RESEARCH ARTICLE
Use of Longitudinal EEG Measures in Estimating
Language Development in Infants With
and Without Familial Risk for Autism
Spectrum Disorder
Carol L. Wilkinson1
, Laurel J. Gabard-Durnam1
, Kush Kapur2
, Helen Tager-Flusberg3,
April R. Levin2
, and Charles A. Nelson1
1Division of Developmental Medicine, Boston Children’s Hospital, 波士顿, 嘛
2神经内科, Boston Children’s Hospital, 波士顿, 嘛
3Department of Psychological and Brain Sciences, 波士顿大学, 波士顿, 嘛
关键词: autism spectrum disorder, electroencephalography (EEG), language development, bio-
标记, 婴儿, 孩子
抽象的
Language development in children with autism spectrum disorder (自闭症谱系障碍) varies greatly among
affected individuals and is a strong predictor of later outcomes. Younger siblings of children
with ASD have increased risk of ASD, but also language delay. Identifying neural markers
of language outcomes in infant siblings could facilitate earlier intervention and improved
结果. This study aimed to determine whether electroencephalography (EEG) 措施
from the first 2 years of life can explain heterogeneity in language development in children at
low and high risk for ASD, and whether associations between EEG measures and language
development are different depending on ASD risk status or later ASD diagnosis. 在这个
prospective longitudinal study, EEG measures collected between 3 和 24 months were used
in a multivariate linear regression model to estimate participants’ 24-month language
发展. Individual baseline longitudinal EEG measures included (1) the slope of EEG
power across 3 到 12 months or 3 到 24 months of life for six canonical frequency bands, (2)
the estimated EEG power at 6 months of age for the same frequency bands, 和 (3) 条款
representing the interaction between ASD risk status and EEG power measures. Modeled
24-month language scores using EEG data from either the first 2 年 (Pearson p = 0.70, 95%
−18) or the first year of life (Pearson p = 0.66, 95% CI [0.540,
CI [0.595, 0.783], p = 1 × 10
−14) were highly correlated with observed scores. All models included
0.761], p = 2.5 × 10
significant interaction effects of risk on EEG measures, suggesting that EEG-language
associations are different depending on risk status, and that different brain mechanisms affect
language development in low- versus high-risk infants.
介绍
Children with autism spectrum disorder (自闭症谱系障碍) have striking heterogeneity in their early lan-
guage development (Anderson et al., 2007; Kjelgaard & Tager-Flusberg, 2001; Pickles,
安德森, & Lord, 2014). Although many children present initially with language delays,
roughly one-fourth will develop age-appropriate skills by school age, 和 30% will remain
minimally verbal throughout life (Anderson et al., 2007; Tager-Flusberg & Kasari, 2013).
此外, language acquisition is one of the best predictors of later outcomes in children
开放访问
杂志
引文: Wilkinson, C. L。, Gabard-
Durnam, L. J。, Kapur, K., Tager-
Flusberg, H。, 莱文, A. R。, & 纳尔逊,
C. A. (2020). Use of longitudinal EEG
measures in estimating language
development in infants with and
without familial risk for autism
spectrum disorder. Neurobiology of
语言, 1(1), 33–53. https://doi.org/
10.1162/nol_a_00002
DOI:
https://doi.org/10.1162/nol_a_00002
支持信息:
https://doi.org/10.1162/nol_a_00002
已收到: 29 行进 2019
公认: 27 九月 2019
利益争夺: 作者有
声明不存在竞争利益
存在.
通讯作者:
Carol L. Wilkinson
Carol.wilkinson@childrens.harvard.
edu
处理编辑器:
Kate Watkins
版权: © 2019
麻省理工学院
在知识共享下发布
归因 4.0 国际的
(抄送 4.0) 执照.
麻省理工学院出版社
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Use of longitudinal EEG to estimate language in infants with ASD risk
with ASD (Billstedt, Gillberg, & Gillberg, 2005; Gotham, Pickles, & Lord, 2012; 磨坊主
等人。, 2017; Szatmari et al., 2000, 2009). Although children with ASD are clearly at risk
for language delay, so are their younger siblings; 例如, longitudinal studies follow-
ing infant siblings of children with ASD (high-risk infants) have reported delays in language
skills as early as 12 months of age in infant siblings, both with and without later diagnoses
of ASD (Estes et al., 2015; Landa & Garrett-Mayer, 2006; Marrus et al., 2018; 斯旺森
等人。, 2017). 像这样, earlier identification of future language delays in toddlers at risk
for ASD would facilitate earlier intervention and increase the likelihood of improving
结果.
Longitudinal large population-based studies have identified various risk factors on the
个人 (性别, prematurity), familial (maternal education), and environmental (收入)
levels that influence language development (Reilly et al., 2007; Schjølberg, Eadie,
Zachrisson, Oyen, & 事先的, 2011). 然而, across studies these factors consistently account
for 4%–7% of variance in language outcomes in 2-year-olds, suggesting that additional factors
play an important role in a child’s early language trajectory. 的确, high-risk infants who do
not have ASD still have a fourfold increased risk of language delay compared to infants with-
out a family history of ASD, suggesting that early language delay is an endophenotype of ASD
with shared genetic liability and possibly shared underlying biology (Marrus et al., 2018).
Therefore studying the neural mechanisms underlying language development in high-risk chil-
dren may shed light on early language delay as a whole, and allow for earlier detection before
delays in language can be measured behaviorally.
To meet this goal, researchers have applied neuroimaging techniques to measure brain-
based changes that support language development in low- and high-risk infants over the first
3 years after birth. A number of studies suggest that the relationship between brain measures
and language outcomes are different depending on ASD diagnosis or ASD risk status
(Lombardo et al., 2015; Seery, Tager-Flusberg, & 纳尔逊, 2014; Swanson et al., 2017). 为了
例子, in infants who show a language delay at a later age, the relationship between lan-
guage ability and reactivity to speech (as measured on fMRI) is reversed depending on ASD
diagnosis (Lombardo et al., 2015). 相似地, using EEG in low- and high-risk infants, signifi-
cant relationships between frontal brain activity and language ability have been observed in
high-risk but not low-risk infants at both 9 和 24 月 (Seery et al., 2014; Wilkinson, 莱文,
Gabard-Durnam, Tager-Flusberg, & 纳尔逊, 2019). These differential brain-behavior associa-
tions suggest that the neural mechanisms affecting language development in high-risk children
may be different than in low-risk children. One reason for this could relate to global brain
differences such as reduced power across frequency bands observed as early as 3 到 6 月
in high-risk infants regardless of ASD outcome (莱文, Varcin, O’Leary, Tager-Flusberg, &
纳尔逊, 2017; Riva et al., 2018; Tierney, Gabard-Durnam, Vogel-Farley, Tager-Flusberg, &
纳尔逊, 2012), or accelerated surface area brain growth observed between 6 和 12 月
of age in high-risk infants with later ASD diagnosis (Hazlett et al., 2017). These early brain
differences could alter typical development of language circuitry. 鉴于这种, predictive
models of language must account for these possible differences between low- and high-risk
人口.
EEG, which measures network level brain activity at the scalp surface, has particular
promise in its use as a clinical biomarker of language development in infants and toddlers as
it is noninvasive, relatively low cost, and already regularly used in the outpatient setting for sei-
zure monitoring (Jeste, Frohlich, & Loo, 2015; 莱文 & 纳尔逊, 2015; Varcin & 纳尔逊, 2016).
Various measures collected during task-related or resting-state EEG have been associated with
Neurobiology of Language
34
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Use of longitudinal EEG to estimate language in infants with ASD risk
language processing and language development in young children. Evoked brain responses to
auditory stimuli presented as early as 6 months of age reflect both current language processing
and future language ability (Jansson-Verkasalo et al., 2010; Kuhl, 2010; Kuhl, Conboy, Padden,
纳尔逊, & Pruitt, 2005). 此外, phase-locking, or entrainment, to both the temporal syllabic
structure of speech (in the delta/theta range 1–8 Hz) and the phonetic structure of speech (在里面
beta/gamma range 20–50 Hz) has been observed in animals and humans during speech process-
英, and is correlated with measures of phonological processing (Giraud & Poeppel, 2012).
Although it is still unknown whether aberrant entrainment in these frequency bands affects lan-
guage acquisition in children, studies in children and adults with ASD have reported differences
in resting and task-evoked theta and gamma power (Jochaut et al., 2015; Rojas & Wilson, 2014;
王等人。, 2013), and it is hypothesized that such differences impair speech processing and
delay language acquisition (Jochaut et al., 2015). Clinically, resting EEG (non-task-related) 是个
simplest to collect, especially in young children or in populations for which longer task-related
paradigms are not well tolerated. 此外, associations between resting frontal gamma power
and language have been observed. 具体来说, resting (IE。, not task-related) frontal gamma
power has been associated positively with receptive and expressive language ability in typically
developing 3- to 4-year-olds (Benasich, Gou, Choudhury, & 哈里斯, 2008; Gou, Choudhury, &
Benasich, 2011; Tarullo et al., 2017), but negatively associated with expressive language in
high-risk 2-year-olds (Wilkinson et al., 2019). Reduced spectral power in delta/theta and gamma
bands during visual processing has also been observed in minimally verbal children with ASD
compared to a typically developing control group (Ortiz-Mantilla, Cantiani, Shafer, & Benasich,
2019). The spectral power of resting EEG has also been evaluated in children with dyslexia, 和
reduced frontal delta power observed in 3-year-olds who are later diagnosed with dyslexia com-
pared with controls (Schiavone et al., 2014), but increased delta and theta power in school-aged
children with dyslexia (Colon, Notermans, de Weerd, & Kap, 1979; Harmony et al., 1995;
Penolazzi, Spironelli, & Angrilli, 2008; Spironelli, Penolazzi, Vio, & Angrilli, 2006), suggesting
that developmental changes in EEG spectral power are also likely relevant predictors of
language-related outcomes.
Although the preceding investigations have found associations between specific frequency
bands and language development in both typically developing and neurodevelopmental dis-
order populations, they have not accounted for the majority of variance in language scores
between individuals. This is likely because by focusing on a single frequency band during a
limited developmental age range, we fail to capture the impact of developmental changes
across the spectrum of frequency bands as it relates to language development.
This study had three main objectives. 第一的, we aimed to evaluate whether a combination of
early and longitudinal EEG measures (spectral power at 6 months of age and the developmen-
tal trajectory of spectral power) can explain the variance in language ability at 2 years of age in
both low- and high-risk children. 换句话说, in a nonclinical longitudinal data sample,
can resting EEG measures sufficiently estimate language ability in toddlers, to support its future
use as a clinical biomarker of language development? 第二, to begin to develop a theoret-
ical framework of how neural oscillations (such as theta and gamma) may influence language
发展, we used a data-driven approach to model building, and then investigated
whether associations between frequency band power and language ability were different be-
tween low- and high-risk infants. 具体来说, in this second aim, we explored whether differ-
ent EEG measures predict language ability in these risk groups, and whether the direction of
these relationships was similar or different between groups. Because differences in EEG power
have been observed as early as 3 到 6 months of age in high-risk infants, we hypothesized that
spectral power at 6 months of age would differentially contribute to model language estimates
Neurobiology of Language
35
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Use of longitudinal EEG to estimate language in infants with ASD risk
in low- versus high-risk groups (LR and HR, 分别). 此外, we hypothesized that
frequency bands important for speech processing (delta/theta and beta/gamma) would most
robustly contribute to language estimates across low- and high-risk groups. 最后, 我们
asked whether these brain-language associations were also different between high-risk in-
fants with and without ASD (HR-ASD and HR-NoASD, 分别).
