Unraveling the Relation between EEG Correlates
of Attentional Orienting and Sound Localization
Performance: A Diffusion Model Approach
Laura-Isabelle Klatt1, Daniel Schneider1, Anna-Lena Schubert2, Christina Hanenberg1,
Jörg Lewald1,3, Edmund Wascher1, and Stephan Getzmann1
Abstrait
■ Understanding the contribution of cognitive processes and
their underlying neurophysiological signals to behavioral phenom-
ena has been a key objective in recent neuroscience research.
Using a diffusion model framework, we investigated to what ex-
tent well-established correlates of spatial attention in the elec-
troencephalogram contribute to behavioral performance in an
auditory free-field sound localization task. Younger and older par-
ticipants were instructed to indicate the horizontal position of a
predefined target among three simultaneously presented distrac-
tors. The central question of interest was whether posterior alpha
lateralization and amplitudes of the anterior contralateral N2 sub-
component (N2ac) predict sound localization performance (accu-
racy, mean RT) and/or diffusion model parameters (drift rate,
boundary separation, non-decision time). Two age groups were
compared to explore whether, in older adults (who struggle with
multispeaker environments), the brain–behavior relationship
would differ from younger adults. Regression analyses revealed
that N2ac amplitudes predicted drift rate and accuracy, alors que
alpha lateralization was not related to behavioral or diffusion
modeling parameters. This was true irrespective of age. The re-
sults indicate that a more efficient attentional filtering and se-
lection of information within an auditory scene, reflected by
increased N2ac amplitudes, was associated with a higher speed
of information uptake (drift rate) and better localization per-
formance (accuracy), while the underlying response criteria
(threshold separation), mean RTs, and non-decisional pro-
cesses remained unaffected. The lack of a behavioral correlate
of poststimulus alpha power lateralization constrasts with the
well-established notion that prestimulus alpha power reflects
a functionally relevant attentional mechanism. This highlights
the importance of distinguishing anticipatory from poststimulus
alpha power modulations. ■
INTRODUCTION
When multiple sources of acoustic information are simul-
taneously present, selective filtering of the available in-
formation is necessary to, par exemple, focus on a talker
of interest while ignoring traffic noise, music playing in
the background, or other peoples’ conversations. Ce
capacity of the human auditory system is especially as-
tonishing, given that the incoming auditory signals often
overlap in time, espace, or spectral content. The behav-
ioral effects of such selective orienting of attention in
noisy, multispeaker environments, usually referred to as
“cocktail party scenarios” (Cherry, 1953), have been stud-
ied for decades (for a review, see Bronkhorst, 2015).
Cependant, the contribution of neural signals to observable
behavioral performance and its underlying cognitive pro-
cesses is still poorly understood. Ici, we investigated
the relationship between well-established correlates of
spatial attention in the electroencephalogram (EEG)
and behavioral performance in an auditory sound local-
1Leibniz Research Centre for Working Environment and Human
Factors, 2Heidelberg University, 3Ruhr-University Bochum
© 2020 Massachusetts Institute of Technology
ization task. En particulier, we specified the role of modu-
lations in the alpha frequency band as well as an anterior
contralateral N2 subcomponent (N2ac; Gamble & Luck,
2011) with respect to sound localization performance.
Lateralized modulations of alpha power amplitude
have been shown to reflect the orienting of spatial atten-
tion in visual (Foster, Sutterer, Serences, Vogel, & Awh,
2017; Ikkai, Dandekar, & Curtis, 2016; Rihs, Michel, &
Thut, 2007; Worden, Foxe, Wang, & Simpson, 2000), tac-
tile (Haegens, Luther, & Jensen, 2012; Haegens, Händel,
& Jensen, 2011), and auditory space (Klatt, Getzmann,
Wascher, & Schneider, 2018b; Wöstmann, Vosskuhl,
Obleser, & Herrmann, 2018; Wöstmann, Herrmann, Maess,
& Obleser, 2016). Typiquement, alpha power is shown to
decrease contralaterally to the attended location (Kelly,
Gomez-Ramirez, & Foxe, 2009; Sauseng et al., 2005) or to in-
crease contralaterally to the unattended or ignored location
(Kelly, Lalor, Reilly, & Foxe, 2006; Worden et al., 2000).
Consistently across different modalities, this lateralized
pattern of alpha-band activity has been shown to be linked
to visual detection performance (Händel, Haarmeier, &
Jensen, 2011; van Dijk, Schoffelen, Oostenveld, & Jensen,
2008; Thut, Nietzel, Brandt, & Pascual-Leone, 2006), tactile
Journal des neurosciences cognitives 32:5, pp. 945–962
https://doi.org/10.1162/jocn_a_01525
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
n
t
p
o
:
un
/
d
/
e
d
m
je
F
r
t
o
p
m
r
c
h
.
p
s
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
2
c
je
5
e
9
–
4
p
5
d
F
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
un
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
/
.
t
F
o
n
0
5
M.
un
oui
2
0
2
1
discrimination acuity (Craddock, Poliakoff, El-deredy,
Klepousniotou, & Lloyd, 2017; Haegens et al., 2011), et
listening performance (Tune, Wöstmann, & Obleser, 2018;
Wöstmann et al., 2016). Going beyond a mere correlational
approche, recent studies applying stimulation techniques,
such as TMS or continuous transcranial alternating current
stimulation, suggest a causal role of alpha oscillations in the
processing of incoming information (Wöstmann et al., 2018;
Romei, Gross, & Thut, 2010). Two major (not necessarily
mutually exclusive) mechanisms have been proposed to
underlie those asymmetric modulations of alpha power
oscillations: target enhancement (Noonan et al., 2016;
Yamagishi, Goda, Callan, Anderson, & Kawato, 2005)
and distractor inhibition (Schneider, Göddertz, Haase,
Hickey, & Wascher, 2019; Rihs et al., 2007; Kelly et al.,
2006; Worden et al., 2000). Although the majority of pre-
vious studies investigated prestimulus alpha oscillations
as an index of anticipatory allocation of spatial attention
in young adults, we focused on poststimulus alpha later-
alization in a sound localization task, simulating a “cocktail
party scenario.” Such an experimental setup more closely
resembles frequent real-life situations, in which a person
searches for a sound of interest (par exemple., a voice or a ringing
phone) without knowing in advance where to look for it. Dans
fact, there is first evidence that distinct attentional mecha-
nisms contribute to the preparation for as opposed to the
ongoing processing of a stimulus (van Ede, Szebényi, &
Maris, 2014). En outre, we explore whether the pro-
posed mechanistic function of alpha oscillations extends
to samples of older participants, which remains an ongoing
matter of debate (Tune et al., 2018; Mok, Myers, Wallis, &
Nobre, 2016; Hong, Sun, Bengson, Mangun, & Tong, 2015;
Vaden, Hutcheson, McCollum, Kentros, & Visscher, 2012).
A second neural measure of interest, indicating the alloca-
tion of attention within an auditory scene, is the N2ac. Le
N2ac has been shown to be evoked in the N2 latency range
(starting at around 200 msec) when detecting or localizing a
target sound in the presence of one or multiple distractor
stimuli, using artificial sounds (Gamble & Luck, 2011), ani-
mal vocalizations (Klatt, Getzmann, Wascher, & Schneider,
2018un; Lewald & Getzmann, 2015), or spoken numerals
(Lewald, Hanenberg, & Getzmann, 2016). Bien que le
N2ac was originally suggested to reflect the allocation of
selective attention to the target (Gamble & Luck, 2011),
analogously to the visual posterior contralateral N2 sub-
component (N2pc; Eimer, 1996; Luck & Hillyard, 1994),
its functional significance remains ambiguous. Ici, nous
aimed to provide further evidence on the functional signif-
icance of the N2ac by investigating its relationship to sound
localization performance.
Dans cette étude, the diffusion modeling approach (Ratcliff,
1978) was applied, allowing for a more detailed under-
standing of behavioral patterns in discrimination tasks (pour
recent reviews, see Voss, Nagler, & Lerche, 2013; Ratcliff &
McKoon, 2008). Although diffusion models are still only
rarely used in cognitive neuroscience research (voir, par exemple.,
Schubert, Nunez, Hagemann, & Vandekerckhove, 2019;
Nunez, Vandekerckhove, & Srinivasan, 2017; Schubert,
Hagemann, Voss, Schankin, & Bergmann, 2015; Ratcliff,
Philiastides, & Sajda, 2009; Philiastides, Ratcliff, & Sajda,
2006), the interest in and the application of this methodo-
logical approach has increased considerably during the past
decade. The general purpose of diffusion models is to
decompose the cognitive processes underlying a binary
décision. As one of the major advantages of the diffusion
model, the estimation procession is not limited to single
mean or median values but takes the whole RT distribution
into account. Spécifiquement, the resulting separation of pro-
cessing components offers an enormous potential to pro-
vide more detailed descriptions of cognitive processes
and to generate more accurate predictions for behavioral
and neurophysiological data (Tourneur, Rodriguez, Norcia,
McClure, & Steyvers, 2016; Ratcliff & McKoon, 2008).
The diffusion model assumes that, in order for a deci-
sion to be made and a reaction to be executed, evidence
for either response is accumulated in the course of a noisy
process until it reaches either the decision boundary of re-
sponse A or response B (voir la figure 2 in Voss et al., 2013
for an illustration of this evidence accumulation process).
The basic diffusion model includes the following pa-
rameters: The drift rate v describes the speed at which
evidence is accumulated (or “the rate of accumulation of
information”; Ratcliff & McKoon, 2008, p. 3), with higher
drift rates resulting in shorter RTs and fewer errors.
Threshold separation a indicates the amount of infor-
mation considered until a decision is made. C'est, con-
servative response criteria that are associated with slower
but more accurate responses result in large estimates of a,
whereas more liberal response criteria result in smaller es-
timates of a. Threshold separation and drift rate have been
shown to be negatively correlated due to the fact that in-
dividuals with higher drift rates tend to allow more liberal
response criteria (c'est à dire., smaller threshold separation values;
Schmiedek, Oberauer, Wilhelm, Süß, & Wittmann, 2007).
A priori biases toward one of the decision thresholds are
reflected by starting point z. Beyond that, the model also
includes non-decisional processing, such as response exe-
cution, working memory access, or stimulus encoding.
The latter is indicated by the non-decision time constant
t0. Typiquement, older adults show a slowing in this decision-
unrelated domain (Ratcliff, Thapar, Gomez, & McKoon,
2004; Ratcliff, Thapar, & McKoon, 2001). Enfin, trial-to-
trial variability in drift rate (sv), non-decision time (st0),
starting point (sz), and the proportion of contaminated tri-
als ( pdiff; par exemple., underlying non-diffusion-like processes) peut
be accounted for.
En résumé, here we aimed at characterizing the rela-
tion between electrophysiological correlates of atten-
tional orienting within a complex auditory scene (c'est à dire.,
alpha lateralization and N2ac) and sound localization
performance, which was assessed by classical RT and accu-
racy measures as well as by diffusion modeling parameters.
We hypothesized that, if the cognitive processes reflected by
alpha power modulations and N2ac amplitudes contribute
946
Journal des neurosciences cognitives
Volume 32, Nombre 5
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
n
t
p
o
:
un
/
d
/
e
d
m
je
F
r
t
o
p
m
r
c
h
.
p
s
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
2
c
je
5
e
9
–
4
p
5
d
F
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
un
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
t
/
F
.
o
n
0
5
M.
un
oui
2
0
2
1
to the successful selection of the target from a sound array
containing simultaneously present distractors, they should
in turn contribute to the information accumulation process
that results in the localization of the target. Ainsi, alpha
power modulations and N2ac amplitudes should predict
drift rate (c'est à dire., the speed of information accumulation)
et, à son tour, RT and accuracy.
The data analyzed here were taken from a separate study
on effects of auditory training on cocktail party listening
performance in younger and older adults (Hanenberg,
Getzmann, & Lewald, unpublished). Exclusively pretraining
data of this study were used. The sample analyzed here in-
cluded both age groups. Although we did not primarily aim
at the investigation of age effects, age differences with re-
spect to sound localization performance, alpha lateralization
and N2ac, as well as the relation between these electro-
physiological correlates of attentional orienting and sound
localization performance were considered. Irrespective of
the expected age-related decline, we proposed the latter
brain–behavior relationship to be true for both age groups.
MÉTHODES
Participants
The original sample included 28 older adults and 24
younger adults. Data for three younger participants were
discarded because of technical problems with the EEG
recording. En outre, two older participants were ex-
cluded from analysis because their performance was be-
faible (14% correct) or very close to (30%) chance level
(25%). Par conséquent, the final sample included 26 older
adultes (âge moyen = 69 années, range = 56–76 years, 13
femmes) et 21 younger adults (âge moyen = 24 années,
range = 19–29 years, 11 femmes). All participants were
right-handed as assessed by the Edinburgh Handedness
Inventory (Oldfield, 1971).
An audiometry, y compris 11 pure-tone frequencies (0.125–
8 kHz; Oscilla USB100, Inmedico) was conducted. Hearing
thresholds in the speech frequency range (<4 kHz) in-
dicated normal hearing (≤25 dB) for all younger par-
ticipants and mild impairments older participants
(≤40 dB). The study was conducted in accordance with
the Declaration of Helsinki approved by the
Ethical Committee the Leibniz Research Centre for
Working Environment Human Factors. All participants
gave their written informed consent participation.
