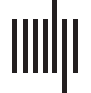SPECIAL ISSUE:
The Role of the Cerebellum in Language Comprehension and Production
Language and the Cerebellum: 结构性
Connectivity to the Eloquent Brain
Katie R. Jobson1
, Linda J. Hoffman1, Athanasia Metoki2
Anthony S. Dick3, Jamie Reilly1,4, and Ingrid R. Olson1
, Haroon Popal1
,
开放访问
杂志
1心理学系, Temple University, 费城, 宾夕法尼亚州, 美国
2神经内科, Washington University in St. 路易斯, 英石. 路易斯, Missouri, 美国
3心理学系, Florida International University, Miami, Florida, 美国
4Department of Speech and Language Sciences, Temple University, 费城, 宾夕法尼亚州, 美国
关键词: cerebellum, diffusion imaging, 额叶, human connectome project, 语言,
tractography, white matter
抽象的
Neurobiological models of receptive language have focused on the left-hemisphere
perisylvian cortex with the assumption that the cerebellum supports peri-linguistic cognitive
processes such as verbal working memory. The goal of this study was to identify language-
sensitive regions of the cerebellum then map the structural connectivity profile of these
地区. Functional imaging data and diffusion-weighted imaging data from the Human
Connectome Project (HCP) were analyzed. We found that (A) 工作记忆, motor activity,
and language comprehension activated partially overlapping but mostly unique subregions of
the cerebellum; (乙) the linguistic portion of the cerebello-thalamo-cortical circuit was more
extensive than the linguistic portion of the cortico-ponto-cerebellar tract; (C) there was a
frontal-lobe bias in the connectivity from the cerebellum to the cerebrum; (d) there was some
degree of specificity; 和 (e) for some cerebellar tracts, individual differences in picture
identification ability covaried with fractional anisotropy metrics. These findings yield insights
into the structural connectivity of the cerebellum as relates to the uniquely human process of
language comprehension.
介绍
One of the most controversial issues in cognitive neuroscience today is whether the cerebel-
lum deserves a seat at the table of the core language network. Neuromotor control of speech,
which is important for both comprehension and production of language (Fischer & Zwaan,
2008), can be affected by cerebellar damage, with ataxic dysarthria being a common outcome
of cerebellar damage (Spencer & Slocomb, 2007). 然而, there is growing evidence sug-
gesting the cerebellum’s involvement in language goes beyond its well-established role in the
motor aspects of speech and comprehension (Mariën et al., 2014). The most compelling find-
ings come from studies of individuals with damage to the posterior cerebellum and a disorder
known as cerebellar cognitive affective syndrome. Individuals with this disorder have disrup-
tions in many cognitive functions, including features of language (Schmahmann & Sherman,
1998). Prior work has found that individuals with this disorder often produce agrammatic lan-
guage output (De Smet et al., 2007; Mariën et al., 2000; Silveri et al., 1994), with concurrent
deficits in sentence comprehension (De Smet et al., 2007; Murdoch & Whelan, 2007; Silveri,
引文: Jobson, K. R。, Hoffman, L. J。,
Metoki, A。, Popal, H。, 迪克, A. S。, Reilly,
J。, & 奥尔森, 我. 右. (2022). 语言和
the cerebellum: 结构连接
to the eloquent brain. Neurobiology of
语言. Advance publication.
https://doi.org/10.1162/nol_a_00085
DOI:
https://doi.org/10.1162/nol_a_00085
支持信息:
https://doi.org/10.1162/nol_a_00085
已收到: 13 四月 2022
公认: 10 十月 2022
利益争夺: 作者有
声明不存在竞争利益
存在.
通讯作者:
Ingrid R. 奥尔森
iolson@temple.edu
处理编辑器:
Julie Fiez
版权: © 2022
麻省理工学院
在知识共享下发布
归因 4.0 国际的
(抄送 4.0) 执照
麻省理工学院出版社
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
d
哦
我
/
我
.
/
/
1
0
1
1
6
2
n
哦
_
A
_
0
0
0
8
5
2
0
5
8
2
9
1
n
哦
_
A
_
0
0
0
8
5
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
Cerebellar white matter and language
2021), anomia as defined by deficits in word generation (De Smet et al., 2007; Mariën et al.,
2000), and diminished verbal fluency (Hoche et al., 2018; Silveri, 2021). Although linguistic
and motoric deficits are often co-morbid in acquired cerebellar injuries, these two domains are
also dissociable (Ahmadian et al., 2019). The cerebellum has, 然而, proven to be an enig-
matic lesion model. 例如, people with acquired cerebellar injuries tend to experience
rapid recovery of speech and language functions, with frank symptoms only apparent during
the acute phase in adults (Fabbro et al., 2004).
Neuroimaging studies have supplemented and extended patient-based case studies. Recent
studies have reported that language comprehension activates Lobule IV, Crus I & 二, and Lobule
IX in the posterior cerebellum (Geva et al., 2021; Vias & 迪克, 2017), with a rightward cere-
bellar bias, due to the fact that cortico-cerebellar-cortical connections are crossed (Bostan
等人。, 2013). What is the computational role of this region in language? Several hypotheses
have been offered. One idea is that the cerebellum modulates timing and sequencing of
language production and language perception (Fiez, 2016; Ivry & Keele, 1989; Leggio &
Molinari, 2015; Mariën et al., 2014; Mariën & Borgatti, 2018; Molinari et al. 1997; Molinari Q1
,
2015; Salman, 2002; Schwartze & Kotz, 2016). Another hypothesis is that the cerebellum is
involved in automatizing a range of behaviors, including language (Doyon et al., 1998; Fiez,
2016; Ramnani, 2014; Vicari et al., 2018; 杨等人。, 2014). A final hypothesis is that the
cerebellum’s role in language is related to the more general role of the cerebellum in verbal
工作记忆 (陈 & 德斯蒙德, 2005; Desmond et al., 1997; Marvel & 德斯蒙德, 2010;
Peterburs et al., 2021).
D
哦
w
n
哦
A
d
e
d
A
r
t
我
C
e
–
p
d
e
d
你
n
哦
F
r
哦
米
H
d
我
r
e
C
t
.
d
哦
米
:
/
/
我
t
.
p
t
t
F
/
我
/
我
/
/
我
我
我
/
/
.
1
0
1
1
6
2
n
哦
_
A
_
0
0
0
8
5
2
0
5
8
2
9
1
n
哦
_
A
_
0
0
0
8
5
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
The comprehensive mapping of cortico-cerebellar connectivity can provide a framework
upon which to understand function. 进一步来说, rather than understanding the role of
the cerebellum in language more broadly, the mapping of the pathways that connect
language-specific regions in the cerebellum and cerebral language regions may show some
specificity. If this connectivity can be shown to be functionally specialized for particular lin-
guistic subdomains, it may show more specifically how the cerebellum contributes to each
linguistic subdomain. For more than a hundred years it has been recognized that certain
white matter tracts play a key role in language (Dejerine & Dejerine-Klumpke, 1895,
1901; Miraillé, 1896). Modern diffusion-weighted imaging (DWI) methods have allowed
investigators to identify structural networks in the cerebrum essential for language (迪克 &
Tremblay, 2012; Duffau, 2015; Krestel et al., 2013; Middlebrooks et al., 2017; Smits et al.,
2014). Whether cerebral language networks are structurally connected with portions of the
cerebellum activated in language tasks is not known. Although functional connectivity
studies have shown correlated activity between linguistically sensitive cortex and regions
of the posterior cerebellum (reviewed in Vias & 迪克, 2017) this method is far removed from
“ground truth” evidence provided by postmortem tract tracing. 的确, findings from func-
tional connectivity often disagree with findings from structural connectivity (for an example,
see Metoki et al., 2021).
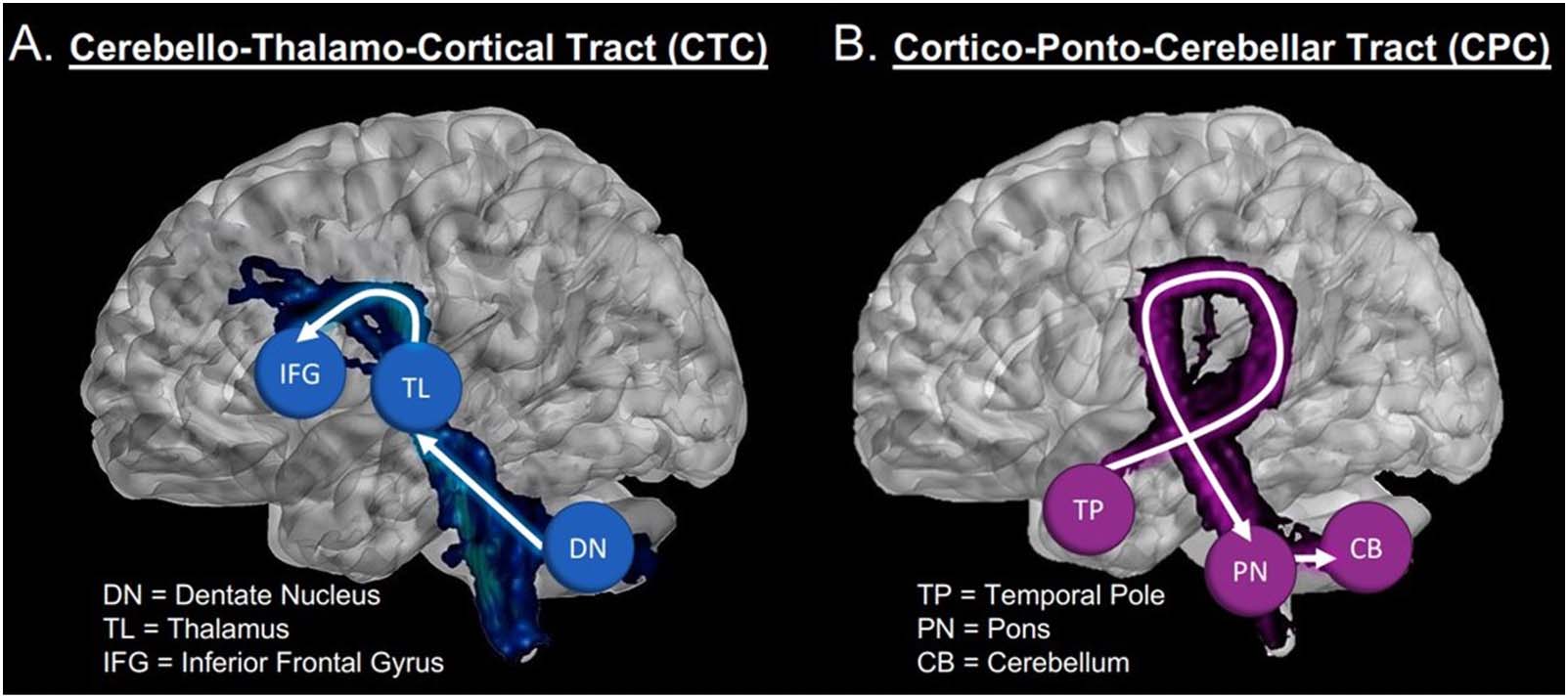
Findings from tract tracing have shown that cerebellar connectivity to and from the
cerebrum is uniquely defined by two major pathways. The cortico-ponto-cerebellar pathway
projects from the cerebral hemispheres with decussation at the pons terminating in the con-
tralateral cerebellar cortex. 相比之下, the cerebello-thalamo-cortical pathway begins in the
cerebellum then crosses over to synapse on the contralateral thalamus, continuing on to
different regions of the cerebral cortex. The dentate nucleus of the cerebellum retains a topo-
graphically ordered pattern of connectivity (Dum & 针织, 2003; Palesi et al., 2021; Steele
等人。, 2017). It is presumed that this connectivity remains orderly throughout the cerebello-
thalamo-cortical and cortico-ponto-cerebellar pathway tracts.
Neurobiology of Language
2
Cerebellar white matter and language
纤维束成像术:
The act of measuring white matter
voxel-by-voxel using various
方法, producing reconstructions
of pathways.
The goal of this study is to use DWI to help disentangle the role of the cerebellum in
language comprehension. DWI provides information about what brain regions are communi-
cating with each other via a mathematical model of axonal connectivity. Information about
axonal connectivity can be used to determine which facets of language are mediated by
the cerebellum.
We used a tractography pipeline that we previously developed for studying the role of the
cerebellum in theory of mind (Metoki et al., 2021). We applied this pipeline to language
regions of interest (ROIs) in the cerebellum and cerebrum. Cerebellar language ROIs were
derived from the sentence comprehension task in the Human Connectome Project (HCP)
task-fMRI data set (Binder et al., 2011). Cerebral ROIs were chosen a priori based on the
sentence comprehension literature (Barch et al., 2013; Binder et al., 2011; Booth et al.,
2007; Fengler et al., 2016; Friederici, 2002; Friederici et al., 2003; Keller et al., 2001; Kieren
& 巴克纳, 2009; Rogalsky et al., 2008). We focused on language comprehension, 而不是
overt language production, with the goal of reducing the potential impact of motor processing.
Because language tends to be strongly left lateralized in the cerebrum (Frost et al., 1999;
Mariën et al., 2001; Takaya et al., 2015) and the cerebellum has crossed structural connectivity
with the cerebrum (Gonzalo-Ruiz & Leichnetz, 1990; Ito et al., 1986; 凯莉 & 针织, 2003), 我们
performed lateralized tractography between language-sensitive ROIs in the right cerebellum to
language-sensitive ROIs in the left cerebrum. Based on prior DWI work (Metoki et al., 2021) 作为
well as monkey histology work (Glickstein et al., 1985; Schmahmann, 1996) we predicted that
there would be significantly more fibers to linguistically sensitive frontal cerebral ROIs, 像
left inferior frontal gyrus (IFG), as compared to linguistically sensitive ROIs in the temporal
叶.
材料和方法
Data Set and Participants
All data used in this study are part of the HCP data set, specifically the WU-Minn HCP Con-
sortium S900 Release (WU-Minn HCP Consortium, 2015). This data set is publicly available
and accessible at https://www.humanconnectome.org. Only participants that completed all
imaging sessions of interest (T1/T2, task fMRI), and DWI scans were included in this study.
We restricted our population to only right-handed subjects using the Edinburgh Handedness
questionnaire (Oldfield, 1971). We chose a random subset of 100 参与者 (50 女性,
M age = 27.89 yr, 标准差= 3.9 yr), as using a large sample size incurs a computational cost, 给定
that probabilistic tractography is mathematically intensive. Unless otherwise stated, all signif-
icant results reported in this study were corrected for multiple comparisons using the false
discovery rate correction (Benjamini & Hochberg, 1995).
Overview of HCP Behavioral Protocol
While known for its neuroimaging data, the HCP protocol also includes several behavioral
assessments conducted outside of the scanner. The NIH Toolbox for Assessment of Neurolog-
ical and Behavioral function was of most interest to us, in particular their tasks related to
language comprehension. The NIH Toolbox Picture Vocabulary Test, a measure of receptive
语言, was our dependent behavioral measure. This particular version of the task was
adaptive, which allowed for more variation in the data (for additional detail see Gershon
等人。, 2014). Note that fMRI tasks are generally designed to produce strong blood oxygen level
dependent (大胆的) signals but often have very little variance in the resultant behavioral out-
来了. The HCP behavioral tasks used outside of the scanner elicited higher variance in the
Neurobiology of Language
3
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
d
哦
我
/
我
.
/
/
1
0
1
1
6
2
n
哦
_
A
_
0
0
0
8
5
2
0
5
8
2
9
1
n
哦
_
A
_
0
0
0
8
5
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
Cerebellar white matter and language
Diffusion imaging:
An MRI scan that measures the
diffusivity of water within the brain—
this method allows us to approximate
white matter pathways.
behavioral outcomes, making them potentially more sensitive for analyses of individual
差异.
