Restoration of fMRI Decodability Does Not Imply
Latent Working Memory States
Sebastian Schneegans and Paul M. Bays
Abstrakt
■ Recent imaging studies have challenged the prevailing
view that working memory is mediated by sustained neural
Aktivität. Using machine learning methods to reconstruct mem-
ory content, these studies found that previously diminished
representations can be restored by retrospective cueing or
other forms of stimulation. These findings have been inter-
preted as evidence for an activity-silent working memory state
that can be reactivated dependent on task demands. Hier, Wir
test the validity of this conclusion by formulating a neural pro-
cess model of working memory based on sustained activity
and using this model to emulate a spatial recall task with retro-
cueing. The simulation reproduces both behavioral and fMRI
results previously taken as evidence for latent states, in par-
ticular the restoration of spatial reconstruction quality follow-
ing an informative cue. Our results demonstrate that recovery
of the decodability of an imaging signal does not provide
compelling evidence for an activity-silent working memory
state. ■
EINFÜHRUNG
The dominant view of the neural mechanism underlying
working memory is that memory representations are
maintained in the sustained spiking activity of neurons
(Chaudhuri & Fiete, 2016; Bays, 2015; Eriksson, Vogel,
Lansner, Bergström, & Nyberg, 2015; Funahashi, Bruce,
& Goldman-Rakic, 1989; Fuster & Alexander, 1971). Das
sustained activity may arise from local self-excitation in
populations of neurons or reverberatory loops between
different cortical areas ( Wang, 2001). Working memory
representations can be modeled as attractor states in
the neural activation dynamics. For continuous features,
such as location, color, or orientation of visual stimuli,
population codes with homogeneous recurrent connec-
tivity form continuous attractors such that each possible
feature value can be maintained in working memory
(Johnson, Simmering, & Buss, 2014; Wei, Wang, & Wang,
2012; Compte, Brunel, Goldman-Rakic, & Wang, 2000).
Errors arise at encoding and when the attractors drift to
neighboring feature values or decay under the influence
of random noise ( Wimmer, Nykamp, Constantinidis, &
Compte, 2014; Burak & Fiete, 2012; Camperi & Wang,
1998).
Models of this type have successfully accounted for
memory-related neural activity at a high level of physio-
logical detail (Wimmer et al., 2014; Compte et al., 2000).
Zusätzlich, they have been used to explain a wide range
of behavioral findings, such as performance and capacity
limits in change detection tasks (Engel & Wang, 2011;
University of Cambridge
Johnson, Spencer, & Schöner, 2009), biases in visual
Arbeitsgedächtnis (Schneegans, Spencer, Schöner, Hwang,
& Hollingworth, 2014; Simmering, Spencer, & Schöner,
2006; Camperi & Wang, 1998), and developmental changes
in working memory performance (Schutte & Spencer,
2009; Edin, Macoveanu, Olesen, Tegnér, & Klingberg,
2007).
A strikingly different account of working memory pro-
poses that memory representations are maintained in
an activity-silent state (d.h., without sustained neural
firing) through biochemical or morphological changes
in neurons (Stokes, 2015; Barak & Tsodyks, 2014). A
prominent candidate mechanism is rapid synaptic plas-
ticity (Erickson, Maramara, & Lisman, 2010), which may
allow sensory stimulation to produce changes in neural
connection patterns over short timescales and create
assemblies of neurons that may later be reactivated by
nonspecific input (Mi, Katkov, & Tsodyks, 2017; Mongillo,
Barak, & Tsodyks, 2008). Proponents of such working
memory mechanisms suggest that persistent neural activ-
ity only reflects sustained attention to the currently task-
relevant item, whereas other items can be held in an
activity-silent state (Rose et al., 2016; LaRocque, Lewis-
Peacock, Drysdale, Oberauer, & Postle, 2013; Lewis-
Peacock, Drysdale, Oberauer, & Postle, 2012).
An important source of information about the neural
basis of working memory is provided by fMRI studies
that measure neural activity during the delay periods of
working memory tasks. Results of early studies were con-
sistent with the sustained firing account, finding elevated
BOLD signals in regions of prefrontal and parietal cortex
(Xu & Chun, 2006; Todd & Marois, 2004; Linden et al.,
© 2017 Massachusetts Institute of Technology. Published under a
Creative Commons Atrribution 3.0 Unportiert (CC BY 3.0) Lizenz.
Zeitschrift für kognitive Neurowissenschaften 29:12, S. 1977–1994
doi:10.1162/jocn_a_01180
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
Ö
6
C
8
N
9
_
6
A
/
_
J
0
Ö
1
C
1
N
8
0
_
A
P
_
D
0
1
B
1
j
8
G
0
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
F
.
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
2003; Courtney, Ungerleider, Keil, & Haxby, 1997). Wie-
immer, the interpretation of these univariate fMRI results is
not necessarily straightforward. More recent studies have
applied multivariate pattern analysis and reconstruction
methods to BOLD signal data to characterize working
memory representation (Sprague, Ester, & Serences,
2014; Riggall & Postle, 2012; Harrison & Tong, 2009;
Serences, Ester, Vogel, & Awh, 2009). Such studies have
found a dissociation between cortical regions with ele-
vated BOLD signal and regions from which the features
of remembered items could be successfully decoded
(Emrich, Riggall, LaRocque, & Postle, 2013; Riggall &
Postle, 2012).
Kürzlich, such reconstruction methods have also been
used to investigate how working memory representa-
tions change when informative cues are provided during
the delay period. In the task used by Sprague, Ester, Und
Serences (2016), participants had to memorize the loca-
tions of two colored disks. In some trials, a color cue
given midway through the memory period indicated
which location would be tested at the end. The recon-
struction quality for the cued item’s location decreased
over the course of the delay period but showed a sig-
nificant restoration following this spatially nonspecific
retro-cue. This was interpreted as evidence for an activity-
silent component in location working memory, welche
cannot be decoded from the BOLD signal data but can
be reactivated (and thereby contribute to reconstruction
quality again) by the retro-cue.
Similar conclusions have been drawn from several
prominent studies applying reconstruction methods to
fMRI and EEG data. Rose et al. (2016) found that the cat-
egory of a currently task-relevant item could be success-
fully decoded from fMRI recordings, but not the category
of a second item concurrently held in working memory
(that became task relevant at a later stage). The authors
concluded that this second item is held in an activity-
silent state and is only restored into a sustained activity
representation when cued as immediately task relevant.
A restoration of decoding quality from EEG data has been
observed following TMS pulses (Rose et al., 2016) oder
presentation of salient, but uninformative, visuelle Reize
( Wolff, Jochim, Akyürek, & Stokes, 2017; Wolff, Ding,
Myers, & Stokes, 2015). This was interpreted as effect
of interactions between the unspecific stimulation and a
silent working memory representation, realized in a
changed neural connectivity pattern.