Experimental Setup, Procedure, Stimuli
The original study, which data were collected (Hanenberg
et al., unpublished), comprised three training sessions on
3 days, with experimental blocks per session (15 min
pretraining, 15 min posttraining, 1 hr posttraining) and
with intervals 1–3 weeks between sessions. For pres-
ent reanalysis, exclusively obtained pre-
training blocks, pooled across sessions, were
used. experiment a dimly lit, echo-
reduced, sound-proof room. Participants seated a
comfortable chair that positioned equal distances
to left, right, front wall Participants’
head position stabilized chin rest. A semicircular
array nine broadband loudspeakers (SC5.9; Visaton;
housing volume 340 cm3) mounted par-
ticipant at distance 1.5 m from participant’s head.
Only four loudspeakers, located azimuthal positions of
−60°, −20°, 20°, 60°, used experimental
setup this study. red light-emitting diode (diameter =3 mm, luminous intensity =0.025 mcd) attached right
below central loudspeaker median plane the
participant’s head eye level. was
continuously on served as fixation point.
The sound localization task applied a
modification multiple-sources approach has
been several previous studies auditory selec-
tive spatial attention “cocktail party scenarios” (Lewald,
2016, 2019; Lewald & Getzmann, 2015; Zündorf, Karnath,
& Lewald, 2011, 2014; Karnath, 2013).
Details present version have been previously
described (Lewald et 2016). Briefly, participants indi-
cated predefined target numeral was
presented simultaneously distractor numerals.
The kept constant each participant was
counterbalanced age groups such
that an number of
times within overall experiment. Four 1-syllable nu-
merals (“eins,” 1; “vier,” 4; “acht,” 8; “zehn,” 10), spoken
by two male (mean pitch =141 Hz) female (mean
pitch =189 native German speakers, sound
stimuli numerals presented
equally often possible posi-
tions (located −60°, 60° azimuth). Numerals
presented trial spoken different
speakers. pressure level sound
arrays 66 dB(A), measured par-
ticipant’s using sound-level meter 0.5-in. free-
field measuring microphone (Types 2226 4175, Brüel &
Kjær).
The trial, posi-
tion, positions, speakers varying between
trials following fixed pseudorandom order. stimu-
lus duration 600 msec, followed response period
of 2 sec intertrial interval 525 resulting in
a total 3.125 sec. given
by pressing one out buttons the
index finger right hand. buttons
were arranged semicircular array, related to four
possible locations (i.e., far inner inner
right, right). Each block consisted 288 trials, re-
sulting block. As already
mentioned above, assessed on
different pooled. Thus, there of
864 trials participant. On 3 partici-
pants completed short sequence 10 trials
Klatt al.
947
D
o
w
n
l
o
a
d
e
d
l
l
>3000 msec) RTs were discarded. Subsequently, data
were log-transformed and z-standardized to exclude all
trials with RTs exceeding ± 3 SDs of the mean for each
individual participant.
The free software fast-dm ( Voss & Voss, 2007) était
used to fit a diffusion model to the RT distributions of
the present data. The model parameters were estimated
based on an iterative permutation process using the
Kolmogrov-Smirnov test statistic. The starting point z
was set to 0.5, presuming that participants were not bi-
ased toward one of the two response categories (correct
target location vs. distractor locations). The parameters
un, v, and t0 were allowed to vary freely. En outre, Pennsylvanie-
rameters sv and st0 were estimated because they led to a
notable improvement of model fit. Trial-to-trial variability
of starting point (sz), the difference in speed of response
execution (d ), as well as the measure for the percentage
of contaminants ( pdiff) were set to 0. To graphically eval-
uate model fit, we plotted observed versus predicted ac-
curacy as well as observed versus predicted values of the
RT distribution for the first (.25), second (.50), and third
(.75) quartile. Predicted parameter values were derived
using the construct-samples tool of fast-dm ( Voss &
948
Journal des neurosciences cognitives
Volume 32, Nombre 5
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
n
t
p
o
:
un
/
d
/
e
d
m
je
F
r
t
o
p
m
r
c
h
.
p
s
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
2
c
je
5
e
9
–
4
p
5
d
F
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
un
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
F
.
t
/
o
n
0
5
M.
un
oui
2
0
2
1
Voss, 2007). C'est, 500 data sets were generated for
each participant based on each individual’s empirical pa-
rameter values and number of trials. Enfin, the mean
quartile values and mean response accuracy were cal-
culated for each participant. Pearson correlations were
calculated to quantify the relationship between empirical
data and model predictions for both age groups. Si le
majority of data points lie close to the line of perfect cor-
relation, good model fit can be assumed.
ERP Analysis
To investigate the N2ac component (Gamble & Luck, 2011),
we computed the mean contralateral and ipsilateral ERP
amplitude at frontocentral electrodes FC3/4 for older
adults and FC5/6 for younger adults. The contralateral
portion comprised the average signal at left hemispheric
electrodes in right target trials and right hemispheric elec-
trodes in left target trials, whereas the ipsilateral portion
included the average signal at left hemispheric electrodes
in left target trials and right hemispheric electrodes in
right target trials. Mean amplitude was measured from
477 à 577 msec relative to sound array onset. The mea-
surement window was based on a 100-msec time window
set around the 50% fractional area latency (FAL; Luck,
2014; Hansen & Hillyard, 1980) in the grand-averaged
contralateral minus ipsilateral difference curve averaged
across age groups and electrodes (50% FAL = 527 msec).
To determine the FAL, the area under the difference
curve was measured in a broad time window ranging from
200 à 800 msec relative to sound array onset. The latency
at which this area is divided in two equal halves denotes
le 50% FAL. We determined a common analysis time
window for both age groups because a prior control anal-
ysis did not reveal any significant differences between the
50% FAL for younger (M = 525.86 msec) and older adults
(M = 517.50 msec), Z = 0.26, p = .80, U3 = 0.48. Le
respective electrodes of interest (c'est à dire., FC3/4 and FC5/6)
were chosen to include the scalp sites with the most
pronounced asymmetry (c'est à dire., peak asymmetry in the age-
specific grand-averaged waveform) for each age group.
This age-specific mean amplitude was measured in the
time window specified above.
Time–Frequency Data
To obtain time–frequency representations of the single-trial
oscillatory power, we convolved the epoched, stimulus-
locked EEG data with three-cycle complex Morlet wavelets.
The number of cycles increased with frequency by a factor
de 0.5, c'est, half as fast as the number of cycles in the
respective fast Fourier transformation. This resulted in
three-cycle wavelets at the lowest frequency (4 Hz) et
11.25-cycle wavelets at the highest frequency (30 Hz). À
quantify asymmetries in the attentional modulation of total
oscillatory power (induced + evoked activity), the alpha lat-
eralization index (ALI) was calculated (Wildegger, van Ede,
Woolrich, Gillebert, & Nobre, 2017; Wöstmann et al., 2016;
Haegens et al., 2011). The latter quantifies the strength of
the ipsilateral minus contralateral difference in alpha power
relative to the total power across both hemispheres:
Þ
ð
ALI ¼ ipsilateral alpha power−contralateral alpha power
Þ
ð
ipsilateral alpha power þ contralateral alpha power
(1)
This normalization controls for potential confounds through
differences in overall power level when comparing the
two age groups. Mean ipsilateral and contralateral power
was extracted in the alpha frequency band (8–12 Hz) à
electrodes PO7/PO8 in a time window ranging from 705
à 902 msec relative to the onset of the sound array. Le
measurement window was based on a 200-msec time win-
dow set around the 50% FAL in the ALI difference curve av-
eraged across age groups (50% FAL = 804 msec). Le 50%
FAL was calculated based on a broad time window ranging
depuis 300 à 1400 msec relative to sound array onset. Nous
determined a common analysis time window for both age
groupes, because a control analysis did not reveal any signif-
icant differences between the 50% FAL for younger (M =
796.00 msec) and older adults (M = 860.57 msec), Z =
−1.27, p = .20, U3 = 0.31. The electrodes sites were se-
lected based on a range of previous studies (par exemple., Klatt
et coll., 2018b; van Driel, Gunseli, Meeter, & Olivier, 2017;
van Ede, Niklaus, & Nobre, 2017; Myers, Walther, Wallis,
Stokes, & Nobre, 2015; Van der Lubbe, Bundt, & Abrahamse,
2014; Gould, Rushworth, & Nobre, 2011; Thut et al., 2006),
revealing a parieto-occipital scalp distribution and show-
ing PO7/8 to be a representative choice of electrodes
when measuring alpha lateralization. To minimize the
family wise error rate, we chose to limit the analysis to
one pair of electrodes. The ALI is positive when alpha
power is higher over the ipsilateral hemisphere (relative
to the target sound) and/or lower over the contralateral
hemisphere. In contrast, negative values indicate higher
alpha power contralateral to the target and/or lower alpha
power over ipsilateral electrode sites. The lateralization in-
dex was calculated using the raw, baseline-uncorrected
power values. ALI values for younger and older adults
were submitted to parametric two-sample t tests, en utilisant
Satterthwaite’s approximation to assess degrees of free-
dom. Subsequently, one-sample t tests were conducted
to test for significance of alpha lateralization within or
across age groups.
Multiple Regression
To investigate to what extent alpha lateralization and
N2ac amplitudes predict behavior in the given auditory
localization task, we applied regression analyses. Sep-
arate multiple linear regression models were evaluated
for mean RT, drift rate v, threshold separation a, et
non-decision time t0 as response variables, using the fitlm
Klatt et al.
949
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
n
t
p
o
:
un
/
d
/
e
d
m
je
F
r
t
o
p
m
r
c
h
.
p
s
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
2
c
je
5
e
9
–
4
p
5
d
F
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
un
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
t
.
/
F
o
n
0
5
M.
un
oui
2
0
2
1
ð
function implemented in the MATLAB Statistics and
Machine Learning toolbox (R2018a). To account for the
fact that accuracy proportions range inbetween 0 et 1,
a beta regression was calculated for accuracy as a re-
sponse variable using the R betareg package by Cribari-
Neto and Zeileis (2010). For all five regression analyses,
N2ac amplitudes, ALI, and age group served as pre-
dictors. En outre, to assess whether the relationship
between electrophysiological correlates and behavioral
outcomes differed between age groups, two interaction
terms were also included (c'est à dire., âge:N2ac, âge:ALI). Effects
coding was used as a contrast scheme for the age group
variable to enable a proper interpretation of lower and
higher order effects. Model assumptions were verified by
examination of residuals plots: Pearson residuals were
plotted against fitted values and against predictor variables
to assess nonconstant error variance (heteroscedasticity)
and deviations from linearity, respectivement. En outre,
normal probability plots were examined to evaluate nor-
mality of residuals. In case of a nonsignificant Durbin–
Watson test, returning a test statistic close to 2, residuals
were assumed to be uncorrelated. Variation inflation fac-
tors were inspected for signs of multicollinearity. Enfin,
to check for influential cases, leverage and cook’s distance
were examined. Values exceeding 1 for cook’s distance
(Cook & Weisberg, 1982) ou 3 × kþ1
Þ
(with k indicating
n
the number of predictors and n indicating the sample
size) for leverage (Pituch & Stevens, 2016) were set as cut-
offs for further inspection. The inspection of residuals
plots indicated deviations from normality for the drift rate
regression model. Refitting the model with a log trans-
formation (to base 10) of the drift rate values (v + 1; un
constant was added to avoid negative values) resulted in
approximately normally distributed residuals. Ainsi, ordi-
nary least square regression was applied. For the models
regarding threshold separation, non-decision time, et
RT, the residual probability plots indicated some outliers.
To reduce outlier effects, we fitted a robust regression
model, using an iterative reweighted least squares proce-
dure and a bisquare weight function. Adjusted R-squared
(denoted as R2) is reported as a goodness-of-fit statistic. À
correct for the fact that we conducted separate multiple
regression analyses for each of the five dependent vari-
ables, p values for regression coefficients were corrected
using a Bonferoni–Holm procedure (Holm, 1979). Note
that in each case the five p values belonging to the same
type of estimate (c'est à dire., intercept, N2ac fixed effect, ALI fixed
effet, age fixed effect, N2ac:age interaction term, or ALI:
age interaction term) were corrected for multiple testing.
To visualize the relationship between single predictors and
résultats, marginal effects plots (ggeffect function from
ggeffects package; Lüdecke, 2018) and adjusted response
les fonctions (plotInteraction and plotAdjustedResponse
les fonctions) were used for the beta regression model (in R)
and linear regression models (in MATLAB), respectivement.
Adjusted response functions describe the relationship be-
tween the fitted response and a specific predictor, alors que
the other predictors are averaged out by averaging the fitted
values over the data used in the fit. Adjusted response values
are computed by adding the residual to the adjusted fitted
value for each observation (The MathWorks, 2019). Quand
plotting marginal effects using “ggeffect,” the other factors are
held constant at an average value (Lüdecke, 2018).