Overview of HCP fMRI Protocol
A detailed description of the HCP data acquisition and preprocessing pipelines can be found
别处 (Barch et al., 2013; Glasser et al., 2013; 史密斯等人。, 2013; Van Essen et al., 2012).
Briefly, the HCP protocol includes acquisition of structural MRI, task-state fMRI, diffusion MRI,
and extensive behavioral testing. The imaging data used in this article are the “minimally pre-
processed” subjects included in the WU-Minn HCP Consortium S900 Release (WU-Minn HCP
Consortium, 2015). This includes standard preprocessing using TOPUP, EDDY, and BED-
POSTX. Details of imaging protocols, preprocessing pipelines, and in-scanner task protocols
can be found in the Supporting Information, which can be found at https://doi.org/10.1162/nol
_a_00085.
Task-state fMRI encompasses seven major domains, three of which were used in this study:
(1) working memory/cognitive control systems; (2) motor (visual, motion, somatosensory, 和
motor systems); 和 (3) language comprehension. The main task of interest was the language
任务 (Binder et al., 2011). Participants in the language task listened to stories adapted from
Aesop’s fables (加州. fourth century B.C.E./2012). Sentences were read aloud by a text-to-speech
program to participants in the scanner. After listening to the stories, the participants were pre-
sented with a two-answer forced choice question about the contents of the story. The question
was meant to probe understanding about the theme of the story, thus evoking activations
related to comprehension (Binder et al., 2011). Participants selected one of the two answers
by pushing a button. Specific details about the contents of the stories such as number of
事件, number of actors, mean sentence length, and duration are described in detail by Binder
and colleagues (2011). The accompanying control task involved participants doing math prob-
莱姆斯. The design of this task was the same as the language task, with the same text-to-speech
method used to present the stimuli. Rather than listening to a story, participants were read math
problems aloud (“six times two equals…”). Participants were presented with a mathematical
problem and were asked a two-answer forced choice question. The difficulty of the math task
was increased after six correct responses and decreased in difficulty after one incorrect
response. Descriptions of the working memory and motor tasks, used in the overlap and
control analyses, are further described in the Supporting Information.
The results of the language fMRI task in the scanner did not yield enough variance to
examine its potential relationship with our white matter pathways. 反而, we included
participants’ performance on the NIH Toolbox Picture Vocabulary Test as our dependent
behavioral measure.
Regions of Interest
We used two sets of ROIs for our analyses: one set in the cerebrum and one in the cerebellum.
Because language is strongly lateralized, we limited our ROIs to the left cerebral hemisphere
and to the right cerebellum. The set of cerebral ROIs were drawn from prior work on language.
The cerebral ROIs included the following: angular gyrus (ANG; Fengler et al., 2016; 凯勒
等人。, 2001; Van Ettinger-Veenstra et al., 2016), dorsolateral prefrontal cortex (DLPFC; 凯勒
等人。, 2001; Kieren & 巴克纳, 2009), superior temporal gyrus (STG; Barch et al., 2013;
Binder et al., 2011; Booth et al., 2007; Fengler et al., 2016; Friederici et al., 2003; 凯勒
等人。, 2001; Turken & Dronkers, 2011), middle temporal gyrus (MTG; Binder et al., 2011;
Keller et al., 2001; Turken & Dronkers, 2011; Van Ettinger-Veenstra et al., 2016), inferior
Neurobiology of Language
4
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
d
哦
我
/
我
.
/
/
1
0
1
1
6
2
n
哦
_
A
_
0
0
0
8
5
2
0
5
8
2
9
1
n
哦
_
A
_
0
0
0
8
5
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
Cerebellar white matter and language
temporal gyrus (ITG; Ikuta et al., 2006), temporal pole (TP; Barch et al., 2013; Binder et al.,
2011), IFG/Broca’s area (Barch et al., 2013; Binder et al., 2011; Booth et al., 2007; 货车
Ettinger-Veenstra et al., 2016), and posterior superior portion of the temporal lobe (PST; Just
等人。, 1996; Mesulam et al., 2015). This area is often considered to be synonymous with Wer-
nicke’s area; 然而, because there is a lack of consensus regarding its location in the field
(Tremblay & 迪克, 2016), throughout this article we will refer to this area as the PST. 虽然
other brain areas also support language, we focused on regions that have been consistently
implicated in sentence comprehension. Coordinates for cerebral ROIs were taken from Neu-
rosynth (https://neurosynth.org/; Yarkoni et al., 2011) by searching for the name of the region
under “terms” and taking the voxel with the highest z-score using a cluster analysis. 使用
term superior posterior produced an activation map closest to the agreed-upon anatomical
location of Wernicke’s area (Tremblay & 迪克, 2016). 很遗憾, the highest z-scored
voxel produced under the term posterior superior was too posterior for the aims of this study.
反而, we took the highest z-scored voxel that was also close enough to Heschl’s gyrus as
justification for using it to represent the PST. See Table A in the Supporting Information for
exact MNI (Minnesota Neurological Institute) coordinates and accompanying z-scores for
each ROI. 随后, each coordinate was then transformed into a 6 mm sphere using
FSL software (Jenkinson et al., 2012), to be used in the analysis.
For the ROIs in the cerebellum, we used a different apporach. Although several studies
have attempted to functionally map the cerebellar cortex (Buckner et al., 2011; Diedrichsen
& Zotow, 2015; Guell, Gabrieli, 等人。, 2018; Guell, Schmahmann, 等人。, 2018; 金等人。,
2019; Kieren & 巴克纳, 2009; Marek et al., 2018; Riedel et al., 2015), there is no consensus
about functional boundaries within the cerebellum. 因此, it was impossible to employ the
same approach we used with the predefined cerebral ROIs. 反而, we used a data-driven
方法. Following the method used by Guell, Gabrieli, 等人. (2018), we transformed FSL’s
等级 2 individual cope files (results of within-subject fixed-effects grayordinate-based analyses
that generate output files that index mean effects for an individual subject averaged across the
two scan runs for a task) into Cohen’s d group maps by first transforming the grayordinate.
dscalar.nii files to NIfTI. We then used FSL commands fslselectvols, to extract the contrast
of interest “story > math” for each individual, and fslmerge, fslmaths -Tmean, -Tstd, 和
-div to merge the individual contrast images, extract the mean and the standard deviation,
and divide the two, ultimately getting group Cohen’s d maps for the contrasts “story > math”
(语言), “2-back > 0-back” (工作记忆), and “average” (motor) based on a larger
sample of 671 subjects. The HCP S900 Release provides level 3 group z-maps, but Cohen’s
d maps made it possible to observe the effect size of each task contrast rather than the signif-
icance of the BOLD signal change. A sample of 671 subjects ensures that a d value higher than
0.5 (科恩, 1988) will be statistically significant even after correction for multiple compari-
儿子们 (d = z/sqrt(n), d > 0.5 we have z > 12.95 for N = 671; analysis of 17,853 cerebellar voxels
would require p < 0.000028 after Bonferroni correction, and p < 0.000028 is equivalent to z >
4.026). 因此, we used FSL’s cluster tool, the Cohen’s d maps, and a threshold of 0.5 到
extract clusters of activation for each task and local maxima within each cluster. After using a
whole cerebellar mask to retain only the clusters and local maxima within the right cerebel-
卢姆, clusters smaller than 100 mm3 were further removed to omit very small clusters that were
considered to be noninformative and would make a comprehensive description of the results
too extensive (见图 1 for a visualization of the three functional tasks). The coordinates of
the remainder local maxima from the “story > math” (语言) contrast within the cerebellum
were used to create two group cerebellar ROIs (spheres, 6 mm radius) in the right hemisphere.
This resulted in two language cerebellar ROIs: right Crus I and right Lobule IX. 相同
Local maxima:
In regard to fMRI data, this refers to
the voxel that has the highest amount
of activity in a brain region of
兴趣.
Neurobiology of Language
5
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
d
哦
我
/
我
/
.
/
1
0
1
1
6
2
n
哦
_
A
_
0
0
0
8
5
2
0
5
8
2
9
1
n
哦
_
A
_
0
0
0
8
5
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
Cerebellar white matter and language
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
d
哦
我
/
我
/
/
.
1
0
1
1
6
2
n
哦
_
A
_
0
0
0
8
5
2
0
5
8
2
9
1
n
哦
_
A
_
0
0
0
8
5
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
数字 1. Functional activation to language tasks in the cerebellum. (A) Functional activations from this study to three tasks (语言, 工作-
ing memory, and motor) displayed on a flatmap of the cerebellum. Only activations within the right cerebellar hemisphere are displayed; (乙) A
figure from King et al. (2019) depicting cerebellar activations to a range of cognitive and motor tasks. Note that there is overlap between their
language activations (labeled primarily as numbers 7 和 8) and our language activations in (A). (C) A figure from LeBel et al. (2021) inves-
tigating sentence-level and word-level semantic language processing. (D) A figure from Guell, Schmahmann et al. (2018) displaying cerebellar
activations to a range of cognitive and motor tasks from the HCP data set.
Neurobiology of Language
6
Cerebellar white matter and language
method was used to extract the cerebellar motor and working memory cerebellar ROIs, 哪个
were used in overlap and control analyses. Seven working memory ROIs were created, 也在
the right cerebellar hemisphere. Four of the seven cerebellar working memory ROIs over-
lapped with each other, so they were removed from the analysis, leaving three working mem-
ory ROIs in the cerebellum. The remaining cerebellar working memory ROIs were primarily
located in Lobule IV, Crus I, and Lobule VIIb.
Cluster Overlap and Euclidean Distances
Given that the HCP data set uses FNIRT (安德森, 2010) registration
to the MNI template,
we calculated the percentage of overlap of each cerebellar cluster by using Diedrichsen’s
FNIRT MNI maximum probability map (Diedrichsen et al., 2009). Previous studies have
already explored the overlap of several other tasks included in the HCP data set (Metoki
等人。, 2021). 因此, we only included three tasks in this analysis: 语言, working mem-
奥里, and motor. We used FSL’s atlasq tool to determine the percent overlap of each cerebellar
cluster in the cerebellar lobes, hence determining the primary location of each cluster. 这
Sørensen–Dice coefficient, which is a statistic measuring the similarity of two samples (Dice,
1945; Sørensen, 1948), was then used to calculate the percentage of overlap between the
functional clusters generated from all tasks and determine their similarity, and Euclidean dis-
tances were calculated to estimate the distances of local maxima within and between clusters.
At the individual level, we thresholded z-scored β-weights of each subject’s activation map
for each task contrast to > 0 to retain only increased activation during the tasks and then ran
Wilcoxon signed-rank tests between each task pair to examine whether there was a statistical
difference between them.
Diffusion Analyses
All diffusion analyses were completed on Temple University’s High-Performance Computing
Cluster, OwlsNest. Probabilistic tractography analyses were performed using FSL’s prob-
trackx2 (probabilistic tracking with crossing fibres, Version 6.0.2; Behrens et al., 2003,
2007) in each subject’s native space. Due to HCP preprocessing steps, this native space we
refer to is the subjects’ T1w space. After tractography, results were then transformed to MNI
standard space using transformation matrices (see Supporting Information for more details). 一个
ROI-to-ROI approach was used with cerebral and cerebellar ROIs used as seeds and targets to
reconstruct each subject’s cerebello-cerebral white matter connections. Fiber tracking was ini-
tialized in both directions separately (from seed to target and vice versa) 和 5,000 streamlines
were drawn from each voxel in each ROI. Tract length correction was also used, as cerebellar
tracts are lengthy due to their polysynaptic nature, and length of a tract can introduce more
false positives to the data (琼斯, 2010). Tractographies were performed to delineate the
cerebello-thalamo-cortical pathway, which projects from the cerebellar cortex to the deep
cerebellar nuclei then crosses over to synapse on the contralateral thalamus, continuing on
to different regions of the cerebral cortex (Middleton & 针织, 1997; Palesi et al., 2015;
Schmahmann & Pandya, 1997) and the cortico-ponto-cerebellar pathway, which projects from
the cerebral hemispheres to the pons, then to the contralateral cerebellar cortex (Palesi et al.,
2017; Ramnani, 2006). For the cerebello-thalamo-cortical tractographies, a binarized mask of
the superior cerebellar peduncle in MNI space from the Johns Hopkins University ICBM-DTI-
81 white-matter labels atlas (Hua et al., 2008; Mori et al., 2005; Wakana et al., 2007) and left
thalamus in MNI space from the Harvard-Oxford subcortical atlas (Desikan et al., 2006;
Frazier et al., 2005; Goldstein et al., 2007; Makris et al., 2006) were used as waypoints respec-
主动地. The binarized contralateral cerebellar and cerebral hemispheres, lobes of non-interest,
Neurobiology of Language
7
Q2
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
d
哦
我
/
我
/
.
/
1
0
1
1
6
2
n
哦
_
A
_
0
0
0
8
5
2
0
5
8
2
9
1
n
哦
_
A
_
0
0
0
8
5
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
Cerebellar white matter and language
and the opposing cerebellar peduncle were set as exclusion masks. All cerebral masks were
created using the Harvard-Oxford cortical atlas (Desikan et al., 2006; Frazier et al., 2005;
Goldstein et al., 2007; Makris et al., 2006). 例如, cerebello-thalamo-cortical tractogra-
phy between the right cerebellum and left DLPFC included the right superior cerebellar
peduncle and left thalamus as waypoints. The exclusion mask was comprised of the left cer-
ebellar hemisphere, right cerebral hemisphere, middle cerebellar peduncle, precentral gyrus,
as well as the temporal, 顶叶, and occipital lobes. For the cortico-ponto-cerebellar pathway
tractographies, a binarized mask of the middle cerebellar peduncle in MNI space from the
same atlas was used as a waypoint. The contralateral cerebellar and cerebral hemispheres,
right superior cerebellar peduncle, and lobes of non-interest were used as exclusion masks.
例如, cortico-ponto-cerebellar pathway tractography between a left temporal lobe
ROI and right cerebellar ROI would entail the middle cerebellar peduncle waypoint mask.
The exclusion mask included the left cerebellar hemisphere, right cerebral hemisphere, 正确的
superior cerebellar peduncle, precentral gyrus, and frontal, 顶叶, and occipital lobes. Exact
regions included in each exclusion mask can be found in Table B in the Supporting
信息. The pons was not selected as an inclusion mask because of its absence from
any standardized atlases. Despite not having the pons as an orthogonal waypoint in the
cortico-ponto-cerebellar pathway tract, we did include the thalamus in the cerebello-thalamo-
cortical reconstruction to follow previous literature (Palesi et al., 2015) and to ensure our
tractography was as anatomically similar to the ground truth as possible. Our exclusion masks
were comprehensive, as we had certain expectations as to where the tract would be traveling
and terminating (席林等人。, 2020). For cerebral ROIs in the frontal lobe, we knew that we
were not interested in fibers that extended into other regions, such as the parietal or temporal
lobes; therefore we incorporated them in the exclusion mask for that ROI. We also excluded
the motor cortex with the knowledge that there are tracts from the cerebellum to the motor
cortex. Exclusion of the motor cortex ensured our results were not due to connections between
cerebral motor regions and our ROIs in the cerebellum.
