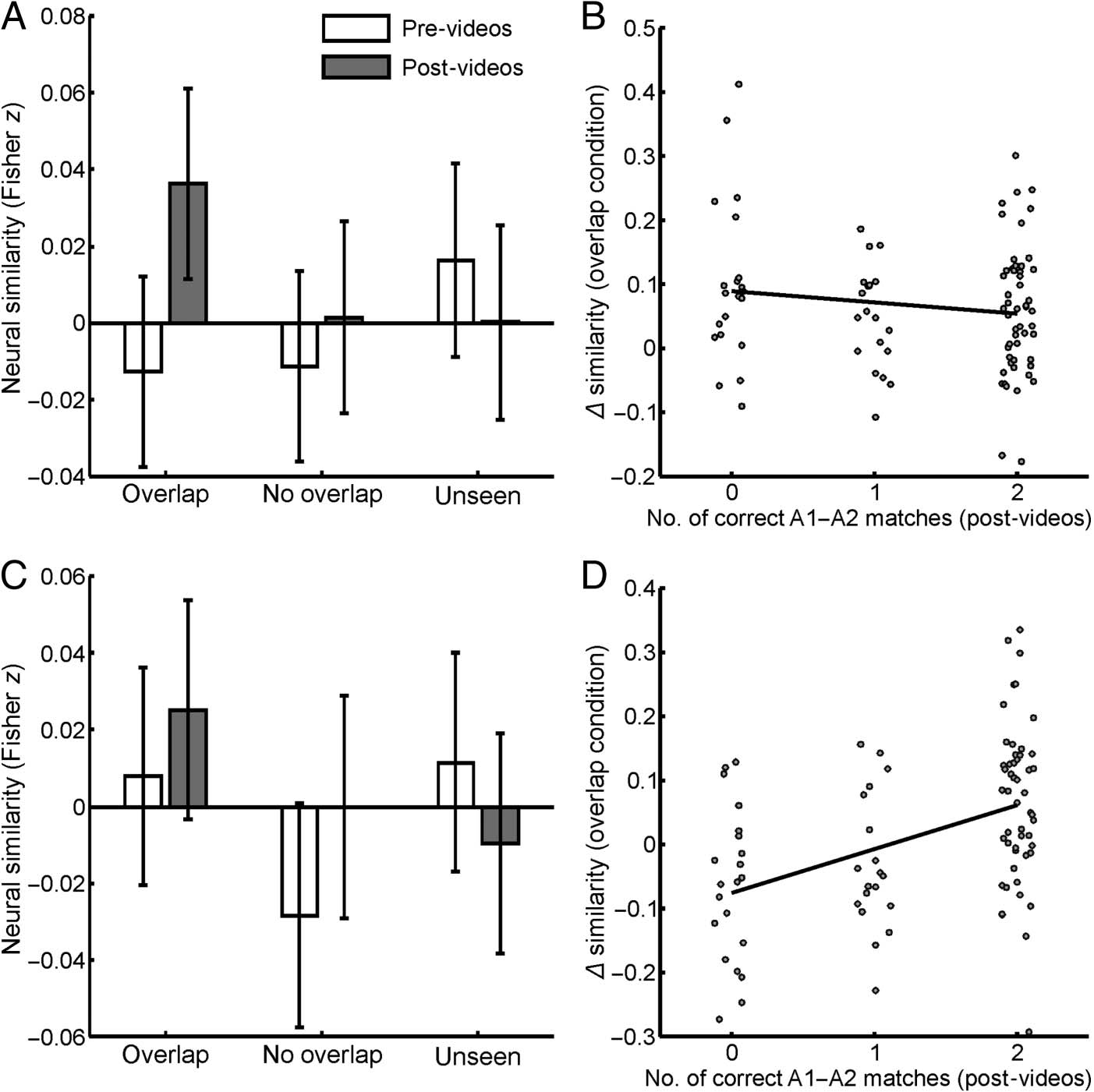
Figure 4. Results of the RSAs in
the right PHC (rPHC; top row)
and right RSC (rRSC; bottom
row). (A) rPHC similarity
estimates of scenes in the pre-
video and post-video sessions,
plotted by experimental
condition. There was a significant
change in similarity estimates
between sessions in the overlap
condition, t(2739) = 2.923, p =
.004, BF01 = 0.167, d = 0.610, that
was not present in the no-overlap
and unseen conditions, t(2739) =
0.756, p = .450, BF01 = 3.533,
d = 0.158, and t(2739) = −0.970,
p = .332, BF01 = 3.001, d =
−0.202, respectively. (B) In the
rPHC, pre-video to post-video
changes in representational
similarity for the overlap
condition plotted against the
number of correct matches
between overlap endpoints in the
post-video behavioral task. This
association was not significant,
t(2739) = −0.892, p = .373,
BF01 = 3.199, d = −0.186. (C and
D) Same as A and B but for the
rRSC ROI. The rRSC showed no
overall similarity changes in any
of the experimental conditions
(t(2728) = 0.870, t(2728) =
1.419, and t(2728) = −1.059 for
the overlap, no-overlap, and
unseen conditions, respectively;
all ps > .156, BF01s > 1.895, ds < 0.296). Nonetheless, there was a significant association between behavioral performance and similarity changes in the
overlap condition, t(2728) = 2.886, p = .004, BF01 = 0.179, d = 0.602. All bars plot baseline-corrected similarity estimates having subtracted out correlations
between nonassociated endpoints (e.g., A1–B1, A1–B2). As such, the zero line in A and C denotes the average similarity of these nonassociated endpoints in
each session. Error bars indicate 95% confidence intervals.
all endpoint pairs and is not evident in the Session 1
Overlap condition alone (osf.io/uxhs9).
Effects in the Right RSC
The Session × Condition and Session × Overlap interac-
tions were not significant in any other ROI (Fs < 2.775,
pFWEs > .250; similarity estimates for the right RSC plotted
in Figure 4C). 然而, we saw a significant Session ×
Overlap × Behavior interaction in the right RSC, t(2728) =
2.886, pFWE = .016, BF01 = 0.179, d = 0.602 (Figure 4D).
This suggests that the RSC only encoded viewpoint-
independent representations when the spatial relation-
ships between endpoints could be retrieved during the
postscanner test. No other ROIs showed a significant
Session × Overlap × Behavior interaction (largest effect:
t = 0.050, pFWE = 1, BF01 = 4.567, d = −0.010).
Differentiating the PHC and RSC
We next assessed whether there was evidence for disso-
ciable roles of the right PHC and RSC, given that both
represented location-based information but were differ-
ently associated with behavioral performance. 具体来说,
we assessed whether location-based representations in
the RSC were significantly more associated with partici-
pants’ ability to match endpoints from the same location
compared to representations in the PHC. This would sug-
gest that the RSC plays a greater role in guiding behavioral
performance than the PHC. We therefore tested whether
the Session × Overlap × Behavior (three-way) effect was
larger in the RSC than the PHC. A comparison of effect sizes
did show evidence for such a dissociation, t(5311.9) =
3.931, p < .001, BF01 = 0.021, d = 0.820.
