Individual Differences in Multisensory Processing Are
Related to Broad Differences in the Balance of
Local versus Distributed Information
Phillip R. Johnston , Claude Alain, and Anthony R. McIntosh*
Abstracto
■ The brain’s ability to extract information from multiple sen-
sory channels is crucial to perception and effective engage-
ment with the environment, but the individual differences
observed in multisensory processing lack mechanistic explana-
ción. Nosotros planteamos la hipótesis de que, from the perspective of information
theory, individuals with more effective multisensory processing
will exhibit a higher degree of shared information among
distributed neural populations while engaged in a multisensory
tarea, representing more effective coordination of information
among regions. To investigate this, healthy young adults com-
pleted an audiovisual simultaneity judgment task to measure
their temporal binding window (TBW), which quantifies the
ability to distinguish fine discrepancies in timing between
auditory and visual stimuli. EEG was then recorded during a
second run of the simultaneity judgment task, and partial least
squares was used to relate individual differences in the TBW
width to source-localized EEG measures of local entropy and
mutual information, indexing local and distributed processing
of information, respectivamente. The narrowness of the TBW,
reflecting more effective multisensory processing, was related
to a broad pattern of higher mutual information and lower
local entropy at multiple timescales. Además, un pequeño
group of temporal and frontal cortical regions, including those
previously implicated in multisensory integration and response
selección, respectivamente, played a prominent role in this pattern.
En general, these findings suggest that individual differences in
multisensory processing are related to widespread individual
differences in the balance of distributed versus local informa-
tion processing among a large subset of brain regions, con
more distributed information being associated with more
effective multisensory processing. The balance of distributed
versus local information processing may therefore be a useful
measure for exploring individual differences in multisensory
Procesando, its relationship to higher cognitive traits, and its
disruption in neurodevelopmental disorders and clinical
condiciones. ■
INTRODUCCIÓN
We experience the world through multiple sensory sys-
TEMS, each sampling different kinds of energy from the
environment and offering complementary information
about the world around us. The brain’s ability to extract
relevant information from across sensory channels is crit-
ical to interacting effectively with the world, as demon-
strated by the numerous behavioral enhancements that
multisensory stimuli provide compared with unisensory
estímulos (p.ej., Shams & Seitz, 2008; Diederich & Colonius,
2004; Hershenson, 1962; Sumby & Pollack, 1954). Estos
enhancements, in combination with converging anatomi-
cal and physiological evidence of extensive multisensory
influences across neural scales, suggest that the brain is
fundamentally organized to facilitate multisensory pro-
cesando, with wide-reaching consequences for perception,
cognición, and action (Conductor & Noesselt, 2008; Ghazanfar
& Schroeder, 2006).
universidad de toronto
*Current affiliation: Simon Fraser University.
© 2022 Instituto de Tecnología de Massachusetts
Multisensory processing is also characterized by its var-
iability across the population, with substantial individual
differences in multisensory tasks and illusions occurring
in healthy individuals (p.ej., Cecere, rees, & Romei, 2015;
Nath & Beauchamp, 2012; stevenson, Zemtsov, & Wallace,
2012; Molinero & D'Esposito, 2005). Además, correla-
tions among various multisensory tasks and audiovisual
illusions suggest that these individual differences may be
expressions of a general mechanism of multisensory pro-
cesando (stevenson & Wallace, 2013; Stevenson et al.,
2012), which may use a set of basic neural operations to
flexibly integrate information between distributed neural
populations in a wide variety of contexts (van Atteveldt,
Murray, Thut, & Schroeder, 2014; Senkowski, Schneider,
Foxe, & ángel, 2008). Sin embargo, prior studies investigating
individual differences in multisensory processing have
largely focused on univariate measures of neural activity
(p.ej., Ferri et al., 2017; Balz et al., 2016; Kaganovich &
Schumaker, 2016; Cecere et al., 2015) without considering
interactions among neural populations (but see Kumar,
Dutta, Talwar, roy, & Banerjee, 2020). Given that multi-
sensory processing involves reconciling information
Revista de neurociencia cognitiva 34:5, páginas. 846–863
https://doi.org/10.1162/jocn_a_01835
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
oh
C
norte
_
a
_
0
1
8
3
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
encoded in different sensory systems to produce percepts
and guide action, we expect that effective multisensory
processing requires information to be processed across
widely distributed neural populations. Tal como, nosotros
hypothesize that individuals capable of more effective
multisensory processing will demonstrate a higher degree
of shared information among distributed cortical regions
while engaged in a multisensory task, likely including
regions of multisensory convergence such as the STS
(jones & Powell, 1970), which is frequently implicated in
audiovisual processing (p.ej., Marchant, Fallar, & Conductor,
2012; Balk et al., 2010; Noesselt et al., 2007; Calvert, Hansen,
Iversen, & Brammer, 2001).
To investigate this possibility, we compared the balance
of local versus distributed information processing to
multisensory performance using information-theoretical
measures of local entropy and mutual information at
source-localized EEG time series. Multisensory perfor-
mance was assessed as audiovisual temporal discrimination,
employing the commonly used audiovisual simultaneity
juicio (SJ) tarea (p.ej., stevenson & Wallace, 2013;
Powers, Hillock, & Wallace, 2009; van Eijk, Kohlrausch,
Juola, & van de Par, 2008). This task allows a temporal bind-
ing window (TBW) to be measured for each participant,
quantifying the probability that an auditory and visual
stimulus will be perceived as asynchronous as a function
of the time difference between them. TBWs are known to
vary substantially between individuals (Stevenson et al.,
2012; Powers et al., 2009; Conrey & Pisoni, 2006; Molinero &
D'Esposito, 2005), and those with narrower TBWs were
considered to have more effective multisensory processing,
reflecting a better ability to resolve timing differences
between sensory channels. Following measurement of the
TBW, participants underwent EEG recording during a
second run of the SJ task, which was calibrated to equalize
difficulty between participants.
To quantify the balance of distributed versus local infor-
procesamiento de datos, source-localized EEG time series were
submitted to a time delay embedding-based algorithm
computing local entropy and mutual information at multi-
ple timescales (Vakorin, Lippé, & McIntosh, 2011). Estos
information-theoretical measures allow the joint informa-
tion of two neural time series to be partitioned into that
which is unique to each population (local entropy) y
that which is shared between them (mutual information).
Tal como, local entropy and mutual information provide
measures of local and distributed information, respetar-
activamente. Partial least squares (PLS; McIntosh & Lobaugh,
2004) was then used to identify components (latent vari-
ables [LVs]) of local entropy and mutual information distri-
bution that maximally covaried with the individual-level
TBW measurements. Además, source-localized power
spectral densities (PSDs) were subjected to the same PLS
analysis to allow comparison with previous electrophysio-
logical work and disambiguate the role of cross-frequency
dependencies and nonlinear autocorrelations in differen-
tiating individuals (Courtiol et al., 2016).
MÉTODOS
Participantes
Twenty-eight healthy young adults were recruited, con
three being excluded from analysis because of failed sig-
moid model fitting after the calibration task (vea abajo)
and three excluded for excessive EEG artifacts. The final
sample of 22 young adults (11 mujer, ages 19–33 years,
mean = 23.6 años, DE = 3.5 años) had an average of
16.2 years of education, and all had normal or corrected-
to-normal vision and hearing. Twenty participants were
right-handed, one was left-handed, and one was ambidex-
trous. No participant reported a diagnosis of dyslexia,
autism spectrum disorder, schizophrenia, or other clinical
condition with noted relevance to multisensory processing
(Hahn, Foxe, & Molholm, 2014; de Boer-Schellekens,
Eussen, & Vroomen, 2013; Martín, Giersch, Huron, &
van Wassenhove, 2013; Kwakye, Foss-Feig, Cascio, Piedra,
& Wallace, 2011; Foucher, Lacambre, Pham, Giersch, &
eliot, 2007; Hairston, Burdette, Flores, Madera, & Wallace,
2005). All participants provided written consent accord-
ing to the guidelines established by Baycrest Centre
and the University of Toronto and were provided mone-
tary compensation for their participation.
This work is a secondary analysis of a data set (collected
by the authors and previously unpublished) intended to
investigate the neural correlates of audiovisual perceptual
binding. The final sample size (22) offered by this data set
surpassed that of similar work using EEG to investigate
multisensory perception during the SJ task ( Yuan, li,
Liu, Yuan, & Huang, 2016 [18]; Kambe, Kakimoto, & Araki,
2015 [14]) and is comparable to other EEG studies inves-
tigating individual differences in multisensory perception
(p.ej., Kumar et al., 2020 [18]; Cecere et al., 2015 [22]).
Behavioral Protocol
Audiovisual SJ Task
Participants first completed a two-alternative forced-
choice SJ task to measure the width of their TBWs, también
as to calibrate the stimuli for the task presented during
EEG recording. The SJ task consisted of a jittered fixation
período (1000–1500 msec), followed by a visual flash and
auditory beep stimulus, and lastly a response prompt
(ver figura 1). The flash and beep stimuli were separated
by a systematically varied SOA, where a negative number
denotes auditory-leading (AV) presentation and a positive
number denotes visual-leading (Virginia) presentación.
The auditory stimulus was a 3500-Hz pure sine tone,
10 msec in length, delivered by a GSI 61 audiometer
through ER-3A insert earphones (Etymotic Research). El
audiometer was calibrated such that a 5-sec tone at
the same frequency produced an intensity of 102 dB SPL.
The visual stimulus was a white annulus flash on a black
fondo, presented for 10 msec and covering 3.8° of
visual angle at a viewing distance of 60 centímetro. It was presented
on a Dell Trinitron CRT monitor at a refresh rate of 100 Hz.
Johnston, Alaín, and McIntosh
847
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
oh
C
norte
_
a
_
0
1
8
3
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
oh
C
norte
_
a
_
0
1
8
3
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. Audiovisual task. Schematic of the audiovisual SJ task depicting the visual-leading ( Virginia) condición.
After an interval of 750 mseg, a prompt was displayed,
and the participant reported whether they perceived the
two stimuli as synchronous (“yes”) or asynchronous (“no”)
by pressing the left or right arrow button (counterba-
lanced between participants) on a standard computer key-
junta. Participants responded with the right hand, y
responses were made within a 2000-msec time limit. Trials
were then separated by a 750-msec intertrial interval
before the next fixation.