Statistical Tests and Effect Sizes
Data were considered normally distributed if the Lilliefors
test (Lilliefors, 1967) yielded insignificant results ( p >
.05). For normally distributed data, parametric two-sample
Welch’s t tests were applied. Degrees of freedom were
estimated using Satterthwaite’s approximation, assuming
unequal variances. Wilcoxon rank-sum test served as the
nonparametric counterpart in case of nonnormality. À
test for significance within an age group, a parametric
one-sample t test or the nonparametric Wilcoxon signed-
rank test was applied. Measures of effect sizes were calcu-
lated using the MES toolbox provided by Hentschke and
Stüttgen (2011). For parametric one- and two-sample t
tests, g1 and Hedge’s g (in the following referred to as g)
are reported, respectivement. For both measures, effect sizes
of ±0.2 are typically referred to as small, values of ±0.5 as
moyen, and values of ±0.8 as large. For nonparametric t
tests, Cohen’s U3 is reported. Cohen’s U3 is a measure of
overlap of two distributions, avec 0.5 indicating minimal
overlap and 0 ou 1 indicating maximal overlap. The sig-
nificance of effects was assessed at a significance level of
α = .05. The Bonferroni–Holm correction procedure
was applied to correct for multiple comparisons when
appropriate (Holm, 1979). Adjusted p values are denoted
as padj.
Given that p values from standard inferential statistics
do not allow any conclusions on whether or not the null
hypothesis is true, we additionally report the Bayes factor
(BF) to strengthen the interpretability of effects in this
étude. En substance, the BF provides a “continuous” mea-
sure, which indicates how much more likely the ob-
served results are under a given hypothesis, compared
with an alternative hypothesis (for an introduction to
Bayesian statistics, see Quintana & Williams, 2018;
Wagenmakers et al., 2018). A BF of 1 indicates that the
results are equally likely under both hypotheses (c'est à dire.,
the null and the alternative hypothesis). A BF < 1 pro-
vides increasing evidence in favor of the null hypothesis
relative to the alternative hypothesis, whereas a BF > 1
provides increasing evidence favoring the alternative
hypothesis over the null hypothesis (Dienes, 2014). À
facilitate the interpretation of BFs, the classification
scheme originally proposed by Jeffreys (1961) is applied:
The latter suggests that a BF > 3 and > 10 provide mod-
erate and strong evidence for the alternative hypothesis,
respectivement, whereas a BF < 0.33 or < 0.1 suggests mod-
erate and strong evidence in favor of the null hypothesis,
950
Journal of Cognitive Neuroscience
Volume 32, Number 5
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
5
e
9
-
4
p
5
d
f
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
a
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
/
.
f
o
n
0
5
M
a
y
2
0
2
1
respectively. Finally, BFs between 0.33 and 3 are inter-
preted in terms of anecdotal evidence. However, it should
be noted that those cutoffs have no absolute meaning
(Dienes, 2014) in that evidence is continuous and it is di-
rectly interpretable in terms of an odds ratio (Quintana &
Williams, 2018). The notation BF10 indicates the Bayes
factor for the alternative hypothesis (i.e., that the means
of the samples are different). BF functions implemented
in MATLAB by Krekelberg (2019) and the BayesFactor
package implemented in R (function: linearReg.R2stat) by
Morey and Rouder (2018) were used to calculate BFs for
t tests and regression, respectively. To obtain a BF for a
specific coefficient in our regression model (BFcoef), the
BF for the full model and the restricted model were com-
pared according to the following formula: BFfull/BFrestr.
BFfull indicates the BF for the full model, including all pre-
dictors, whereas BFrestr indicates the BF for the restricted
model, omitting the coefficient of interest. Default priors,
that is, the Jeffrey–Zellner–Siow Prior for t tests and a
mixture of g-priors according to Liang, Paulo, Molina,
Clyde, and Berger (2008) for regression, were applied.
Because those packages do not support the calcula-
tion of BFs for beta regression, no Bayesian statistics
are provided for the regression analysis of accuracy
data.
RESULTS
Behavioral Results
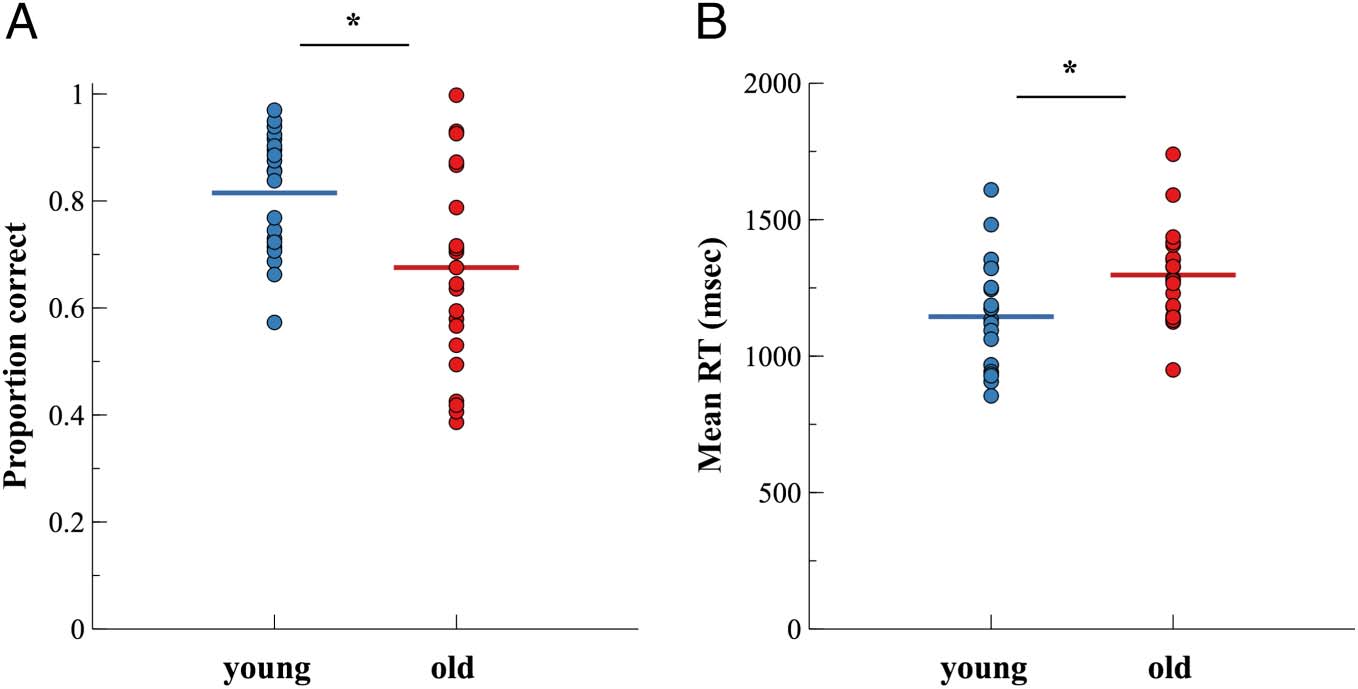
Figure 1 shows the proportion of correct responses
(Figure 1A) as well as mean RTs (Figure 1B) separately
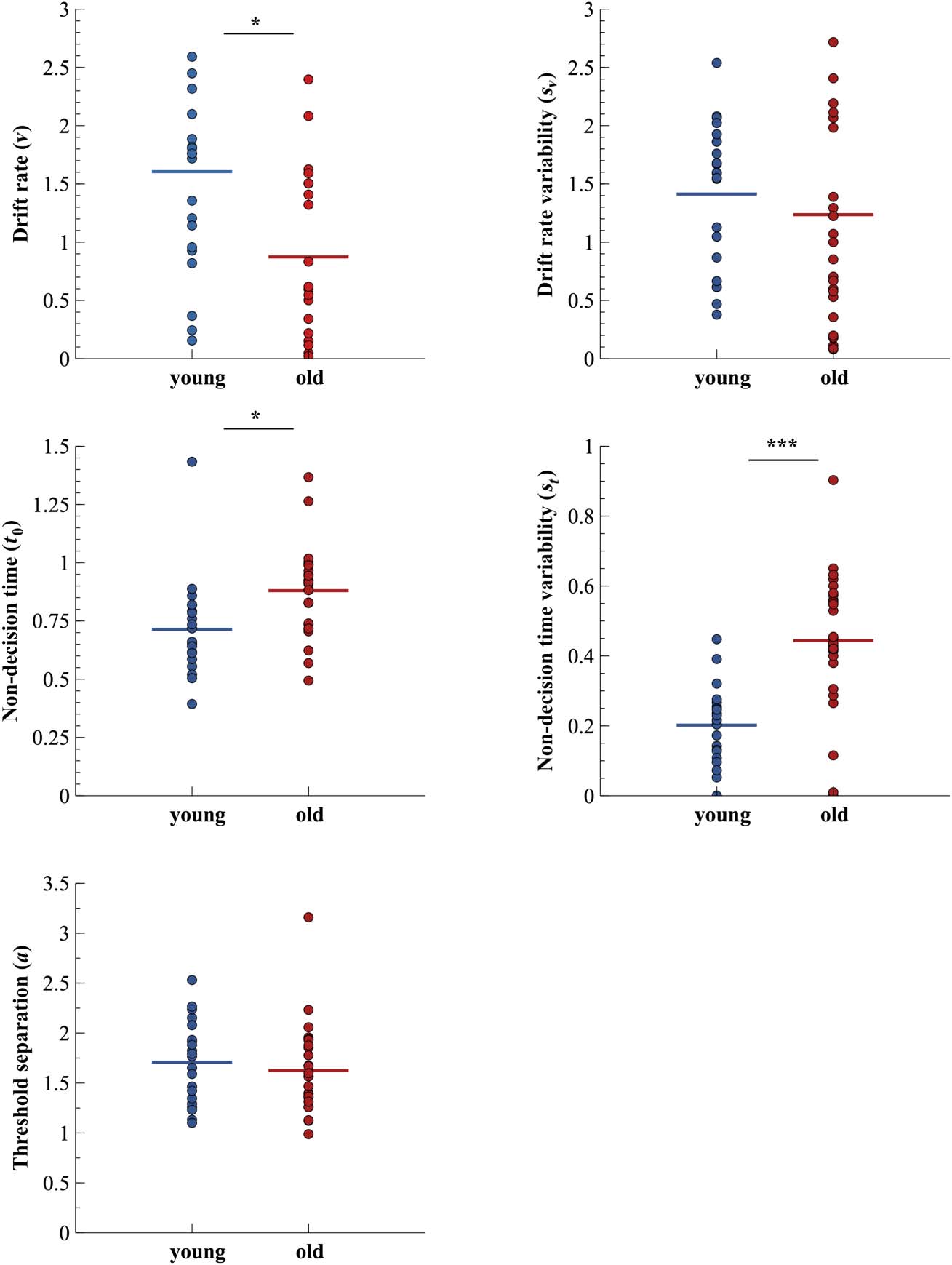
for both age groups. Diffusion parameters are depicted
in Figure 2. On average, younger adults showed higher
accuracy (t(43.05) = −3.36, p = .002, padj = .01, g =
0.92, BF10 = 14.21) and faster responses than older
adults (t(38.56) = 2.80, p = .008, padj = .038, g =
−0.83, BF10 = 6.93). The BFs indicated that the alterna-
tive model was around 14 times and six times more likely
than the null model, respectively, thus providing strong
and moderate support for a difference between age
groups.