In this study, we critically examine the interpretation
that the restoration of reconstruction quality following
a cue is evidence for activity-silent working memory
Staaten. We implement a neural model of working memory
based on the principle of sustained activity through self-
excitation in neural populations and derive simulated
BOLD signal data from this model. We then apply the
same kind of reconstruction method as was previously
used for the analysis of experimental data. Focusing on
the study by Sprague et al. (2016), we demonstrate that
the neural model can reproduce both the behavioral and
fMRI reconstruction results, despite relying only on active
memory representations. Insbesondere, in a minimal neu-
ral model of the retro-cue task, a spatially nonspecific
color cue can selectively strengthen the reconstruction
of the cued item’s location. The neural model demon-
strates that the assumption of activity-silent WM states
is not necessary to explain these experimental results
and highlights the importance of considering the func-
tional neural architectures involved in the task when
interpreting imaging data.
METHODEN
Neural Model
We aim to provide a minimal neural model that can
perform the delayed estimation task with retro-cueing
(Figure 1A) and account for the behavioral and fMRI
results from the experimental study of Sprague et al.
(2016). We formulate this model within the framework
of dynamic neural fields, a form of recurrent neural net-
work that describes the continuous evolution of activity
distributions in neural populations through differential
Gleichungen (Schneegans & Schöner, 2008). Neural fields
are continuous in space: Rather than simulating the ac-
tivity of individual neurons, each point in a neural field
reflects the mean activity of neurons with the corre-
sponding spatial selectivity. Working memory is realized
in this framework through localized peaks of activation
that are sustained through self-excitation (balanced by
surround inhibition), forming attractor states in the neu-
ral dynamics. Neural field models generate continuous
time courses of neural activity patterns, making them well
suited for modeling fMRI data ( Wijeakumar, Ambrose,
Spencer, & Curtu, 2016).
We employ a single combined perceptual and working
memory representation to emulate neural activation in
the task (Figure 1B). Stimulus locations are represented
in a population code over 2-D visual space. Wichtig,
the model also needs to represent the item color asso-
ciated with each location to select the correct memo-
rized item for response following a color cue. Following
a recent model of feature binding in visual working
Erinnerung, we employ a conjunctive code for color–location
bindings (Schneegans & Bays, 2017; Schneegans,
Spencer, & Schöner, 2016). To keep the model simple,
we represent only the two stimulus colors used in the
original experiment (red and blue), yielding two neural
fields for these discrete colors over the same 2-D visual
Raum. These can be interpreted as slices from a full
population code for possible color–location conjunc-
tionen (compare Richter, Lins, Schneegans, & Schöner,
2014; Lipinski, Schneegans, Sandamirskaya, Spencer, &
Schöner, 2012).
The activation in these fields, which can be equated
with the mean membrane potential of neurons with the
1978
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 12
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
F
/
T
T
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
Ö
6
C
8
N
9
_
6
A
/
_
J
0
Ö
1
C
1
N
8
0
_
A
P
_
D
0
1
B
1
j
8
G
0
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
F
/
.
T
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
interactions within the model, and ξ is a random noise
Begriff.
The lateral interactions are based on the current field
output, a continuous homologue of the neural firing rate,
which is computed from the activation a via a sigmoid
(logistic) function f:
f að Þ ¼
1
1 þ e−a
(2)
The output is close to zero for low (negative) Aktivierung
Werte, rises for activation values around zero, and satu-
rates at a value of one for higher activations. This pro-
duces the effect that only regions with high activation
contribute to interactions within the field.
Lateral interactions consist of three components: A
local excitatory component, described by a normalized
2-D Gaussian function ϕ(X; σexc) with mean zero and
width parameter σexc that is scaled with an excitatory
weight cexc; a uniform inhibitory component within each
color field with weight cinhc; and a global inhibitory com-
ponent over both fields with weight cinhg. The input from
lateral interactions for each field position is computed as
Z
D
g l; X; T
Þ ¼
f a l; x0; T
D
D
Þ
dx0 − cinhg
D
Z
Þ cexcϕ x − x0; σexc
D
X
f a l0; x0; T
D
Þ
D
Þ − cinhc
Þ dx0
Þ
(3)
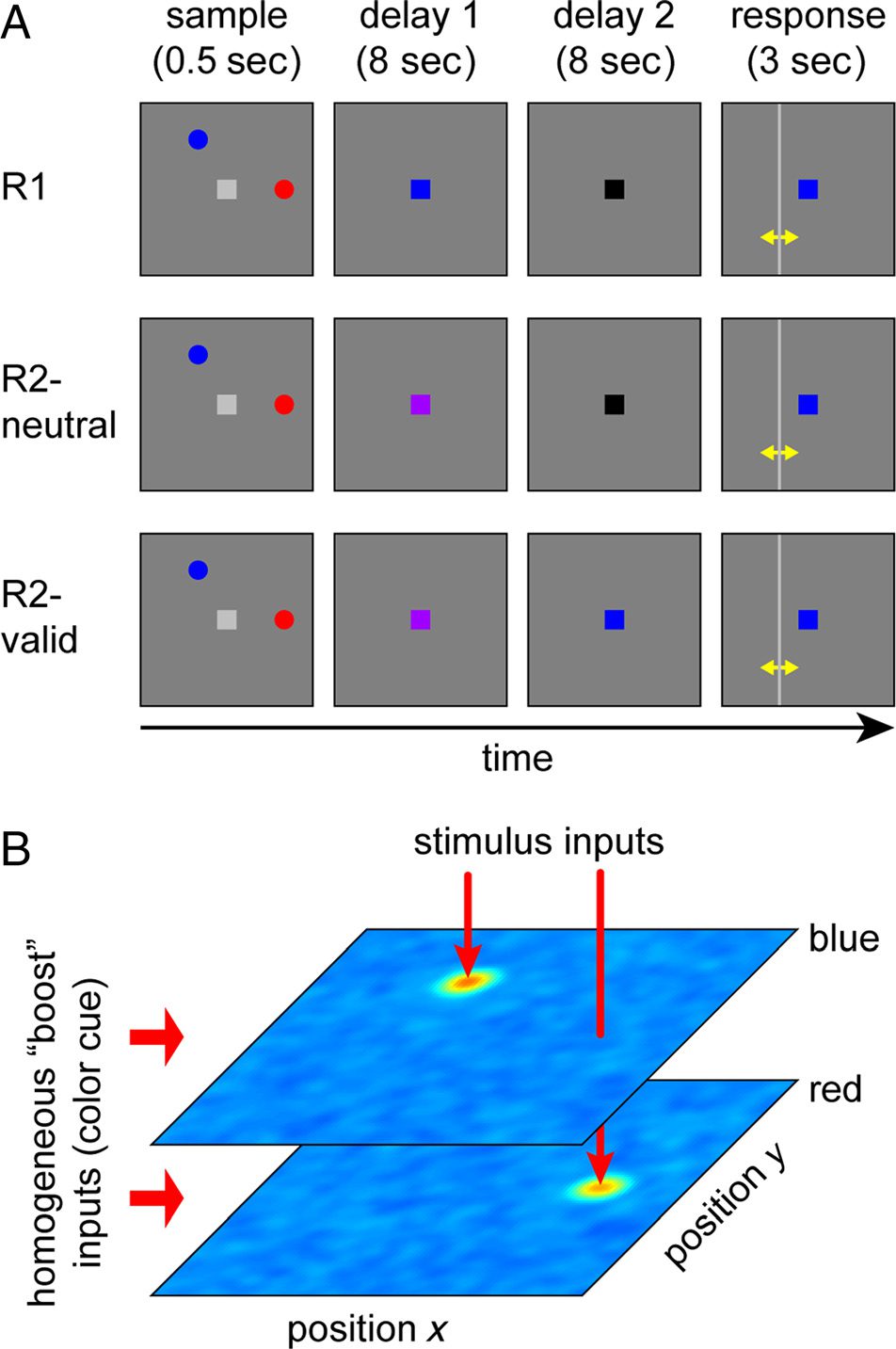
Figur 1. (A) Experimental task of Sprague et al. (2016). Teilnehmer
view a sample array of two colored discs and have to report the location
of one item (the target) by adjusting a vertical or horizontal bar (Gelb
arrows are not part of the experimental display). The color of the
fixation point can change at different times during the delay period
to indicate the target item. (B) Architecture of the neural model.
