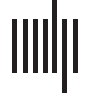ARTICLE DE RECHERCHE
Investigating the Link Between Linguistic and
Non-Linguistic Cognitive Control in Bilinguals
Using Laplacian-Transformed Event
Related Potentials
Martha N. Mendoza1
Robert T. Knight1
, Henrike K. Blumenfeld2,4
, and Stephanie K. Ries2,3,4
,
1Helen Wills Neuroscience Institute and Department of Psychology, Université de Californie, Berkeley, Berkeley, Californie, Etats-Unis
2School of Speech, Language, and Hearing Sciences, San Diego State University, San Diego, Californie, Etats-Unis
3Center for Clinical and Cognitive Neuroscience, San Diego State University, San Diego, Californie, Etats-Unis
4SDSU-UCSD Joint Doctoral Program in Language and Communicative Disorders, San Diego, Californie, Etats-Unis
Mots clés: bilingual language control, cognitive control, cross-linguistic interference, Laplacian
transformation, event-related potentials
ABSTRAIT
Bilinguals’ need to suppress the activation of their other language while speaking has been
proposed to result in enhanced cognitive control abilities outside of language. Several studies
therefore suggest shared cognitive control processes across linguistic and non-linguistic tasks.
Here we investigate this potential overlap using scalp electroencephalographic recordings and
the Laplacian transformation, providing an estimation of the current source density and
enabling the separation of EEG components in space. Fourteen Spanish-English bilinguals
performed a picture-word matching task contrasting incongruent trials using cross-linguistic
false cognates (par exemple., a picture – foot, overlaid with distractor text: the English word PIE, c'est à dire., le
false cognate for the Spanish pie meaning “foot”) with congruent trials (matching English
picture names and words, c'est à dire., a picture – foot, with overlaid text: the English word FOOT ),
and an unrelated control condition. En outre, participants performed an arrow-version of the
Eriksen flanker task. Worse behavioral performance was observed in incongruent compared to
congruent trials in both tasks. In the non-linguistic task, we replicated the previously observed
congruency effect on a medial-frontal event-related potential (ERP) peaking around 50 ms
before electromyography (EMG) début. A similar ERP was present in the linguistic task, était
sensitive to congruency, and peaked earlier, autour 150 ms before EMG onset. En outre,
another component was found in the linguistic task at a left lateralized anterior frontal site
peaking around 200 ms before EMG onset, but was absent in the non-linguistic task. Notre
results suggest a partial overlap between linguistic and non-linguistic cognitive control
processes and that linguistic conflict resolution may engage additional left anterior frontal
control processes.
INTRODUCTION
Investigating the cognitive consequences of bilingualism has become a source of great interest
in the past few decades. Evidence from language perception and production tasks (par exemple., lexical
décision, picture naming) indicates that bilinguals’ languages are active in parallel (Colomé,
un accès ouvert
journal
Citation: Mendoza, M.. N., Blumenfeld,
H. K., Knight, R.. T., & Ries, S. K. (2021).
Investigating the link between linguistic
and non-linguistic cognitive control in
bilinguals using Laplacian-transformed
event related potentials. Neurobiology
of Language, 2(4), 605–627. https://est ce que je
.org/10.1162/nol_a_00056
EST CE QUE JE:
https://doi.org/10.1162/nol_a_00056
Informations complémentaires:
https://doi.org/10.1162/nol_a_00056
Reçu: 1 Février 2021
Accepté: 10 Septembre 2021
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
Stephanie K. Ries
sries@sdsu.edu
Éditeur de manipulation:
Manuel Carreiras
droits d'auteur: © 2021
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
un
_
0
0
0
5
6
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
2001; Costa et al., 1998; Dijkstra & van Heuven, 1998; Vert, 1998; Kroll et al., 2008; Marian
et coll., 2003; Marian & Spivey, 2003). In order to speak or understand an intended language,
bilinguals must therefore be able to selectively inhibit or filter non-target language represen-
tations (for reviews, see La Heij, 2005; van Heuven & Dijkstra, 2010). The unique need bilin-
guals have to constantly regulate two languages has been proposed to result in enhanced
cognitive control abilities that expand beyond the language domain (Bialystok et al., 2004,
2008; Bialystok & Craik, 2010; Kroll & Bialystok, 2013), and has been coined the bilingual
advantage (BA). In support of this claim, a large number of studies have shown that bilinguals
outperform monolinguals in non-linguistic tasks that make use of cognitive control processes
(Bialystok et al., 2004, 2008; Costa et al., 2008, 2009). The underlying assumption is that these
cognitive control processes are shared between language and other functions. Donc, ces
cognitive control processes are adaptively “trained” by the constant need bilinguals have to
suppress their other language or switch between their languages (Vert & Abutalebi, 2013).
Cependant, a significant number of studies have also reported results in which bilinguals show
no such advantage in the same non-linguistic tasks that have been used to show the BA (pour
reviews see Lehtonen et al., 2018; Paap & Greenberg, 2013). Donc, the existence of these
shared cognitive control processes in bilinguals has been questioned.
Brain imaging studies have sought to answer the question of whether there is a possible func-
tional overlap between cognitive control and language tasks in bilinguals by investigating
whether similar brain regions or electrophysiological components may be engaged in both
types of tasks. Cependant, determining when functional overlap across linguistic and non-
linguistic tasks can be interpreted as constituting domain-generality in processing can be a
matter of debate. Nozari and Novick (2017) argued that two criteria must be met for determin-
ing functional overlap and inferring domain-generality in processing: “shared computational
principles” and “shared neural implementation.” Shared neural implementation can be cap-
tured, Par exemple, through the overlap of brain activation across tasks in functional magnetic
resonance imaging (IRMf) études. Shared computational principles refer to similar processes
operating in different domains on different domain-specific representations. As argued in
Nozari and Novick (2017), it is undisputed that representations are domain-specific and are
stored by different cortical regions. Cependant, the processes operating upon these representa-
tions may be common across domains. In the comparison between non-linguistic and linguistic
tasks, these processes are what is generally targeted. Even though shared neural implementa-
tion can be an argument in favor of shared computational principles, it may not always be
sufficient. Additional information from imaging modalities beyond fMRI can help further sup-
port the existence of shared computational principles across domains. Par exemple, the shared
computational principles criterion may be supported by identifying event-related potential
(ERP) components modulated in the same way across domains and emerging from the same
general areas as those showing shared neural implementation. Below, we review fMRI and
EEG studies investigating the functional overlap between linguistic and non-linguistic tasks
in bilinguals, and discuss how the present study can bring further information in this respect.
FMRI studies in bilinguals have revealed functional overlap in terms of brain activation
between cognitive control and language tasks (Coderre et al., 2016; Garbin et al., 2010;
van Heuven & Dijkstra, 2010). Surtout, several brain imaging studies have found that
bilinguals activate a brain network typically associated with executive control, y compris
medial frontal regions such as the anterior cingulate gyrus (ACC), the pre-supplementary motor
area (pre-SMA), left inferior prefrontal cortex, and the left caudate nucleus when they need to
manage cross-linguistic conflict arising from the automatic lexical activation of the non-target
langue (Rodriguez-Fornells et al., 2005; van Heuven et al., 2008). Bilinguals have also been
Neurobiology of Language
606
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
un
_
0
0
0
5
6
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
shown to activate brain regions typically associated with language tasks, such as the left infe-
rior frontal gyrus, when performing non-linguistic cognitive control tasks (par exemple., a non-linguistic
switching task; Garbin et al., 2010). This cognitive control network underlying bilingual
processing has been confirmed by large meta-analytic studies combining data on bilingual
traitement (Abutalebi & Vert, 2007; Sulpizio et al., 2020). Dans l'ensemble, these studies suggest that
similar brain regions are modulated by cognitive control demands in the linguistic and
non-linguistic domains in bilinguals. En outre, the current literature linking language control
during bilingual processing to a domain-general executive control network (for a review, voir
Calabria et al., 2018) aligns well with Fedorenko and Thompson-Schill’s (2014) view of a spe-
cialized “core” language system that is overlaid with a domain-general “peripheral” system.
In examining the overlap between linguistic and non-linguistic cognitive control networks
in bilinguals, a central strategy is to identify language tasks where cross-linguistic interference
has been documented during processing. False cognates, also known as interlingual homo-
graphs or homophones, provide a lens into bilingual processing contexts where cross-
linguistic interference is present. Par exemple, the English word for a baked treat, pie, moyens
“foot” in Spanish, thus activating distinct semantic representations in each language of
Spanish-English bilinguals. Behavioral evidence shows lower accuracy rates and longer reac-
tion times in bilinguals for interlingual homograph processing relative to control words (par exemple.,
van Heuven et al., 2008; Vanlangendonck et al., 2019; von Studnitz & Vert, 2002). Ce
finding has been interpreted as reflecting crosslinguistic interference at the lexico-semantic
level as participants select a language-specific response. Using fMRI with a lexical decision
task in a monolingual context, van Heuven et al. (2008) found enhanced blood-oxygen-level-
dependent (AUDACIEUX) signal in areas of the executive control network, including the pre-SMA
and ACC when Dutch-English bilinguals processed interlingual homographs (par exemple., the word
room is part of a house in English but means “cream” in Dutch; van Heuven et al., 2008).
In a similar lexical decision study in Dutch-English bilinguals, Peeters et al. (2019) also iden-
tified activation in pre-SMA and left inferior frontal gyrus (IFG) associated with processing of
interlingual homographs compared to English control words. Cependant, showing a direct neu-
ral implementation overlap between linguistic and cognitive control networks requires using
linguistic and non-linguistic tasks directly in the same participants. So far, only a few studies
have used this direct comparison in the same participants (Coderre et al., 2016; De Baene
et coll., 2015; Ye & Zhou, 2009), and only Coderre et al. (2016) tested bilinguals specifically.
En particulier, overlapping activations in the arrow version of the Eriksen flanker task (Eriksen &
Eriksen, 1974; Stoffels & van der Molen, 1988) and a semantic categorization task have been
found in the left inferior frontal gyrus in bilinguals but not in monolinguals (Coderre et al.,
2016), suggesting shared neural implementation across domains in bilinguals.
Although fMRI studies can inform us about whether or not similar brain regions are engaged
in linguistic and non-linguistic control, studying the time point at which this functional overlap
takes place in the bilingual brain requires temporally resolved techniques, which could help
identify shared computational principles. Previous studies using EEG and, in particular, ERPs
have indirectly and directly investigated this issue using a variety of paradigms, y compris
language switching paradigms (par exemple., G. M.. Jackson et al., 2001), go/no-go tasks manipulating
interference at different levels (Rodriguez-Fornells et al., 2006), and the negative priming par-
adigm (Dash & Kar, 2020). These studies have often reported fronto-central activity peaking
entre 200 et 500 ms post-stimulus onset (N2 and N400 components) as being larger in
situations requiring more control (par exemple., switch trials, no-go trials, cross-linguistic incongruent
trials) compared to situations requiring less control (par exemple., non-switch trials, go trials, cross-
linguistic congruent trials) in linguistic and non-linguistic tasks (S. R.. Jackson et al., 1999).
False cognates:
Words that overlap in form but not
meaning in different languages; aussi
known as interlingual homophones
and homographs.
Event-related potentials (ERPs):
Voltage fluctuations recorded
through electroencephalography
at the scalp that are time-locked
to an event.
Neurobiology of Language
607
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
un
_
0
0
0
5
6
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
Laplacian transformation:
The mathematical process by which
estimates of radial current flow at the
scalp, or current source density, sont
derived from the scalp-recorded
electroencephalogram.
Cependant, similarly as for fMRI studies, few studies have directly compared ERPs associated
with cognitive control processes in linguistic and non-linguistic tasks in the same participants.
A recent study (Dash & Kar, 2020) used the negative priming paradigm and an animacy judg-
ment task, with words and pictures in the linguistic versus non-linguistic versions respectively.
The results showed modulations of the N2 amplitude in the linguistic and non-linguistic
versions alike, suggesting a computational (traitement) overlap between linguistic and non-
linguistic domains in bilinguals. While these results are informative, the non-linguistic task
used pictures representing nameable items, which may not have been ideal given the possible
internal word retrieval process induced by the stimuli. Another possible more general limitation
stems from the limited spatial resolution of traditional ERP studies, making it difficult to observe
components that could be differentially affected by linguistic compared to non-linguistic inter-
ference at similar time points.
