Unified Coding of Spectral and Temporal Phonetic Cues:
Electrophysiological Evidence for Abstract
Phonological Features
Philip J. Monahan1,2
, Jessamyn Schertz2,3
, Zhanao Fu2
, and Alejandro Pérez1,4
Abstract
■ Spoken word recognition models and phonological theory
propose that abstract features play a central role in speech pro-
cessing. It remains unknown, however, whether auditory cortex
encodes linguistic features in a manner beyond the phonetic
properties of the speech sounds themselves. We took advantage
of the fact that English phonology functionally codes stops and
fricatives as voiced or voiceless with two distinct phonetic cues:
Fricatives use a spectral cue, whereas stops use a temporal cue.
Evidence that these cues can be grouped together would indi-
cate the disjunctive coding of distinct phonetic cues into a func-
tionally defined abstract phonological feature. In English, the
voicing feature, which distinguishes the consonants [s] and [t]
from [z] and [d], respectively, is hypothesized to be specified
only for voiceless consonants (e.g., [s t]). Here, participants lis-
tened to syllables in a many-to-one oddball design, while their
EEG was recorded. In one block, both voiceless stops and frica-
tives were the standards. In the other block, both voiced stops
and fricatives were the standards. A critical design element was
the presence of intercategory variation within the standards.
Therefore, a many-to-one relationship, which is necessary to
elicit an MMN, existed only if the stop and fricative standards
were grouped together. In addition to the ERPs, event-related
spectral power was also analyzed. Results showed an MMN effect
in the voiceless standards block—an asymmetric MMN—in a
time window consistent with processing in auditory cortex, as
well as increased prestimulus beta-band oscillatory power to
voiceless standards. These findings suggest that (i) there is an
auditory memory trace of the standards based on the shared
[voiceless] feature, which is only functionally defined; (ii) voiced
consonants are underspecified; and (iii) features can serve as a
basis for predictive processing. Taken together, these results
point toward auditory cortex’s ability to functionally code dis-
tinct phonetic cues together and suggest that abstract features
can be used to parse the continuous acoustic signal. ■
INTRODUCTION
Spoken language comprehension requires listeners to rec-
ognize words embedded in the continuous acoustic
speech signal, which in turn involves identifying the seg-
mental units of words, phonemes (Kazanina, Bowers, &
Idsardi, 2018). The identity of different phonemes primar-
ily depends on specific spectral and temporal properties of
the auditory signal (Liberman, Cooper, Shankweiler, &
Studdert-Kennedy, 1967). These are the frequency profile
of the acoustic signal and the time intervals of major acous-
tic events (e.g., aspiration, vowel duration), respectively.
In linguistics, these distinctive spectral and temporal
characteristics are represented as features (Halle, 2002;
Clements & Hume, 1995; Chomsky & Halle, 1968;
Jakobson, Fant, & Halle, 1961). Ultimately, features are
the units of speech sound representations that link articu-
latory and acoustic characteristics of a given phoneme
(Baković, 2014; Halle, 1983). Some phonetic features code
for a spectral property, whereas others code for a temporal
1University of Toronto Scarborough, Canada, 2University of
Toronto, Canada, 3University of Toronto Mississauga, Canada,
4Cambridge University, United Kingdom
property. Different features have been postulated to
describe speech sounds in the field of linguistics, where
the notation [feature] refers to a specific feature. As an
example, sounds produced with primary airflow through
the nasal cavity (i.e., [m n ŋ]), which causes the attenuation
of acoustic energy in higher frequencies, all share the feature
[nasal]. In addition to their importance in phonological
theory, features have played a central role as perceptual units
in various models of spoken word recognition (Hickok,
2014; Poeppel & Monahan, 2011; Gow, 2003; Stevens,
2002; McClelland & Elman, 1986; Halle & Stevens, 1962).
Neuroimaging and neurophysiological studies support
the idea that features also play a central role in how audi-
tory cortex represents speech sounds (see the work of
Monahan, 2018, for a review). The spectral and temporal
characteristics of speech sounds are encoded by different
spectro-temporal receptive fields in superior temporal
gyrus (STG; Fox, Leonard, Sjerps, & Chang, 2020; Yi,
Leonard, & Chang, 2019; Hullett, Hamilton, Mesgarani,
Schreiner, & Chang, 2016; Mesgarani, Cheung, Johnson,
& Chang, 2014). Different populations in STG code dis-
tinct phonetic properties of speech sounds (Mesgarani
et al., 2014), including voice onset time (VOT; Fox et al.,
© 2022 Massachusetts Institute of Technology. Published under
a Creative Commons Attribution 4.0 International (CC BY 4.0) license.
Journal of Cognitive Neuroscience 34:4, pp. 618–638
https://doi.org/10.1162/jocn_a_01817
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
2020). Moreover, Khalighinejad, da Silva, and Mesgarani
(2017) and Mesgarani et al. (2014) showed that stops
and fricatives are coded by distinct neuronal populations,
independent of their voicing status. Collectively, phonetic
classes are distributively coded by auditory cortex. It is
important to note, however, that all stops share a common
phonetic characteristic in both articulation and acoustics:
the presence of a burst followed by a release. All fricatives
also share a common phonetic characteristic: a narrow
constriction in the vocal tract, resulting in turbulent acous-
tic energy. In other words, different stop consonants are
phonetically linked, and different fricative consonants
are phonetically linked. These studies demonstrate that
auditory cortex encodes phonetic sound classes; however,
it is still unknown if auditory cortex encodes features in a
manner that goes beyond the phonetic properties of the
speech sounds themselves.
Direct evidence of whether auditory cortex functionally
links different phonetic sound classes of the same phonol-
ogical class is key to determining whether auditory cortex
can represent abstract phonological features. Here, we
take advantage of the functionally defined English feature
[voiceless] to determine whether two distinct phonetic
classes of speech sounds, that is, stops and fricatives, can
be grouped together when they share voicing. The feature
[voiceless] in word-initial English stops (i.e., [ph th kh]) is
primarily cued by a temporal dimension: a long duration
VOT compared to a short duration VOT for voiced stops
(Lisker & Abramson, 1964). Meanwhile, in fricatives
(e.g., [f s]), the feature [voiceless] is primarily cued by a
spectral dimension: the absence of periodic low-frequency
energy that is present in their voiced counterparts because
of glottal adduction (Smith, 1997). Overall, English pro-
vides a strong test case for our question of interest, as
opposed to languages where voicing in fricatives and stops
are cued by the same phonetic property. For example,
Spanish uses periodic low-frequency energy to cue voicing
in both stops and fricatives (Lisker & Abramson, 1964).
Specifically, the distinction between voicing in English
and Spanish indicates that whether a speech sound is
phonologically voiced or voiceless is not tied directly to
a single acoustic pattern or phonetic cue.
The question addressed here is whether spectral and
temporal phonetic features can be disjunctively coded
together into a larger phonological category. To address
this question, we performed an auditory MMN experi-
ment. The MMN is a component in the ERP, sensitive to
language-specific properties of speech sounds in auditory
memory ( Winkler, Kujala, Alku, & Näätänen, 2003;
Näätänen, 2001; Phillips et al., 2000; Sharma & Dorman,
1999, 2000; Winkler et al., 1999; Näätänen et al., 1997). It
is usually evoked in an oddball paradigm, which consists
of a series of repeating (standard) stimuli interrupted
by an infrequent (deviant) stimulus. The auditory MMN
typically peaks between 150 and 350 msec postonset of a
deviant stimulus and is maximal over fronto-central elec-
trode sites. The source of the auditory MMN response
consistently localizes to the auditory cortex (Alho, 1995;
Aulanko, Hari, Lounasmaa, Näätänen, & Sams, 1993; Sams,
Kaukoranta, Hämäläinen, & Näätänen, 1991). The MMN is
a useful tool to study whether auditory cortex perceptually
groups over distinct cues. For example, Gomes, Ritter, and
Vaughan (1995) studied the MMN response to pure-tone
sinusoids and observed that the duration cue alone can be
extracted from a series of standards that also varied in fre-
quency and intensity. This suggests that auditory memory
representations can be built based on a single cue, even
when other cues are varying.
Here, English-speaking participants took part in a
many-to-one oddball experiment. Their EEG was recorded
as they listened to a series of frequent, standard consonant–
vowel syllables, interrupted by an infrequent, deviant
consonant–vowel syllable. The standards and deviants in
each block were sampled from five different phones (i.e.,
voiceless: [ph th kh f s], voiced: [b d ɡ v z]). In the voiceless
standards block, participants heard voiceless consonants
as the standards, interrupted by occasional voiced devi-
ants. In the voiced standards block, participants heard
voiced consonants as the standards, interrupted by occa-
sional voiceless deviants. As we used multiple phonetic
categories with differing manners of articulation in the
standards, a many-to-one oddball relationship among
the standards only exists if the stops and fricatives that
share voicing are grouped together along the basis of their
abstract voicing representation. An MMN effect would
indicate that distinct phonetic cues group together per-
ceptually based on an abstract phonological feature.
In nearly all MMN studies of speech perception, the
standard category in one block serves as the deviant in
the other block. If both categories are native to the lan-
guage of the participants and contrastive in the language,
then equal-sized MMNs are predicted across the two
blocks. This is not always the case, however. In fact,
MMN evidence for features largely arises from findings that
demonstrate that some speech sounds elicit larger MMNs
than others. These asymmetric results have been observed
for vowels (de Rue, Snijders, & Fikkert, 2021; Yu & Shafer,
2021; Scharinger, Monahan, & Idsardi, 2012, 2016;
Cornell, Lahiri, & Eulitz, 2011; Eulitz & Lahiri, 2004),
consonants (Fu & Monahan, 2021; Hestvik, Shinohara,
Durvasula, Verdonschot, & Sakai, 2020; Schluter, Politzer-
Ahles, Al Kaabi, & Almeida, 2017; Hestvik & Durvasula,
2016; Schluter, Politzer-Ahles, & Almeida, 2016; Cornell,
Lahiri, & Eulitz, 2013; Maiste, Wiens, Hunt, Scherg, &
Picton, 1995), and lexical tone (Politzer-Ahles, Schluter,
Wu, & Almeida, 2016). These asymmetries are often—
although not exclusively (see the work of Maiste et al.,
1995)—taken to reflect the underlying featural content
of the two categories consistent with underspecified rep-
resentations (Lahiri & Reetz, 2002, 2010). Traditionally,
features were conceptualized as binary. That is, the
sound [n] was [+nasal], whereas the sound [d] was
[−nasal]. Underspecified features posit that only con-
trastive aspects of a speech sound are represented.
Monahan et al.
619
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Therefore, [n] is specified for nasality, possessing the fea-
ture [nasal], while [d] lacks any specification for [nasal] in
memory and, thus, does not contain the feature [−nasal]
(Archangeli, 1988; Steriade, 1995). A larger MMN is
observed when the standard is specified for a given fea-
ture and the deviant mismatches with that feature. When
the standard is underspecified, there is no mismatch
between the standard and deviant, and as such, a smaller
or no MMN is observed. In studies that have tested voic-
ing in English stops (Hestvik & Durvasula, 2016) and
English fricatives (Schluter et al., 2017), larger MMNs
are observed when voiceless consonants are the standard
relative to when the voiced consonants are the standard,
suggesting that voiceless consonants are specified with
the feature [voiceless].
These neurophysiological results align with proposals
from phonological theory, which posit that English voice-
less stops and fricatives are specified for their voicing sta-
tus and contain the feature [voiceless], whereas voiced
consonants are unmarked and underspecified for voicing
(Avery & Idsardi, 2001). Linguistic evidence for this claim
arises from the findings that (i) voiced English obstruents
are less consistent in their phonetic realization relative to
voiceless obstruents (Smith, 1997; Docherty, 1992) and
(ii) voiced English obstruents do not trigger assimilation,
whereas voiceless English obstruents do (Iverson &
Salmons, 1995). Both observations are consistent with
voiceless English obstruents possessing a marked feature
for voicing and voiced obstruents lacking such a marked
feature. That voiceless obstruents are marked, whereas
voiced obstruents are unmarked, however, is language-
specific; languages with different voicing systems (e.g.,
Spanish, Japanese, Thai, Hindi) could have distinct voicing
specifications. The above asymmetry in representation
makes predictions for conditions in which we anticipate
observing an MMN. We expect an MMN when the speci-
fied [voiceless] consonants are the standards but not
when the underspecified voiced consonants are the
standards.
It is important to note that standard–deviant designs
entail a matching procedure of the upcoming physical
stimulus with an auditory memory representation
(Näätänen & Kreegipuu, 2012; Näätänen, Jacobsen, &
Winkler, 2005; Näätänen, 2001). Thus, anticipating what
sound comes next is inherent in oddball designs evoking
the MMN component ( Winkler, 2007). If we observe dif-
ferences between specified standards and underspecified
standards before hearing it, this suggests predictive pro-
cessing that is at least partially based on that feature. In
other words, different neural states preceding the onset
of different types of repetitive sounds would indicate that
the expectancy between these sounds varies depending
on the segregating characteristic, that is, the feature. In
fact, differences before the onset of different kinds of
standards have been observed (Scharinger et al., 2016).
Those differences were evident in the induced neural
oscillatory activity, which is neurophysiological activity
that—unlike ERPs—is time-locked but not phase-locked
to endogenous sensory events. Neuronal oscillations play
a key role in auditory perception of verbal input (Morillon
& Schroeder, 2015; Doelling, Arnal, Ghitza, & Poeppel,
2014; Arnal & Giraud, 2012; Obleser & Weisz, 2012; Arnal,
Wyart, & Giraud, 2011), with various rhythms ascribed to
specific functional roles. Brain oscillations refer to rhyth-
mic fluctuations in the excitability of neuronal popula-
tions, sometimes in response to sensory stimulation.
Beta-frequencies (approximately 15–30 Hz) are dominant
in propagating information flow from higher cortical areas
and levels of representation (Riddle, Hwang, Cellier,
Dhanani, & D’Esposito, 2019; Lewis, Schoffelen, Schriefers,
& Bastiaansen, 2016; Fontolan, Morillon, Liegeois-Chauvel,
& Giraud, 2014; Arnal et al., 2011; Wang, 2010). For exam-
ple, descending information flow from higher-order brain
areas in an individual listening to sentences is frequency
specific (approximately 15–30 Hz), suggesting that beta-
frequencies are dominant in top–down propagation
(Fontolan et al., 2014). Scharinger et al. (2016) observed
an increase in beta-power in a prestimulus time window
when the standards were specified (i.e., the high-vowel
[ɪ]) relative to when the standards were underspecified
(i.e., the mid-vowel [ɛ]). This prestimulus increase in
beta-power is taken to reflect predictive mechanisms
associated with the MMN, as presumably, only specified
features can be used to predict the upcoming stimulus.
Here, we also investigated beta-band oscillatory activity
before hearing the stimulus, to provide complementary
evidence to the neural implementation of features. In
the context of predictive processing, if features can be
used as the basis for upcoming predictions, we expect
beta-band power increases to the specified voiceless
standards, consistent with Scharinger et al. (2016).