Two neural fields over retinal space, with selectivity matching the
two colors (blue and red) used in the task, are shown as color-coded
activation distributions. Each colored sample stimulus provides localized
input to the corresponding field, generating a peak of activity that
encodes the stimulus location. The informative color cue generates a
homogeneous input to the field of matching color, boosting activity
throughout the population.
corresponding selectivity (Amari, 1977), is governed by
the differential equation
τ _a l; X; T
D
D
Þ ¼ −a l; X; T
D
þ ξ l; X; T
Þ þ h þ s l; X; T
Þ
D
D
Þ þ g l; X; T
Þ
(1)
where a(l, X, T) is the activation for color l 2 {Rot, Blau},
spatial position x = (X, j), and time t. The notation _a
indicates the rate of change of activation over time, mit
a time constant τ. The scalar variable h is the global resting
level of activation that the field will relax to in the absence
of input and lateral interactions, s is the local external
input to each field location, g is the input from lateral
l0
The noise term ξ describes spatially correlated random
noise with amplitude cnoise, generated from white noise
by convolution with a Gaussian kernel (using the same
width parameter σexc as in the lateral interactions):
Z
D
ξ l; X; T
Þ ¼ cnoise
v l; x0; T
D
D
v l; X; T
Þϕ x − x0; σexc
D
Þ
Þ ∼ N 0; 1D
Þdx0;
(4)
Model parameters were adjusted manually to produce a
close fit to the behavioral and fMRI reconstruction data
and are fixed across task conditions. All parameter values
are listed in Table 1. The model was simulated numerically
by sampling the visual space in the range [−6°, 6°] ×
[−6°, 6°] mit 100 × 100 sampling points and approxi-
mating the temporal evolution of activation using the
Euler method with a fixed time step of 10 ms.
Behavioral Task and Emulation in the Model
We use the neural model to emulate the delayed estima-
tion task used by Sprague et al. (2016), shown in
Figure 1A. Participants had to memorize the locations
of two colored discs (one red and one blue, diameter
0.15°) presented for 500 ms. The discs were located
on an invisible circle (radius 3.5°) around a central fixa-
tion point, offset from each other by ±60°, ±120°, oder
180°. An additional small trial-to-trial variation of relative
positions was introduced by adding a random value uni-
formly distributed in [−0.3°, 0.3°] independently to each
horizontal and vertical stimulus position. Participants had
Schneegans and Bays
1979
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
Ö
6
C
8
N
9
_
6
A
/
_
J
0
Ö
1
C
1
N
8
0
_
A
P
_
D
0
1
B
1
j
8
G
0
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
T
F
.
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
Tisch 1. Parameter Values for Neural Model and Generation of
Simulated BOLD Signals
Parameter
Description
Wert
τ
H
cexc
σexc
cinhc
cinhg
cnoise
cstim
σstim
ccue
cforget
σvox
cnvox
Time constant of neural dynamics
100 ms
Field resting level
Lateral excitatory weight
Width of lateral excitation
Inhibitory weight (within color field)
Inhibitory weight (global)
Noise amplitude
Stimulus amplitude
Stimulus width
Color cue amplitude
Deactivation amplitude after trial
Voxel range parameter
Voxel noise amplitude
−5
20
0.25°
2.6
0.52
55
50
2°
17.5
5
1.5
2.5
to remember stimulus locations over a delay interval of
16 sec in total, then report the location of one stimulus
cued by its color.
Memory load in this task was manipulated across three
Bedingungen: In the “remember one” condition (R1), Die
fixation point changed color to either red or blue imme-
diately after offset of the sample stimuli, indicating with
100% reliability which item was to be tested. The fixation
point then changed to a neutral color (black) nach 8 Sek.
In the “remember two” condition, the color of the fixa-
tion point changed to purple at the beginning of the
delay period, indicating that either item could be tested.
Nach 8 Sek, the fixation point’s color then either changed
to black (R2-neutral), or it changed to one of the item
colors, acting as a retro-cue that indicated the item to
be tested with 100% reliability (R2-valid). In all condi-
tionen, the fixation point took the color of the tested item
at the beginning of the response period (after the 16-sec
delay). The participant then had to adjust either a hori-
zontal or vertical line displayed on the screen to report
the horizontal or vertical location of the cued item.
In the neural model, memory sample stimuli are emu-
lated as (nonnormalized) Gaussian inputs (with ampli-
tude cstim and width parameter σstim), centered at the
stimulus location ml for a stimulus with color l. Der
input for each colored stimulus only drives activation
in the field for the matching color and remains active
for a 500-msec interval [Tonne, toff]:
D
s l; X; T
Þ ¼ cstim exp −
!
jjx − mljj2
2σ2
stim
for ton ≤ t ≤ toff
(5)
The informative color cue is modeled as a homogeneous
“boost” input with amplitude ccue to the field with match-
ing color lcue that is active for 500 msec after onset of the
cue (see Lipinski et al., 2012, for an analogous mecha-
nism at the perceptual level):
D
s lcue; X; T
Þ ¼ ccue for tcue ≤ t ≤ tcue þ 500 ms
(6)
Behavioral results in retro-cue experiments indicate that
processing of a retro-cue takes about 300–500 msec
(Souza & Oberauer, 2016; Souza, Rerko, & Oberauer,
2014; Tanoue & Berryhill, 2012), and we assume that
the cue has no further effect after this, even though it
remains visible. The uninformative cues (black or purple)
are not modeled explicitly.