This implies that the right PHC might have exhibited
above-baseline pattern similarity between A1 and A2
endpoints even when those endpoints were not subse-
quently remembered as belonging to the same location.
We directly tested this by rerunning the RSA having ex-
cluded A1–A2 pairs that were consistently remembered
as belonging to the same location (i.e., having two correct
responses during the postscanner test). Despite these
exclusions, pattern similarity differences in the overlap
condition remained significant, t(1188) = 2.364, p = .018,
454
Journal of Cognitive Neuroscience
Volume 33, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
o
c
n
_
a
_
0
1
6
5
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
BF01 = 0.528, d = 0.493, and were not seen in any other
condition (no-overlap: t(1188) = 0.324, p = .746, BF01 =
4.359, d = 0.068; unseen: t(1188) = −0.585, p = .559,
BF01 = 3.915, d = −0.122; see Figure 4B that plots the
size of the Session × Overlap effect in the right PHC at
each level of behavioral performance. In contrast, the right
RSC only showed above-baseline pattern similarity
when the endpoints were consistently remembered as
belonging to the same location. Rerunning the RSA on
these remembered pairs alone revealed similarity increases
between consistently remembered endpoints in the
overlap condition, t(1538) = 2.449, p = .014, BF01 =
0.402, d = 0.511 (see Figure 4D), that were not seen in
any other condition (no-overlap: t(1538) = 1.107, p =
.269, BF01 = 2.651, d = 0.230; unseen: t(1538) = −1.316,
p = .188, BF01 = 2.134, d = −0.274).
In summary, we saw an increase in pattern similarity in
the right PHC and right RSC between different scenes of
the same location after they had been presented in an over-
lap video. Furthermore, we observed a dissociation
between the PHC and the RSC. Whereas the PHC showed
increased pattern similarity regardless of performance on
the postscanner test, the RSC only showed increased pat-
tern similarity when participants were able to subsequently
identify those scenes as belonging to the same location.
Across-Session Decreases in Pattern Similarity
Our mixed-effects regression models were conducted on
the raw Fisher z scores computed from each pair of
endpoints. This ensured that effects were not driven by
complex data manipulation or scaling, and so the data were
not adjusted to account for across-session shifts in the
similarity of all multivariate patterns (see Methods).
Interestingly however, we did observe that Fisher z scores
decreased from pre-video to post-video across all pairs of
endpoints regardless of condition, in each ROI (see figure
at osf.io/2y3pm). This is reflected by a notable session
effect in each mixed-effects model indicating reduced
levels of similarity between nonassociated endpoints (i.e.,
endpoints not belonging to the same location, video, or
experimental condition; minimum effect size: t(2736) =
−1.529, p = .126, BF01 = 1.655, d = 0.319). As the size
of this session effect was relatively large, the Session ×
Overlap and Session × Overlap × Behavior interactions
involved less of a decrease in similarity scores relative to
all other conditions (see Figure 3).
Given that similarity scores decrease across all endpoint
pairs, it is unlikely that the Session effect was a direct result
of our video manipulation (i.e., learning-induced neural
differentiation). A mass differentiation on this scale would
imply implausibly large amounts of information gain as the
uniqueness (or entropy) of all neural representations
would have to increase. Instead, it is more likely that the
reduced levels of similarity were caused by systematically
higher levels of noise in the second session. Most signifi-
cantly, increases in temperature caused by radio frequency
absorption during scanning will shift the thermal equilib-
rium that governs how many hydrogen nuclei are aligned
to the external magnetic field (B0) and can therefore con-
tribute to the MR signal (see osf.io/8kns6/). In this case, we
would expect to see similar shifts in the level of similarity
across the entire brain. To test this, we measured pattern
similarity in the genu of the corpus callosum, a region that
should exhibit negligible levels of BOLD activity. On the ba-
sis of a seed voxel at Montreal Neurological Institute of [0,
26, 6], multivariate patterns were taken from the 122 white-
matter voxels closest to that seed in native space. The size
of this ROI was chosen to reflect the average size of our a
priori ROIs. A mixed-effects regression model of these data
did indeed show reduced levels of neural similarity from
Session 1 to Session 2, t(2739) = −2.167, p = .030, BF01 =
0.651, d = −0.452 (similar in magnitude to the session
effect in all other regions; see osf.io/p9qzx/).
In summary, we conclude that the overall decrease in
pattern similarity across sessions was not driven by any
meaningful change in neural representations and, once con-
trolled for, reveals a significant increase in pattern similarity
in both the right PHC and RSC in the overlap condition,
indicative of viewpoint-independent representations.
Laterality of RSA Effects
The above analyses identified location-based representa-
tions in both right-hemisphere ROIs but no similar effects
in the left hemisphere. Given this, we explored whether
each RSA effect was significantly stronger in the right versus
left hemisphere. Comparing the Session × Overlap effects
in the PHC did indeed reveal a significantly stronger effect in
the right hemisphere, t(5390.5) = 3.798, p < .001, BF01 =
0.028, d = 0.792. Similarly, comparing the Session ×
Overlap × Behavior interactions in the RSC revealed a signif-
icantly stronger effect in the right hemisphere, t(5427.4) =
2.708, p = .007, BF01 = 0.251, d = 0.565. Note that
Robertson et al. (2016) collapsed their analyses across hemi-
sphere, potentially masking laterality effects. These results
are consistent with observations and theoretical models that
the right hemisphere may preferentially process spatial in-
formation in humans as a consequence of predominantly
left-lateralized language function (Shulman et al., 2010;
Vallortigara & Rogers, 2005; Smith & Milner, 1981).
Searchlight RSA
The searchlight analysis that tested for a Session × Overlap
interaction across the whole brain revealed one small
cluster in the right inferior occipital gyrus (Area V4), t(21) =
4.78, pFWE = .010, k = 38, BF01 < 0.003, d = 0.997. However,
when BOLD similarity in the cluster was modeled with the
full mixed-effects analysis described above, the Session ×
Overlap effect was found to not be significant, t(2740) =
1.734, p = .083 (uncorrected), BF01 = 1.259, d = 0.361.
Model parameter estimates suggested that the searchlight
effect was driven by below-baseline BOLD similarity in the
Berens, Joensen, and Horner
455
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
o
c
n
_
a
_
0
1
6
5
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
overlap condition before the videos were shown (95% CI
[−0.116, −0.021]), a result that is not consistent with any
effect of interest. No other areas showed a significant
Session × Overlap or Session × Overlap × Behavior inter-
action in the searchlight analysis. Nonetheless, both of the
previously reported effects in the PHC and RSC are evi-
dent in the searchlight analysis at subthreshold levels,
t(21) > 2, d > 0.417 (see neurovault.org/collections/4819/).
Univariate Responses to Endpoints
We investigated whether each of our ROIs produced uni-
variate BOLD activations consistent with a Session ×
Condition interaction or a three-way interaction with
行为. No such effects were found, all Fs < 1.140,
ps > .288. 此外, a mass univariate analysis testing
for these effects at the whole-brain level yielded no sig-
nificant activations.