In short, this study aims to determine if auditory cortex
constructs disjunctive groupings for linguistic features in
auditory memory and whether features have predictive
power in auditory processing. We employ an MMN design
that adopted intercategory variation in the standards. In
one block, the standards were voiced consonants, and
the deviants were voiceless consonants. In the other
block, the standards were voiceless consonants, and the
deviants were voiced consonants. Importantly, the stan-
dards and deviants included both stops and fricatives. Elic-
iting the MMN suggests that auditory cortex disjunctively
codes the temporal and spectral auditory cues for voicing
into a coherent auditory memory trace. For the abstract
phonological feature [voiceless], we predict an MMN in
the voiceless standards block, as [voiceless] appears to
be the specified feature, whereas voiced consonants are
underspecified for voicing. Finally, given that increased
beta-power has been observed in blocks whose standards
are putatively specified for a given feature (see the work of
Scharinger et al., 2016), we predict an increase in prestim-
ulus beta-power in the voiceless standards block and a
reduction in prestimulus beta-power in the voiced stan-
dards block.
620
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
METHODS
Participants
Thirty native speakers of English participated. All participants
were recruited from the University of Toronto Scarborough.
Data from three participants were excluded because of tech-
nical issues during data acquisition. This left 27 participants
(mean age = 20.3 years, SD = 2.4 years, 17 women) included
in the analysis. All participants were right-handed, as
assessed by the Edinburgh Handedness Survey (Oldfield,
1971) and self-reported no known history of auditory,
language, or neurological deficits. Finally, all participants pro-
vided written informed consent before the experiment and
were remunerated for their time. This study was approved
by the research ethics board at the University of Toronto.
Materials
The experimental stimuli included 14 consonant–vowel
syllables in a [Cɑ] frame. There were five voiced ([b d ɡ
v z]) and five voiceless consonants ([ph th kh f s]), pro-
duced 3 times each. Each group of voiced and voiceless
consonants included three stop consonants and two frica-
tives. In addition, the syllables [vi vu pho fu] were included
as fillers. Stimuli were recorded by a female native speaker
of North American English in a sound-attenuated cabin.
Recordings were made with an Audio-Technica AT3035
microphone at a sampling rate of 44.1 kHz and 16-bit
depth. The syllables were produced at a natural speaking
rate (mean duration = 540 msec, SD = 58 msec). Stimuli
intensity was normalized to a mean intensity of 75 dB SPL.
Cosine2 offset ramps were also applied to the offset of the
auditory stimuli; this was not applied to the onset to pre-
serve the burst properties of the stop consonants.
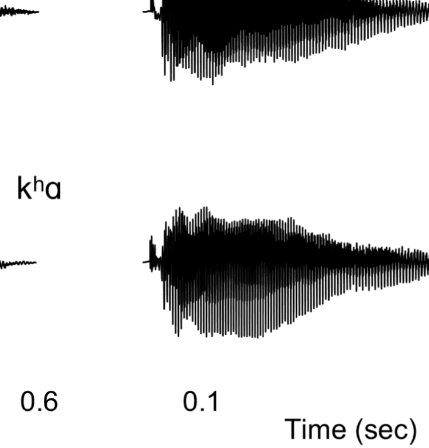
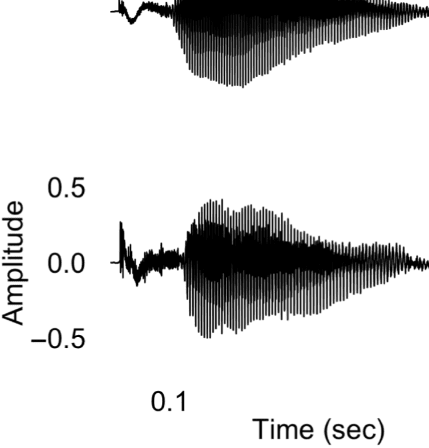
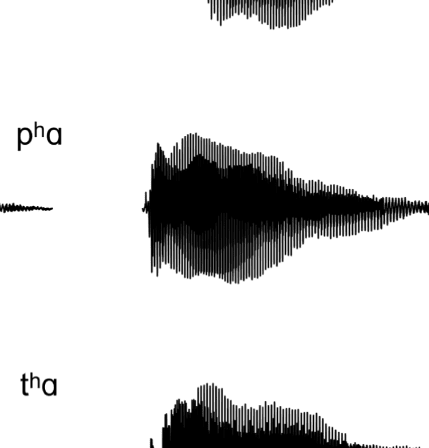
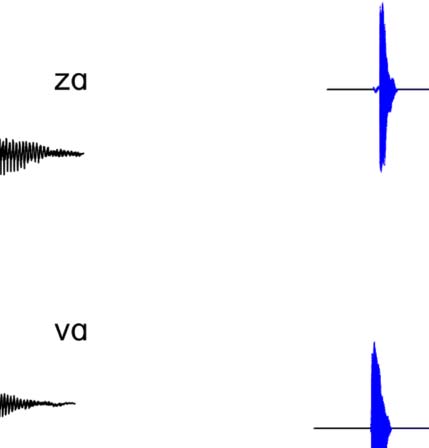
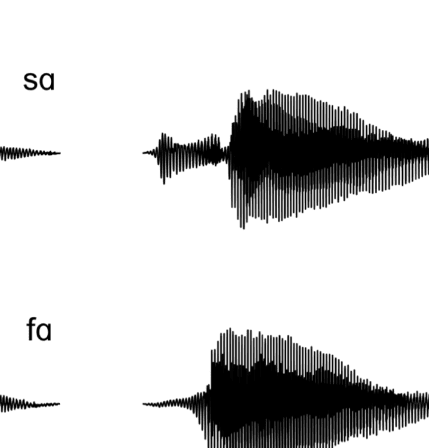
Figure 1A presents example acoustic waveforms for
10 representative experimental syllables, one for each
category, used in the experiment. The voiced fricatives
included periodic acoustic energy during the fricative, as
is common in syllable-initial position in North American
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. (A) Waveforms of sample stimuli for each phonetic category used in the experiment. Voiceless consonants are on the left, and voiced
consonants are on the right. The top four waveforms are of fricatives used in the experiment, whereas the bottom six waveforms are of stops used in
the experiment. The voiced fricatives contain periodic energy before the vowel, as is evident in both [zɑ] and [vɑ]. The voiceless fricatives lack this
periodic energy. In the stop consonants, the voiceless stops have a longer interval between the release of the burst and the onset of the vowel; that is,
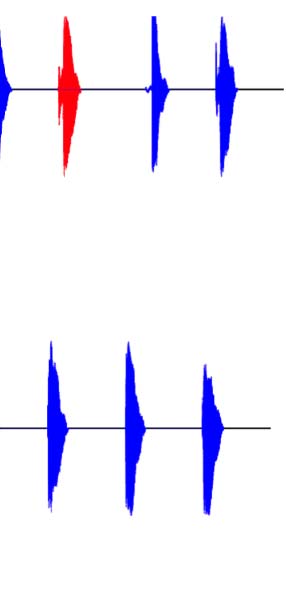
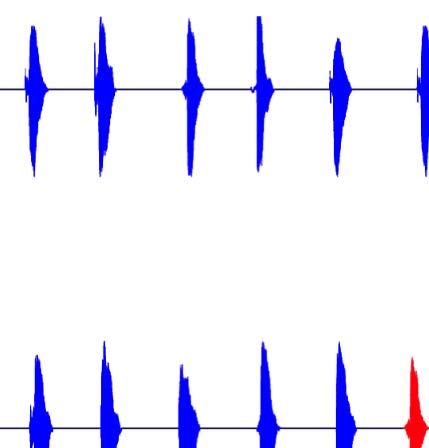
they contain a longer VOT. The VOT for voiced stops is considerably shorter. (B) Schematic of the many-to-one oddball paradigm used in the
experiment for both the voiceless (top) and voiced (bottom) standard blocks. The standards (blue) in each block included intercategory variation,
interrupted by an infrequent deviant (red) sampled from the other voicing category. (C) Predictions for the MMN experiment for the voiceless block
only. If the intercategory varying standards are grouped along a shared voicing feature (top), we predict a many-to-one relationship and,
consequently, the presence of an MMN. If grouping is not possible (bottom), then no many-to-one relationship exists, and no MMN is predicted.
Monahan et al.
621
English (Smith, 1997). This was absent in the voiceless fric-
atives. The voiceless stops had a longer interval of time,
that is, the VOT, between the release of the stop burst
and the onset of the vowel as compared with the voiced
stops (Lisker & Abramson, 1964). Note that there is no
physical property that binds voiceless stops and voiceless
fricatives together, only a linguistic property, namely,
voicing.
As noted above, the primary phonetic cues to phonolo-
gical voicing are distinct in stops and fricatives. There are,
however, secondary acoustic–phonetic cues on which
voiced and voiceless consonants systematically differ,
although in a less reliable manner (Lisker, 1986). To deter-
mine whether such secondary acoustic–phonetic cues
could robustly indicate voicing category membership in
our stimuli, we measured several acoustic dimensions
known to be secondary cues to the English voicing con-
trast. Specifically, we measured total syllable duration,
consonant duration, vowel duration, fundamental (f0)
and first formant (F1) frequency at vowel onset, and inten-
sity of the following vowel. Table 1 provides the means and
standard deviations, and Figure 2 provides the distribu-
tions for each of these cues in our stimuli. The average
values showed expected cross-category patterns, but as
is evident by the overlap in the distributions, a value from
a single token is not informative about its voicing category
membership. Specifically, the distributions for voiced
stops and fricatives always overlap with the distributions
for voiceless stops and fricatives. In short, these secondary
cues do not reliably indicate phonological voicing category
membership and, therefore, are unlikely to contribute to
effects found in the current work.
Procedure
Participants were seated in a quiet room and passively
listened to the stimuli, while watching a silent movie to
maintain an awake state and reduce excessive ocular
movements (Tervaniemi et al., 1999). Two blocks were
presented with a break in between. One block contained
voiceless standards and voiced deviants. The other block
contained voiced standards and voiceless deviants. The
order of blocks was counterbalanced across participants.
Figure 1B represents the many-to-one oddball paradigm
used in the voiceless (top) and the voiced block (bottom).
The standards (blue) in each block included intercategory
variation, interrupted by an infrequent deviant (red) sam-
pled from the other voicing category.
In each block, participants heard approximately 785 stim-
uli, consisting primarily of standards (e.g., voiceless conso-
nants in the voiceless standards block) interspersed with
deviants (e.g., voiced consonants in the voiceless standards
block) at semiregular intervals. There was, on average, a 7-
to-1 standard-to-deviant ratio, with approximately 676 stan-
dard and exactly 105 deviant stimuli per block. In each block,
three distinct acoustic tokens of the five standard syllables
were presented to require participants to abstract over
token-specific acoustic properties (Hestvik & Durvasula,
2016; Scharinger et al., 2012, 2016; Kazanina, Phillips, &
Idsardi, 2006; Phillips et al., 2000; Aulanko et al., 1993).
For the deviants, only one acoustic token of each of the
five deviant syllables was presented, and each deviant cat-
egory was presented 15 times. This led to 75 deviants per
block. Moreover, there were also two filler deviant sylla-
bles per block. These filler deviants matched the standards
in their voicing but differed in their vowel. They were pre-
sented 15 times each, for an additional 30 deviant trials.
Filler deviants were included to ensure that there would
be detectable differences between tokens as the vowel
changes are likely more salient than the consonant
changes given the intercategory variation in the standard
consonants. Each deviant stimulus was preceded by sev-
eral standard stimuli (range: 4–10). As few as three to four
consecutive standards allow for the elicitation of an MMN
response (e.g., Dehaene-Lambertz, 2000; Cowan, Winkler,
Teder, & Näätänen, 1993). The number of standards was
randomly drawn from a uniform distribution, and the stan-
dard stimuli themselves were randomly sampled from the
Table 1. Means for the Measured Secondary Acoustic–Phonetic Cues
Voicing
Manner
Total Duration
(msec)
Consonant Duration
(msec)
Vowel Duration
(msec)
f0 (Hz)
F1 (Hz)
Intensity
(dB SPL)
Voiced
519 (62.5)
Fricative
565 (75.1)
Stop
488 (25.4)
Voiceless
562 (45.3)
Fricative
590 (60.4)
Stop
543 (17.4)
55 (56.5)
116 (38.7)
14 (4.3)
121 (36.1)
157 (27.2)
98 (15.1)
464 (34.6)
206 (14.3)
571 (64.0)
75.9 (0.35)
449 (43.3)
209 (11.2)
574 (81.2)
76.3 (0.26)
474 (25.5)
203 (16.6)
569 (53.7)
75.7 (0.17)
441 (28.8)
224 (10.2)
743 (86.4)
76.5 (0.27)
433 (37.0)
220 (9.0)
661 (38.8)
76.7 (0.09)
445 (23.1)
227 (10.5)
805 (50.6)
76.3 (0.21)
One standard deviation of the mean is presented in parentheses. Durations for stop consonants were measured from the offset of the burst to the
onset of the vowel. Total duration and vowel duration were measured until the end of periodic voicing in the vowel. f0 and F1 frequency measure-
ments were taken 10 msec after vowel onset. Intensity refers to the intensity of the vowel alone.
622
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
–
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Distributions for each secondary acoustic–phonetic cue measured in the stimuli. Each panel represents one cue: (A) total stimulus
duration, (B) consonant duration, (C) vowel duration, (D) f0, (E) F1 frequency, (F) vowel intensity. Points indicate each of the 30 stimuli.
five consonants of its voicing category. Given that the
standard token categories were randomly sampled, the
number of tokens of a given category in single standard
train was not controlled. The duration of the ISI was ran-
domly sampled from a uniform distribution between 1.0
and 1.5 sec. ISI durations in this time range have been
shown to reinforce phonological-level processing ( Werker
& Logan, 1985).
EEG Recording and Analysis
Continuous EEG recordings were acquired from 32-
channel actiCAP active electrodes connected to an acti-
CHamp amplifier (Brain Products GmbH). The EEG signal
was digitized at a 500-Hz sampling frequency with a 200-Hz
on-line low-pass filter. Electrodes were positioned on the
scalp according to the International 10–20 system. Posi-
tions included Fp1/2, F3/4, F7/8, FC1/2, FC5/6, FT9/10,
C3/4, T7/8, CP1/2, CP5/6, TP9/10, P3/4, P7/8, O1/2, Oz,
Fz, Cz, and Pz. A ground electrode was placed at Fpz.
The EEG signal was referenced to the right mastoid
(TP10) on-line. Impedances were reduced below 20 kΩ
at each electrode site before recording. Four additional
bipolar electrodes were placed on the temples and above
and below the right eye to monitor the EOG. The auditory
stimuli were delivered to participants through RHA
MA750i noise-isolating in-ear headphones. In addition to
the EEG channels, the auditory signal was also sent to
the amplifier. This provides the opportunity for off-line
correction of any temporal delay between the delivery of
the auditory stimulus and sending of the digital trigger
marker (Pérez, Monahan, & Lambon Ralph, 2021).