The task was built with PsychoPy software ( Versión
1.90.3; Peirce et al., 2019) and presented on a Dell Preci-
sion T3600 computer. Stimulus timing was verified to be
accurate within ±4 msec using a Tektronix TDS210 two-
channel oscilloscope.
For the initial calibration run, 19 SOAs were used in
total, ranging from −300 to 300 mseg, to estimate each
participant’s TBW. Específicamente, SOAs of 0, 10, 20, 50, 80,
100, 150, 200, 250, y 300 msec were presented in both
the AV and VA cases (Stevenson et al., 2012). The task was
broken into four blocks, wherein each SOA was presented
four times in a pseudorandom order, for a total of 16 pre-
sentations per SOA. Two hundred eighty-five trials were
presented over the course of the whole task, which lasted
apenas 15 min in total. Participants were offered a self-
timed break between each block.
EEG was recorded during a second run of the SJ task.
This version of the task, originally designed to investigate
the neural correlates of audiovisual binding, presented six
SOAs, four of which were calibrated for each participant
individually based on the results of the preceding behav-
ioral run. para hacerlo, the rate of synchrony perception for
each SOA was calculated as the number of “synchronous”
responses divided by the total number of presentations
(16). Two psychometric sigmoid functions—one for the
AV SOAs (−300 to 0 mseg) and one for the VA SOAs (0–
300 mseg)—were fit to the resulting rates using the lmfit
function in Python (least squares mode; stevenson &
Wallace, 2013; Hillock-Dunn & Wallace, 2012; Powers
et al., 2009). For the AV stimuli, “A50” and “A95” SOAs were
calibrated to produce asynchrony perception 50% y
95% del tiempo, respectivamente, by solving the AV psycho-
metric sigmoid for values of 0.5 y 0.95. “V50” and
“V95” trials were calibrated in the same way for VA stimuli.
Por último, “A10” and “V10” trials used a fixed SOA of 10 mseg
with auditory- and visual-leading stimuli, respectivamente. A
total of 512 trials were presented in a pseudorandom
orden, broken into four blocks of 128 trials each. Dentro
each block, A50 and V50 trials were presented 32 veces
cada, and A10, V10, A95, and V95 trials were presented
16 times each. This balance of trial types was chosen to
equalize subjective difficulty by presenting participants
with a high proportion of ambiguous stimuli while pre-
venting guessing or adaptation by offsetting these with
more obviously synchronous (A10, V10) and asynchro-
Nosotros (A95, V95) estímulos.
Observer Model
Although an arbitrary psychometric sigmoid was used
to calibrate the ambiguous stimuli at the time of data
recopilación, a more sophisticated observer model was
applied post hoc to parameterize the TBW. The four
parameter observer model proposed by Yarrow et al.
( Yarrow, 2018, 2020; Yarrow, Jahn, Durant, & arnold,
2011) posits that SJs are the product of a latent decision pro-
impuesto, where a noisy internal representation of the difference
in timing between stimuli is compared with lower (auditivo-
leading) and upper (visual-leading) decision criteria. El
TBW is therefore estimated as the difference of two
cumulative Gaussians representing the noisy boundaries
separating simultaneous from nonsimultaneous judg-
mentos. Each Gaussian is defined by a mean (mAV and
mVA; the estimated mean SOA values of the decision
criteria) and standard deviation (σAV and σVA; representante
a combination of sensory and criterion noise). Tal como,
mAV and mVA define the width of the TBW and are
848
Revista de neurociencia cognitiva
Volumen 34, Número 5
analogous to other point estimates of TBW width, cual
have previously shown individual differences (p.ej.. stevenson
et al., 2012; Powers et al., 2009; Conrey & Pisoni, 2006;
Molinero & D'Esposito, 2005), whereas σAV and σVA deter-
mine the slope of the TBW. In line with previous work,
mAV and mVA were considered the primary measures of
interest quantifying individual differences in the multisen-
sory SJ task and will hereafter be referred to simply as the
AV and VA TBW width, respectivamente.
To assess the fit of the observer model, it was compared
with a simpler two-parameter “guessing” model, diseñado
to determine whether participants were simply guessing
and whether they were presented with a sufficient range
of SOAs to adequately sample their transitions from simul-
taneous to nonsimultaneous judgments. Full details of the
observer model, the guessing model, and their implemen-
tation are available in Yarrow et al. ( Yarrow, 2018, 2020;
Yarrow et al., 2011).
Self-report Measures
Participants completed a posttask questionnaire, el
results of which were not analyzed here. Participantes
reflected on the SJ task and tried to quantify potential
biases in their responding, as well as guess the intention
of the study. Additional questions assessed discomfort
and whether participants fell asleep at any point during
la tarea, as well as clinical or neurodevelopmental diagno-
ses (p.ej., dyslexia, autism spectrum disorder, and schizo-
phrenia). Finalmente, participants reported their experience
with and time spent playing video games, as well as musi-
cal training and proficiency.
Electrophysiological Analysis
EEG Recording and Preprocessing
EEG was recorded with a BioSemi ActiveTwo acquisition
sistema (BioSemi Instrumentation) at a sampling rate of
2048 Hz and bandwidth (−3 dB) of DC-400 Hz and then
decimated to 512 Hz in ActiView acquisition software.
Sixty-six scalp electrodes were employed, using BioSemi’s
64 + 2 electrode cap configuration based on the 10/20 sys-
tema. Ten additional electrodes were applied in pairs to the
mastoids, preauricular points, upper cheeks, outer canthi
of the eyes, and inferior orbit of the eyes. These provided
better coverage of the scalp, as well as an accurate record
of eye movements for later artifact removal. All recordings
took place in a dimly lit, sound-attenuating room.
All EEG preprocessing was performed in Brainstorm
( Version 07-Apr-2020; Tadel, Baillet, Mosher, Pantazis, &
Leahy, 2011), an open-source application for M/EEG data
processing and visualization. Continuous recordings were
digitally band-pass filtered at 0.5–90 Hz (linear-phase FIR,
stopband attenuation = 60 db, transition band = 0 Hz),
with a notch filter centered at 60 Hz to attenuate line noise
(second-order IIR, 3-dB notch bandwidth = 2 Hz). Bad
electrodes and contaminated segments of continuous data
were manually rejected from subsequent processing, y
the remaining data were rereferenced to the average of all
remaining electrodes. Eye- and cardiac-related artifact
components were then detected and removed using
Brainstorm’s implementation of Infomax independent
component analysis (Makeig, Campana, Junk, & Sejniano,
1996) applied to the longest available continuous segment
of data without major artifacts (minimum of 5 min or
153,600 muestras).
We assessed individual differences in neural activity
during the multisensory task by analyzing data from the
intertrial interval, with epochs spanning 1750 msec start-
ing immediately after the response and ending before
the onset of the next stimulus. This interval was chosen
to capture individual differences in the functional organi-
zation of brain activity that emerges during the multisen-
sory task while minimizing spillover from the stimuli
ellos mismos, which varied between participants and could
not be directly compared. Implications of this choice are
addressed in the Discussion section. After rejection of
contaminated segments, an average of 471.4 epochs of
1750 mseg (13.7 min of data in total) were analyzed for
cada participante.
EEG Source Estimation
The cortical current sources of the EEG signals were esti-
mated with Brainstorm using sLORETA (Pascual-Marqui,
2002), with one dipole modeled normal to the cortical sur-
face at each vertex, using an OpenMEEG BEM forward
modelo (Gramfort, Papadopoulo, Olivi, & Clerc, 2010) com-
puted on the default MNI/ ICBM152 anatomy in Brain-
storm (fonov, evans, Mckinstry, almíli, & collins, 2009).
The inversion kernel computed on the full cortical surface
(15,002 vértices) was used to downsample the results to
the Desikan-Killiany atlas (Desikan et al., 2006), resulting
in one time series for each of the 68 regions of the atlas.
PSD Computation
PSDs were computed using a smoothed fast Fourier trans-
form implemented in the neurodsp package in Python
(“medfilt” method, frequency range = 1–90 Hz, frequency
resolution = 0.57 Hz, median filter length = 1 Hz; Col,
Donoghue, gao, & Voytek, 2019). PSDs were computed
on each individual epoch and then averaged over epochs,
resulting in one average PSD associated with each source.
Local Entropy and Mutual Information
Entropy H(X ) is a measure of information or uncertainty
associated with a single random variable X. Similarmente, el
joint entropy H(X, Y ) is the entropy of the joint probability
distribution of two random variables X and Y. By concep-
tualizing neural time series as random variables, informa-
tion theory provides tools to partition the joint entropy of
Johnston, Alaín, and McIntosh
849
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
oh
C
norte
_
a
_
0
1
8
3
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
oh
C
norte
_
a
_
0
1
8
3
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 2. Mutual information and local entropy. The total entropy H(X, Y ) of two neural time series (X and Y ) can be partitioned into shared and
unique quantities. Mutual information, I(X; Y ), is the shared entropy between X and Y and is therefore a measure of affiliation between the two
variables. h(X|Y ) y H(Y|X ) are estimates of the local (unique) entropy of each variable, representing the remaining entropy after knowledge of
the other variable (mutual information) is excluded.
decomposition (SVD) to extract orthogonal patterns of
maximal covariance, called LVs, between two data matrices.
PLS can therefore identify components of a brain data
matrix that maximally covary with behavioral measure-
mentos. Específicamente, the relationship between brain and
behavior data, represented by their cross-product matrix,
is decomposed using SVD, producing three new matrices,
containing (1) behavior saliences, (2) brain saliences, y
(3) singular values. The behavior saliences can be thought
of as contrasts that specify the relationship between the
elements of the behavioral matrix, and the “brain saliences”
are the extracted patterns of brain data that best character-
ize the cross-product matrix. The singular values are the
square roots of the eigenvalues and are proportional to
the amount of covariance in the cross-product matrix cap-
tured by each LV. Finalmente, because the extraction of these
brain–behavior relationships via SVD takes place across
the entire brain in a single mathematical step, correction
for multiple comparisons is not necessary (McIntosh &
Lobaugh, 2004; McIntosh et al., 1996).
two neural time series into that which is unique to one or
the other (local entropy) and that which is shared between
a ellos (mutual information; ver figura 2).
Partitioning of the joint entropy of X and Y can be
described with the following formula:
Þ þ H Y jXð
Þ ¼ H XjYð
d
H X; Y
d
Þ þ I X; Y
Þ
The conditional entropies H(X|Y ) y H(Y|X ), referred to
here as local entropies, represent the entropy of each time
series with knowledge of the other excluded. Lo mutuo
information I(X; Y ) between X and Y therefore represents
the amount of information shared between the two
variables.