Although mean RTs do not offer any insights into the
underlying causes of prolonged RTs, diffusion parame-
ters allow for a closer look at different possible explana-
tions for the observed difference between age groups,
including a slowdown of information update (i.e., higher
drift rate v), a more conservative response criterion (i.e.,
higher threshold separation a), or delayed response exe-
cution (i.e., higher response constant t0). In our sample,
older adults showed a significantly reduced drift rate
(t(44.89) = −2.51, p = .016, padj = .047, g = 0.70,
BF10 = 3.01), higher non-decision time (t(41.31) =
2.81, p = .008, padj = .038, g = −0.82, BF10 =
6.59), as well as higher variability of non-decision time
(t(40.26) = 5.25, p < .001, padj < .001, g = −1.43, BF10 =
153.9). Threshold separation values (t(44.24) = −0.66,
p = .513, padj = .513, g = 0.19, BF10 = 0.35) and trial-
to-trial variability of drift rate (Z = −1.21, p = .226, padj =
.453, U3 = 0.29, BF10 = 0.35) did not differ significantly
between age groups. Although the BFs supported clas-
sical inferential statistics for significant results (BFs > 3),
for insignificant results they fell short of the criterion for
moderate evidence for equivalence (BFs > 0.33). To graph-
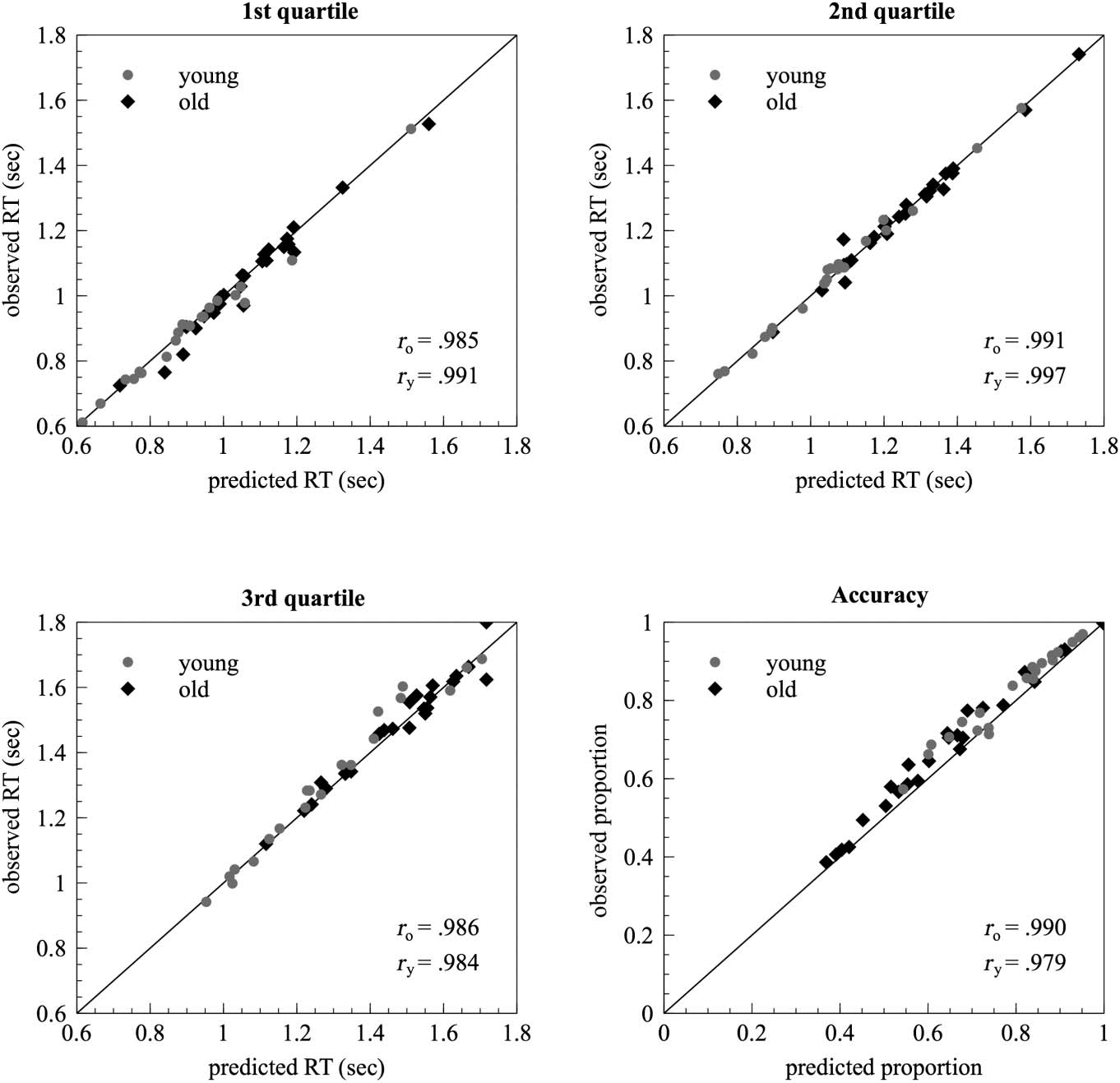
ically assess the fit of the estimated diffusion models, ob-
served RT quartiles (.25, .5, .75) and observed accuracy
were plotted against the corresponding value of the pre-
dicted distributions. As can be seen in Figure 3, the major-
ity of data points lie close to the line of perfect correlation,
indicating adequate model fit.
N2 Anterior Contralateral Component
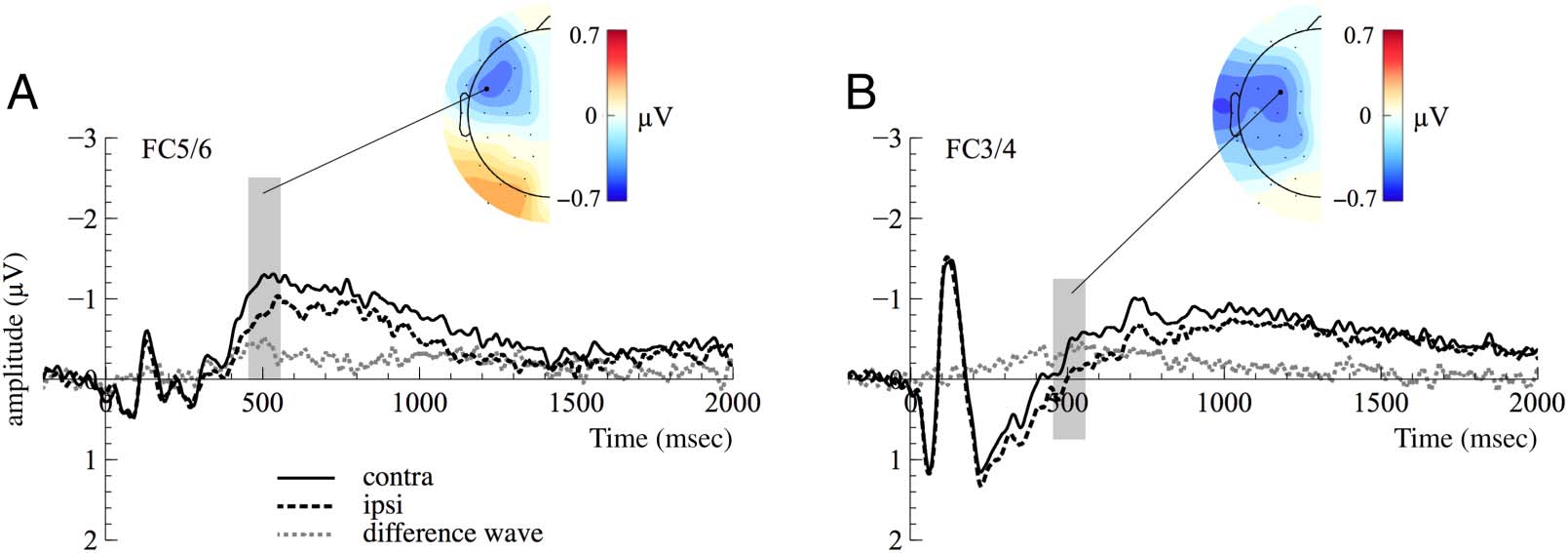
Chiffre 4 presents the ERPs at frontocentral electrodes
FC3/4 for older adults and electrodes FC5/6 for younger
adultes. En outre, the corresponding topographies
based on the contralateral minus ipsilateral difference
wave in the analysis time window are depicted. N2ac am-
plitudes (c'est à dire., contralateral minus ipsilateral differences)
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
n
t
p
o
:
un
/
d
/
e
d
m
je
F
r
t
o
p
m
r
c
h
.
p
s
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
2
c
je
5
e
9
–
4
p
5
d
F
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
un
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
t
/
.
F
Chiffre 1. Proportion of (UN)
correct responses and (B) mean
RTs for younger and older
adultes. Colored horizontal lines
indicate the respective group
mean. Dots indicate individual
participants’ mean values.
*padj < .05.
o
n
0
5
M
a
y
2
0
2
1
Klatt et al.
951
Figure 2. Diffusion model
parameter estimates for
younger and older participants.
Dots represent single subject
data. Colored horizontal lines
show the mean model
parameters within age groups.
*padj < .05, ***padj < .001.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
5
e
9
-
4
p
5
d
f
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
a
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
/
f
.
o
n
0
5
M
a
y
2
0
2
1
did not differ significantly between younger (M = −0.37,
SD = 0.47) and older adults (M = −0.38, SD = 0.40, t
(39.37) = −0.09, p = .926, g = 0.03, BF10 = 0.29).
The BF of 0.29 can be interpreted as insufficient evi-
dence, supporting neither the null nor the alternative hy-
pothesis. A one-sample t test confirmed that across both
age groups, N2ac amplitudes were significantly different
from zero (t(46) = −6.02, p < .001, padj < .001, g1 =
−0.88, BF10 > 1000). Cependant, it should be noted that
the original analysis time window was based on the 50%
FAL in the grand-averaged difference waveform across
both age groups; thus, this procedure favors a significant
result when testing overall N2ac amplitudes against zero.
To avoid this problem of “double dipping,” we performed
a second one-sample t test, using a broader analysis time
window of 400–600 msec post sound array onset. The lat-
ter yielded comparable results (t(46) = −4.41, p < .001,
padj < .001, g1 = −0.64, BF10 > 1000). Consistently, le
BF provided strong evidence in favor of the presence of
an N2ac component across both age groups.
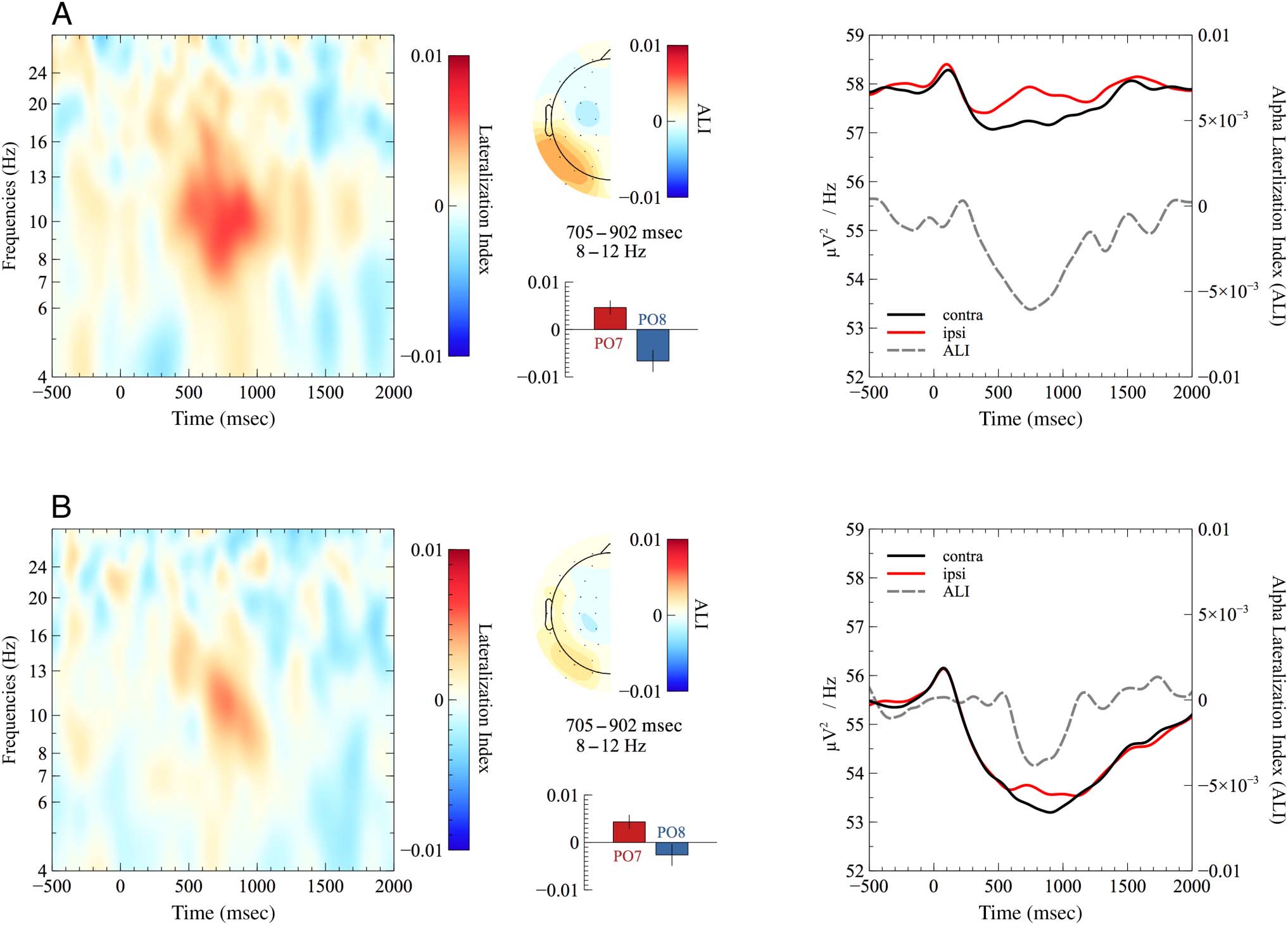
Alpha Lateralization
The time–frequency plots in Figure 5 illustrate the asym-
metric modulation of alpha power (8–12 Hz) at electrodes
PO7/8 time-locked to sound array onset for younger
(Figure 5A) and older adults (Figure 5B), respectivement. Dans
addition, the corresponding topographies based on the
normalized ipsilateral minus contralateral difference in al-
pha power are depicted. Although younger adults appeared
to show larger alpha power lateralization than older adults,
the analysis revealed no significant difference in alpha
power lateralization between age groups (t(41.23) =
−1.43, p = .161, g = 0.42, BF10 = 1.13). The BF suggested
that the data were insensitive to distinguish the null (Non
amplitude difference between groups) from the alternative
952
Journal des neurosciences cognitives
Volume 32, Nombre 5
Chiffre 3. Graphical analysis of
model fit. Scatter plots show the
observed proportion of correct
responses as well as the first
three quartiles (.25, .5, .75) de
the observed RT distribution as
a function of the corresponding
value from the predicted
distribution. Dots and
diamonds represent single
subject data for younger and
older participants. r denotes the
corresponding Pearson
correlation coefficients,
separately for younger (ry) et
older (ro) adultes.
hypothèse (difference in amplitudes between age groups).