Two metrics were extracted to be used as dependent measures in our analyses. Volume was
extracted from the streamline density map using FSL’s fslstats and normalized using intracra-
nial volume (ICV; Voevodskaya et al., 2014). Intracranial volume was calculated using the
method of Malone et al. (2015). This involves isolating the skull-stripped T1-weighted image
provided by HCP in the same native space as the diffusion data and segmenting the brain using
SPM12 into gray matter, white matter, and cerebral spinal fluid (in liters). The summed
volumes of these tissue types yield the whole-brain intracranial volume, which when trans-
formed into mm3 yields the metric for probabilistic tractography reported by FSL. 此外,
fractional anisotropy (F A) was extracted from respective cerebellar peduncles and included as
a dependent variable. This was accomplished by taking whole-brain FA scalar data in native
空间 (the result of the command dtifit, which is run on eddy-corrected data) and using each of
the peduncles to mask the data. This produced FA maps exclusively within each peduncle.
That data was then transformed into standard space, which allowed us to extract microstruc-
tural information of each tract within each peduncle. We chose to extract microstructural
indices from only the peduncles following previous literature, as the peduncles are generally
a good point in the tract to evaluate white matter cohesion (Jossinger et al., 2021; Taoka et al.,
2007; 王等人。, 2003). Selecting only one region along the resulting tractography was
expected to reduce some of the noise that would be introduced at the termination of the tract,
which is a concern for such lengthy fiber pathways. This also allowed us to isolate the con-
tribution of the cerebellar pathways in behavior, without picking up on cerebral white matter
pathways (such as the arcuate fasciculus).
Neurobiology of Language
8
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
d
哦
我
/
我
/
/
.
1
0
1
1
6
2
n
哦
_
A
_
0
0
0
8
5
2
0
5
8
2
9
1
n
哦
_
A
_
0
0
0
8
5
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
Cerebellar white matter and language
结果
Functional Domains in the Cerebellum
We first examined whether there was overlap between language, 工作记忆, and motor
activations in the right cerebellum. We found that there was a significant difference in local-
izations for all three task pairs. Overall motor activations were localized to the anterior cere-
战争 (Lobules I–IV), with some activation in the posterior cerebellum (Lobules VIIB and
VIIIA). Overall language activations were localized to Crus I/II and Lobule IX. Overall working
memory activations were found in Crus I/II, Lobule VI, Lobule VIIb, and Lobule VIIIa. 这
β-weights of each subject’s activation map that were previously extracted, we performed
Wilcoxon signed-rank tests for each task pair to examine whether there was a statistical
difference between them. The language and motor activations had no overlap, 而
language and working memory functional activations had minimal overlap (0.005%; 看
桌子 1). After conducting the analyses on the overall activations from these tasks, 峰值
activation was extracted from the group activation maps and transformed into 6 mm spheres in
FSL to be used in subsequent analyses.
Cerebellar Structural Connections: Language–Language
下一个, we asked whether language-sensitive regions in the cerebellum, which are poorly
understood and have received little attention, are structurally connected to language-sensitive
areas in the cerebrum that have been studied for over a century. We ran probabilistic tracto-
graphy to reconstruct the cerebello-thalamo-cortical and cortico-ponto-cerebellar pathway
white matter pathways between ROIs in the right cerebellum and left cerebral cortex. 体积
was extracted for the cerebello-thalamo-cortical and cortico-ponto-cerebellar pathways and
F A (microstructure) was extracted for the superior (cerebello-thalamo-cortical tracts) or middle
(cortico-ponto-cerebellar tract) peduncles.
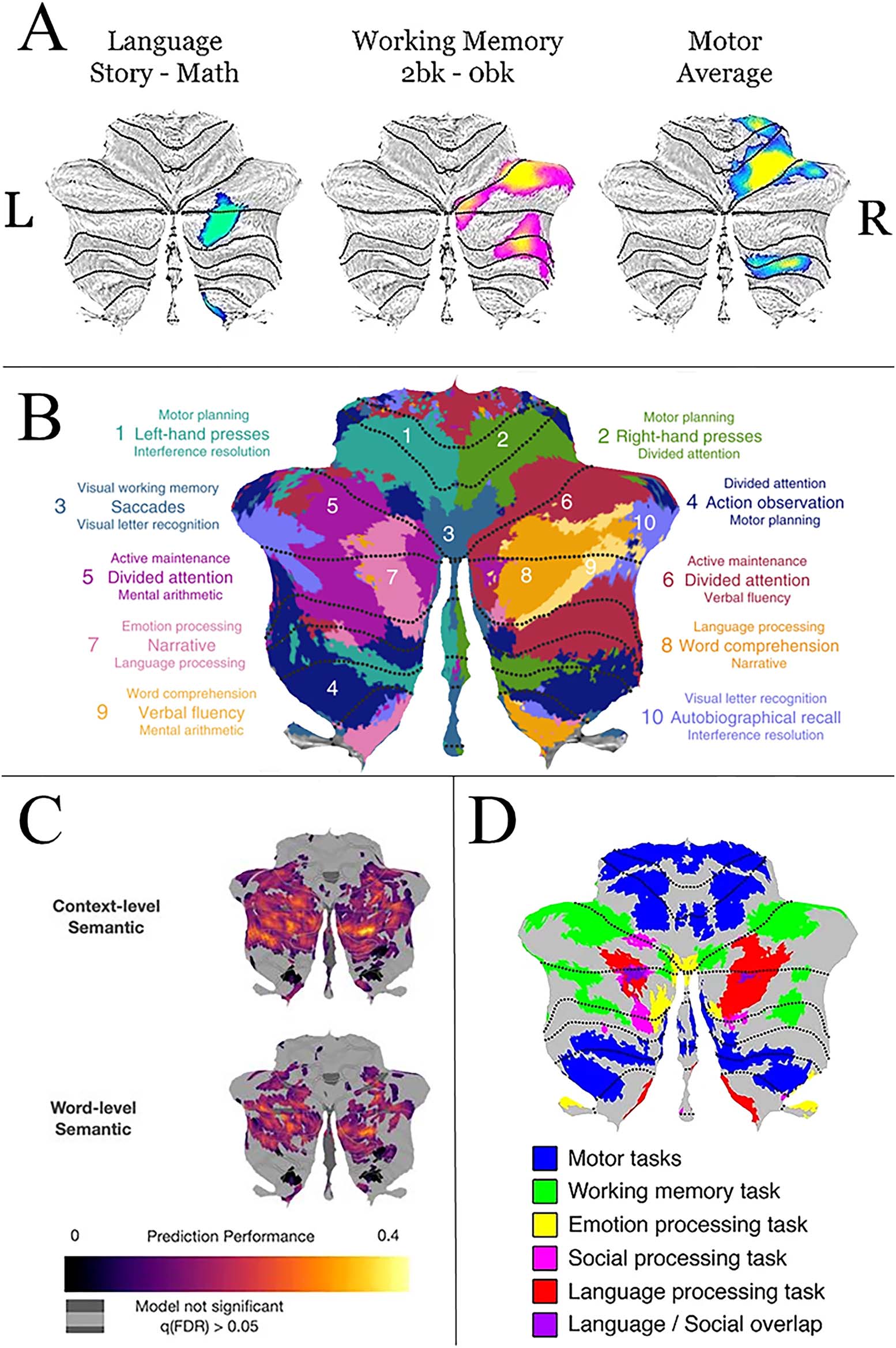
The average volume for tracts (before correction for intracranial volume) ranged from
74,714.32 mm to 240,083.92 毫米. There were significant differences between the
cerebello-thalamo-cortical and cortico-ponto-cerebellar pathways with lower volume in the
cortico-ponto-cerebellar pathways (见表 2 和图 2; W = 8,614, p < 0.001). For
the cerebello-thalamo-cortical pathway, we examined whether there was a difference in the
average volume between each cerebellar ROI seed (Crus I and Lobule IX). For this analysis, we
intended to isolate the average volume when a tract was seeded in a cerebellar region to iden-
tify if one seed had a relatively greater volume of tracts being sent to the cerebrum. We accom-
plished this by taking each tract seeded in its respective cerebellar ROI to each of the eight
cerebral targets (IFG, DLFPC, ANG, TP, STG, MTG, PST, ITG) and averaging the numbers.
Using a paired Wilcoxon signed-rank test, we found no significant difference in tract volume
Table 1.
for the tasks of interest in the right cerebellar hemisphere.
The Sørensen–Dice coefficient (in percentages) between the three functional activations
fMRI task activation
Language
Working memory
Motor
Language
Working memory
Motor
0.0051
0
0.1507
Note. All three tasks showed very little overlap. Representation of the activations is depicted in Figure 1.
Neurobiology of Language
9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
.
/
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Cerebellar white matter and language
Table 2.
cortical (CTC) connections to language-related cortical regions.
A summary comparison of the volumes and standard deviations for the cortico-ponto-cerebellar (CPC) and cerebello-thalamo-
CTC (seeded in Crus I)
CPC (seeded in Crus I)
Uncorrected Vol.
Language ROI
STG
Mean
150664.72
SD
55440.85
ICV Corrected Vol.
SD
Mean
0.04
0.10
MTG
ITG
ANG
TP
IFG
DLPFC
PST
150863.84
55557.13
150506.24
55179.80
150000.88
54587.71
150073.92
55120.73
226984.88
74864.76
228344.00
75418.98
151325.60
55894.72
0.10
0.10
0.10
0.10
0.16
0.16
0.10
0.04
0.04
0.04
0.04
0.06
0.06
0.04
Uncorrected Vol.
Mean
91803.12
SD
26480.96
85077.28
30491.50
74621.92
27725.44
117644.72
30662.77
123634.80
28211.70
175971.60
41002.20
122568.64
45010.97
71870.72
29796.13
ICV Corrected Vol.
SD
Mean
0.02
0.06
0.05
0.05
0.08
0.09
0.08
0.12
0.05
0.02
0.02
0.02
0.02
0.03
0.03
0.02
Note. The cerebellar region of interest for all tracts is Crus I for the delineated descriptives. ICV = intracranial volume.
based on cerebellar seed. Paired Wilcoxon signed-rank tests were then used to look at
differences in volume based on target cerebral ROIs for cerebello-thalamo-cortical pathways.
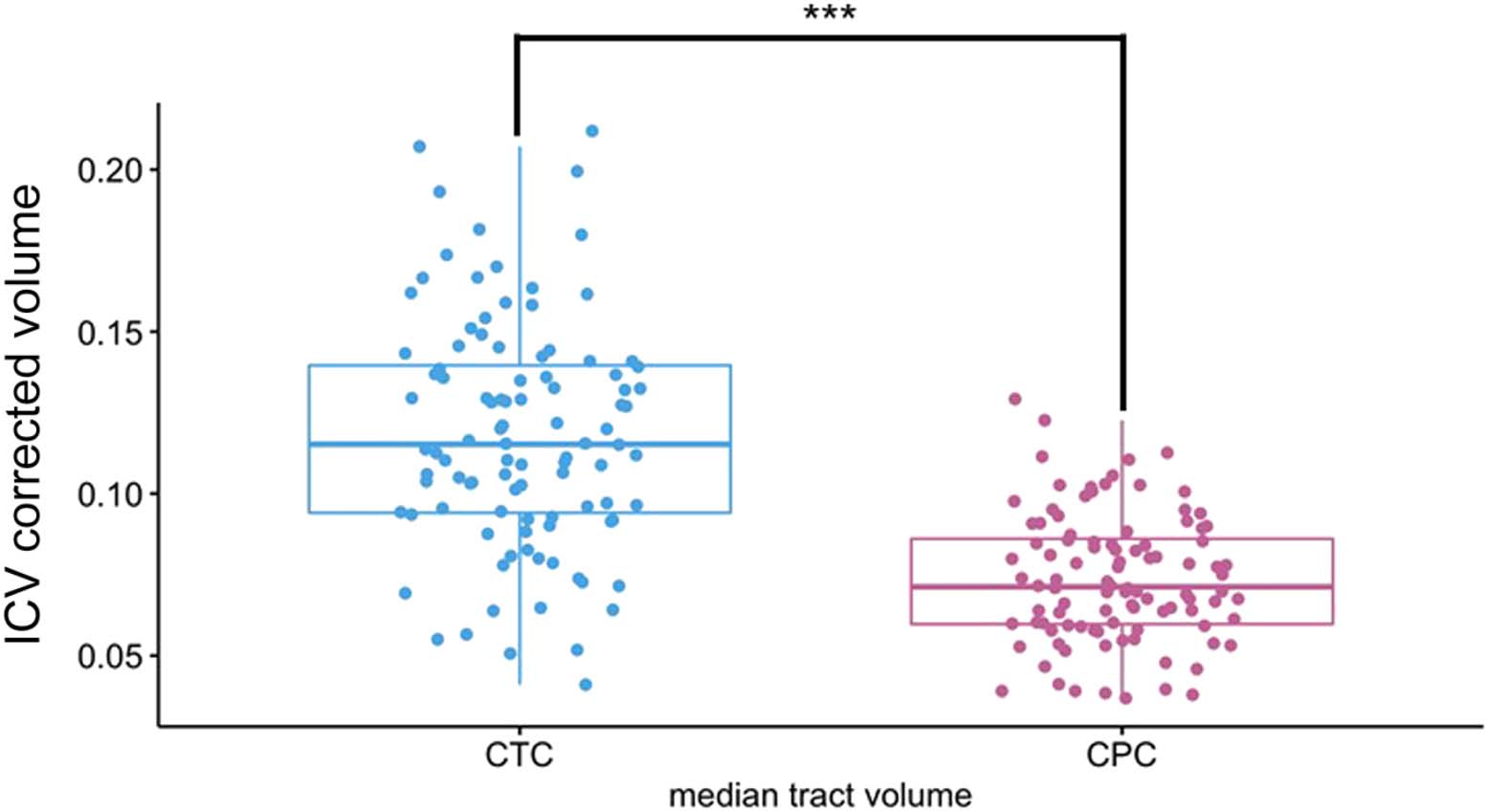
We found that tracts terminating in frontal lobe ROIs (IFG and DLPFC) had significantly higher
volumes than those of any tracts terminating in other lobe (see Figure 3A). There was
no difference between the parietal and temporal lobe tract volumes, but we found some
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Median volume of the linguistic portion of the cerebello-thalamo-cortico (CTC) and
cortico-ponto-cerebellar (CPC) tracts. Each pathway was created by averaging the volume of all
tracts across all eight cerebral regions of interest (ROIs), resulting in a single average volume per
tract for each participant. For example, the CTC seeded in Crus I had tracts calculated for each of
the right cerebral ROIs. All eight of those tract volumes were averaged to create the CTC plot shown
in this figure. Note that the size of the ROIs was identical in both analyses. For this particular exam-
ple, pathways seeded/terminated in Crus I. This was done for illustration purposes. Each dot repre-
sents a single participant; *** indicates p < 0.001.
Neurobiology of Language
10
Cerebellar white matter and language
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Volume comparison between tracts. (A) Tract beginning in right Crus I and ending in different left-lateralized cerebral language
targets. (B) Tracts projecting from cerebral language ROIs to the target in Crus I. For this particular example, pathways seeded/terminated in
Crus I. This was done for illustration purposes. For further information about each individual pairwise comparison, see Table C in the
Supporting Information. CTC = cerebello-thalamo-cortical pathway; CPC = cortico-ponto-cerebellar pathway; ICV = intracranial volume;
IFG = inferior frontal gyrus/Broca’s area; DLPFC = dorsolateral prefrontal cortex; ANG = angular gyrus; TP = temporal pole; STG = superior
temporal gyrus; MTG = middle temporal gyrus; PST = posterior superior temporal lobe; ITG = inferior temporal gyrus. *** indicates p < 0.001,
** indicates p < 0.05; ns = nonsignificant.
diversity among temporal lobe ROIs. We found a significant difference between MTG and PST
(V = 1139, p = 0.037), ITG and PST (V = 880, p < 0.001), as well as between the TP and PST
(V = 997, p < 0.001). There was also a significant difference between IFG and DLPFC volumes,
with IFG having the greater volume between the two (V = 699, p < 0.001).