方法
Study Design and Population
Participants were part of a prospective longitudinal study of early neurocognitive development of infant
siblings of children with ASD, conducted at Boston University and Boston Children’s Hospital. 这
study was approved by the institutional review board (#X06-08-0374) and written informed
consent was obtained from a parent or guardian prior to each child’s participation in the study.
High-risk infants were defined as having at least one full sibling with a community diagno-
sis of ASD, which was confirmed using the Social Communication Questionnaire (Rutter
贝利 & Lord, 2003) and/or the Autism Diagnostic Observation Schedule (ADOS; Lord &
Rutter, 2012). Low-risk infants were defined by having a typically developing older sibling
and no first- or second-degree family members with ASD. All infants had a minimum gesta-
tional age of 36 weeks, no history of prenatal or postnatal medical or neurological problems,
and no known genetic disorders. For this analysis, all infants were also from households speak-
ing primarily English (English spoken more than 75% 当时的).
A total of 220 participants were enrolled who met the above inclusion and exclusion
criteria (97LR, 123 人力资源). Only a portion of enrolled participants had sufficient quality EEG re-
corded (see EEG Rejection Criteria), developmental testing completed at the 24-month visit,
and ADOS completed at 24 和/或 36 月. 此外, three low-risk male participants
went on to meet Diagnostic and Statistical Manual of Mental Disorders, Fifth Edition (DSM-5)
criteria for ASD and were not included in this analysis. This study includes data from a total of
58 LR and 72 HR infants (桌子 1). 的 72 HR infants, 21 (29%) met criteria for ASD based
on assessments described in Language and Behavioral Assessments (桌子 1).
Measures
Language and behavioral assessments
At multiple time points, participants were assessed using the Mullen Scales of Early Learning
(MSEL). For this analysis, the MSEL Verbal Developmental Quotient (VDQ) 在 24 months was
calculated from the receptive and expressive language subscales. This time point, instead of the
later 36-month time point, was used as it included the most participants enrolled in the study.
Final ASD outcomes were determined for all infants using the ADOS administered at 24 和 36
months of age. For those children meeting criteria on the ADOS, or coming within 3 points of
cutoffs, a licensed clinical psychologist reviewed scores and video recordings of concurrent and
previous behavioral assessments, and using DSM-5 criteria provided a best estimate clinical
判断.
EEG data collection
Baseline, non-task-related EEG data were collected at 6 访问 (3, 6, 9, 12, 18, 和 24 months of
年龄). The infant was held by their seated caregiver in a dimly lit, sound-attenuated, electrically
shielded room while a research assistant ensured that the infant remained calm and still by
blowing bubbles and/or showing toys. Continuous EEG was recorded for 2 到 5 minutes.
Neurobiology of Language
36
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
桌子 1. Sample characteristics
LR
N= 58
30中号, 28F
HR-NoASD
N= 51
25中号, 26F
HR-ASD
N= 21
14中号, 7F
p Value by outcome (LR vs
HR-NoASD vs HR-ASD)
0.4
0.060
p Value by risk
(LR vs HRa)
0.86
0.04
Use of longitudinal EEG to estimate language in infants with ASD risk
Sexb
Parental educationb, n (%)
Not answered
<4-year college degree 4-year degree >4-year college degree
Household incomeb, n (%)
Not answered
<$75,000 >$75,000
Raceb, n (%)
Non-white
Ethnicityb, n (%)
6 (10)
3 (5)
10 (17)
39 (67)
8 (14)
9 (15)
2 (4)
10 (20)
10 (20)
29 (57)
3 (6)
6 (12)
5 (24)
3 (14)
6 (29)
7 (33)
6 (29)
2 (9)
41 (71)
42 (82)
13 (62)
7 (12)
3 (6)
4 (19)
Hispanic or Latino
1 (2)
3 (6)
3 (14)
Number of EEG time pointsc
4.03 ± 1.1
3.64 ± 1.2
3.9 ± 1.3
Mean ± SD
24m MSEL VDQd Mean ± SD
118.0 ± 15.3
112.0 ± 14.3
96 ± 22.6
24m MSEL NVDQd Mean ± SD
110.1 ± 15.0
107.0 ± 11.9
97 ± 22.6
24m ADOS severity scoree
1.61 ± 0.86
1.91 ± 1.13
5.25 ± 2.47
Mean ± SD
Participant EEG data included in analysis, n (%)
3 月
6 月
9 月
12 月
18 月
24 月
9 (15)
44 (76)
48 (83)
52 (90)
37 (64)
44 (76)
15 (29)
30 (59)
37 (73)
33 (65)
32 (62)
39 (77)
7 (33)
13 (62)
15 (71)
19 (91)
14 (67)
14 (67)
0.82
0.44
0.28
0.08
0.23
0.0007
0.02
0.0001
0.57
0.13
0.13
0.03
0.14
0.003
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
笔记. ASD = autism spectrum disorder; LR = low-risk without ASD; HR-NoASD = high-risk without ASD; HR-ASD = high-risk with ASD; ADOS = Autism
Diagnostic Observation Schedule.
aHR = HR-NoASD + HR-ASD; n = 72. bFisher exact. cOne-way ANOVA. dOne-way ANOVA with parental education as covariate. eKruskal-Wallis.
Neurobiology of Language
37
Use of longitudinal EEG to estimate language in infants with ASD risk
EEG data were collected using either a 64-channel Geodesic Sensor Net System or a 128-
channel Hydrocel Geodesic Sensor Nets (Electrical Geodesics, 公司, 尤金, 或者) 连接的
to a DC-coupled amplifier (Net Amps 200 or Net Amps 300, Electrical Geodesics, 公司). 那里
was no difference in distribution of net type at each time point between outcome groups. 数据
were sampled at 250 Hz or 500 赫兹, and collected using a 0.1-Hz high-pass analog (IE。, 难的-
器皿) filter and referenced online to a single vertex electrode (Cz), with impedances kept be-
low 100kΩ. Electrooculographic electrodes were removed to improve the child’s comfort.
EEG preprocessing
Raw EEG data collected in NetStation (Electrical Geodesics, 公司) were exported to MATLAB
(version R2017a) for preprocessing and subsequent power analysis.
All files were batch processed using the Batch EEG Automated Processing Platform (莱文,
Méndez Leal, Gabard-Durnam, & O’Leary, 2018), to ensure uniform analysis regardless of
when the EEG was acquired or which risk group they were in.
A 1-Hz high-pass filter and 100-Hz low-pass filter were applied and then data sampled at
500 Hz were resampled using interpolation to 250 赫兹. Both experimental and participant-
induced artifacts were then identified and removed using the Harvard Automated Pre-
processing Pipeline for EEG (HAPPE; Gabard-Durnam, Mendez Leal, Wilkinson, & 莱文,
2018). HAPPE is a MATLAB-based preprocessing pipeline optimized for developmental data
with short recordings and/or high levels of artifact, to automate preprocessing and artifact re-
moval, and to evaluate data quality in the processed EEG recordings (Gabard-Durnam et al.,
2018). HAPPE artifact identification and removal includes removing 60-Hz line noise, 坏的
channel rejection, and participant-produced artifact (eye blinks, movement, and muscle activ-
性) through wavelet-enhanced independent component analysis (ICA) and multiple artifact re-
jection algorithm (MARA; Winkler, Debener, 穆勒, & Tangermann, 2015; Winkler, Haufe, &
Tangermann, 2011). MARA has excellent detection and removal of muscle artifact compo-
尼特, which can affect higher frequency band signal (Gabard-Durnam et al., 2018; Winkler
等人。, 2011). The following channels, in addition to the 10–20 electrodes, were used for
MARA: 64-channel net – 2, 3, 8, 9, 12, 16, 21, 25, 50, 53, 57, 58; and 128-channel net – 3,
4, 13, 19, 20, 23, 27, 28, 40, 41, 46, 47, 75, 98, 102, 103, 109, 112, 117, 118, 123. 后
artifact removal using HAPPE, data were re-referenced to an average reference (calculated
using the same channels used for MARA), detrended using the signal mean, and then regions
of high-amplitude signal (> 40 uV was used to account for the reduce signal amplitude which
occurs during the wavelet-enhanced ICA step of HAPPE preprocessing) were removed prior
to segmenting the remaining data into 2 s windows to allow for power calculations using
multitaper spectral analysis (Babadi & 棕色的, 2014). Noncontinuous data were not
concatenated.
EEG rejection criteria
HAPPE data output quality measures were used to systematically reject poor quality data that
were unfit for further analyses. EEG recordings were rejected if they had fewer than 20 细分市场
(40 s of total EEG), or were more than 3 standard deviations (标清) from the mean on the following
HAPPE data quality output parameters: percent good channels (3 标清: < 82%), mean retained
artifact probability (3 SD: > 0.3), median retained artifact probability (3 标清: > 0.35), 的百分比
independent components rejected as artifact (3 标清: > 84%), and percent of EEG signal vari-
ance retained after artifact removal (3 标清: < 32%). Based on the preceding criteria, 69 of 674
(10.2%) of EEG recordings collected between 3 and 24 months were rejected. In addition, any
Neurobiology of Language
38
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
o
_
a
_
0
0
0
0
2
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Use of longitudinal EEG to estimate language in infants with ASD risk
EEG with a frequency band power greater than 2 SD above or below the mean of their asso-
ciated outcome group (LR, HR-NoASD, or HR-ASD) mean were reviewed, resulting in the re-
jection of an additional 23 EEG studies (3.4%). HAPPE quality metrics and visual inspection
rejection rates did not differ significantly between groups (all p’s > 0.1). A full description of
HAPPE quality metrics and visualization based on this longitudinal dataset were published
previously (Gabard-Durnam et al., 2018). Gabard-Durnam et al. (2018) also provide examples
of EEG recordings from this dataset pre- and post-HAPPE processing, power spectra after each
step of processing, and comparison of HAPPE versus other approaches to artifact rejection
using this dataset.