Univariate Responses to Videos
最后, we investigated whether univariate BOLD re-
sponses to the video clips differed between the overlap
and no-overlap conditions or as a function of scene mem-
ory in the postscanner test. A group-level model was spec-
ified with predictors for (1) video type (overlap vs.
no-overlap), (2) post-video performance in matching A1
and A2 endpoints, 和 (3) the interaction between video
type and behavioral performance. This revealed two clus-
ters that produced significantly greater BOLD responses
during overlap versus no-overlap videos (数字 5, hot
颜色). The largest of these peaked in the medial pFC
and extended into the anterior cingulate, left frontal pole,
and left middle frontal gyrus, t(21) = 5.53, pFWE < .001, k =
600, BF01 < 0.001, d = 1.153. The second cluster peaked in
the left supramarginal gyrus, t(21) = 5.40, pFWE = .004, k =
185, BF01 = 0.001, d = 1.126, adjacent to a smaller, sub-
threshold effect in the left angular gyrus.
No effects for the reverse contrast (i.e., no overlap >
重叠) reached statistical significance at the whole-brain
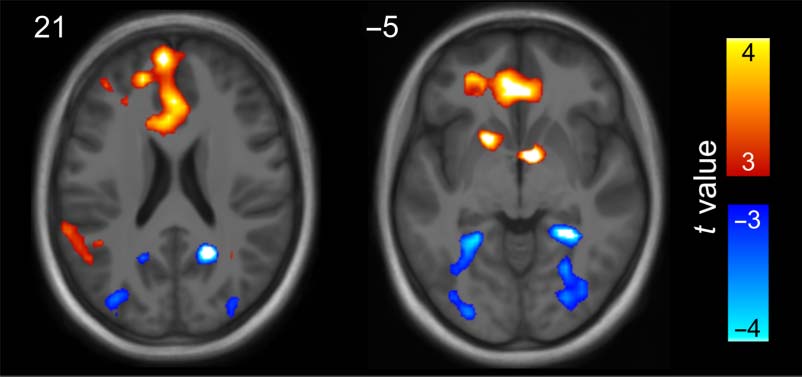
数字 5. Univariate BOLD effects showing differences in activity
between the two video conditions (thresholded at t(21) > 3, p < .004
uncorrected, BF01 < 0.143, d > 0.626). Hot colors indicate areas
showing a greater response to overlap versus no-overlap videos. Cool
colors indicate areas showing a greater response to no-overlap versus
overlap videos. An unthresholded statistical map of this contrast is
available at neurovault.org/collections/4819.
等级 (subthreshold effects shown in Figure 5, cool colors).
然而, a small volume correction for the PHC and RSC
bilaterally revealed two clusters with a significant no-
overlap > overlap effect. These were found in the right
RSC, t(21) = −4.84, pFWE = .032, k = 26, BF01 = 0.003,
d = −1.001, and right PHC, t(21) = −4.77, pFWE = .026,
k = 30, BF01 = 0.003, d = −0.995, extending into the fusi-
form gyrus. Subthreshold effects for the no-overlap > over-
lap contrast were also evident in the left RSC and PHC.
These results were mirrored by a linear mixed-effects
model contrasting overlap and no-overlap video responses
averaged across each ROI in native space. 这里, 两者都
right PHC and right RSC exhibited greater BOLD activity
in the no-overlap video condition relative to the overlap
状况, t(42) = −3.638, pFWE = .003, BF01 = 0.039,
d = −0.759, and t(42) = −3.499, pFWE = .004, BF01 =
0.052, d = −0.730, 分别. Effects in the left PHC
and left RSC were below threshold and considerably
weaker, t(42) = −1.828, pFWE = .299, BF01 = 1.101, d =
−0.381, and t(42) = −2.212, pFWE = .130, BF01 = 0.605,
d = −0.461, 分别. Neither the whole-brain analysis
nor the mixed-effects model identified BOLD responses
to the videos that significantly correlated with memory
performance in the postscanner test.
总之, we saw greater activity in the medial pFC
during the overlap videos relative to the no-overlap videos.
相比之下, the PHC and RSC showed greater activity during
the no-overlap relative to overlap videos. 换句话说,
the medial posterior regions that showed increased pattern
similarity after presentation of the overlap video showed
decreased activity while participants were watching the
视频.
讨论
We show that scene-selective brain regions rapidly learn
location-based representations of novel environments by
integrating information across different viewpoints. 一次
participants observed the spatial relationship between
two viewpoints from a given location, BOLD pattern sim-
ilarity between viewpoints increased in the right PHC and
RSC, implying the emergence of location-based represen-
tations. In the right PHC, these representations appeared
regardless of whether participants could identify which
scenes were from the same location. 相比之下, 代表-
sentations in the right RSC only emerged for scene pairs
that participants could subsequently identify as being
from the same location.
The results provide further evidence that the PHC and
RSC support spatial representations that are not solely
driven by visual features in a scene (Robertson et al., 2016;
Marchette et al., 2015; Vass & 爱泼斯坦, 2013; 比照. 沃森,
Hartley, & Andrews, 2017). Using a similar panoramic video
manipulation, Robertson et al. (2016) suggested that the
RSC and OPA maintain viewpoint-independent representa-
tions but found a more general associative effect in the
PHC. Our results further identify the PHC in this process
456
认知神经科学杂志
体积 33, 数字 3
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
哦
C
n
_
A
_
0
1
6
5
4
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
and highlight that RSC representations are more tightly
linked to behavior. Note that the OPA was not one of our
a priori ROIs, and we therefore make no claims in relation
to this region supporting location-based representations
(see ROIs section for further details). Our results also place
constraints on models that describe how location-based
representations are used. Unlike Robertson et al., we show
that viewpoint-independent representations are evoked
during passive viewing, in the absence of any explicit
memory task (although we cannot rule out the possibility
that participants engaged in active imagery, as explicitly re-
quired in Robertson et al.; 见下文).
此外, we show that the learning of location-
based representations can take place rapidly (in a single
scanning session), with few exposures to the spatial layout
of a location. Consistent with this, the firing fields of place
cells have been shown to emerge rapidly in the rodent
hippocampus (Monaco et al., 2014). Novel locations,
where rats engaged in head-scanning behavior (IE。, explo-
配给), were associated with place fields the next time the
rat visited the same location. Our results provide evidence
that location-based representations form after only three
learning exposures to the videos. Although we were specif-
ically interested in the emergence of viewpoint-independent
spatial representations, a similar approach could be used to
track the emergence of viewpoint-independent represen-
tations of other stimulus categories (例如, objects or faces;
see Clarke et al., 2016, for a similar approach), opening
the door to understanding how such representations are
形成的, or modulated, across the visual system.