Encore, a one-sample t test confirmed that alpha lateralization
across both age groups was significantly different from zero
(t(46) = 6.07, p < .001, padj < .001, g1 = 0.89, BF10 >
1000), and the BF consistently suggested strong evidence
for the alternative hypothesis. As mentioned above (cf.
N2 Anterior Contralateral Component section), the analysis
time window (determined based on the 50% FAL in the
grand-averaged waveform) favors a significant result when
testing across age groups, against zero. Ainsi, a second
one-sample t test was performed, based on a broader anal-
ysis time window of 600–900 msec post sound array onset,
yielding comparable results (t(46) = 5.91, p < .001, padj <
.001, g1 = 0.86, BF10 > 1000).
Regression Analyses
We examined the relationship between mean alpha
power lateralization, N2ac amplitudes, and behavioral
performance (including diffusion model parameters)
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
n
t
p
o
:
un
/
d
/
e
d
m
je
F
r
t
o
p
m
r
c
h
.
p
s
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
2
c
je
5
e
9
–
4
p
5
d
F
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
un
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
F
/
t
.
o
n
0
5
M.
un
oui
2
0
2
1
Chiffre 4. N2ac component at frontocentral electrodes FC5/6 for (UN) younger and at FC3/4 for (B) older participants. Contralateral and ipsilateral
portions of the signal as well as the resulting difference wave (contralateral minus ipsilateral) are depicted. Scalp topographies show the
distribution of voltage differences based on the contralateral minus ipsilateral difference wave in the time window used for statistical analysis
(highlighted in gray in ERP figures).
Klatt et al.
953
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
n
t
p
o
:
un
/
d
/
e
d
m
je
F
r
t
o
p
m
r
c
h
.
p
s
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
2
c
je
5
e
9
–
4
p
5
d
F
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
un
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
F
.
t
/
o
n
0
5
M.
un
oui
2
0
2
1
Chiffre 5. Grand-average time–frequency plots of lateralization indices at electrodes PO7/8 for (UN) younger and (B) older adults. The scalp
topographies are based on normalized differences of ipsilateral minus contralateral alpha power in the time window used for statistical analysis.
Bar graphs show the mean difference (left minus right) for the left (c'est à dire., PO7) and right (c'est à dire., PO8) hemisphere. Error bars indicate the SEM. Doubler
plots (droite) illustrate the contralateral and ipsilateral portion of the raw ERSPs as well as the resulting ALI.
using multiple linear regression. The estimated param-
eters are provided in Table 1. Participants with greater
N2ac amplitudes showed higher accuracy (Z = −3.93,
p < .001, padj < .001) and higher drift rate (t(41) =
−2.79, p = .008, padj = .032, BFcoef = 7.75), whereas
there was no significant effect of alpha lateralization on
those performance outcomes (accuracy: Z = −1.54, p =
.124, padj = .499; drift rate: t(41) = −0.37, p = .712, padj =
1.067, BFcoef = 0.43). For both models, there was no
significant interaction with age (all padj > .160). The cor-
responding BFs (only available for the drift rate model; cf.
Statistical Tests and Effect Sizes section) were below 3
(BFcoef ≤ 0.65) but above 0.33, thus lending insufficient
evidence for the null or the alternative hypotheses. Le
full models, including all predictors, explained 26% et
36% of variance in drift rate (R2
adj = .26, F(5, 41) =
4.15, p = .004) and accuracy (pseudo-R2 = .36, precision
parameter phi = 9.73, SE = 1.96, z = 4.97, Pr(>|z|) <
.001), respectively. For all other models tested, nei-
ther N2ac amplitudes nor alpha power lateralization or
their interaction with age groups served as statistically
significant predictors (all padj > .095; cf. Tableau 1). For all
but one parameter, the corresponding BFs were in-
conclusive (3 < BFcoef > 0.33), providing no substantial
support for the alternative hypothesis, but neither for
the null hypothesis. Cependant, for the regression model
predicting non-decision time, the BF for the interaction
term N2ac*Age ( p = .095) lend moderate evidence in fa-
vor of the alternative hypothesis (BFcoef = 5.92), suggérer-
ing that, in older adults, less pronounced N2ac amplitudes
were associated with higher non-decision times. In con-
trast, the latter relationship appeared absent in younger
adultes. Age group, not surprisingly, significantly predicted
non-decision time (t(41) = 3.00, p = .005, padj = .018,
BFcoef = 15.27), accuracy (Z = 3.03, p = .002, padj =
.012), and drift rate (t(41) = −2.86, p = .007, padj =
.020, BFcoef = 8.78). Although age group failed to serve
as a significant predictor for RT in the regression model
framework (t(41) = 1.82, p = .075, padj = .151, BFcoef =
2.38), the results largely confirm the behavioral age dif-
ferences reported in the Behavioral Results section. The BF
de 2.38 suggests that the data may simply be underpowered
954
Journal des neurosciences cognitives
Volume 32, Nombre 5
Tableau 1. Estimated Parameters, Standard Errors, Confidence Intervals, and t Test (or z Test) Statistics for Each Predictor in the Linear (or Beta) Regression Model
Résultat
v
un
t0
Accuracy
RT
Prédicteurs
b (SE) [95% CI]
t
b (SE) [95% CI]
t
b (SE) [95% CI]
t
b (SE)
z
b (SE) [95% CI]
t
Intercept
0.22*** (0.05)
[0.12 0.31]
−0.22* (0.08)
[−0.38 −0.06]
−2.52 (6.77)
[−16.20 11.16]
−0.14* (0.05)
[−0.24 −0.04]
−0.16 (0.08)
[−0.32 0.00]
−4.67 (6.78)
[−18.35 9.02]
N2ac
ALI
Age
N2ac*Age
ALI*Age
Adjusted/
pseudo-R2
4.43, p < .001
padj < .001
−2.79, p = .008
padj = .032
1.55*** (0.09)
[1.36 1.74]
16.47, p < .001
padj < .001
0.77*** (0.04)
[0.70 0.86]
19.77, p < .001
padj < .001
0.89***
(0.16)
5.63, p < .001
padj < .001
1.24*** (0.04)
[1.15 1.33]
28.95, p < .001
padj < .001
−0.32 (0.15)
[−0.62 −0.01]
−2.10, p = .042
padj = .126
0.08 (0.06)
[−0.05 0.21]
1.29, p = 0.203
padj = .407
−1.02***
(0.26)
−3.93, p < .001
padj < .001
−0.37, p = .712
padj = 1.067
−10.87 (13.05)
[−37.22 15.49]
−0.83, p = .410
padj = 1.230
10.18 (5.45)
[−0.83 21.19]
1.87, p = .069
padj = .345
−33.48
(21.81)
−1.54, p = .124
padj = .499
0.08 (0.07)
[−0.06 0.22]
3.73 (5.94)
[−8.27 15.74]
−2.86, p = .007
padj = .020
−2.02, p = .050
padj = .160
−0.69, p = .495
padj = .965
−0.07 (0.09)
[−0.26 0.12]
−0.11 (0.15)
[−0.41 0.20]
−0.76, p = .452
padj = .452
−0.72, p = .474
padj = .474
0.12* (0.04)
[0.03 0.20]
0.15 (0.06)
[0.03 0.28]
3.00, p = .005
padj = .018
2.44, p = .018
padj = .095
−0.48*
(0.16)
−0.53
(0.26)
3.03, p = .002
padj = .012
0.08 (0.04)
[−0.01 0.16]
−2.05, p = .039
padj = .160
0.09 (0.04)
[−0.04 0.24]
−10.06 (13.05)
[−36.41 16.30]
−0.77, p = .445
padj = 1.336
11.23 (5.45)
[0.22 22.24]
2.06, p = .046
padj = .229
−15.30
(21.78)
−0.70, p = .483
padj = 1.336
9.19 (5.94)
[−2.81 21.19]
1.14, p = .261
padj = .407
0.63, p = .533
padj = 1.230
1.82, p = .075
padj = .151
1.43, p = .160
padj = .319
1.55, p = .130
padj = .519
.26
.03
.32
.36
−
.14
F(5, 41) = 2.46, p = .048
F-statistic
F(5, 41) = 4.15, p = .004
F(5, 41) = 1.25, p = .302
F(5, 41) = 5.29, p = .001
v, a, t0, and RT denote drift rate, threshold separation, non-decision time, and mean RTs, respectively. SE = standard error, CI = confidence interval. Adjusted R2 is given for linear regression models (v, a,
t0, and RT); pseudo-R2 is given for beta-regression (accuracy). p denotes uncorrected p values; padj denotes p values corrected for multiple comparison using a Bonferroni–Holm correction procedure
(Holm, 1979). Asterisks denote significant estimates with adjusted p values as *padj < .05, ***padj < .001.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
5
e
9
-
4
p
5
d
f
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
a
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
l
K
a
t
t
e
t
a
l
.
9
5
5
o
n
0
5
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
5
e
9
-
4
p
5
d
f
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
a
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
0
5
M
a
y
2
0
2
1
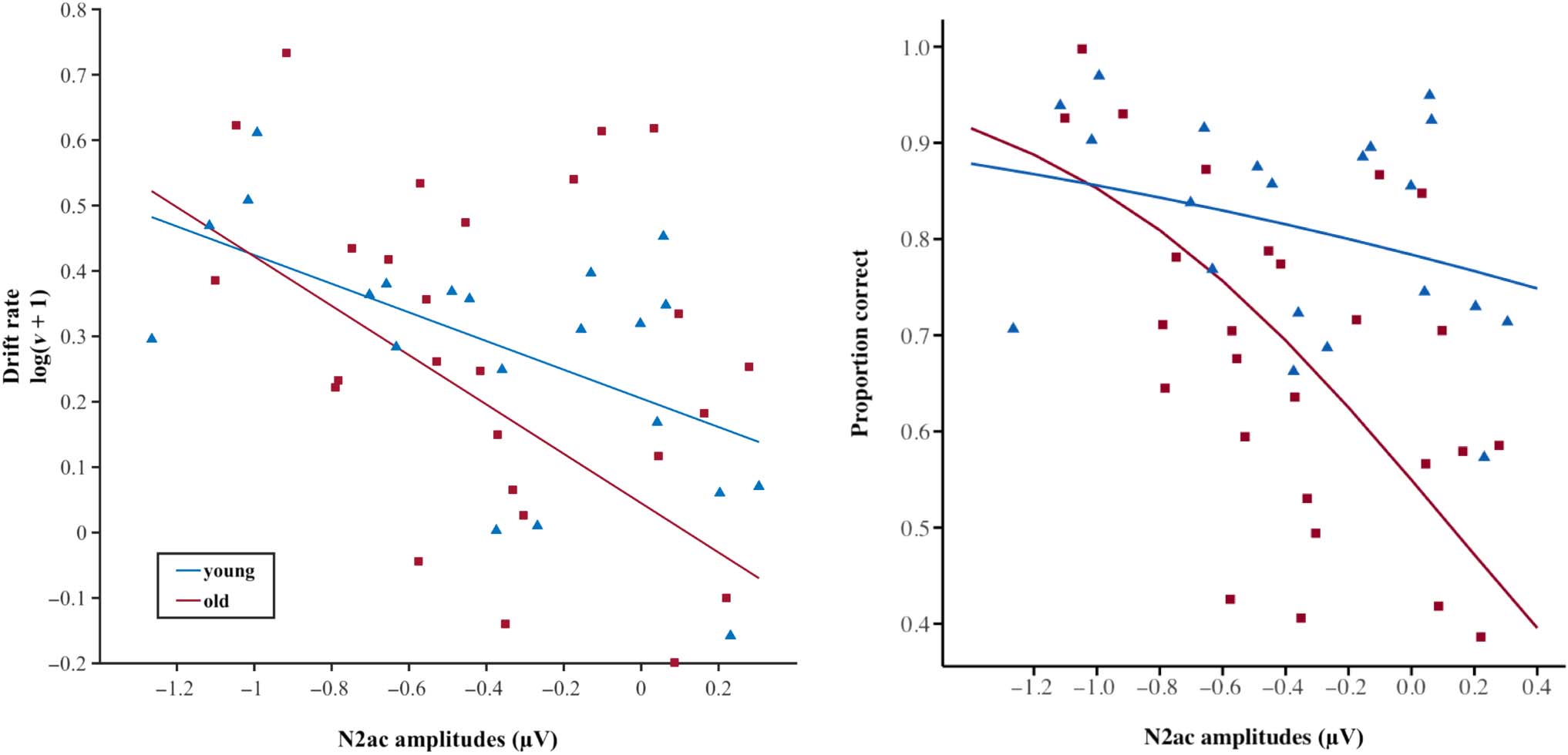
Figure 6. Participants’ drift rate and mean accuracy as a function of mean N2ac amplitude. Triangles represent younger participants (n = 21);
squares represent older participants (n = 26). For the linear drift rate regression model, an adjusted response function describes the relationship
between the fitted response and N2ac amplitudes, whereas the other predictors are averaged out by averaging the fitted values over the data used in
the fit. Adjusted response data points are computed by adding the residual to the adjusted fitted value for each observation. For the accuracy
beta regression model, the marginal effect of the interaction N2ac amplitude by age group is displayed, holding the other factors constant at an
average value.
to reveal a relation between RT and age group in the present
regression model. Figure 6 visualizes the reported results for
those outcomes that were significantly predicted by N2ac
amplitudes.
that the data is inconclusive with respect to age effects
in the electrophysiological data. Age differences in behav-
ioral performance are briefly reviewed below.