EEG power analysis
A multitaper fast Fourier transform, using three orthogonal tapers (Thomson, 1982) was used to
decompose EEG signal into power for each 2 s segment of data for selected frontal electrodes:
64-channel net – 2, 3, 8, 9, 12, 13, 58, 62; and 128-channel net – 3, 4, 11, 19, 20, 23, 24, 27,
118, 123, 124 (Supplemental Figure 1). Frontal electrodes chosen a priori based on findings
from our previous work (Levin et al., 2017; Tierney et al., 2012; Wilkinson et al., 2019) 和
that of others (Benasich et al., 2008; Gou et al., 2011; Schiavone et al., 2014) 表明
frontal power in both low- and high-frequency bands is associated with language development
in typically developing and ASD populations. The summed power was calculated across all
frequencies within commonly used frequency bands in infant EEG studies: delta (2–3.99 Hz),
theta (4–5.99 Hz), low-alpha (6–8.99 Hz), high-alpha (9–12.99 Hz), beta (13–29.99 Hz), 和
gamma (30–50 Hz). As a minor point, we opted not to examine power < 2 Hz to allow use of
a 1-Hz high-pass filter required by HAPPE. Power was then averaged across all 2 s seg-
ments and then across the frontal electrodes for each participant to obtain their average
frontal power at each time point. We report absolute power values, normalized by a log
10 transform. Power spectra at each time point for low- and high-risk groups are shown in
Supplemental Figure 2.
EEG data reduction
Longitudinal baseline EEG data from participants were reduced to two parameters for each
frequency band per participant: (a) estimated power at 6 months (intercept at 6 months) and
(b) estimated linear slope of power over logarithmic age, calculated either over 3 to 24 months
(Models 1 and 2) or 3 to 12 months (Model 3). Given the nonlinear, logarithmic trajectories of
EEG power over early brain development observed in this dataset (Figure 1) and by other
labs (Cornelissen, Kim, Purdon, Brown, & Berde, 2015; Jing, Gilchrist, Badger, & Pivik, 2010),
we employed the ordinary least squares to model a linear relationship between the log-
transformed EEG power as a function of log-age for each individual. Ordinary least squares
regression equations utilized all the EEG data available from each individual from 3 to 24
months. This linear transform allowed us to include longitudinal EEG data from any individual
with EEG recordings from at least two time points across the 3- to 24-month period (Models 1
and 3) or 3- to 12-month period (Model 2), allowing us to maximize the sample size included in
the model. The average number of EEG studies from each participant was 3.86 ± 1.2 and did
not differ significantly between groups (Table 1; p > 0.1). The estimated 6-month intercept was
used instead of a 3-month intercept because not all infants were enrolled at 3 months of age (这
time of enrollment changed from 6 到 3 months during the study), and we aimed to use a time
point that was inclusive of the full group’s study visit parameters.
Neurobiology of Language
39
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Use of longitudinal EEG to estimate language in infants with ASD risk
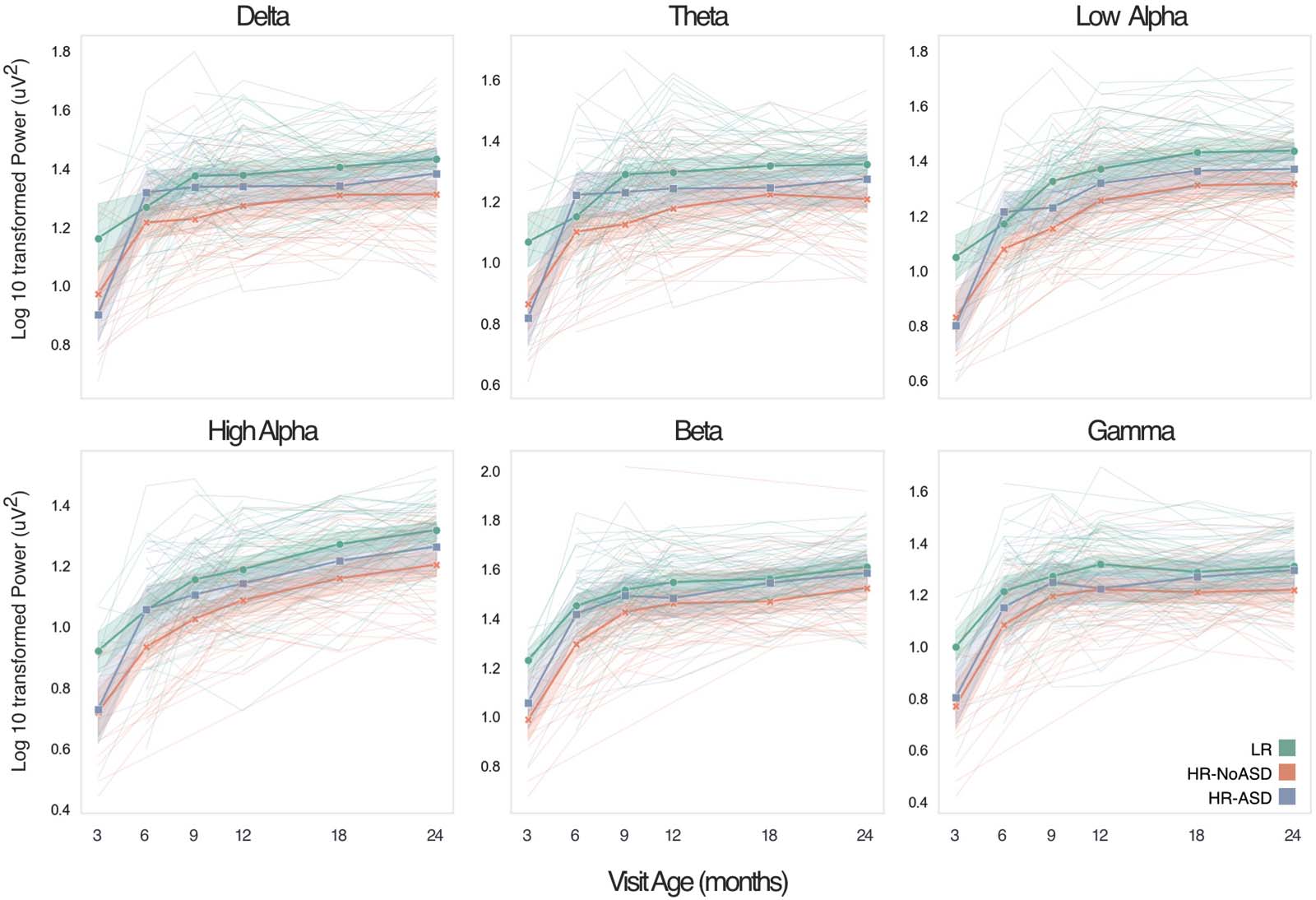
数字 1. Developmental trajectories of frontal electroencephalography (EEG) power across multiple frequency bands from 3 到 24 月.
Longitudinal trajectories of log 10 transformed absolute frontal EEG power across six frequency bands for individuals from low-risk (LR; 绿色的,
n = 58), high-risk without ASD (HR-NoASD; orange, n = 51), and high-risk with ASD (HR-ASD; 蓝色的, n = 21) are shown. Both individual and
mean trajectories by group are shown. Shaded regions represent the 95% confidence interval.
Statistical Analysis
Group comparisons of descriptive characteristics
A chi-square test (or Fisher exact test in case the cell value was < 5) was used to characterize differences in the categorical demographic data between groups. One-way analysis of vari- ance (ANOVA) was used to assess group differences in average number of EEG studies per participant and MSEL scores. Kruskal-Wallis followed by post hoc Dunn’s test was performed on non-normally distributed ADOS severity scores. Model Development Multivariate linear regression was used to characterize the relationship between the EEG pa- rameters and 24-month language outcomes across groups. Three models were examined, all using 24-month VDQ as the dependent variable. Potential EEG predictors for each model in- cluded the 6-month intercepts, the slopes, and the interaction terms between intercept and slope for each of the six frequency bands (18 possible variables). For all adjusted models, pa- rental education (averaged maternal and paternal education when both available), which is known to affect language development, was included as a covariate. Given the marginally significant increased number of male participants in the HR-ASD group, sex was also included as a covariate in adjusted models. Unadjusted models (Supplemental Materials) included only EEG parameters and interactions with either risk or ASD outcome. Neurobiology of Language 40 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u n o / l / l a r t i c e - p d f / / / / / 1 1 3 3 1 8 6 7 8 7 6 n o _ a _ 0 0 0 0 2 p d . l f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 Use of longitudinal EEG to estimate language in infants with ASD risk Table 2. Predictive models of Mullen Verbal Developmental Quotient Model 1 (adjusted R2 = 0.329) Low risk High risk Difference (high – low risk) B Coefficient (SE) p Value B Coefficient (SE) p Value B Coefficient (SE) p Value 6-month intercept Delta Theta Low alpha High alpha Beta Gamma Slope Delta Theta Low alpha High alpha Beta Gamma Intercept × Slope Delta Theta Low alpha High alpha Beta Gamma Covariates 3.55 (6.03) −7.04 (6.22) − 7.92 (5.55) 0.08 (4.66) −0.76 (2.37) −2.91 (5.56) −13.50 (4.33) −0.21 (3.57) 42.55 (16.68) 0.56 0.26 − 0.15 0.98 0.75 0.60 0.002* 0.95 0.01* 20.07 (6.59) −24.47 (6.69) − 4.51 (4.40) −0.1 (2.86) −0.76 (2.37) −2.91 (5.56) −13.50 (4.33) 5.41 (2.66) 8.97 (8.54) −77.88 (19.78) <0.001* −17.19 (9.35) 0.003* <.001* − 0.31 0.97 0.75 0.60 0.002* 0.05 0.3 0.07 16.77 (6.18) 0.008* 16.77 (6.18) 0.008* 0.65 (0.32) 0.04 0.65 (0.32) 0.04 − − − − − − −3.28 (1.48) 0.03 −0.04 (0.87) 5.33 (1.24) <0.001* 0.81 (0.72) −1.33 (0.51) 0.01* −1.33 (0.51) − − 0.97 0.26 0.01 Sex, reference = female −6.96 (2.93) 0.02 −6.96 (2.93) 0.02 Parental education College degree 13.05 (4.91) <0.01* 13.05 (4.91) <0.01* >College degree
16.94 (4.37)