Data analysis was carried out using the EEGLAB toolbox
v14.1.1 (Delorme & Makeig, 2004) and custom programs
running in MATLAB ( Version 2017a, The MathWorks Inc.).
First, offset delays between the trigger and the auditory
stimulus presentation were corrected to ensure millisecond-
precise stimulus-digital trigger synchrony. This was done
by cross-correlating the original stimuli waveforms and
the auditory track in the EEG recording. Second, only
standards that followed at least two other standard trials
were retained. This was done to ensure that the averaged
standard responses included only trials that were pre-
ceded by multiple stimuli from the same voicing category.
Next, the EEG signal was rereferenced to the linked mas-
toids, which provide the most robust MMN responses
(Mahajan, Peter, & Sharma, 2017). Subsequently, a high-
pass filter (finite impulse response; FIR) at 1 Hz was
applied. The PREP toolbox (Bigdely-Shamlo, Mullen,
Kothe, Su, & Robbins, 2015) was used to identify and
Monahan et al.
623
remove bad channels (mean = 1.5, SD = 1.4). These bad
channels were then interpolated. Because of the relatively
small channel montage (i.e., 32 channels), the spherical
spline interpolation method was utilized for bad channel
interpolation, which is more accurate for low-density elec-
trode montages (Perrin, Pernier, Bertrand, & Echallier,
1989). Next, an artifact subspace reconstruction (ASR)
algorithm adapted from the EEGLAB software (Mullen
et al., 2013) was implemented to remove high-amplitude
artifacts. ASR transforms a sliding window of the EEG data
with a principal component analysis to identify channels
of high variance. This is accomplished via a statistical com-
parison of clean EEG data containing minimal artifacts.
This clean data were automatically identified inside each
individual EEG recording and subsequently used as the
calibration data for the ASR. Corrupted channels (or sub-
spaces of multiple channels) were reconstructed from
neighboring channels using a mixing matrix, which is com-
puted from the covariance matrix of the calibration data.
In this study, a sliding window of 500 msec and a variance
threshold of 3 SDs were used to identify corrupted
subspaces. Portions of the data that were not possible to
reconstruct because of the presence of multiple artifacts
were removed.
Subsequently, an adaptive mixture-independent com-
ponent analysis (AMICA) technique (Palmer, Kreutz-
Delgado, & Makeig, 2012) was applied to the cleaned
EEG data from each participant to separate the EEG
recordings, a combination of individual brain and non-
brain sources mixed by volume conduction, into spatially
static independent components of maximal temporal
independence. AMICA maximizes mutual information
reduction and the dipolarity of scalp projections following
decomposition (Delorme, Palmer, Onton, Oostenveld, &
Makeig, 2012). AMICA was performed on 45 min of EEG
data for each participant. Thus, the number of time points
used to estimate the weighting matrix ranged from 161 to
177 times the number of channels squared, exceeding rec-
ommendations for satisfactory decomposition (Delorme
et al., 2012). Next, an equivalent dipole current source
was fit to each independent component from a three-shell
boundary element model using the DIPFIT toolbox in
EEGLAB (Oostenveld & Oostendorp, 2002). The EEG elec-
trode positions were aligned to fit the standard Montreal
Neurological Institute brain. Then, individual components
accounting mainly for electrooculographic, electrocardio-
graphic, electromyographic, or line noise were removed
from the data. Component rejection was performed man-
ually guided by the following criteria: (i) component’s
topography, (ii) component’s time series, (iii) compo-
nent’s power spectrum properties, and (iv) properties of
the dipole associated to each component: localization
outside of the head or close to the eyes jointly with low
variance (up to 5%). On average, we removed two compo-
nents per participant (range: 1–5). From this step, the
ERPs and event-related spectral perturbations (ERSPs)
were computed.
After data preprocessing, in the voiceless standards
block, there were, on average, 69 voiced deviants (SD =
9.2) and 440 voiceless standards (SD = 61.3). In the voiced
standards block, there were, on average, 71 voiceless devi-
ants (SD = 7.1) and 435 voiced standards (SD = 51.1). Over-
all, there were, on average, 438 standards (SD = 55.9) and
70 real deviants (SD = 8.2) per block, and the number of
trials were approximately equivalent across blocks. More-
over, data preprocessing did not disproportionately affect
the different real deviant categories (percentage of trials
retained after data preprocessing; voiced deviant stop:
92.9%; voiced deviant fricative: 91.6%; voiceless deviant
stop: 93.7%; voiceless deviant fricative: 95.4%).
ERP Analysis
First, the continuous EEG signal was down-sampled to
250 Hz. Next, epochs were extracted with a 100-msec
prestimulus baseline and a 700-msec poststimulus onset
time window. Epochs were baseline corrected using the
100-msec prestimulus baseline. Average ERPs were calcu-
lated for each condition. The difference ERP waveform was
obtained for each block by subtracting the ERPs of the
standard from the deviant. Because MMNs are largest over
fronto-central scalp areas when referenced to linked
mastoids (Näätänen, Paavilainen, Rinne, & Alho, 2007),
the eight fronto-central electrode sites (i.e., Fz, FC1/2,
Cz, C3/4, CP1/2) were collapsed. The resulting channel-
averaged ERPs for each condition were statistically
compared at each time point. Statistical analyses were
conducted in the −100- to 700-msec time window with
permutation tests on the t statistic and a false discovery
rate (FDR) correction for multiple comparisons. The num-
ber of permutations was set to 2000. Differences with
pFDR < .05 are reported as statistically significant.
To ensure that the observed ERP responses are driven
by the standard–deviant relationship, we also included a
control analysis. In this analysis, the ERP to the deviant is
compared with the ERP to the standard version of the
same stimulus (Hestvik & Durvasula, 2016; Peter,
McArthur, & Thompson, 2010; Pulvermüller & Shtyrov,
2006; McGee et al., 2001; Deacon, Gomes, Nousak, Ritter,
& Javitt, 2000). This type of analysis is referred as the iden-
tity MMN (iMMN). The iMMN for the voiced consonants
was calculated by comparing the ERP responses when they
were the standard (voiced standard block) compared to
when they were the deviant (voiceless standard block).
Similarly, the voiceless iMMN was calculated by comparing
the ERP of the voiceless standards in the voiceless stan-
dard block versus the ERPs of the voiceless deviants in
the voiced standard block. Despite substantial variation
in the standards and deviants in the current design, it is
possible that any differences in the within-block compari-
son may arise because of the ERPs being different to
voiceless and voiced consonants and not because of the
standard–deviant relationship. The iMMN comparison
potentially eliminates this confound. The presence of an
624
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
iMMN suggests that the differences observed in the within-
block MMN cannot be solely attributed to intrinsic ERP
response differences to voiceless versus voiced conso-
nants, but that the MMN is also driven by the standard–
deviant relationship.
ERSP Analysis
For the ERSP analysis, epochs were extracted from the
continuous EEG signal with a 1-sec baseline and 2-sec
poststimulus onset time window. The time–frequency
decomposition of each epoch was computed using a
wavelet window Morlet taper. The number of cycles in
each Morlet increased linearly in 0.8 cycles beginning at
3 cycles at 3 Hz. Single-trial normalization at each time–
frequency bin was performed by dividing frequency spe-
cific power averaged from −444 to −300 msec. Finally,
ERSPs containing estimations from 3 to 30 Hz and −300
to 700 msec were averaged across all channels for each
condition. Statistical analyses comparing the voiced and
voiceless standards were conducted with bootstrap tests.
The number of random samples used in the bootstrap was
2000. Differences with pFDR < .05 are reported as statisti-
cally significant.
RESULTS
ERPs
Average waveforms elicited over fronto-central electrode
sites in each block are presented in Figure 3. Figure 3A
shows the grand average for the voiceless standard block.
The permutation test revealed significant differences in
the time window from 116 to 196 msec and a later time
window, from 316 to 440 msec. Figure 3B shows the grand
average in the voiced standard block. The response to the
voiceless deviant was relatively more negative in the 216-
to 240-msec time window and at two very brief later time
windows (i.e., 532–556 msec, 636–648 msec); however,
the voiceless deviant elicited a larger positivity in the
292- to 364-msec time window relative to the voiced
standard. Figure 3C provides a comparison of the two
standards. The voiced standard stimuli elicited a larger
negativity in the 68- to 138-msec time window and a larger
positivity in the 208- to 248-msec time window. In turn, the
voiceless standard stimuli elicited a larger positivity in the
292- to 436-msec time window. A comparison of ERP
responses to the standards based on manner (i.e., stop,
fricative) and voicing (i.e., voiced, voiceless) is provided
in Appendix A. Finally, Figure 3D compares the difference
waves between the two blocks (deviant minus standard).
The difference between the deviants and standards across
the two blocks is evident between 96 and 188 msec and
between 288 and 412 msec.
Overall, in the voiceless standards block, the voiced
deviant eliciting a larger negative deflection in the ERP rel-
ative to the voiceless standard is consistent with typical
observations in an MMN paradigm. In the voiced standards
block, the most robust difference is during the 292- to
364-msec time window. There, the deviant elicited a larger
positive response relative to the standard, which is oppo-
site of the characteristic MMN pattern. The presence of the
early and late negative ERP deflections in the voiceless
standard block suggests that auditory cortex was able to
perceptually group the voiceless consonants together.
Next, we conducted an iMMN analysis to establish
whether the MMN effect is driven by the standard–deviant
relationship. Figure 4 shows that there was no difference
when the voiceless consonants were the standards relative
to when they were the deviant. The panels beneath the ERP
waveforms show the difference waves (deviant minus stan-
dard) for each block. In the voiced consonants iMMN com-
parison, voiced consonants as the deviant elicited a larger
negativity in the 196- to 212-msec time window compared
to when they served as the standards. Results from iMMN
analysis suggests that the differences in the within-block
MMN comparison above are not solely because of intrinsic
ERP responses to voiced and voiceless consonants alone
but also reflect the standard–deviant relationship.
Figure 3. Average ERP responses for the within-block comparisons. ERPs are the average of eight fronto-central scalp electrodes (Fz, FC1/2, Cz, C3/4,
CP1/2). (A) ERP responses in the voiceless standards block: voiceless standard and voiced deviant; (B) ERP responses in the voiced standards block:
voiced standard and voiceless deviant; (C) ERP responses to the standards: voiceless standard and voiced standard; (D) comparison of ERP difference
waves (deviant minus standard) in each block (voiced refers to the voiced standards block, voiceless refers to the voiceless standards block). Rug
plots along the x axes indicate time points when the comparison was statistically significant (see text for analysis procedures). Shaded regions
represent the 95% confidence interval of the mean.
Monahan et al.
625
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. iMMN ERP responses averaged over eight fronto-central scalp electrodes (Fz, FC1/2, Cz, C3/4, CP1/2) for (left) voiceless standards and
deviants and (right) voiced standards and deviants. The bottom panels are the difference ERP waves, computed as the response to the standard
subtracted from the response to the deviant, for the (left) voiceless consonants and (right) voiced consonants. Rug plots along the x axes indicate
time points when the comparison was statistically significant (see text for analysis procedures). Shaded regions represent the 95% confidence interval
of the mean.
Thus far, the MMN comparisons have focused on fronto-
central electrode sites. Analyzing the topographic distribution
allows us to (i) identify the extent over the scalp in which
these differences are observed and (ii) confirm that the
overall distribution of our responses is consistent with
the typical MMN topographical patterns. Given that such
extensive intercategory variation in the standards is rarely
tested, it is also important to provide a more complete pic-
ture of the nature of these responses. The two distinct
time windows selected are within the typical MMN range:
125–225 msec and 300–400 msec. These time windows
also align with the within-block MMN and iMMN results.
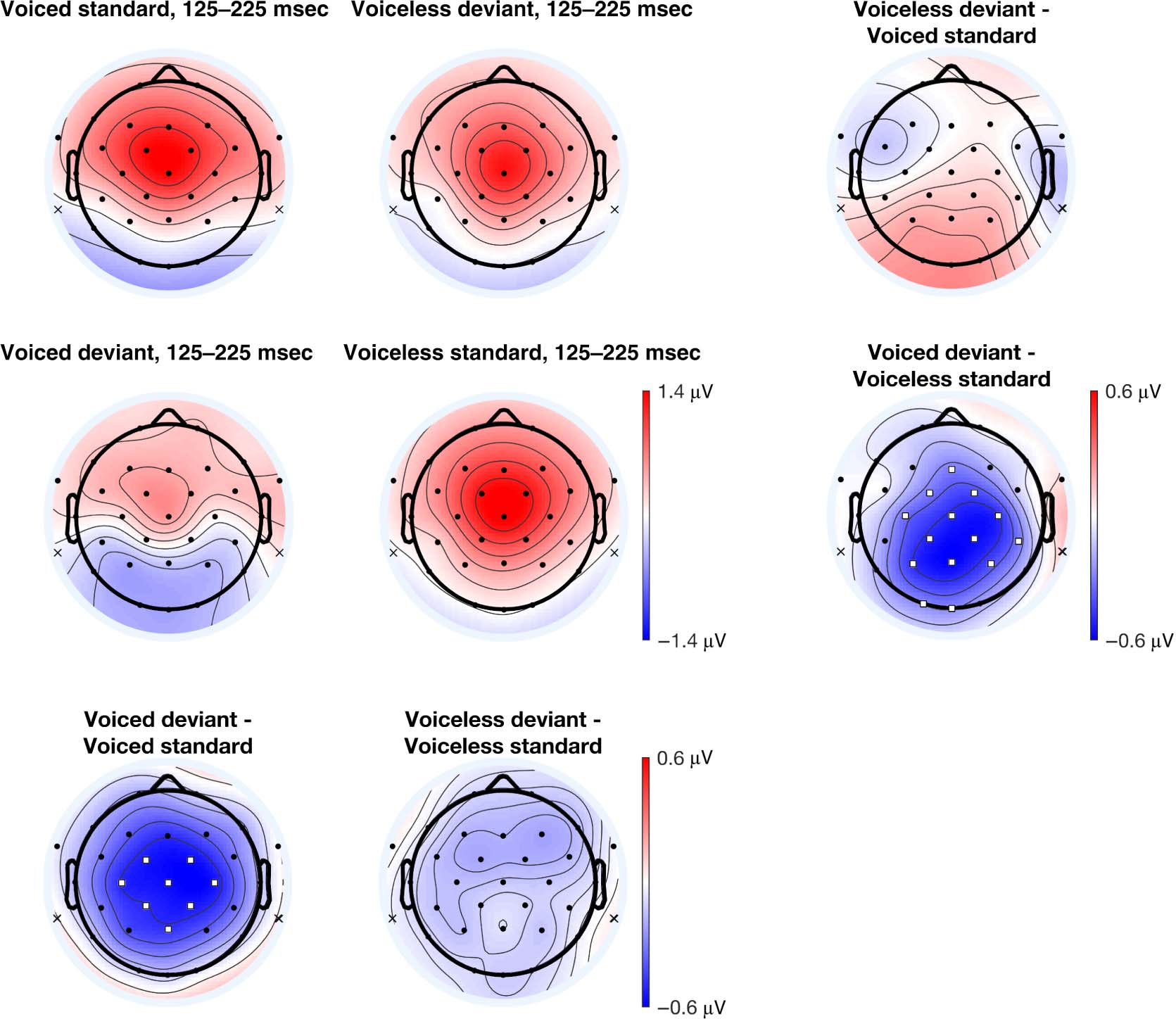
Figure 5A presents each condition’s topographical ERP
distribution in the earlier 125- to 225-msec time window.