We found a different pattern of connection for the cortico-ponto-cerebellar pathway path-
way (see Figure 3B). Again using paired Wilcoxon signed-rank tests, we found that there was
still a statistically significant difference between the IFG and all other cerebral ROIs. Addition-
ally, there were differences in volume in the temporal and parietal ROIs. Rather than the frontal
ROIs being greater than the rest and all other tracts being equal to each other, we found that
tracts from the ANG, STG, and TP seeds had higher volumes than in the cerebello-thalamo-
cortical tract. The overall pattern was IFG > DLPFC ⩭ ANG ⩭ TP > STG ⩭ MTG > WER ⩭ ITG
Neurobiology of Language
11
Cerebellar white matter and language
桌子 3. A summary comparison of the volumes and standard deviations for subsections of the cortico-ponto-cerebellar (CPC) and cerebello-
thalamo-cortical (CTC) connecting to working memory ( WM) 地区.
CTC (projecting to IFG)
CPC (seeded in IFG)
Uncorrected Vol.
WM ROI
Lobule IV
意思是
173764.88
标清
41627.86
Crus I
175658.40
41211.88
Lobule VIIb
174411.04
40735.06
ICV Corrected Vol.
标清
意思是
0.04
0.09
0.10
0.14
0.04
0.05
Uncorrected Vol.
意思是
138546.08
标清
56472.37
135630.24
61385.95
200515.76
68625.34
ICV Corrected Vol.
标清
意思是
0.03
0.12
0.12
0.12
0.03
0.03
笔记. The CPC projections to WM regions in the cerebellum are seeded in the IFG, while in the CTC WM cerebellar regions are projecting to IFG. ROI = region
of interest; IFG = inferior frontal gyrus; ICV = intracranial volume.
(with ⩭ signaling statistical equivalence). Overall for both tracts, the tracts whose cerebral ROIs
were IFG/Broca’s area had the highest volume (significant at p < 0.001 in all pairwise com-
parisons; see Table C in the Supporting Information for full list of comparisons).
Cerebellar Structural Connections: Language–Working Memory
To examine the specificity of these white matter connections, we ran probabilistic tractography
between the cerebellar working memory ROIs and cerebral language ROIs using the exact
same waypoints and exclusion masks as in the language–language analyses. The language–
language connections were then compared to the working memory–language connections.
We examined the white matter tracts projecting to and from IFG, as this was our highest vol-
ume target and seed. Results showed that the white matter pathways from the working memory
cerebellar ROIs to the language cerebral ROIs had significantly lower volumes for tracts going
to IFG in the cerebello-thalamo-cortical tracts (all comparisons p < 0.001; see Table C in the
Supporting Information for all p values and effect sizes, and Table 3 for working memory tract
means). Interestingly, in the cortico-ponto-cerebellar pathway tracts, there was one exception:
working memory ROI in right Lobule VI was not significantly different in ICV corrected volume
than language ROI right Crus I (W = 2,725, p = 0.667). These results suggest that there are
specific structural connections between the linguistic cerebellar and cerebral areas, with the
possibility that multiple regions of the cerebellum receive input from the same cerebral areas.
Cerebellar Microstructure: Brain-Behavior Correlations
Next, we investigated the microstructural properties of each tract to determine if we could find
a relation between white matter and behavior. We used Spearman’s rho to correlate FA
extracted from the peduncles for each tract and the behavioral outcomes of the NIH Toolbox
Picture Vocabulary Test from the HCP data set. Spearman’s rho was used as some of our dif-
fusion data from the cortico-ponto-cerebellar tract violated tests of normality (STG to Crus I:
W = 0.964, p = 0.024; MTG to Crus I: W = 0.970, p = 0.057); STG to Lobule IX: W = 0.968,
p = 0.043; MTG to Lobule IX: W = 0.977, p = 0.058). No significant correlations were found
between microstructure of the cerebello-thalamo-cortical tract and picture naming ability.
However, within the cortico-ponto-cerebellar tract there were several trending or significant
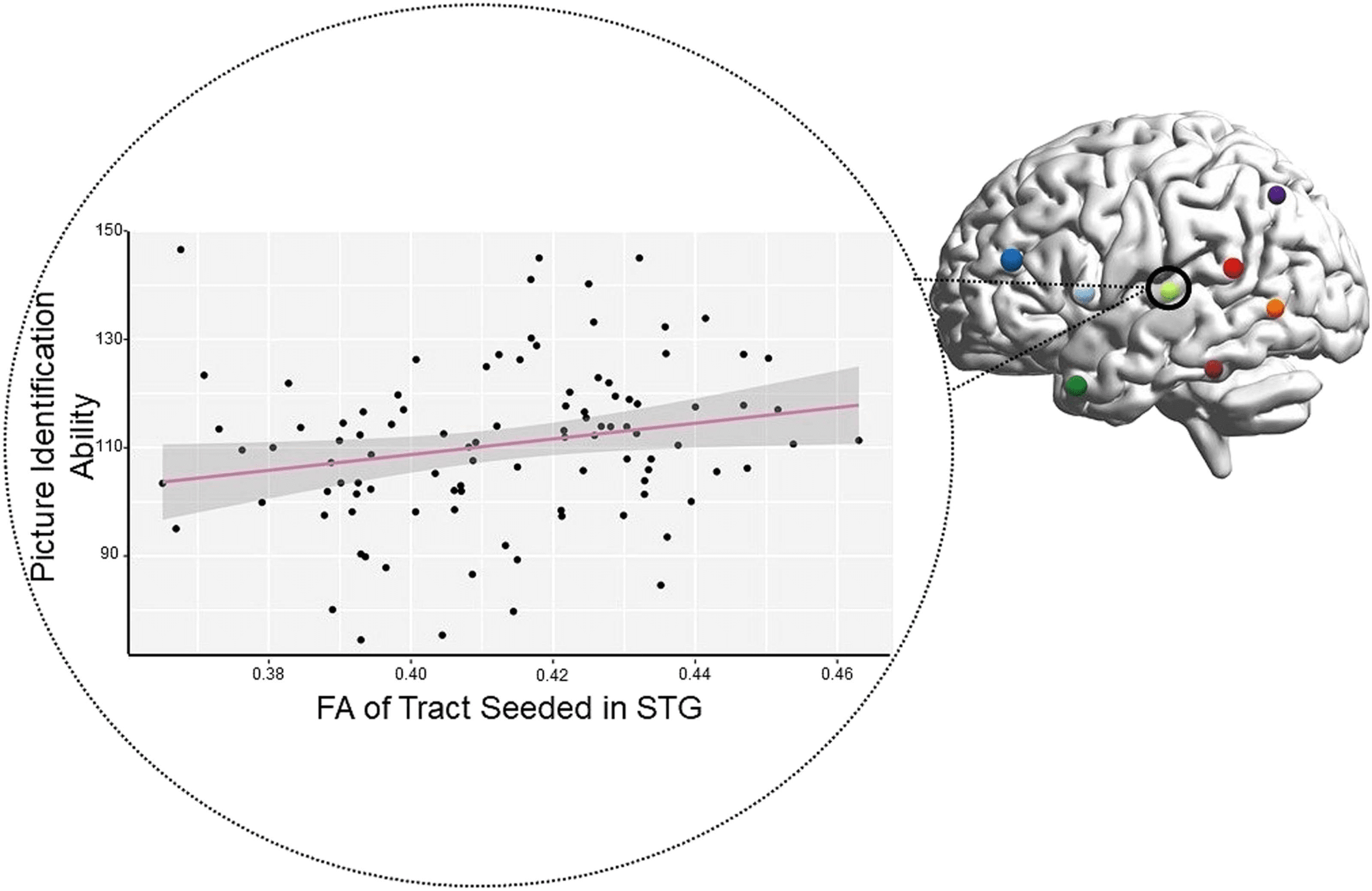
correlations. For the tracts that had Crus I as the target, tracts seeded in: IFG (r = 0.23, p =
0.058), STG (r = 0.27, p = 0.020), and TP (r = 0.24, p = 0.041) were either trending or signif-
icant. For tracts with Lobule IX as the target, tracts seeded in: IFG (r = 0.24, p = 0.044), STG
(r = 0.26, p = 0.024), and TP (r = 0.23, p = 0.050) were also significant (see Figure 5 in the
Discussion section).
Neurobiology of Language
12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Cerebellar white matter and language
DISCUSSION
In this study we mapped structural pathways between language-sensitive regions in the cere-
bellum and language-sensitive regions in the cerebral cortex. Our first step was to identify
regions of the cerebellum sensitive to verbal comprehension and then to examine overlap with
motor and working memory task activations. The regions activated to the sentence compre-
hension contrast were localized to Crus I/Crus II, and Lobule IX (see Figure 1) of the posterior
cerebellum. The location of these activations is consistent with prior work that was conducted
on different participants and using different stimuli (Geva et al., 2021; King et al., 2019). It
should be noted that the activation in Lobule IX has been a controversial addition to the lin-
guistic cerebellum. However, there is one piece of intriguing evidence linking it to receptive
language: Geva and colleagues (2021) found that in their sample of six cerebellar lesion
patients, damage to Lobule IX was the only lesion location that produced lasting deficits in
sentence comprehension. Future research should examine this relationship more closely.
We found marginal overlap between the language and working memory tasks in Crus I,
which is consistent with past literature (Marvel & Desmond, 2010). Τhis overlap could be
attributed to the nature of the language task. Participants listened to fables and later made
judgments about the fables. This required them to remember the gist of five to nine sentences,
a task that invokes verbal working memory. The fact that these fables also involve social sce-
narios explains our previous finding that the language task has nearly 50% overlap with acti-
vations from the theory of mind task (Metoki et al., 2021). There is a sizable literature linking
portions of the cerebellum to normal and abnormal social cognition as observed in autism
spectrum disorder (Stoodley & Tsai, 2021; Van Overwalle et al., 2021). Whether language
tasks that are less social and have lower working memory loads (e.g., single word processing)
would show overlap with language activations in the cerebellum is unknown. In contrast,
there was no overlap between the language comprehension task and the motor task.
One explanation of the social and language overlap could be derived from the social
sequencing hypothesis (Heleven et al., 2021; Van Overwalle et al., 2019; Van Overwalle
et al., 2021). This hypothesis states that the cerebellum is involved in the prediction of social
interaction by forming internal models (a representation of the predicted world based on pre-
vious experiences) about how sequential social events should unfold. The cerebellum uses
associative information to identify what may come next in a sequence of events, such as
another person’s reaction or response. This relates to language because language is inherently
social. Not only do we use it to relay information to other individuals, but the content of what
we are relaying is often social. One study reported stronger activations in the cerebellum to
social as compared to nonsocial sentences (Pu et al., 2020), adding credence to this idea.
While the focus of our study was to parse receptive language and working memory, future
work will need to be done to parse language and social cognition in the cerebellum.
The observed overlap between language and working memory led us to conduct a distance
analysis. This showed that each cluster had distinct local maxima. We also analyzed the
β-weights for each task at the individual level and found that all task pairs have a significantly
different localization of activation. Overall, our results provide support for the hypothesis that
the cerebellum contains domain-specific mapping of cognitive functions including language
comprehension.
If these cerebellar regions truly play a role in language, they should be structurally con-
nected to regions involved in language in the cerebrum. Histology methods in macaques have
revealed an extensive network of fiber paths between the anterior cerebellum and nearly all
nonmotor regions of the frontal lobe (Clower et al., 2005; Dum & Strick, 2003; Ito, 1984; Kelly
Neurobiology of Language
13
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Cerebellar white matter and language
& Strick, 2003; Leiner et al., 1993; Middleton & Strick, 1997; Schmahmann & Pandya, 1997).
In regards to the parietal lobe, gold-standard histology studies have found connections
between the cerebellum and BA 5 and 7 in the macaque parietal lobe (Clower et al., 2005;
Glickstein et al., 1985). Other histology studies in macaques have reported the existence
of structural connections between the superior temporal lobe (e.g. the length of the STG,
excepting A1, as well as the depths of the superior temporal sulcus), with the cerebellum
(Schmahmann & Pandya, 1991). However, there are few to no connections between most
of the MTG and ITG and the cerebellum (Schmahmann & Pandya, 1991). Thus, histological
findings in macaques predict that we should find strong structural connectivity between the
cerebellum and language ROIs in the frontal lobe and superior temporal lobe, but weaker
structural connections between inferior parietal lobe and inferior temporal lobe ROIs and
the cerebellum. This prediction must be tempered by the fact that language is a uniquely
human trait, and changes have occurred in the human temporal and inferior parietal lobes
through the evolution of language.
The results of our probabilistic tractography analysis partially confirmed the above predic-
tions. First, we found that the volume of the language-specific cerebello-thalamo-cortical path-
way was greater than the volume of the language-specific cortico-ponto-cerebellar pathway.
This was not entirely unexpected as previous research found this bias when looking at the
social portions of the cerebello-thalamo-cortical and cortico-ponto-cerebellar tracts (Metoki
et al., 2021). Interpretation of this bias awaits further investigation.
Second, we found that in the cerebello-thalamo-cortical pathway, frontal lobe ROIs cap-
tured the majority of the projections from the cerebellum (IFG followed by the DLPFC; see
Figure 3 and Figure 4). There was some elevated connectivity to the PST compared to other
temporal lobe ROIs, but nothing compared to the volume of the pathways projecting to the
frontal lobe, as was expected based on previous literature (Palesi et al., 2015).
However, the cortico-ponto-cerebellar pathway showed a different, more complex pattern
of connectivity. While IFG still held the highest number of connections to the cerebellum,
DLPFC, ANG, STG, and TP had elevated connectivity compared to other temporal lobe ROIs.
The ANG and TP structural connections captured on average the same amount of volume
as the DLPFC. The results involving the ANG and STG are not surprising, based on our review
Figure 4. Visualization of the cerebello-thalamo-cortical and cortico-ponto-cerebellar tracts. (A) The CTC tract running from Crus I of the
cerebellum to inferior frontal gyrus (blue sphere). (B) The portion of the CPC tract, running from the temporal pole (red sphere) to cerebellar
Crus I. Visualizations using 3D slcer (Fedorov et al., 2012; Kikinis et al., 2014).
Neurobiology of Language
14
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Cerebellar white matter and language
of the tract tracing and histology literature (Ramnani, 2012; Schmahmann & Pandya, 1991).
However, finding elevated volume in the projections coming from the TP were somewhat
unexpected. There is one study in macaques showing that the temporal polar section of the
superior temporal sulcus (TPO1) sends projections to the pons (Schmahmann, 1996), and
likely continues onward into the cerebellum. However, we are not aware of any other ground
truth studies.
These results provide new insight on the “closed-loop” nature of cerebellum-to-cerebrum
connections. Computational models describing the cerebellum’s functional contribution to
motor processes have described a closed loop system in which motor regions in the cerebrum
are connected to specific regions within the cerebellum, which in turn are connected to the
same regions back in the cerebrum (Ito, 2008; Kawato & Gomi, 1992; Wolpert et al., 1998).