For the generation of the spatial response in the model,
the response color cue input is presented for 500 ms
in the same fashion as described above and 500 msec after
the offset of the cue; the spatial center of mass (xresp,
yresp) of the field outputs (summed over colors) is deter-
mined as
Z
X
xresp ¼
l
X
Z
l
Z
X
yresp ¼
l
X
Z
l
D
x f a l; X; T
D
Þ
Þdx
D
f a l; X; T
D
Þ
Þdx
D
y f a l; X; T
D
Þ
Þdx
D
f a l; X; T
D
Þ
Þdx
(7)
Note that under normal conditions, there will be a peak
of activation in only one of the two fields after the pre-
sentation of the response cue (with low homogeneous
activity in the other field), so this method yields the
estimated location of the cued item. Response error is
determined as absolute deviation of the report location
from the location of the cued stimulus, either in the
horizontal or vertical direction (randomly selected) Zu
emulate the experimental response procedure. Nach
response generation, the activation levels of both fields
are transiently decreased by a value cforget for 500 ms
(beginnend 500 msec after response time) to extinguish any
existing activation peaks.
We numerically simulated 10 blocks of 216 Versuche (72 Versuche
per condition) of this task, with trials in each block fully
counterbalanced for possible stimulus positions, angular
distance between stimuli, and cued item color. Stimulus
parameters directly emulate the original study, und das
total number of trials approximates the number of trials
across participants in that study.
In the original fMRI study, an additional mapping task
was performed as basis for the reconstruction of spatial
representations from voxel BOLD signals. In this task,
participants memorized the location of a single stimulus,
then a flickering checkerboard disc (radius 1.083°) War
presented for 3000 ms, covering the location of the
1980
Zeitschrift für kognitive Neurowissenschaften
Volumen 29, Nummer 12
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
Ö
6
C
8
N
9
_
6
A
/
_
J
0
Ö
1
C
1
N
8
0
_
A
P
_
D
0
1
B
1
j
8
G
0
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
T
F
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
memory stimulus. After this presentation, a probe stimulus
wurde gezeigt, and participants had to report whether the
memorized stimulus location was to the left or right (oder
above or below) the probe. The checkerboard stimulus
was shown in 36 locations on a hexagonal grid, welche
was again rotated in 12 different ways across sessions.
We emulate the checkerboard stimulus as a Gaussian
input with the same amplitude cstim and width σstim as
the stimuli in the main task that is applied to the same
location in the fields for both colors. We do not emulate
the working memory component of the mapping task,
because the simple model architecture does not allow
differentiation between perceptual and working memory
Artikel. This does not affect the reconstruction procedure,
because only the checkerboard stimulus locations were
taken into account for computing the reconstructions
in the original study. We performed four sets of this map-
ping task for each block of working memory trials, rotat-
ing the hexagonal stimulus grid by increments of 15°
across sets and by increments of 5° across blocks.
Generating Simulated BOLD Signals
The original study measured BOLD signals for individual
voxels and used an additional localizer task to assign
these voxels to different cortical areas of interest (inkl-
ing V1, V3A, IPS0, IPS2, and sPCS). We do not aim to cap-
ture this division into different cortical regions in the
model and only generate a single set of BOLD signals
to compare it to the combined results from the fMRI
Studie (which contains all of the key effects reported in
that study).
The interpretation of the original study’s fMRI results
as evidence for activity-silent working memory is based
on the assumption that the BOLD signal reflects neural
spiking activity. We follow this assumption here and de-
rive the simulated BOLD signal directly from the field
output in the neural model (the continuous homologue
of the firing rate). We note that this is most likely an
oversimplification, but we believe that it allows the most
direct test of the conclusions drawn from the original
Studie. We define a simulated voxel by selecting a ran-
dom sample of points from the two fields, {(l1, x1),…,
(ln, xn)}. Building on the assumption that the visual
areas under investigation are at least partly organized
retinotopically, spatial locations of sampling points are
drawn from a mixture of a Gaussian and a uniform
distribution,
D
p li; xi
D
Þ ∝ 0:5 þ 0:5ϕ x − xvox; σvox
Þ
(8)
The center xvox = (xvox, yvox) of the Gaussian distribu-
tion for each voxel is chosen uniformly from the space
[−6°, 6°] × [−6°, 6°] and applies to both color fields.
The color li for each sampling point is chosen with
equal probabilities from {Rot, Blau}.
from the neural model at these sampling points and
convolving this signal in time with the canonical hemo-
dynamic response function h:
Z ∞
D
b tscan
Þ ¼
D
f a li; xi; tscan − t
D
Þ
Þdt
X
h tð Þ 1
N
D
0
ich
þ cnvoxχ tscan
Þ
(9)
Hier, χ ∼ N (0,1) is random noise added to each voxel
to reflect neural activity not directly related to the task.
The hemodynamic response function, as defined by
Lindquist, Loh, Atlas, and Wager (2009), Ist
h tð Þ ¼ t α1−1
D
Þβα1 e −βt
D
Þ
Γ α1ð
Þ
− c
Þ
D
t α2−1
Þβα2 e −βt
D
Þ
Γ α2ð
(10)
with standard parameters α1 = 6, α2 = 16, β = 1, Und
c = 1/6.
Emulating the procedure in the original study, Wir
generated BOLD signals for 10 time points in each trial
of the working memory task, spaced in intervals of
2250 msec beginning at memory stimulus onset. Im
mapping task, a single set of BOLD signals was generated
for each trial. For each block of trials in the working
memory task (and the associated trials of the mapping
Aufgabe), a new mapping of voxels to points in the neural
fields was randomly generated. Wir verwendeten 1000 voxels with
1000 points per voxel. All simulated BOLD signals were
transformed into z scores before further analysis, inde-
pendently between blocks of trials and between mapping
task and working memory task.
Reconstruction and Analysis of
Spatial Representations
The reconstruction of neural representations from simu-
lated BOLD signals is performed in the same way as the
reconstruction from actual fMRI data in the original
Studie (see Sprague et al., 2016, for details). In brief, Die
z-transformed BOLD signal for each voxel is modeled as
a linear combination of 37 information channels (spatial
filters over 2-D visual space). The data from the spatial
mapping task is used to estimate a weight matrix that
maps expected filter responses (based on the known
stimulus locations in this task) onto the observed BOLD
Signale. This matrix is then inverted to compute channel
activations for each trial and time point in the working
memory task from measured BOLD signals.
For further analysis and visualization, reconstructed
activation distributions over visual space are generated
as superpositions of the spatial filters of each channel
weighted with the estimated channel activation. Target
positions in different trials can be aligned with each
other in the reconstructions by rotating, flipping, Und
shifting the pattern of spatial filters associated with each
channel.
D
Ö
w
N
l
Ö
A
D
e
D
F
R
Ö
M
l
l
/
/
/
/
J
T
T
F
/
ich
T
.
:
/
/
H
T
T
P
:
/
D
/
Ö
M
w
ich
N
T
Ö
P
A
R
D
C
e
.
D
S
F
ich
R
Ö
l
M
v
e
H
R
C
P
H
A
D
ich
ich
R
R
e
.
C
C
T
.