Local entropy and mutual information were estimated
with time delay embedding (embedding dimension d =
2, embedding delay τ = 1, characteristic scale length r =
1) using the method previously described in Vakorin et al.
(2011). Given that entropy estimates are scale dependent
(Costa, Goldberger, & Peng, 2005; zhang, 1991), estos
quantities were computed at multiple timescales. Hacer
entonces, each time series was downsampled by averaging within
nonoverlapping windows of successively increasing length
before time delay embedding. A maximum window length
de 18 samples was chosen to ensure that a minimum of
acerca de 50 puntos de datos (896 samples/18 = 49.78 windows)
were included in the estimation.
To characterize the dynamic interactions of the whole-
brain network, multiscale local entropy and mutual infor-
mation were computed between each pair of sources at
each epoch and then averaged across epochs. Local
entropy for a given node X was then taken to be the aver-
age of local entropy values, h(X|Y ), computed between
that node and all other nodes Y.
PLS Analysis
PLS (McIntosh & Lobaugh, 2004; McIntosh, Bookstein,
Haxby, & grady, 1996) is a multivariate statistical technique,
similar to canonical correlation, which uses singular value
Cifra 3. TBW estimates. (A) Average TBW for the whole sample. Por
convention, negative SOAs denote auditory-leading (AV) estímulos y
positive SOAs denote visual-leading ( Virginia) estímulos. Shading depicts SEM.
(B) Relationship between AV and VA TBW width estimates. TBW widths
(AV and VA) were quantified for each participant as the means of the
two cumulative Gaussians from that participant’s observer model fit
( Yarrow, 2018, 2020).
850
Revista de neurociencia cognitiva
Volumen 34, Número 5
Aquí, behavior PLS implemented in MATLAB (McIntosh
& Lobaugh, 2004) was used to identify LVs relating individ-
ual differences in TBW width to patterns in (1) local
entropía, (2) mutual information, y (3) PSDs. A
explore the maximal experimental effects in the data,
the rotated version of PLS was used, which produces a
set of LVs based on mutually orthogonal contrasts in a
data-driven manner. Statistical assessment of the resulting
LVs was performed with a two-stage resampling proce-
desdén. Primero, statistical significance of each LV was esti-
mated using permutation testing (1000 resamples),
where observations of the behavioral matrix were ran-
domly reassigned without replacement. La resultante
p value represents the proportion of permuted singular
values that exceeded the observed singular values. El
reliability of each element was then estimated using boot-
strap resampling (1000 resamples), where the standard
error of each element salience was estimated by recom-
puting PLS on a set of observations resampled with
replacement (maintaining the mapping between brain
and behavioral observations). The bootstrap ratio (el
ratio of the salience to the standard error computed
through resampling) captures how dependent the ele-
ment salience is on the particular makeup of the sample
(sansón, Streissguth, barr, & Bookstein, 1989), y es
roughly equivalent to a z score when the bootstrap
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
oh
C
norte
_
a
_
0
1
8
3
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
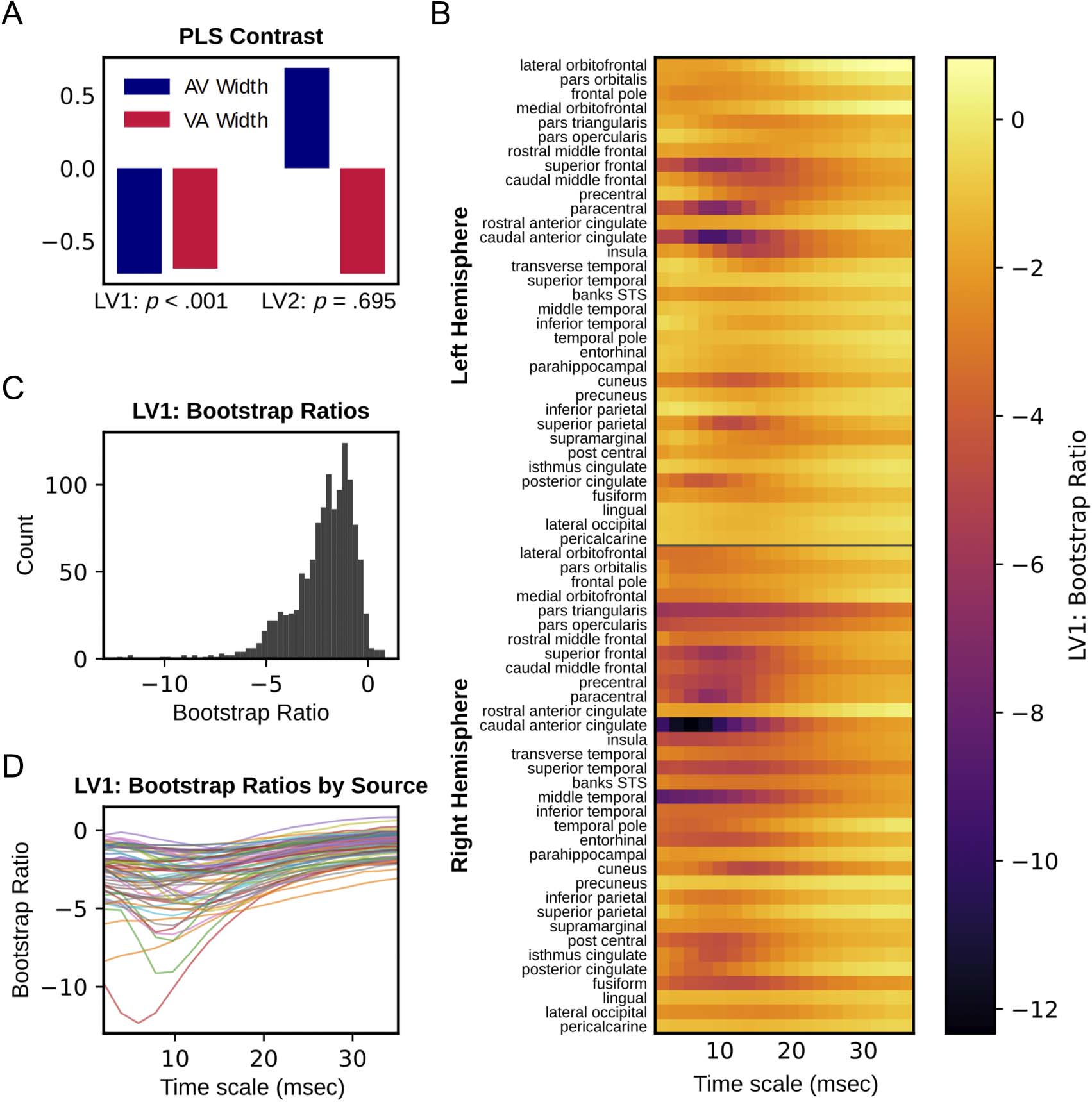
Cifra 4. Local entropy PLS results. (A) Data-driven contrast for PLS relating AV and VA TBW width to local entropy, with p values derived from
1000 permutations. (B) LV1 bootstrap ratios at each source and timescale. Bootstrap ratios quantify the stability of the contrast at each source
and timescale as determined through bootstrap resampling. Negative values here indicate sources and timescales where narrower TBWs are
associated with less local entropy. (C) Distribution of LV1 bootstrap ratios across all sources. (D) LV1 bootstrap ratios at each source as a function
of timescale.
Johnston, Alaín, and McIntosh
851
distribution is normal (Efron & Ruidoso, 1986). Nosotros
therefore interpret the bootstrap ratios as measures of
fiabilidad, using the permutation test for null hypothesis
pruebas.
Mesa 1. El 10 Sources with the Most Extreme Median PLS
Bootstrap Ratios for (A) Local Entropy and (B) Mutual
Información, Computed Across All Timescales
Fuente
Median Bootstrap Ratio
RESULTADOS
Observer Model Fitting
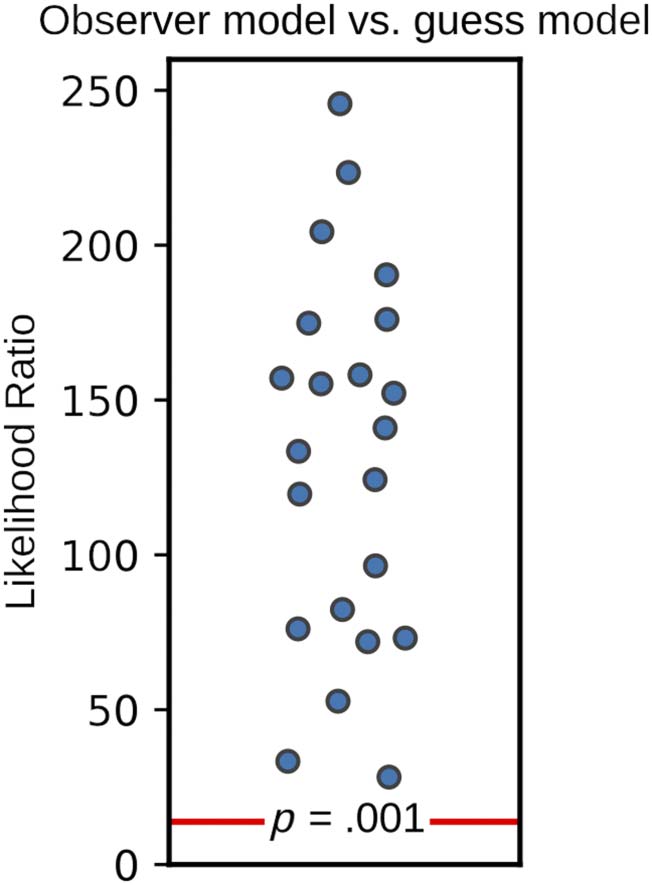
The observer model provided a significant improvement
in fit over the guessing model in all cases ( pag < .001;
Appendix A), suggesting that participants were not guess-
ing, and SOAs were adequately sampled.
The mean TBW width estimated with the observer
model was 162.1 msec (SD = 59.3 msec) for AV stimuli
and 236.0 msec (SD = 86.44 msec) for VA stimuli, repro-
ducing the commonly reported finding that the TBW is
wider for VA stimuli than AV stimuli on average, t(21) =
−5.9, p < .001.1 Figure 3A depicts the average TBW for
all participants in the sample. As described previously
(e.g., Stevenson et al., 2012), AV and VA TBW widths were
strongly correlated within individuals, r(20) = .75, p <
.001 (see Figure 3B).