We also found that the right RSC only exhibited location-
based representations when participants were able to iden-
tify which scenes belonged to that location in a postscanner
测试 (PHC representations emerged regardless of behav-
ioral performance on the postscanner test). This implies
that the ability to identify differing scenes as from the same
location is perhaps more dependent on representations
in the RSC than PHC. Computational models hold that
medial posterior and temporal regions (包括
PHC and RSC) perform distinct but complementary func-
tions in support of spatial navigation and imagery
(Bicanski & 伯吉斯, 2018; 伯恩等人。, 2007). 具体来说,
the PHC is thought to represent allocentric information
related to the spatial geometry of the environment.
反过来, the posterior parietal cortex supports egocen-
tric representations that allow the organism to actively
navigate. The RSC transforms allocentric representations
in the MTL into egocentric representations in the parietal
cortex (and vice versa). Critically, the models predict that
spatial navigation and planning is carried out in an egocen-
tric reference frame. 因此, the RSC is critical to the transla-
tion of allocentric to more behaviorally relevant, egocentric
信息.
Our task required participants to match distinct scenes
from the same location. This likely requires transformation
from the presented egocentric viewpoint to an allocentric
表示 (egocentric-to-allocentric; IE。, A1 to the
allocentric representation A*). 反过来, the allocentric
representation may allow for the retrieval of the associ-
ated viewpoint from the same location (allocentric-to-
egocentric; IE。, A* to A2). Under this assumption, 这
RSC is likely to be more tightly coupled to behavior relative
to the PHC, as shown in the present data. 这是因为
allocentric representations in the PHC only require the ini-
tial egocentric-to-allocentric transformation to be retrieved
(A1–A*). If only the egocentric-to-allocentric transforma-
tion occurs, participants will not be able to perform the
任务. 像这样, it is possible to see evidence for allocentric
PHC representations in the absence of accurate behavior.
For allocentric representations in the RSC to be retrieved,
both the initial egocentric-to-allocentric (A1–A*) and sub-
sequent allocentric-to-egocentric (A*–A2), 转型
is required. If both transformations occur, then partici-
pants should be able to perform the task accurately.
因此, location-based representations in the RSC may only
be seen in the presence of accurate behavior and may
reflect the transformation between reference frames rather
than reflecting an allocentric representation per se.
A related possibility is that, during the passive viewing of
specific scenes, participants engaged in active imagery of
the associated scenes, leading to subsequent improve-
ments in behavior for scenes from the same location.
然而, we note that the task did not explicitly require
memory retrieval; participants responded to oddball tar-
gets leaving little time for active imagery (see Linde-
Domingo, Treder, Kerrén, & Wimber, 2019). 此外,
participants would only be able to engage in active imagery
on the overlap trials alone. Despite this, we did not ob-
serve any univariate BOLD effects indicative of additional
processing on these trials. 像这样, the activation of these
representations does not appear to depend on any task-
specific memory demands. It is possible that the retrieval
of PHC representations (IE。, egocentric-to-allocentric
映射) occurs relatively automatically, consistent with
the proposal that allocentric representations in the MTL
are automatically updated during self-motion in an envi-
罗门特 (Bicanski & 伯吉斯, 2018; 伯恩等人。, 2007).
然而, the retrieval of associated egocentric represen-
tations (IE。, allocentric-to-egocentric mapping) may not
occur automatically during passive viewing, consistent
with the observation that viewpoint-independent repre-
sentations in the RSC are abolished when participants en-
gage in a task that prevents them from active retrieval of
空间信息 (Marchette et al., 2015). 重要的,
both of the above accounts are consistent with the pro-
posal that the RSC plays a critical role in mapping between
allocentric and egocentric representations.
Although consistent with models of allocentric process-
英, it is possible that the location-based representations
we observed reflect other forms of associative learning
(例如, O’Reilly & Rudy, 2001). On this view, Scene A1 may
become bound to A2 via a simple associative representa-
tion such that, after seeing the videos, A2 is covertly
retrieved when presented with A1 (leading to increased
Berens, Joensen, 和霍纳
457
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
哦
C
n
_
A
_
0
1
6
5
4
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
pattern similarity). 然而, contrary to our findings, 这
simple account may also predict increased similarity in the
no-overlap condition, where B1 and C2 are shown in the
same video—particularly given that models of associative
learning often rely on prediction error signals to account
for incidental encoding (Den Ouden, 弗里斯顿, Daw,
McIntosh, & Stephan, 2009), which could be strongest in
the no-overlap condition. A second possibility is that the
overlapping content in the overlap videos (relative to the
no-overlap videos) increases the probability of a direct
association between A1 and A2. 的确, it is the overlap-
ping content that likely drives the increase in pattern
similarity between overlap endpoints. Our current study
is not able to discern whether the resulting “location-
based” representations are associative, or truly allocentric,
in nature.
In terms of associative learning, a related possibility is
that the overlapping content supports a more complex tran-
sitive representation (例如, A1–AX and A2–AX where X is the
overlapping scene in the center of the panorama). On this
帐户, presentation of A1 cues the retrieval of AX and
subsequently A2 (similar in nature to AB–AC inference par-
adigms; see Joensen, Gaskell, & 霍纳, 2020; Schlichting,
Mumford, & 普雷斯顿, 2015; 霍纳 & 伯吉斯, 2014;
Schlichting, Zeithamova, & 普雷斯顿, 2014; Zeithamova,
Dominick, & 普雷斯顿, 2012). Representations that encode
these transitive relationships between scenes are possible
and may support spatial navigation but are not directly pre-
dicted by models of spatial memory (Bicanski & 伯吉斯,
2018; 伯恩等人。, 2007). 此外, the hippocampus
and medial PFC (mPFC) are more typically associated with
transitive inference (Schlichting et al., 2014, 2015;
Zeithamova et al., 2012), yet we only found evidence of
location-based representations in scene-selective regions.
此外, Robertson et al. have demonstrated that asso-
ciative memory for scenes belonging to different locations
is poor (comparable to their no-overlap condition) 甚至
when those scenes are presented in a “morphed” panorama
such that they are associated with a common context. 作为
这样的, our data are suggestive of processes that go beyond
associative or transitive learning and provide support for
models of allocentric processing, although we cannot rule
out an “associative” explanation.
最后, it is noteworthy that certain nonspatial models
may be able to account for our findings. 尤其,
models of directed attention may predict increased levels
of pattern similarity in the overlap condition if the over-
lap videos alerted participants to visual features that are
shared across scenes (例如, Luo, Roads, & Love, 2020; Mack,
普雷斯顿, & Love, 2013). Further work will be needed
to fully establish the true nature of the location-based
representations that we report here. To fully match all
visual features across scenes in each condition, one pos-
sibility would be to experimentally manipulate the central
section of continuous panoramas so that no coherent
spatial representation can be learned. 此外, to fully
distinguish between allocentric and transitive (A1–AX–A2)
陈述, an imaging study incorporating the pano-
ramic morph manipulation used by Robertson et al. 可能
be used.