Evidence of closed-loop cerebello-cerebral connections that would be critical for nonmotor
processes have also been found (Palesi et al., 2017; Salmi et al., 2010; Strick et al., 2009),
including closed-loops between the posterior cerebellum and the DLPFC (Kelly & Strick,
2003). However, as noted by Moberget and Ivry (2016, p. 12), “… a close examination of
the primate data suggests that [closed loops] may not always be the case.” The asymmetry
between the cerebello-thalamo-cortical and cortico-ponto-cerebellar pathway in our function-
ally defined ROIs suggest that multiple regions across the cerebral cortex may be providing
input into a specific region of the cerebellum, which then outputs to a subset of those input
regions. This finding highlights that the closed-loop nature of cerebellar connectivity may be
more in line with a network modulation function—a many-to-one—rather than a single
region-to-region, or one-to-one, modulation. Individual regions of the cerebellum would be
well suited to receive inputs from a diverse set of brain regions, as cerebellum “microcom-
plexes,” thought to be the computational building blocks of the cerebellum, include an initial
layer of cells that are well suited for high-dimensional inputs (Hull, 2020; Raymond & Medina,
2018).
To date, only a small number of studies have examined the cerebello-thalamo-cortical and
cortico-ponto-cerebellar pathways in vivo (Jissendi et al., 2008; Karavasilis et al., 2019; Keser
et al., 2015; Sokolov et al., 2014). This is likely due to the technical challenges present in
doing tractography along a long, polysynaptic pathway that has crossovers and sharp turns.
Thus most of the existing literature has simply looked at white matter within the cerebellum
(e.g., the peduncles) rather than between the cerebellum and cerebrum. Our study is unique in
that we carefully measured the linguistically relevant portion of the ouroboros loop in a large
sample with excellent data quality.
Previous studies that reconstructed cerebello-thalamo-cortical and cortico-ponto-cerebellar
pathways relied solely upon diffusion tensor imaging, which has had notable issues with cross-
ing fibers (Lee et al., 2015). Crossing fibers are in abundance in the brainstem and within these
tracts of interest (Ford et al., 2013). Our tractography has improved upon this with the use of
the ball-and-stick model, which is better at resolving crossing fibers (Behrens et al., 2007). We
modeled our methods after Palesi et al. (2017), who found evidence of projections of the
cortico-ponto-cerebellar pathway originating from the temporal lobe with constrained spher-
ical deconvolution. This method also has a better ability to solve for crossing fibers (Daducci
et al., 2014; Tournier et al., 2008; Tournier et al., 2012). Palesi and colleagues’ analysis of all
projections from association cortices showed that the temporal lobe sends projects to the cer-
ebellum via the cortico-ponto-cerebellar pathway, even though it may not receive projections
from the cerebello-thalamo-cortical. Schmahmann and Pandya (1991) also observed projec-
tions from the superior temporal sulcus to the cortico-ponto-cerebellar pathway in rhesus mon-
keys, so the results are not completely unfounded. It is possible these findings may be the result
Neurobiology of Language
15
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
.
/
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Cerebellar white matter and language
of the software and not true projections found in the human brain, but converging evidence
across multiple methodologies may suggest some shared truth.
In an exploratory analysis of DWI microstructure data, we found that individual differ-
ences in FA values in the cortico-ponto-cerebellar pathway correlated with receptive vocab-
ulary ability. In other words, individuals with higher FA in the middle cerebellar peduncle of
the cortico-ponto-cerebellar tract had better ability to identify a picture that matched a word
with which they were presented. Note that all tracts projecting from regions of the temporal
lobe, except for PST, had weak to moderate positive correlations with the receptive language.
This suggests that individuals who have faster and more efficient transmission of auditory-
linguistic information to the cerebellum perform better on single-word comprehension tasks
(Figure 5). This is reflected in our findings with the TP and the STG. The correlation of the
MTG ROI may be a little less straightforward. Some evidence has shown that the posterior
MTG is involved in semantic control (Davey et al., 2015), while others have postulated that
it could be involved in some form of integration between lobes (Turken & Dronkers, 2011).
In addition, IFG also showed weak to moderate positive correlation with the task, which
makes sense with its role in lexical and semantic processing (Dapretto & Bookheimer,
1999; Friederici et al., 2000).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Significant brain-behavior correlation: single-word comprehension performance correlated with fractional anisotropy (FA) of tract
connecting the left superior temporal gyrus (STG) to the right cerebellum. Tracts seeded in the inferior frontal gyrus and temporal pole were also
significant. Each colored sphere on the brain represents a cortical region of interest. Each black dot on the graph represents a single participant.
Neurobiology of Language
16
Cerebellar white matter and language
How selective are these findings? Our results show that there is some domain-specific con-
nectivity to the cerebrum, even within large regions such as Crus I. Tracts from working mem-
ory ROIs in the cerebellum to language ROIs in the cerebrum were volumetrically smaller than
the language–language tracts. This occurred even when the cerebellar ROIs from the different
tasks were in the same lobule of the cerebellum. Some specificity was lost in the cortico-
ponto-cerebellar tract (with fibers projecting to regions derived from the working memory
task). Tracts seeded in IFG and projecting to working memory cerebellar regions Crus I were
not significantly different in volume from the tracts projecting to the language ROI (also
located in Crus I). The longer tracts get, the less accurate they are, so this result may just be
due to the fact that the ROIs we had were located in the same lobule of the cerebellum. The
specificity that these white matter tracts hold cannot be completely resolved without a more
in-depth knowledge of what these pathways truly look like in humans. In addition, our findings
help localize what type of cerebellar damage should cause language comprehension deficits:
lesions to right Crus I or Lobule IX, as well as to the polysynaptic fiber paths connecting these
regions to the cerebrum. Individuals with lesions that preserve these regions are not expected
to exhibit any language comprehension deficits.
Limitations and Future Directions
First, the task set included in the HCP data set is limited, which limited the cerebellar seed
regions used in our tractography analysis. Future studies should test the limits of our findings
by employing a range of language production and reception tasks involving lower- and higher-
level processing, mapping the functional ROIs, and conducting tractography. Second, diffu-
sion MRI tractography has been criticized for having a high rate of false positives (Maier-Hein
et al., 2017; Reveley et al., 2015; Thomas et al., 2014). The best solution to this problem is to
conduct histology-guided tractography analyses followed by replication in independent sam-
ples. Third, the relationship between structural connectivity, as described here, and functional
connectivity, as reported in many prior studies (reviewed in Vias & Dick, 2017) in regards to
the linguistic cerebellum needs to be investigated in a larger-scale study. Last, like all neuro-
imaging analyses, results can depend on various factors including, for example, the selection
of ROIs, waypoints, thresholding, and software choices. Every analytical decision made in this
study was based on prior work exploring similar pathways (Metoki et al., 2021) with a small
number of updates based on our understanding of best practices in diffusion imaging. How-
ever we are cognizant that diffusion imaging methods are relatively young, and standards are
in flux. We hope that future investigators attempt replications of our work and that our findings
are robust to different choices made in analytical parameters.
Future directions include the investigation of the specificity of the mentalizing and lan-
guage cerebellar pathways. While it was shown that there was little functional overlap in
the cerebellum between the language and working memory tasks, this was not the same for
the language and mentalizing tasks. In order to further identify whether the language-specific
pathways are truly language-specific, further research must be done between the language and
mentalizing tasks in regards to diffusion tractography.
Conclusions
This neuroimaging study investigated the structural connectivity between language cerebellar
and cerebral areas. We found projections from the cerebellum to language regions of the
frontal lobe. There was also evidence of input to the cerebellum from other regions of the
cerebrum, such as the ANG and the TP portion of the superior temporal sulcus. These findings
Neurobiology of Language
17
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Cerebellar white matter and language
suggest that regions of the posterior cerebellum play a key role in language comprehension.
We also found that these white matter tracts were, at least in part, specific to language-
sensitive areas of the cerebellum and cerebrum.
ACKNOWLEDGMENTS
We would like to thank Angela Piecyk, Marah Dormuth, Sarah Johnson, Giovanna Arantes De
Oliveira Campos, and Jason Konadu for their work as research assistants on this project. We
would also like to thank Huiling Peng for assistance with tractography. Last, we would like to
thank three anonymous reviewers for their helpful commentary.
This research includes calculations carried out on HPC resources supported in part by the
National Science Foundation through major research instrumentation grant number 1625061
and by the US Army Research Laboratory under contract number W911NF-16-2-0189. The
content is solely the responsibility of the authors and does not necessarily represent the official
views of the National Institute of Mental Health or the National Institutes of Health.
FUNDING INFORMATION
Ingrid R. Olson, National Institutes of Health (https://dx.doi.org/10.13039/100000002), Award
ID: R21 HD098509. Ingrid R. Olson, National Institutes of Health (https://dx.doi.org/10.13039
/100000002), Award ID: R01 NICHD. Ingrid R. Olson, National Institute of Child Health and
Human Development ([institution doi]), Award ID: R01HD099165. Ingrid R. Olson, National
Institute of Mental Health (https://dx.doi.org/10.13039/100000025), Award ID: MH091113.
AUTHOR CONTRIBUTIONS
Katie R. Jobson: Conceptualization: Lead; Data curation: Lead; Formal analysis: Lead; Meth-
odology: Lead; Software: Lead; Validation: Lead; Visualization: Lead; Writing—original draft:
Lead; Writing—review & editing: Supporting. Linda J. Hoffman: Formal analysis: Equal;
Methodology: Supporting; Validation: Supporting; Writing—original draft: Supporting;
Writing—review & editing: Equal. Athanasia Metoki: Formal analysis: Supporting; Methodol-
ogy: Supporting; Writing—original draft: Equal; Writing—review & editing: Equal. Haroon
Popal: Conceptualization: Supporting; Writing—original draft: Supporting; Writing—review
& editing: Equal. Anthony S. Dick: Methodology: Supporting; Writing—review & editing:
Equal. Jamie Reilly: Conceptualization: Supporting; Methodology: Supporting;
Writing—review & editing: Equal. Ingrid R. Olson: Conceptualization: Equal; Formal analysis:
Supporting; Funding acquisition: Lead; Resources: Lead; Supervision: Equal; Writing—original
draft: Equal; Writing—review & editing: Equal.
Q3
REFERENCES
Aesop. (2012). Fables ( J. R. Long, Comp.). Aesop’s Fables online
collection. https://aesopfables.com
Ahmadian, N., van Baarsen, K., van Zandvoort, M., & Robe, P. A.
(2019). The cerebellar cognitive affective syndrome—A
meta-analysis. Cerebellum, 18(5), 941–950. https://doi.org/10
.1007/s12311-019-01060-2, PubMed: 31392563
Andersson, J. L. R., Jenkinson, M., & Smith, S. (2007). Non-linear
registration, aka spatial normalisation (FMRIB Technical Report
TR07JA2). FMRIB Centre. Oxford, UK.
Barch, D. M., Burgess, G. C., Harms, M. P., Petersen, S. E.,
Schlaggar, B. L., Corbetta, M., Glasser, M. F., Curtiss, S., Dixit,
S., Feldt, C., Nolan, D., Bryant, E., Hartley, T., Footer, O., Bjork,
J. M., Poldrack, R., Smith, S., Johansen-Berg, H., Snyder, A. Z., …
WU-Minn HCP Consortium. (2013). Function in the human con-
nectome: Task-fMRI and individual differences in behavior. Neu-
roImage, 80, 169–189. https://doi.org/10.1016/j.neuroimage
.2013.05.033, PubMed: 23684877
Behrens, T. E. J., Johansen-Berg, H., Jbabdi, S., Rushworth, M. F. S.,
& Woolrich, M. W. (2007). Probabilistic diffusion tractography
with multiple fibre orientations: What can we gain? NeuroImage,
34(1), 144–155. https://doi.org/10.1016/j.neuroimage.2006.09
.018, PubMed: 17070705
Neurobiology of Language
18
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Cerebellar white matter and language
Behrens, T. E. J., Woolrich, M. W., Jenkinson, M., Johansen-Berg, H.,
Nunes, R. G., Clare, S., Matthews, P. M., Brady, J. M., & Smith,
S. M. (2003). Characterization and propagation of uncertainty in
diffusion-weighted MR imaging. Magnetic Resonance in Medi-
cine, 50(5), 1077–1088. https://doi.org/10.1002/mrm.10609,
PubMed: 14587019
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society: Series B (Methodological),
57(1), 289–300. https://doi.org/10.1111/j.2517-6161.1995
.tb02031.x
Binder, J. R., Gross, W. L., Allendorfer, J. B., Bonilha, L., Chapin, J.,
Edwards, J. C., Grabowski, T. J., Langfitt, J. T., Loring, D. W.,
Lowe, M. J., Koenig, K., Morgan, P. S., Ojemann, J. G., Rorden,
C., Szaflarski, J. P., Tivarus, M. E., & Weaver, K. E. (2011). Map-
ping anterior temporal lobe language areas with fMRI: A multi-
center normative study. NeuroImage, 54(2), 1465–1475. https://
doi.org/10.1016/j.neuroimage.2010.09.048 , PubMed:
20884358
Booth, J. R., Wood, L., Lu, D., Houk, J. C., & Bitan, T. (2007). The
role of the basal ganglia and cerebellum in language processing.
Brain Research, 1133(1), 136–144. https://doi.org/10.1016/j
.brainres.2006.11.074, PubMed: 17189619
Bostan, A. C., Dum, R. P., & Strick, P. L. (2013). Cerebellar networks
with the cerebral cortex and basal ganglia. Trends in Cognitive
Sciences, 17(5), 241–254. https://doi.org/10.1016/j.tics.2013.03
.003, PubMed: 23579055
Brodal, P., & Bjaalie, J. G. (1997). Salient anatomic features of the
cortico-ponto-cerebellar pathway. Progressive Brain Research,
114, 227–250. https://doi.org/10.1016/S0079-6123(08)63367-1
Buckner, R. L., Krienen, F. M., Castellanos, A., Diaz, J. C., & Yeo,
B. T. T. (2011). The organization of the human cerebellum
estimated by intrinsic functional connectivity. Journal of Neuro-
physiology, 106(5), 2322–2345. https://doi.org/10.1152/jn.00339
.2011, PubMed: 21795627
Buckner, R. L., Krienen, F. M., & Yeo, B. T. T. (2013). Opportunities
and limitations of intrinsic functional connectivity MRI. Nature
Neuroscience, 16(7), 832–837. https://doi.org/10.1038/nn.3423,
PubMed: 23799476
Caplan, D., & Waters, G. (1999). Verbal working memory and
sentence comprehension. Behavioral and Brain Sciences, 22(1),
77–94. https://doi.org/10.1017/S0140525X99001788, PubMed:
11301522
Chen, S. H., & Desmond, J. E. (2005). Cerebrocerebellar networks
during articulatory rehearsal and verbal working memory tasks.
NeuroImage, 24(2), 332–338. https://doi.org/10.1016/j
.neuroimage.2004.08.032, PubMed: 15627576
Clower, D. M., Dum, R. P., & Strick, P. L. (2005). Basal ganglia and
cerebellar inputs to AIP. Cerebral Cortex, 15(7), 913–920. https://
doi.org/10.1093/cercor/bhh190, PubMed: 15459083
Cohen, J. (1988). Statistical power analysis for the behavioral
sciences (2nd ed.). Lawrence Erlbaum.
Daducci, A., Canales-Rodríguez, E. J., Descoteaux, M., Garyfallidis,
E., Gur, Y., Lin, Y.-C., Mani, M., Merlet, S., Paquette, M., Ramirez-
Manzanares, A., Reisert, M., Reis Rodrigues, P., Sepehrband, F.,
Caruyer, E., Choupan, J., Deriche, R., Jacob, M., Menegaz, G.,
Prčkovska, V., … Thiran, J.-P. (2014). Quantitative comparison
of reconstruction methods for intra-voxel fiber recovery from
diffusion MRI. IEEE Transactions on Medical Imaging, 33(2),
384–399. https://doi.org/10.1109/TMI.2013.2285500, PubMed:
24132007
Dapretto, M., & Bookheimer, S. Y. (1999). Form and content: Dis-
sociating syntax and semantics in sentence comprehension.