Previous non-linguistic electrophysiological studies using the Laplacian transformation have
described ERPs associated with cognitive control mechanisms time-locked to the response. Le
Laplacian transformation provides an estimate of the current source density and therefore more
focal topographic resolution than traditional ERPs (Babiloni et al., 2001). Using this technique
has enabled the dissociation of activity occurring at neighboring sites but attributable to differ-
ent cognitive processes. One of these components, the N-40, is of particular interest here. Le
N-40 is a negative going wave that peaks around 40 ms prior to electromyographic (EMG) début
leading to the response (c'est à dire., the button press), maximal over medial-frontal electrodes (FCz),
and preceding the components associated with response execution and inhibition recorded
over the contralateral and ipsilateral motor cortices respectively (Vidal et al., 2003). Critique,
these later motor components cannot be dissociated from the N-40 in traditional monopolar
ERP analyses, which may lead to a confound between these activities if Laplacian transforma-
tion is not used (for a demonstration, see Burle et al., 2015). By using Laplacian transformation
and recording the EMG activity associated with the response, this component was found to be
more closely aligned in time to the onset of the EMG activity leading to the response than to
the button press itself (Vidal et al., 2003). The N-40 is larger in incongruent than congruent
trials in the Eriksen flanker task (Roger, 2009). The N-40, along with the inhibition of the ipsi-
lateral motor cortex associated with the hand not being used, also disappears when there is no
choice to be made between two possible responses (in go/no-go tasks; Vidal et al., 2011). Dans
addition, the amplitude of this component is reduced when biasing information about the
response to be produced is available to the participant (Carbonnell et al., 2004). The N-40
has therefore been associated with decision-making processes, and in particular response
selection, in choice reaction time tasks (Vidal et al. 2011), where participants must decide
between two conflicting responses, usually a left or right button press. Surtout, the N-40
is dissociable from the lateralized readiness potential, which is lateralized and peaks around
response onset (for a review, see Vidal et al., 2018). De plus, as mentioned earlier, the N-40
peaks around 40 ms prior to EMG onset leading to the response (c'est à dire., the button press), et
hence is also different from the error-related negativity (ERN), which peaks after the response,
and which has been associated with action monitoring, including language output monitoring
(par exemple., Riès et al., 2011, 2020; Riès, Xie, et coll., 2013) and conflict monitoring (par exemple., Masaki et al.,
2012; Yeung et al., 2004). Although Laplacian transformation does not allow the identification
of specific neural sources, the fronto-central distribution of the N-40 is compatible with the
engagement of medial frontal regions in cognitive control (for a review, see Ridderinkhof
et coll., 2011).
Dans cette étude, we used the same Laplacian transformation technique and compared a linguistic
decision-making task, indexing cross-linguistic interference using interlingual homographs, to a
Neurobiology of Language
608
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
un
_
0
0
0
5
6
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
non-linguistic decision-making task. In the current study, we will refer to interlingual homographs
as false cognates to focus on the competing meanings that are activated by these words across
languages. We hypothesize that if bilinguals use shared cognitive control processes in the
language domain and outside of language, then this overlap may be visible on the N-40 component.
More specifically, we hypothesize that a negative component will be observed in our linguistic
decision-making task at the same fronto-central recording site as in the non-linguistic decision-
making task and that this component will be sensitive to the congruency manipulations in both tasks.
Finding a similar fronto-central decision-making mechanism engaged in linguistic and non-linguistic
domains would provide support for the idea that similar cognitive control processes are involved in
language and outside of language, providing a further argument in favor of shared neural computa-
tions in language and outside of language in bilinguals. We have previously used Laplacian trans-
formation to examine the brain dynamics underlying picture naming and found two main frontal
components, one fronto-central peaking between 300 et 200 ms before vocal onset, ainsi que
a left frontal component peaking around vocal onset (Riès, Janssen, et coll., 2013). While this pre-
vious study was mainly descriptive, did not target bilinguals, and did not include a non-linguistic
task, we are expecting to find similar medial frontal and left frontal components in the linguistic
task in the present study. Whether or not these components will be similarly sensitive to non-
linguistic and linguistic manipulations will be tested here.
The Current Study
We recorded scalp EEG in 17 Spanish-English participants as they performed a picture-word
matching (PWM) task, and the arrow version of the Eriksen flanker task (Eriksen & Eriksen,
1974; Stoffels & van der Molen, 1988). In the PWM task, participants decided whether a
picture, and a word superimposed on it, corresponded to the same word or not. The PWM
contrasted incongruent trials using false cognates (par exemple., picture – foot; distractor: English word
PIE, c'est à dire., false cognate for the Spanish pie meaning “foot”) with congruent trials (matching
picture name and word) and an unrelated control condition. We also employed the arrow
version of the flanker task, where participants decided on the direction of a central arrow while
ignoring flanking arrows. In incongruent trials, flanking arrows were in the opposite direction of
the central arrow, while they matched in congruent trials. We preferred the arrow version of the
Eriksen flanker task over the original version (using letter stimuli) as we aimed to make this task
as non-linguistic as possible.
We aligned the linguistic PWM task with the non-linguistic Eriksen flanker task in terms of
their underlying loci of cognitive control. Spécifiquement, the incongruent conditions on both
tasks were characterized as including stimulus-level and response-level conflict, sources of
conflict that are considered separable during bilingual processing (Dijkstra & van Heuven,
2002) and in non-linguistic cognitive control tasks (Kornblum et al., 1999). Stimulus- et
response-based conflict was specifically examined in false cognates by van Heuven et al.
(2008): Stimulus-based conflict (c'est à dire., the two possible interpretations of false cognates across
languages) was associated with activity in anterior and posterior left inferior prefrontal cortex;
response-based conflict (c'est à dire., the conflict generated when participants had to explicitly identify
language membership of the false cognates at the response level) was associated with activity
in pre-SMA and ACC. De la même manière, the incongruent condition of our flanker task contains
stimulus-based conflict (c'est à dire., the presence of right-pointing and left-pointing arrows around a
central arrow on the display) and response-level conflict (par exemple., the mapping of a right-hand
response, despite the presence of left-pointing arrows on the display).
In both the PWM and flanker tasks, participants pressed buttons using their thumbs to give
their answers, and EMG activity of the corresponding muscles ( flexor pollicis brevis) était
Neurobiology of Language
609
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
un
_
0
0
0
5
6
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
recorded along with scalp EEG (as in, par exemple., Riès et al., 2011; Roger, 2009; Vidal et al., 2011).
EMG recordings were used to mark the onset of the muscular activity associated with pressing
the response buttons. Laplacian-transformed ERPs were time-locked to EMG onset to investi-
gate the potential overlap between medial frontal cognitive control processes in linguistic and
non-linguistic decision-making.
We focused on the medial frontal and left frontal components previously described in linguistic
(Riès, Janssen, et coll., 2013) and non-linguistic tasks (Vidal et al., 2011). En particulier, we focused on
the N-40 component as this component has been previously described in non-linguistic tasks
focusing on cognitive control, and its amplitude has been shown to be modulated by congruency
in the flanker task (Roger, 2009). We hypothesized that a larger N-40 would be observed in
incongruent than in congruent trials in the flanker task as in Roger (2009). If the same underlying
mechanism is similarly engaged in linguistic decision-making and is sensitive to cross-linguistic
interference, then the N-40 should also be larger in the false-cognate compared to the congruent
(c'est à dire., matching) condition in the PWM task.
We further investigated whether additional left frontal activity would be sensitive to congruency
in the PWM task and whether this left frontal activity would be selectively engaged in the
linguistic task. Based on the results from the bilingual cognitive control literature (Abutalebi &
Vert, 2007; Sulpizio et al., 2020), we expected to find a left-lateralized ERP component sensitive
to congruency in both the linguistic and non-linguistic tasks and reflecting the overlap in cognitive
control function subserved by the left inferior frontal cortex. Left frontal activity has been previously
reported in picture naming using the Laplacian transformation (Riès, Janssen, et coll., 2013);
therefore we expected to find similar activity in the present study in the linguistic task. Si
or not this same component is also present in non-linguistic tasks and is sensitive to cognitive
control demands in linguistic and non-linguistic tasks remains to be investigated.
MÉTHODES
Participants
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
un
_
0
0
0
5
6
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The study was performed in agreement with the Declaration of Helsinki. All participants gave
informed consent approved by the University of California, Berkeley, Committee for Protection
of Human Subjects before the experiment.
A total of 17 right-handed Spanish-English bilinguals with normal or corrected to normal
vision participated in the experiment (âge moyen = 20.41 années, SD = 1.12 années). The data of
3 participants were not included in the analysis because of failing to complete the experiment
(n = 1) or technical difficulties during EEG recording causing the EEG data to be unusable (n = 1)
or absent in one of the tasks (n = 1). Ainsi, the data of 14 participants (4 males, âge moyen =
20.21 années, SD = 1.05 années) was processed and analyzed for the current study. This number of
participants is in line with previous studies of decision-making and response selection using
linguistic and non-linguistic tasks describing similar components using EEG and Laplacian
transformation (Carbonnell et al., 2004; Riès, Janssen, et coll., 2013; Vidal et al., 2003, 2011).
All participants were students at the University of California, Berkeley. They were recruited
through the Research Participatory Program (RPP) from the psychology department and
received course credit or monetary compensation for their participation. The recruitment pro-
cess was conducted in English, and participants were recruited specifically because they were
Spanish-English bilinguals (Spanish was their first/native language). More specifically, le
recruitment language indicated that participants must be at least 18 years of age, be a
Spanish-English bilingual (specifying that Spanish must be their first language), be fluent in
Neurobiology of Language
610
Bilingual cognitive control as revealed by Laplacian transformed ERPs
English and Spanish and use both languages at similar levels, be right-handed, and have
normal or corrected-to-normal vision. All of the participants reported learning Spanish as their
first language and English as their second language. The mean age for acquisition of English
était 5.89 years of age (SD = 3.37). They all self-reported being more dominant in English at the
time of testing, although speaking Spanish regularly. All interactions, questions, and instruc-
tions were kept in English throughout the participants’ involvement in the experiment.
Materials and Design
Flanker task
Stimuli consisted of five arrows presented in white on a black background at the center of the
screen, presented in free viewing within a visual angle of 7°. There were four stimulus condi-
tion: congruent right (>>>>>), congruent left (<<<<<), incongruent right (<<><<), and incon-
gruent left (>><>>). In analyses, congruent trials were compared to incongruent trials, avec
each condition including right- and left-facing arrow trials. There was a total of eight blocks
containing 60 trials each, and a total of 120 trials per condition (c'est à dire., congruent left, congruent
droite, incongruent left, and incongruent right) overall, hence the ratio of congruent to incon-
gruent trials was 1 à 1 in this task. Stimuli were pseudorandomized, such that identical trials
were not repeated more than 5 times in a row.
Picture-word matching task
Thirty-two colored pictures of common objects were selected as stimuli from the Bank of Stan-
dardized Stimuli (BOSS; Brodeur et al., 2010). All stimuli were presented at the center of a
computer monitor.
Stimuli consisted of color pictures of common items fitted to a 2,000 × 2,000 pixel, blanc
square background, superimposed with words centered to the center of the square, written in
black bolded Arial font, 180 pixels high, over a transparent white rectangle (opacity: 65%),
and were presented in free viewing within a visual angle of 7°. There were three stimulus con-
ditions: a Congruent or Identity (ID) condition, in which the picture and the word matched
(c'est à dire., picture – foot; word – foot); an Incongruent or False-Cognate (FC) condition, where the
picture and the word did not match in the target language, English, but where the word was a
false-cognate to the Spanish picture-name (c'est à dire., picture – foot; word – pie; Spanish picture-
name = pie meaning “foot”; false cognate: English pie and Spanish pie); and an Unrelated
control (UR) condition, where the picture and the word did not match in either language
(c'est à dire., picture – foot; word – bread ) (voir la figure 1). Thus in the FC condition, while bilingual
participants might have been tempted to respond that the picture and overlaid word were
matches since the English word PIE was also the Spanish translation equivalent of foot, ce
response would have been incorrect. Across the stimulus set of 32 target pictures with their
corresponding false cognate distractor words, three false cognates were identical homographs
of the picture’s Spanish translation equivalent (bread – PAN; foot – PIE; net – RED). (See the
Appendix, located in the Supporting Information at https://doi.org/10.1162/nol_a_00056.) Le
remaining false cognates were near-homographs of the picture’s Spanish translation equivalent
(par exemple., candle [Spanish vela] – VEIL; elbow [Spanish codo] – CODE ). There was a total of four
blocks with 96 trials each. Each stimulus of the 32 unique stimuli was repeated 3 times per
block, once in each condition (ID, FC, UR); hence, the rate of match (“yes” responses on ID
trials) compared to no-match responses (“no” responses on FC and UR trials) était 1 à 2 in this
task. Stimuli were pseudorandomized within blocks such that 2 consecutive stimuli did not
share the same phonological onset and were not semantically related.