Ö
M
M
/
J
e
Ö
D
u
C
N
Ö
/
C
A
N
R
A
T
R
ich
T
ich
C
C
l
e
e
–
P
–
D
P
D
2
F
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
Ö
6
C
8
N
9
_
6
A
/
_
J
0
Ö
1
C
1
N
8
0
_
A
P
_
D
0
1
B
1
j
8
G
0
u
.
e
P
S
T
D
Ö
F
N
B
0
j
8
S
M
e
ICH
P
T
e
M
L
ich
B
B
e
R
R
A
2
R
0
2
ich
3
e
S
/
J
.
T
F
/
u
S
e
R
Ö
N
1
7
M
A
j
2
0
2
1
The simulated BOLD signal b for one voxel at time tscan
is then computed by averaging over the output signal
To visualize and compare spatial representations over
time in the different task conditions, trials are grouped by
Schneegans and Bays
1981
condition and absolute angular distance between memory
Reize (60°, 120°, and 180° ), and reconstructions are
generated, aligned with respect to the positions of the
two stimuli (ignoring the small random offset in stimulus
locations from trial to trial). For further analysis, Versuche
are grouped by condition only, and reconstructions are
generated, aligned by the exact location of the cued stim-
ulus (half of the trials with a stimulus separation of 180°
are omitted here to obtain equal numbers of stimulus
separations −120°, −60°, 60°, 120°, and 180°). Eins-
dimensional reconstructions over an angular space are
extracted from the obtained activation distributions
over visual space by interpolating and averaging acti-
vations within a ring of 2.9°–4.1° eccentricity around the
center.
We apply the same analyses on these reconstructions
as employed in the experimental study. A fidelity mea-
sure F of target representation in the 1-D reconstructions
R(θ) (aligned with the cued stimulus position always at
null) is defined as the mean product of the reconstruc-
tion with a cosine function,
Z
F ¼ 1
2π
r θð Þ cos θð Þdθ
(11)
Activation profiles in the 1-D reconstructions are quan-
tified by fitting with a function
8
<
(cid:2)
f θð Þ ¼ b þ a 0:5 þ 0:5 cos
:
b;
(cid:3)
j
7
2π θ − μ
j
σfit
;
if θ − μ
j
j < σfit
otherwise
(12)
where b is baseline activation, a is amplitude of a Gaussian-
like component, σfit is the width parameter for this com-
ponent, and μ is an angular bias of the peak limited to a
small range around zero. For analysis of the model be-
havior, we also apply this function fitting directly to the
field output in the neural model (averaged across trials,
without generation of simulated BOLD signals and re-
construction from them).
To determine significance of fidelity and differences in
fitting parameters, we used a bootstrapped sign test. Over
1000 iterations, we resampled with replacement from the
1-D single-trial reconstructions (with 720 trials per condi-
tion) and computed fidelity and function fit for the aver-
age over resampled reconstructions. For comparing fitting
parameters between two conditions or time periods, we
computed the proportion in which the difference between
these variables was greater than zero and the proportion
in which it was less than zero and doubled the smaller of
these proportions to obtain the p value. For testing whether
the fidelity measure was greater than zero, we performed
a one-tailed sign test and report the proportion of re-
sampling iterations with F < 0.
RESULTS
Model Mechanism and Behavioral Results
We applied a neural model of working memory based on
sustained activity in neural populations to emulate the
retro-cue task used by Sprague et al. (2016). In this task
(Figure 1A), participants were presented with two col-
ored disks, which they memorized in three different
conditions: On R1 trials, a colored cue at the start of
the trial indicated one of the items whose location was
to be remembered; on R2-neutral trials, both items had
to be remembered to the end of the trial; and on the
critical R2-valid trials, a retro-cue delivered midway
through the trial indicated which item would be tested,