Neuron, 24(2), 427–432. https://doi.org/10.1016/S0896-6273
(00)80855-7, PubMed: 10571235
Davey, J., Cornelissen, P. L., Thompson, H. E., Sonkusare, S.,
Hallam, G., Smallwood, J., & Jefferies, E. (2015). Automatic
and controlled semantic retrieval: TMS reveals distinct contribu-
tions of posterior middle temporal gyrus and angular gyrus. Jour-
nal of Neuroscience, 35(46), 15230–15239. https://doi.org/10
.1523/JNEUROSCI.4705-14.2015, PubMed: 26586812
Dejerine, J., & Dejerine-Klumpke, A. M. K. (1895). Anatomie des
centres nerveux. Rueff et Cie.
Dejerine, J., & Dejerine-Klumpke, A. M. K. (1901). Anatomie des
centres nerveux. Rueff et Cie.
Desikan R. S., Ségonne, F., Fischl, B., Quinn, B. T., Dickerson,
B. C., Blacker, D., Buckner, R. L., Dale, A. M., Maguire, R. P.,
Hyman, B. T., Albert, M. S., & Killiany, R. J. (2006). An automated
labeling system for subdividing the human cerebral cortex on
MRI scans into gyral based regions of interest. NeuroImage,
31(3), 968–80. https://doi.org/10.1016/j.neuroimage.2006.01
.021, PubMed: 16530430
De Smet, H. J., Baillieux, H., De Deyn, P. P., Mariën, P., & Paquier,
P. (2007). The cerebellum and language: The story so far. Folia
Phoniatrica et Logopaedica, 59(4), 165–170. https://doi.org/10
.1159/000102927, PubMed: 17627124
Desmond, J. E., Gabrieli, J. D., Wagner, A. D., Ginier, B. L., &
Glover, G. H. (1997). Lobular patterns of cerebellar activation in
verbal working-memory and finger-tapping tasks as revealed by
functional MRI. Journal of Neuroscience, 17(24), 9675–9685.
https://doi.org/10.1523/ JNEUROSCI.17-24-09675.1997,
PubMed: 9391022
Dice, L. R. (1945). Measures of the amount of ecologic association
between species. Ecology, 26(3), 297–302. https://doi.org/10
.2307/1932409
Dick, A. S., & Tremblay, P. (2012). Beyond the arcuate fasciculus:
Consensus and controversy in the connectional anatomy of
language. Brain, 135(Part 12), 3529–3550. https://doi.org/10
.1093/brain/aws222, PubMed: 23107648
Diedrichsen, J., Balsters, J. H., Flavell, J., Cussans, E., & Ramnani,
N. (2009). A probabilistic MR atlas of the human cerebellum.
NeuroImage, 46(1), 39–46. https://doi.org/10.1016/j.neuroimage
.2009.01.045, PubMed: 19457380
Diedrichsen, J., & Zotow, E. (2015). Surface-based display of
volume-averaged cerebellar imaging data. PLOS One, 10(7),
Article e0133402. https://doi.org/10.1371/journal.pone
.0133402, PubMed: 26230510
Doyon, J., Laforce, R., Jr., Bouchard, G., Gaudreau, D., Roy, J.,
Poirier, M., Bédard, P. J., Bédard, F., & Bouchard, J. P. (1998).
Role of the striatum, cerebellum and frontal lobes in the autom-
atization of a repeated visuomotor sequence of movements.
Neuropsychologia, 36(7), 625–641. https://doi.org/10.1016
/S0028-3932(97)00168-1, PubMed: 9723934
Duffau, H. (2015). Stimulation mapping of white matter tracts to
study brain functional connectivity. Nature Reviews Neurology,
11(5), 255–265. https://doi.org/10.1038/nrneurol.2015.51,
PubMed: 25848923
Dum, R. P., & Strick, P. L. (2003). An unfolded map of the cerebellar
dentate nucleus and its projections to the cerebral cortex. Journal
of Neurophysiology, 89 (1), 634–639. https://doi.org/10.1152/jn
.00626.2002, PubMed: 12522208
Elam, J. (2021). HCP-YA Data Dictionary—Updated for the 1200
Subject Release [Instrument worksheet]. HCP Wiki—Public
Data. https://wiki.humanconnectome.org/display/ PublicData
/ HCP-YA+Data+Dictionary-+Updated+for+the+1200+Subject
+Release
Neurobiology of Language
19
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Cerebellar white matter and language
Fabbro, F., Tavano, A., Corti, S., Bresolin, N., De Fabritiis, P, &
Borgatti, R. (2004). Longterm neuropsychological deficits after
cerebellar infarctions in two young twins. Neuropsychologia,
42(4), 536–545. https://doi.org/10.1016/j.neuropsychologia
.2003.09.006, PubMed: 14728925
Fedorov, A., Beichel, R., Kalpathy-Cramer, J., Finet, J., Fillion-
Robin, J.-C., Pujol, S., Bauer, C., Jennings, D., Fennessy, F. M.,
Sonka, M., Buatti, J., Aylward, S. R., Miller, J. V., Pieper, S., &
Kikinis, R. (2012). 3D slicer as an image computing platform
for the quantitative imaging network. Magnetic Resonance
Imaging, 30(9), 1323–1341. https://doi.org/10.1016/j.mri.2012
.05.001, PubMed: 22770690
Fengler, A., Meyer, L., & Friederici. A. D. (2016). How the brain
attunes to sentence processing: Relating behavior, structure,
and function. NeuroImage, 129, 268–278. https://doi.org/10
.1016/j.neuroimage.2016.01.012, PubMed: 26777477
Fiez, J. A. (2016). The cerebellum and language: Persistent themes
and findings. Brain and Language, 161, 1–3. https://doi.org/10
.1016/j.bandl.2016.09.004, PubMed: 27663062
Fischer, M. H., & Zwaan, R. A. (2008). Embodied language:
A review of the role of the motor system in language compre-
hension. Quarterly Journal of Experimental Psychology, 61(6),
825–850. https://doi.org/10.1080/17470210701623605,
PubMed: 18470815
Ford, A. A., Colon-Perez, L., Triplett, W. T., Gullett, J. M., Mareci,
T. H., & FitzGerald, D. B. (2013). Imaging white matter in human
brainstem. Frontiers in Human Neuroscience, 7, 400. https://doi
.org/10.3389/fnhum.2013.00400, PubMed: 23898254
Frazier, J. A., Chiu, S., Breeze, J. L., Makris, N., Lange, N., Kennedy,
D. N., Herbert, M. R., Bent, E. K., Koneru, V. K., Dieterich, M. E.,
Hodge, S. M., Rauch, S. L., Grant, P. E., Cohen, B. M., Seidman,
L. J., Caviness, V. S., & Biederman J. (2005). Structural brain
magnetic resonance imaging of limbic and thalamic volumes
in pediatric bipolar disorder. American Journal of Psychiatry,
162(7), 1256–1265. https://doi.org/10.1176/appi.ajp.162.7
.1256, PubMed: 15994707
Friederici, A. D. (2002). Towards a neural basis of auditory
sentence processing. Trends in Cognitive Neuroscience, 6(2),
78–84. https://doi.org/10.1016/S1364-6613(00)01839-8,
PubMed: 15866191
Friederici, A. D., Opitz, B., & von Cramon, D. Y. (2000). Segregat-
ing semantic and syntactic aspects of processing in the human
brain: An fMRI investigation of different word types. Cerebral
Cortex, 10(7), 698–705. https://doi.org/10.1093/cercor/10.7
.698, PubMed: 10906316
Friederici, A. D., Rüschemeyer, S. A., Hahne, A., & Fiebach, C. J.
(2003). The role of left inferior frontal and superior temporal
cortex in sentence comprehension: Localizing syntactic and
semantic processes. Cerebral Cortex, 13(2), 170–177. https://
doi.org/10.1093/cercor/13.2.170, PubMed: 12507948
Frost, J. A., Binder, J. R., Springer, J. A., Hammeke, T. A., Bellgowan,
P. S. F., Rao, S. M., & Cox, R. W. (1999). Language processing is
strongly left lateralized in both sexes: Evidence from functional
MRI. Brain, 122(Part 2), 199–208. https://doi.org/10.1093/brain
/122.2.199, PubMed: 10071049
Gershon, R. C., Cook, K. F., Mungas, D., Manly, J. J., Slotkin, J.,
Beaumont, J. L., & Weintraub, S. (2014). Language measures of
the NIH Toolbox Cognition Battery. Journal of the International
Neuropsychological Society, 20(6), 642–651. https://doi.org/10
.1017/S1355617714000411, PubMed: 24960128
Gershon, R. C., Slotkin, J., Manly, J. J., Blitz, D. L., Beaumont, J. L.,
Schnipke, D., Wallner-Allen, K., Golinkoff, R. M., Gleason, J. B.,
Hirsh-Pasek, K., Adams, M. J., & Weintraub, S. (2013). NIH
Toolbox Cognition Battery (CB): Measuring language (vocabulary
comprehension and reading decoding). Monographs of the Soci-
ety for Research in Child Development, 78(4), 49–69. https://doi
.org/10.1111/mono.12034, PubMed: 23952202
Geva, S., Schneider, L. M., Roberts, S., Green, D. W., & Price, C. J.
(2021). The effect of focal damage to the right medial posterior
cerebellum on word and sentence comprehension and produc-
tion. Frontiers in Human Neuroscience, 15, Article 664650.
https://doi.org/10.3389/fnhum.2021.664650, PubMed:
34093152
Glasser, M. F., Sotiropoulos, S. N., Wilson, J. A., Coalson, T. S.,
Fischl, B., Andersson, J. L., Xu, J., Jbabdi, S., Webster, M.,
Polimeni, J. R., Van Essen, D. C., Jenkinson, M., & WU-Minn
HCP Consortium. (2013). The minimal preprocessing pipelines
for the human connectome project. NeuroImage, 80, 105–124.
https://doi.org/10.1016/j.neuroimage.2013.04.127, PubMed:
23668970
Glickstein, M., May, J. G., III, & Mercier, B. E. (1985). Corticopon-
tine projection in the macaque: The distribution of labelled
cortical cells after large injections of horseradish peroxidase in
the pontine nuclei. Journal of Comparative Neurology, 235(3),
343–359. https://doi.org/10.1002/cne.902350306, PubMed:
3998215
Goldstein, J. M., Seidman, L. J., Makris, N., Ahern, T., O’Brien,
L. M., Caviness, V. S., Jr., Kennedy, D. N., Faraone, S. V., &
Tsuang, M. T. (2007). Hypothalamic abnormalities in schizophre-
nia: Sex effects and genetic vulnerability. Biological Psychiatry,
61(8), 935–945. https://doi.org/10.1016/j.biopsych.2006.06
.027, PubMed: 17046727
Gonzalo-Ruiz, A., & Leichnetz, G. R. (1990). Connections of the
caudal cerebellar interpositus complex in a new world monkey
(Cebus apella). Brain Research Bulletin, 25(6), 919–927. https://
doi.org/10.1016/0361-9230(90)90189-7, PubMed: 2289174
Griffanti, L., Salimi-Khorshidi, G., Beckmann, C. F., Auerbach, E. J.,
Douaud, G., Sexton, C. E., Zsoldos, E., Ebmeier, K. P., Filippini,
N., Mackay, C. E., Moeller, S., Xu, J., Yacoub, E., Baselli, G.,
Ugurbil, K., Miller, K. L., & Smith, S. M. (2014). ICA-based arte-
fact removal and accelerated fMRI acquisition for improved rest-
ing state network imaging. NeuroImage, 95, 232–247. https://doi
.org/10.1016/j.neuroimage.2014.03.034, PubMed: 24657355
Guell, X., Gabrieli, J. D. E., & Schmahmann, J. D. (2018). Triple
representation of language, working memory, social and emotion
processing in the cerebellum: Convergent evidence from task
and seed-based resting-state fMRI analyses in a single large
cohort. NeuroImage, 172, 437–449. https://doi.org/10.1016/j
.neuroimage.2018.01.082, PubMed: 29408539
Guell, X., Schmahmann, J. D., & Gabrieli, J. D. E., & Ghosh, S. S.
(2018). Functional gradients of the cerebellum. eLife, 7, Article
e36652. https://doi.org/10.7554/eLife.36652, PubMed:
30106371
Heleven, E., van Dun, K., De Witte, S., Baeken, C., & Van Overwalle,
F. (2021). The role of the cerebellum in social and non-social
action sequences: A preliminary LF-rTMS study. Frontiers in
Human Neuroscience, 15, Article 593821. https://doi.org/10
.3389/fnhum.2021.593821, PubMed: 33716690
Hoche, F., Guell, X., Vangel, M. G., Sherman, J. C., & Schmahmann,
J. D. (2018). The cerebellar cognitive affective/Schmahmann
syndrome scale. Brain, 141(1), 248–270. https://doi.org/10.1093
/brain/awx317, PubMed: 29206893
Hua, K., Zhang, J., Wakana, S., Jiang, H., Li, X., Reich, D. S.,
Calabresi, P. A., Pekar, J. J., van Zijl, P. C. M., & Mori, S.
(2008). Tract probability maps in stereotaxic spaces: Analyses
of white matter anatomy and tract-specific quantification.
Neurobiology of Language
20
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Cerebellar white matter and language
NeuroImage, 39(1), 336–347. https://doi.org/10.1016/j.neuroimage
.2007.07.053, PubMed: 17931890
Hull, C. (2020). Prediction signals in the cerebellum: Beyond super-
vised motor learning. eLife, 9, Article e54073. https://doi.org/10
.7554/eLife.54073, PubMed: 32223891
Ikuta, N., Sugiura, M., Sassa, Y., Watanabe, J., Akitsuki, Y., Iwata, K.,
Miura, N., Okamoto, H., Watanabe, Y., Sato, S., Horie, K.,
Matsue, Y., & Kawashima, R. (2006). Brain activation during
the course of sentence comprehension. Brain and Language,
97(2), 154–161. https://doi.org/10.1016/j.bandl.2005.10.006,
PubMed: 16298427
Ito, H., Murakami, T., Fukuoka, T., & Kishida, R. (1986). Thalamic fiber
connections in a teleost (Sebastiscus marmoratus): Visual somato-
sensory, octavolateral, and cerebellar relay region to the telenceph-
alon. Journal of Comparative Neurology, 250(2), 215–227. https://
doi.org/10.1002/cne.902500208, PubMed: 3745513
Ito, M. (1984). The cerebellum and neural control. Raven Press.
Ito, M. (2008). Control of mental activities by internal models in the
cerebellum. Nature Reviews Neuroscience, 9(4), 304–313.
https://doi.org/10.1038/nrn2332, PubMed: 18319727
Ivry, R. B., & Keele, S. W. (1989). Timing functions of the cerebel-
lum. Journal of Cognitive Neuroscience, 1(2), 136–152. https://
doi.org/10.1162/jocn.1989.1.2.136, PubMed: 23968462
Jenkinson, M., Beckmann, C. F., Behrens, T. E. J., Woolrich, M. W.,
& Smith, S. M. (2012). FSL. NeuroImage, 62, 782–790. https://doi
.org/10.1016/j.neuroimage.2011.09.015, PubMed: 21979382
Jissendi, P., Baudry, S., & Balériaux, D. (2008). Diffusion tensor
imaging (DTI) and tractography of the cerebellar projections to
prefrontal and posterior parietal cortices: A study at 3T. Journal
of Neuroradiology, 35(1), 42–50. https://doi.org/10.1016/j
.neurad.2007.11.001, PubMed: 18206240
Jones, D. K. (2010). Challenges and limitations of quantifying brain
connectivity in vivo with diffusion MRI. Imaging in Medicine,
2(3), 341–355. https://doi.org/10.2217/iim.10.21
Jossinger, S., Kronfeld-Duenias, V., Zislis, A., Amir, O., & Ben-
Shachar, M. (2021). Speech rate association with cerebellar
white-matter diffusivity in adults with persistent developmental
stuttering. Brain Structure & Function, 226(3), 801–816. https://
doi.org/10.1007/s00429-020-02210-7, PubMed: 33538875
Just, M. A., Carpenter, P. A., Keller, T. A., Eddy, W. F., & Thulborn,
K. R. (1996). Brain activation modulated by sentence compre-
hension. Science, 274(5284), 114–116. https://doi.org/10.1126
/science.274.5284.114, PubMed: 8810246
Karavasilis, E., Christidi, F., Velonakis, G., Giavri, Z., Kelekis, N. L.,
Efstathopoulos, E. P., Evdokimidis, I., & Dellatolas, G. (2019).