Neurobiology of Language
611
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
un
_
0
0
0
5
6
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
Chiffre 1. Picture-word matching task example stimuli in the Identity (ID) condition (the picture name and the distractor word match), le
False Cognate (FC) condition (the picture name and the distractor word do not match and are false cognates, pie means “foot” in Spanish but
not in English), and the Unrelated (UR) condition (the picture name and the distractor word do not match and are unrelated phonologically
and semantically). In the FC condition, while participants might be tempted to respond that the picture and overlaid word are matches since
the English word PIE is a perfect homograph with the Spanish translation equivalent of foot, this response would be incorrect.
Procedure
Behavioral recordings
Reaction times and accuracy were recorded using E-Prime’s Serial Response Box (SRB).
Participants were first familiarized with the stimuli and the conditions in each task. Le
E-prime 2.0 Professional software (Psychology SoftwareTools, Inc., Pittsburg, Pennsylvanie) was used
to control stimulus presentation for both tasks. Participants were comfortably seated at eye
level to the computer monitor on which the stimuli were presented in a soundproof, dimly
lit room. All participants completed the flanker task and the PWM task. Half of the partic-
ipants performed the flanker task first. Participants were instructed to respond as fast and as
accurately as possible. They were also told that the target language for the PWM task was
English, and all interactions between the experimenters and participants were in English.
En outre, participants were instructed to remain as still as possible during experimental
blocks in order to prevent electromyographic (EMG) activity from contaminating the EEG
signal as much as possible. Participants were able to rest for as long as needed between blocks
in each task.
Flanker task. A trial consisted in the following: (1) a fixation cross (a picture of a white plus
sign on a black background) was displayed for 1,000 ms at the center of the screen; (2) le
stimulus was presented on a black background for 120 ms; (3) a blank black screen displayed
pour 1,000 ms after stimulus offset. Participants were instructed to indicate the direction of the
central target arrow by pressing a left or right button on the response box (the buttons were
marked “left” and “right”). Responses could be made during stimulus presentation or during
the blank screen, but the stimulus and blank screen remained even after the response had been
made in order to maintain coherent trial structure.
Picture-word matching task. A trial consisted in the following: (1) a fixation cross (a picture of a
black plus sign on a white background) was presented for 1,000 ms at the center of the screen;
(2) the stimulus was presented for 1,000 ms; (3) a blank screen was presented for 2,000 ms
after the stimulus offset. Participants were instructed to indicate whether the picture and the
word matched by pressing a right or left button marked “Match” or “No Match” on the
Neurobiology of Language
612
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
un
_
0
0
0
5
6
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
corresponding SRB keys. Similar to the flanker task, responses could be made during stimulus
presentation or during the blank screen.
Electrophysiological recordings
Electroencephalography was recorded using 64 Ag/AgCI pre-amplified electrodes (BIOSEMI,
Amsterdam, Netherlands; 10–20 system positions). The sampling rate was 1024 Hz (avec
acquisition filters: DC to 208 Hz, 3 db/octave). Two surface electrodes (Ag/AgCl) were placed
autour 2 cm apart on the skin of the thenar eminence to record the EMG activity from the thumb
muscle, flexor pollicis brevis. The vertical electrooculogram (EOG) was recorded by two sur-
face electrodes (Ag/AgCl) placed above and below the left eye. Horizontal EOG was recorded
by two electrodes placed next to the outer canthi. The passive reference electrode was placed
over the right mastoid.
Data Preprocessing
Behavioral data
Trials were considered errors when participants pressed the wrong button, did not produce a
response, or when the produced response occurred later than 1,120 ms for the flanker task or
3,000 ms for the PWM task, respectivement, corresponding to the time the stimulus and following
white screen were presented in each task. Surtout, these cutoffs were well over 3 standard
deviations above the mean reaction time (RT) in each task (Mean RT + 3 * SD in flanker =
701.5 ms; Mean RT + 3 * SD in PWM = 1,284.6 ms). Reaction times were measured between
the time of stimulus presentation and the time of the button press.
EEG and EMG data
After data acquisition, EMG data were filtered (high pass = 10 Hz; low-pass = 300 Hz), et
rectified. EMG onsets were manually marked on a trial-by-trial basis. Surtout, EMG onsets
were clearly discriminable from the baseline. Trials containing more than one EMG burst
before the button press were rejected from further analyses.
The EEG data were resampled to 256 Hz and vertical eye movements (c'est à dire., eye blinks) étaient
removed using independent component analysis as implemented in EEGLAB (Delorme &
Makeig, 2004). A blind source separation algorithm based on canonical correlation analysis
(BSS-CCA; De Clercq et al., 2006; de Vos et al., 2010) was applied on non-overlapping con-
secutive 30-sec time windows on monopolar recordings in order to remove any EMG activity
(due to frowning or other muscular tension) that may have contaminated the EEG data.
Following the BSS-CCA procedure, the data were carefully inspected on a trial-by-trial
basis, and all other artifacts were manually removed on monopolar recordings. Monopolar
recordings for correct trials were then averaged to EMG onset, and a Laplacian transformation
(approximating the current source density) was implemented in BrainVision Analyzer (Cerveau
Products™, Munich) and applied to each participant’s average (as in Riè s et al., 2011,
2015, 2020; Riè s, Janssen, et coll., 2013; Riè s, Xie, et coll., 2013). The main advantage of using
Laplacian transformation is that it is reference-free, and it improves the spatial definition as it
enhances the separation of EEG components on the scalp, providing a good estimation of the
corticogram (Nunez & Srinivasan, 2006). The baseline for EMG-locked averages was taken
depuis 500 ms until 300 ms before EMG onset although the measures we used for analyses were
independent from the baseline choice.
Neurobiology of Language
613
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
un
_
0
0
0
5
6
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
Analysis
Statistical analyses were performed within R version 3.6.3 (R Core Team, 2020).
Behavioral data
We used the package lme4 to compute generalized linear (for reaction times) and logistic (pour
accuracy rates) mixed-effects models (Baayen et al., 2008; Jaeger, 2008), which rely on single-
trial data rather than on averages over participants or items, and are also free from the assump-
tions of homogenous variance and sphericity that are inherent to the more classic ANOVA
(Pinheiro & Bates, 2000). The individual reaction times were inversed to reduce skewness
and approach a normal distribution. The analyses were performed on inversed reaction times
and accuracy. We tested for a fixed effect of Condition (Congruent vs. Incongruent in the flanker
task, and Identity vs. False Cognate vs. Unrelated in the PWM task) and controlled for random
effects of picture name and participant, as well as by-item and by-participant random slopes for
Condition. The p values were obtained using type-II analyses of deviance tables providing Wald
chi-square ( Wald χ2) tests and associated p values for the fixed effects in the generalized linear
mixed-effects models, using the R package car (Fox & Weisberg, 2011). For all models, nous
report Wald χ2 values and p values from the analysis of deviance tables as well as raw
beta estimates (βraw), 95% confidence intervals around these beta estimates, standard errors,
t values for reaction times, Wald z values for accuracy rates, and associated p values.
Enfin, we calculated Spearman correlation coefficients to examine a possible relation
between congruency effects on flanker and PWM tasks on average reaction times and on
error rates.
EEG data
We focused our analysis on the Laplacian-transformed EEG components time-locked to EMG
onset on two sets of recoding sites: (1) the medial-frontal electrode, FCz, previously associated
with cognitive control mechanisms in non-linguistic tasks (Vidal et al., 2011), et (2) the left
frontal electrodes including F3, F7, AF3, and AF7, given activity seen on the grand averages at
these recording sites. Even though our previous study found activity at FC5 (Riè s, Janssen, et coll.,
2013), no activity was visible at this recording site on the grand averages in the present study. Dans
order to assess the presence of an activity at these recording sites, we first compared the slope of
the waveforms to zero (as in Riès et al., 2011, 2020; Riè s, Janssen, et coll., 2013; Riè s, Xie, et coll.,
2013). Alors, in order to assess whether any effects were present on the signal recorded at these
électrodes, we calculated the peak-to-peak amplitudes and the latencies of the peaks of interest
of the observed activities. These measures are known to be independent from the baseline. Ce
analysis was performed on activities present within 500 ms prior to EMG onset. Peak latencies
and peak-to-peak amplitude measures were defined as follows: The latencies of the peaks were
measured on smoothed data, using a 40-ms-long sliding smoothing window to minimize the
impact of background noise, on the grand average activities in the False Cognate and Incon-
gruent conditions respectively as these conditions yielded the largest components on the grand
averages. The surfaces between the x axis and the peaks of interest were calculated on 40-ms
time windows around these latencies on the non-smoothed data for all conditions per partici-
pant. Enfin, the peak-to-peak amplitude was calculated by finding the difference between the
positive and negative surfaces (as in Riè s et al., 2011, 2020; Riè s, Janssen, et coll., 2013; Riè s, Xie,
et coll., 2013). These measures were calculated for electrodes F3 (the only left frontal electrode
showing significant activity on the slope analyses) and FCz in the PWM and flanker tasks. All
measures were compared using two-tailed paired Student’s t tests.
Neurobiology of Language
614
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
un
_
0
0
0
5
6
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
RÉSULTATS
We present the results for the flanker task first given our aim to compare cognitive control
processes supporting language to cognitive control processes engaged outside of language.
Behavioral Results
Flanker task
The mean RTs, accuracy rates, and standard deviations are presented per condition in
Table 1A. There was a significant effect of condition on reaction times ( Wald χ2(1) = 141.35,
p < 0.001). Reaction times were longer in the Incongruent condition than in the Congruent
condition (βraw = −2.13 × 10−4, 95% CI [−2.48 × 10−4, −1.78 × 10−4], SE = 1.79 × 10−5, t =
−11.89, p < 0.001). There was also a significant effect of condition on accuracy ( Wald χ2(2) =
36.25, p < 0.001). The accuracy rate was lower in the Incongruent condition than in the
Congruent condition (βraw = −1.45, 95% CI [−1.92, −0.98], SE = 0.241, z = −6.02, p < 0.001).
Picture-word matching task
The mean RTs, accuracy rates, and standard deviations are presented per condition in Table 1B.
There was a significant effect of condition on reaction times ( Wald χ2(2) = 15.13, p < 0.001).
Reaction times were longer in the False Cognate condition than in the Identity condition (βraw =
−1.58 × 10−5, 95% CI [−4.10 × 10−5, 9.42 × 10−6], SE = 1.29 × 10−5, t = −1.23), and longer
in the False Cognate condition than in the Unrelated condition (βraw = −1.75 × 10−5, 95%
CI [−3.25 × 10−5, −2.53 × 10−6], SE = 7.64 × 10−6, t = −2.29). There was also a significant
effect of condition on accuracy rates ( Wald χ2(2) = 14.32, p < 0.001). The accuracy was lower in
the False Cognate condition than in the Unrelated condition (βraw = −0.86, 95% CI [−1.31,
−0.41], SE = 0.23, z = −3.76, p < 0.001), and higher in the False Cognate than in the Identity
condition (βraw = 0.58, 95% CI = [0.11, 1.05], SE = 0.24, z = 2.44, p = 0.015). We note, however,
that the median error rates were smaller than 5% across conditions (see Table 1).
Flanker versus PWM congruency effect comparison
There was no significant correlation between the size of the congruency effect in the flanker
(Incongruent − Congruent) versus the PWM (False Cognate − Identity) tasks on reaction times
(rho = −0.14, S = 518, p = 0.638) or accuracy rates (rho = −0.41, S = 643, p = 0.142). In
Table 1. Mean reaction time and accuracy rate per condition and per task with standard deviations
around the means in parenthesis.