In the within-block comparison (Figure 5C), the voiced
deviant elicited a relatively larger negativity compared to
the voiceless standard in the voiceless standard block. This
difference was distributed over frontal, central, parietal,
and occipital electrode sites. There were no differences
between the voiceless deviant and the voiced standard
in the voiced standards block and no difference between
the iMMN difference waves. In the iMMN comparison
(Figure 5B), the voiced deviant elicited a relatively larger
negativity compared to the voiced standard over central
and parietal electrode sites. No difference was observed
in the voiceless comparison. The iMMN differences are
distributed more squarely over central electrode sites in
the voiced consonants comparison. In the within-block
MMN comparison, the differences observed in the voice-
less standards block extends beyond the fronto-central
electrode sites in the ERP waveform analysis to include
posterior and occipital sites.
To assess the nature of the ERP differences in the later
time window, we also conducted an analysis in the 300- to
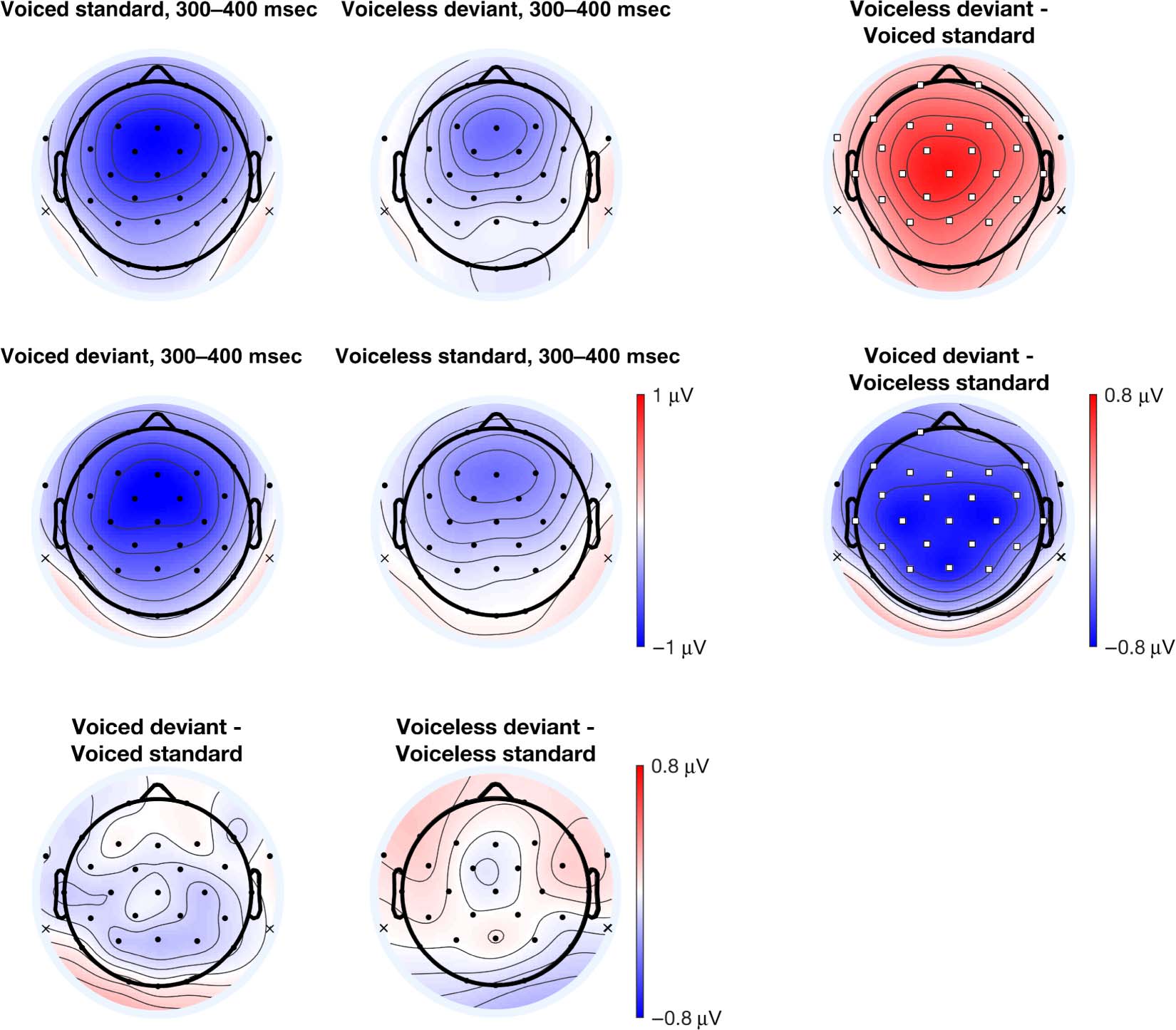
400-msec time window (Figure 6). In the within-block
comparison (Figure 6C), we observed differences in both
blocks. In the voiceless standards block, the voiced devi-
ant elicited a larger negativity relative to the voiceless stan-
dard. In the voiced standards block, the voiceless deviant
elicited a relatively larger positivity compared to the
voiced standard. This larger positivity is inconsistent with
typical MMN reports. Again, this effect was widely distrib-
uted over nearly all electrode sites. No iMMN (Figure 6B)
was observed in the later time window. In summary, the
extent of the topographic differences over the scalp in
the later time window are even greater than those in the
626
Journal of Cognitive Neuroscience
Volume 34, Number 4
earlier time window; the voiceless standards block shows
the relatively larger negativity consistent with the typical
MMN distribution, whereas the voiced standards block
shows the opposite pattern, inconsistent with the typical
MMN distribution.
Summarizing, for the within-block comparisons, the
voiceless standard block showed the canonical MMN pat-
tern in two time windows, an earlier (125–225 msec) and
later time window (300–400 msec). That is, voiced devi-
ants elicited a larger negativity when preceded by a series
of voiceless standards. Moreover, the iMMN response to
voiced stimuli when they were a deviant resulted in a
larger negativity compared to when they were the stan-
dard. On the other hand, voiceless deviants showed a
larger positivity when preceded by a series of voiced stan-
dards in the later time window but not in the earlier time
window. The timing and distribution of the larger negativ-
ity to the voiced deviants are consistent with previous
MMN reports (Näätänen et al., 2007). The larger positivity
to the voiceless deviants in the voiced standard block is
not likely an MMN, but is also consistent with previous
reports that have used an intercategory variation many-
to-one oddball paradigm for the unmarked standards
(Fu & Monahan, 2021).
ERSP
To assess whether neural activity preceding the voiced and
voiceless standards is different, we analyzed the oscillatory
power of the responses to the standards. Although we
were particularly interested in the prestimulus period
and beta-band frequencies, the analysis included a more
complete exploration of the oscillatory activity, until
700-msec poststimulus onset and frequencies between 3
and 30 Hz. Figure 7A presents these results. First, consis-
tent with our predictions, we observed decreased beta-
band oscillatory power in the prestimulus baseline
(−140 to −40 msec) for the putatively underspecified
voiced standards relative to the specified voiceless stan-
dards (see the work of Scharinger et al., 2016). Next,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. (A) Voltage topographic maps for the 125- to 225-msec time window. (B) iMMN comparisons between standards and deviants across
experimental blocks. (C) Within-block MMN comparisons between standards and deviants within experimental blocks. Electrodes highlighted in
white squares denote electrode sites with significant differences using a permutation test ( pFDR < .05). Topographic plots use the linked mastoid
reference, whose locations are marked with an ‘x’. See the Methods section for analysis procedures.
Monahan et al.
627
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
Figure 6. (A) Voltage topographic maps for the 300- to 400-msec time window. (B) iMMN comparisons between standards and deviants across
experimental blocks. (C) Within-block MMN comparisons between standards and deviants within experimental blocks. Electrodes highlighted in
white squares denote electrode sites with significant differences using a permutation test ( pFDR < 0.05). Topographic plots use the linked mastoid
reference, whose locations are marked with an “x.” See the Methods section for analysis procedures.
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. (A) ERSP for both the voiced standards (left) and voiceless standards (center). Third panel illustrates the results of a statistical comparison
between the two ERSPs. The ERSP response to the voiced standards has higher oscillatory power compared to the ERSP response to the voiceless
standards in theta-band (3–10 Hz) between 150 and 200 msec. In the prestimulus period, beta-band oscillatory power differences (13–18 Hz, marked
with a dashed rectangle) are observed between the two standard conditions. The voiced standards show lower oscillatory power relative to voiceless
standards. (B) Distribution of significant channel responses (marked in red) to beta-band (13–18 Hz) oscillatory power in the prestimulus period
(−140 to −40 msec).
628
Journal of Cognitive Neuroscience
Volume 34, Number 4
voiced standards showed increased oscillatory power on
frequencies in the theta-band range (3–7 Hz) approxi-
mately from stimulus onset until 400-msec poststimulus
onset. The opposite pattern, larger oscillatory power in
the voiceless standards relative to the voiced standards,
was observed between 11 and 16 Hz in the 200- to
250-msec time window. Figure 7B provides the topo-
graphical distribution of channels that show a difference
between the two standard conditions in the beta-band
between −140- and −40-msec prestimulus onset. This
time window was chosen based on the bootstrap statistical
test. The beta-power differences localized over fronto-
central electrode sites. These differences in prestimulus
beta-power are detected using a bootstrap test and
pFDR < .05 but disappear with the more conservative per-
mutation test. In general, these results suggest the voice-
less segments provide predictive information.
DISCUSSION
The goal of the current experiment was to determine
whether auditory cortex represents abstract phonological
features. Specifically, we tested English voicing, which
codes stops and fricatives as voiced or voiceless. We used
a many-to-one oddball MMN design to test if listeners con-
struct a unified auditory memory representation of distinct
phonetic cues that are functionally linked to one phonol-
ogical feature. The key feature of the design is that inter-
category variation is introduced into the standards. A
many-to-one relationship only exists if spectral and tempo-
ral phonetic cues to voicing are grouped together. This
many-to-one relationship would lead to an MMN effect.
The absence of an MMN would suggest no perceptual
grouping. Below, we discuss the three primary findings
from the current study.
First, in the context of voiceless standards, the neuro-
physiological response reflects a disjunctive encoding of
a spectral and temporal phonetic cue that is functionally
linked: periodic, low-frequency spectral energy in frica-
tives, and VOT in stops. A larger negative deflection
over fronto-central electrode sites was observed in two
distinct time windows to the voiced deviants in the con-
text of a series of voiceless standards: 116–196 msec and
316–440 msec. The timing and topographic distribution
of these responses indicate that these are the MMN
(Näätänen & Kreegipuu, 2012; Näätänen et al., 2007;
Näätänen, 2001). Observing the MMN suggests that the
auditory memory trace for the standards was constructed
based on the voicing feature, despite its distinct phonetic
implementation in stops versus fricatives. That is, spectral
and temporal phonetic cues can be disjunctively coded
when they are functionally linked in the phonology.
Second, an MMN was observed only in the voiceless
standards block. Asymmetric MMNs of this type are not
uncommon. As noted above, previous MMN results have
found that in English, voiced stop (Hestvik & Durvasula,
2016) and fricative (Schluter et al., 2017) deviants elicit a
larger MMN in the context of a series of voiceless stops or
fricatives, respectively. The conclusion drawn is that in
English, voiceless consonants are specified for their voic-
ing with the feature [voiceless], whereas voiced conso-
nants are underspecified for voicing (Avery & Idsardi,
2001; Iverson & Salmons, 1995). Considering these previ-
ous findings, the observation in the current study that the
voiced deviant elicited a negative deflection in the voice-
less standards block was predicted. Differences were larg-
est over fronto-central electrode sites, where the MMN is
typically observed. The presence of an MMN in the 100- to
200-msec time window is consistent with the time course
of a number of studies using the MMN to investigate
speech sound categories (Cornell et al., 2011; Ylinen,
Shestakova, Huotilainen, Alku, & Näätänen, 2006; Phillips
et al., 2000; Winkler et al., 1999; Näätänen et al., 1997). The
later negative deflection has also been observed in oddball
experiments that expose listeners to intercategory varia-
tion in the standards (Fu & Monahan, 2021), which might
require listeners to tap into putatively phonemic levels of
representation (Kazanina et al., 2006) or perceptually group
experimental stimuli based on dialect (Lanwermeyer et al.,
2016; Scharinger, Monahan, & Idsardi, 2011).
In a comparison of the standards, the voiced standards
elicited a larger negativity in an early time window
(∼100-msec poststimulus onset) and a later time window
(∼300- to 430-msec poststimulus onset). It is possible that
the differences in the standards comparison are because
of differences in the obligatory auditory evoked potential
responses to the voiced consonants as a class compared to
the voiceless consonants as a class. First, however, MMN
and repetition suppression (Larsson & Smith, 2012;
Gruber, Malinowski, & Müller, 2004) experiments suggest
that repetition fundamentally alters responses to exoge-
nous stimuli. Moreover, it is possible, for example, that
repetition intrinsically affects one category differently
from the other. This is especially relevant in designs where
asymmetric responses are predicted, as in the current
experiment. Second, the responses in the late time
window here are consistent with previous findings (Fu &
Monahan, 2021). There, the deviant in the underspecified
standard block elicited a positivity in the ∼300- to
400-msec time window, and the deviant in the specified
standard block elicited a negativity in a similar time win-
dow. That experiment and the current experiment used
two distinct features (i.e., [retroflex] vs. [voiceless]) from
two distinct languages (i.e., Mandarin Chinese vs. English).
That said, we cannot entirely rule out an auditory evoked
potential account of the standards in the within-block
analysis.
To address this possibility, an iMMN analysis was con-
ducted. While the within-block MMN contrast compares
standards and deviants from within the same block
(Scharinger et al., 2012, 2016; Kazanina et al., 2006;
Näätänen et al., 1997; Kraus, McGee, Sharma, Carrell, &
Nicol, 1992), it is possible that the MMN could be partially
driven by intrinsic ERP responses to different acoustic–
Monahan et al.
629
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
phonetic (or even phonological) properties of the stimuli.