Although we directly link to computational models of
spatial navigation and imagery, as well as rodent studies
on spatial navigation, it is important to note that we have
assessed pattern similarity during visual presentation of
static scenes. This is a common approach in human
功能磁共振成像 (Bonner & 爱泼斯坦, 2017; Robertson et al., 2016;
Marchette et al., 2015; Marchette et al., 2014), as it allows
one to control for many potential experimental con-
founds that might be present in a more ecologically valid
experimental setting (例如, using virtual reality; Julian
等人。, 2016; Doeller, Barry, & 伯吉斯, 2010). 然而,
this approach has the issue of being further removed
from real-world spatial navigation. 有趣的是, we saw
evidence for increases in pattern similarity despite using
a low-level attentional task, speaking to the potential
automaticity of retrieving more location-based represen-
tations. Across the literature, there are numerous exam-
ples of evidence for spatial learning in humans and
rodents during goal-directed navigation (Aoki, Igata,
Ikegaya, & Sasaki, 2019; Howard et al., 2014), non-goal-
directed navigation (例如, 奥基夫 & Dostrovsky, 1971;
Tolman, 1948), mental imagery (例如, Bellmund, Deuker,
Schröder, & Doeller, 2016; 霍纳, Bisby, Zotow, 衬套,
& 伯吉斯, 2016), and viewing of static images (例如,
Robertson et al., 2016; Marchette et al., 2015; Vass &
爱泼斯坦, 2013). Our study adds to this growing literature
suggesting that these representations can be assessed
across diverse experimental environments with multiple
methodologies.
The PHC has been proposed to represent several com-
plementary spatial representations, including geometric
information regarding one’s location in relation to bear-
ings and distances to environmental features (例如,
边界; 公园, 布雷迪, Greene, & Oliva, 2011). 这
representations that we observed in PHC may reflect en-
riched spatial representations relating specific scenes to
environmental features outside the current field of view.
Also consistent with our results, the PHC may represent
spatial contexts more broadly (爱泼斯坦 & Vass, 2014). 这
experimental manipulation used here could be modified
to learn novel locations in the same spatial context,
potentially dissociating between the above accounts. A
further proposal is that viewpoint-independent represen-
tations in the PHC reflect prominent landmarks that are
visible across viewpoints (Marchette et al., 2015). 虽然
this proposal yields similar predictions to above, it is less
able to account for our finding of shared representations
of views that did not contain any of the same landmarks.
Our PHC results are somewhat inconsistent with those
of Robertson et al. (2016). Whereas our similarity increases
were specific to the overlap condition, Robertson et al. saw
effects in both their overlap and no-overlap conditions.
One possibility is that our results reflect a Type II error,
in that we failed to find an effect in the no-overlap
458
认知神经科学杂志
体积 33, 数字 3
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
哦
C
n
_
A
_
0
1
6
5
4
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
condition when one is present. A second possibility is that
Robertson et al. 任何一个 (1) found an effect in the no-overlap
condition when one is not present (IE。, a Type I error) 或者
(2) failed to find a similarity effect in the overlap condition
that was significantly larger than in the no-overlap condi-
的 (a Type II error). 尤其, the “overlap > no-overlap”
effect size that we observed in the PHC is considerably
larger than the same effect reported by Robertson et al.
(0.610 relative to −0.062) and more in line with their RSC
and OPA effects (0.470 和 0.415, 分别). 因此, 它
seems plausible that the disagreement stems from a Type
I error in Robertson et al. Despite this, without further
信息, it is not possible to draw clear conclusions.
然而, one important caveat is that we also saw
evidence for a “same-location” effect in the PHC before
learning had occurred. This effect was seen despite con-
trolling for visual similarity across stimuli using the GIST
descriptor, accounting for pixel-wise correlations in lumi-
nance and color content, and despite participants being
unable to identify which endpoints were from the same lo-
cation before the videos. It is therefore possible that the
PHC effects in Robertson et al. could have been driven
by a similar effect not dependent on learning. This under-
lines the importance of including a prelearning versus post-
learning estimate of pattern similarity, to definitively rule
out trivial effects driven by preexisting similarities between
images that are difficult to control for.
RSC representations may reflect the retrieval of spatial or
conceptual information associated with the environment
(Marchette et al., 2015). Further evidence suggests that
the RSC contains multiple viewpoint-dependent and
viewpoint-independent (Vass & 爱泼斯坦, 2013), 也
local and global (Jacob et al., 2017; Marchette et al., 2014),
spatial representations. This multitude of representations
fits with the proposed role of the RSC as a transformation
circuit, mapping between allocentric and egocentric repre-
句子. The heterogeneity of representations, relative
to the PHC, may also be a further reason why we did not
see clear evidence for location-based representations with-
out taking behavior into account. Our RSC results are con-
sistent with those of Robertson et al. in that they saw a clear
effect after more extensive learning across 2 天 (在哪里
behavioral performance was likely higher than in our study).
然而, we extend these findings to show that these effects
are specifically associated with the locations that each
individual participant has learned (IE。, a within-participant
correlation that is consistent across participants). Regardless
of the exact nature of such representations, our results
provide clear evidence that we can track their emergence
in both the PHC and RSC.
Although more explorative, we also examined activity
during learning of new spatial relationships (IE。, 视频
presentation). BOLD activations in medial posterior brain
地区 (including but not limited to the PHC and RSC
ROIs) were greater for no-overlap videos compared to
overlap videos. This effect perhaps reflects greater fMRI
adaptation during the overlap videos because they
presented the central viewpoint of the panorama more
frequently than no-overlap videos (数字 1). 然而,
it is interesting that the same cortical regions that
showed increased pattern similarity after presentation
of the overlap video showed decreased activity when par-
ticipants were watching the videos. This underlines the
complex relationship between univariate activity during
learning and resultant changes in patterns of activity after
学习. More theoretically driven research would be
needed to provide a robust explanation for this finding.
此外, we found that mPFC showed a greater
BOLD response in the overlap than no-overlap condition.
This may reflect a mnemonic integration process that
guides the learning of viewpoint-independent represen-
tations. Similar effects in mPFC have been observed in
tasks that require integrating overlapping memories to
support inference and generalization (Milivojevic, Vicente-
Grabovetsky, & Doeller, 2015; Schlichting et al., 2015).
的确, mPFC has been implicated in detecting new
information that is congruent with previously learnt
materials so that it can be integrated into a generalized
表示 (van Kesteren, Ruiter, Fernández, &
Henson, 2012). Our results are broadly in line with this
proposal, where mPFC may be detecting the presence of
overlapping spatial information during the overlap videos,
resulting in the integration of previously learnt represen-
tations into more coherent viewpoint-independent repre-
sentations in posterior medial regions. Despite this, 我们的
results do not exclude the possibility that mPFC activa-
tions reflect disinhibition from medial–posterior inputs
(which showed reduced activity) or attentional differences
related to the behavioral task.
We have shown that brain regions in the scene network,
specifically the right PHC and RSC, rapidly learn represen-
tations of novel environments by integrating information
across different viewpoints. They appear to be relatively
viewpoint-independent in that they become active regard-
less of which part of an environment is in the current field
的观点. We show that the PHC and RSC have potentially
dissociable roles, consistent with models that propose
the RSC plays a role in translating viewpoint-independent
representations into a behaviorally relevant egocentric
reference frame. 最后, our experimental approach allows
for tracking the emergence of viewpoint-independent
representations across the visual system.