A. Flanker task
Congruent
Incongruent
Mean reaction time
394 ms (SD = 55 ms)
Mean accuracy rate
99% (SD = 2%)
466 ms (SD = 51 ms)
82% (SD = 16%)
B. Picture-word matching task
Mean reaction time
Mean accuracy rate
Identity
False-cognate
Unrelated
715 ms (SD = 104 ms)
747 ms (SD = 97 ms)
714 ms (SD = 84 ms)
94% (SD = 8%)
94% (SD = 7%)
98% (SD = 3%)
615
Neurobiology of Language
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
a
_
0
0
0
5
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
addition, the random slopes of the corresponding condition effects per participant extracted
from the mixed effect models were not correlated (RTs: rho = −0.068, S = 486, p = 0.820;
accuracy rates: rho = −0.138, S = 518, p = 0.638).
Electrophysiological Results
EEG results in the flanker task
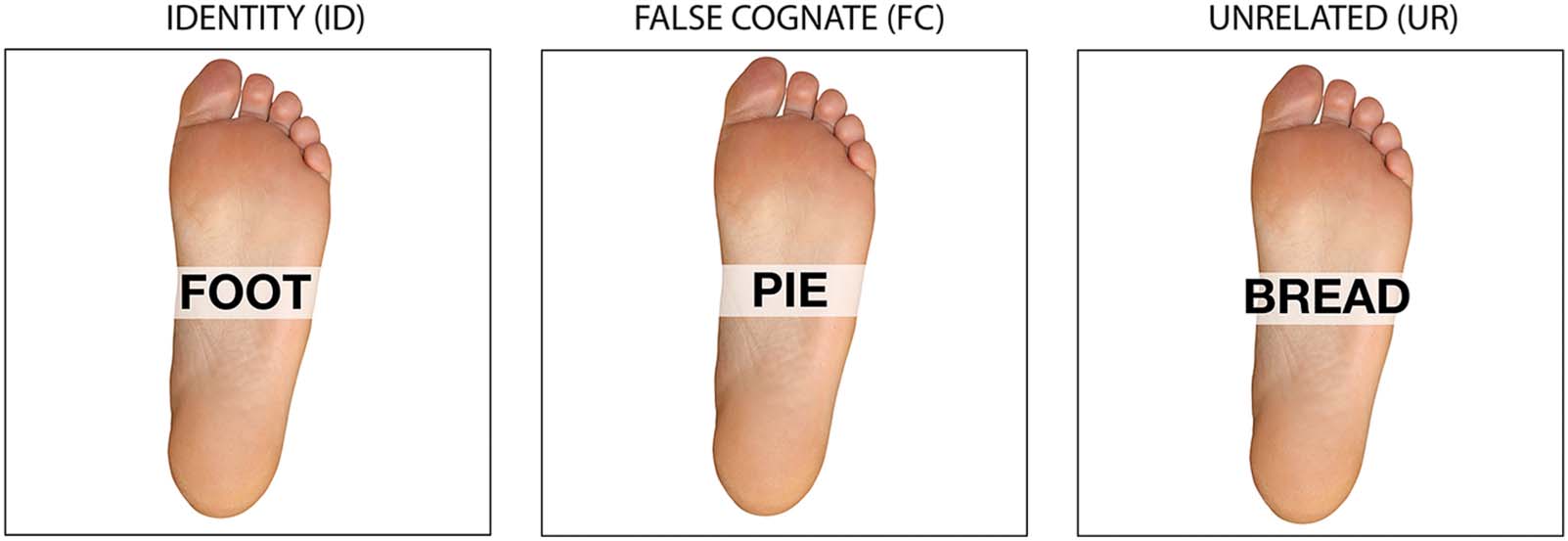
We observed a negativity at the fronto-central recording site, FCz, peaking on average 52 ms
(SD = 32 ms) before EMG onset in incongruent trials (Figure 2A). The slope of this negativity
was significantly different from zero between 150 and 50 ms before EMG onset in incongruent
trials (t (13) = − 2.51, p = 0.026), replicating the N-40 component described in prior studies
(e.g., Vidal et al., 2011). The peak-to-peak amplitude of this negativity was significantly larger
in incongruent than in congruent trials (t (13) = 3.58, p = 0.003, with measures taken around
the peak latencies in the Incongruent condition). Interestingly, in congruent trials, there was no
negative component peaking around 40 ms prior to EMG onset. Indeed, the slope was not
different from zero between 150 and 50 ms before EMG onset in congruent trials (t (13) =
1.07, p = 0.305), indicating that the N-40 component was sensitive to congruency manipulation.
There was a later negativity in congruent trials peaking on average 4 ms (SD = 32 ms)
before EMG onset (the slope of the negativity was significantly different from zero between
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
a
_
0
0
0
5
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Flanker task EEG results. (A) Medial frontal activity in the flanker task. On the left, waveforms of the medial frontal activity at FCz in
the Congruent (blue) and Incongruent (red) conditions time-locked to EMG onset (in ms). On the right, topographies of the medial frontal
activity in each condition on a 50-ms time window centered around the peak latency in the Incongruent condition. (B) Absence of the left
anterior frontal activity at F3; waveforms are shown on the left and on the right are shown the topographies on a 50-ms time window centered
around the peak latency in the False Cognate condition in the picture-word matching task.
Neurobiology of Language
616
Bilingual cognitive control as revealed by Laplacian transformed ERPs
100 before EMG onset and EMG onset, t (13) = −2.92, p = 0.012). However, the peak latency
of this later negativity appears incompatible with the N-40 given that the N-40 typically peaks
earlier (i.e., around 40 ms before EMG onset; e.g., Vidal et al., 2003). Of note, this later neg-
ativity was smaller in amplitude than the negativity peaking around 50 ms before EMG onset
in incongruent trials (t (13) = 2.44, p = 0.030).
Following EMG onset, a negativity peaking around 100 ms post-EMG onset is visible in
both conditions, which likely corresponds to the ERN previously shown to be present in
correct trials as well as in incorrect trials, only of larger amplitude in incorrect trials (Vidal
et al., 2000, 2003; Roger et al., 2010). However, the ERN was not the focus of the present
study, hence no analyses were performed for this component.
The topography in Figure 2 indicates the fronto-central activity may also be present at Fz,
anterior to FCz. We therefore also analyzed the activity recorded at Fz (see Figure S1) and
found that the slope of the negativity at Fz between 150 and 50 ms before EMG onset was
significantly different from zero in the Incongruent condition (t (13) = −4.62, p < 0.001), but
not in the Congruent condition (t (13) = −0.96, p = 0.353). There was a significant difference
between the slope of the negativity in incongruent versus congruent trials (t (13) = −3.89, p =
0.002; it was more negative-going in the Incongruent compared to the Congruent condition),
but there was no significant difference between conditions for the peak-to-peak amplitude
(t (13) = 2.05, p = 0.062).
There was no left anterior component at electrode F3 in the flanker task (Figure 2B). The
slope of the waveforms was not significantly different from zero between 300 and 200 ms
before EMG onset in the Congruent or Incongruent conditions (t (13) < 1). There was also
no amplitude difference between the two waveforms (t (13) < 1; surface measures were taken
between 200 and 100 ms before EMG onset).
EEG results in the PWM task
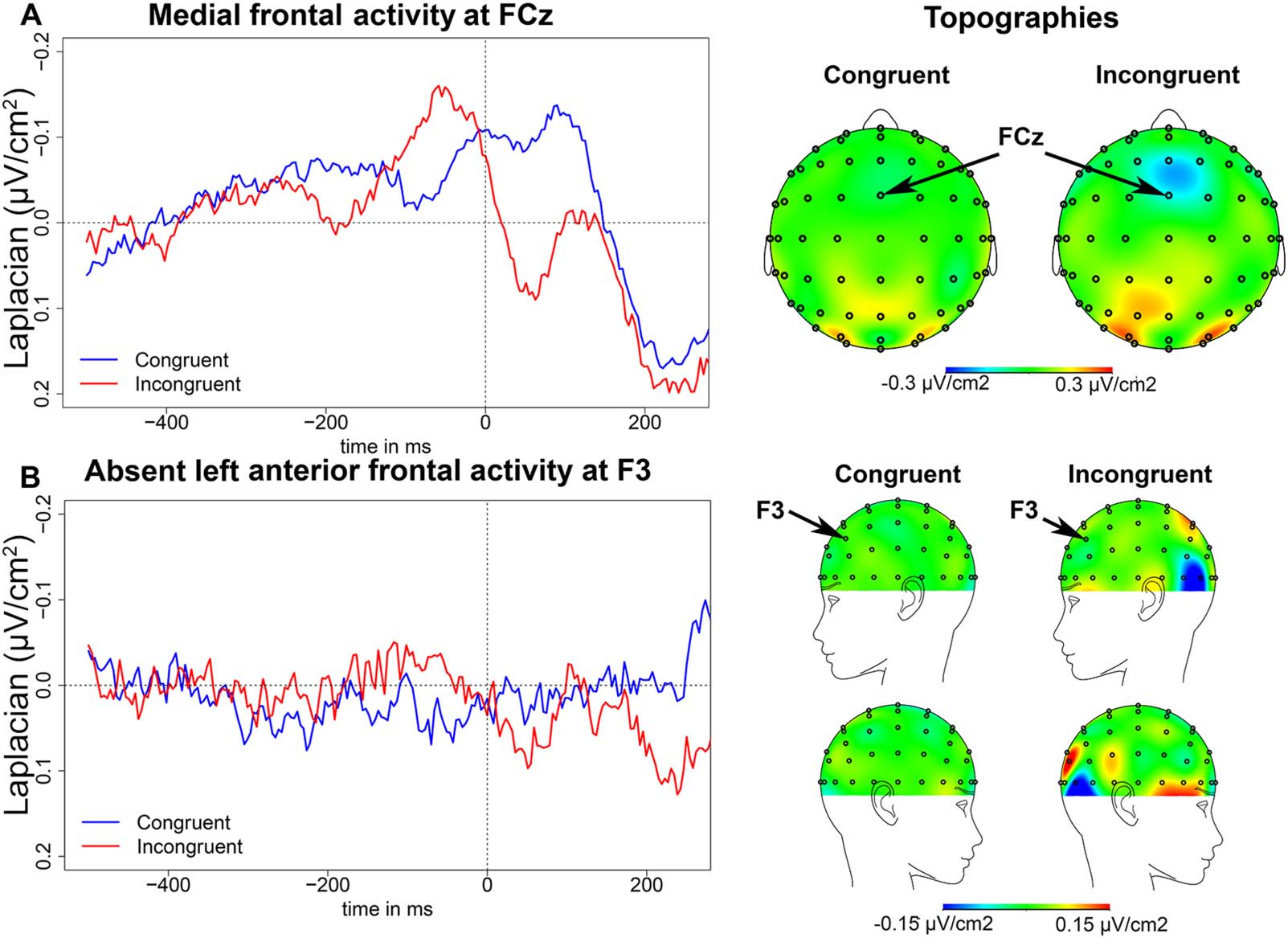
As in the flanker task, we observed negativities at the fronto-central recording site, FCz,
peaking around 150 ms before EMG onset in the PWM task. For a parallel comparison with
the flanker task, we first compare the False Cognate to the Identity condition and then describe
the results of the Unrelated condition separately.
The negativity peaked on average 191 ms (SD = 79 ms) before EMG onset in the Identity
condition, and 137 ms (SD = 69 ms) before EMG onset in the False Cognate condition
(Figure 3A). There was an effect of condition on the latency of the peak (t (13) = 2.53, p =
0.025), as the negativity peaked later in the False Cognate condition than in the Identity con-
dition. The slope of this negativity between 300 and 200 ms before EMG onset was signifi-
cantly different from zero in the False Cognate condition (t (13) = −2.60, p = 0.022), but not
in the Identity condition (t (13) = −1.45, p = 0.172). However, there was no significant differ-
ence between conditions on the slope of the negativity (t (13) = −1.25, p = 0.232). Finally, the
peak-to-peak amplitude was larger in the False Cognate condition than in the Identity condi-
tion (t (13) = 2.47, p = 0.028). The topographies in Figure 3 indicate the fronto-central activity
was well-centered at FCz.