Local Entropy
Behavior PLS relating local entropy values to TBW width
identified one significant LV (LV1: p < .001, singular
value = 18.58; LV2: p = .70, singular value = 3.28).
Figure 4 illustrates the relationship between TBW width
and local entropy estimates across sources and timescales
captured by this LV. The data-driven contrast values for AV
and VA TBW width had the same sign (see Figure 4A, left),
indicating that this LV captures commonalities between
the two measures in their relation to local entropy. For this
LV, correlations [95% CI] between brain scores and TBW
width were AV: −.75 [−.89, −.67] and VA: −.67 [−.85,
−.55]. Bootstrap ratios quantify the stability of this con-
trast at each source and timescale, where positive ratios
indicate support for the contrast and negative ratios
indicate support for the inverse of the contrast. Because
the contrast values in this case are negative, negative boot-
strap ratios indicate where TBW width and local entropy
are positively related. Bootstrap ratios were overwhelm-
ingly negative across sources and timescales (see
Figure 4B–D), indicating that narrower TBWs were associ-
ated broadly with lower local entropy, and therefore,
wider TBWs were associated with higher local entropy.
Furthermore, the effect was most stable at finer time-
scales, with the 11.7 msec timescale having the most
extreme median bootstrap ratio. Table 1A lists the sources
demonstrating the highest median bootstrap ratios across
timescales.
In addition to the width parameters, the slope parame-
ters were also compared with local entropy values using
behavior PLS. Whether relating local entropy to width
and slope together (Appendix Figure B1, left side) or slope
(A) Local Entropy
Pars triangularis R
Caudal anterior cingulate R
Middle temporal R
Superior temporal R
Caudal anterior cingulate L
Superior frontal L
Pars opercularis R
Caudal middle frontal R
Superior frontal R
Fusiform R
(B) Mutual Information
Middle temporal R
Rostral middle frontal R
Pars triangularis R
Pars orbitalis R
Caudal anterior cingulate R
Paracentral R
Superior temporal R
Superior frontal L
Rostral middle frontal L
Pars triangularis L
−4.98
−4.75
−4.49
−4.38
−4.28
−4.22
−3.80
−3.72
−3.59
−3.56
2.60
2.46
2.44
2.14
2.13
2.12
2.10
2.05
2.03
1.97
Sources in bold are within the top 10 for both local entropy and mutual
information. R = right; L = left.
alone (Appendix Figure B1, right side), the pattern that
emerged was largely the same as that described above.
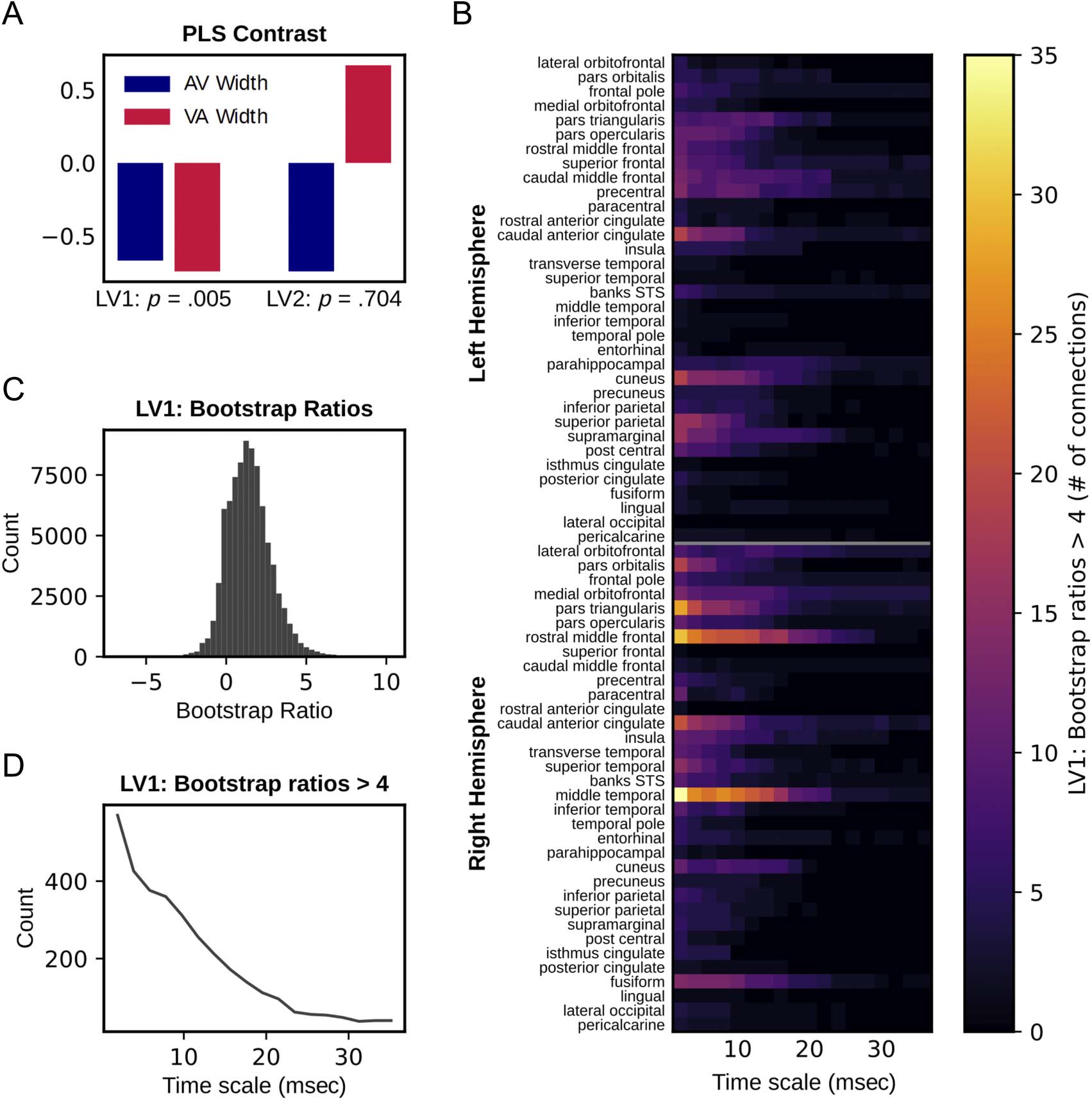
Mutual Information
PLS relating mutual information to TBW width identified
one significant LV (LV1: p = .005, singular value =
125.96; LV2: p = .70, singular value = 27.88). Figure 5
depicts the expression of this LV across timescales and
source-to-source connections. The data-driven contrast
values for AV and VA TBW width again had the same sign
(see Figure 5A, left), indicating that this LV captures
commonalities between the two measures in their rela-
tionship to mutual information. For this LV, correlations
[95% CI] between brain scores and TBW width were AV:
−.63 [−.85, −.54] and VA: −.71 [−.88, −.65]. In con-
trast to local entropy, the bootstrap ratios for mutual
852
Journal of Cognitive Neuroscience
Volume 34, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
o
c
n
_
a
_
0
1
8
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
o
c
n
_
a
_
0
1
8
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Mutual information PLS results. (A) Data-driven contrast for PLS relating AV and VA TBW width to mutual information, with p values
derived from 1000 permutations. (B) Number of times the given source was involved in a highly reliable (bootstrap ratio > 4) connection with
any other source. Bootstrap ratios here quantify the stability of the contrast at each source-to-source connection and timescale as determined
through bootstrap resampling. Positive values here indicate connections where narrower TBWs are associated with higher mutual information.
(C) Distribution of LV1 bootstrap ratios across all source-to-source connections. (D) Number of source-to-source connections with LV1 bootstrap
ratios exceeding 4 as a function of timescale.
information were predominantly positive, Indicando que
narrower TBWs were largely associated with higher
mutual information across timescales and connections,
and therefore, wider TBWs were associated with lower
mutual information. De nuevo, this effect was most stable
at finer timescales. Table 1B lists the sources with the
highest median bootstrap ratios across timescales, cual
are most prominently involved in this pattern, y
Figure 5B depicts the number of times a given source
is involved in a highly reliable (bootstrap ratio > 4)
connection with any other source. To illustrate the pat-
tern of connectivity itself, Cifra 6 displays the pattern
of connections exhibiting the highest reliability at the
11.7-msec timescale.
The slope parameters were also compared with mutual
information using behavior PLS. Relating mutual informa-
tion to width and slope together revealed a similar pattern
to the one described above (Appendix Figure C1, izquierda
lado); sin embargo, slope alone did not yield any significant
LVs (Appendix Figure C1, right side).
PSD
PLS relating PSDs to TBW width extracted two LVs, but per-
mutation testing indicated that neither was significant
(LV1: pag = .28, singular value = 31.41; LV2: pag = .73, singular
value = 9.71).
Johnston, Alaín, and McIntosh
853
Cifra 6. Mutual information
PLS results: conectividad. Circle
plot depicting the 1% de
connections with the highest
bootstrap ratios for the
11.7 msec timescale. Positive
values indicate connections
where narrower TBWs are
associated with higher mutual
información. Top left inset:
Connections with LV1 bootstrap
ratios exceeding 4 as a function
of timescale. Red line marks
the timescale featured in the
circle plot.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
oh
C
norte
_
a
_
0
1
8
3
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
DISCUSIÓN
Narrower TBW Is Broadly Associated with Mutual
Information and Negatively Associated with
Local Entropy
We found that individual differences in audiovisual tempo-
ral discrimination ability, as measured by TBW width
covaried with differences in local entropy and mutual
information while participants engaged in an audiovisual
tarea. Widespread differences were observed, with better
temporal discrimination (narrower TBW) being associated
with higher mutual information and lower local entropy
broadly across cortical sources and timescales. Taken
juntos, these results support the hypothesis that better
multisensory processing abilities are associated with a pro-
pensity for greater shared information among distributed
cortical sources and, en cambio, less effective processing
is associated with more localized processing of informa-
ción. A potential interpretation of this finding is that the
ability to extract stimulus features (p.ej., momento) encoded
in separate modalities, compare them, and respond
appropriately is facilitated in individuals who show broad
integration of information involving the coordination of
diverse neural populations while engaged in a multisen-
sory task.