<0.001* 16.94 (4.37) <0.001* Neurobiology of Language 16.53 (8.15) −17.43 (8.26) − −3.41 (6.44) −0.18 (4.68) − − − 5.61 (4.13) −33.58 (18.02) 0.05 0.04 − 0.60 0.97 − − − 0.18 0.07 60.70 (20.29) 0.004* − − − − − − − − 3.24 (1.69) −4.53 (1.42) 0.06 0.002* − − − − − − − − 41 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . e d u n o / l / l a r t i c e - p d f / / / / / 1 1 3 3 1 8 6 7 8 7 6 n o _ a _ 0 0 0 0 2 p d . l f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 Use of longitudinal EEG to estimate language in infants with ASD risk Table 2. (continued) Model 2 (adjusted R2 = 0.249) Low risk High risk Difference (high – low risk) B Coefficient (SE) p Value B Coefficient (SE) p Value B Coefficient (SE) p Value 6-month intercept Delta Theta Low alpha High alpha Beta Gamma Slope Delta Theta Low alpha High alpha Beta Gamma Intercept × Slope Delta Theta Low alpha High alpha Beta Gamma Covariates −1.16 (5.48) 7.25 (7.33) −2.41 (3.86) − −4.28 (4.08) 5.45 (3.46) − 1.05 (1.93) 8.21 (5.01) − −19.09 (7.39) 6.11 (4.09) − − −0.70 (0.44) − 1.41 (0.49) −0.51 (0.32) 0.83 0.33 0.54 − 0.30 0.12 − 0.59 0.11 − 0.01 0.14 − − 0.12 − 0.005 0.12 11.17 (6.19) −16.07 (6.14) 1.76 (3.44) − 2.52 (2.86) 0.61 (2.87) − −2.75 (1.78) −3.06 (2.82) − −1.39 (7.65) 6.11 (4.09) − − 0.47 (0.21) − 0.22 (0.56) −0.51 (0.32) 0.08 0.01 0.61 − 0.38 0.83 − 0.13 0.28 − 0.86 0.14 − − 0.03 − 0.69 0.12 Sex, reference = female −7.14 (3.11) 0.02 −7.14 (3.11) 0.02 Parental Education College degree 5.73 (4.93) >College degree
13.40 (4.55)
0.24
0.004
5.73 (4.93)
13.40 (4.55)
0.24
0.004
12.33 (8.04)
−23.32 (9.44)
4.16 (5.24)
-
6.80 (4.28)
−4.84 (3.84)
-
−3.80 (2.57)
−11.27 (5.82)
-
0.13
0.02
0.43
-
0.12
0.21
-
0.14
0.06
-
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
17.70 (8.79)
0.048
-
-
-
1.17 (0.50)
-
−1.18 (0.62)
-
-
-
-
-
-
-
0.02
-
0.06
-
-
-
-
Neurobiology of Language
42
Use of longitudinal EEG to estimate language in infants with ASD risk
桌子 2. (continued)
模型 3 (adjusted R2 = 0.530)
No autism
Autism
Difference (autism − no autism)
B Coefficient (SE)
p Value
B Coefficient (SE)
p Value
B Coefficient (SE)
p Value
34.59 (14.37)
0.02
22.10 (15.53)
−53.11 (18.94)
0.008*
−37.16 (20.02)
6-month intercept
Delta
Theta
Low alpha
High alpha
Beta
Gamma
Slope
Delta
Theta
Low alpha
High alpha
Beta
Gamma
Intercept × Slope
Delta
Theta
Low alpha
High alpha
Beta
Gamma
Covariates
12.49 (5.78)
−15.95 (6.42)
−3.23 (3.91)
9.08 (4.58)
−7.13 (3.48)
7.66 (3.74)
-
−2.89 (4.34)
−13.04 (5.42)
9.19 (4.89)
12.39 (10.88)
5.26 (3.39)
-
-
0.04
0.02
0.41
0.06
0.047
0.048
-
0.51
0.02
0.07
0.26
0.13
-
-
7.12 (7.34)
9.08 (4.58)
20.73 (7.81)
−15.56 (6.19)
-
−23.17 (11.93)
4.97 (11.10)
9.19 (4.89)
28.84 (12.81)
−4.63 (6.19)
-
-
0.34
0.06
0.01
0.02
-
0.06
0.66
0.07
0.03
0.46
-
-
1.30 (0.40)
0.002*
1.30 (0.40)
0.002*
-
−1.61 (0.77)
-
-
0.04
-
-
−1.61 (0.77)
-
-
0.04
-
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
0.16
0.07
0.20
-
0.003*
0.003*
-
0.11
0.11
-
0.08
0.17
-
-
-
-
-
-
-
-
-
10.35 (7.97)
-
27.86 (8.63)
−23.22 (7.35)
-
−20.29 (12.29)
18.01 (10.92)
-
16.46 (9.02)
−9.89 (7.07)
-
-
-
-
-
-
-
-
-
性别, reference = female
−6.50 (3.62)
0.08
−6.50 (3.62)
0.08
Parental Education
College degree
9.25 (5.44)
>College degree
11.80 (4.64)
0.09
0.015
9.25 (5.44)
11.80 (4.64)
0.09
0.015
笔记. B Coefficient are scaled by a factor of 10. 例如, B Coefficient for the 6-month intercept represents change in Mullen Scales of Early Learning
Verbal Development Quotient for every 0.1uV increase in spectral power. p Values in bold are significant. SE = Standard error.
*Maintained significance after false discovery rate (FDR) correction, using an FDR at alpha = 0.05.
Neurobiology of Language
43
Use of longitudinal EEG to estimate language in infants with ASD risk
For each of the models, we performed data-driven selection from the potential parameters
using a hybrid stepwise approach and minimization of the Akaike information criterion (AIC)
(Hastie, Tibshirani, & 弗里德曼, 2009). This model selection uses an iterative process that
compares AIC values across candidate models in order to minimize both information loss
and overfitting. Once model selection was complete, secondary post hoc hypothesis testing
was performed simultaneously on all parameters to determine which parameters were rela-
tively robust predictors of MSEL VDQ and whether interaction effects were significant.
Correction for multiple comparisons within the secondary analysis was performed using
the Benjamini-Hochberg procedure (Benjamini & Hochberg, 1995) to limit the false discov-
ery rate to alpha level 0.05 across tests, and p values surviving significance after correction
are marked with an asterisk.
模型 1 included interactions between risk (LR vs. 人力资源) 和 15 的 18 EEG parameters cal-
culated from data collected between 3 和 24 月. 模型 2 included interactions between
风险 (LR vs. 人力资源) 和所有 18 EEG parameters calculated from data collected from 3 到 12 月
年龄. 模型 3, using data only from high-risk infants, included interactions between ASD
outcome and 12 的 18 EEG parameters (桌子 2). Due to the limited sample size, we could
not simultaneously include interactions between EEG parameters and both risk and ASD out-
come. In all models, B coefficients were scaled by a factor of 10, so that they represent the
change in MSEL score after a 0.1-unit change in EEG intercept or slope.
结果
Participant Characteristics
Demographic data are shown in Table 1 for the full sample, with statistical analyses comparing
both outcome groups (LR vs. HR-NoASD vs. HR-ASD) and risk groups (LR vs. 人力资源, with HR
consisting of combined HR-NoASD and HR-ASD infants).
Developmental EEG Trajectories
Developmental trajectories of EEG power across the six frequency bands are shown in
数字 1, subdivided by outcome group (LR, HR-NoASD, and HR-ASD).
Longitudinal EEG measures explain language variability in low- versus high-risk infants (模型 1)
To test whether EEG predictors of language were different in low- versus high-risk infants,
模型 1 allowed for potential two-way interactions between risk status and all EEG parameters
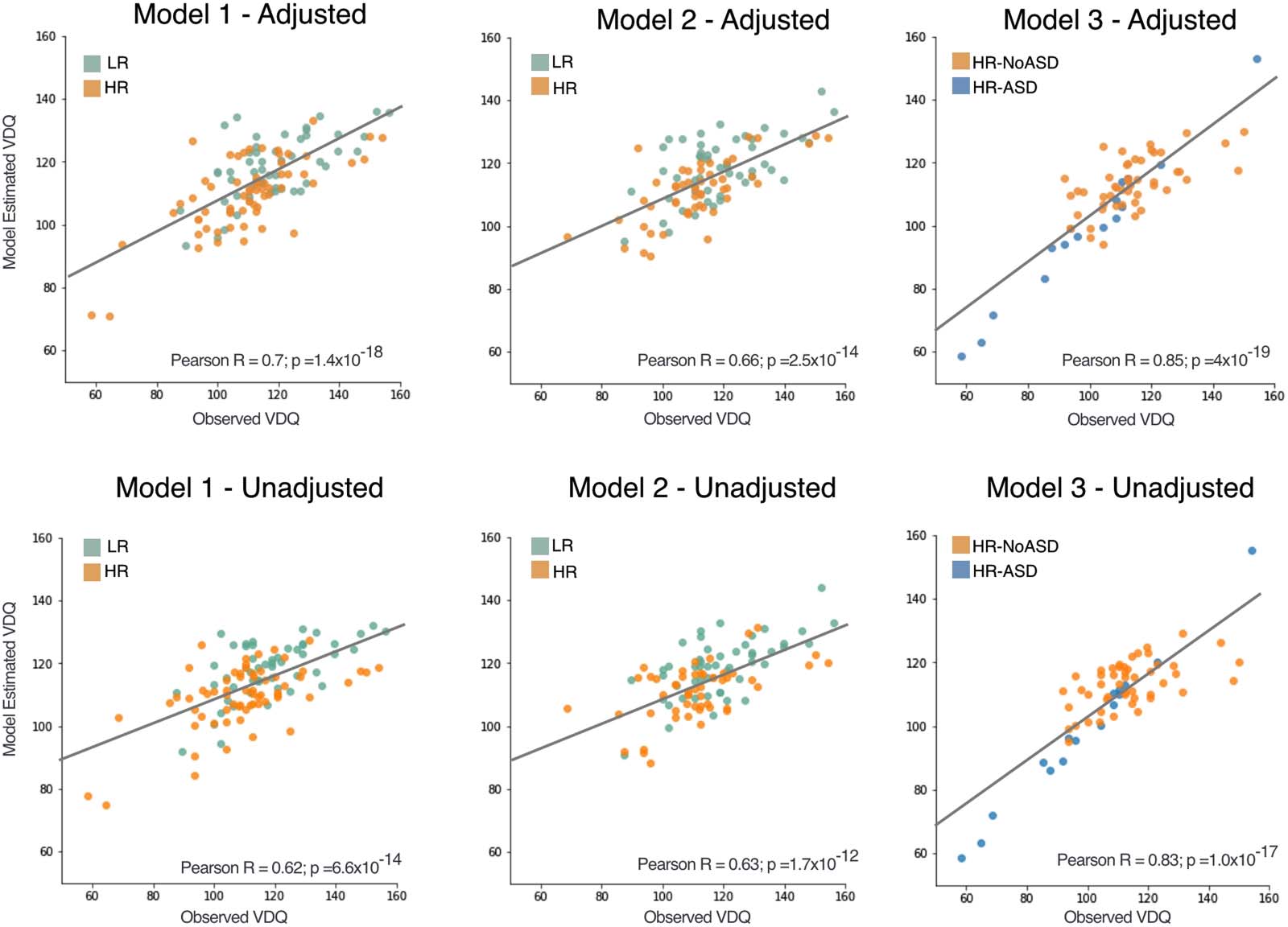
included in the model. Pearson correlations between the modeled language scores and ob-
served language scores were all significant, both collapsed across low- and high-risk subjects
−18) 什么时候
(Model Adjusted R2 = 0.329; Pearson R = 0.70, 95% CI [0.595, 0.783]; p = 1 × 10
−6) and high-risk (Pearson R =
low-risk (Pearson R = 0.617, 95% CI [0.414, 0.762]; p = 1 × 10
−11) groups were analyzed separately. Pearson corre-
0.709, 95% CI [0.542, 0.812]; p = 4 × 10
lation for the unadjusted model (Model Adjusted R2 = 0.211), using only EEG parameters, 曾是
−14). Scatterplots of
also significant (Pearson R = 0.62, 95% CI [0.497, 0.722]; p = 6.6 × 10
model-estimated and observed language scores are shown in Figure 2.