DISCUSSION
In this study, we investigated the contribution of post-
stimulus alpha power lateralization and N2ac amplitudes
to sound localization performance in a sample of younger
and older adults. Both measures have been associated
with the deployment of attention in auditory space. We
hypothesized that if the cortical processes reflected by
alpha lateralization and N2ac amplitudes contribute to
successful target selection, their magnitudes should be
related to the information accumulation process (i.e.,
drift rate; cf. diffusion model framework, as outlined in
the Introduction) and in turn to localization accuracy
and RTs. In fact, what we found only partially confirmed
this hypothesis: N2ac amplitudes significantly predicted
both drift rate and accuracy, whereas alpha lateralization
was not associated with any of the behavioral outcomes.
We thus proposed that N2ac and alpha lateralization re-
flect distinct aspects of attentional orienting in auditory
scenes. Classical frequentist inferential statistics suggested
that the observed relationship did not depend on age and
that both age groups showed comparable neural signa-
tures. However, Bayesian alternatives to classical hypothe-
ses testing raised doubts about these claims, suggesting
Cocktail Party Sound Localization in Older and
Younger Adults
As expected, older adults showed fewer correct responses
and slower RTs than younger adults. This is in line with the
often-described difficulties of older people to follow a con-
versation in noisy (“cocktail party”) environments, which
depends on the integrity of both sensory and cognitive
functions (Shinn-Cunningham, 2017). Declined perfor-
mance in older adults in the present task is likely to be
related to age-related deficits in concurrent sound segre-
gation (Hanenberg, Getzmann, & Lewald, 2019; Alain &
McDonald, 2007; Snyder & Alain, 2005). Traditionally,
such deficits have been interpreted as a result of a gen-
eral sensory-cognitive decline (e.g., Pichora-Fuller, Alain,
& Schneider, 2017), assuming all aspects of processing in
an experimental task to be globally slowed in aging adults
(Myerson, Hale, Wagstaff, Poon, & Smith, 1990). The diffu-
sion model allows to differentiate between different aspects
of processing that might be affected by age (Ratcliff, Spieler,
& McKoon, 2000): Consistent with previous results (Ratcliff,
Thapar, & McKoon, 2003, 2011; Ratcliff et al., 2001), we
found an increase in non-decision time for older adults. In
addition, older participants varied more strongly in their
non-decision time from trial to trial, indicating that this
956
Journal of Cognitive Neuroscience
Volume 32, Number 5
process was noisier in older adults (Spaniol, Madden, &
Voss, 2006). However, rather untypically, the two age
groups did not differ in their threshold separation values.
This contradicts the wide-spread assumption that older
adults usually aim to minimize errors (leading to more con-
servative response criteria) whereas younger adults focus on
balancing speed and accuracy (Starns & Ratcliff, 2010). The
observed lack of differences in response criteria between
older and younger adults could be due to the relatively
long response period in this study, potentially inducing
a change in task goals in younger adults. Alternatively,
as the corresponding BFs were rather inconclusive, we
cannot exclude that the data are simply underpowered
and therefore fail to reveal significant differences in our
sample. Furthermore, supporting a line of evidence that
showed differences in the rate of information accumulation
in some contexts (Ratcliff et al., 2004, 2011; Spaniol et al.,
2006), older adults had significantly decreased drift rates.
Given the current state of research, the conditions under
which drift rate decreases with age are still hard to grasp.
Here, drift rate was significantly predicted by N2ac ampli-
tudes. In participants with higher N2ac amplitudes (i.e.,
more negative difference waves) drift rates were higher,
whereas participants with lower N2ac amplitudes tended
to have lower drift rates. Hence, differences in drift rate
may reflect the differences in the ability to extract rele-
vant information from a perceptual scene (in this case,
an array of concurrently presented sounds). In the fol-
lowing section, we will discuss this relationship in more
detail.
N2ac Amplitudes Predict Drift Rate and Accuracy
To date, little is known about the functional relevance of
the N2ac component. The regression analysis conducted
here revealed that N2ac amplitudes significantly pre-
dicted variations in accuracy as well as drift rate, while
they were unrelated to mean RTs, threshold separation,
or non-decision time. These findings add to the sparse
literature that has so far investigated the N2ac com-
ponent in different contexts (Klatt et al., 2018b; Lewald
et al., 2016; Gamble & Woldorff, 2015a, 2015b; Lewald &
Getzmann, 2015; Gamble & Luck, 2011). In addition, to
our best knowledge, this is the first study to show an
N2ac component in a sample of older adults. Gamble
and Luck (2011) originally proposed that the N2ac arises
to resolve the competition between simultaneously pres-
ent stimuli and reflects the attentional orienting toward a
target. They further elucidated that this may be based on
the biasing of neural coding toward the attended stimulus,
as observed in the visual modality. In fact, the observed
relationship of N2ac amplitudes and drift rate may support
this line of reasoning: Drift rate conceptually reflects the
quality of relevant information derived from sensory input
that eventually drives the decision process (Ratcliff et al.,
2000). Hence, the better participants may be able to re-
solve competition between concurrent sounds by focusing
on the target (i.e., N2ac amplitude), the better the quality
of information that prompts participants to make a deci-
sion (i.e., drift rate; or in other words, the higher the rate
of evidence accumulation in favor of a given response). In
turn, it logically follows that the better or more consis-
tently participants are able to focus their attention onto
a relevant target sound (i.e., N2ac amplitude), the higher
their overall accuracy.
Interestingly, in addition to the similar N2ac ampli-
tudes for younger and older adults, we found no signifi-
cant interactions between N2ac amplitudes and age,
neither for accuracy nor for drift rate (cf. Table 1). This
may suggest that the variances within age groups contrib-
ute more strongly to the observed relationship than the
variance between age groups. However, the difficulties of
interpreting a null effect, such as a missing interaction
with age, need to be considered as a caveat here. Al-
though regression lines in Figure 6 show a trend toward
an interaction of N2ac amplitude and age group, the cal-
culated BFs (cf. Regression Analyses section) suggest the
data to be insensitive to age group differences, providing
no substantial evidence in favor of the null or alternative
hypothesis. Nevertheless, one may raise the question, if
lower N2ac amplitudes result in lower drift rates and
decreased performance, why did older adults not show
reduced N2ac amplitudes, given that they performed sig-
nificantly worse than the younger adults? On the one
hand, the well-pronounced N2ac component in older
adults may, at least in part, have resulted from the re-
cruitment of additional top–down resources to allow for
more efficient target selection. This interpretation would
be in line with the decline-compensation hypothesis
(Cabeza, Anderson, Locantore, & McIntosh, 2002; for a re-
view, see Schneider, Pichora-Fuller, & Daneman, 2010),
proposing that age-related declines in peripheral and
central auditory processing are compensated for by in-
creased allocation of cognitive resources. Increases in at-
tentional focusing, however, might not be sufficient to
completely compensate for the reduced performance of
the older group. On the other hand, we cannot exclude
that we simply failed to find a significant difference in
N2ac amplitudes due to a lack of power, as the calcula-
tion of BFs provided no substantial evidence in favor of
the null hypothesis.
Is Poststimulus Alpha Power Lateralization
Functionally Relevant?
This study also investigated alpha lateralization as a mea-
sure of attentional orienting within an auditory scene.
Typically, alpha lateralization manifests in a bilateral de-
crease of alpha power, which is more pronounced over
the contralateral hemisphere (relative to a target or a
cue). This spatially specific modulation of oscillatory activ-
ity has been repeatedly associated with the top–down con-
trolled voluntary allocation of attention (Ikkai et al., 2016;
Haegens et al., 2011; Thut et al., 2006; Foxe, Simpson,
Klatt et al.
957
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
5
e
9
-
4
p
5
d
f
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
a
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
t
.
o
n
0
5
M
a
y
2
0
2
1
& Ahlfors, 1998). Here, we replicated this consistently
observed response in the alpha frequency band in a sam-
ple of younger and older participants who performed an
auditory localization task, requiring them to indicate the
location of a predefined target stimulus among three con-
cordantly presented distractors. Our results suggested that
older adults may, in principle, be able to recruit the same
oscillatory mechanisms as younger adults when searching
for a target among simultaneously present distractors
(Klatt et al., 2018b). Although Bayesian statistics were in-
decisive in whether the nonsignificant difference in alpha
lateralization between age groups presents a true null
effect, the preserved poststimulus alpha lateralization cor-
roborated a number of studies, showing intact alpha later-
alization in older adults when anticipating an upcoming
(lateralized) stimulus (Heideman et al., 2018; Leenders,
Lozano-Soldevilla, Roberts, Jensen, & De Weerd, 2018;
Tune et al., 2018). However, recent studies did not find
alpha lateralization in older adults, although they were still
able to perform the task as well as their younger counter-
parts (van der Waal, Farquhar, Fasotti, & Desain, 2017;
Hong et al., 2015). This poses the question to what extent
lateralized alpha dynamics are functionally relevant for
behavior.
It is relatively undisputed that alpha power lateraliza-
tion tracks the locus and timing of spatial attention
(Bae & Luck, 2018; Foster et al., 2017; Samaha, Iemi, &
Postle, 2017). In addition, a growing body of evidence
supports the notion that the alpha rhythm as a correlate
of spatial attention, so far predominantly investigated in
the visual attention literature, analogously operates in dif-
ferent modalities (Klatt et al., 2018a, 2018b; Wöstmann
et al., 2016, 2018; Thorpe, D’Zmura, & Srinivasan, 2012;
Haegens et al., 2011). Yet, what remains a matter of de-
bate is (1) how alpha power lateralization aids selective
spatial attention and (2) whether it reflects a necessary
prerequisite for successful behavioral performance.
Regarding the how, two prevailing views exist: The
gating by inhibition theory, proposed by Jensen and
Mazaheri (2010), suggested that the relative increase of
alpha power over the ipsilateral hemisphere inhibits re-
gions processing irrelevant information. Alternatively, it
has been suggested that the relative decrease of alpha
power over the contralateral hemisphere results in in-
creased cortical excitability, allowing for enhanced pro-
cessing of the targets (Noonan et al., 2016; Yamagishi
et al., 2005). Both mechanisms are not necessarily mutu-
ally exclusive. In fact, Foster and Awh (2019) just recently
pointed out that a lot of the empirical evidence is com-
patible with either the target enhancement or the distrac-
tor suppression account. Recent evidence suggested that
both mechanisms might independently contribute to at-
tentional orienting (Schneider et al., 2019). In line with
those latter findings, Capilla, Schoffelen, Paterson, Thut,
and Gross (2014) proposed distinct sources and behav-
ioral correlates for the ipsilateral and contralateral por-
tion of the alpha power signal.
Adressing the second question—Does alpha lateraliza-
tion reflect a necessary prerequisite for successful behav-
ioral performance?—a range of spatial-cueing studies has
provided compelling evidence showing behavioral per-
formance to be predicted by the degree of alpha lateral-
ization (Haegens et al., 2011; Kelly et al., 2009; Thut et al.,
2006). On the contrary, our findings question the notion
that alpha power lateralization reflects a behaviorally rel-
evant attentional mechanism: Surprisingly, we did not
find any association between alpha lateralization and dif-
fusion model parameters, mean RTs, or accuracy. This
could be explained by the fact that this study differed
from the majority of previous studies in that it investi-
gated alpha power modulations following stimulus presen-
tation. That is, although alpha lateralization may in fact be
necessary to successfully shift one’s attention in anticipa-
tion of an upcoming stimulus, it does not appear to be a
required neural response in the attentional processing
following the presentation of a multisound array. This is
in line with the proposal previously made by van Ede
et al. (2014), who similarly concluded that the relevance
of attentional modulations might be “restricted to situa-
tions in which attention influences perception through an-
ticipatory processes” (p. 139). However, in contrast to our
results, these authors found that alpha lateralization was
completely abolished during the processing of an ongoing
tactile stimulus.
Alternatively, the lack of a relationship with behavioral
performance may be due to the fact that we calculated a
relative measure of alpha amplitudes, that is, the differ-
ence between ipsilateral and contralateral alpha power.
In a cued somatosensory detection task, van Ede et al.
(2014) found only contralateral alpha power amplitudes
to be related to tactile detection performance, whereas
fluctuations in the contralateral minus ipsilateral differ-
ence failed to predict performance. Similarly, other stud-
ies using a relative index of alpha power modulations did
not find a strong relationship with behavioral performance
(Tune et al., 2018; Limbach & Corballis, 2017). These find-
ings or rather null findings might strengthen the emerging
view that both target enhancement (i.e., contralateral
alpha power decrease) and distractor suppression (i.e.,
ipsilateral alpha power increase) differentially contribute
to task performance (Schneider et al., 2019) and that this
should be taken into account when analyzing the contribu-
tion of alpha power oscillations to behavior. Yet, it should
be noted that there are studies that successfully demon-
strated an effect of the relative strength of alpha lateraliza-
tion on task performance (Haegens et al., 2011; Kelly et al.,
2009), suggesting the reasons for those diverging results
are likely to be more complex than just a methodological
artifact. Also, it has to be noted that the respective BFs
(below 1, but above 0.33) were rather indecisive; thus,
although our data do not seem to support a significant
relationship between alpha lateralization and behavioral
performance, they cannot provide compelling evidence
for a true null effect either.