致谢
We thank all the staff at the York Neuroimaging Centre for their
assistance in running this project. We are also grateful to Tim
Andrews and Tom Hartley for early discussions regarding exper-
imental design. 乙. H. J. was funded a PhD studentship awarded
by the Department of Psychology, 约克大学.
Reprint requests should be sent to Sam C. Berens or Aidan J.
霍纳, 心理学系, 约克大学, 约克
YO10 5DD, 英国, or via e-mails: s.berens@sussex.ac.uk, aidan
.horner@york.ac.uk.
Berens, Joensen, 和霍纳
459
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
哦
C
n
_
A
_
0
1
6
5
4
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
Author Contributions
Sam C. Berens: 概念化; 数据管理; 正式的
分析; 调查; 方法; 项目管理;
可视化; Writing—original draft; Writing—review &
编辑. Bárður H. Joensen: 概念化; Data cura-
的; 方法; Writing—review & 编辑. Aidan J.
霍纳: 概念化; 形式分析; 资金
acquisition; 调查; 方法; Project adminis-
翻译; 监督; 可视化, Writing—original
草稿; Writing—review & 编辑.
Funding Information
Aidan J. 霍纳: Wellcome Trust (http://dx.doi.org/10
.13039/100004440), Grant number: 204277/Z/16/Z,
Economic and Social Research Council (http://dx.doi.org
/10.13039/501100000269), Grant number: ES/R007454/1.
参考
安德森, J. L. R。, Hutton, C。, Ashburner, J。, 车工, R。, &
弗里斯顿, K. (2001). Modeling geometric deformations in EPI
time series. Neuroimage, 13, 903–919. DOI: https://doi.org
/10.1006/nimg.2001.0746, PMID: 11304086
Aoki, Y。, Igata, H。, Ikegaya, Y。, & Sasaki, 时间. (2019). The integration
of goal-directed signals onto spatial maps of hippocampal
place cells. Cell Reports, 27, 1516–1527. DOI: https://doi.org
/10.1016/j.celrep.2019.04.002, PMID: 31042477
Ashburner, J. (2007). A fast diffeomorphic image registration
algorithm. Neuroimage, 38, 95–113. DOI: https://doi.org
/10.1016/j.neuroimage.2007.07.007, PMID: 17761438
Baayen, 右. H。, 戴维森, D. J。, & Bates, D. 中号. (2008). Mixed-
effects modeling with crossed random effects for subjects
and items. 记忆与语言杂志, 59, 390–412.
DOI: https://doi.org/10.1016/j.jml.2007.12.005
Bellmund, J. L. S。, Deuker, L。, Schröder, 时间. N。, & Doeller, C. F.
(2016). Grid-cell representations in mental simulation. 电子生活,
5, e17089. DOI: https://doi.org/10.7554/eLife.17089, PMID:
27572056, PMCID: PMC5005038
Bicanski, A。, & 伯吉斯, 氮. (2018). A neural-level model of spatial
memory and imagery. 电子生活, 7, e33752. DOI: https://doi.org/10
.7554/eLife.33752, PMID: 30176988, PMCID: PMC6122954
Bonner, 中号. F。, & 爱泼斯坦, 右. A. (2017). Coding of navigational
affordances in the human visual system. 诉讼程序
美国国家科学院, 美国。, 114, 4793–4798. DOI:
https://doi.org/10.1073/pnas.1618228114, PMID: 28416669,
PMCID: PMC5422815
伯吉斯, N。, Becker, S。, 国王, J. A。, & 奥基夫, J. (2001).
Memory for events and their spatial context: Models and
实验. Philosophical Transactions of the Royal Society
of London, Series B, Biological Sciences, 356, 1493–1503.
DOI: https://doi.org/10.1098/rstb.2001.0948, PMID:
11571039, PMCID: PMC1088531
Byrne, P。, Becker, S。, & 伯吉斯, 氮. (2007). Remembering the
past and imagining the future: A neural model of spatial
memory and imagery. 心理评论, 114, 340–375.
DOI: https://doi.org/10.1037/0033-295X.114.2.340, PMID:
17500630, PMCID: PMC2678675
Calton, J. L。, & Taube, J. S. (2009). Where am I and how will I
get there from here? A role for posterior parietal cortex in the
integration of spatial information and route planning.
Neurobiology of Learning and Memory, 91, 186–196. DOI:
https://doi.org/10.1016/j.nlm.2008.09.015, PMID: 18929674,
PMCID: PMC2666283
克拉克, A。, Pell, 磷. J。, Ranganath, C。, & Tyler, L. K. (2016).
Learning warps object representations in the ventral
temporal cortex. 认知神经科学杂志, 28,
1010–1023. DOI: https://doi.org/10.1162/jocn_a_00951,
PMID: 26967942
Den Ouden, H. 乙. M。, 弗里斯顿, K. J。, Daw, 氮. D ., McIntosh, A. R。,
& Stephan, K. 乙. (2009). A dual role for prediction error in
associative learning. 大脑皮层, 19, 1175–1185. DOI:
https://doi.org/10.1093/cercor/bhn161, PMID: 18820290,
PMCID: PMC2665159
Doeller, C. F。, Barry, C。, & 伯吉斯, 氮. (2010). Evidence for grid
cells in a human memory network. 自然, 463, 657–661.
DOI: https://doi.org/10.1038/nature08704, PMID: 20090680,
PMCID: PMC3173857
Eichenbaum, H. (2004). Hippocampus: Cognitive processes
and neural representations that underlie declarative memory.
神经元, 44, 109–120. DOI: https://doi.org/10.1016/j.neuron
.2004.08.028, PMID: 15450164
爱泼斯坦, 右. A。, Patai, 乙. Z。, Julian, J. B., & Spiers, H. J. (2017).
The cognitive map in humans: Spatial navigation and
超过. 自然神经科学, 20, 1504–1513. DOI:
https://doi.org/10.1038/nn.4656, PMID: 29073650, PMCID:
PMC6028313
爱泼斯坦, 右. A。, & Vass, L. K. (2014). Neural systems for landmark-
based wayfinding in humans. Philosophical Transactions of
the Royal Society of London, Series B, Biological Sciences,
369, 20120533. DOI: https://doi.org/10.1098/rstb.2012.0533,
PMID: 24366141, PMCID: PMC3866451
Gelman, A。, Jakulin, A。, Pittau, 中号. G。, & Su, Y.-S. (2008). A
weakly informative default prior distribution for logistic and
other regression models. Annals of Applied Statistics, 2,
1360–1383. DOI: https://doi.org/10.1214/08-AOAS191
Hannula, D. E., & Ranganath, C. (2009). The eyes have it:
Hippocampal activity predicts expression of memory in eye
movements. 神经元, 63, 592–599. DOI: https://doi.org/10
.1016/j.neuron.2009.08.025, PMID: 19755103, PMCID:
PMC2747814
Henson, 右. N。, & Gagnepain, 磷. (2010). Predictive, 交互的
multiple memory systems. Hippocampus, 20, 1315–1326.