allowing participants to forget the other item.
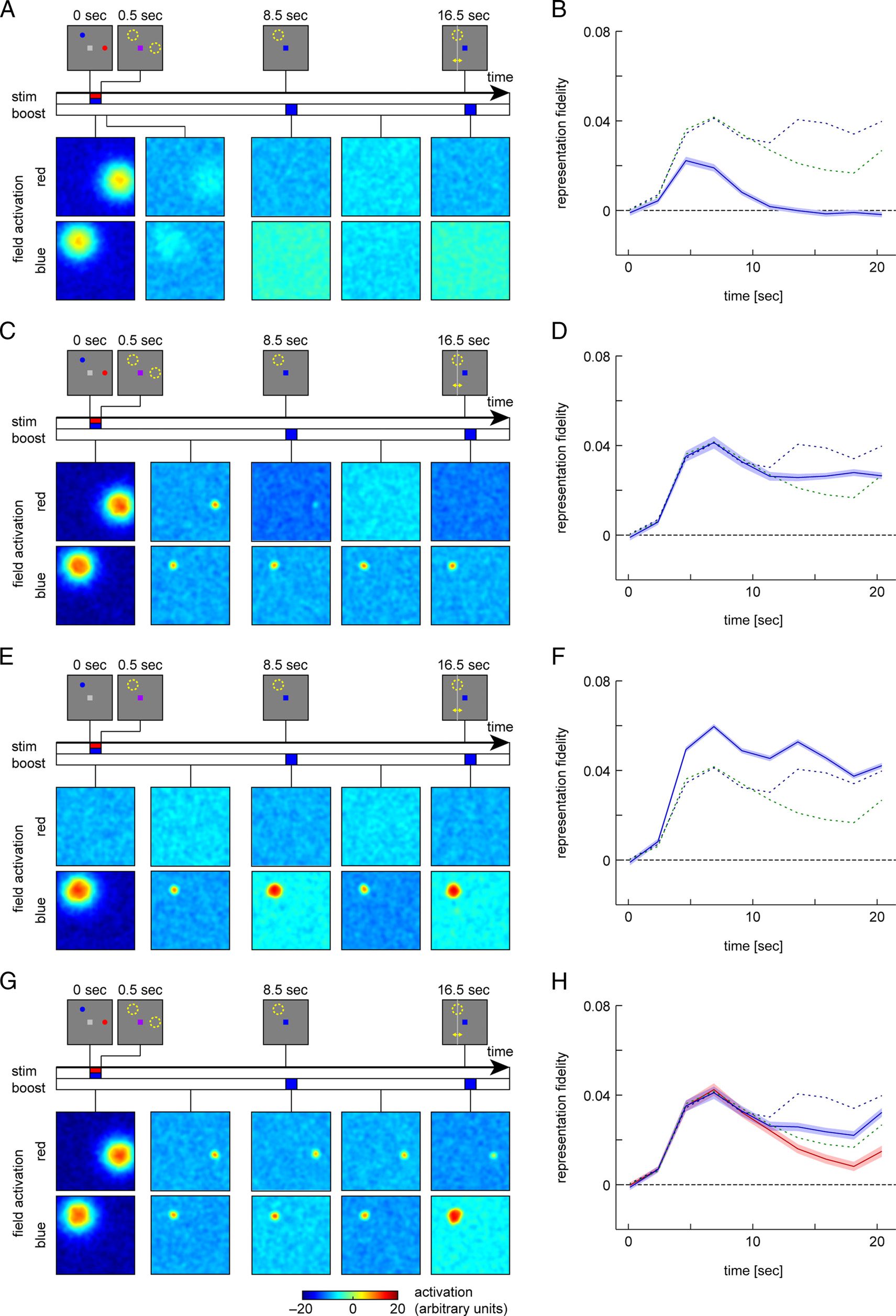
The model (Figure 1B) describes activity of two spatial-
and color-selective populations (neural “fields”) with
color selectivity matching the two colors used in the
task. There is localized lateral excitation within each field
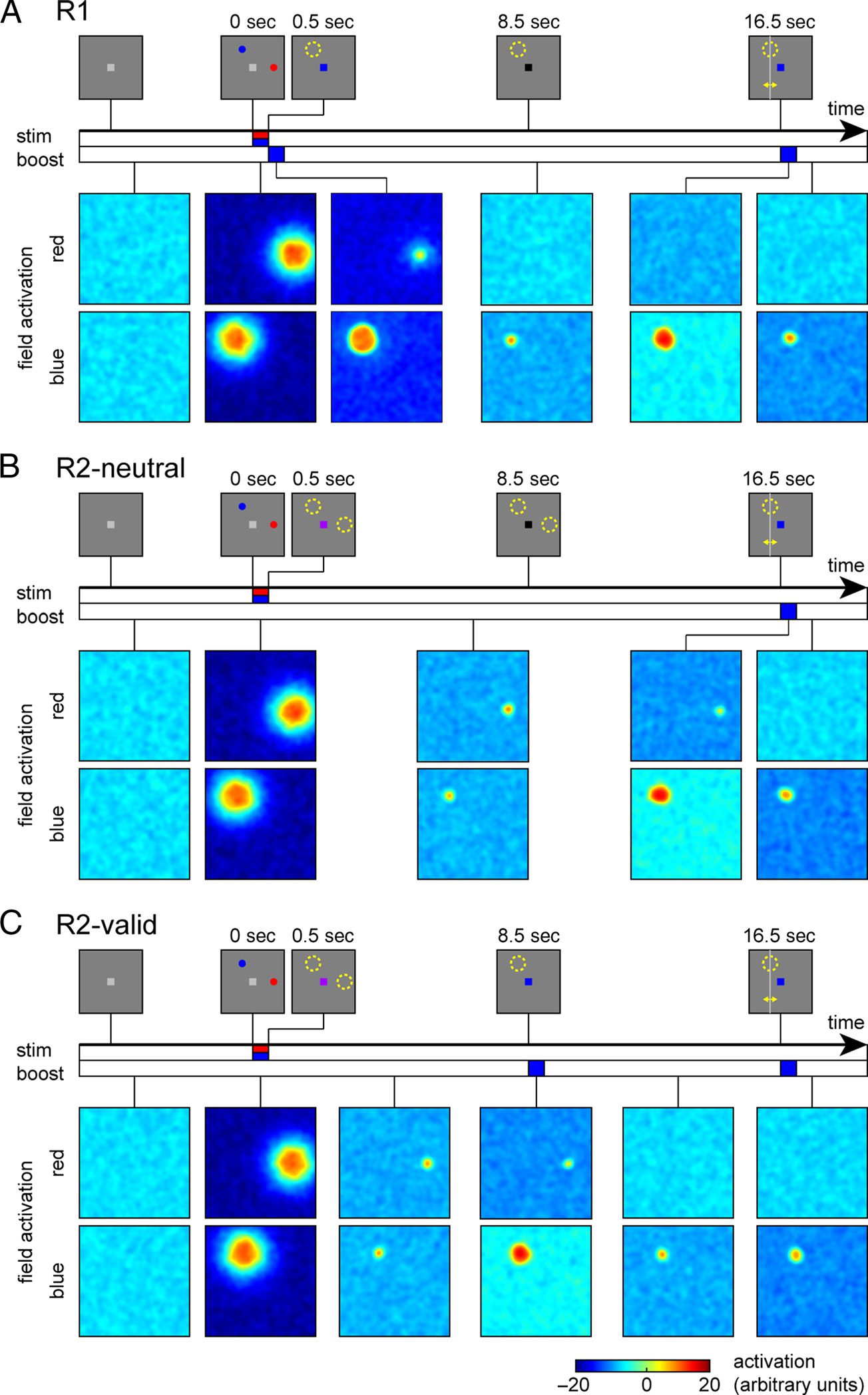
and global inhibition within and between fields. Figure 2
illustrates the activation time course in the model for a
single trial in each task condition to demonstrate the basic
mechanisms of the model. The figure shows the activa-
tion distributions in both fields at different points in time
(at this stage, without generation of simulated BOLD sig-
nals or reconstructions from them).
At the beginning of the R1 trial (Figure 2A), both fields
show uniformly low activation, with small fluctuations
due to random noise. Presentation of the memory sam-
ple stimuli induces strong activation in both fields that is
localized at the stimulus positions and specific for the
color of each stimulus. Immediately following the sample
stimuli, a blue color cue is presented in this trial, which
homogeneously raises activation levels in the blue color
field. Because of the effects of lateral interactions, this
strengthens the existing peak of activation in this field,
whereas activation in other parts of the field is held rela-
tively low by the increased global inhibition arising from
this peak. In addition, the global inhibitory effect acts on
the activation levels in the red color field. It reduces the
strength of the existing activation peak in that field such
that it ceases to generate sufficient self-excitation and
ultimately collapses.
After the color cue input is turned off, the activation
level in the blue field decreases, but a smaller localized
peak of activation remains stable throughout the delay
period, sustained by local self-excitation and surround
inhibition within the field. During the response period,
the color cue is activated again and strengthens the re-
maining peak but does not qualitatively alter the activa-
tion patterns in the model. After this second cue input
has ceased, the response location is determined as the
center of mass of the field output over visual space
(summed over both fields).
The R2-neutral trial begins in the same way as the R1
trial (Figure 2B), but here no informative color cue is given
until the response period. Following the presentation of
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
o
6
c
8
n
9
_
6
a
/
_
j
0
o
1
c
1
n
8
0
_
a
p
_
d
0
1
b
1
y
8
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
1982
Journal of Cognitive Neuroscience
Volume 29, Number 12
Figure 2. Evolution of
activation patterns in the neural
model during a single trial in
each condition. Stimulus
displays during different phases
of the task are shown at the top
of each panel. Colored bars
along the time axes indicate
duration of stimulus and cue
inputs in the model. Activation
patterns in the two fields of the
model (for colors red and blue)
are shown color-coded below
for different points during the
course of each trial.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
o
6
c
8
n
9
_
6
a
/
_
j
0
o
1
c
1
n
8
0
_
a
p
_
d
0
1
b
1
y
8
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
the sample array, the activation peaks in both fields settle
at a lower activation level, but they both remain stable
throughout the delay period. When the color cue is pre-
sented at the end of the trial, it raises activation levels in
the blue field, and the activation in the red field is sup-
pressed through mutual inhibition. Only the activation
peak for the blue item remains and forms the basis for
the response generation as before.