An iMMN analysis, however, putatively eliminates the
influence of stimulus intrinsic properties by comparing
the responses to the same category (or class of speech
categories) to itself when in the standard and deviation
position. Similar experiments testing phonetic and pho-
nological representations have also reported the iMMN
(Fu & Monahan, 2021; Hestvik & Durvasula, 2016; Cornell
et al., 2011, 2013). In the current iMMN analysis, the ERP to
the voiced deviants was more negative relative to the ERP
to the voiced standards around 200-msec poststimulus
onset. No such differences were observed in the compar-
ison of the voiceless consonants. The difference in the
iMMN suggests that the results of the within-block MMN
comparison cannot be solely because of intrinsic physical
differences between voiced stops and fricatives as a class,
relative to voiceless stops and fricatives.
As noted above, Fu and Monahan (2021) also report an
asymmetric MMN with intercategory variation in the
standards but tested retroflex consonants in Mandarin
Chinese. There, Mandarin-speaking participants were
exposed to intercategory variation in the standards. An
MMN was observed only in the retroflex standard block,
where retroflex consonants were the standards and non-
retroflex consonants were the deviants. No MMN was
observed when nonretroflex consonants were the stan-
dards and retroflex consonants were the deviants. It was
concluded that the feature [retroflex] was extracted from
the sequence of standards in the retroflex standards block
and served as the basis for the auditory memory trace.
Whereas intercategory variation was used in the standards,
stops, affricates and approximants all acoustically code
retroflex similarly, that is, lower spectral energy ranges.
Like the current experiment, an MMN in the retroflex
standard block was observed in a later time window
(256–380 msec). There, it was argued that the later nega-
tivity might reflect the added task difficulty involved with
having to group various phonetic categories with one
another based on a single phonetic feature. Negative
deflections in this study also occurred around 316- to
440-msec poststimulus onset, which is later than the
normal MMN window (Strotseva-Feinschmidt, Cunitz,
Friederici, & Gunter, 2015; Martynova, Kirjavainen, &
Cheour, 2003; Čeponienė et al., 2002; Cheour, Korpilahti,
Martynova, & Lang, 2001). Late negativities have been
observed in both speech (Fu & Monahan, 2021; Datta
et al., 2020; Hestvik & Durvasula, 2016) and nonspeech
paradigms (Peter, McArthur, & Thompson, 2012; Zachau
et al., 2005). These late negativities typically appear with
the within-block MMN but can also appear independently
(Strotseva-Feinschmidt et al., 2015). Bishop, Hardiman,
and Barry (2011) suggest that late negativities might
appear as a result of additional processing required by
certain features of stimuli that are difficult to detect.
Gomes et al. (1995), who included variation across mul-
tiple cues in the standards, observed an MMN at approxi-
mately 250-msec poststimulus onset. This is later than
other studies that used sinusoids as their stimuli (e.g.,
Sams, Paavilainen, Alho, & Näätänen, 1985). In another
varying standards design, Hu, Gu, Wong, Tong, and
Zhang (2020) observed a visual MMN in the 230- to
290-msec time window when participants could group dis-
tinct, visually presented lexical items belonging to the
same semantic class (e.g., colors vs. tastes). The later time
course of differences in the current experiment might
reflect the more complex integration of multiple catego-
ries into a single memory trace. Alternatively, these late
negativities might also reflect the increased time taken
to accumulate sufficient information to identify the stimu-
lus as deviating from the standard. It should also be noted
that in the current experiment, we observed a positivity in
the voiced standards block, which is putatively the block in
which the standards could not be grouped together if
voiced consonants lack a specification for voicing. In
another study, we also observed a larger positivity to the
deviant in the underspecified standard block in a similar
time window, that is, 320–364 msec (Fu & Monahan,
2021). The topography and time course of the positivity
response are similar across the two experiments. Previous
experiments on speech sound categories have reported
differences in the P3 time window (Friedman, Cycowicz,
& Gaeta, 2001; Escera, Alho, Schröger, & Winkler, 2000),
but these often include overt tasks that require attention
even if they are irrelevant to the standard or deviant stimuli
( Winkler et al., 2003). Given the passive nature of the cur-
rent design, the P3a, which is often elicited as an orienting
response in oddball tasks with an overt task (Polich, 1993,
2007), was not predicted and is also not present in the cur-
rent iMMN comparisons. Moreover, the substantive pho-
netic variation in our standards and deviants likely reduced
the saliency of a change from standard to deviant and, as
such, was potentially too subtle to draw participants’ atten-
tion. To summarize our second finding, we observed an
asymmetric MMN in both the within-block MMN and
iMMN analyses, consistent with previous findings on
English voicing in stops and fricatives, independently, as
well as previous findings that have used intercategory
variation in the standards. Moreover, the effects observed
in the later time windows may reflect the additional
processing demands and complexity that result from the
current design.
Third, linguistic features appear to be used to generate
predictions about the incoming auditory stimulus. This
predictiveness occurs despite physical variation in the
repeating standard auditory stimuli ( Winkler et al.,
1990). Predictions are created even when the standards
remain constant along one physical dimension while vary-
ing in others, for example, when remaining constant in
duration but varying in intensity and frequency (Nousak,
Deacon, Ritter, & Vaughan, 1996; Gomes et al., 1995).
This result is consistent with previous findings of infor-
mation propagation from higher-level regions of the
functional hierarchy observable in prestimulus intervals
(Bastos et al., 2015) and in the activation of higher-level
630
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
linguistic representations during language comprehension
(Molinaro, Monsalve, & Lizarazu, 2016). This prestimulus
differential neural activity in beta-band could be consid-
ered a hallmark of predictive coding mechanisms. That
is, abstract characteristics of the speech sound repre-
sentation are being predicted, perhaps demonstrating
an instance of “what” cortical hypothesis generation
(Doelling et al., 2014). The MMN is argued to be based
on predictive neurophysiological models, where sensory
cues and abstract information combine to produce predic-
tions about upcoming sounds (Winkler, 2007). If the MMN
fundamentally reflects predictive mechanisms (Wacongne
et al., 2011; Garrido, Kilner, Kiebel, & Friston, 2009;
Winkler, 2007; Friston, 2005), the response to the deviant
then indicates a predictive coding error with the con-
structed neural representation of the standard. As such, it
is predicted that differences in beta-band oscillations
should be observed within an MMN paradigm.
Beta-power in electrophysiological responses has been
taken to indicate information propagation from higher-
order brain areas and levels of representation (Riddle
et al., 2019; Bidelman, 2015; Lewis, Wang, & Bastiaansen,
2015; Fontolan et al., 2014; Arnal & Giraud, 2012; Arnal
et al., 2011; Buschman & Miller, 2007; Engel & Fries,
2010; Wang, 2010). In addition, beta-band oscillatory
power increases if the system maintains the actual lan-
guage stimuli in memory, and it will decrease if the current
cognitive state is interrupted by novel and/or unexpected
stimuli. Consequently, a beta-band oscillatory power
decrease predicts the probability of new processing
demands (Engel & Fries, 2010). Scharinger et al. (2016)
observed increased prestimulus beta-power when the
standard was specified (i.e., [ɪ]) and decreased prestimu-
lus beta-power when the standard was underspecified
(i.e., [ɛ]). Here, we also observed a similar increase in
prestimulus beta-power in the block where the standards
were the specified, that is, voiceless consonants, and a
decrease in beta-power prestimulus onset when the stan-
dards were the underspecified, that is, voiced consonants.
The specification for voiceless consonants likely forms the
basis of the auditory memory trace and is used to predict
the upcoming speech sound. Moreover, it also potentially
reflects the added maintenance of linguistic stimuli in
memory, as they are marked with the feature [voiceless].
This is compared to the voiced consonants, which are
underspecified for voicing. Furthermore, we observed
reduced theta-power in the voiceless standards relative
to the voiced standards, which is consistent with the con-
struction of a short-term auditory memory trace in the
voiceless standards block. Although we did not have pre-
dictions regarding differences in theta-band power, this is
in a similar time window and frequency band as the MMN.
Positing that voiceless consonants are marked is consis-
tent with proposals from linguistic theory (Avery & Idsardi,
2001; Iverson & Salmons, 1995). Given the ERP findings,
the voiced deviants elicited a larger MMN in the context
of the series of voiceless standards. If the voiceless
standards are marked with the feature [voiceless] and this
feature results in greater habituation because of the
many-to-one organization of the standards, we expected
increased beta-band oscillatory power in the voiceless
standards block relative to the voiced standards block. In
the ERSP results, the voiceless standards did elicit an
increase in prestimulus beta-power compared to the
voiced standards. Consistent with the ERP results, where
an MMN was elicited only in the voiceless standards block,
we attribute this increase in prestimulus beta-power to the
voiceless standards in the oscillatory response to the
marked feature [voiceless] that could be extracted and
grouped. Again, this is potentially because of the construc-
tion of a short-term auditory memory trace to the voiceless
standards, considering that the voiceless standards had
the marked feature [voiceless] that could be extracted
and grouped.
Not all accounts of neurophysiological and perceptual
asymmetries rely on featural underspecification. Previous
neurophysiological asymmetries have been attributed to
low-level feature detectors based on specific acoustic
properties that differ between standards and deviants
(see the work of Maiste et al., 1995, for spectral differences
between [ba] and [da]; Hisagi, Shafer, Strange, &
Sussman, 2010, 2015; see the work of Ylinen et al., 2006,
for vowel duration differences). Given that we did not find
reliable unified differences separating voiced and voiceless
stimuli in any of the acoustic dimensions that we mea-
sured in our stimuli, this is unlikely to be able to provide
a plausible explanation for the effects found in this study.
Moreover, if voiceless sounds are indeed specified for
their voicing and voiced sounds are underspecified, our
results are opposite of the work of Bishop, O’Reilly, and
McArthur (2005). There, an MMN was observed only when
frequency-modulated tones—which are presumably
marked for an additional acoustic property—served as
deviants in the context of unmodulated sinusoid stan-
dards, whereas no MMN was observed in the opposite
configuration.
Perceptual asymmetries have also been observed in
vowel perception, with discrimination performance
depending on the order in which the stimuli are pre-
sented. Several behavioral studies have shown that it is
easier to discriminate contrasts that move from more cen-
tral to more peripheral, and thus more acoustically focal,
regions of the vowel space than when the same stimuli are
presented in the opposite direction (Masapollo, Polka,
Molnar, & Ménard, 2017; Polka & Bohn, 2003, 2011); how-
ever, neurophysiological evidence for these asymmetries
is less robust (de Rue et al., 2021; Polka, Molnar, Zhao, &
Masapollo, 2021; Riedinger, Nagels, Werth, & Scharinger,
2021). In addition to vowel peripherality and focality,
prototypicality and frequency also result in perceptual
asymmetries (Kuhl et al., 2008; Kuhl, Williams, Lacerda,
Stevens, & Lindblom, 1992). It is unclear, however, how
easily notions of peripherality, centrality, and prototypi-
cally in a two-dimensional vowel space translate into the
Monahan et al.
631
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
consonant domain, where the space is less continuous
and potentially more multidimensional.
Frequency might also be a factor in the current results.
Both lexical frequency, that is, how common a phoneme
or a word is in a language, and cross-linguistic frequency,
that is, how common a phoneme is across the world’s lan-
guages, have been shown to affect MMN results. Lexical
frequency appears to result in asymmetric neurophysio-
logical responses ( Jacobsen et al., 2004, 2021; Alexandrov,
Boricheva, Pulvermüller, & Shtyrov, 2011; Jacobsen,
Schröger, Winkler, & Horváth, 2005). If larger MMNs are
observed to higher-frequency deviants relative to lower-
frequency deviants (Alexandrov et al., 2011), then we
might predict a larger MMN to the voiceless deviants, as
the voiceless consonants in our study are more frequent
in English than the voiced consonants ( Wang & Crawford,
1960). We, however, found the opposite pattern: Voiced
deviants in the context of voiceless standards resulted in
larger MMNs. It is also possible that the cross-linguistic fre-
quency of a given phoneme or phoneme class might affect
MMN responses. For example, Shafer, Schwartz, and
Kurtzberg (2004) observed an earlier negativity in Hindi
participants compared to English participants when retro-
flex [ɖ] served as the standard and [b] was the deviant,
compared to the opposite configuration. They posit that
this asymmetry might be because retroflex consonants
are cross-linguistically less common. In the context of
the current experiment, voiced consonants are cross-
linguistically less common than voiceless consonants
(Maddieson, 1984). Given the Shafer et al. (2004) results,
this might predict earlier/ larger MMNs to the higher-
frequency voiceless deviants in the context of the lower-
frequency voiced standards. Again, we observed the
opposite pattern, suggesting that neither lexical frequency
nor cross-linguistic frequency is principally responsible
for the effects observed in the current experiment.
There is now emerging evidence favoring not only
underspecified features being stored, as described above,
but also cross-linguistic evidence that auditory cortex sup-
ports featural underspecification in distinct laryngeal sys-
tems. Using Japanese, where [voiced] is the putatively
marked feature, Hestvik et al. (2020) observed a larger
MMN when voiced stops were the standard compared to
when voiceless stops were the standard. And using
Danish, Højlund, Gebauer, McGregor, and Wallentin
(2019) observed a larger MMN when [d] was the standard
compared to when [th] was the standard. They posit that
this is potential evidence for the claim that voiced conso-
nants are marked in Danish, whereas voiceless consonants
are unmarked.
Future research could include designs that synthetically
minimize acoustic differences between different speech
sound classes. The goal would be to further isolate the
contribution of the abstract phonological feature [voice-
less] in the elicitation of the MMN in intercategory oddball
designs. For the items in the current experiment, individ-
ual secondary acoustic–phonetic cues to voicing do not
reliably predict voicing category membership (see
Table 1 and Figure 2); however, it is possible that multiple
secondary cues (e.g., f0 and F1, duration and f0) might
conspire to provide a reliable signal-driven cue to phonol-
ogical voicing. If true, this would indicate that participants
could rely on a combination of acoustic–phonetic cues to
identify phonological category membership. This could be
tested with a design systematically controlling all acoustic
cues in the stimuli. Moreover, different languages could be
brought to bear on the current research questions. For
example, Spanish and English speakers could be com-
pared on the Spanish and English voicing contrasts. Recall
that Spanish voiced stops and voiced fricatives use the
same phonetic cue to phonological voicing, whereas
English stops and fricatives use distinct phonetic cues.
Such cross-linguistic comparisons could further empha-
size the role that (i) the native language voicing inventory
plays in the construction of auditory memory traces that
reflect abstract phonological features and (ii) the distribu-
tion of different phonetic cues plays in the construction of
phonological features. If acoustic–phonetic features,
rather than an abstract phonological feature, were the pri-
mary drivers of the MMN effect, we would expect to find
similar performance by the two language groups when
listening to the same stimuli. On the other hand, if pho-
nology is the main driver, as we expect, we would posit
different performance across the two groups.