Significant differences between risk groups were observed for the effects of beta slope and
its interaction with its 6-month intercept as well as the effect of 6-month theta intercept on
language outcomes (桌子 2, 模型 1). Four EEG parameters were significantly associated
with language outcome in both low- and high-risk groups and had the same B coefficient:
theta slope, gamma slope, delta intercept × slope, and gamma intercept × slope. 尤其, 风险
Neurobiology of Language
44
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Use of longitudinal EEG to estimate language in infants with ASD risk
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
数字 2. Correlations of observed and model estimated verbal developmental quotient (VDQ). Observed and estimated Mullen Scales for
Early Learning VDQ for each model. Adjusted models include sex and parental education as covariates. Regression line shown represents
estimate based on all individuals included in the model. Each data point is colored based on the independent variable included in the model
(楷模 1 和 2 = risk status; 模型 3 = ASD diagnosis).
itself was not a significant predictor of 24-month language in either the adjusted or unadjusted
型号 (Adjusted Model 1: B coefficient = 19.31, SE 52.22; p = 0.71; Unadjusted Model 1: 乙
coefficient = −33.9, SE 58.64; p = 0.56). Parental education and sex covariates were signifi-
cant predictors in the adjusted model (桌子 2, 模型 1). The unadjusted Model 1 had similar
patterns of significance (data shown in Supplemental Table 1).
Estimating 2-year language from EEG measures over the first year of life (模型 2)
We next examined how well the longitudinal EEG measures restricted to visits from the first
year of life predicted language ability at 24 months across low- and high-risk infants. 模型 2
used EEG data from both low- and high-risk infants, but now restricted to data collected
之间 3 和 12 月. Pearson correlations between estimated and actual MSEL scores
were significant (Model Adjusted R2 = 0.249; Pearson R = 0.66, 95% CI [0.540, 0.761]; p =
−14; 数字 2), although not as accurate as Model 1. As in Model 1, beta slope and its
2.5 × 10
interaction with the beta 6-month intercept were significantly associated with future lan-
guage ability only for low-risk infants, and 6-month theta intercept was associated with fu-
ture language ability only for high-risk infants; 然而, neither remained statistically
significant after controlling for multiple comparisons. Gamma slope and the interaction be-
tween intercept and slope were no longer significant predictors in this model, suggesting that
the trajectory of gamma power beyond 12 months may be more relevant to language devel-
opment than its trajectory over the first 12 月.
Neurobiology of Language
45
Use of longitudinal EEG to estimate language in infants with ASD risk
Differences in how EEG measures explain language variability in high-risk infants with and without ASD
(模型 3)
Next we asked whether high-risk infants with ASD (HR-ASD) have different brain-language
associations than high-risk infants without ASD (HR-NoASD). 模型 3 therefore used EEG data
only from high-risk infants, allowing for two-way interactions between ASD outcome and EEG
参数. This model had improved predictive accuracy as measured by the correlation be-
tween model-estimated and measured language outcomes (数字 2) across high-risk infants
−19;
(Adjusted Model: Adjusted R2 = 0.530; Pearson R = 0.849, 95% CI [0.763, 0.905]; i = 4 × 10
−17).
Unadjusted Model: Adjusted R2 = 0.480; Pearson R = 0.83, 95% CI [0.737, 0.894; p = 1 × 10
正如预期的那样, ASD diagnosis negatively contributed to language outcomes, but was only margin-
ally significant as a predictor (B coefficient = −123.0, p = 0.053). In this adjusted model, 二
brain-language associations were significantly different between high-risk infants with and with-
out ASD: beta and gamma 6-month intercepts. For each of these, the brain-language association
was significant within each group, but directionally opposite (桌子 2, 模型 3). Higher estimated
6-month gamma power was associated with increased 24-month language in HR-NoASD infants,
and decreased 24-month language in HR-ASD infants.
此外, several longitudinal EEG measures contributed similarly across both groups.
Consistent with Model 1, both HR-NoASD and HR-ASD groups had significant and consistent
associations between 6-month intercepts of theta and delta and 24-month language measures.
此外, the effect of the interaction between intercept and slope for the low alpha and beta
frequency bands were the same across groups.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
讨论
This work represents our effort to utilize longitudinal EEG measurements to predict a function-
ally relevant developmental outcome in ASD—language ability. 此外, we were able to
further dissect differences in brain-language associations between low- and high-risk infants
crucial to furthering our understanding of the neural underpinnings of language delay in this
人口.
Use of EEG as a Predictor of Language Ability
All three models were remarkably accurate in within-model estimates of language ability. 的
兴趣, unadjusted models (which did not include parental education and sex covariates) 还
showed a high correlation between model-estimated and observed language scores, empha-
sizing the importance of the brain measures in the predictive models. 此外, in our ad-
justed Model 1, ASD risk status was not a significant predictor of language ability. 这
suggests that when the model accounts for differences in brain-language associations related
to risk status, as well as sex and parental education, the remaining effect of high-risk ASD sta-
tus on language development is no longer significant. 此外, significant interactions be-
tween several EEG variables and risk status were present in the model, supporting our
hypothesis that there are significant brain differences between low- and high-risk infants,
and that these brain differences rather than risk status contribute to language development.
Although EEG parameters in this study sample could estimate language scores with reason-
able accuracy, there are several major caveats that need to be addressed before we are fully
convinced that EEG measures can be used clinically to accurately predict future language risk.
第一的, relatively few participants in this study sample had below average language skills. 到
improve generalizability, a much larger sample with diverse language ability is required.
第二, the ability of EEG to be used as a clinical biomarker is reliant on EEG measures being
Neurobiology of Language
46
Use of longitudinal EEG to estimate language in infants with ASD risk
reproducible across different locations and different collection environments. 第三, 虽然
we used resting power for this study, other measures such as intertrial phase coherence, phase
amplitude coupling, or newly developed measures such as fitting oscillations & one over f, 或者
FOOOF (Haller et al., 2018), should also be evaluated to determine which measures best
discriminate language outcomes. 最后, external validity of model performance needs to
be tested in the future.
Further insights into neural mechanisms of early language development
A secondary goal of this study was to evaluate differences in brain-language associations be-
tween low- and high-risk infants. To do this we performed post hoc analyses of the predictors
included in the models to determine which EEG measures most robustly contributed to lan-
guage estimates and whether there were significant differences between low- and high-risk
infants in how a measure contributed to language estimates. In our models, possible EEG
measures included both a developmentally early measure of EEG power (estimated 6-month
intercept) and a measure of how EEG power changes over development (slope as a function of
log-age). We specifically included estimated 6-month power, as we have previously reported
reduced EEG power across frequency bands at this young age in the high-risk population
(Levin et al., 2017; Tierney et al., 2012) and hypothesized that this reduced power may influ-
ence the development of language circuitry. 的确, in Models 1 和 2, early estimated 6-month
EEG power measures (in delta and theta) contributed significantly to language scores only in the
high-risk group. One possible explanation for this is that these early power measures reflect the
degree of aberrant brain development or differences in cortical maturation in this at-risk popu-
关系. Consistent with this, in our model restricted only to high-risk infants (模型 3), estimated
6-month power of multiple frequency bands (delta, theta, beta, 和伽玛) played prominent
roles in predicting language.
We also hypothesized that specific frequency bands known to be important during speech
加工 (delta/theta and beta/gamma) would be significant predictors of language within
our models; 然而, we were uncertain whether brain-language associations in these fre-
quency bands would be different between risk groups, as differences in resting state and
task-evoked theta and gamma power have been observed in children and adults with ASD.
然而, in evaluating specific frequency bands it is important to recognize that within the
multivariate linear regression models developed, the contribution of an individual variable ex-
ists in the context of the other variables included (EEG measures, risk status, 性别, parental ed-
教育). 所以, the effect attributed to a particular variable reflects only the unique portion
of that variable that does not overlap with the other variables included in the model. 因此,
interpreting the direction and weight of B coefficients within these models must be done with
caution, as the effects of any frequency band may be different in the multivariate model con-
text than when the same frequency band is examined in isolation. With this caveat in mind,
several observations related to theta and gamma warrant further discussion.
Within Model 1, which included data from 3 到 24 months from low- and high-risk infants,
the slope of theta and gamma were significant predictors of language and did not differ be-
tween risk groups. 此外, estimated 6-month delta and theta power were also significant
predictors of language in this model, but only for the high-risk group. 尤其, estimated
6-month theta power was a consistent significant predictor in all three models for high-risk
婴儿, both adjusted and unadjusted, with increased theta power associated with worse
24-month language ability. 尤其, increased theta power has been associated with learning
and attention disorders (Barry, 克拉克, & Johnstone, 2003), and increased theta has been
Neurobiology of Language
47
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Use of longitudinal EEG to estimate language in infants with ASD risk
observed in institutionalized infants and toddlers with increased risk for developmental delays
(马歇尔, Bar-Haim, & 狐狸, 2002). There are several biological explanations for theta’s asso-
ciation with language outcomes. Perhaps most intriguing is the observation that during speech
加工, neural oscillations in the delta and theta range phase-lock to the syllabic rhythm of
speech, which typically peaks in the theta range (Giraud & Poeppel, 2012; 力量, Mead,
巴恩斯, & Goswami, 2012). Aberrant phase-locking in the delta and theta range has been pro-
posed as a possible mechanism for abnormal language acquisition (Goswami, 2011). 在阿迪-
的, theta oscillations are known to influence gamma activity and this coupling between
frequencies is thought to facilitate the alignment of speech components during speech decod-
英 (Giraud & Poeppel, 2012). In adults with ASD, atypical coordination between theta and
gamma responses to speech has been reported, and correlated with clinical measures of ASD
(Jochaut et al., 2015). Future investigation of theta oscillations and their coupling with gamma
oscillations, at rest and in response to speech, at this young age in neurodevelopmental pop-
ulations at risk for significant language delays may provide specific insight related to auditory
processing abnormalities affecting language development. Because theta oscillations have
also been implicated in attention, future comparison of models developed to estimate other
developmental nonverbal skills (例如, visual-spatial, motor, 注意力) would provide more
information of the specificity of these EEG patterns on language development.
Alterations in theta and gamma power have been observed in children and adults with ASD
who also have language delays; therefore it is possible that the association between these fre-
quency bands and language within Model 1 may be related to their shared variance with ASD.
To further separate out EEG measures that predict language, beyond their association with
自闭症谱系障碍, we developed Model 3, which includes ASD diagnosis as a predictor and interaction
term with EEG measures. 这里, having an ASD diagnosis was a marginally significant inde-
pendent predictor of language (p = 0.05) with a large negative B coefficient, suggesting that
factors associated with ASD, but not captured by other variables in our model, have a negative
impact on language development. Such factors could include other structural and functional
brain-derived measures besides resting EEG power, or reduced eye contact or aberrant sensory
processing that may not be captured by resting EEG measures. 此外, with the ASD var-
iable in the model, shared variance between ASD status and the EEG measures is no longer
reflected in the individual parameters but does contribute to the model’s overall ability to dif-
ferentiate language scores. It is notable that within Model 3, significant and opposing differ-
ences in brain-language associations of estimated 6-month beta and gamma power were
observed when ASD status was included in the model. Here we observe that when ASD
diagnosis and its effect on language outcomes are accounted for, reduced 6-month gamma
power positively contributes to estimated 24-month language ability. 然而, 当。。。的时候
effects of ASD on language are not included in the model, reduced 6-month gamma power
negatively contributes to estimated 24-month language ability.