958
Journal of Cognitive Neuroscience
Volume 32, Number 5
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
5
e
9
-
4
p
5
d
f
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
a
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
.
f
/
o
n
0
5
M
a
y
2
0
2
1
Critically, one question remains unanswered: If alpha
lateralization is not a necessary component of poststimu-
lus attentional processing in an auditory scene, what does
it reflect? It might be that poststimulus alpha lateraliza-
tion is an “optional response” that may result in more ef-
fective target enhancement or distractor inhibition, when
a specific strategy is applied. Hence, because of different
strategies used by different participants, there might be no
overall relationship between alpha lateralization and be-
havior when analyzed across all participants (Limbach &
Corballis, 2017; Rihs, Michel, & Thut, 2009). Alterna-
tively, as shown in a previous study using a very similar
task design, auditory poststimulus alpha lateralization
might be more closely related to the spatial specificity
of the task (Klatt et al., 2018b). In the latter study, a
lateralization of alpha power was only evident when par-
ticipants were instructed to localize (instead of to simply
detect) a target sound within a multisound array. Hence,
we proposed that, in poststimulus attentional processing,
the lateralization of alpha power indexes the access to a
spatiotopic template that is used to generate a spatially
specific response (Klatt et al., 2018b). If alpha lateraliza-
tion reflects such a process, one may argue that there
should be no or a substantially reduced alpha lateraliza-
tion in incorrect trials, and thus, alpha lateralization
should in fact be associated with behavioral performance.
Such differences in ALI amplitudes for correct versus
incorrect trials have in fact been reported (Tune et al.,
2018; Wöstmann et al., 2016, 2018). The fact that we cal-
culated ALIs based on each participant’s mean alpha
power in correctly answered trials may explain why
we fail to capture such differences for a rather coarse,
dichotic measure of behavioral performance such as
accuracy.
Conclusion
In summary, fluctuations in N2ac amplitude predicted the
rate of information accumulation (i.e., drift rate) as well as
overall accuracy. We conclude that the N2ac component
reflected the participants’ ability to resolve competition be-
tween co-occurring sounds by focusing on the target. This,
in turn, determined the quality of the information accu-
mulated during the decision-making process and thereby
affected overall accuracy levels. In contrast, alpha lateraliza-
tion was unrelated to behavioral performance, suggesting
that successful attentional orienting within an auditory
scene (as opposed to in anticipation of an upcoming target
sound), does not rely on alpha lateralization. Our findings
strengthen the proposal that alpha lateralization is not spe-
cific to the visual domain but may reflect a supramodal at-
tentional mechanism that generalizes to the auditory
domain (Thorpe et al., 2012; Kerlin, Shahin, & Miller,
2010). Yet, we highlight that it is important to distinguish
between cue-related, anticipatory modulations of alpha
power and poststimulus alpha power lateralization.
Acknowledgments
This work was supported by the German Federal Ministry of
Education and Research in the framework of the TRAIN-STIM
project (Grant Number 01GQ1424E). The authors are grateful to
David Schmude, Jonas Heyermann, Stefan Weber, and Michael-
Christian Schlüter for their help in running the experiments; to
Peter Dillmann and Tobias Blanke for preparing software and parts
of the electronic equipment; and to two anonymous reviewers for
valuable comments on a previous version of this manuscript.
Reprint requests should be sent to Laura-Isabelle Klatt, Leibniz
Research Centre for Working Environment and Human Factors,
Ardeystraße 67, 44139 Dortmund, Germany, or via e-mail: klatt@
ifado.de.
REFERENCES
Alain, C., & McDonald, K. L. (2007). Age-related differences in
neuromagnetic brain activity underlying concurrent sound
perception. Journal of Neuroscience, 27, 1308–1314.
Bae, G.-Y., & Luck, S. J. (2018). Dissociable decoding of spatial
attention and working memory from EEG oscillations and
sustained potentials. Journal of Neuroscience, 38, 409–422.
Bronkhorst, A. W. (2015). The cocktail party problem revisited:
Early processing and selection of multi-talker speech.
Attention, Perception, & Psychophysics, 77, 1465–1487.
Cabeza, R., Anderson, N. D., Locantore, J. K., & McIntosh, A. R.
(2002). Aging gracefully: Compensatory brain activity in high-
performing older adults. Neuroimage, 17, 1394–1402.
Capilla, A., Schoffelen, J.-M., Paterson, G., Thut, G., & Gross,
J. (2014). Dissociated α-band modulations in the dorsal
and ventral visual pathways in visuospatial attention and
perception. Cerebral Cortex, 24, 550–561.
Cherry, E. C. (1953). Some experiments on the recognition of
speech, with one and with two ears. Journal of the Acoustical
Society of America, 25, 975–979.
Cook, R. D., & Weisberg, S. (1982). Residuals and influence in
regression. New York: Chapman & Hall.
Craddock, M., Poliakoff, E., El-deredy, W., Klepousniotou, E., &
Lloyd, D. M. (2017). Pre-stimulus alpha oscillations over
somatosensory cortex predict tactile misperceptions.
Neuropsychologia, 96, 9–18.
Cribari-Neto, F., & Zeileis, A. (2010). Beta regression in R.
Journal of Statistical Software, 34, 1–24.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21.
Dienes, Z. (2014). Using Bayes to get the most out of non-
significant results. Frontiers in Psychology, 5, 781.
Eimer, M. (1996). The N2pc component as an indicator of
attentional selectivity. Electroencephalography and Clinical
Neurophysiology, 99, 225–234.
Foster, J. J., & Awh, E. (2019). The role of alpha oscillations in
spatial attention: Limited evidence for a suppression account.
Current Opinion in Psychology, 29, 34–40.
Foster, J. J., Sutterer, D. W., Serences, J. T., Vogel, E. K., & Awh,
E. (2017). Alpha-band oscillations enable spatially and
temporally resolved tracking of covert spatial attention.
Psychological Science, 28, 929–941.
Foxe, J. J., Simpson, G. V., & Ahlfors, S. P. (1998). Parieto-
occipital ∼10 Hz activity reflects anticipatory state of visual
attention mechanisms. NeuroReport, 9, 3929–3933.
Gamble, M. L., & Luck, S. J. (2011). N2ac: An ERP component
associated with the focusing of attention within an auditory
scene. Psychophysiology, 48, 1057–1068.
Klatt et al.
959
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
5
e
9
-
4
p
5
d
f
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
a
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
.
/
f
o
n
0
5
M
a
y
2
0
2
1
Gamble, M. L., & Woldorff, M. G. (2015a). Rapid context-based
identification of target sounds in an auditory scene. Journal
of Cognitive Neuroscience, 27, 1675–1684.
Gamble, M. L., & Woldorff, M. G. (2015b). The temporal
cascade of neural processes underlying target detection and
attentional processing during auditory search. Cerebral
Cortex, 25, 2456–2465.
Gould, I. C., Rushworth, M. F., & Nobre, A. C. (2011). Indexing
the graded allocation of visuospatial attention using
anticipatory alpha oscillations. Journal of Neurophysiology,
105, 1318–1326.
Haegens, S., Händel, B. F., & Jensen, O. (2011). Top–down
controlled alpha band activity in somatosensory areas
determines behavioral performance in a discrimination task.
Journal of Neuroscience, 31, 5197–5204.
Haegens, S., Luther, L., & Jensen, O. (2012). Somatosensory
anticipatory alpha activity increases to suppress distracting
input. Journal of Cognitive Neuroscience, 24, 677–685.
Händel, B. F., Haarmeier, T., & Jensen, O. (2011). Alpha
oscillations correlate with the successful inhibition of
unattended stimuli. Journal of Cognitive Neuroscience, 23,
2494–2502.
Kerlin, J. R., Shahin, A. J., & Miller, L. M. (2010). Attention gain
control of ongoing cortical speech representations in a
“cocktail party.” Journal of Neuroscience, 30, 620–628.
Klatt, L.-I., Getzmann, S., Wascher, E., & Schneider, D. (2018a).
Searching for auditory targets in external space and in
working memory: Electrophysiological mechanisms
underlying perceptual and retroactive spatial attention.
Behavioural Brain Research, 353, 98–107.
Klatt, L.-I., Getzmann, S., Wascher, E., & Schneider, D. (2018b).
The contribution of selective spatial attention to sound
detection and sound localization: Evidence from event-
related potentials and lateralized alpha oscillations.
Biological Psychology, 138, 133–145.
Krekelberg, B. (2019). bayesFactor. GitHub. Retrieved from
https://www.github.com/klabhub/bayesFactor.
Leenders, M. P., Lozano-Soldevilla, D., Roberts, M. J., Jensen, O.,
& De Weerd, P. (2018). Diminished alpha lateralization
during working memory but not during attentional cueing in
older adults. Cerebral Cortex, 28, 21–32.
Lewald, J. (2016). Modulation of human auditory spatial scene
analysis by transcranial direct current stimulation.
Neuropsychologia, 84, 282–293.
Hanenberg, C., Getzmann, S., & Lewald, J. (2019). Transcranial
Lewald, J. (2019). Bihemispheric anodal transcranial direct-
direct current stimulation of posterior temporal cortex
modulates electrophysiological correlates of auditory
selective spatial attention in posterior parietal cortex.
Neuropsychologia, 131, 160–170.
Hanenberg, C., Getzmann, S., & Lewald, L. (unpublished).
Audiovisual-congruency training modulates EEG correlates of
spatial auditory selective attention.
Hansen, J. C., & Hillyard, S. A. (1980). Endogeneous brain
potentials associated with selective auditory attention.
Electroencephalography and Clinical Neurophysiology, 49,
277–290.
Heideman, S. G., Rohenkohl, G., Chauvin, J. J., Palmer, C. E.,
van Ede, F., & Nobre, A. C. (2018). Anticipatory neural
dynamics of spatial-temporal orienting of attention in
younger and older adults. Neuroimage, 178, 46–56.
Hentschke, H., & Stüttgen, M. C. (2011). Computation of
measures of effect size for neuroscience data sets. European
Journal of Neuroscience, 34, 1887–1894.
Holm, S. (1979). A simple sequentially rejective multiple test
procedure. Scandinavian Journal of Statistics, 6, 65–70.
Hong, X., Sun, J., Bengson, J. J., Mangun, G. R., & Tong, S.
(2015). Normal aging selectively diminishes alpha
lateralization in visual spatial attention. Neuroimage, 106,
353–363.
Ikkai, A., Dandekar, S., & Curtis, C. E. (2016). Lateralization
in alpha-band oscillations predicts the locus and spatial
distribution of attention. PLoS One, 11, e0154796.
Jeffreys, H. (1961). Theory of probability (3rd ed.). Oxford:
Clarendon Press.
Jensen, O., & Mazaheri, A. (2010). Shaping functional
architecture by oscillatory alpha activity: Gating by inhibition.
Frontiers in Human Neuroscience, 4, 186.
Kavanagh, R. N., Darcey, T. M., Lehmann, D., & Fender, D. H.
(1978). Evaluation of methods for three-dimensional
localization of electrical sources in the human brain. IEEE
Transactions on Biomedical Engineering, 25, 421–429.
Kelly, S. P., Gomez-Ramirez, M., & Foxe, J. J. (2009). The
strength of anticipatory spatial biasing predicts target
discrimination at attended locations: A high-density EEG
study. European Journal of Neuroscience, 30, 2224–2234.
Kelly, S. P., Lalor, E. C., Reilly, R. B., & Foxe, J. J. (2006).
Increases in alpha oscillatory power reflect an active
retinotopic mechanism for distracter suppression during
sustained visuospatial attention. Journal of Neurophysiology,
95, 3844–3851.
current stimulation over temporal cortex enhances auditory
selective spatial attention. Experimental Brain Research,
237, 1539–1549.
Lewald, J., & Getzmann, S. (2015). Electrophysiological
correlates of cocktail party listening. Behavioural Brain
Research, 292, 157–166.
Lewald, J., Hanenberg, C., & Getzmann, S. (2016). Brain
correlates of the orientation of auditory spatial attention onto
speaker location in a “cocktail party” situation.
Psychophysiology, 53, 1484–1495.
Liang, F., Paulo, R., Molina, G., Clyde, M. A., & Berger, J. O.
(2008). Mixtures of g priors for Bayesian variable selection.
Journal of the American Statistical Association, 103,
410–423.
Lilliefors, H. W. (1967). On the Kolmogorov-Smirnov test for
normality with mean and variance unknown. Journal of the
American Statistical Association, 62, 399–402.
Limbach, K., & Corballis, P. M. (2017). Alpha-power modulation
reflects the balancing of task requirements in a selective
attention task. Psychophysiology, 54, 224–234.
Luck, S. J. (2014). An introduction to the event-related
potential technique (2nd ed.). Cambridge, MA: MIT Press.
Luck, S. J., & Hillyard, S. A. (1994). Spatial filtering during
visual search: Evidence from human electrophysiology.
Journal of Experimental Psychology: Human Perception
and Performance, 20, 1000–1014.
Lüdecke, D. (2018). ggeffects: Tidy data frames of marginal
effects from regression models. Journal of Open Source
Software, 3, 772.
MathWorks. (2019). Statistics and Machine Learning ToolboxTM
user’s guide (R 2019b).