A limitation of the current study is that given the num-
ber of deviants used in the current design, it is difficult to
know whether stops or fricatives played distinct roles in
the perceptual grouping of voicing or the elicitation of
the MMN. Each block contained 75 deviants. Forty-five of
these deviants were stops and 30 were fricatives. This
results in lower than desired power to determine their
independent effects. For example, given the distinct
nature of how voicing is coded in stops versus fricatives,
it might be the case that voiced stops resulted in a larger
MMN than voiced fricatives in the context of voiceless
standards; however, the ratios of stops to fricatives were
similar in the standards and deviants across both blocks,
and so this is unlikely to be the case. At this point, we do
not have clear predictions about which manner of articu-
lation might have an increased effect on the MMN, but the
current design makes it difficult to ascertain. We leave this
possibility for future research.
The current results support theories of language pro-
cessing that posit that speech sounds are stored in terms
of featural representations and that these features are
utilized during on-line spoken word recognition (Hickok,
2014; Poeppel & Monahan, 2011; Gow, 2003; Stevens,
2002; McClelland & Elman, 1986; Halle & Stevens, 1962).
Whether researchers should search for linguistic units of
representation in perception is contentious. In particular,
Goldinger and Azuma (2003) and Samuel (2020) have
argued against positing the phoneme and, to a lesser
extent, the feature as a unit of language processing and
perception. Their position is that the search for identifying
632
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
units as conceptualized in linguistics within perception is
misguided. As reviewed above, however, there is a body
of neurophysiological evidence that is consistent with
feature-based speech sound representations. Moreover,
various models of feature representations (e.g., underspe-
cification) make specific predictions regarding neurophys-
iological response asymmetries, and these patterns have
been replicated across several speech sound classes and
in different languages (see the work of Monahan, 2018,
for a review). In particular, the current results indicate a
brain signature underlying abstract groupings of speech
sounds and are consistent with predictive theories of
processing that are at least partially based on phonological
features, such as analysis by synthesis (Halle & Stevens,
1962; Poeppel & Monahan, 2011).
Conclusion
The current experiment provides neurophysiological evi-
dence that distinct phonetic cues that are functionally
linked in a language’s phonology are grouped together.
This is shown by the presence of an MMN when hearing
deviants that differ from the standards in their phonolo-
gical voicing. This is true even when the speech sounds
encompass phonetically distinct realizations of a single
phonological feature. The MMN was asymmetric, consis-
tent with previous studies arguing for underspecified
featural representations. Here, the results support the
conclusion that voiceless consonants in English are speci-
fied as [voiceless], whereas voiced consonants are under-
specified for voicing. Moreover, we observed a difference
between the voiced and voiceless standards in the prestim-
ulus baseline, suggesting that predictions regarding
which sounds come next can be constructed from speci-
fied features. These results point toward auditory cortex’s
ability to disjunctively code spectral and temporal
phonetic cues when they are functionally joined and,
ultimately, to encode abstract phonological features. In
summary, the current work demonstrates that linguistic
units and, specifically, features do have a role to play in
understanding the cognitive neuroscience of language.
APPENDIX A
We calculated the ERPs to the four standard conditions
(i.e., voiced fricative standards, voiced stop standards,
voiceless fricative standards, and voiceless stop stan-
dards). The ERPs are presented in Figure A1 (rug plot,
top: voicing; rug plot, bottom: manner). As expected,
there are differences in the responses between the four
conditions. Most time points showed a main effect of
manner. A main effect of voicing was observed in three
principal time windows: 68–160 msec, 212–252 msec,
and 292–448 msec. There is a question as to whether
certain subcategories of speech sounds (e.g., voiced frica-
tives) could drive the MMN responses reported in the
article. To address this, we computed the ratios of stops
Figure A1. Average evoked ERP responses over fronto-central electrode
sites (i.e., Fz, FC1/2, Cz, C3/4, CP1/2) to the voiced fricatives, voiced
stops, voiceless fricatives, and voiceless stops. The ERP responses
to fricatives are indicated with solid lines, and the ERP responses to
stops are indicated with dotted lines. The ERP responses to voiceless
consonants are indicated in orange, and the ERP responses to voiced
consonants are indicated in purple. Rug plots along the x axes indicate
time points when the comparison was statistically significant (see text
for analysis procedures). Rug plots along the top of the figure indicate
time points in which there is a main effect of voicing. Rug plots along
the bottom of the figure indicate time points in which there is a main
effect of manner. Shaded regions represent the 95% confidence interval
of the mean.
to fricatives in the standards and deviants after data pre-
processing. If there were substantial imbalances between
these ratios in the standards and deviants, then this could
potentially result in some subcategories driving the MMN
response more than others. The ratio of stops to fricatives
in the voiced standards (1:1.44) is similar to the ratio of
stops to fricatives in the voiceless deviants (1:1.47). The
same is true for the voiceless standards (1:1.52) and voiced
deviants (1:1.54). The differences in stop–fricative ratios in
the iMMN comparisons are slightly larger (voiceless stan-
dards: 1:1.52, voiceless deviants: 1:1.47; voiced standards:
1:1.44, voiced deviants: 1:1.54), but again, quite similar.
Given that the stop–fricative ratios are similar between
standards and deviants, it is unlikely that any one subcate-
gory is driving the MMN when comparing standards to
deviants.
Acknowledgments
We thank members of the Computation and Psycholinguistics
Laboratory at the University of Toronto Scarborough for their
assistance.
Reprint requests should be sent to Philip J. Monahan, Depart-
ment of Language Studies, University of Toronto Scarborough,
1265 Military Trail, Toronto ON M1C 1A4, Canada, or via e-mail:
philip.monahan@utoronto.ca.
Author Contributions
Philip J. Monahan: Conceptualization; Formal analysis;
Funding acquisition; Investigation; Methodology;
Monahan et al.
633
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Resources; Supervision; Visualization; Writing—Original
draft; Writing—review & editing. Jessamyn Schertz:
Conceptualization; Investigation; Methodology; Writing—
review & editing. Zhanao Fu: Formal analysis; Investigation;
Methodology; Visualization; Writing—review & editing.
Alejandro Pérez: Formal analysis; Investigation; Method-
ology; Visualization; Writing—Original draft; Writing—
review & editing.
Funding Information
A. P. received funding from the European Union’s Horizon
2020 Research and Innovation Programme under the
Marie Skłodowska-Curie grant number: 840885. This
work was supported in part by the Social Sciences and
Humanities Research Council (SSHRC) (http://dx.doi
.org/10.13039/501100000155) of Canada, grant number:
IDG 430-15-00647, and in part by the Natural Sciences
and Engineering Research Council (NSERC) of Canada,
grant number: RGPIN-2017-06053.
Data Availability Statement
Stimuli, acoustic measurements, and EEG datasets are
publicly available in the Open Science Framework:
https://osf.io/jn5cr/.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN ) during this period
were M(an)/M = .407, W(oman)/M = .32, M/W = .115, and
W/W = .159, the comparable proportions for the articles
that these authorship teams cited were M/M = .549,
W/M = .257, M/W = .109, and W/W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly
when selecting which articles to cite and gives them the
opportunity to report their article’s gender citation
balance. The authors of this article report its proportions
of citations by gender category to be as follows: M/M =
.606, W/M = .213, M/ W = .096, and W/ W = .085.
REFERENCES
Alexandrov, A. A., Boricheva, D. O., Pulvermüller, F., & Shtyrov,
Y. (2011). Strength of word-specific neural memory traces
assessed electrophysiologically. PLoS One, 6, e22999. https://
doi.org/10.1371/journal.pone.0022999, PubMed: 21853063
Alho, K. (1995). Cerebral generators of mismatch negativity
(MMN) and its magnetic counterpart (MMNm) elicited by
sound changes. Ear and Hearing, 16, 38–51. https://doi.org
/10.1097/00003446-199502000-00004, PubMed: 7774768
Archangeli, D. (1988). Aspects of underspecification
theory. Phonology, 5, 183–207. https://doi.org/10.1017
/S0952675700002268
Arnal, L. H., & Giraud, A.-L. (2012). Cortical oscillations and
sensory predictions. Trends in Cognitive Sciences, 16,
390–398. https://doi.org/10.1016/j.tics.2012.05.003, PubMed:
22682813
Arnal, L. H., Wyart, V., & Giraud, A.-L. (2011). Transitions in
neural oscillations reflect prediction errors generated in
audiovisual speech. Nature Neuroscience, 14, 797–801.
https://doi.org/10.1038/nn.2810, PubMed: 21552273
Aulanko, R., Hari, R., Lounasmaa, O. V., Näätänen, R., & Sams,
M. (1993). Phonetic invariance in the human auditory
cortex. NeuroReport, 4, 1356–1358. https://doi.org/10.1097
/00001756-199309150-00018, PubMed: 8260620
Avery, P., & Idsardi, W. J. (2001). Laryngeal dimensions,
completion and enhancement. In T. A. Hall (Ed.), Distinctive
feature theory (pp. 41–70). Berlin: Walter de Gruyter. https://
doi.org/10.1515/9783110886672.41
Baković, E. (2014). Phonemes, segments and features. Language,
Cognition and Neuroscience, 29, 21–23. https://doi.org/10
.1080/01690965.2013.848992
Bastos, A. M., Vezoli, J., Bosman, C. A., Schoffelen, J.-M.,
Oostenveld, R., Dowdall, J. R., et al. (2015). Visual areas
exert feedforward and feedback influences through distinct
frequency channels. Neuron, 85, 390–401. https://doi.org/10
.1016/j.neuron.2014.12.018, PubMed: 25556836
Bidelman, G. M. (2015). Induced neural beta oscillations predict
categorical speech perception abilities. Brain and Language,
141, 62–69. https://doi.org/10.1016/j.bandl.2014.11.003,
PubMed: 25540857
Bigdely-Shamlo, N., Mullen, T., Kothe, C., Su, K.-M., & Robbins,
K. A. (2015). The PREP pipeline: Standardized preprocessing for
large-scale EEG analysis. Frontiers in Neuroinformatics, 9, 16.
https://doi.org/10.3389/fninf.2015.00016, PubMed: 26150785
Bishop, D. V. M., Hardiman, M. J., & Barry, J. G. (2011). Is
auditory discrimination mature by middle childhood? A study
using time–frequency analysis of mismatch responses from
7 years to adulthood: Is auditory discrimination mature?
Developmental Science, 14, 402–416. https://doi.org/10.1111
/j.1467-7687.2010.00990.x, PubMed: 22213909
Bishop, D. V. M., O’Reilly, J., & McArthur, G. M. (2005).
Electrophysiological evidence implicates automatic low-level
feature detectors in perceptual asymmetry. Cognitive Brain
Research, 24, 177–179. https://doi.org/10.1016/j.cogbrainres
.2004.12.007, PubMed: 15922169
Buschman, T. J., & Miller, E. K. (2007). Top–down versus
bottom–up control of attention in the prefrontal and
posterior parietal cortices. Science, 315, 1860–1862. https://
doi.org/10.1126/science.1138071, PubMed: 17395832
Čeponienė, R., Yaguchi, K., Shestakova, A., Alku, P., Suominen,
K., & Näätänen, R. (2002). Sound complexity and
‘speechness’ effects on pre-attentive auditory discrimination
in children. International Journal of Psychophysiology, 43,
199–211. https://doi.org/10.1016/S0167-8760(01)00172-6,
PubMed: 11850086
Cheour, M., Korpilahti, P., Martynova, O., & Lang, A.-H. (2001).
Mismatch negativity and late discriminative negativity in
investigating speech perception and learning in children and
infants. Audiology and Neuro-Otology, 6, 2–11. https://doi
.org/10.1159/000046804, PubMed: 11173771
Chomsky, N., & Halle, M. (1968). The sound pattern of English.
Cambridge, MA: MIT Press.
Clements, G. N., & Hume, E. V. (1995). The internal organization
of speech sounds. In J. A. Goldsmith (Ed.), The handbook of
phonological theory (pp. 245–306). Oxford: Blackwell.
Cornell, S. A., Lahiri, A., & Eulitz, C. (2011). “What you encode is
not necessarily what you store”: Evidence for sparse feature
634
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
representations from mismatch negativity. Brain Research,
1394, 79–89. https://doi.org/10.1016/j.brainres.2011.04.001,
PubMed: 21549357
Cornell, S. A., Lahiri, A., & Eulitz, C. (2013). Inequality across
consonantal contrasts in speech perception: Evidence from
mismatch negativity. Journal of Experimental Psychology:
Human Perception and Performance, 39, 757–772. https://
doi.org/10.1037/a0030862, PubMed: 23276108
Cowan, N., Winkler, I., Teder, W., & Näätänen, R. (1993).
Memory prerequisites of mismatch negativity in the auditory
event-related potential (ERP). Journal of Experimental
Psychology: Learning, Memory, and Cognition, 19, 909–921.
https://doi.org/10.1037//0278-7393.19.4.909, PubMed: 8345328
Datta, H., Hestvik, A., Vidal, N., Tessel, C., Hisagi, M., Wróblewski,
M., et al. (2020). Automaticity of speech processing in
early bilingual adults and children. Bilingualism: Language
and Cognition, 23, 429–445. https://doi.org/10.1017
/S1366728919000099, PubMed: 32905492
Deacon, D., Gomes, H., Nousak, J. M., Ritter, W., & Javitt, D.
(2000). Effect of frequency separation and stimulus rate on
the mismatch negativity: An examination of the issue of
refractoriness in humans. Neuroscience Letters, 287,
167–170. https://doi.org/10.1016/S0304-3940(00)01175-7,
PubMed: 10863021
Dehaene-Lambertz, G. (2000). Cerebral specialization for
speech and non-speech stimuli in infants. Journal of
Cognitive Neuroscience, 12, 449–460. https://doi.org/10.1162
/089892900562264, PubMed: 10931771
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21. https://doi.org/10.1016/j.jneumeth.2003
.10.009, PubMed: 15102499
Delorme, A., Palmer, J., Onton, J., Oostenveld, R., & Makeig, S.
(2012). Independent EEG sources are dipolar. PLoS One, 7,
e30135. https://doi.org/10.1371/journal.pone.0030135,
PubMed: 22355308
de Rue, N. P. W. D., Snijders, T. M., & Fikkert, P. (2021).
Contrast and conflict in Dutch vowels. Frontiers in Human
Neuroscience, 15, 629648. https://doi.org/10.3389/fnhum
.2021.629648, PubMed: 34163338
Docherty, G. J. (1992). The timing of voicing in British English
obstruents. Berlin: Mouton de Gruyter.
Doelling, K. B., Arnal, L. H., Ghitza, O., & Poeppel, D. (2014).
Acoustic landmarks drive delta–theta oscillations to enable
speech comprehension by facilitating perceptual parsing.