Gamma oscillations are associated with many processes thought to be relevant to language pro-
cessing, including visual and auditory sensory integration (Senkowski, 施耐德, Foxe, & 恩格尔,
2008), 注意力 (薯条, Nikolić, & 歌手, 2007; 泰勒, Mandon, Freiwald, & Kreiter, 2005), 和
工作记忆 (Howard et al., 2003; Pesaran, Pezaris, Sahani, 米特拉, & 安徒生, 2002). 在
婴儿, induced gamma-band power in response to native phonemes matures between 3
和 6 months of age (Ortiz-Mantilla, Hämäläinen, Musacchia, & Benasich, 2013; Peña,
Pittaluga, & Mehler, 2010). Resting frontal gamma power has also been positively associated
with language ability in toddlers and preschool-aged children (Benasich et al., 2008; Brito,
Fifer, 迈尔斯, Elliott, & 高贵, 2016; Gou et al., 2011; Tarullo et al., 2017). 然而, 增加
gamma power has been observed in older children with ASD and other ASD-related
Neurobiology of Language
48
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Use of longitudinal EEG to estimate language in infants with ASD risk
neurodevelopmental disorders such as Fragile X syndrome, and this aberrant increase in
gamma power is thought to reflect imbalances in the excitatory and inhibitory systems of the
脑 (Buzsáki & 王, 2012; Sohal, 2012; Traub et al., 2003; Whittington & Traub, 2003).
此外, in Fragile X syndrome, increased baseline gamma power has been associated with
decreased stimulus-induced phase-locked gamma activity, and therefore has been hypothesized
to represent an hyperexcitable system with reduced ability to synchronize to a stimulus (Ethridge
等人。, 2016). In minimally verbal children with ASD, reduced frontal gamma during visual pro-
cessing has been observed and hypothesized to be modulated by attention, although how this
related to baseline gamma has not been reported (Ortiz-Mantilla et al., 2019). 因此, one hypoth-
esis for the difference between gamma-language associations in HR-NoASD versus HR-ASD in-
fants, is that reduced gamma power in an individual with underlying excitatory/inhibitory
imbalance (HR-ASD) may represent successful compensation of early aberrant neurocircuitry,
whereas reduced gamma power in an individual without such underlying imbalances (人力资源-
NoASD) may reflect delayed neural maturation as it relates to speech processing. Future investi-
gation of how differences in resting gamma may influence evoked gamma across development
may inform both the timing and development of therapeutics.
最后, sex and parental education both were significantly associated with language in ad-
justed models, supporting an independent role of each in language development. 这不是
surprising as previous studies have observed relationships between both sex (Campbell et al.,
2003; Choudhury & Benasich, 2003; Reilly et al., 2007; Schjølberg et al., 2011; Zubrick,
泰勒, 米, & Slegers, 2007) and parental education (Campbell et al., 2003; Schjølberg
等人。, 2011; Tomblin, 哈代, & Hein, 1991; Tomblin, 史密斯, & 张, 1997) 和语言
发展. 然而, unadjusted models that used EEG data alone also estimated language
scores that were highly correlated with actual scores, suggesting that longitudinal EEG mea-
sures may also reflect differences in sex and parental education. Of interest, in Model 3, 哪个
was restricted to high-risk infants and included ASD outcome, sex continued to have a mar-
ginally significant effect on language outcomes. Given the growing evidence of sex differences
in early brain development within ASD (Baron-Cohen, 2010; Kim et al., 2013; Lai et al., 2017;
Mottron et al., 2015; Werling & Geschwind, 2013), it is possible that within the high-risk
infant-sibling population, there are additional brain-language differences between males
and females. A larger sample size is needed to fully tease apart the effects of sex and parental
education on brain development as it relates to language development in this complex
人口.
Limitations
Although this is a comparatively large infant-sibling study, we were limited in our analyses by
our sample size. To maximize the number of individuals included in model development, 我们
included individuals with only two EEG time points (n = 21). Requiring more time points could
provide more stable estimations of trajectories; 然而, it would also significantly reduce the
sample size. 的确, limiting Model 1 to individuals with at least three EEG sessions, reduced
model fit (adjusted R2 = 0.2574 与. 0.3295) and accuracy (Pearson correlation coefficient 0.65
与. 0.70). This study also limited its analysis to frontal power. The decision to use frontal power
was made a priori based on previous resting-state EEG literature in both the ASD and language
fields. 然而, it is possible that power analysis using a different set of electrodes could be
more informative. To evaluate this possibility, models using whole brain or temporal elec-
trodes were built from the same individuals included in Model 1. Both of these models had
reduced adjusted R2 values (whole brain, 0.2065; 颞, 0.2334; frontal, 0.3295) 和
lower Pearson correlation coefficients for predicted versus observed Mullen scores (颞,
Neurobiology of Language
49
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Use of longitudinal EEG to estimate language in infants with ASD risk
0.59; whole brain – 0.60, frontal – 0.70). 最后, as discussed earlier, in order to improve
generalizability of future predictive models, a larger and more diverse study sample is re-
询问, followed by external validity testing of model performance.
结论
This study supports the possible use of EEG measures as predictive biomarkers for language
development in infants. It also provides further insight into which neurobiological substrates
may specifically relate to language development, potentially informing future work and ther-
apeutic interventions. 此外, our findings support further investigation of brain differences
in high-risk infants at, or prior to, 6 months of age, and how such differences affect future lan-
guage development and response to services. 要做到这一点, collaboration across laboratories is
needed in order to collect larger datasets for accurate model development that can then be
leveraged clinically. 最终, early prediction of developmental outcomes will require im-
proved knowledge of how underlying genetic and environmental risk factors affect neural
measures and their association with outcomes.
致谢
We thank all the families and staff who were involved in this study.
资金信息
H. Tager-Flusberg, National Institute on Deafness and Other Communication Disorders
(https://doi.org/10.13039/100000055), 奖项ID: R01-DC010290. C. A. 纳尔逊, 国家的
Institute on Deafness and Other Communication Disorders (https://doi.org/10.13039/
100000055), 奖项ID: R01-DC010290. H. Tager-Flusberg, National Institute on Deafness
and Other Communication Disorders (https://doi.org/10.13039/100000055), 奖项ID: R21
DC08637. C. L. Wilkinson, National Institute of Mental Health (https://doi.org/10.13039/
100000025), 奖项ID: 1T32MH112510. A. 右. 莱文, American Brain Foundation (我们). C. L.
Wilkinson, Autism Science Foundation (https://doi.org/10.13039/100008152). L. J. Gabard-
Durnam, Autism Science Foundation (https://doi.org/10.13039/100008152). A. 右. 莱文,
Autism Science Foundation (https://doi.org/10.13039/100008152). H. Tager-Flusberg,
Autism Speaks (https://doi.org/10.13039/100000073), 奖项ID: 1323. A. 右. 莱文, Brain
and Behavior Research Foundation (https://doi.org/10.13039/100000874). C. L. Wilkinson,
FRAXA Research Foundation (https://doi.org/10.13039/100000297). A. 右. 莱文, 氮. Lurie
Marks Family Foundation (https://doi.org/10.13039/100007429). C. L. Wilkinson, Thrasher
Research Fund (https://doi.org/10.13039/100005627). L. J. Gabard-Durnam, Rett Syndrome
Research Trust. Charles A Nelson, Simons Foundation (https://doi.org/10.13039/
100000893), 奖项ID: 137186.
作者贡献
C. L. Wilkinson: Conceptualization – lead; Data curation – lead; 形式分析; 方法;
Writing – original draft. L. J. Gabard-Durnam: 概念化; 方法; 软件;
写作——复习 & 编辑. K. Kapur: 形式分析; Methodology – lead; 写作——复习 &
编辑. H. Tager-Flusberg: 概念化; 资金获取; 调查; Super-
想象; 写作——复习 & 编辑. A. 右. 莱文: 方法; 软件; 监督; Writing –
review & 编辑. C. A. 纳尔逊: 概念化; 资金获取; 调查; 项目
administration – lead; 监督; 写作——复习 & 编辑.
Neurobiology of Language
50
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Use of longitudinal EEG to estimate language in infants with ASD risk
参考
安德森, D. K., Lord, C。, 创新, S。, DiLavore, 磷. S。, 舒尔曼, C。,
Thurm, A。, … Pickles, A. (2007). Patterns of growth in verbal
abilities among children with autism spectrum disorder. 杂志
of Consulting and Clinical Psychology, 75(4), 594–604. https://
doi.org/10.1037/0022-006X.75.4.594
Babadi, B., & 棕色的, 乙. 氮. (2014). A review of multitaper spectral
分析. IEEE Transactions on Biomedical Engineering, 61(5),
1555–1564. https://doi.org/10.1109/TBME.2014.2311996
Baron-Cohen, S. (2010). Empathizing, systemizing, and the ex-
treme male brain theory of autism. 脑研究进展,
186(C), 167–175. https://doi.org/10.1016/B978-0-444-53630-
3.00011-7
Barry, 右. J。, 克拉克, A. R。, & Johnstone, S. J. (2003). A review of
electrophysiology in attention-deficit/hyperactivity disorder: 我.
Qualitative and quantitative electroencephalography. Clinical
Neurophysiology, 114(2), 171–183. https://doi.org/10.1016/
S1388-2457(02)00362-0
Benasich, A. A。, Gou, Z。, Choudhury, N。, & 哈里斯, K. D. (2008). 早期的
cognitive and language skills are linked to resting frontal gamma
power across the first 3 年. Behavioural Brain Research, 195,
215–222. https://doi.org/10.1016/j.bbr.2008.08.049
Benjamini, Y。, & Hochberg, 是. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Journal of the Royal Statistical Society B. https://doi.org/10.1111/
j.2517-6161.1995.tb02031.x
Billstedt, E., Gillberg, 我. C。, & Gillberg, C. (2005). Autism after ad-
olescence: Population-based 13- to 22-year follow-up study of
120 individuals with autism diagnosed in childhood. 杂志
Autism and Developmental Disorders, 35(3), 351–360. https://
doi.org/10.1007/s10803-005-3302-5
Brito, 氮. H。, Fifer, 瓦. P。, 迈尔斯, 中号. M。, Elliott, A. J。, & 高贵, K. G.
(2016). Associations among family socioeconomic status, EEG power
出生时, and cognitive skills during infancy. Developmental Cog-
nitive Neuroscience, 19, 144–151. https://doi.org/10.1016/j.
dcn.2016.03.004
Buzsáki, G。, & 王, X.-J. (2012). Mechanisms of gamma oscilla-
系统蒸发散. Annual Review of Neuroscience, 35(1), 203–225. https://
doi.org/10.1146/annurev-neuro-062111-150444
坎贝尔, 时间. F。, Dollaghan, C. A。, Rockette, H. E., Paradise, J. L。,
费尔德曼, H. M。, Shriberg, L. D ., … Kurs-Lasky, 中号. (2003). 风险
factors for speech delay of unknown origin in 3-year-old chil-
德伦. Child Development, 74(2), 346–357. https://doi.org/
10.1111/1467-8624.7402002
Choudhury, N。, & Benasich, A. A. (2003). A family aggregation
学习: The influence of family history and other risk factors on
language development. Journal of Speech, 语言, 和
Hearing Research: JSLHR, 46(2), 261–272. https://doi.org/
10.1044/1092-4388(2003/021)
Colon, 乙. J。, Notermans, S. L. H。, de Weerd, J. 磷. C。, & Kap, J.