Además, for both local entropy and mutual infor-
formación, only the LV capturing similarities between the
AV and VA TBW widths passed statistical assessment,
suggesting that the pattern of mutual information and
local entropy identified in the brain data applies to both
types of stimulus. This finding, along with the strong
correlation observed between AV and VA TBWs, accords
with previous work proposing that the TBW width indexes
a general mechanism of multisensory processing that
applies across stimulus types (Stevenson et al., 2012),
although further work comparing multiple tasks will be
necessary to confirm this. Además, if local entropy and
mutual information balance do reflect domain-general
multisensory processing ability, further work will need to
consider how this more general process interacts with
possible stimulus-specific mechanisms, which may pro-
duce the observed difference in neural responses to AV
and VA stimuli (p.ej., Cecere, Bruto, willis, & Thut, 2017)
and differences in the malleability of the AV and VA TBWs
(Cecere, Bruto, & Thut, 2016; Powers et al., 2009).
Although TBW width was our primary measure of inter-
Este, the observer model also provides the slope of the
TBW, and these two parameters have different theoretical
interpretaciones. Under the assumptions of the observer
modelo, width indexes the position of the boundary
between simultaneous and nonsimultaneous judgments,
whereas slope is hypothesized to capture the “noisiness”
of internal representations of stimulus timing and decision
criteria. Despite these theoretical differences, nuestros resultados
suggest that both width and slope relate to largely the
same pattern of individual differences in local entropy
and mutual information (Appendices B and C). It could
be the case that the observer model is correct in its inter-
pretation of these parameters, but they are too strongly
correlated to disambiguate (es decir., noisy observers may tend
to also have noisy criteria; Magnotti, Mamá, & Beauchamp,
2013). Alternativamente, the model may not reflect the
854
Revista de neurociencia cognitiva
Volumen 34, Número 5
underlying process that produced these simultaneity
judgments accurately enough to afford such specific
interpretation of its parameters. In either case, podemos
only conclude that the individual differences observed
in the brain data correspond to general performance on
the SJ task, rather than criterion setting or the noisiness
of internal representations specifically.
Temporal and Frontal Regions Exhibit Most
Reliable Differences in Local Entropy and
Mutual Information
Although narrower TBWs were associated with more
distributed and less localized information processing
broadly throughout the cortex, the most reliable differ-
ences were observed primarily in temporal and frontal
corteza (ver tabla 1). Several of these temporal regions
(right superior temporal gyrus and right middle temporal
gyrus) and frontal regions (right pars triangularis, bien
caudal anterior cingulate, and left superior frontal gyrus)
were among those that most reliably demonstrated an
effect in both the local entropy and mutual information
analiza.
Of particular interest is the STS, which has historically
been considered a key region for multisensory processing
because of its converging auditory and visual inputs
(jones & Powell, 1970), and the nonhuman primate work
demonstrating the presence of neurons responsive to
both modalities there (Schroeder & Foxe, 2002; bruce,
Desimona, & Bruto, 1981; Benevento, Caer sobre, davis, &
Rezak, 1977). In humans, numerous fMRI studies have
implicated STS (Marchant et al., 2012; Balk et al., 2010;
Noesselt et al., 2007; Calvert et al., 2001), as well as nearby
superior temporal gyrus (Marchant et al., 2012; stevenson,
VanDerKlok, Pisoni, & Jaime, 2011; stevenson, Altieri,
kim, Pisoni, & Jaime, 2010; Noesselt et al., 2007) en el
integration or temporal discrimination of asynchronous
auditory and visual stimuli using a variety of task para-
digms, which motivated our prediction that STS would
be linked to individual differences on this task. Dado
the limited spatial resolution of the present analysis, es
plausible that the prominent differences observed in the
right superior temporal and right middle temporal sources
could originate in the STS and superior temporal gyrus. Si
entonces, these results would suggest that individual variability in
audiovisual temporal discrimination may in part reflect the
degree to which these putatively multisensory temporal
regions exchange information with a wide array of other
cortical regions (ver figura 6). Sin embargo, it is important
to note that these sources are based on atlas parcellations
encompassing large areas of cortex, likely including
multiple subregions that play diverse roles in cognition
(Hein & Caballero, 2008). Por lo tanto, the sources used here
may capture different processes than those reported in
the fMRI literature. Además, in contrast to previous
findings, here the most reliable effects in temporal cortex
were observed in the right hemisphere, with little
difference emerging in the left hemisphere. A pesar de
the fMRI literature shows largely left-hemispheric or
bilateral effects at the group level, the emphasis here on
individual differences means that this is not necessarily a
contradictory finding (but see Marchant et al., 2012). Este
discrepancy may be the result of heterogeneity in task
design as well as inherent differences between the BOLD
signal and information-theoretical measures used here.
Alternativamente, this finding could indicate that left multi-
sensory temporal cortex is more consistently implicated
in audiovisual processing tasks, but the degree to which
right multisensory temporal cortex is involved varies
between individuals and may be more closely related to
actuación. This individual variability in the involvement
of right multisensory temporal cortex could therefore
explain why it is less consistently identified using BOLD
signal contrasts computed at the group level. Trabajo futuro
seeking to clarify the networks involved in multisensory
processing may therefore benefit from considering how
individual differences in these networks may relate to
task performance.
In frontal cortex, a particularly strong relationship was
identified between bilateral caudal anterior cingulate and
TBW width. En particular, higher local entropy in this
region was strongly associated with wider TBWs, especialmente
at finer timescales. Involvement of this region in a forced-
choice task is not unexpected, given that ACC has been
implicated in decision-making and action selection in
general and is thought to play a flexible role in integrating
behaviorally relevant information from its prefrontal,
parietal, and subcortical connections (Monosov, Haber,
Leuthardt, & Jezzini, 2020). Además, there is evi-
dence that ACC is capable of integrating information over
long timescales, including those spanning multiple trials
(Spitmaan, SEO, Sotavento, & Soltani, 2020). Although still spec-
ulative, this result could suggest that individual differences
in TBW may, hasta cierto punto, reflect differences in
response selection processes mediated by ACC and its net-
trabajar, perhaps involving the way information from previ-
ous trials is integrated to affect response selection.
Differences in Local Entropy and Mutual
Information Most Evident at Fine Timescales
In addition to spatial information, the current method gives
insight into the temporal scales where individual differ-
ences in the information-theoretical measures most reli-
ably correspond to individual differences in TBW. A pesar de
some sources exhibited differences across all timescales,
the finer timescales were most consistently identified for
both local entropy and mutual information, with median
bootstrap ratios peaking at the 11.7 msec timescale for
local entropy and decreasing monotonically from fine to
coarse timescales for mutual information. Because the
downsampling procedure removes the influence of fast-
changing activity as the coarse-graining windows become
Johnston, Alaín, and McIntosh
855
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
oh
C
norte
_
a
_
0
1
8
3
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
más grande, fine timescales represent the entropy of both fast-
and slow-changing activity, whereas coarser timescales rep-
resent only slow-changing activity (Courtiol et al., 2016).
This suggests that the key individual differences in local
entropy and mutual information are likely attributable to
relatively fast-changing elements of the neural signals. Este
finding would seem to corroborate the view that gamma
synchronization between distinct neuronal groups may
play an instrumental role in structuring information within
cortical networks in general (Fries, 2009; ángel, Fries, &
Cantante, 2001) as well as the orchestration of multisensory
interactions more specifically (Keil & Senkowski, 2018).
Sin embargo, relating power spectra to the width of the
TBW with PLS did not show differences in power in the
gamma range or any other, highlighting the fact that
straightforward comparisons between the (multiescala)
entropy measure used here and the power spectrum
are likely not possible. Simulation work has demonstrated
that multiscale entropy (Costa et al., 2005), a similar
technique to that used here, is sensitive to nonlinear
autocorrelations in the time series, as well as cross-
frequency dependencies, although this is not true of the
power spectrum (Courtiol et al., 2016; McIntosh, Kovacevic,
& Itier, 2008). These differences confirm that the
information-theoretical measures provide additional
insight into the temporal structure of neural signals at fine
timescales, which is not captured in the power spectrum.
Implications of Task Design
Given the substantial variability in performance on the SJ
tarea, the SOAs were calibrated to equalize difficulty across
Participantes (Yuan et al., 2016; Kambe et al., 2015). Para el
present analysis, the choice to vary stimuli between partic-
ipants comes with a trade-off, a saber, that stimulus-related
activity cannot be compared directly between individuals.
Although this is not ideal, the particularities of the neural
responses to each type of stimulus (es decir., AV and VA) is of
secondary interest to individual differences in the broader
context of information processing that emerged during
la tarea, which we judged more likely to relate to general
(es decir., nonstimulus specific) mechanisms of multisensory
Procesando. Además, focusing on the intertrial interval
allowed longer epochs without motor contamination to be
analyzed and therefore allowed longer timescales to be
assessed. Por último, equalizing task difficulty across participants
is intended to prevent the boredom or blind guessing
that comes with a task that is too easy or too difficult,
respectivamente, which are problematic when participants are
assumed to be comparably engaged in the task.
The use of the intertrial interval in this analysis, bastante
than task-free resting-state data, poses additional ques-
tions for interpretation. Task-free resting-state data are
typically assumed to better reflect inherent differences in
functional organization between individuals, as opposed
to the more transient brain states related to a specific task.
Sin embargo, the assumption that task-free resting-state
provides an inherently neutral baseline reading of brain
organization or that such a baseline can exist has been
questioned on both procedural and theoretical grounds
(Duncan & Northoff, 2013; Morcom & Fletcher, 2007),
and evidence from fMRI work suggests that engaging
participants in a task may in fact enhance behaviorally rel-
evant individual differences compared with rest (Finn
et al., 2017). Teniendo esto en cuenta, we judged the intertrial
interval sufficient to identify the task-related individual
differences of interest; sin embargo, future work may need
to consider both recordings made during multisensory
tasks as well as task-free rest to disambiguate this issue.