Ipsilateral and contralateral cerebro-cerebellar white matter con-
nections: A diffusion tensor imaging study in healthy adults. Jour-
nal of Neuroradiology, 46(1), 52–60. https://doi.org/10.1016/j
.neurad.2018.07.004, PubMed: 30098370
Kawato, M., & Gomi, H. (1992). A computational model of four
regions of the cerebellum based on feedback-error learning. Bio-
logical Cybernetics, 68(2), 95–103. https://doi.org/10.1007
/BF00201431, PubMed: 1486143
Keller, T. A., Carpenter, P. A., & Just, M. A. (2001). The neural bases
of sentence comprehension: A fMRI examination of syntactic and
lexical processing. Cerebral Cortex, 11(3), 223–237. https://doi
.org/10.1093/cercor/11.3.223, PubMed: 11230094
Kelly, R. M., & Strick, P. L. (2003). Cerebellar loops with motor
cortex and prefrontal cortex of a nonhuman primate. Journal of
Neuroscience, 23(23), 8432–8444. https://doi.org/10.1523
/JNEUROSCI.23-23-08432.2003, PubMed: 12968006
Keser, Z., Hasan, K. M., Mwangi, B. I., Kamali, A., Ucisik-Keser,
F. E., Riascos, R. F., Yozbatiran, N., Francisco, G. E., & Narayana,
P. A. (2015). Diffusion tensor imaging of the human cerebellar
pathways and their interplay with cerebral macrostructure. Fron-
tiers in Neuroanatomy, 9, 41. https://doi.org/10.3389/fnana.2015
.00041, PubMed: 25904851
Kieren, F. M., & Buckner, R. L. (2009). Segregated fronto-cerebellar
circuits revealed by intrinsic functional connectivity. Cerebral
Cortex, 19(10), 2485–2497. https://doi.org/10.1093/cercor
/bhp135, PubMed: 19592571
Kikinis, R., Pieper, S. D., & Vosburgh, K. (2014) 3D slicer: A
platform for subject-specific image analysis, visualization, and
clinical support. In F. Jolesz (Ed.), Intraoperative imaging image-
guided therapy (pp. 277–289). Springer. https://doi.org/10.1007
/978-1-4614-7657-3_19
King, M., Hernandez-Castillo, C. R., Poldrack, R. A., Ivry, R. B., &
Diedrichsen, J. (2019). Functional boundaries in the human cer-
ebellum revealed by a multi-domain task battery. Nature Neuro-
science, 22(8), 1371–1378. https://doi.org/10.1038/s41593-019
-0436-x, PubMed: 31285616
Krestel, H., Annoni, J.-M., & Jagella, C. (2013). White matter in
aphasia: A historical review of Dejerines’ studies. Brain and Lan-
guage, 127(3), 526–532. https://doi.org/10.1016/j.bandl.2013.05
.019, PubMed: 23895939
LeBel, A., Jain, S., & Huth, A G. (2021). Voxelwise encoding
models show that cerebellar language representations are highly
conceptual. Journal of Neuroscience, 41(50), 10341–10355.
https://doi.org/10.1523/ JNEUROSCI.0118-21.2021, PubMed:
34732520
Lee, D.-H., Park, J. W., Park, S.-H., & Hong, C. (2015). Have you
ever seen the impact of crossing fiber in DTI?: Demonstration of
the corticospinal tract pathway. PLOS One, 10(7), Article
e0112045. https://doi.org/10.1371/journal.pone.0112045,
PubMed: 26135795
Leggio, M., & Molinari, M. (2015). Cerebellar sequencing: A trick
for predicting the future. Cerebellum, 14(1), 35–38. https://doi
.org/10.1007/s12311-014-0616-x, PubMed: 25331541
Leiner, H. C., Leiner, A. L., & Dow, R. S. (1993). Cognitive and lan-
guage functions of the human cerebellum. Trends in Neurosci-
ences, 16(11), 444–447. https://doi.org/10.1016/0166-2236(93)
90072-T, PubMed: 7507614
Maier-Hein, K. H., Neher, P. F., Houde, J.-C., Côté, M.-A.,
Garyfallidis, E., Zhong, J., Chamberland, M., Yeh, F.-C., Lin, Y.-C.,
Ji, Q., Reddick, W. E., Glass, J. O., Chen, D. Q., Feng, Y., Gao, C.,
Wu, Y., Ma, J., He, R., Li, Q., … Descoteaux, M. (2017). The chal-
lenge of mapping the human connectome based on diffusion
tractography. Nature Communications, 8(1), 1349. https://doi
.org/10.1038/s41467-017-01285-x, PubMed: 29116093
Makris, N., Goldstein, J. M., Kennedy, D., Hodge, S. M., Caviness,
V. S., Faraone, S. V., Tsuang, M. T., & Seidman, L. J. (2006).
Decreased volume of left and total anterior insular lobule in
schizophrenia. Schizophrenia Research, 83(2–3), 155–171.
https://doi.org/10.1016/j.schres.2005.11.020, PubMed:
16448806
Malone, I. B., Leung, K. K., Clegg, S., Barnes, J., Whitwell, J. L.,
Ashburner, J., Fox, N. C., & Ridgway, G. R. (2015). Accurate
automatic estimation of total intracranial volume: A nuisance var-
iable with less nuisance. NeuroImage, 104, 366–372. https://doi
.org/10.1016/j.neuroimage.2014.09.034, PubMed: 25255942
Marek, S., Siegel, J. S., Gordon, E. M., Raut, R. V., Gratton, C.,
Newbold, D. J., Ortega, M., Laumann, T. O., Adeyemo, B.,
Miller, D. B., Zheng, A., Lopez, K. C., Berg, J. J., Coalson,
R. S., Nguyen, A. L., Dierker, D., Van, A. N., Hoyt, C. R.,
McDermott, K. B., … Dosenbach, N. U. F. (2018). Spatial and
temporal organization of the individual human cerebellum.
Neurobiology of Language
21
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Cerebellar white matter and language
Neuron, 100(4), 977–993. https://doi.org/10.1016/j.neuron.2018
.10.010, PubMed: 30473014
Mariën, P., Ackermann, H., Adamaszek, M., Barwood, C. H.,
Beaton, A., Desmond, J., De Witte, E., Fawcett, A. J., Hertrich,
I., Küper, M., Leggio, M., Marvel, C., Molinari, M., Murdoch,
B. E., Nicolson, R. I., Schmahmann, J. D., Stoodley, C. J.,
Thürling, M., Timmann, D., Wouters, E., & Ziegler, W. (2014).
Consensus paper: Language and the cerebellum: An ongoing
enigma. Cerebellum, 13(3), 386–410. https://doi.org/10.1007
/s12311-013-0540-5, PubMed: 24318484
Mariën, P., & Borgatti, R. (2018). Language and the cerebellum. In
Handbook of clinical neurology (Vol. 154, pp. 181–202). https://
doi.org/10.1016/ B978-0-444-63956-1.00011-4, PubMed:
29903439
Mariën, P., Engelborghs S., Fabbro F., & De Deyn P. P. (2001). The
lateralized linguistic cerebellum: A review and a new hypothesis.
Brain and Language, 79(3), 580–600. https://doi.org/10.1006
/brln.2001.2569, PubMed: 11781058
Mariën, P., Engelborghs, S., Pickut, B. A., & De Deyn, P. P. (2000).
Aphasia following cerebellar damage: Fact or fallacy? Journal of
Neurolinguistics, 13(2–3), 145–171. https://doi.org/10.1016
/S0911-6044(00)00009-9
Marvel, C. L., & Desmond, J. E. (2010). Functional topography of
the cerebellum in verbal working memory. Neuropsychology
Review, 20(3), 271–279. https://doi.org/10.1007/s11065-010
-9137-7, PubMed: 20563894
Mesulam, M.-M., Thompson, C. K., Weintraub, S., & Rogalski, E. J.
(2015). The Wernicke conundrum and the anatomy of language
comprehension in primary progressive aphasia. Brain, 138(8),
2423–2437. https://doi.org/10.1093/ brain/awv154, PubMed:
26112340
Metoki, A., Wang, Y., & Olson, I. R. (2021). The social cerebellum:
A large-scale investigation of functional and structural specificity
and connectivity. Cerebral Cortex, 32(5), 987–1003. https://doi
.org/10.1093/cercor/bhab260, PubMed: 34428293
Middlebrooks, E. H., Yagmurlu, K., Szaflarski, J. P., Rahman, M., &
Bozkurt, B. (2017). A contemporary framework of language
processing in the human brain in the context of preoperative
and intraoperative language mapping. Neuroradiology, 59(1),
69–87. https://doi.org/10.1007/s00234-016-1772-0, PubMed:
28005160
Middleton, F. A., & Strick, P. L. (1997). Cerebellar output channels.
International Review of Neurobiology, 41, 61–82. https://doi.org
/10.1016/S0074-7742(08)60347-5, PubMed: 9378611
Miraillé, C. (1896). De l’aphasie sensorielle. G. Steinheil.
Moberget, T., & Ivry, R. B. (2016). Cerebellar contributions to motor
control and language comprehension: Searching for common
computational principles. Annals of the New York Academy of
Sciences, 1369(1), 154–171. https://doi.org/10.1111/nyas
.13094, PubMed: 27206249
Molinari, M., Leggio, M. G., Solida, A., Ciorra, R., Misciagna, S.,
Silveri, M. C., & Petrosini, L. (1997). Cerebellum and procedural
learning: Evidence from focal cerebellar lesions. Brain, 120(10),
1753–1762. https://doi.org/10.1093/ brain/120.10.1753,
PubMed: 9365368
Mori, S., Wakana, S., Nagae-Poetscher, L. M., & van Zijl, P. C. M.
(2005). MRI atlas of human white matter (1st ed.). Springer.
https://doi.org/10.1016/B978-044451741-8/50001-9
Murdoch, B. E., & Whelan, B.-M. (2007). Language disorders sub-
sequent to left cerebellar lesions: A case for bilateral cerebellar
involvement in language? Folia Phoniatrica et Logopaedica,
59(4), 184–189. https://doi.org/10.1159/000102930, PubMed:
17627127
Oldfield, R. C. (1971). The assessment and analysis of handedness:
The Edinburgh Inventory. Neuropsychologia, 9(1), 97–113.
https://doi.org/10.1016/0028-3932(71)90067-4, PubMed:
5146491
Palesi, F., De Rinaldis, A., Castellazzi, G., Calamante, F., Muhlert,
N., Chard, D., Tournier, J. D., Magenes, G., D’Angelo, E., &
Wheeler-Kingshott, C. A. M. G.
(2017). Contralateral
cortico-ponto-cerebellar pathways reconstruction in humans
in vivo: Implications for reciprocal cerebro-cerebellar structural
connectivity in motor and non-motor areas. Science Reports,
7(1), Article 12841. https://doi.org/10.1038/s41598-017-13079-8,
PubMed: 28993670
Palesi, F., Ferrante, M., Gaviraghi, M., Misiti, A., Savini, G.,
Lascialfari, A., D’Angelo, E., & Wheeler-Kingshott, C. A. M. G.
(2021). Motor and higher-order functions topography of the
human dentate nuclei identified. Human Brain Mapping,
42(13), 4348–4361. https://doi.org/10.1002/ hbm.25551,
PubMed: 34087040
Palesi, F., Tournier, J.-D., Calamante, F., Muhlert, N., Castellazzi,
G., Chard, D., D’Angelo, E., & Wheeler-Kingshott, C. A. M. G.
(2015). Contralateral cerebello-thalamo-cortical pathways with
prominent involvement of associative areas in humans in vivo.
Brain Structure and Function, 220(6), 3369–3384. https://doi
.org/10.1007/s00429-014-0861-2, PubMed: 25134682
Peterburs, J., Liang, Y., Cheng, D. T., & Desmond, J. E. (2021).
Sensory acquisition functions of the cerebellum in verbal work-
ing memory. Brain Structure and Function, 226(3), 833–844.
https://doi.org/10.1007/s00429-020-02212-5, PubMed:
33481104
Pu, M., Heleven, E., Delplanque, J., Gibert, N., Ma, Q., Funghi, G.,
& Van Overwalle, F. (2020). The posterior cerebellum supports
the explicit sequence learning linked to trait attribution. Cogni-
tive Affective & Behavioral Neuroscience, 20(4), 798–815.
https://doi.org/10.3758/s13415-020-00803-7, PubMed:
32495270
Ramnani, N. (2014). Automatic and controlled processing in the
corticocerebellar system. Progress in Brain Research, 210,
255–285. https://doi.org/10.1016/ B978-0-444-63356-9.00010
-8, PubMed: 24916296
Ramnani, N. (2006). The primate cortico-cerebellar system:
Anatomy and function. Nature Reviews Neuroscience, 7(7),
511–522. https://doi.org/10.1038/nrn1953, PubMed: 16791141
Ramnani, N. (2012). Frontal lobe and posterior parietal contri-
butions to the cortico-cerebellar system. Cerebellum, 11(2),
366–383. https://doi.org/10.1007/s12311-011-0272-3,
PubMed: 21671065
Raymond, J. L., & Medina, J. F. (2018). Computational principles of
supervised learning in the cerebellum. Annual Review of Neuro-
science, 41, 233–253. https://doi.org/10.1146/annurev-neuro
-080317-061948, PubMed: 29986160
Reveley, C., Seth, A. K., Pierpaoli, C., Silva, A. C., Yu, D., Saunders,
R. C., Leopold, D. A., & Ye, F. Q. (2015). Superficial white matter
fiber systems impede detection of long-range cortical connec-
tions in diffusion MR tractography. Proceedings of the National
Academy of Sciences, 112(21), E2820–E2828. https://doi.org/10
.1073/pnas.1418198112, PubMed: 25964365
Riedel, M. C., Ray, K. L., Dick, A. S., Sutherland, M. T., Hernandez,
Z., Fox, P. M., Eickhoff, S. B., Fox, P. T., & Laird, A. R. (2015).
Meta-analytic connectivity and behavioral parcellation of the
human cerebellum. NeuroImage, 117, 327–342. https://doi.org
/10.1016/j.neuroimage.2015.05.008, PubMed: 25998956
Rogalsky, C., Matchin, W., & Hickok, G. (2008). Broca’s area, sen-
tence comprehension, and working memory: An fMRI study.