(1979). The discriminating role of EEG power spectra in dyslexic
孩子们. Journal of Neurology, 221(4), 257–262. https://doi.org/
10.1007/BF00314642
Cornelissen, L。, Kim, S.-E., Purdon, 磷. L。, 棕色的, 乙. N。, & Berde, C. 乙.
(2015). Age-dependent electroencephalogram (EEG) patterns dur-
ing sevoflurane general anesthesia in infants. 亿生活, 4. https://土井.
org/10.7554/eLife.06513
Estes, A。, Zwaigenbaum, L。, Gu, H。, St John, T。, Paterson, S。, Elison,
J. T。, … IBIS Network. (2015). Behavioral, cognitive, and adap-
tive development in infants with autism spectrum disorder in the
第一的 2 years of life. Journal of Neurodevelopmental Disorders, 7
(1), 24. https://doi.org/10.1186/s11689-015-9117-6
Ethridge, L。, 白色的, S。, Mosconi, M。, 王, J。, Byerly, M。, &
Sweeney, J. (2016). Reduced habituation of auditory evoked
potentials indicate cortical hyper-excitability in Fragile X
Syndrome. Translational Psychiatry, 6. https://doi.org/10.1038/
tp.2016.48
薯条, P。, Nikolić, D ., & 歌手, 瓦. (2007). The gamma cycle.
Trends in Neurosciences, 30(7), 309–316. https://doi.org/
10.1016/j.tins.2007.05.005
Gabard-Durnam, L. J。, Mendez Leal, A. S。, Wilkinson, C. L。, &
莱文, A. 右. (2018). The Harvard Automated Processing
Pipeline for Electroencephalography (HAPPE): Standardized pro-
cessing software for developmental and high-artifact data.
Frontiers in Neuroscience, 12, 97. https://doi.org/10.3389/FNINS.
2018.00097
Giraud, A.-L., & Poeppel, D. (2012). Cortical oscillations and
speech processing: Emerging computational principles and oper-
ations. 自然神经科学, 15(4), 511–517. https://doi.org/
10.1038/nn.3063
Goswami, U. (2011). A temporal sampling framework for develop-
mental dyslexia. 认知科学的趋势, 15(1), 3–10.
https://doi.org/10.1016/J.TICS.2010.10.001
Gotham, K., Pickles, A。, & Lord, C. (2012). Trajectories of autism
severity in children using standardized ADOS scores. Pediatrics,
130(5), e1278–e1284. https://doi.org/10.1542/peds.2011-3668
Gou, Z。, Choudhury, N。, & Benasich, A. A. (2011). Resting frontal
gamma power at 16, 24, 和 36 months predicts individual dif-
ferences in language and cognition at 4 和 5 年. Behavioural
脑研究, 220(2), 263–270. https://doi.org/10.
1016/j.bbr.2011.01.048
Haller, M。, 多诺霍, T。, 彼得森, E., Varma, P。, Sebastian, P。,
高, R。, … Voytek, 乙. (2018). Parameterizing neural power
spectra. bioRxiv. https://doi.org/10.1101/299859
Harmony, T。, Marosi, E., Becker, J。, Rodríguez, M。, Reyes, A。,
Fernández, T。, … Bernal, J. (1995). Longitudinal quantitative EEG
study of children with different performances on a reading-writing
测试. Electroencephalography and Clinical Neurophysiology, 95(6),
426–433. https://doi.org/10.1016/0013-4694(95)00135-2
Hastie, T。, Tibshirani, R。, & 弗里德曼, J. (2009). The elements of sta-
tistical learning (2nd 版。). 纽约, 纽约: Springer and Science
Business Media. Retrieved from https://link.springer.com/10.1007/
978-0-387-84858-7
Hazlett, H. C。, Gu, H。, Munsell, 乙. C。, Kim, S. H。, Styner, M。, Wolff,
J。, … Piven, J. (2017). Early brain development in infants at high
risk for autism spectrum disorder. 自然, 542(7641), 348–351.
https://doi.org/10.1038/nature21369
霍华德, 中号. W., Rizzuto, D. S。, Caplan, J. B., Madsen, J. R。, Lisman,
J。, Aschenbrenner-Scheibe, R。, … Kahana, 中号. J. (2003). Gamma
oscillations correlate with working memory load in humans.
大脑皮层, 13(12), 1369–1374. https://doi.org/10.1093/
cercor/bhg084
Jansson-Verkasalo, E., Ruusuvirta, T。, Huotilainen, M。, Alku, P。,
Kushnerenko, E., Suominen, K., … Hallman, 中号. (2010). Atypical
perceptual narrowing in prematurely born infants is associated
with compromised language acquisition at 2 年龄. BMC
神经科学, 11, 88. https://doi.org/10.1186/1471-2202-11-88
Jeste, S. S。, Frohlich, J。, & Loo, S. K. (2015). Electrophysiological
biomarkers of diagnosis and outcome in neurodevelopmental
disorders. Current Opinion in Neurology, 28(2), 110–116.
https://doi.org/10.1097/WCO.0000000000000181
Jing, H。, Gilchrist, J. M。, Badger, 时间. M。, & Pivik, 右. 时间. (2010). A longi-
tudinal study of differences in electroencephalographic activity
Neurobiology of Language
51
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Use of longitudinal EEG to estimate language in infants with ASD risk
among breastfed, milk formula-fed, and soy formula-fed infants dur-
ing the first year of life. Early Human Development, 86(2), 119–125.
https://doi.org/10.1016/j.earlhumdev.2010.02.001
Jochaut, D ., Lehongre, K., Saitovitch, A。, Devauchelle, A.-D.,
Olasagasti, 我。, Chabane, N。, … Giraud, A.-L. (2015). Atypical co-
ordination of cortical oscillations in response to speech in autism.
Frontiers in Human Neuroscience, 9, 171. https://doi.org/10.3389/
fnhum.2015.00171
Kim, K. C。, Kim, P。, Go, H. S。, Choi, C. S。, 公园, J. H。, Kim, H. J。, ……
Shin, C. 是. (2013). Male-specific alteration in excitatory post-
synaptic development and social interaction in pre-natal valproic
acid exposure model of autism spectrum disorder. 杂志
Neurochemistry, 124(6), 832–843. https://doi.org/10.1111/
jnc.12147
Kjelgaard, 中号. M。, & Tager-Flusberg, H. (2001). An investigation of
language impairment in autism: Implications for genetic sub-
团体. Language and Cognitive Processes, 16(2–3), 287–308.
https://doi.org/10.1080/01690960042000058
Kuhl, 磷. K. (2010). Brain mechanisms in early language acquisition.
神经元, 67(5), 713–727. https://doi.org/10.1016/j.neuron.2010.08.038
Kuhl, 磷. K., Conboy, 乙. T。, Padden, D ., 纳尔逊, T。, & Pruitt, J.
(2005). Language learning and development early speech per-
ception and later language development: Implications for the
“Critical Period.” Language Learning and Development, 1(3–4),
237–254. https://doi.org/10.1080/15475441.2005.9671948
Lai, 中号. C。, Lerch, J. P。, Floris, D. L。, Ruigrok, A. 氮. V, Pohl, A。,
Lombardo, 中号. 五、, & Baron-Cohen, S. (2017). Imaging sex/gender
and autism in the brain: Etiological implications. 杂志
Neuroscience Research, 95(1–2), 380–397. https://doi.org/
10.1002/jnr.23948
Landa, R。, & Garrett-Mayer, 乙. (2006). Development in infants with
autism spectrum disorders: A prospective study. Journal of Child
Psychology and Psychiatry, 47(6), 629–638. https://doi.org/
10.1111/j.1469-7610.2006.01531.x
莱文, A. R。, Méndez Leal, A. S。, Gabard-Durnam, L. J。, & O’Leary,
H. 中号. (2018). BEAPP: The Batch Electroencephalography
Automated Processing Platform. Frontiers in Neuroscience, 12.
https://doi.org/10.3389/fnins.2018.00513
莱文, A. R。, & 纳尔逊, C. A. (2015). Inhibition-based biomarkers for
autism spectrum disorder. Neurotherapeutics, 12(3), 546–552.
https://doi.org/10.1007/s13311-015-0350-1
莱文, A. R。, Varcin, K. J。, O’Leary, H. M。, Tager-Flusberg, H。, &
纳尔逊, C. A. (2017). EEG power at 3 months in infants at high
familial risk for autism. Journal of Neurodevelopmental
Disorders, 9(1), 34. https://doi.org/10.1186/s11689-017-9214-9
Lombardo, 中号. 五、, Pierce, K., Eyler, L. T。, Carter Barnes, C。, Ahrens-
Barbeau, C。, Solso, S。, … Courchesne, 乙. (2015). Different func-
tional neural substrates for good and poor language outcome in
自闭症. 神经元, 86(2), 567–577. https://doi.org/10.1016/j.
neuron.2015.03.023
Lord, C。, & Rutter, 中号. (2012). ADOS-2: Autism diagnostic observa-
tion schedule (2nd 版。). 天使们, CA: Western Psychological
Services.
Marrus, N。, 大厅, L. P。, Paterson, S. J。, Elison, J. T。, Wolff, J. J。, &
斯旺森, 中号. 右. (2018). Language delay aggregates in toddler sib-
lings of children with autism spectrum disorder. 杂志
Neurodevelopmental Disorders, 10(1), 29. https://doi.org/10.1186/
s11689-018-9247-8
马歇尔, 磷. J。, Bar-Haim, Y。, & 狐狸, 氮. A. (2002). Development of the
EEG from 5 months to 4 年龄. Clinical Neurophysiology, 113(8),
1199–1208. https://doi.org/10.1016/S1388-2457(02)00163-3
磨坊主, L. E., 伯克, J. D ., Troyb, E., Knoch, K., Herlihy, L. E., & Fein,
D. A. (2017). Preschool predictors of school-age academic
achievement in autism spectrum disorder. The Clinical Neuropsy-
chologist, 31(2), 382–403. https://doi.org/10.1080/13854046.2016.
1225665
Mottron, L。, Duret, P。, Mueller, S。, 摩尔, 右. D ., Forgeot d’Arc, B.,
Jacquemont, S。, & Xiong, L. (2015). Sex differences in brain plas-
ticity: A new hypothesis for sex ratio bias in autism. Molecular
Autism, 6(1), 33. https://doi.org/10.1186/s13229-015-0024-1
Ortiz-Mantilla, S。, Cantiani, C。, Shafer, V. L。, & Benasich, A. A. (2019).