DOI: https://doi.org/10.1002/hipo.20857, PMID: 20928831
霍纳, A. J。, Bisby, J. A。, Zotow, E., 衬套, D ., & 伯吉斯, 氮.
(2016). Grid-like processing of imagined navigation. 当前的
生物学, 26, 842–847. DOI: https://doi.org/10.1016/j.cub
.2016.01.042, PMID: 26972318, PMCID: PMC4819517
霍纳, A. J。, & 伯吉斯, 氮. (2014). Pattern completion in
multielement event engrams. 现代生物学, 24, 988–992.
DOI: https://doi.org/10.1016/j.cub.2014.03.012, PMID:
24746796, PMCID: PMC4012134
霍华德, L. R。, Javadi, A. H。, 于, Y。, Mill, 右. D ., Morrison, L. C。,
骑士, R。, 等人. (2014). The hippocampus and entorhinal
cortex encode the path and Euclidean distances to goals
during navigation. 现代生物学, 24, 1331–1340. DOI:
https://doi.org/10.1016/j.cub.2014.05.001, PMID: 24909328,
PMCID: PMC4062938
Hutton, C。, Bork, A。, Josephs, 奥。, Deichmann, R。, Ashburner, J。,
& 车工, 右. (2002). Image distortion correction in fMRI: A
quantitative evaluation. Neuroimage, 16, 217–240. DOI:
https://doi.org/10.1006/nimg.2001.1054, PMID: 11969330
雅各布, P.-Y., Casali, G。, Spieser, L。, 页, H。, Overington, D ., &
Jeffery, K. (2017). An independent, landmark-dominated
head-direction signal in dysgranular retrosplenial cortex.
自然神经科学, 20, 173–175. DOI: https://doi.org
/10.1038/nn.4465, PMID: 27991898, PMCID: PMC5274535
Joensen, 乙. H。, Gaskell, 中号. G。, & 霍纳, A. J. (2020). United we
fall: All-or-none forgetting of complex episodic events. 杂志
of Experimental Psychology: General, 149, 230–248. DOI:
460
认知神经科学杂志
体积 33, 数字 3
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
哦
C
n
_
A
_
0
1
6
5
4
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
https://doi.org/10.1037/xge0000648, PMID: 31305093,
PMCID: PMC6951107
Julian, J. B., Fedorenko, E., 韦伯斯特, J。, & Kanwisher, 氮. (2012).
An algorithmic method for functionally defining regions
of interest in the ventral visual pathway. Neuroimage, 60,
2357–2364. DOI: https://doi.org/10.1016/j.neuroimage.2012
.02.055, PMID: 22398396
Julian, J. B., Keinath, A. T。, Marchette, S. A。, & 爱泼斯坦, 右. A.
(2018). The neurocognitive basis of spatial reorientation.
现代生物学, 28, R1059–R1073. DOI: https://doi.org
/10.1016/j.cub.2018.04.057, PMID: 30205055, PMCID:
PMC6161705
Julian, J. B., Ryan, J。, 汉密尔顿, 右. H。, & 爱泼斯坦, 右. A. (2016).
The occipital place area is causally involved in representing
environmental boundaries during navigation. 当前的
生物学, 26, 1104–1109. DOI: https://doi.org/10.1016/j.cub
.2016.02.066, PMID: 27020742, PMCID: PMC5565511
Kass, 右. E., & 椽子, A. 乙. (1995). 贝叶斯因素. Journal of the
American Statistical Association, 90, 773–795. DOI: https://
doi.org/10.1080/01621459.1995.10476572
Kumaran, D ., Hassabis, D ., Spiers, H. J。, Vann, S. D ., Vargha-
Khadem, F。, & Maguire, 乙. A. (2007). Impaired spatial and
non-spatial configural learning in patients with hippocampal
pathology. Neuropsychologia, 45, 2699–2711. DOI: https://
doi.org/10.1016/j.neuropsychologia.2007.04.007, PMID:
17507060, PMCID: PMC2572909
Linde-Domingo, J。, Treder, 中号. S。, Kerrén, C。, & Wimber, 中号.
(2019). Evidence that neural information flow is reversed
between object perception and object reconstruction from
记忆. Nature Communications, 10, 179. DOI: https://
doi.org/10.1038/s41467-018-08080-2, PMID: 30643124,
PMCID: PMC6331625
Luo, X。, Roads, 乙. D ., & Love, 乙. C. (2020). The costs and
benefits of goal-directed attention in deep convolutional
神经网络. ArXiv Preprint arXiv:2002.02342. Retrieved
from http://arxiv.org/abs/2002.02342.
Mack, 中号. L。, 普雷斯顿, A. R。, & Love, 乙. C. (2013). Decoding the
brain’s algorithm for categorization from its neural
implementation. 现代生物学, 23, 2023–2027. DOI:
https://doi.org/10.1016/j.cub.2013.08.035, PMID: 24094852,
PMCID: PMC3874407
Malcolm, G. L。, Silson, 乙. H。, 亨利, J. R。, & 贝克, C. 我. (2018).
Transcranial magnetic stimulation to the occipital place area
biases gaze during scene viewing. Frontiers in Human
神经科学, 12, 189. DOI: https://doi.org/10.3389/fnhum
.2018.00189, PMID: 29867413, PMCID: PMC5953332
Marchette, S. A。, Ryan, J。, & 爱泼斯坦, 右. A. (2017). Schematic
representations of local environmental space guide goal-
directed navigation. 认识, 158, 68–80. DOI: https://土井
.org/10.1016/j.cognition.2016.10.005, PMID: 27814459,
PMCID: PMC5123926
Marchette, S. A。, Vass, L. K., Ryan, J。, & 爱泼斯坦, 右. A. (2014).
Anchoring the neural compass: Coding of local spatial
reference frames in human medial parietal lobe. 自然
神经科学, 17, 1598–1606. DOI: https://doi.org/10.1038
/nn.3834, PMID: 25282616, PMCID: PMC4309016
Marchette, S. A。, Vass, L. K., Ryan, J。, & 爱泼斯坦, 右. A. (2015).
Outside looking in: Landmark generalization in the human
navigational system. 神经科学杂志, 35, 14896–14908.
DOI: https://doi.org/10.1523/JNEUROSCI.2270-15.2015, PMID:
26538658, PMCID: PMC4635136
Milivojevic, B., Vicente-Grabovetsky, A。, & Doeller, C. F. (2015).
Insight reconfigures hippocampal–prefrontal memories.
现代生物学, 25, 821–830. DOI: https://doi.org/10.1016
/j.cub.2015.01.033, PMID: 25728693
摩纳哥, J. D ., 饶, G。, Roth, 乙. D ., & Knierim, J. J. (2014).
Attentive scanning behavior drives one-trial potentiation of
hippocampal place fields. 自然神经科学, 17, 725–731.