Conclusion and Future Directions
Partitioning entropy into local and distributed compo-
nents provided new insight into the neural correlates of
individual differences in multisensory processing com-
monly observed in healthy adults and served as a proof
of principle for the utility of information-theoretical mea-
sures for investigating multisensory processing. En general,
more effective multisensory processing, here operational-
ized as a narrower audiovisual TBW, was associated with a
widespread pattern of higher mutual information and
lower local entropy while participants were engaged in
an audiovisual SJ task. This pattern provides support for
the hypothesis that more effective multisensory process-
ing requires sharing of information between widely distrib-
uted neural populations. Además, several regions
were strongly implicated in this pattern, including tempo-
ral and frontal regions that have previously been linked to
multisensory integration and response selection, respetar-
activamente. This suggests that the involvement of these regions
within the larger pattern of information exchange could be
important determinants of individual differences in multi-
sensory processing.
To more definitively establish the causal role that the
distribution versus localization of information plays in indi-
vidual multisensory performance, future work will need to
adopt an experimental approach. Prior work has shown
that the TBW can be narrowed with training (stevenson,
wilson, Powers, & Wallace, 2013; Powers et al., 2009)
and that such training produces changes in resting-state
and task-related BOLD functional connectivity among a
network of unisensory and multisensory areas, incluido
the posterior STS (Powers, Hevey, & Wallace, 2012). Tren-
ing could be combined with the information-theoretical
approach used here to test whether pre- and posttraining
comparisons identify similar differences in local and dis-
tributed information processing throughout the brain.
Además, comparing local entropy and mutual infor-
mation across multiple multisensory tasks could distin-
guish effects related to the particularities of each task from
those related to a more general mechanism of multisen-
sory processing and/or response selection underlying
the correlation between tasks (van Atteveldt et al., 2014;
stevenson & Wallace, 2013; Stevenson et al., 2012).
856
Revista de neurociencia cognitiva
Volumen 34, Número 5
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
oh
C
norte
_
a
_
0
1
8
3
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Por último, given the widespread nature of the individual
differences in local entropy and mutual information,
this perspective may help explain the links between
individual differences in multisensory processing and
higher-order cognitive abilities (see Wallace, Woynaroski,
& stevenson, 2020, para una revisión), such as those between
multisensory RT benefit and intelligence in children
(Barutchu et al., 2011), TBW width and problem solving
in young adults (Zmigrod & Zmigrod, 2016), y
audiovisual detection and mild cognitive impairment in
adultos mayores (Murray et al., 2018). Similarmente, este enfoque
may also shed light on mechanisms of dysfunction in
clinical and neurodevelopmental groups with abnormali-
ties in multisensory processing such as schizophrenia
(Martin et al., 2013; Foucher et al., 2007), autism spec-
trum disorder (de Boer-Schellekens et al., 2013; Kwakye
et al., 2011), and dyslexia (Hahn et al., 2014; Hairston
et al., 2005).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
APPENDIX A
Figure A1. Observer model
versus guessing model
likelihood ratios. The observer
model fit was compared with a
two-parameter guessing model
for each participant ( Yarrow,
2018, 2020). The likelihood
ratio was calculated as
likelihood ratio = −2 log
likelihoodguess model − (−2 log
likelihoodobserver model), y el
resulting ratio was compared
with the chi-square distribution
(df = 2). The observer model
provided a superior fit for all
Participantes (all likelihood ratios
>13.82, pag < .001), indicating
that participants were likely
not guessing, and the range of
SOAs used adequately sampled
both transitions (AV and VA)
from simultaneous to
nonsimultaneous perception.
/
j
o
c
n
_
a
_
0
1
8
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Johnston, Alain, and McIntosh
857
APPENDIX B
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
o
c
n
_
a
_
0
1
8
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure B1. Local entropy and observer model parameters—extended PLS results. Left side: Behavior PLS relating all four observer model
parameters (AV and VA width and slope) to local entropy. Only the first LV was significant (LV1: p < .001, singular value = 22.11; LV2: p = .79,
singular value = 5.89, LV3: p = .92, singular value = 4.48, LV4: p = .71, singular value = 3.23). Note that AV slope and VA slope represent
the observer model parameters σAV and σVA, respectively, therefore lower values represent steeper slopes and a sharper transition between
simultaneous and nonsimultaneous judgments. (A) Data-driven contrast for PLS relating all four observer model parameters to local entropy (first LV
only), with p values derived from 1000 permutations. (B) LV1 bootstrap ratios relating all four observer model parameters to local entropy at each
source as a function of timescale. Right side: Behavior PLS relating just the AV and VA slope observer model parameters to local entropy. Only
the first LV was significant (LV1: p = .037, singular value = 13.05; LV2: p = .50, singular value = 5.26). (C) Data-driven contrast for PLS relating the
observer model slope parameters to local entropy (first LV only), with p values derived from 1000 permutations. (D) LV1 bootstrap ratios relating
the observer model slope parameters to local entropy at each source as a function of timescale.
858
Journal of Cognitive Neuroscience
Volume 34, Number 5
APPENDIX C
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
o
c
n
_
a
_
0
1
8
3
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure C1. Mutual information and observer model parameters—extended PLS results. Left side: Behavior PLS relating all four observer model
parameters (AV and VA width and slope) to mutual information. Only the first LV was significant (LV1: p = .001, singular value = 146.91; LV2: p = .97,
singular value = 47.73; LV3: p = .91, singular value = 34.56; LV4: p = .72, singular value = 27.61). Note that AV slope and VA slope represent
the observer model parameters σAV and σVA, respectively; therefore, lower values represent steeper slopes and a sharper transition between
simultaneous and nonsimultaneous judgments. (A) Data-driven contrast for PLS relating all four observer model parameters to mutual information
(first LV only), with p values derived from 1000 permutations. (B) Number of times the given source was involved in a highly reliable (bootstrap
ratio > 4) connection with any other source. Right side: Behavior PLS relating just the AV and VA slope observer model parameters to mutual
información. Neither of the resulting LVs were significant (LV1: pag = .16, singular value = 88.48; LV2: pag = .81, singular value = 36.69). (C) Data-driven
contrast for PLS relating the observer model slope parameters to mutual information (first LV only), with p values derived from 1000 permutations.
(D) Number of times the given source was involved in a highly reliable (bootstrap ratio > 4) connection with any other source.
Johnston, Alaín, and McIntosh
859
Reprint requests should be sent to Phillip R. Johnston, Salir-
ment of Psychology, universidad de toronto, toronto, ontario
M5S 3G3, Canada, o por correo electrónico: phil.johnston@mail.utoronto.ca.
Información de financiación
This work was supported by the Natural Sciences and
Engineering Research Council of Canada (https://dx.doi
.org/10.13039/501100000038), números de subvención: CGS M
and CGS D to P. R. J., RGPIN-2018-04457 to A. R. METRO., y
RGPIN-2016-05523 to C. A.
Diversidad en las prácticas de citas
Análisis retrospectivo de las citas en cada artículo publicado.-
publicado en esta revista de 2010 a 2021 revela una persistente
patrón de desequilibrio de género: Aunque las proporciones de
equipos de autoría (categorizado por iden de género estimado-
tificación del primer autor/último autor) publicando en la revista-
final de neurociencia cognitiva ( JoCN) durante este período
fueron M(un)/m = .407, W.(Omán)/m = .32, M/M = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/M = .109, and W/ W = .085 (Postle y
fulvio, JoCN, 34:1, páginas. 1–3). Como consecuencia, JoCN anima-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance. El
authors of this article report its proportions of citations by
gender category to be as follows: M/M = .574, W/M = .296,
M/M = .093, and W/ W = .037.
Nota
1. Nearly identical results were obtained from the arbitrary
sigmoid fit at the time of data collection, with the width of
the sigmoid at 50% simultaneity perception correlating almost
perfectly with the width estimated from the observer model
(Pearson’s r = .99 for AV stimuli and r = .98 for VA stimuli).
REFERENCIAS
Balk, METRO. h., Ojanen, v., Pekkola, J., Autti, T., Sams, METRO., &
Jääskeläinen, I. PAG. (2010). Synchrony of audio-visual speech
stimuli modulates left superior temporal sulcus. NeuroReport,
21, 822–826. https://doi.org/10.1097/ WNR.0b013e32833d138f,
PubMed: 20588202
Balz, J., Keil, J., Roa Romero, y., Mekle, r., Schubert, F., Aydin,
S., et al. (2016). GABA concentration in superior temporal
sulcus predicts gamma power and perception in the
sound-induced flash illusion. Neuroimagen, 125, 724–730.
https://doi.org/10.1016/j.neuroimage.2015.10.087, PubMed:
26546865
Barutchu, A., Crewther, S. GRAMO., Fifer, J., Shivdasani, METRO. NORTE.,
Innes-Brown, h., Toohey, S., et al. (2011). The relationship
between multisensory integration and IQ in children.
Developmental Psychology, 47, 877–885. https://doi.org/10
.1037/a0021903, PubMed: 21142364
Benevento, l. A., Caer sobre, J., davis, B. J., & Rezak, METRO. (1977).
Auditory–visual interaction in single cells in the cortex of the
superior temporal sulcus and the orbital frontal cortex of the
macaque monkey. Experimental Neurology, 57, 849–872.
https://doi.org/10.1016/0014-4886(77)90112-1, PubMed:
411682
bruce, C., Desimona, r., & Bruto, C. GRAMO. (1981). Visual
properties of neurons in a polysensory area in superior
temporal sulcus of the macaque. Revista de neurofisiología,
46, 369–384. https://doi.org/10.1152/jn.1981.46.2.369,
PubMed: 6267219
Calvert, GRAMO. A., Hansen, PAG. C., Iversen, S. D., & Brammer, METRO. j.
(2001). Detection of audio-visual integration sites in humans
by application of electrophysiological criteria to the BOLD
efecto. Neuroimagen, 14, 427–438. https://doi.org/10.1006
/nimg.2001.0812, PubMed: 11467916
Cecere, r., Bruto, J., & Thut, GRAMO. (2016). Behavioural evidence
for separate mechanisms of audiovisual temporal binding as a
function of leading sensory modality. European Journal of
Neurociencia, 43, 1561–1568. https://doi.org/10.1111/ejn
.13242, PubMed: 27003546
Cecere, r., Bruto, J., willis, A., & Thut, GRAMO. (2017). Being first
matters: Topographical representational similarity analysis
of ERP signals reveals separate networks for audiovisual
temporal binding depending on the leading sense. Diario
of Neuroscience, 37, 5274–5287. https://doi.org/10.1523
/JNEUROSCI.2926-16.2017, PubMed: 28450537
Cecere, r., rees, GRAMO., & Romei, V. (2015). Individual differences
in alpha frequency drive crossmodal illusory perception.