Mognon, A., Jovicich, J., Bruzzone, L., & Buiatti, M. (2011).
ADJUST: An automatic EEG artifact detector based on the
joint use of spatial and temporal features. Psychophysiology,
48, 229–240.
Mok, R. M., Myers, N. E., Wallis, G., & Nobre, A. C. (2016).
Behavioral and neural markers of flexible attention over
working memory in aging. Cerebral Cortex, 26, 1831–1842.
Morey, R. D., & Rouder, J. N. (2018). BayesFactor: Computation
of Bayes factors for common designs (R package version
0.9.12-4.2). Retrieved from https://cran.r-project.org/
package=BayesFactor.
Myers, N. E., Walther, L., Wallis, G., Stokes, M. G., & Nobre,
A. C. (2015). Temporal dynamics of attention during
encoding versus maintenance of working memory:
960
Journal of Cognitive Neuroscience
Volume 32, Number 5
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
5
e
9
-
4
p
5
d
f
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
a
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
0
5
M
a
y
2
0
2
1
Complementary views from event-related potentials and
alpha-band oscillations. Journal of Cognitive Neuroscience,
27, 492–508.
Myerson, J., Hale, S., Wagstaff, D., Poon, L. W., & Smith, G. A.
(1990). The information-loss model: A mathematical theory
of age-related cognitive slowing. Psychological Review, 97,
475–487.
Noonan, M. P., Adamian, N., Pike, A., Printzlau, F., Crittenden,
B. M., & Stokes, M. G. (2016). Distinct mechanisms for
distractor suppression and target facilitation. Journal of
Neuroscience, 36, 1797–1807.
Nunez, M. D., Vandekerckhove, J., & Srinivasan, R. (2017). How
attention influences perceptual decision making: Single-trial
EEG correlates of drift-diffusion model parameters. Journal
of Mathematical Psychology, 76, 117–130.
Oldfield, R. C. (1971). The assessment and analysis of
handedness: The Edinburgh inventory. Neuropsychologia, 9,
97–113.
Onton, J., & Makeig, S. (2006). Information-based modeling of
event-related brain dynamics. In C. Neuper & W. Klimesch
(Eds.), Progress in brain research (Vol. 159, pp. 99–120).
Amsterdam: Elsevier.
Philiastides, M. G., Ratcliff, R., & Sajda, P. (2006). Neural
representation of task difficulty and decision making during
perceptual categorization: A timing diagram. Journal of
Neuroscience, 26, 8965–8975.
Pichora-Fuller, M. K., Alain, C., & Schneider, B. A. (2017). Older
adults at the cocktail party. In J. C. Middlebrooks, J. Z. Simon,
A. N. Popper, & R. R. Fay (Eds.), The auditory system at the
cocktail party (Vol. 60, pp. 227–259). Cham: Springer.
Pituch, K. A., & Stevens, J. P. (2016). Applied multivariate
statistics for the social sciences (6th ed.). Mahwah, NJ:
Erlbaum.
Quintana, D. S., & Williams, D. R. (2018). Bayesian alternatives
for common null-hypothesis significance tests in psychiatry: A
non-technical guide using JASP. BMC Psychiatry, 18, 178.
Ratcliff, R. (1978). A theory of memory retrieval. Psychological
Review, 85, 59–108.
Ratcliff, R., & McKoon, G. (2008). The diffusion decision model:
Theory and data for two-choice decision tasks. Neural
Computation, 20, 873–922.
Ratcliff, R., Philiastides, M. G., & Sajda, P. (2009). Quality of
evidence for perceptual decision making is indexed by trial-
to-trial variability of the EEG. Proceedings of the National
Academy of Sciences, U.S.A., 106, 6539–6544.
Ratcliff, R., Spieler, D., & McKoon, G. (2000). Explicitly modeling
the effects of aging on response time. Psychonomic
Bulletin & Review, 7, 1–25.
Ratcliff, R., Thapar, A., Gomez, P., & McKoon, G. (2004). A
diffusion model analysis of the effects of aging in the lexical-
decision task. Psychology and Aging, 19, 278–289.
Ratcliff, R., Thapar, A., & McKoon, G. (2001). The effects of
aging on reaction time in a signal detection task. Psychology
and Aging, 16, 323–341.
Ratcliff, R., Thapar, A., & McKoon, G. (2003). A diffusion model
analysis of the effects of aging on brightness discrimination.
Perception & Psychophysics, 65, 523–535.
Ratcliff, R., Thapar, A., & McKoon, G. (2011). Effects of aging
and IQ on item and associative memory. Journal of
Experimental Psychology: General, 140, 464–487.
Rihs, T. A., Michel, C. M., & Thut, G. (2007). Mechanisms
of selective inhibition in visual spatial attention are indexed
by α-band EEG synchronization. European Journal of
Neuroscience, 25, 603–610.
Rihs, T. A., Michel, C. M., & Thut, G. (2009). A bias for posterior
α-band power suppression versus enhancement during
shifting versus maintenance of spatial attention. Neuroimage,
44, 190–199.
Romei, V., Gross, J., & Thut, G. (2010). On the role of
prestimulus alpha rhythms over occipito-parietal areas in
visual input regulation: Correlation or causation? Journal of
Neuroscience, 30, 8692–8697.
Samaha, J., Iemi, L., & Postle, B. R. (2017). Prestimulus alpha-
band power biases visual discrimination confidence, but not
accuracy. Consciousness and Cognition, 54, 47–55.
Sauseng, P., Klimesch, W., Stadler, W., Schabus, M.,
Doppelmayr, M., Hanslmayr, S., et al. (2005). A shift of visual
spatial attention is selectively associated with human EEG
alpha activity. European Journal of Neuroscience, 22,
2917–2926.
Schmiedek, F., Oberauer, K., Wilhelm, O., Süß, H.-M., &
Wittmann, W. W. (2007). Individual differences in
components of reaction time distributions and their relations
to working memory and intelligence. Journal of
Experimental Psychology: General, 136, 414–429.
Schneider, D., Göddertz, A., Haase, H., Hickey, C., &
Wascher, E. (2019). Hemispheric asymmetries in EEG
alpha oscillations indicate active inhibition during
attentional orienting within working memory. Behavioural
Brain Research, 359, 38–46.
Schneider, B., Pichora-Fuller, M. K., & Daneman, M. (2010).
Effects of senescent changes in audition and cognition on
spoken language comprehension. In S. Gordon-Salant, D. R.
Frisina, A. N. Popper, & R. R. Fay (Eds.), The aging auditory
system (Vol. 34, pp. 167–210). New York: Springer.
Schubert, A.-L., Hagemann, D., Voss, A., Schankin, A., &
Bergmann, K. (2015). Decomposing the relationship between
mental speed and mental abilities. Intelligence, 51, 28–46.
Schubert, A.-L., Nunez, M. D., Hagemann, D., &
Vandekerckhove, J. (2019). Individual differences in cortical
processing speed predict cognitive abilities: A model-based
cognitive neuroscience account. Computational Brain &
Behavior, 2, 64–84.
Shinn-Cunningham, B. (2017). Cortical and sensory causes of
individual differences in selective attention ability among
listeners with normal hearing thresholds. Journal of Speech,
Language, and Hearing Research, 60, 2976–2988.
Snyder, J. S., & Alain, C. (2005). Age-related changes in neural
activity associated with concurrent vowel segregation.
Cognitive Brain Research, 24, 492–499.
Spaniol, J., Madden, D. J., & Voss, A. (2006). A diffusion model
analysis of adult age differences in episodic and semantic
long-term memory retrieval. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 32, 101–117.
Starns, J. J., & Ratcliff, R. (2010). The effects of aging on the
speed–accuracy compromise: Boundary optimality in the
diffusion model. Psychology and Aging, 25, 377–390.
Thorpe, S., D’Zmura, M., & Srinivasan, R. (2012). Lateralization
of frequency-specific networks for covert spatial attention
to auditory stimuli. Brain Topography, 25, 39–54.
Thut, G., Nietzel, A., Brandt, S. A., & Pascual-Leone, A. (2006).
α-band electroencephalographic activity over occipital cortex
indexes visuospatial attention bias and predicts visual target
detection. Journal of Neuroscience, 26, 9494–9502.
Tune, S., Wöstmann, M., & Obleser, J. (2018). Probing the limits
of alpha power lateralisation as a neural marker of selective
attention in middle-aged and older listeners. European
Journal of Neuroscience, 48, 2537–2550.
Turner, B. M., Rodriguez, C. A., Norcia, T. M., McClure, S. M., &
Steyvers, M. (2016). Why more is better: Simultaneous
modeling of EEG, fMRI, and behavioral data. Neuroimage,
128, 96–115.
Vaden, R. J., Hutcheson, N. L., McCollum, L. A., Kentros, J., &
Visscher, K. M. (2012). Older adults, unlike younger adults,
do not modulate alpha power to suppress irrelevant
information. Neuroimage, 63, 1127–1133.
Klatt et al.
961
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
5
e
9
-
4
p
5
d
f
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
a
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
f
t
.
o
n
0
5
M
a
y
2
0
2
1
Van der Lubbe, R. H. J., Bundt, C., & Abrahamse, E. L. (2014).
Internal and external spatial attention examined with
lateralized EEG power spectra. Brain Research, 1583,
179–192.
van der Waal, M., Farquhar, J., Fasotti, L., & Desain, P. (2017).
Preserved and attenuated electrophysiological correlates of
visual spatial attention in elderly subjects. Behavioural Brain
Research, 317, 415–423.
van Dijk, H., Schoffelen, J.-M., Oostenveld, R., & Jensen, O.
(2008). Prestimulus oscillatory activity in the alpha band
predicts visual discrimination ability. Journal of
Neuroscience, 28, 1816–1823.
van Driel, J., Gunseli, E., Meeter, M., & Olivers, C. N. L. (2017).
Local and interregional alpha EEG dynamics dissociate
between memory for search and memory for recognition.
Neuroimage, 149, 114–128.
van Ede, F., Niklaus, M., & Nobre, A. C. (2017). Temporal
expectations guide dynamic prioritization in visual working
memory through attenuated α oscillations. Journal of
Neuroscience, 37, 437–445.
van Ede, F., Szebényi, S., & Maris, E. (2014). Attentional
modulations of somatosensory alpha, beta and gamma
oscillations dissociate between anticipation and stimulus
processing. Neuroimage, 97, 134–141.
Voss, A., Nagler, M., & Lerche, V. (2013). Diffusion models in
experimental psychology: A practical introduction.
Experimental Psychology, 60, 385–402.
Voss, A., & Voss, J. (2007). Fast-dm: A free program for efficient
diffusion model analysis. Behavior Research Methods, 39,
767–775.
Voss, A., Voss, J., & Lerche, V. (2015). Assessing cognitive
processes with diffusion model analyses: A tutorial based on
fast-dm-30. Frontiers in Psychology, 6, 336.
Wagenmakers, E.-J., Marsman, M., Jamil, T., Ly, A., Verhagen, J.,
Love, J., et al. (2018). Bayesian inference for psychology.
Part I: Theoretical advantages and practical ramifications.
Psychonomic Bulletin & Review, 25, 35–57.
Wildegger, T., van Ede, F., Woolrich, M., Gillebert, C. R., &
Nobre, A. C. (2017). Preparatory α-band oscillations reflect
spatial gating independently of predictions regarding target
identity. Journal of Neurophysiology, 117, 1385–1394.
Worden, M. S., Foxe, J. J., Wang, N., & Simpson, G. V. (2000).
Anticipatory biasing of visuospatial attention indexed by
retinotopically specific α-band electroencephalography
increases over occipital cortex. Journal of Neuroscience, 20,
RC63.
Wöstmann, M., Herrmann, B., Maess, B., & Obleser, J. (2016).
Spatiotemporal dynamics of auditory attention synchronize
with speech. Proceedings of the National Academy of
Sciences, U.S.A., 113, 3873–3878.
Wöstmann, M., Vosskuhl, J., Obleser, J., & Herrmann, C. S.
(2018). Opposite effects of lateralised transcranial alpha
versus gamma stimulation on auditory spatial attention.
Brain Stimulation, 11, 752–758.
Yamagishi, N., Goda, N., Callan, D. E., Anderson, S. J., & Kawato,
M. (2005). Attentional shifts towards an expected visual target
alter the level of alpha-band oscillatory activity in the human
calcarine cortex. Cognitive Brain Research, 25, 799–809.
Zündorf, I. C., Karnath, H.-O., & Lewald, J. (2011). Male advantage
in sound localization at cocktail parties. Cortex, 47, 741–749.
Zündorf, I. C., Karnath, H.-O., & Lewald, J. (2014). The effect of
brain lesions on sound localization in complex acoustic
environments. Brain, 137, 1410–1418.
Zündorf, I. C., Lewald, J., & Karnath, H.-O. (2013). Neural
correlates of sound localization in complex acoustic
environments. PLoS One, 8, e64259.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
5
e
9
-
4
p
5
d
f
2
0
/
1
3
3
2
3
/
6
5
2
/
9
o
4
c
5
n
/
_
a
1
_
8
0
6
1
1
5
7
2
7
5
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
2
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
t
f
.
o
n
0
5
M
a
y
2
0
2
1
962
Journal of Cognitive Neuroscience
Volume 32, Number 5