The model simulations of the R2-valid trials combine
aspects from the two previous conditions (Figure 2C).
Following the presentation of the sample stimuli, local-
ized activation peaks form in both fields and remain sta-
ble during the first half of the delay period, analogous to
the R2-neutral trial. When the color retro-cue is then pre-
sented, it strengthens activation in the blue field and, in
turn, suppresses activation in the red field. The activation
peak for the blue item then returns again to a lower
activation level when the homogeneous input ceases,
although it remains slightly stronger than during the first
memory period because it no longer experiences inhibi-
tion from a second activation peak. The response period
then proceeds in the same way as in the R1 condition.
Schneegans and Bays
1983
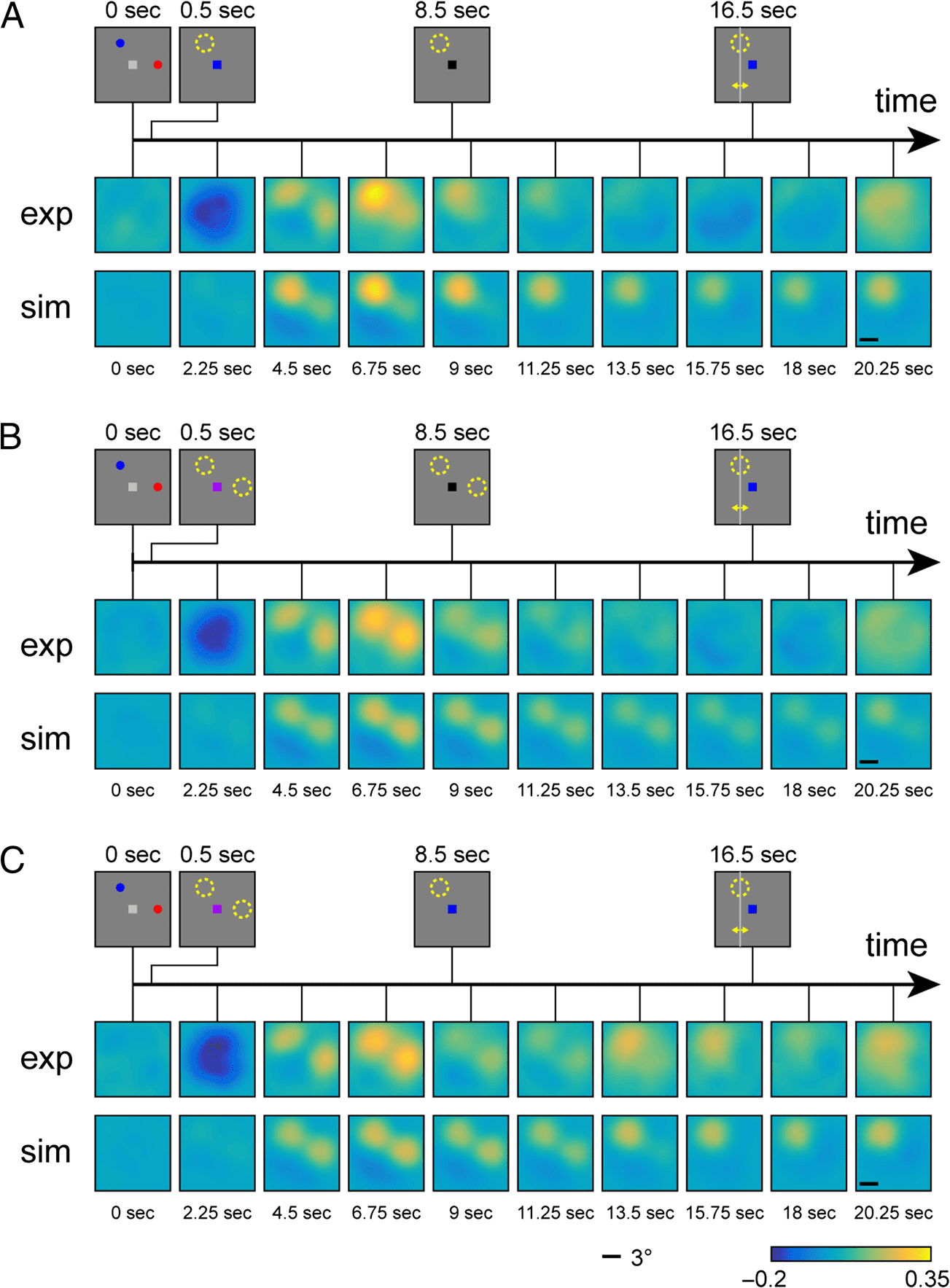
In the averaged and spatially aligned reconstructions
over all trials for each condition (shown for one stimulus
separation in Figure 4), the representations of stimulus
locations are clearly discernible, and the reconstructions
qualitatively match the activation patterns and changes
over time from the fMRI study. In the R1 condition, the
location of the cued and uncued stimulus can be seen
with some delay after stimulus presentation (reconstruc-
tion for time point 4.5 sec), but only the location of the
cued item remains active in the subsequent time points,
with activation slowly diminishing over time. It should be
noted that this slow decrease in activation levels does not
reflect the time course in the neural model itself (where
the activation levels settle into a stable state within less
than one second after stimulus offset) but is created by
the low-pass filter properties of the hemodynamic re-
sponse function.
In the R2-neutral condition, both memory stimuli are
equally represented in reconstructions throughout most
of the trial, appearing with some delay after stimulus pre-
sentation and slowly diminishing in activation until the
response period. The activation time course in the R2-
valid condition is indistinguishable from that in the R2-
neutral condition during the first half of the trial, as
would be expected. After the presentation of the infor-
mative retro-cue, however, the activation increases for
the location of the cued item and disappears for the
location of the uncued item. This reproduces the key
observation in the experimental study that a spatially un-
informative retro-cue can lead to a selective strengthen-
ing of the cued item’s spatial reconstruction in fMRI.
Quantitative Analysis of Reconstructions
Additional quantitative analyses are based on 1-D re-
constructions of the spatial representations, which reflect
activation distributions in an annulus around the fixation
point. These reconstructions are aligned such that the
position of the cued stimulus (the target) is always at
zero and are averaged over all stimulus separations for
each condition. The fidelity of target representations in
these reconstructions is then determined from the con-
centration of activation at the target location.