Neuroimage, 85, 761–768. https://doi.org/10.1016/j
.neuroimage.2013.06.035, PubMed: 23791839
Engel, A. K., & Fries, P. (2010). Beta-band oscillations—Signalling
the status quo? Current Opinion in Neurobiology, 20, 156–165.
https://doi.org/10.1016/j.conb.2010.02.015, PubMed: 20359884
Escera, C., Alho, K., Schröger, E., & Winkler, I. W. (2000).
Involuntary attention and distractibility as evaluated with
event-related brain potentials. Audiology and Neurotology,
5, 151–166. https://doi.org/10.1159/000013877, PubMed:
10859410
Eulitz, C., & Lahiri, A. (2004). Neurobiological evidence for
abstract phonological representations in the mental lexicon
during speech recognition. Journal of Cognitive Neuroscience,
16, 577–583. https://doi.org/10.1162/089892904323057308,
PubMed: 15185677
Fontolan, L., Morillon, B., Liegeois-Chauvel, C., & Giraud, A.-L.
(2014). The contribution of frequency-specific activity to
hierarchical information processing in the human auditory
cortex. Nature Communications, 5, 4694. https://doi.org/10
.1038/ncomms5694, PubMed: 25178489
Fox, N. P., Leonard, M., Sjerps, M. J., & Chang, E. F. (2020).
Transformation of a temporal speech cue to a spatial neural
code in human auditory cortex. eLife, 9, e53051. https://doi
.org/10.7554/eLife.53051, PubMed: 32840483
Friedman, D., Cycowicz, Y. M., & Gaeta, H. (2001). The novelty
P3: An event-related brain potential (ERP) sign of the brain’s
evaluation of novelty. Neuroscience & Biobehavioral
Reviews, 25, 355–373. https://doi.org/10.1016/S0149-7634(01)
00019-7, PubMed: 11445140
Friston, K. (2005). A theory of cortical responses. Philosophical
Transactions of the Royal Society of London, Series B:
Biological Sciences, 360, 815–836. https://doi.org/10.1098
/rstb.2005.1622, PubMed: 15937014
Fu, Z., & Monahan, P. J. (2021). Extracting phonetic features
from natural classes: A mismatch negativity study of Mandarin
Chinese retroflex consonants. Frontiers in Human
Neuroscience, 15, 609898. https://doi.org/10.3389/fnhum
.2021.609898, PubMed: 33841113
Garrido, M. I., Kilner, J. M., Kiebel, S. J., & Friston, K. J. (2009).
Dynamic causal modeling of the response to frequency
deviants. Journal of Neurophysiology, 101, 2620–2631.
https://doi.org/10.1152/jn.90291.2008, PubMed: 19261714
Goldinger, S. D., & Azuma, T. (2003). Puzzle-solving science:
The quixotic quest for units in speech perception. Journal of
Phonetics, 31, 305–320. https://doi.org/10.1016/S0095-4470
(03)00030-5, PubMed: 29093608
Gomes, H., Ritter, W., & Vaughan, H. G. (1995). The nature of
preattentive storage in the auditory system. Journal of
Cognitive Neuroscience, 7, 81–94. https://doi.org/10.1162
/jocn.1995.7.1.81, PubMed: 23961755
Gow, D. W. (2003). Feature parsing: Feature cue mapping in
spoken word recognition. Perception & Psychophysics, 65,
575–590. https://doi.org/10.3758/BF03194584, PubMed:
12812280
Gruber, T., Malinowski, P., & Müller, M. M. (2004). Modulation
of oscillatory brain activity and evoked potentials in a
repetition priming task in the human EEG. European
Journal of Neuroscience, 19, 1073–1082. https://doi.org/10
.1111/j.0953-816X.2004.03176.x, PubMed: 15009155
Halle, M. (1983). On distinctive features and their articulatory
implementation. Natural Language & Linguistic Theory, 1,
91–105. https://doi.org/10.1007/BF00210377
Halle, M. (2002). From memory to speech and back: Papers on
phonetics and phonology 1954–2002. Berlin: Walter de
Gruyter. https://doi.org/10.1515/9783110871258
Halle, M., & Stevens, K. N. (1962). Speech recognition: A model
and a program for research. IRE Transactions on
Information Theory, 8, 155–159. https://doi.org/10.1109/TIT
.1962.1057686
Hestvik, A., & Durvasula, K. (2016). Neurobiological evidence
for voicing underspecification in English. Brain and
Language, 152, 28–43. https://doi.org/10.1016/j.bandl.2015
.10.007, PubMed: 26705957
Hestvik, A., Shinohara, Y., Durvasula, K., Verdonschot, R. G., &
Sakai, H. (2020). Abstractness of human speech sound
representations. Brain Research, 1732, 146664. https://doi
.org/10.1016/j.brainres.2020.146664, PubMed: 31930995
Hickok, G. (2014). The architecture of speech production and
the role of the phoneme in speech processing. Language,
Cognition and Neuroscience, 29, 2–20. https://doi.org/10
.1080/01690965.2013.834370, PubMed: 24489420
Hisagi, M., Shafer, V. L., Strange, W., & Sussman, E. S. (2010).
Perception of a Japanese vowel length contrast by
Japanese and American English listeners: Behavioral and
electrophysiological measures. Brain Research, 1360,
89–105. https://doi.org/10.1016/j.brainres.2010.08.092,
PubMed: 20816759
Hisagi, M., Shafer, V. L., Strange, W., & Sussman, E. S. (2015).
Neural measures of a Japanese consonant length
discrimination by Japanese and American English listeners:
Monahan et al.
635
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Effects of attention. Brain Research, 1626, 218–231.
https://doi.org/10.1016/j.brainres.2015.06.001, PubMed:
26119918
Højlund, A., Gebauer, L., McGregor, W. B., & Wallentin, M.
(2019). Context and perceptual asymmetry effects on the
mismatch negativity (MMNm) to speech sounds: An MEG
study. Language, Cognition and Neuroscience, 34, 545–560.
https://doi.org/10.1080/23273798.2019.1572204
Hu, A., Gu, F., Wong, L. L. N., Tong, X., & Zhang, X. (2020).
Visual mismatch negativity elicited by semantic violations in
visual words. Brain Research, 1746, 147010. https://doi.org
/10.1016/j.brainres.2020.147010, PubMed: 32663455
Hullett, P. W., Hamilton, L. S., Mesgarani, N., Schreiner, C. E., &
Chang, E. F. (2016). Human superior temporal gyrus
organization of spectrotemporal modulation tuning derived
from speech stimuli. Journal of Neuroscience, 36,
2014–2026. https://doi.org/10.1523/JNEUROSCI.1779-15
.2016, PubMed: 26865624
Iverson, G. K., & Salmons, J. C. (1995). Aspiration and laryngeal
representation in Germanic. Phonology, 12, 369–396. https://
doi.org/10.1017/S0952675700002566
Jacobsen, T., Bäß, P., Roye, A., Winkler, I., Schröger, E., &
Horváth, J. (2021). Word class and word frequency in the MMN
looking glass. Brain and Language, 218, 104964. https://doi
.org/10.1016/j.bandl.2021.104964, PubMed: 33964668
Jacobsen, T., Horváth, J., Schröger, E., Lattner, S., Widmann, A.,
& Winkler, I. (2004). Pre-attentive auditory processing of
lexicality. Brain and Language, 88, 54–67. https://doi.org/10
.1016/S0093-934X(03)00156-1, PubMed: 14698731
Jacobsen, T., Schröger, E., Winkler, I., & Horváth, J. (2005).
Familiarity affects the processing of task-irrelevant auditory
deviance. Journal of Cognitive Neuroscience, 17, 1704–1713.
https://doi.org/10.1162/089892905774589262, PubMed:
16269107
Jakobson, R., Fant, G., & Halle, M. (1961). Preliminaries
to speech analysis: The distinctive features and their
correlates. Cambridge, MA: MIT Press.
Kazanina, N., Bowers, J. S., & Idsardi, W. J. (2018). Phonemes:
Lexical access and beyond. Psychonomic Bulletin & Review,
25, 560–585. https://doi.org/10.3758/s13423-017-1362-0,
PubMed: 28875456
Kazanina, N., Phillips, C., & Idsardi, W. J. (2006). The influence of
meaning on the perception of speech sounds. Proceedings of
the National Academy of Sciences, U.S.A., 103, 11381–11386.
https://doi.org/10.1073/pnas.0604821103, PubMed: 16849423
Khalighinejad, B., da Silva, G. C., & Mesgarani, N. (2017).
Dynamic encoding of acoustic features in neural responses to
continuous speech. Journal of Neuroscience, 37, 2176–2185.
https://doi.org/10.1523/JNEUROSCI.2383-16.2017, PubMed:
28119400
Kraus, N., McGee, T., Sharma, A., Carrell, T., & Nicol, T. (1992).
Mismatch negativity event-related potential elicited by
speech stimuli. Ear and Hearing, 13, 158–164. https://doi
.org/10.1097/00003446-199206000-00004, PubMed: 1397755
Kuhl, P. K., Conboy, B. T., Coffey-Corina, S., Padden, D., Rivera-
Gaxiola, M., & Nelson, T. (2008). Phonetic learning as a
pathway to language: New data and native language magnet
theory expanded (NLM-e). Philosophical Transactions of the
Royal Society of London, Series B: Biological Sciences, 363,
979–1000. https://doi.org/10.1098/rstb.2007.2154, PubMed:
17846016
Kuhl, P. K., Williams, K. A., Lacerda, F., Stevens, K. N., &
Lindblom, B. (1992). Linguistic experience alters phonetic
perception in infants by 6 months of age. Science, 255,
606–608. https://doi.org/10.1126/science.1736364, PubMed:
1736364
Lahiri, A., & Reetz, H. (2002). Underspecified recognition. In
C. Gussenhoven & N. Warner (Eds.), Laboratory phonology
(Vol. 7, pp. 637–675). Berlin: Mouton de Gruyter. https://doi
.org/10.1515/9783110197105.637
Lahiri, A., & Reetz, H. (2010). Distinctive features: Phonological
underspecification in representation and processing. Journal
of Phonetics, 38, 44–59. https://doi.org/10.1016/j.wocn.2010
.01.002
Lanwermeyer, M., Henrich, K., Rocholl, M. J., Schnell, H. T.,
Werth, A., Herrgen, J., et al. (2016). Dialect variation
influences the phonological and lexical-semantic word
processing in sentences. Electrophysiological evidence from
a cross-dialectal comprehension study. Frontiers in
Psychology, 7, 739. https://doi.org/10.3389/fpsyg.2016.00739,
PubMed: 27303320
Larsson, J., & Smith, A. T. (2012). fMRI repetition suppression:
Neuronal adaptation or stimulus expectation? Cerebral
Cortex, 22, 567–576. https://doi.org/10.1093/cercor/bhr119,
PubMed: 21690262
Lewis, A. G., Schoffelen, J.-M., Schriefers, H., & Bastiaansen, M.
(2016). A predictive coding perspective on beta oscillations
during sentence-level language comprehension. Frontiers in
Human Neuroscience, 10, 85. https://doi.org/10.3389/fnhum
.2016.00085, PubMed: 26973500
Lewis, A. G., Wang, L., & Bastiaansen, M. (2015). Fast oscillatory
dynamics during language comprehension: Unification
versus maintenance and prediction? Brain and Language,
148, 51–63. https://doi.org/10.1016/j.bandl.2015.01.003,
PubMed: 25666170
Liberman, A. M., Cooper, F. S., Shankweiler, D., & Studdert-
Kennedy, M. (1967). Perception of the speech code.
Psychological Review, 74, 431–461. https://doi.org/10.1037
/h0020279, PubMed: 4170865
Lisker, L. (1986). “Voicing” in English: A catalogue of acoustic
features signaling /b/ versus /p/ in trochees. Language and
Speech, 29, 3–11. https://doi.org/10.1177/002383098602900102,
PubMed: 3657346
Lisker, L., & Abramson, A. S. (1964). A cross-language study of
voicing in initial stops: Acoustical measurements. Word, 20,
384–422. https://doi.org/10.1080/00437956.1964.11659830,
PubMed: 3657346
Maddieson, I. (1984). Patterns of Sounds. Cambridge:
Cambridge University Press. https://doi.org/10.1017
/CBO9780511753459
Mahajan, Y., Peter, V., & Sharma, M. (2017). Effect of EEG
referencing methods on auditory mismatch negativity.
Frontiers in Neuroscience, 11, 560. https://doi.org/10.3389
/fnins.2017.00560, PubMed: 29066945
Maiste, A. C., Wiens, A. S., Hunt, M. J., Scherg, M., & Picton,
T. W. (1995). Event-related potentials and the categorical
perception of speech sounds. Ear and Hearing, 16, 68–89.
https://doi.org/10.1097/00003446-199502000-00006, PubMed:
7774771
Martynova, O., Kirjavainen, J., & Cheour, M. (2003). Mismatch
negativity and late discriminative negativity in sleeping
human newborns. Neuroscience Letters, 340, 75–78. https://
doi.org/10.1016/S0304-3940(02)01401-5, PubMed: 12668240
Masapollo, M., Polka, L., Molnar, M., & Ménard, L. (2017).
Directional asymmetries reveal a universal bias in adult vowel
perception. Journal of the Acoustical Society of America,
141, 2857–2869. https://doi.org/10.1121/1.4981006, PubMed:
28464636
McClelland, J. L., & Elman, J. L. (1986). The TRACE model of
speech perception. Cognitive Psychology, 18, 1–86. https://
doi.org/10.1016/0010-0285(86)90015-0
McGee, T., King, C., Tremblay, K., Nicol, T., Cunningham, J.,
& Kraus, N. (2001). Long-term habituation of the
speech-elicited mismatch negativity. Psychophysiology, 38,
653–658. https://doi.org/10.1111/1469-8986.3840653,
PubMed: 11446578
636
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Mesgarani, N., Cheung, C., Johnson, K., & Chang, E. F. (2014).