In the Unrelated condition, the negativity peaked on average 160 ms (SD = 60 ms) before
EMG onset, and the slope of this negativity between 300 and 200 ms before EMG onset was
significantly different from zero (t (13) = 4.54, p < 0.001). Finally, as expected, the negativity
was larger in the False Cognate condition than in the Unrelated condition (t (13) = 2.48, p =
0.027), but not larger in the Unrelated than in the Identity condition (t (13) < 1).
Neurobiology of Language
617
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
a
_
0
0
0
5
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
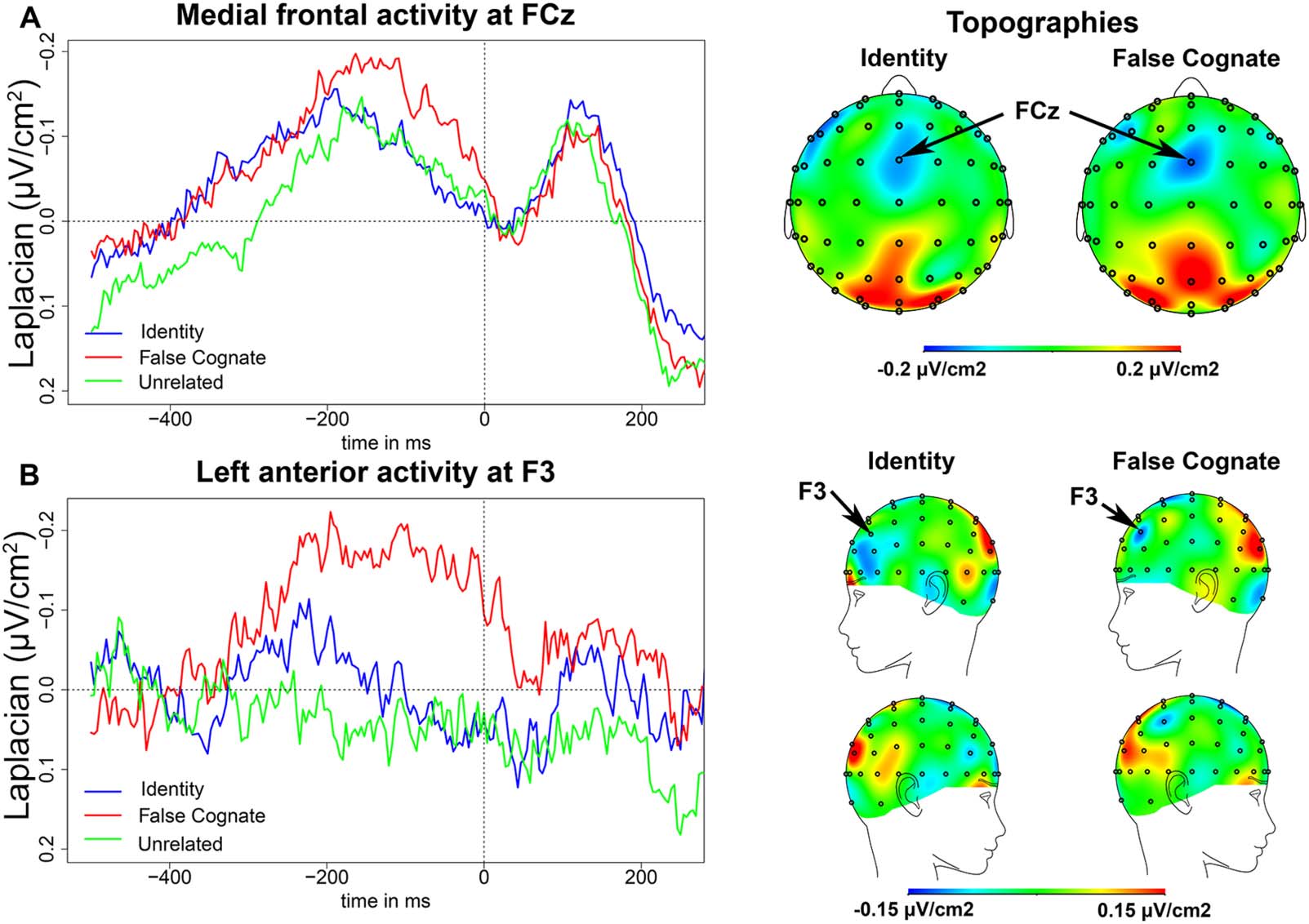
Figure 3. Picture-word matching task EEG results. (A) Medial frontal activity in the picture-word matching task. On the left, waveforms of the
medial frontal activity at FCz in the Identity (blue), False Cognate (red), and Unrelated (green) conditions time-locked to EMG onset (in ms). On
the right, topographies of the medial frontal activity in the Identity and False Cognate conditions on a 50-ms time window centered around the
peak latency in each condition. (B) Left anterior frontal activity in the picture-word matching task. On the left, waveforms of the left anterior
frontal activity at F3 in the Identity (blue), False Cognate (red), and Unrelated (green) conditions time-locked to EMG onset (in ms). On the
right, topographies of the left anterior frontal activity in the Identity and False Cognate conditions on a 50-ms time window centered around the
peak latency in each condition.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
a
_
0
0
0
5
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
A left-lateralized component, at electrode F3, was found in the PWM task (Figure 3B). This
negativity peaked on average 190 ms (SD = 37 ms) before EMG onset in the False Cognate
condition and on average 232 ms (SD = 26 ms) before EMG onset in the Identity condition.
There was an effect of condition on the latency of the peak (t (13) = 3.91, p = 0.002), as the
negativity peaked later in the False Cognate condition than in the Identity condition. The slope
of this negativity between 300 and 200 ms before EMG onset was significantly different from
zero in the False Cognate condition (t (13) = −2.51, p = 0.026), but not in the Identity condition
(t (13) = −0.29, p = 0.218). However, there was no significant difference between conditions
on the slope of the negativity (t (13) = −1.57, p = 0.140). Finally, there was no significant dif-
ference between the peak-to-peak amplitude in the False Cognate condition and the Identity
condition (t (13) = 1.00, p = 0.334).
In the Unrelated condition, there was no apparent negativity at electrode F3; the slope of
the EEG waveform was not significantly different from zero between 300 and 200 ms before
EMG onset (t (13) < 1). The topographies in Figure 3 indicate that the left frontal activity may be
more anterior and inferior in the Identity condition compared to the False Cognate condition.
We therefore also examined the activity recorded at F5, F7, AF3, and AF7 (see Figure S2) and
found that the slope of the waveforms at these electrodes was not significantly different from
zero in any of the conditions under analysis (all t s were between 0.28 and −1.09, except for
Neurobiology of Language
618
Bilingual cognitive control as revealed by Laplacian transformed ERPs
F5 in the Identity condition: t (13) = −1.77, p = 0.099, and F7 in the Identity condition: t (13) =
−1.88, p = 0.082).
DISCUSSION
This study’s aim was to compare frontal cognitive control processes as engaged in linguistic
versus non-linguistic decision-making in Spanish-English bilinguals using a PWM task involving
false cognates and the arrow version of the Eriksen flanker task. Behavioral results showed that
congruency effects were present in both the linguistic and non-linguistic tasks, but that the size of
the congruency effects in the two tasks was not significantly correlated across participants. EEG
results revealed a medial frontal component corresponding to the N-40 in the non-linguistic task.
Its amplitude was modulated by congruency. A similar potential, although peaking earlier,
around 150 ms before EMG onset, was found at the same electrode in the linguistic task and
was modulated by linguistic congruency. Moreover, a left frontal potential was found in the
linguistic task peaking around 200 ms before EMG onset and was sensitive to congruency. This
component was absent in the non-linguistic task. These findings suggest a partial overlap
between the control networks underlying the non-linguistic and linguistic tasks.
Behavioral Results
Although the expected congruency effects were present in both the linguistic and non-
linguistic tasks, the reaction times and the size of the congruency effects were different in
the two tasks. The reaction times were shorter in the flanker task (between 394 and 466 ms
on average) than in the PWM task (between 714 and 747 ms on average), but the congruency
effect was larger in the flanker task (a 70 ms difference and 17% less accurate on average in
the Incongruent compared to the Congruent condition) than in the PWM task (32 ms difference
and no difference in accuracy on average in the Incongruent, i.e., False Cognate, compared to
the Congruent, i.e., Identity, condition). The size of the congruency effect is on par with what
was reported in the original study using the arrow version of the flanker task (74 ms in Stoffels
& van der Molen, 1988), and the reaction times were in the same general order (between 350
and 450 ms). The size of the cross-linguistic interference effect is more variable in the literature
varying between over 100 ms to less than 10 ms (van Heuven et al., 2008; Vanlangendonck
et al., 2019; von Studnitz & Green, 2002); however, the tasks used in these studies were
different than in ours and often used lexical decision to assess cross-linguistic interference
rather than picture-word matching. In addition, interference resolution abilities have been
shown to be modulated by language proficiency and dominance in bilinguals (Robinson
Anthony & Blumenfeld, 2019), which may be another factor underlying the variability in
the size of the cross-linguistic interference effect across studies. In a recent review, Calabria
et al. (2018) note that how control is exercised is likely task-dependent and depends on bilin-
gual profiles, calling for continued research into the exact manner in which bilingual control
and executive function networks are linked in particular contexts.
Possible reasons why reaction times are longer overall in the PWM task could be linked to
the fact that there are more possible response alternatives in this task overall (i.e., more than
one possible name for a picture, and co-activation of linguistic representations that are related
but not directly relevant to the experimental context), whereas there are only two possible
responses in the flanker task. In addition, arrows are highly learned symbols and have very
strong associations with their corresponding directions (Ridderinkhof et al., 2020). The asso-
ciations between pictures and their names are generally not as high. Finally, it is possible that,
as the central target arrow on the flanker task is spatially separated from its flankers, it was
Neurobiology of Language
619
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
a
_
0
0
0
5
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
easier for participants to focus on the central target arrow and ignore flankers during flanker
task performance than to focus on the picture and ignore the superimposed word during the
PWM task.
Concerning the difference in the size of the congruency effect, one reason could be that in
the flanker task, the alternative response is highly salient as the central arrow is flanked by four
arrows going in the opposite direction in the incongruent trials. In addition, the association
between these flanking arrows pointing in the direction opposite to the central arrow and
the associated response is very strong, making the overcoming of this interference in response
selection difficult. In contrast, in the PWM task, there is only one stimulus calling for the alter-
native response on the screen. In fluent readers, the superimposed text that cues the alternative
response also yields an automatic reading response; however, this response is likely not as
tightly associated with verbal output as the arrows in the flanker task are associated with right /
left motor responses. Relatedly, bilinguals need to inhibit the other language constantly in
everyday life and therefore likely have more practice in overcoming cross-linguistic interfer-
ence associated with the superimposed text on the PWM task than non-linguistic interference
as assessed in the flanker task.
Medial Frontal Component
In the flanker task, we observed longer reaction times and lower accuracy rates in the Incon-
gruent compared to the Congruent condition, replicating the well-established congruency
effect in the arrow version of the Eriksen flanker task (Stoffels & van der Molen, 1988; for a
review, see Ridderinkhof et al., 2020). In addition, our EEG results revealed a negative
component peaking around 50 ms before EMG onset in the Incongruent condition. In the
Congruent condition, there was no significant component peaking around the same latency,
instead there was a later negativity peaking right around EMG onset. Previous reports have
described a negative component peaking around 40 ms prior to EMG onset, at the same
fronto-central recording site as in our study, in tasks requiring a choice between two possible
responses (Carbonnell et al., 2004; Vidal et al., 2003, 2011). This component is thought to
originate from the medial frontal cortex, including the SMAs (Vidal et al., 2003), and develops
prior to the activation of the primary motor areas in tasks involving a choice to make between
different possible manual responses (Carbonnell et al., 2004; Vidal et al., 2003). This compo-
nent was also found to be modulated in amplitude by congruency in the Eriksen flanker task
(Roger, 2009) and was found to be reduced in amplitude in situations when information about
the response to be produced was available to the participant ahead of stimulus presentation
(Carbonnell et al., 2004).