Biología actual, 25, 231–235. https://doi.org/10.1016/j.cub
.2014.11.034, PubMed: 25544613
Col, S., Donoghue, T., gao, r., & Voytek, B. (2019). NeuroDSP:
A package for neural digital signal processing. Diario de
Open Source Software, 4, 1272. https://doi.org/10.21105
/joss.01272
Conrey, B., & Pisoni, D. B. (2006). Auditory–visual speech
perception and synchrony detection for speech and
nonspeech signals. Journal of the Acoustical Society of
America, 119, 4065–4073. https://doi.org/10.1121/1.2195091,
PubMed: 16838548
Costa, METRO., Goldberger, A. l., & Peng, C. k. (2005). Multiescala
entropy analysis of biological signals. Revisión física e, 71,
1–18. https://doi.org/10.1103/PhysRevE.71.021906, PubMed:
15783351
Courtiol, J., Perdikis, D., Betkoski, S., Müller, v., Huys, r.,
Sleimen-Malkoun, r., et al. (2016). The multiscale entropy:
Guidelines for use and interpretation in brain signal
análisis. Journal of Neuroscience Methods, 273, 175–190.
https://doi.org/10.1016/j.jneumeth.2016.09.004, PubMed:
27639660
de Boer-Schellekens, l., Eussen, METRO., & Vroomen, j. (2013).
Diminished sensitivity of audiovisual temporal order in autism
spectrum disorder. Frontiers in Integrative Neuroscience,
7, 1–8. https://doi.org/10.3389/fnint.2013.00008, PubMed:
23450453
Desikan, R. S., Ségonne, F., pescado, B., quinn, B. T., Dickerson,
B. C., Blacker, D., et al. (2006). An automated labeling system
for subdividing the human cerebral cortex on MRI scans into
gyral based regions of interest. Neuroimagen, 31, 968–980.
https://doi.org/10.1016/j.neuroimage.2006.01.021, PubMed:
16530430
Diederich, A., & Colonius, h. (2004). Bimodal and trimodal
multisensory enhancement: Effects of stimulus onset and
intensity on reaction time. Percepción & Psicofísica, 66,
1388–1404. https://doi.org/10.3758/BF03195006, PubMed:
15813202
Conductor, J., & Noesselt, t. (2008). Multisensory interplay reveals
crossmodal influences on “sensory-specific” brain regions,
860
Revista de neurociencia cognitiva
Volumen 34, Número 5
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
oh
C
norte
_
a
_
0
1
8
3
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
neural responses, and judgments. Neurona, 57, 11–23.
https://doi.org/10.1016/j.neuron.2007.12.013, PubMed:
18184561
Duncan, norte. w., & Northoff, GRAMO. (2013). Overview of potential
procedural and participant-related confounds for
neuroimaging of the resting state. Journal of Psychiatry
and Neuroscience, 38, 84–96. https://doi.org/10.1503/jpn
.120059, PubMed: 22964258
Efron, B., & Ruidoso, R. (1986). Bootstrap methods for
errores estándar, confidence intervals and other measures of
statistical accuracy. Statistical Science, 1, 54–77. https://doi
.org/10.1214/ss/1177013815
ángel, A., Fries, PAG., & Cantante, W.. (2001). Dynamic predictions:
Oscillations and synchrony in top–down processing. Naturaleza
Reseñas Neurociencia, 2, 704–716. https://doi.org/10.1038
/35094565, PubMed: 11584308
Ferri, F., Nikolova, Y. S., Perrucci, METRO. GRAMO., Costantini, METRO., Ferretti,
A., Gatta, v., et al. (2017). A neural “tuning curve” for
multisensory experience and cognitive-perceptual
schizotypy. Schizophrenia Bulletin, 43, 801–813. https://doi
.org/10.1093/schbul/sbw174, PubMed: 28168302
Finn, mi. S., Scheinost, D., Finn, D. METRO., shen, X., Papademetris,
X., & Constable, R. t. (2017). Can brain state be
manipulated to emphasize individual differences in
conectividad funcional? Neuroimagen, 160, 140–151. https://
doi.org/10.1016/j.neuroimage.2017.03.064, PubMed:
28373122
fonov, V. S., evans, A. C., Mckinstry, R. C., almíli, C. r., & collins,
D. l. (2009). Unbiased nonlinear average age-appropriate
brain templates from birth to adulthood. Neuroimagen, 47,
S102. https://doi.org/10.1016/S1053-8119(09)70884-5
Foucher, j. r., Lacambre, METRO., Pham, B. T., Giersch, A., & eliot,
METRO. A. (2007). Low time resolution in schizophrenia.
Lengthened windows of simultaneity for visual, auditivo
and bimodal stimuli. Schizophrenia Research, 97, 118–127.
https://doi.org/10.1016/j.schres.2007.08.013, PubMed:
17884350
Hillock-Dunn, A., & Wallace, METRO. t. (2012). Developmental
changes in the multisensory temporal binding window
persist into adolescence. Developmental Science, 15,
688–696. https://doi.org/10.1111/j.1467-7687.2012.01171.x,
PubMed: 22925516
jones, mi. GRAMO., & Powell, t. PAG. S. (1970). An anatomical study of
converging sensory pathways within the cerebral cortex of
the monkey. Cerebro, 93, 793–820. https://doi.org/10.1093
/brain/93.4.793, PubMed: 4992433
Kaganovich, NORTE., & Schumaker, j. (2016). Electrophysiological
correlates of individual differences in perception of
audiovisual temporal asynchrony. Neuropsicología, 86,
119–130. https://doi.org/10.1016/j.neuropsychologia.2016.04
.015, PubMed: 27094850
Kambe, J., Kakimoto, y., & Araki, oh. (2015). Phase reset
affects auditory–visual simultaneity judgment. Cognitivo
Neurodynamics, 9, 487–493. https://doi.org/10.1007/s11571
-015-9342-4, PubMed: 26379799
Keil, J., & Senkowski, D. (2018). Neural oscillations
orchestrate multisensory processing. neurocientífico,
24, 1–18. https://doi.org/10.1177/1073858418755352,
PubMed: 29424265
Kumar, V. GRAMO., Dutta, S., Talwar, S., roy, D., & Banerjee, A.
(2020). Biophysical mechanisms governing large-scale
brain network dynamics underlying individual-specific
variability of perception. European Journal of Neuroscience,
52, 3746–3762. https://doi.org/10.1111/ejn.14747, PubMed:
32304122
Kwakye, l. D., Foss-Feig, j. h., Cascio, C. J., Piedra, W.. l., &
Wallace, METRO. t. (2011). Altered auditory and multisensory
temporal processing in autism spectrum disorders. Frontiers
in Integrative Neuroscience, 4, 1–11. https://doi.org/10.3389
/fnint.2010.00129, PubMed: 21258617
Magnotti, j. F., Mamá, W.. J., & Beauchamp, METRO. S. (2013).
Causal inference of asynchronous audiovisual speech.
Fronteras en psicología, 4, 798. https://doi.org/10.3389/fpsyg
.2013.00798, PubMed: 24294207
Fries, PAG. (2009). Neuronal gamma-band synchronization as a
Makeig, S., Campana, A. J., Junk, T.-P., & Sejniano, t. j. (1996).
fundamental process in cortical computation. Annual Review
of Neuroscience, 32, 209–224. https://doi.org/10.1146
/annurev.neuro.051508.135603, PubMed: 19400723
Ghazanfar, A. A., & Schroeder, C. mi. (2006). Is neocortex
essentially multisensory? Tendencias en Ciencias Cognitivas, 10,
278–285. https://doi.org/10.1016/j.tics.2006.04.008, PubMed:
16713325
Gramfort, A., Papadopoulo, T., Olivi, MI., & Clerc, METRO. (2010).
OpenMEEG: Opensource software for quasistatic
bioelectromagnetics. Biomedical Engineering Online, 9, 45.
https://doi.org/10.1186/1475-925X-9-45, PubMed: 20819204
Hahn, NORTE., Foxe, j. J., & Molholm, S. (2014). Impairments of
multisensory integration and cross-sensory learning as
pathways to dyslexia. Neuroscience and Biobehavioral
Reseñas, 47, 384–392. https://doi.org/10.1016/j.neubiorev
.2014.09.007, PubMed: 25265514
Hairston, W.. D., Burdette, j. h., Flores, D. l., Madera, F. B.,
& Wallace, METRO. t. (2005). Altered temporal profile of
visual-auditory multisensory interactions in dyslexia.
Experimental Brain Research, 166, 474–480.
https://doi.org/10.1007/s00221-005-2387-6, PubMed:
16028030
Hein, GRAMO., & Caballero, R. t. (2008). Superior temporal sulcus—It’s
my area: Or is it? Revista de neurociencia cognitiva, 20,
2125–2136. https://doi.org/10.1162/jocn.2008.20148,
PubMed: 18457502
Hershenson, METRO. (1962). Reaction time as a measure of
intersensory facilitation. Revista de Psicología Experimental,
63, 289–293. https://doi.org/10.1037/h0039516, PubMed:
13906889
Independent component analysis of electroencephalographic
datos. En D. S. Touretzky, METRO. C. Mozer, & METRO. mi. Hasselmo
(Editores.), Advances in neural information processing systems
(volumen. 8, páginas. 145–151). Cambridge, MAMÁ: CON prensa.
Marchant, j. l., Fallar, C. C., & Conductor, j. (2012). Audiovisual
synchrony enhances BOLD responses in a brain network
including multisensory STS while also enhancing
target-detection performance for both modalities. Humano
Mapeo cerebral, 33, 1212–1224. https://doi.org/10.1002/hbm
.21278, PubMed: 21953980
Martín, B., Giersch, A., Huron, C., & van Wassenhove, V. (2013).
Temporal event structure and timing in schizophrenia:
Preserved binding in a longer “now.” Neuropsychologia, 51,
358–371. https://doi.org/10.1016/j.neuropsychologia.2012.07
.002, PubMed: 22813430
McIntosh, A. r., Bookstein, F. l., Haxby, j. v., & grady, C. l.
(1996). Spatial pattern analysis of functional brain images
using partial least squares. Neuroimagen, 157, 143–157.
https://doi.org/10.1006/nimg.1996.0016, PubMed: 9345485
McIntosh, A. r., Kovacevic, NORTE., & Itier, R. j. (2008). Increased
brain signal variability accompanies lower behavioral
variability in development. Biología Computacional PLoS,
4, e1000106. https://doi.org/10.1371/journal.pcbi.1000106,
PubMed: 18604265
McIntosh, A. r., & Lobaugh, norte. j. (2004). Partial least squares
analysis of neuroimaging data: Applications and advances.