Neurobiology of Language
22
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
8
5
2
0
5
8
2
9
1
n
o
_
a
_
0
0
0
8
5
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Cerebellar white matter and language
Frontiers in Human Neuroscience, 2, 14. https://doi.org/10.3389
/neuro.09.014.2008, PubMed: 18958214
Salman, M. S. (2002). The cerebellum: It’s about time! But timing is
not everything: New insights into the role of the cerebellum in
timing motor and cognitive tasks. Journal of Child Neurology,
17(1), 1–9. https://doi.org/10.1177/088307380201700101,
PubMed: 11913561
Salmi, J., Pallesen, K. J., Neuvonen, T., Brattico, E., Korvenoja, A.,
Salonen, O., & Carlson, S. (2010). Cognitive and motor loops of
the human cerebro-cerebellar system. Journal of Cognitive Neu-
roscience, 22(11), 2663–2676. https://doi.org/10.1162/jocn.2009
.21382, PubMed: 19925191
Schilling, K. G., Petit, L., Rheault, F., Remedios, S., Pierpaoli, C.,
Anderson, A. W., Landman, B. A., & Descoteaux, M. (2020).
Brain connections derived from diffusion MRI tractography can
be highly anatomically accurate—If we know where white mat-
ter pathways start, where they end, and where they do not go.
Brain Structure and Function, 225(8), 2387–2420. https://doi
.org/10.1007/s00429-020-02129-z, PubMed: 32816112
Schmahmann, J. D. (1996). From movement to thought: Anatomic
substrates of the cerebellar contribution to cognitive processing.
Human Brain Mapping, 4(3), 174–198. https://doi.org/10.1002
/(SICI )1097-0193(1996)4:3<174::AID-HBM3>3.0.一氧化碳;2-0,
考研: 20408197
Schmahmann, J. D ., & Pandya, D. 氮. (1991). Projections to the
basis pontis from the superior temporal sulcus and superior
temporal region in the rhesus monkey. Journal of Comparative
Neurology, 308(2), 224–248. https://doi.org/10.1002/cne
.903080209, 考研: 1716269
Schmahmann, J. D ., & Pandya, D. 氮. (1997). The cerebrocere-
bellar system. International Review of Neurobiology, 41, 31–60.
https://doi.org/10.1016/S0074-7742(08)60346-3, 考研:
9378595
Schmahmann, J. D ., & Sherman, J. C. (1998). The cerebellar cogni-
tive affective syndrome. Brain, 121(部分 4), 561–579. https://土井
.org/10.1093/brain/121.4.561, 考研: 9577385
Schwartze, M。, & Kotz, S. A. (2016). Contributions of cerebellar
event-based temporal processing and preparatory function to
speech perception. Brain and Language, 161, 28–32. https://土井
.org/10.1016/j.bandl.2015.08.005, 考研: 26362972
Silveri, 中号. C. (2021). Contribution of the cerebellum and the basal
ganglia to language production: Speech, word fluency, 和
sentence construction—evidence from pathology. 小脑,
20(2), 282–294. https://doi.org/10.1007/s12311-020-01207-6,
考研: 33120434
Silveri, 中号. C。, Leggio, 中号. G。, & Molinari, 中号. (1994). The cerebel-
lum contributes to linguistic production: A case of agrammatic
speech following a right cerebellar lesion. Neurology, 44(11),
2047–2050. https://doi.org/10.1212/ WNL.44.11.2047,
考研: 7969957
史密斯, S. M。, Vidaurre, D ., 贝克曼, C. F。, Glasser, 中号. F。, 詹金森,
M。, 磨坊主, K. L。, Nichols, 时间. E., 罗宾逊, 乙. C。, Salimi-Khorshidi,
G。, 伍尔里奇, 中号. W., Barch, D. M。, Uğurbil, K., & Van Essen,
D. C. (2013). Functional connectomics from resting-state fMRI.
认知科学的趋势, 17(12), 666–682. https://doi.org/10
.1016/j.tics.2013.09.016, 考研: 24238796
Smits, M。, Jiskoot, L. C。, & Papma, J. 中号. (2014). White matter tracts
of speech and language. Seminars in Ultrasound, CT, and MR,
35(5), 504–516. https://doi.org/10.1053/j.sult.2014.06.008,
考研: 25217302
Sokolov, A. A。, Erb, M。, Grodd, W., & Pavlova, 中号. A. (2014).
Structural loop between the cerebellum and the superior tempo-
ral sulcus: Evidence from diffusion tensor imaging. Cerebral
Cortex, 24(3), 626–632. https://doi.org/10.1093/cercor/bhs346,
考研: 23169930
Sørensen, 时间. (1948). A method of establishing groups of equal
amplitude in plant sociology based on similarity of species con-
tent and its application to analyses of the vegetation on Danish
commons. Biologiske Skrifter/Kongelige Danske Videnskabernes
Selskab, 5, 1–34.
索蒂罗普洛斯, A. N。, Jbabdi, S。, 徐, J。, 安德森, J. L。, Moeller, S。,
Aurbach, 乙. J。, Glasser, 中号. F。, Hernandez, M。, Sapiro, G。,
詹金森, M。, 范伯格, A。, Yacoub, E., Lenglet, C。, Van Essen,
D. C。, Uğurbil, K., & 贝伦斯, 时间. 乙. J. (2013). Advances in diffu-
sion MRI acquisition and processing in the Human Connectome
项目. 神经影像, 80, 125–143. https://doi.org/10.1016/j
.neuroimage.2013.05.057, 考研: 23702418
Spencer, K. A。, & Slocomb, D. L. (2007). The neural basis of ataxic
dysarthria. 小脑, 6(1), 58–65. https://doi.org/10.1080
/14734220601145459, 考研: 17366266
Steele, C. J。, 安旺德, A。, Bazin, P.-L., Trampel, R。, Schaefer, A。,
车工, R。, Ramnani, N。, & 狂野的召唤者, A. (2017). Human cerebel-
lar sub-millimeter diffusion imaging reveals the motor and
non-motor topography of the dentate nucleus. 大脑皮层,
27(9), 4537–4548. https://doi.org/10.1093/cercor/ bhw258,
考研: 27600851
Stoodley, C. J。, & Tsai, 磷. 时间. (2021). Adaptive prediction for social
上下文: The cerebellar contribution to typical and atypical
social behaviors. Annual Review of Neuroscience, 44,
475–493. https://doi.org/10.1146/annurev-neuro-100120
-092143, 考研: 34236892
针织, 磷. L。, Dum, 右. P。, & Fiez, J. A. (2009). Cerebellum and non-
motor function. Annual Review of Neuroscience, 32, 413–434.
https://doi.org/10.1146/annurev.neuro.31.060407.125606,
考研: 19555291
Takaya, S。, Kuperberg, G. R。, 刘, H。, Greve, D. N。, 马克里斯, N。, &
Stufflebeam, S. 中号. (2015). Asymmetric projections of the arcuate
fasciculus to the temporal cortex underlie lateralized language
function in the human brain. Frontiers in Neuroanatomy, 9, 119.
https://doi.org/10.3389/fnana.2015.00119, 考研: 26441551
Taoka, T。, Kin, T。, Nakagawa, H。, Hirano, M。, Sakamoto, M。, Wada,
T。, Takayama, K., Wuttikul, C。, Iwasaki, S。, Ueno, S。, &
Kichikawa, K. (2007). Diffusivity and diffusion anisotropy of
cerebellar peduncles in cases of spinocerebellar degenerative
疾病. 神经影像, 37(2), 387–393. https://doi.org/10.1016/j
.neuroimage.2007.05.028, 考研: 17583535
托马斯, C。, 叶, F. Q., Irfanoglu, 中号. 奥。, Modi, P。, Saleem, K. S。,
Leopold, D. A。, & 皮尔保利, C. (2014). Anatomical accuracy of
brain connections derived from diffusion MRI tractography is
inherently limited. 美国国家科学院院刊
科学, 111(46), 16574–16579. https://doi.org/10.1073/pnas
.1405672111, 考研: 25368179
图尔尼耶, J. D ., 卡拉曼特, F。, & 康纳利, A. (2012). MRtrix:
Diffusion tractography in crossing fiber regions. 国际的
Journal of Imaging Systems and Technology, 22(1), 53–66.
https://doi.org/10.1002/ima.22005.
图尔尼耶, J.-D., Yeh, C.-H., 卡拉曼特, F。, 给, K.-H., 康纳利, A。,
& 林, C.-O. (2008). Resolving crossing fibres using constrained
spherical deconvolution: Validation using diffusion-weighted
imaging phantom data. 神经影像, 42(2), 617–625. https://
doi.org/10.1016/j.neuroimage.2008.05.002, 考研:
18583153
Tremblay, P。, & 迪克, A. S. (2016). Broca and Wernicke are dead, 或者
moving past the classic model of language neurobiology. Brain
and Language, 162, 60–71. https://doi.org/10.1016/j.bandl
.2016.08.004, 考研: 27584714
Neurobiology of Language
23
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
d
哦
我
/
我
.
/
/
1
0
1
1
6
2
n
哦
_
A
_
0
0
0
8
5
2
0
5
8
2
9
1
n
哦
_
A
_
0
0
0
8
5
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
Cerebellar white matter and language
Turken, A. U。, & Dronkers, 氮. F. (2011). The neural architecture of
the language comprehension network: Converging evidence
from lesion and connectivity analyses. Frontiers in Systems Neu-
roscience, 5, 1. https://doi.org/10.3389/fnsys.2011.00001,
考研: 21347218
Van Essen, D. C。, Uğurbil, K., 奥尔巴赫, E., Barch, D ., 贝伦斯,
时间. 乙. J。, Bucholz, R。, 张, A。, 陈, L。, 科尔贝塔, M。, Curtiss,
S. W., 笔下的, S。, 范伯格, D ., Glasser, 氮. H。, Herath,
A. C。, Larson-Prior, L。, 马库斯, D ., Michalareas, G。, Moeller,
S。, Oostenveld, R。, … WU-Minn HCP Consortium. (2012). 这
Human Connectome Project: A data acquisition perspective.
神经影像, 62(4), 2222–2231. https://doi.org/10.1016/j
.neuroimage.2012.02.018, 考研: 22366334
Van Ettinger-Veenstra, H。, McAllister, A。, Lundberg, P。, Karlsson, T。,
& Engström, 中号. (2016). Higher language ability is related to
angular gyrus activation increase during semantic processing,
independent of sentence incongruency. Frontiers in Human
神经科学, 10, 110. https://doi.org/10.3389/fnhum.2016
.00110, 考研: 27014040
Van Overwalle, F。, Ma, Q., & Heleven, 乙. (2020). The posterior crus
II cerebellum is specialized for social mentalizing and emotional
self-experiences: A meta-analysis. Social Cognitive and Affective
神经科学, 15(9), 905–928. https://doi.org/10.1093/scan
/nsaa124, 考研: 32888303
Van Overwalle, F。, Manto, M。, 卡塔内奥, Z。, Clausi, S。, Ferrari, C。,
Gabrieli, J. D. E., Guell, X。, Heleven, E., Lupo, M。, Ma, Q.,
Michelutti, 中号. Olivito, G。, Pu, M。, 米, L. C。, Schmahmann,
J. D ., Siciliano, L, Sokolov, A. A。, Stoodley, C. J。, van Dun, K.,
Vandervert, L。, & Leggio, 中号. (2020). Consensus paper: Cerebel-
lum and social cognition. 小脑, 19(6), 833–868. https://
doi.org/10.1007/s12311-020-01155-1, 考研: 32632709
Van Overwalle, F。, Manto, M。, Leggio, M。, & Delgado-García, J. 中号.
(2019). The sequencing process generated by the cerebellum
crucially contributes to social interactions. Medical Hypotheses,
128, 33–42. https://doi.org/10.1016/j.mehy.2019.05.014,
考研: 31203906
Van Overwalle, F。, Pu, M。, Ma, Q., 李, M。, Haihambo, N。, Baetens,
K., Deroost, N。, Baeken, C。, & Heleven, 乙. (2021). The involve-
ment of the posterior cerebellum in reconstructing and predicting
social action sequences. 小脑, 21(5), 733–741. https://土井
.org/10.1007/s12311-021-01333-9, 考研: 34694590
Vias, C。, & 迪克, A. S. (2017). Cerebellar contributions to language
in typical and atypical development: A review. Developmental
Neuropsychology, 42(6), 404–421. https://doi.org/10.1080
/87565641.2017.1334783, 考研: 28885046
Vicari, S。, Piccini, G。, Mercuri, E., Battini, R。, Chieffo, D ., Bulgheroni,
S。, Pecini, C。, Lucibello, S。, Lenzi, S。, Moriconi, F。, Pane, M。,
D’Amico, A。, Astrea, G。, Baranello, G。, Riva, D ., Cioni, G。, &
Alfieri, 磷. (2018). Implicit learning deficit in children with
Duchenne muscular dystrophy: Evidence for a cerebellar
cognitive impairment? PLOS One, 13(1), Article e0191164.
https://doi.org/10.1371/journal.pone.0191164, 考研:
29338029
Voevodskaya, 奥。, Simmons, A。, Nordenskjöld, R。, Kullberg, J。,
Ahlström, H。, Lind, L。, Wahlund, L.-O., Larsson, E.-M., 韦斯特曼,
E., & Alzheimer’s Disease Neuroimaging Initiative. (2014). 这
effects of intracranial volume adjustment approaches on multiple
regional MRI volumes in healthy aging and Alzheimer’s disease.
Frontiers in Aging Neuroscience, 6, 264. https://doi.org/10.3389
/fnagi.2014.00264, 考研: 25339897
Wakana, S。, Caprihan, A。, Panzenboeck, 中号. M。, Fallon, J. H。, Perry,
M。, Gollub, 右. L。, Hua, K., 张, J。, Jiang, H。, Dubey, P。, Blitz,
A。, van Zijl, P。, & 森, S. (2007). Reproducibility of quantitative
tractography methods applied to cerebral white matter. Neuro-
图像, 36(3), 630–644. https://doi.org/10.1016/j.neuroimage
.2007.02.049, 考研: 17481925
王, F。, Sun, Z。, Du, X。, 王, X。, Cong, Z。, 张, H。, 张,
D ., & 洪, D. (2003). A diffusion tensor imaging study of mid-
dle and superior cerebellar peduncle in male patients with
schizophrenia. Neuroscience Letters, 348(3), 135–138. https://
doi.org/10.1016/S0304-3940(03)00589-5, 考研: 12932812
Wolpert, D. M。, Miall, 右. C。, & Kawato, 中号. (1998). Internal models
in the cerebellum. 认知科学的趋势, 2(9), 338–347.
https://doi.org/10.1016/S1364-6613(98)01221-2, 考研:
21227230
WU-Minn HCP Consortium. (2015). Appendix I – Protocol
guidance and HCP session protocols. In WU-Minn HCP 900
subjects data release: Reference manual (PP. 1–55). WU-Minn
Consortium of the Human Connectome Project. https://万维网
.humanconnectome.org/storage/app/media/documentation/s900
/HCP_S900_Release_Appendix_I.pdf
哪个, Z。, 吴, P。, 翁, X。, & Bandettini, 磷. A. (2014). 小脑
engages in automation of verb-generation skill. 杂志
Integrative Neuroscience, 13(1), 1–17. https://doi.org/10.1142
/S0219635214500010, 考研: 24738536
Yarkoni, T。, Poldrack, R。, Nichols, T。, Van Essen, D ., & Wager, 时间.
(2011, September 4–6). NeuroSynth: A new platform for
large-scale automated synthesis of human functional neuroim-
aging data [Poster presentation]. 4th INCF Congress of Neuroin-
formatics, 波士顿, 嘛.
Neurobiology of Language
24
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
d
哦
我
/
我
/
.
/
1
0
1
1
6
2
n
哦
_
A
_
0
0
0
8
5
2
0
5
8
2
9
1
n
哦
_
A
_
0
0
0
8
5
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3