Our results are therefore largely in agreement with these findings, even if we did not find a
negativity peaking at around the same latency in congruent trials. We note, however, that in
contrast with the Incongruent condition, the slope of the waveform in the Congruent condition
reported in Roger (2009) was also not different from zero between 90 and 60 ms pre-EMG
onset, even though it was significantly different from zero in the later time window, between
60 and 30 ms pre-EMG onset. This is compatible with our observations even if the negativity
peaked at around the same latency in congruent and incongruent trials in Roger (2009).
Moreover, other studies investigating this component have reported an absence of negativity
in the Congruent or easier condition, contrasting with the presence of a negativity peaking
around 40 ms before EMG onset in the Incongruent or more difficult condition (Experiment 1,
Roger, 2009; Carbonnell et al., 2004). Therefore, our results fall within the range of expected
findings based on the literature.
Neurobiology of Language
620
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
a
_
0
0
0
5
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
In the linguistic task, we observed the expected cross-linguistic interference effect on the
behavioral results. Performance was worse in the False Cognate condition than in the Unrelated
and Identity conditions, as in other studies (e.g., van Heuven et al., 2008; Vanlangendonck et al.,
2019; von Studnitz & Green, 2002). EEG results revealed a fronto-central negative component at
the same recording site as in the non-linguistic task (FCz) peaking around 160 ms before EMG
onset in the Unrelated condition. This component peaked around 30 ms earlier in the Identity
condition, and 20 ms later in the False Cognate condition. The peak latency of ERP components
has been reported to be modulated by factors such as task difficulty, age, and stimulus-onset
asynchrony (e.g., Eddy & Holcomb, 2010; Kutas & Federmeier, 2011; Picton, 1992). The fact
the negativity peaked later in the most difficult condition and earlier in the easiest condition
is therefore not surprising. This is also in agreement with the results of Rodriguez-Fornells
et al. (2006). This study found increased amplitude for the fronto-central negativity in the
cross-linguistic phonological interference condition in both go and no-go trials, but also found
a delayed peak latency for the same component in the no-go trials, requiring enhanced inhib-
itory control, compared to the go trials. Similarly, as in the non-linguistic task, the amplitude of
this negativity was larger in the False Cognate condition (analogous to the Incongruent condition
in the non-linguistic task) than in the Identity condition (analogous to the Congruent condition
in the non-linguistic task). In addition, the amplitude of this component was larger in the False
Cognate condition than in the Unrelated condition.
As mentioned in the Introduction, previous studies have found that bilinguals engage medial
frontal regions typically associated with executive control, including the ACC and the pre-SMA,
when faced with cross-linguistic interference (e.g., Rodriguez-Fornells et al., 2005; van Heuven
et al., 2008). For example, van Heuven et al.’s (2008) neuroimaging findings suggested that
pre-SMA / ACC areas showed greater activity for Dutch-English interlingual homographs than
for control stimuli in a lexical decision task that included response-level conflict. However, we
note that similar medial frontal activity peaking around 250 ms before vocal onset was also
previously reported in picture naming, without manual responses, using the same techniques
and source modeling as in the current study (Riè s, Janssen, et al., 2013), and using magneto-
encephalography (Salmelin et al., 1994), even though these studies did not specifically target
bilinguals and did not manipulate cross-linguistic interference. That these activities peaked
earlier than in the present study may be due to task difficulty as these studies used a simple
picture naming task with no overlapping distractors. In addition, activity in the pre-SMA and
SMA has also been reported preceding vocal onset in language production fMRI studies not
targeting bilinguals (e.g., Alario et al., 2006; Tremblay & Gracco, 2010). Moreover, high fre-
quency repetitive transcranial magnetic stimulation of the pre-SMA has been shown to interfere
with the volitional selection of words and oral gestures (Tremblay & Gracco, 2009). As has been
previously proposed (e.g., Tremblay and Gracco, 2009; Riè s, Janssen, et al., 2013), our results
suggest that the medial frontal cortex is active when a choice has to be made, whether it be
linguistic or not, and that the amplitude of this activity is modulated by the difficulty of response
selection, suggesting the brain mechanism underlying the medial frontal activity is shared across
the linguistic and non-linguistic domains. In bilinguals, this medial frontal activity seems to be
similarly engaged in linguistic and non-linguistic response selection, suggesting a functional and
anatomical overlap between the two domains. However, we do not believe that the engagement
of this medial frontal activity in language is unique to bilinguals, but instead is present across all
speakers and sensitive to response selection difficulty across domains. The current data suggest
that in bilinguals activity in this area correlated with interference resolution across languages in
addition to the other response selection contexts already documented in monolinguals. The
results of our study do not allow us to say whether or not this medial frontal activity is larger
in bilinguals than monolinguals as we did not include a monolingual group.
Neurobiology of Language
621
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
a
_
0
0
0
5
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
Left Anterior Frontal Component
A left anterior component was also observed at electrode F3 in the linguistic task. This com-
ponent peaked later in the False Cognate condition than in the Identity condition (on average
190 ms before EMG onset in the False Cognate condition and 232 ms before EMG onset in the
Identity condition). This effect of cross-linguistic interference on peak latency is similar to the
one observed on the medial frontal component at FCz. Although there was no significant dif-
ference in amplitude between the False Cognate and Identity conditions, the slope of the neg-
ativity was significantly different from zero only in the False Cognate condition. Interestingly,
this component was found at a more anterior site than in a previous picture naming study (at
FC5 in Riè s, Janssen, et al., 2013). In addition, the left anterior frontal activity in the present
study peaked earlier than in Riè s, Janssen, et al. (2013), where it peaked right around vocal
onset, and was therefore interpreted as being associated with response preparation. The fact
that the activity we report here peaked earlier and was sensitive to cross-linguistic interference
suggests it is associated with process(es) preceding response preparation, such as response
selection or cognitive control processes helping to resolve cross-linguistic interference. This
could be consistent with van Heuven et al.’s (2008) finding of activation in anterior left inferior
prefontal cortex linked to stimulus-based conflict resolution in cross-linguistic false cognates,
even though the spatial resolution of EEG is limited even after Laplacian transformation and
hence prevents us from making definitive claims regarding the source of the component
we observed. Our results are also more generally in agreement with a role of the left PFC in
resolving cross-linguistic interference, as previously proposed (Abutalebi & Green, 2007).
This left-lateralized component was absent in the non-linguistic task, suggesting the under-
lying brain activity may be specific to the linguistic task. We aimed to align our linguistic PWM
and non-linguistic flanker tasks in terms of loci of conflict, with both stimulus-based and
response-based conflict expected to be present in both tasks. However, it is possible that
the stimulus-based conflict that arose in the linguistic task was more extensive. The longer
reaction times in the linguistic task are in agreement with the fact that a wider array of repre-
sentations was accessed during response selection in the linguistic task. In the non-linguistic
task, there were only two possible stimulus dimensions that could receive activation, those
corresponding to the left and right arrows. In the linguistic task, however, many different word
representations likely received activation from the picture to be identified and from the over-
lapping distractor word, and the availability of multiple conceptual representations likely inter-
fered with selection. For example, a picture of a foot with the word PIE overlaid is likely to
activate the linguistic representations for foot in both English and Spanish, as well as the baked
treat pie in English. In addition, adjacent phonological and semantic representations may be
activated in both languages (e.g., Shook & Marian, 2013; van Heuven & Dijkstra, 2010).
A related explanation for the presence of the left anterior frontal component in the linguistic
but not the non-linguistic task could be linked to the rostro-caudal organization of cognitive
control functions in the frontal cortex (Badre & D’Esposito, 2009; Badre & Nee, 2018), with
more abstract or higher order rules being encoded in the more anterior regions, and more
concrete or lower order rules being encoded in more posterior regions such as the premotor
cortices. Indeed, the PWM task engages linguistic word retrieval and phonological matching
processes leading to selecting the response corresponding to a match or a non-match between
the picture and the overlapping word. The flanker task instead engages more simple stimulus-
to-response mapping where only two possible responses can be activated, selected, and exe-
cuted, without any complex matching process between the stimulus and the response. We
must, however, remain cautious when interpreting spatial localization differences in our study
given the low spatial resolution of EEG, even if we did use Laplacian transformation. This result
Neurobiology of Language
622
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
a
_
0
0
0
5
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
also suggests that additional cognitive control may be engaged in the linguistic task as com-
pared to the non-linguistic task, therefore suggesting that the functional overlap between the
cognitive control networks engaged in language and outside of language in bilinguals is only
partial, at least as revealed by the tasks we used. This could be linked to inherent differences
between linguistic and non-linguistic decision-making processes or to differences between the
tasks used. Indeed, our results differ from those of Coderre et al. (2016), who contrasted a
semantic categorization task to a flanker task and found overlap in the left inferior frontal gyrus
in bilinguals. Our results also differ from those of De Baene et al. (2015), who found that highly
similar brain regions, including the lateral and medial PFC, were engaged in linguistic and
non-linguistic switching using closely matched linguistic and non-linguistic tasks. In this study,
the linguistic task consisted in naming pictures in different languages as cued by a preceding
symbol. Importantly, there were no overlapping written distractor words on the picture. In
closer alignment to our results, a magnetoencephalography study that was not specifically tar-
geting bilinguals found left superior frontal activity between 350 and 650 ms post-stimulus
onset was found to be larger in the semantically related compared to the unrelated condition
on a picture-word interference paradigm (Piai et al., 2014). This activity was interpreted as
reflecting lexical competition resolution. The left anterior frontal activity we report here fol-
lows a similar time course even if we studied it time-locked to the response and not the stim-
ulus. Given that we also used a picture-word interference paradigm, our left anterior frontal
activity may reflect a similar competition resolution mechanism, which would be needed only
in the linguistic and not in the non-linguistic task.
Limitation of the Current Study
The relatively low number of participants included in the final analyses (n = 14) is lower than
what is typically recommended in ERP studies examining potential cognitive consequences of
bilingualism (Cespón & Carreiras, 2020). We, however, note that the number of participants
needed may be dependent on the type of signal processing and analysis performed. Previous
studies of response selection components using EEG and Laplacian transformation have shown
reliable results using a similar or even smaller number of participants (n = 12 in Carbonnell
et al., 2004; Riès, Janssen, et al., 2013; Vidal et al., 2003, 2011). Nevertheless, increasing the
number of participants would be ideal in future studies and could clarify the results reported
here (e.g., the nonsignificant difference in amplitude between conditions in the linguistic task
for the left frontal component). Therefore, the results of the present study should be considered
as preliminary given that Laplacian transformation has not been broadly used before in the
context of research on cognitive underpinnings of bilingualism.
Conclusions
To conclude, our study indicates that in bilinguals medial frontal activity preceding response
execution is engaged in cross-linguistic and non-linguistic cognitive control associated with
response selection at different time points, further supporting shared neural implementation
but also suggesting shared computational principles between linguistic and non-linguistic
domains at the level of response selection. An additional left anterior frontal component
sensitive to cross-linguistic interference was also present in the linguistic but not in the non-
linguistic task. Our results therefore suggest a partial functional overlap between linguistic and
non-linguistic cognitive control processes as engaged in the tasks we used, and that linguistic
conflict resolution may engage additional left anterior frontal control processes in parallel with
more domain-general response selection processes in the medial frontal cortex. These findings
align with the notion that when bilinguals resolve interference across languages, they engage a
Neurobiology of Language
623
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
a
_
0
0
0
5
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
neural network that is in part domain-general. It has been argued that such an overlap between
linguistic and non-linguistic networks is the basis for a mechanism that may drive changes in
cognitive control associated with bilingualism. Specifically, the engagement of non-linguistic
networks during bilingual processing may yield further use and strengthening of such networks
(e.g., Bialystok & Craik, 2010).
ACKNOWLEDGMENTS
This work was supported by a grant from the McNair Scholars Program awarded to Martha N.
Mendoza; NIDCD grant 1R21DC016985 to Stephanie K. Ries; and NINDS grant
5R37NS021135 to Robert T. Knight. (The content is solely the responsibility of the authors
and does not necessarily represent the official views of the National Institutes of Health.)
The authors would like to thank the participants for their involvement in this study.
FUNDING INFORMATION
Stephanie Ries, National Institute on Deafness and Other Communication Disorders (https://dx
.doi.org/10.13039/100000055), Award ID: 1R21DC016985. Robert T. Knight, National Insti-
tute of Neurological Disorders and Stroke (https://dx.doi.org/10.13039/100000065), Award ID:
2R37NS21135.