Phonetic feature encoding in human superior temporal
gyrus. Science, 343, 1006–1010. https://doi.org/10.1126
/science.1245994, PubMed: 24482117
Molinaro, N., Monsalve, I. F., & Lizarazu, M. (2016). Is there a
common oscillatory brain mechanism for producing and
predicting language? Language, Cognition and
Neuroscience, 31, 145–158. https://doi.org/10.1080/23273798
.2015.1077978
Monahan, P. J. (2018). Phonological knowledge and speech
comprehension. Annual Review of Linguistics, 4, 21–47.
https://doi.org/10.1146/annurev-linguistics-011817-045537
Morillon, B., & Schroeder, C. E. (2015). Neuronal oscillations as
a mechanistic substrate of auditory temporal prediction:
Neuronal oscillations and temporal predictions. Annals of the
New York Academy of Sciences, 1337, 26–31. https://doi.org
/10.1111/nyas.12629, PubMed: 25773613
Mullen, T., Kothe, C., Chi, Y. M., Ojeda, A., Kerth, T., Makeig, S.,
et al. (2013). Real-time modeling and 3D visualization of
source dynamics and connectivity using wearable EEG. 2013
35th Annual International Conference of the IEEE
Engineering in Medicine and Biology Society (EMBC),
2184–2187. https://doi.org/10.1109/EMBC.2013.6609968
Näätänen, R. (2001). The perception of speech sounds by the
human brain as reflected by the mismatch negativity (MMN)
and its magnetic equivalent (MMNm). Psychophysiology, 38,
1–21. https://doi.org/10.1111/1469-8986.3810001, PubMed:
11321610
Näätänen, R., Jacobsen, T., & Winkler, I. (2005). Memory-based
or afferent processes in mismatch negativity (MMN):
A review of the evidence. Psychophysiology, 42, 25–32.
https://doi.org/10.1111/j.1469-8986.2005.00256.x, PubMed:
15720578
Näätänen, R., & Kreegipuu, K. (2012). The mismatch negativity
(MMN). In Emily S. Kappenman & Steven J. Luck (Eds.), The
Oxford handbook of event-related potential components
(pp. 143–157). Oxford: Oxford University Press. https://doi
.org/10.1093/oxfordhb/9780195374148.013.0081
Näätänen, R., Lehtokoski, A., Lennes, M., Cheour, M.,
Huotilainen, M., Iivonen, A., et al. (1997). Language-specific
phoneme representations revealed by electric and magnetic
brain responses. Nature, 385, 432–434. https://doi.org/10
.1038/385432a0, PubMed: 9009189
Näätänen, R., Paavilainen, P., Rinne, T., & Alho, K. (2007). The
mismatch negativity (MMN) in basic research of central
auditory processing: A review. Clinical Neurophysiology,
118, 2544–2590. https://doi.org/10.1016/j.clinph.2007.04.026,
PubMed: 17931964
Nousak, J. M. K., Deacon, D., Ritter, W., & Vaughan, H. G.
(1996). Storage of information in transient auditory memory.
Cognitive Brain Research, 4, 305–317. https://doi.org/10
.1016/S0926-6410(96)00068-7, PubMed: 8957572
Obleser, J., & Weisz, N. (2012). Suppressed alpha oscillations
predict intelligibility of speech and its acoustic details.
Cerebral Cortex, 22, 2466–2477. https://doi.org/10.1093
/cercor/bhr325, PubMed: 22100354
Oldfield, R. C. (1971). The assessment and analysis of
handedness: The Edinburgh Inventory. Neuropsychologia,
9, 97–113. https://doi.org/10.1016/0028-3932, PubMed:
5146491
Oostenveld, R., & Oostendorp, T. F. (2002). Validating the
boundary element method for forward and inverse EEG
computations in the presence of a hole in the skull. Human
Brain Mapping, 17, 179–192. https://doi.org/10.1002/hbm
.10061, PubMed: 12391571
Palmer, J. A., Kreutz-Delgado, K., & Makeig, S. (2012). AMICA:
An adaptive mixture of independent component analyzers
with shared components (p. 15, Technical Report). Swartz
Center for Computational Neuroscience, University of
California San Diego.
Pérez, A., Monahan, P. J., & Lambon Ralph, M. A. (2021). Joint
recording of EEG and audio signals in hyperscanning and
pseudo-hyperscanning experiments. MethodsX, 8, 101347.
https://doi.org/10.1016/j.mex.2021.101347, PubMed:
34430250
Perrin, F., Pernier, J., Bertrand, O., & Echallier, J. F. (1989).
Spherical splines for scalp potential and current density
mapping. Electroencephalography and Clinical
Neurophysiology, 72, 184–187. https://doi.org/10.1016/0013
-4694(89)90180-6, PubMed: 2464490
Peter, V., McArthur, G., & Thompson, W. F. (2010). Effect of
deviance direction and calculation method on duration and
frequency mismatch negativity (MMN). Neuroscience Letters,
482, 71–75. https://doi.org/10.1016/j.neulet.2010.07.010,
PubMed: 20630487
Peter, V., McArthur, G., & Thompson, W. F. (2012).
Discrimination of stress in speech and music: A mismatch
negativity (MMN) study. Psychophysiology, 49, 1590–1600.
https://doi.org/10.1111/j.1469-8986.2012.01472.x, PubMed:
23066846
Phillips, C., Pellathy, T., Marantz, A., Yellin, E., Wexler, K.,
Poeppel, D., et al. (2000). Auditory cortex accesses
phonological categories: An MEG mismatch study. Journal of
Cognitive Neuroscience, 12, 1038–1055. https://doi.org/10
.1162/08989290051137567, PubMed: 11177423
Poeppel, D., & Monahan, P. J. (2011). Feedforward and
feedback in speech perception: Revisiting analysis by
synthesis. Language and Cognitive Processes, 26, 935–951.
https://doi.org/10.1080/01690965.2010.493301
Polich, J. (1993). Cognitive brain potentials. Current Directions
in Psychological Science, 2, 175–179. https://doi.org/10.1111
/1467-8721.ep10769728
Polich, J. (2007). Updating P300: An integrative theory of P3a
and P3b. Clinical Neurophysiology, 118, 2128–2148. https://
doi.org/10.1016/j.clinph.2007.04.019, PubMed: 17573239
Politzer-Ahles, S., Schluter, K. T., Wu, K., & Almeida, D. (2016).
Asymmetries in the perception of Mandarin tones: Evidence
from mismatch negativity. Journal of Experimental Psychology:
Human Perception and Performance, 42, 1547–1570.
https://doi.org/10.1037/xhp0000242, PubMed: 27195767
Polka, L., & Bohn, O.-S. (2003). Asymmetries in vowel
perception. Speech Communication, 41, 221–231. https://doi
.org/10.1016/S0167-6393(02)00105-X
Polka, L., & Bohn, O.-S. (2011). Natural referent vowel (NRV)
framework: An emerging view of early phonetic development.
Journal of Phonetics, 39, 467–478. https://doi.org/10.1016/j
.wocn.2010.08.007
Polka, L., Molnar, M., Zhao, T. C., & Masapollo, M. (2021).
Neurophysiological correlates of asymmetries in vowel
perception: An English–French cross-linguistic event-related
potential study. Frontiers in Human Neuroscience, 15, 274.
https://doi.org/10.3389/fnhum.2021.607148, PubMed:
34149375
Pulvermüller, F., & Shtyrov, Y. (2006). Language outside
the focus of attention: The mismatch negativity as a
tool for studying higher cognitive processes. Progress in
Neurobiology, 79, 49–71. https://doi.org/10.1016/j.pneurobio
.2006.04.004, PubMed: 16814448
Riddle, J., Hwang, K., Cellier, D., Dhanani, S., & D’Esposito, M.
(2019). Causal evidence for the role of neuronal oscillations
in top–down and bottom–up attention. Journal of Cognitive
Neuroscience, 31, 768–779. https://doi.org/10.1162/jocn_a
_01376, PubMed: 30726180
Riedinger, M., Nagels, A., Werth, A., & Scharinger, M. (2021).
Asymmetries in accessing vowel representations are driven
by phonological and acoustic properties: Neural and
Monahan et al.
637
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
behavioral evidence from natural German minimal pairs.
Frontiers in Human Neuroscience, 15, 612345. https://doi
.org/10.3389/fnhum.2021.612345, PubMed: 33679344
Sams, M., Kaukoranta, E., Hämäläinen, M., & Näätänen, R.
(1991). Cortical activity elicited by changes in auditory
stimuli: Different sources for the magnetic N100m and
mismatch responses. Psychophysiology, 28, 21–29. https://doi
.org/10.1111/j.1469-8986.1991.tb03382.x, PubMed: 1886961
Sams, M., Paavilainen, P., Alho, K., & Näätänen, R. (1985).
Auditory frequency discrimination and event-related
potentials. Electroencephalography and Clinical
Neurophysiology/Evoked Potentials Section, 62, 437–448.
https://doi.org/10.1016/0168-5597(85)90054-1
Samuel, A. G. (2020). Psycholinguists should resist the allure of
linguistic units as perceptual units. Journal of Memory and
Language, 111, 104070. https://doi.org/10.1016/j.jml.2019
.104070
Scharinger, M., Monahan, P. J., & Idsardi, W. J. (2011). You had
me at “Hello”: Rapid extraction of dialect information from
spoken words. Neuroimage, 56, 2329–2338. https://doi.org
/10.1016/j.neuroimage.2011.04.007, PubMed: 21511041
Scharinger, M., Monahan, P. J., & Idsardi, W. J. (2012).
Asymmetries in the processing of vowel height. Journal of
Speech, Language, and Hearing Research, 55, 903–918.
https://doi.org/10.1044/1092-4388(2011/11-0065), PubMed:
22232394
Scharinger, M., Monahan, P. J., & Idsardi, W. J. (2016). Linguistic
category structure influences early auditory processing:
Converging evidence from mismatch responses and cortical
oscillations. Neuroimage, 128, 293–301. https://doi.org/10
.1016/j.neuroimage.2016.01.003, PubMed: 26780574
Schluter, K. T., Politzer-Ahles, S., Al Kaabi, M., & Almeida, D.
(2017). Laryngeal features are phonetically abstract:
Mismatch negativity evidence from Arabic, English, and
Russian. Frontiers in Psychology, 8, 746. https://doi.org/10
.3389/fpsyg.2017.00746, PubMed: 28555118
Schluter, K. T., Politzer-Ahles, S., & Almeida, D. (2016). No
place for /h/: An ERP investigation of English fricative place
features. Language, Cognition and Neuroscience, 31,
728–740. https://doi.org/10.1080/23273798.2016.1151058,
PubMed: 27366758
Shafer, V. L., Schwartz, R. G., & Kurtzberg, D. (2004). Language-
specific memory traces of consonants in the brain. Cognitive
Brain Research, 18, 242–254. https://doi.org/10.1016/j
.cogbrainres.2003.10.007, PubMed: 14741311
Sharma, A., & Dorman, M. F. (1999). Cortical auditory evoked
potential correlates of categorical perception of voice-onset
time. Journal of the Acoustical Society of America, 106,
1078–1083. https://doi.org/10.1121/1.428048, PubMed:
10462812
Sharma, A., & Dorman, M. F. (2000). Neurophysiologic
correlates of cross-language phonetic perception. Journal of
the Acoustical Society of America, 107, 2697–2703. https://
doi.org/10.1121/1.428655, PubMed: 10830391
Smith, C. L. (1997). The devoicing of /z/ in American English:
Effects of local and prosodic context. Journal of Phonetics,
25, 471–500. https://doi.org/10.1006/jpho.1997.0053
Steriade, D. (1995). Underspecification and markedness.
In J. A. Goldsmith (Ed.), Handbook of phonological theory
(pp. 114–174). Oxford: Blackwell.
Stevens, K. N. (2002). Toward a model for lexical access based
on acoustic landmarks and distinctive features. Journal of the
Acoustical Society of America, 111, 1872–1891. https://doi
.org/10.1121/1.1458026, PubMed: 12002871
Strotseva-Feinschmidt, A., Cunitz, K., Friederici, A. D., &
Gunter, T. C. (2015). Auditory discrimination between
function words in children and adults: A mismatch negativity
study. Frontiers in Psychology, 6, 1930. https://doi.org/10
.3389/fpsyg.2015.01930, PubMed: 26733918
Tervaniemi, M., Kujala, A., Alho, K., Virtanen, J., Ilmoniemi, R. J.,
& Näätänen, R. (1999). Functional specialization of the
human auditory cortex in processing phonetic and musical
sounds: A magnetoencephalographic (MEG) study.
Neuroimage, 9, 330–336. https://doi.org/10.1006/nimg.1999
.0405, PubMed: 10075902
Wacongne, C., Labyt, E., van Wassenhove, V., Bekinschtein, T.,
Naccache, L., & Dehaene, S. (2011). Evidence for a hierarchy
of predictions and prediction errors in human cortex.
Proceedings of the National Academy of Sciences, U.S.A.,
108, 20754–20759. https://doi.org/10.1073/pnas.1117807108,
PubMed: 22147913
Wang, X.-J. (2010). Neurophysiological and computational
principles of cortical rhythms in cognition. Physiological
Reviews, 90, 1195–1268. https://doi.org/10.1152/physrev
.00035.2008, PubMed: 20664082
Wang, W. S.-Y., & Crawford, J. (1960). Frequency studies of
English consonants. Language and Speech, 3, 131–139.
https://doi.org/10.1177/002383096000300302
Werker, J. F., & Logan, J. S. (1985). Cross-language evidence
for three factors in speech perception. Perception
& Psychophysics, 37, 35–44. https://doi.org/10.3758
/BF03207136, PubMed: 3991316
Winkler, I. (2007). Interpreting the mismatch negativity.
Journal of Psychophysiology, 21, 147–163. https://doi.org/10
.1027/0269-8803.21.34.147
Winkler, I., Kujala, T., Alku, P., & Näätänen, R. (2003). Language
context and phonetic change detection. Cognitive Brain
Research, 17, 833–844. https://doi.org/10.1016/S0926-6410
(03)00205-2, PubMed: 14561466
Winkler, I., Lehtokoski, A., Alku, P., Vainio, M., Czigler, I.,
Csépe, V., et al. (1999). Pre-attentive detection of vowel
contrasts utilizes both phonetic and auditory memory
representations. Cognitive Brain Research, 7, 357–369. https://
doi.org/10.1016/S0926-6410(98)00039-1, PubMed: 9838192
Winkler, I., Paavilainen, P., Alho, K., Reinikainen, K., Sams, M., &
Näätanen, R. (1990). The effect of small variation of the
frequent auditory stimulus on the event-related brain
potential to the infrequent stimulus. Psychophysiology, 27,
228–235. https://doi.org/10.1111/j.1469-8986.1990.tb00374.x,
PubMed: 2247552
Yi, H. G., Leonard, M. K., & Chang, E. F. (2019). The encoding
of speech sounds in the superior temporal gyrus. Neuron,
102, 1096–1110. https://doi.org/10.1016/j.neuron.2019.04
.023, PubMed: 31220442
Ylinen, S., Shestakova, A., Huotilainen, M., Alku, P., & Näätänen,
R. (2006). Mismatch negativity (MMN) elicited by changes in
phoneme length: A cross-linguistic study. Brain Research,
1072, 175–185. https://doi.org/10.1016/j.brainres.2005.12.004,
PubMed: 16426584
Yu, Y. H., & Shafer, V. L. (2021). Neural representation of the
English vowel feature [high]: Evidence from /ε/ vs. /ɪ/.
Frontiers in Human Neuroscience, 15, 629517. https://doi
.org/10.3389/fnhum.2021.629517, PubMed: 33897394
Zachau, S., Rinker, T., Körner, B., Kohls, G., Maas, V.,
Hennighausen, K., et al. (2005). Extracting rules: Early and
late mismatch negativity to tone patterns. NeuroReport, 16,
2015–2019. https://doi.org/10.1097/00001756-200512190
-00009, PubMed: 16317345
638
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
1
8
1
9
9
6
9
6
7
/
j
o
c
n
_
a
_
0
1
8
1
7
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3