Minimally-verbal children with autism show deficits in theta and
gamma oscillations during processing of semantically-related visual
信息. Scientific Reports, 9(1), 5072. https://doi.org/10.1038/
s41598-019-41511-8
Ortiz-Mantilla, S。, Hämäläinen, J. A。, Musacchia, G。, & Benasich,
A. A. (2013). Enhancement of gamma oscillations indicates pref-
erential processing of native over foreign phonemic contrasts in
婴儿. The Journal of Neuroscience: The Official Journal of the
Society for Neuroscience, 33(48), 18746–18754. https://doi.org/
10.1523/JNEUROSCI.3260-13.2013
Peña, M。, Pittaluga, E., & Mehler, J. (2010). Language acquisition in
premature and full-term infants. 国家会议录
美国科学院, 107,
3823–3828. https://doi.org/10.1073/pnas.0914326107
Penolazzi, B., Spironelli, C。, & Angrilli, A. (2008). Delta EEG ac-
tivity as a marker of dysfunctional linguistic processing in de-
velopmental dyslexia. Psychophysiology, 45(6), 1025–1033.
https://doi.org/10.1111/j.1469-8986.2008.00709.x
Pesaran, B., Pezaris, J. S。, Sahani, M。, 米特拉, 磷. P。, & 安徒生, 右. A.
(2002). Temporal structure in neuronal activity during working
memory in macaque parietal cortex. 自然神经科学, 5(8),
805–811. https://doi.org/10.1038/nn890
Pickles, A。, 安德森, D. K., & Lord, C. (2014). Heterogeneity and
plasticity in the development of language: A 17-year follow-up of
children referred early for possible autism. Journal of Child
Psychology and Psychiatry, 55(12), 1354–1362. https://doi.org/
10.1111/jcpp.12269
力量, A. J。, Mead, N。, 巴恩斯, L。, & Goswami, U. (2012). Neural
entrainment to rhythmically presented auditory, visual, 和
audio-visual speech in children. 心理学前沿, 3.
https://doi.org/10.3389/fpsyg.2012.00216
Reilly, S。, Wake, M。, Bavin, 乙. L。, 事先的, M。, 威廉姆斯, J。, Bretherton, L。,
… Ukoumunne, 氧. C. (2007). Predicting language at 2 年龄:
A prospective community study. Pediatrics, 120(6), e1441–e1449.
https://doi.org/10.1542/peds.2007-0045
Riva, 五、, Cantiani, C。, Mornati, G。, Gallo, M。, Villa, L。, Mani, E., ……
Molteni, 中号. (2018). Distinct ERP profiles for auditory process-
ing in infants at-risk for autism and language impairment. Sci-
entific Reports, 8(1), 715. https://doi.org/10.1038/s41598-017-
19009-y
Rojas, D. C。, & Wilson, L. 乙. (2014). γ-Band abnormalities as
markers of autism spectrum disorders. Biomarkers in Medicine,
8(3), 353–368. https://doi.org/10.2217/bmm.14.15
Rutter Bailey, A。, & Lord, C. 中号. (2003). SCQ: Social communication
questionnaire. 天使们, CA: Western Psychological Services.
Schiavone, G。, 林肯卡尔-汉森, K., Maurits, 氮. M。, Plakas, A。,
Maassen, 乙. A. M。, Mansvelder, H. D ., … van Zuijen, 时间. L.
(2014). Preliteracy signatures of poor-reading abilities in rest-
ing-state EEG. Frontiers in Human Neuroscience, 8, 735. https://
doi.org/10.3389/fnhum.2014.00735
Schjølberg, S。, Eadie, P。, Zachrisson, H. D ., Oyen, A.-S., & 事先的, 中号.
(2011). Predicting language development at age 18 月: 数据
from the Norwegian Mother and Child Cohort Study. 杂志
Developmental and Behavioral Pediatrics: JDBP, 32(5), 375–383.
https://doi.org/10.1097/DBP.0b013e31821bd1dd
Neurobiology of Language
52
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Use of longitudinal EEG to estimate language in infants with ASD risk
Seery, A。, Tager-Flusberg, H。, & 纳尔逊, C. A. (2014). Event-related
potentials to repeated speech in 9-month-old infants at risk for
autism spectrum disorder. Journal of Neurodevelopmental
Disorders, 6(1), 43. https://doi.org/10.1186/1866-1955-6-43
Senkowski, D ., 施耐德, 时间. R。, Foxe, J. J。, & 恩格尔, A. K. (2008).
Crossmodal binding through neural coherence: Implications for
multisensory processing. Trends in Neurosciences. https://土井.
org/10.1016/j.tins.2008.05.002
Sohal, V. S. (2012). Insights into cortical oscillations arising from
optogenetic studies. Biological Psychiatry, 71(12), 1039–1045.
https://doi.org/10.1016/j.biopsych.2012.01.024
Spironelli, C。, Penolazzi, B., Vio, C。, & Angrilli, A. (2006). Inverted
EEG theta lateralization in dyslexic children during phonological
加工. Neuropsychologia, 44(14), 2814–2821. https://土井.
org/10.1016/j.neuropsychologia.2006.06.009
斯旺森, 中号. R。, 沉, 中号. D ., Wolff, J. J。, Elison, J. T。, Emerson, 右. W.,
Styner, 中号. A。, … Piven, J. (2017). Archival report subcortical brain
and behavior phenotypes differentiate infants with autism versus
language delay. https://doi.org/10.1016/j.bpsc.2017.07.007
Szatmari, P。, Bryson, S。, Duku, E., Vaccarella, L。, Zwaigenbaum,
L。, Bennett, T。, & Boyle, 中号. H. (2009). Similar developmental
trajectories in autism and Asperger syndrome: From early child-
hood to adolescence. Journal of Child Psychology and
Psychiatry, 50(12), 1459–1467. https://doi.org/10.1111/j.1469-
7610.2009.02123.X
Szatmari, P。, Bryson, S. E., Streiner, D. L。, Wilson, F。, Archer, L。, &
Ryerse, C. (2000). Two-year outcome of preschool children with au-
tism or Asperger’s syndrome. American Journal of Psychiatry,
157(12), 1980–1987. https://doi.org/10.1176/appi.ajp.157.12.1980
Tager-Flusberg, H。, & Kasari, C. (2013). Minimally verbal school-
aged children with autism spectrum disorder: The neglected
end of the spectrum. Autism Research, 6(6), 468–478. https://
doi.org/10.1002/aur.1329
Tarullo, A. R。, Obradović, J。, Keehn, B., Rasheed, 中号. A。, Siyal, S。,
纳尔逊, C. A。, & Yousafzai, A. K. (2017). Gamma power in rural
Pakistani children: Links to executive function and verbal ability.
Developmental Cognitive Neuroscience, 26, 1–8. https://doi.org/
10.1016/j.dcn.2017.03.007
泰勒, K., Mandon, S。, Freiwald, 瓦. A。, & Kreiter, A. K. (2005).
Coherent oscillatory activity in monkey area V4 predicts success-
ful allocation of attention. 大脑皮层, 15(9), 1424–1437.
https://doi.org/10.1093/cercor/bhi023
Thomson, D. J. (1982). Spectrum estimation and harmonic analysis.
Proceedings of the IEEE, 70(9), 1055–1096. https://doi.org/10.
1109/PROC.1982.12433
Tierney, A. L。, Gabard-Durnam, L。, Vogel-Farley, 五、, Tager-Flusberg,
H。, & 纳尔逊, C. A. (2012). Developmental trajectories of resting
EEG power: An endophenotype of autism spectrum disorder. 公共科学图书馆
一, 7(6), e39127. https://doi.org/10.1371/journal.pone.0039127
Tomblin, J. B., 哈代, J. C。, & Hein, H. A. (1991). Predicting poor-
communication status in preschool children using risk factors
present at birth. Journal of Speech and Hearing Research, 34(5),
1096–1105. https://doi.org/10.1044/jshr.3405.1096
Tomblin, J. B., 史密斯, E., & 张, X. (1997). Epidemiology of spe-
cific language impairment: Prenatal and perinatal risk factors.
Journal of Communication Disorders, 30(4), 325–344. https://
doi.org/10.1016/S0021-9924(97)00015-4
Traub, 右. D ., 坎宁安, 中号. 奥。, Gloveli, T。, LeBeau, F. 乙. N。, Bibbig,
A。, Buhl, 乙. H。, & Whittington, 中号. A. (2003). GABA-enhanced col-
lective behavior in neuronal axons underlies persistent gamma-
frequency oscillations. 美国国家科学院院刊
Sciences of the United States of America, 100(19), 11047–11052.
https://doi.org/10.1073/pnas.1934854100
Varcin, K. J。, & 纳尔逊, C. A. (2016). A developmental neurosci-
ence approach to the search for biomarkers in autism spectrum
紊乱. Current Opinion in Neurology, 29(2), 123–129. https://
doi.org/10.1097/WCO.0000000000000298
王, J。, Barstein, J。, Ethridge, L. E., Mosconi, 中号. W., Takarae, Y。,
& Sweeney, J. A. (2013). Resting-state EEG abnormalities in au-
tism spectrum disorders. Journal of Neurodevelopmental
Disorders, 5(1), 1–14. https://doi.org/10.1186/1866-1955-5-24
Werling, D. M。, & Geschwind, D. H. (2013). Sex differences in au-
tism spectrum disorders. Current Opinion in Neurology, 26(2),
146–153. https://doi.org/10.1097/WCO.0b013e32835ee548
Whittington, 中号. A。, & Traub, 右. D. (2003). Interneuron Diversity
系列: Inhibitory interneurons and network oscillations in vitro.
Trends in Neurosciences, 26(12), 676–682. https://doi.org/
10.1016/j.tins.2003.09.016
Wilkinson, C. L。, 莱文, A. R。, Gabard-Durnam, L. J。, Tager-
Flusberg, H。, & 纳尔逊, C. A. (2019). Reduced frontal gamma
power at 24 months is associated with better expressive language
in toddlers at risk for autism. Autism Research, 12, 1211–1224.
https://doi.org/10.1002/aur.2131
Winkler, 我。, Debener, S。, 穆勒, K. R。, & Tangermann, 中号. (2015).
On the influence of high-pass filtering on ICA-based artifact re-
duction in EEG-ERP. In Proceedings of the Annual International
Conference of the IEEE Engineering in Medicine and Biology Society
(EMBC), 米兰, 意大利, 4101–4105. https://doi.org/10.1109/
EMBC.2015.7319296
Winkler, 我。, Haufe, S。, & Tangermann, 中号. (2011). Automatic classi-
fication of artifactual ICA-components for artifact removal in EEG
signals. Behavioral and Brain Functions, 7(1), 30. https://doi.org/
10.1186/1744-9081-7-30
Zubrick, S. R。, 泰勒, C. L。, 米, 中号. L。, & Slegers, D. 瓦. (2007). Late
language emergence at 24 月: An epidemiological study of prev-
alence, predictors, and covariates. Journal of Speech, 语言, 和
Hearing Research: JSLHR, 50(6), 1562–1592. https://doi.org/10.1044/
1092-4388(2007/106)
Neurobiology of Language
53
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
1
1
3
3
1
8
6
7
8
7
6
n
哦
_
A
_
0
0
0
0
2
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3