AUTHOR CONTRIBUTIONS
Martha N. Mendoza: Conceptualization: Equal; Data curation: Lead; Formal analysis: Support-
ing; Writing – review & editing: Equal. Henrike K. Blumenfeld: Investigation: Supporting;
Writing – original draft: Equal; Writing – review & editing: Supporting. Robert T. Knight: Funding
acquisition: Lead; Project administration: Supporting; Resources: Lead; Supervision: Supporting;
Writing – original draft: Supporting; Writing – review & editing: Supporting. Stephanie K. Ries:
Conceptualization: Lead; Formal analysis: Lead; Funding acquisition: Supporting; Investigation:
Equal; Methodology: Lead; Project administration: Lead; Resources: Equal; Supervision: Lead;
Validation: Equal; Visualization: Equal; Writing – original draft: Equal; Writing – review &
editing: Equal.
REFERENCES
Abutalebi, J., & Green, D. (2007). Bilingual language production:
The neurocognition of language representation and control. Jour-
nal of Neurolinguistics, 20(3), 242–275. https://doi.org/10.1016/j
.jneuroling.2006.10.003
Alario, F.-X., Chainay, H., Lehericy, S., & Cohen, L. (2006). The role
of the supplementary motor area (SMA) in word production.
Brain Research, 1076(1), 129–143. https://doi.org/10.1016/j
.brainres.2005.11.104, PubMed: 16480694
Baayen, R. H., Davidson, D. J., & Bates, D. M. (2008). Mixed-
effects modeling with crossed random effects for subjects and
items. Journal of Memory and Language, 59(4), 390–412.
https://doi.org/10.1016/j.jml.2007.12.005
Babiloni, F., Cincotti, F., Carducci, F., Rossini, P. M., & Babiloni, C.
(2001). Spatial enhancement of EEG data by surface Laplacian
estimation: The use of magnetic resonance imaging-based head
models. Clinical Neurophysiology, 112(5), 724–727. https://doi
.org/10.1016/s1388-2457(01)00494-1, PubMed: 11336885
Badre, D., & D’Esposito, M. (2009). Is the rostro-caudal axis of the
frontal lobe hierarchical? Nature Reviews Neuroscience, 10(9),
659–669. https://doi.org/10.1038/nrn2667, PubMed: 19672274
Badre, D., & Nee, D. E. (2018). Frontal cortex and the hierarchical
control of behavior. Trends in Cognitive Sciences, 22(2),
170–188. https://doi.org/10.1016/j.tics.2017.11.005, PubMed:
29229206
Bialystok, E., & Craik, F. I. M. (2010). Cognitive and linguistic
processing in the bilingual mind. Current Directions in Psycho-
logical Science, 19(1), 19–23. https://doi.org/10.1177
/0963721409358571
Bialystok, E., Craik, F. I. M., Klein, R., & Viswanathan, M. (2004).
Bilingualism, aging, and cognitive control: Evidence from the
Simon task. Psychology and Aging, 19(2), 290–303. https://doi
.org/10.1037/0882-7974.19.2.290, PubMed: 15222822
Bialystok, E., Craik, F., & Luk, G. (2008). Cognitive control and lex-
ical access in younger and older bilinguals. Journal of Experi-
mental Psychology: Learning, Memory, and Cognition, 34(4),
859–873. https://doi.org/10.1037/0278-7393.34.4.859,
PubMed: 18605874
Brodeur, M. B., Dionne-Dostie, E., Montreuil, T., & Lepage, M.
(2010). The Bank of Standardized Stimuli (BOSS), a new set of
480 normative photos of objects to be used as visual stimuli in
Neurobiology of Language
624
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
a
_
0
0
0
5
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
cognitive research. PLOS ONE, 5(5), Article e10773. https://doi
.org/10.1371/journal.pone.0010773, PubMed: 20532245
Burle, B., Spieser, L., Roger, C., Casini, L., Hasbroucq, T., & Vidal,
F. (2015). Spatial and temporal resolutions of EEG: Is it really
black and white? A scalp current density view. International Jour-
nal of Psychophysiology, 97(3), 210–220. https://doi.org/10.1016
/j.ijpsycho.2015.05.004, PubMed: 25979156
Calabria, M., Costa, A., Green, D. W., & Abutalebi, J. (2018).
Neural basis of bilingual language control. Annals of the
New York Academy of Sciences, 1426(1), 221–235. https://doi
.org/10.1111/nyas.13879, PubMed: 29917244
Carbonnell, L., Hasbroucq, T., Grapperon, J., & Vidal, F. (2004).
Response selection and motor areas: A behavioural and elec-
trophysiological study. Clinical Neurophysiology, 115(9),
2164–2174. https://doi.org/10.1016/j.clinph.2004.04.012,
PubMed: 15294220
Cespón, J., & Carreiras, M. (2020). Is there electrophysiological evi-
dence for a bilingual advantage in neural processes related to
executive functions? Neuroscience & Biobehavioral Reviews,
118, 315–330. https://doi.org/10.1016/j.neubiorev.2020.07.030,
PubMed: 32758515
Coderre, E. L., Smith, J. F., van Heuven, W. J. B., & Horwitz, B.
(2016). The functional overlap of executive control and language
processing in bilinguals. Bilingualism (Cambridge, England),
19(3), 471–488. https://doi.org/10.1017/S1366728915000188,
PubMed: 27695385
Colomé, À. (2001). Lexical activation in bilinguals’ speech produc-
tion: Language-specific or language-independent? Journal of
Memory and Language, 45(4), 721–736. https://doi.org/10.1006
/jmla.2001.2793
Costa, A., Cutler, A., & Sebastián-Gallés, N. (1998). Effects of
phoneme repertoire on phoneme decision. Perception & Psycho-
physics, 60(6), 1022–1031. https://doi.org/10.3758/BF03211936,
PubMed: 9718960
Costa, A., Hernandez, M., Costa-Faidella, J., & Sebastian-Galles, N.
(2009). On the bilingual advantage in conflict processing: Now
you see it, now you don’t. Cognition, 113(2), 135–149. https://
doi.org/10.1016/j.cognition.2009.08.001, PubMed: 19729156
Costa, A., Hernández, M., & Sebastián-Gallés, N. (2008). Bilingual-
ism aids conflict resolution: Evidence from the ANT task. Cogni-
tion, 106(1), 59–86. https://doi.org/10.1016/j.cognition.2006.12
.013, PubMed: 17275801
Dash, T., & Kar, B. R. (2020). Behavioural and ERP correlates of
bilingual language control and general-purpose inhibitory con-
trol predicted by L1 and L2 proficiency. Journal of Neurolinguis-
tics, 56, Article 100914. https://doi.org/10.1016/j.jneuroling
.2020.100914
De Baene, W., Duyck, W., Brass, M., & Carreiras, M. (2015).
Brain circuit for cognitive control is shared by task and lan-
guage switching. Journal of Cognitive Neuroscience, 27(9),
1752–1765. https://doi.org/10.1162/jocn_a_00817, PubMed:
25901448
De Clercq, W., Vergult, A., Vanrumste, B., Paesschen, W. V., &
Huffel, S. V. (2006). Canonical correlation analysis applied to
remove muscle artifacts from the electroencephalogram. IEEE
Transactions on Biomedical Engineering, 53(12), 2583–2587.
https://doi.org/10.1109/ TBME.2006.879459, PubMed:
17153216
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including inde-
pendent component analysis. Journal of Neuroscience Methods,
134(1), 9–21. https://doi.org/10.1016/j.jneumeth.2003.10.009,
PubMed: 15102499
de Vos, M., Riès, S., Vanderperren, K., Vanrumste, B., Alario, F.-X.,
Huffel, V. S., & Burle, B. (2010). Removal of muscle artifacts from
EEG recordings of spoken language production. Neuroinfor-
matics, 8(2), 135–150. https://doi.org/10.1007/s12021-010
-9071-0, PubMed: 20480401
Dijkstra, T., & van Heuven, W. J. B. (1998). The BIA model and
bilingual word recognition. In J. Grainger & A. M. Jacobs
(Eds.), Localist connectionist approaches to human cognition
(pp. 189–225). Lawrence Erlbaum Associates Publishers.
Dijkstra, T., & van Heuven, W. J. B. (2002). The architecture of the
bilingual word recognition system: From identification to deci-
sion. Bilingualism: Language and Cognition, 5(3), 175–197.
https://doi.org/10.1017/S1366728902003012
Eddy, M. D., & Holcomb, P. J. (2010). The temporal dynamics of
masked repetition picture priming effects: Manipulations of
stimulus-onset asynchrony (SOA) and prime duration. Brain
Research, 1340, 24–39. https://doi.org/10.1016/j.brainres.2010
.04.024, PubMed: 20403342
Eriksen, B. A., & Eriksen, C. W. (1974). Effects of noise letters upon
the identification of a target letter in a nonsearch task. Perception
& Psychophysics, 16(1), 143–149. https://doi.org/10.3758
/BF03203267
Fedorenko, E., & Thompson-Schill, S. L. (2014). Reworking the
language network. Trends in Cognitive Sciences, 18(3), 120–126.
https://doi.org/10.1016/j.tics.2013.12.006, PubMed: 24440115
Fox, J., & Weisberg, S. (2011). An R companion to applied regres-
sion (2nd ed.). Sage. https://socialsciences.mcmaster.ca/jfox
/Books/Companion-2E/index.html
Garbin, G., Sanjuan, A., Forn, C., Bustamante, J. C., Rodriguez-
Pujadas, A., Belloch, V., Hernandez, M., Costa, A., & Avila, C.
(2010). Bridging language and attention: Brain basis of the
impact of bilingualism on cognitive control. NeuroImage, 53(4),
1272–1278. https://doi.org/10.1016/j.neuroimage.2010.05.078,
PubMed: 20558314
G r e e n , D . W.
t h e b i l i n g u a l
( 1 9 9 8 ) . M e n t a l c o n t r o l o f
lexico-semantic system. Bilingualism: Language and Cognition,
1(2), 67–81. https://doi.org/10.1017/S1366728998000133
Green, D. W., & Abutalebi, J. (2013). Language control in bilin-
guals: The adaptive control hypothesis. Journal of Cognitive Psy-
chology (Hove, England), 25(5), 515–530. https://doi.org/10
.1080/20445911.2013.796377, PubMed: 25077013
Jackson, G. M., Swainson, R., Cunnington, R., & Jackson, S. R.
(2001). ERP correlates of executive control during repeated lan-
guage switching. Bilingualism: Language and Cognition, 4(2),
169–178. https://doi.org/10.1017/S1366728901000268
Jackson, S. R., Jackson, G. M., & Roberts, M. (1999). The selection
and suppression of action: ERP correlates of executive control in
humans. NeuroReport: For Rapid Communication of Neurosci-
ence Research, 10(4), 861–865. https://doi.org/10.1097
/00001756-199903170-00035, PubMed: 10208561
Jaeger, T. F. (2008). Categorical data analysis: Away from ANOVAs
(transformation or not) and towards logit mixed models. Journal
of Memory and Language, 59(4), 434–446. https://doi.org/10
.1016/j.jml.2007.11.007, PubMed: 19884961
Kornblum, S., Stevens, G. T., Whipple, A., & Requin, J. (1999). The
effects of irrelevant stimuli: 1. The time course of stimulus–
stimulus and stimulus–response consistency effects with Stroop-
like stimuli, Simon-like tasks, and their factorial combinations.
Journal of Experimental Psychology: Human Perception and
Performance, 25(3), 688–714. https://doi.org/10.1037/0096
-1523.25.3.688
Kroll, J. F., & Bialystok, E. (2013). Understanding the consequences
of bilingualism for language processing and cognition. Journal of
Neurobiology of Language
625
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
a
_
0
0
0
5
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
Cognitive Psychology (Hove, England), 25(5). https://doi.org/10
.1080/20445911.2013.799170, PubMed: 24223260
Kroll, J. F., Bobb, S. C., Misra, M., & Guo, T. (2008). Language
selection in bilingual speech: Evidence for inhibitory processes.