Neuroimagen, 23, 250–263. https://doi.org/10.1016/j
.neuroimage.2004.07.020, PubMed: 15501095
Molinero, l. METRO., & D'Esposito, METRO. (2005). Perceptual fusion
and stimulus coincidence in the cross-modal integration
Johnston, Alaín, and McIntosh
861
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
oh
C
norte
_
a
_
0
1
8
3
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
of speech. Revista de neurociencia, 25, 5884–5893. https://
doi.org/10.1523/JNEUROSCI.0896-05.2005, PubMed:
15976077
of Sciences, EE.UU., 117, 22522–22531. https://doi.org/10.1073
/pnas.2005993117, PubMed: 32839338
stevenson, R. A., Altieri, norte. A., kim, S., Pisoni, D. B., & Jaime, t. W..
Monosov, I. MI., Haber, S. NORTE., Leuthardt, mi. C., & Jezzini, A.
(2020). Anterior cingulate cortex and the control of
dynamic behavior in primates. Biología actual, 30,
R1442–R1454. https://doi.org/10.1016/j.cub.2020.10.009,
PubMed: 33290716
Morcom, A. METRO., & Fletcher, PAG. C. (2007). Does the brain have a
base? Why we should be resisting a rest. Neuroimagen, 37,
1073–1082. https://doi.org/10.1016/j.neuroimage.2006.09
.013, PubMed: 17681817
Murray, METRO. METRO., Eardley, A. F., Edginton, T., Oyekan, r.,
Smyth, MI., & Matusz, PAG. j. (2018). Sensory dominance
and multisensory integration as screening tools in aging.
Informes Científicos, 8, 8901. https://doi.org/10.1038/s41598-018
-27288-2, PubMed: 29891964
Nath, A. r., & Beauchamp, METRO. S. (2012). A neural basis for
interindividual differences in the McGurk effect, a
multisensory speech illusion. Neuroimagen, 59, 781–787.
https://doi.org/10.1016/j.neuroimage.2011.07.024, PubMed:
21787869
Noesselt, T., Rieger, j. w., Schoenfeld, METRO. A., Kanowski, METRO.,
Hinrichs, h., Heinze, H.-J., et al. (2007). Audiovisual
temporal correspondence modulates human multisensory
superior temporal sulcus plus primary sensory
cortices. Revista de neurociencia, 27, 11431–11441.
https://doi.org/10.1109/SENSOR.2007.4300668, PubMed:
17942738
Pascual-Marqui, R. D. (2002). Standardized low resolution brain
electromagnetic tomography (sLORETA): Technical details.
Methods and Findings in Experimental and Clinical
Pharmacology, 24 Suplemento. D, 5–12.
Peirce, J., Gris, j. r., Simpson, S., Macaskill, METRO., Höchenberger, r.,
Sogo, h., et al. (2019). PsychoPy2: Experiments in behavior
made easy. Métodos de investigación del comportamiento, 51, 195–203.
https://doi.org/10.3758/s13428-018-01193-y, PubMed:
30734206
Powers, A. r., Hevey, METRO. A., & Wallace, METRO. t. (2012). Neural
correlates of multisensory perceptual learning. Diario de
Neurociencia, 32, 6263–6274. https://doi.org/10.1523
/jneurosci.6138-11.2012, PubMed: 22553032
Powers, A. r., Hillock, A. r., & Wallace, METRO. t. (2009). Perceptivo
training narrows the temporal window of multisensory
binding. Revista de neurociencia, 29, 12265–12274. https://
doi.org/10.1523/JNEUROSCI.3501-09.2009, PubMed:
19793985
sansón, PAG. D., Streissguth, PAG., barr, h. METRO., & Bookstein, F. l.
(1989). Neurobehavioural effects of prenatal alcohol.
Part II. Partial least squares analysis. Neurotoxicology and
Teratology, 11, 477–491. https://doi.org/10.1016/0892-0362
(89)90025-1, PubMed: 2593987
Schroeder, C. MI., & Foxe, j. j. (2002). The timing and laminar
profile of converging inputs to multisensory areas of the
macaque neocortex. Cognitive Brain Research, 14, 187–198.
https://doi.org/10.1016/s0926-6410(02)00073-3, PubMed:
12063142
Senkowski, D., Schneider, t. r., Foxe, j. J., & ángel, A. k.
(2008). Crossmodal binding through neural coherence:
Implications for multisensory processing. Tendencias en
Neurosciences, 31, 401–409. https://doi.org/10.1016/j.tins
.2008.05.002, PubMed: 18602171
Shams, l., & Seitz, A. R. (2008). Benefits of multisensory
aprendiendo. Tendencias en Ciencias Cognitivas, 30, 1–7. https://doi
.org/10.1016/j.tics.2008.07.006, PubMed: 18805039
Spitmaan, METRO., SEO, h., Sotavento, D., & Soltani, A. (2020). Múltiple
timescales of neural dynamics and integration of task-relevant
signals across cortex. Proceedings of the National Academy
(2010). Neural processing of asynchronous audiovisual
speech perception. Neuroimagen, 49, 3308–3318.
https://doi.org/10.1016/j.neuroimage.2009.12.001, PubMed:
20004723
stevenson, R. A., VanDerKlok, R. METRO., Pisoni, D. B., & Jaime, t. W..
(2011). Discrete neural substrates underlie complementary
audiovisual speech integration processes. Neuroimagen, 55,
1339–1345. https://doi.org/10.1016/j.neuroimage.2010.12
.063, PubMed: 21195198
stevenson, R. A., & Wallace, METRO. t. (2013). Multisensory temporal
integración: Task and stimulus dependencies. Experimental
Brain Research, 227, 249–261. https://doi.org/10.1007/s00221
-013-3507-3, PubMed: 23604624
stevenson, R. A., wilson, METRO. METRO., Powers, A. r., & Wallace, METRO. t.
(2013). The effects of visual training on multisensory
temporal processing. Experimental Brain Research, 225,
479–489. https://doi.org/10.1007/s00221-012-3387-y, PubMed:
23307155
stevenson, R. A., Zemtsov, R. K., & Wallace, METRO. t. (2012).
Individual differences in the multisensory temporal binding
window predict susceptibility to audiovisual illusions.
Revista de Psicología Experimental: Percepcion humana
y rendimiento, 38, 1517–1529. https://doi.org/10.1037
/a0027339, PubMed: 22390292
Sumby, W.. h., & Pollack, I. (1954). Visual contribution to
speech intelligibility in noise. Journal of the Acoustical
Sociedad de América, 26, 212–215. https://doi.org/10.1121/1
.1907309
Tadel, F., Baillet, S., Mosher, j. C., Pantazis, D., & Leahy, R. METRO.
(2011). Brainstorm: A user-friendly application for MEG/EEG
análisis. Computational Intelligence and Neuroscience,
2011, 1–13. https://doi.org/10.1155/2011/879716, PubMed:
21584256
Vakorin, V. A., Lippé, S., & McIntosh, A. R. (2011). Variabilidad
of brain signals processed locally transforms into higher
connectivity with brain development. Revista de neurociencia,
31, 6405–6413. https://doi.org/10.1523/JNEUROSCI.3153-10
.2011, PubMed: 21525281
van Atteveldt, NORTE., Murray, METRO. METRO., Thut, GRAMO., & Schroeder, C. mi.
(2014). Multisensory integration: Flexible use of general
operaciones. Neurona, 81, 1240–1253. https://doi.org/10.1016/j
.neuron.2014.02.044, PubMed: 24656248
van Eijk, R. l. J., Kohlrausch, A., Juola, j. F., & van de Par, S.
(2008). Audiovisual synchrony and temporal order
judgments: Effects of experimental method
and stimulus type. Percepción & Psicofísica, 70,
955–968. https://doi.org/10.3758/PP.70.6.955, PubMed:
18717383
Wallace, METRO. T., Woynaroski, t. GRAMO., & stevenson, R. A. (2020).
Multisensory integration as a window into orderly and
disrupted cognition and communication. Annual Review of
Psicología, 71, 193–219. https://doi.org/10.1146/annurev
-psych-010419-051112, PubMed: 31518523
Yarrow, K., (2018). Collecting and interpreting judgments
about perceived simultaneity: A model-fitting tutorial.
En. Vatakis, F. Balcı, METRO. Di Luca, & Á. Correa (Editores.),
Timing and time perception: Procedures,
measures, & aplicaciones (páginas. 295–325). Leiden,
Los países bajos: Rodaballo. https://doi.org/10.1163
/9789004280205_014
Yarrow, K., (2020). MATLAB code to implement fits of
psychophysical observer models to data from
behavioural timing tasks (SJ, TOJ, 2xSJ, ternary)
( Versión 1). City, Universidad de Londres. https://doi.org/10
.25383/city.11848263.v1
862
Revista de neurociencia cognitiva
Volumen 34, Número 5
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
oh
C
norte
_
a
_
0
1
8
3
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Yarrow, K., Jahn, NORTE., Durant, S., & arnold, D. h. (2011). Shifts of
criteria or neural timing? The assumptions underlying timing
perception studies. Conciencia y cognición, 20,
1518–1531. https://doi.org/10.1016/j.concog.2011.07.003
Yuan, X., li, h., Liu, PAG., Yuan, h., & Huang, X. (2016).
Pre-stimulus beta and gamma oscillatory power predicts
perceived audiovisual simultaneity. Revista internacional de
Psychophysiology, 107, 29–36. https://doi.org/10.1016/j
.ijpsycho.2016.06.017, PubMed: 27393081
zhang, Y. (1991). Complexity and 1/f noise. A phase space
acercarse. Journal de Physique I, 1, 971–977. https://doi.org
/10.1051/jp1:1991180
Zmigrod, l., & Zmigrod, S. (2016). On the temporal precision
of thought: Individual differences in the multisensory
temporal binding window predict performance on
verbal and nonverbal problem solving tasks. Multisensory
Investigación, 29, 679–701. https://doi.org/10.1163/22134808
-00002532
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
5
8
4
6
2
0
0
4
5
8
8
/
j
oh
C
norte
_
a
_
0
1
8
3
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Johnston, Alaín, and McIntosh
863