DOI: https://doi.org/10.1038/nn.3687, PMID: 24686786,
PMCID: PMC4036486
Motley, S. E., Grossman, 是. S。, Janssen, 瓦. G. M。, Baxter, 中号. G。,
Rapp, 磷. R。, Dumitriu, D ., 等人. (2018). Selective loss of thin
spines in area 7a of the primate intraparietal sulcus predicts
age-related working memory impairment. 杂志
神经科学, 38, 10467–10478. DOI: https://doi.org/10
.1523/jneurosci.1234-18.2018, PMID: 30355632, PMCID:
PMC6284109
奥基夫, J。, & 伯吉斯, 氮. (2005). Dual phase and rate coding
in hippocampal place cells: Theoretical significance and
relationship to entorhinal grid cells. Hippocampus, 15,
853–866. DOI: https://doi.org/10.1002/hipo.20115, PMID:
16145693, PMCID: PMC2677681
奥基夫, J。, & Dostrovsky, J. (1971). The hippocampus as a
spatial map: Preliminary evidence from unit activity in the
freely-moving rat. 脑研究, 34, 171–175. DOI: https://
doi.org/10.1016/0006-8993(71)90358-1, PMID: 5124915
Oliva, A。, & Torralba, A. (2001). Modeling the shape of the
场景: A holistic representation of the spatial envelope.
International Journal of Computer Vision, 42, 145–175.
DOI: https://doi.org/10.1023/A:1011139631724
O’Reilly, 右. C。, & Rudy, J. 瓦. (2001). Conjunctive representations
in learning and memory: Principles of cortical and hippocampal
function. 心理评论, 108, 311–345. DOI: https://
doi.org/10.1037/0033-295x.108.2.311, PMID: 11381832
公园, S。, 布雷迪, 时间. F。, Greene, 中号. R。, & Oliva, A. (2011).
Disentangling scene content from spatial boundary:
Complementary roles for the parahippocampal place area
and lateral occipital complex in representing real-world
场景. 神经科学杂志, 31, 1333–1340. DOI:
https://doi.org/10.1523/JNEUROSCI.3885-10.2011, PMID:
21273418, PMCID: PMC6623596
罗伯逊, C. E., Hermann, K. L。, Mynick, A。, Kravitz, D. J。, &
Kanwisher, 氮. (2016). Neural representations integrate the
current field of view with the remembered 360° panorama in
scene-selective cortex. 现代生物学, 26, 2463–2468. DOI:
https://doi.org/10.1016/j.cub.2016.07.002, PMID: 27618266
Schlichting, 中号. L。, Mumford, J. A。, & 普雷斯顿, A. 右. (2015).
Learning-related representational changes reveal dissociable
integration and separation signatures in the hippocampus
and prefrontal cortex. Nature Communications, 6, 8151.
DOI: https://doi.org/10.1038/ncomms9151, PMID: 26303198,
PMCID: PMC4560815
Schlichting, 中号. L。, Zeithamova, D ., & 普雷斯顿, A. 右. (2014). CA1
subfield contributions to memory integration and inference.
Hippocampus, 24, 1248–1260. DOI: https://doi.org/10.1002
/hipo.22310, PMID: 24888442, PMCID: PMC4159432
舒尔曼, G. L。, Pope, D. L. W., Astafiev, S. 五、, McAvoy, 中号. P。,
斯奈德, A. Z。, & 科尔贝塔, 中号. (2010). Right hemisphere
dominance during spatial selective attention and target
detection occurs outside the dorsal frontoparietal network.
神经科学杂志, 30, 3640–3651. DOI: https://土井
.org/10.1523/jneurosci.4085-09.2010, PMID: 20219998,
PMCID: PMC2872555
Silson, 乙. H。, Steel, A. D ., & 贝克, C. 我. (2016). Scene selectivity
and retinotopy in medial parietal cortex. Frontiers in Human
神经科学, 10, 412. DOI: https://doi.org/10.3389/fnhum
.2016.00412, PMID: 27588001, PMCID: PMC4988988
史密斯, 中号. L。, & Milner, 乙. (1981). The role of the right hippocampus
in the recall of spatial location. Neuropsychologia, 19, 781–793.
DOI: https://doi.org/10.1016/0028-3932(81)90090-7, PMID:
7329524
Tolman, 乙. C. (1948). Cognitive maps in rats and men. Psychological
审查, 55, 189–208. DOI: https://doi.org/10.1037/h0061626,
PMID: 18870876
Tzourio-Mazoyer, N。, Landeau, B., Papathanassiou, D ., Crivello,
F。, Etard, 奥。, Delcroix, N。, 等人. (2002). Automated anatomical
Berens, Joensen, 和霍纳
461
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
哦
C
n
_
A
_
0
1
6
5
4
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
labeling of activations in SPM using a macroscopic anatomical
parcellation of the MNI MRI single-subject brain. Neuroimage,
15, 273–289. DOI: https://doi.org/10.1006/nimg.2001.0978,
PMID: 11771995
Vallortigara, G。, & 罗杰斯, L. J. (2005). Survival with an
asymmetrical brain: Advantages and disadvantages of cerebral
lateralization. Behavioral and Brain Sciences, 28, 575–589.
DOI: https://doi.org/10.1017/s0140525x05000105, PMID:
16209828
van Kesteren, 中号. 时间. R。, Ruiter, D. J。, Fernández, G。, & Henson,
右. 氮. (2012). How schema and novelty augment memory
形成. Trends in Neurosciences, 35, 211–219. DOI:
https://doi.org/10.1016/j.tins.2012.02.001, PMID: 22398180
Vass, L. K., & 爱泼斯坦, 右. A. (2013). Abstract representations of
location and facing direction in the human brain. 杂志
神经科学, 33, 6133–6142. DOI: https://doi.org/10.1523
/JNEUROSCI.3873-12.2013, PMID: 23554494, PMCID:
PMC3656495
沃森, D. M。, Hartley, T。, & Andrews, 时间. J. (2017). Patterns of
response to scrambled scenes reveal the importance of visual
properties in the organization of scene-selective cortex.
Cortex, 92, 162–174. DOI: https://doi.org/10.1016/j.cortex
.2017.04.011, PMID: 28499144
Welch, 乙. L. (1947). The generalization of “student’s” problem
when several different population variances are involved.
Biometrika, 34, 28–35. DOI: https://doi.org/10.1093/biomet
/34.1-2.28, PMID: 20287819
Zeithamova, D ., Dominick, A. L。, & 普雷斯顿, A. 右. (2012).
Hippocampal and ventral medial prefrontal activation during
retrieval-mediated learning supports novel inference. 神经元,
75, 168–179. DOI: https://doi.org/10.1016/j.neuron.2012
.05.010, PMID: 22794270, PMCID: PMC3398403
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
3
4
4
5
1
8
6
2
5
9
2
/
j
哦
C
n
_
A
_
0
1
6
5
4
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
462
认知神经科学杂志
体积 33, 数字 3