Acta Psychologica, 128(3), 416–430. https://doi.org/10.1016/j
.actpsy.2008.02.001, PubMed: 18358449
Kutas, M., & Federmeier, K. D. (2011). Thirty years and counting:
Finding meaning in the N400 component of the event-related
brain potential (ERP). Annual Review of Psychology, 62(1),
621–647. https://doi.org/10.1146/annurev.psych.093008
.131123, PubMed: 20809790
La Heij, W. (2005). Selection processes in monolingual and
bilingual lexical access. In J. F. Kroll & A. M. B. de Groot
(Eds.), Handbook of bilingualism: Psycholinguistic approaches
(pp. 289–307). Oxford University Press.
Lehtonen, M., Soveri, A., Laine, A., Järvenpää, J., de Bruin, A., &
Antfolk, J. (2018). Is bilingualism associated with enhanced
executive functioning in adults? A meta-analytic review. Psycho-
logical Bulletin, 144(4), 394–425. https://doi.org/10.1037
/bul0000142, PubMed: 29494195
Marian, V., & Spivey, M. (2003). Competing activation in bilingual
language processing: Within- and between-language competi-
tion. Bilingualism: Language and Cognition, 6(2), 97–115.
https://doi.org/10.1017/S1366728903001068
Marian, V., Spivey, M., & Hirsch, J. (2003). Shared and separate
systems in bilingual language processing: Converging evidence
from eyetracking and brain imaging. Brain and Language, 86(1),
70–82. https://doi.org/10.1016/S0093-934X(02)00535-7,
PubMed: 12821416
Masaki, H., Murphy, T. I., Desjardins, J. A., & Segalowitz, S. J.
(2012). The error-related negativity associated with different
strength of stimulus-response interference. Clinical Neurophysi-
ology, 123(4), 689–699. https://doi.org/10.1016/j.clinph.2011
.07.043, PubMed: 21873110
Nozari, N., & Novick, J. (2017). Monitoring and control in
language production. Current Directions in Psychological
S c i e n c e , 2 6 ( 5 ) , 4 0 3 – 4 1 0 . h t t p s : / / d o i . o r g / 1 0 . 1 1 7 7
/0963721417702419
Nunez, P. L., & Srinivasan, R. (2006). Electric fields of the brain: The
neurophysics of EEG. Oxford University Press. https://doi.org/10
.1093/acprof:oso/9780195050387.001.0001
Paap, K. R., & Greenberg, Z. I. (2013). There is no coherent
evidence for a bilingual advantage in executive processing. Cog-
nitive Psychology, 66(2), 232–258. https://doi.org/10.1016/j
.cogpsych.2012.12.002, PubMed: 23370226
Peeters, D., Vanlangendonck, F., Rueschemeyer, S.-A., & Dijkstra,
T. (2019). Activation of the language control network in bilingual
visual word recognition. Cortex, 111, 63–73. https://doi.org/10
.1016/j.cortex.2018.10.012, PubMed: 30458296
Piai, V., Roelofs, A., Jensen, O., Schoffelen, J.-M., & Bonnefond, M.
(2014). Distinct patterns of brain activity characterise lexical acti-
vation and competition in spoken word production. PLOS ONE,
9(2), Article e88674. https://doi.org/10.1371/journal.pone
.0088674, PubMed: 24558410
Picton, T. W. (1992). The P300 wave of the human event-related
potential. Journal of Clinical Neurophysiology: Official Publica-
tion of the American Electroencephalographic Society, 9(4),
456–479. https://doi.org/10.1097/00004691-199210000-00002,
PubMed: 1464675
Pinheiro, J. C., & Bates, D. M. (Eds.). (2000). Linear mixed-effects
models: Basic concepts and examples. In Mixed-effects models
in S and S-PLUS (pp. 3–56). Springer. https://doi.org/10.1007/0
-387-22747-4_1
R Core Team. (2020). The R project for statistical computing. https://
www.r-project.org/index.html
Ridderinkhof, K. R., Forstmann, B. U., Wylie, S. A., Burle, B., & van
den Wildenberg, W. P. M. (2011). Neurocognitive mechanisms of
action control: Resisting the call of the Sirens. Wiley Interdisci-
plinary Reviews. Cognitive Science, 2(2), 174–192. https://doi
.org/10.1002/wcs.99, PubMed: 26302009
Ridderinkhof, K. R., Wylie, S. A., van den Wildenberg, W. P. M.,
Bashore, T. R., & van der Molen, M. W. (2020). The arrow of
time: Advancing insights into action control from the arrow ver-
sion of the Eriksen flanker task. Attention, Perception, & Psycho-
physics, 83, 700–721. https://doi.org/10.3758/s13414-020
-02167-z, PubMed: 33099719
Riès, S. K., Fraser, D., McMahon, K. L., & de Zubicaray, G. I. (2015).
Early and late electrophysiological effects of distractor frequency
in picture naming: Reconciling input and output accounts. Jour-
nal of Cognitive Neuroscience, 27(10), 1936–1947. https://doi
.org/10.1162/jocn_a_00831, PubMed: 26042502
Riès, S. [K.], Janssen, N., Burle, B., & Alario, F.-X. (2013). Response-
locked brain dynamics of word production. PLOS ONE, 8(3),
Article e58197. https://doi.org/10.1371/journal.pone.0058197,
PubMed: 23554876
Riès, S. [K.], Janssen, N., Dufau, S., Alario, F.-X., & Burle, B. (2011).
General-purpose monitoring during speech production. Journal
of Cognitive Neuroscience, 23(6), 1419–1436. https://doi.org/10
.1162/jocn.2010.21467, PubMed: 20350170
Riès, S. K., Nadalet, L., Mickelsen, S., Mott, M., Midgley, K. J.,
Holcomb, P. J., & Emmorey, K. (2020). Pre-output language
monitoring in sign production. Journal of Cognitive Neurosci-
ence, 32(6), 1079–1091. https://doi.org/10.1162/jocn_a_01542,
PubMed: 32027582
Riès, S. K., Xie, K., Haaland, K. Y., Dronkers, N. F., & Knight, R. T.
(2013). Role of the lateral prefrontal cortex in speech monitoring.
Frontiers in Human Neuroscience, 7, Article 703. https://doi.org
/10.3389/fnhum.2013.00703, PubMed: 24194708
Robinson Anthony, J. J. D., & Blumenfeld, H. K. (2019). Language
dominance predicts cognate effects and inhibitory control in
young adult bilinguals. Bilingualism: Language and Cognition,
22(5), 1068–1084. https://doi.org/10.1017/S1366728918001013
Rodriguez-Fornells, A., De Diego Balaguer, R., & Münte, T. F.
(2006). Executive control in bilingual language processing.
Language Learning, 56(s1), 133–190. https://doi.org/10.1111/j
.1467-9922.2006.00359.x
Rodriguez-Fornells, A., van der Lugt, A., Rotte, M., Britti, B.,
Heinze, H.-J., & Münte, T. F. (2005). Second language interferes
with word production in fluent bilinguals: Brain potential and
functional imaging evidence. Journal of Cognitive Neuroscience,
17(3), 422–433. https://doi.org/10.1162/0898929053279559,
PubMed: 15814002
Roger, C. (2009). Supervision de l’action et optimisation des
comportements: Etudes électrophysiologiques et IRMf [Action
supervision and behavior optimization: Electrophysiological
studies and fMRI] [Unpublished doctoral thesis, Université de
Provence - Aix-Marseille I]. CCSD. https://tel.archives-ouvertes
.fr/tel-00448018
Roger, C., Bénar, C. G., Vidal, F., Hasbroucq, T., & Burle, B.
(2010). Rostral cingulate zone and correct response monitor-
ing: ICA and source localization evidences for the unicity of
correct- and error-negativities. NeuroImage, 51, 391–403.
https://doi.org/10.1016/j.neuroimage.2010.02.005, PubMed:
20152906
Salmelin, R., Hari, R., Lounasmaa, O. V., & Sams, M. (1994).
Dynamics of brain activation during picture naming. Nature,
Neurobiology of Language
626
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
a
_
0
0
0
5
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual cognitive control as revealed by Laplacian transformed ERPs
368(6470), 463–465. https://doi.org/10.1038/368463a0,
PubMed: 8133893
Shook, A., & Marian, V. (2013). The bilingual language interaction
network for comprehension of speech. Bilingualism: Language and
Cognition, 16(2). https://doi.org/10.1017/S1366728912000466,
PubMed: 24363602
Stoffels, E. J., & van der Molen, M. W. (1988). Effects of visual and
auditory noise on visual choice reaction time in a continuous-flow
paradigm. Perception & Psychophysics, 44(1), 7–14. https://doi.org
/10.3758/BF03207468, PubMed: 3405732
Sulpizio, S., Del Maschio, N., Fedeli, D., & Abutalebi, J. (2020).
Bilingual language processing: A meta-analysis of functional
neuroimaging studies. Neuroscience & Biobehavioral Reviews,
108, 834–853. https://doi.org/10.1016/j.neubiorev.2019.12.014,
PubMed: 31838193
Tremblay, P., & Gracco, V. L. (2009). Contribution of the pre-SMA
to the production of words and non-speech oral motor gestures,
as revealed by repetitive transcranial magnetic stimulation
(rTMS). Brain Research, 1268, 112–124. https://doi.org/10.1016
/j.brainres.2009.02.076, PubMed: 19285972
Tremblay, P., & Gracco, V. L. (2010). On the selection of words and
oral motor responses: Evidence of a response-independent
fronto-parietal network. Cortex, 46(1), 15–28. https://doi.org/10
.1016/j.cortex.2009.03.003, PubMed: 19362298
van Heuven, W. J. B., & Dijkstra, T. (2010). Language comprehen-
sion in the bilingual brain: FMRI and ERP support for psycholin-
guistic models. Brain Research Reviews, 64(1), 104–122. https://
doi.org/10.1016/j.brainresrev.2010.03.002, PubMed: 20227440
van Heuven, W. J. B., Schriefers, H., Dijkstra, T., & Hagoort, P.
(2008). Language conflict in the bilingual brain. Cerebral Cortex,
18(11), 2706–2716. https://doi.org/10.1093/cercor/ bhn030,
PubMed: 18424776
Vanlangendonck, F., Peeters, D., Rueschemeyer, S.-A., & Dijkstra,
T. (2019). Mixing the stimulus list in bilingual lexical decision
turns cognate facilitation effects into mirrored inhibition effects.
Bilingualism: Language and Cognition, 23(4), 836–844. https://
doi.org/10.1017/S1366728919000531
von Studnitz, R. E., & Green, D. W. (2002). Interlingual homograph
interference in German–English bilinguals: Its modulation and
locus of control. Bilingualism: Language and Cognition, 5(1),
1–23. https://doi.org/10.1017/S1366728902000111
Vidal, F., Burle, B., Grapperon, J., & Hasbroucq, T. (2011). An ERP
study of cognitive architecture and the insertion of mental
processes: Donders revisited. Psychophysiology, 48(9),
1242–1251. https://doi.org/10.1111/j.1469-8986.2011.01186.x,
PubMed: 21342207
Vidal, F., Burle, B., & Hasbroucq, T. (2018). The way we do the
things we do: How cognitive contexts shape the neural dynamics
of motor areas in humans. Frontiers in Psychology, 9, Article
1296. https://doi.org/10.3389/fpsyg.2018.01296, PubMed:
30100890
Vidal, F., Grapperon, J., Bonnet, M., & Hasbroucq, T. (2003). The
nature of unilateral motor commands in between-hand choice
tasks as revealed by surface Laplacian estimation. Psychophysi-
ology, 40(5), 796–805. https://doi.org/10.1111/1469-8986
.00080, PubMed: 14696733
Vidal, F., Hasbroucq, T., Grapperon, J., & Bonnet, M. (2000). Is the
“error negativity” specific to errors? Biological Psychology, 51(2–3),
109–128. https://doi.org/10.1016/S0301-0511(99)00032-0,
PubMed:10686362
Ye, Z., & Zhou, X. (2009). Executive control in language processing.
Neuroscience & Biobehavioral Reviews, 33(8), 1168–1177.
https://doi.org/10.1016/j.neubiorev.2009.03.003, PubMed:
19747595
Yeung, N., Botvinick, M. M., & Cohen, J. D. (2004) The neural basis
of error detection: Conflict monitoring and the error-related neg-
ativity. Psychological Review, 111(4), 931–959. https://doi.org/10
.1037/0033-295x.111.4.939, PubMed: 15482068
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
4
6
0
5
1
9
7
9
6
9
0
n
o
_
a
_
0
0
0
5
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
627