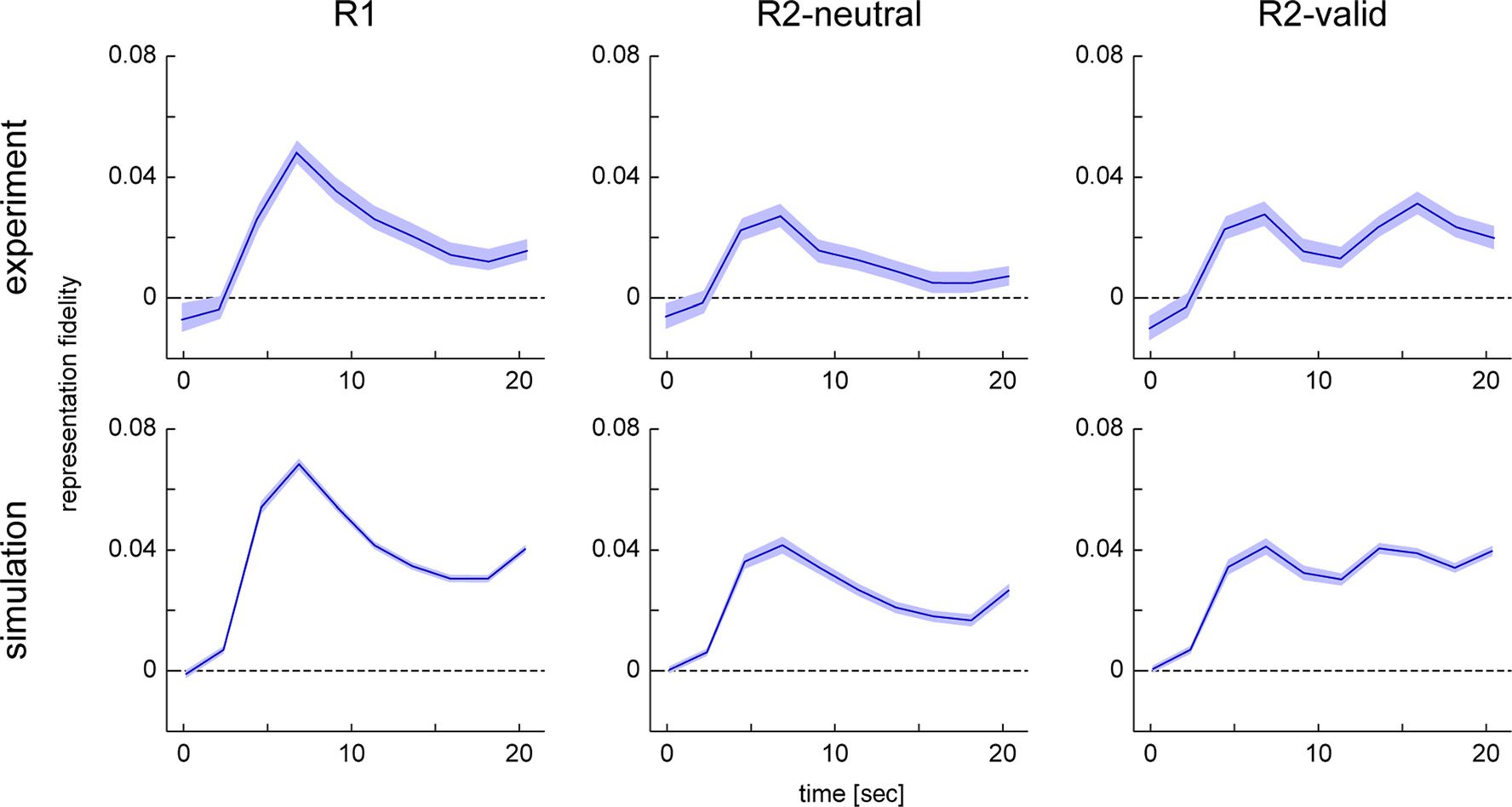
The time course of target representation fidelity for
the three task conditions is shown in Figure 5. Fidelity
of target representation is significantly greater than zero
in all conditions and for all time points starting 2.25 sec
after stimulus presentation. This is to be expected given
that the cued item was continuously represented as a
sustained activation peak in the neural model in all trials
(except for the small proportion of trials in which the peak
collapsed). The fidelity time courses quantitatively repro-
duce the results from the experimental study. Fidelity rises
sharply following the presentation of the stimulus, then
decreases over time in the R1 and R2-neutral conditions,
with overall higher fidelity levels in the R1 than R2-neutral
condition. Critically, in the R2-valid condition, fidelity rises
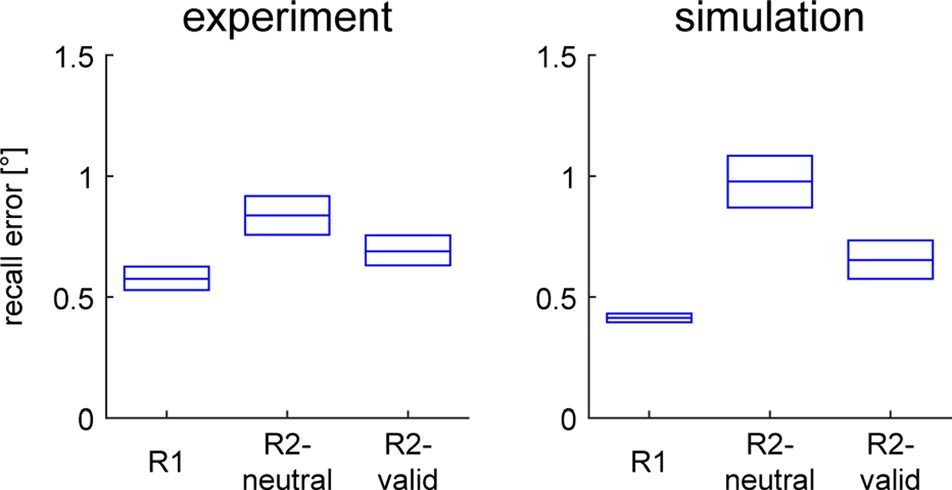
Figure 3. Mean recall errors for different task conditions in experimental
and simulation results. Boxes show 95% confidence interval based on
resampling.
Figure 3 shows the mean recall errors generated by
the model. The model reproduces the pattern of mean
response errors in the fMRI study, with lowest errors in
the R1 condition, highest in the R2-neutral condition, and
intermediate in the R2-valid condition (resampling pro-
cedure producing significant difference for all pairwise
comparisons, p < .001). Deviations of the response posi-
tion from the position of the sample stimulus are gener-
ally caused by random noise in the model. Noise causes
deviations between an external input and the induced
activation peak during encoding and random drift in
the peak location over time (Camperi & Wang, 1998). If
it transiently lowers the activation level in a region by
a sufficient amount, it can also cause the collapse of
a peak. This is much more likely to occur when two lo-
cations are held in memory simultaneously such that
the corresponding activation peaks inhibit each other.
Consequently, the rate of peak collapse in the model is
higher in the R2-neutral condition (21.7% of trials) than
the R2-valid condition (11.6%) and the R1 condition
(0%). In the case of a peak collapse, the homogeneous
input for the color cue during the response phase typi-
cally induces a new activation peak at a random location,
producing higher mean response errors.
Reconstruction of Spatial Representations
To compare our simulation results with the results of the
fMRI study, we first generated simulated voxel BOLD sig-
nals from the output signals (homologue of neural firing
rate) in the model and then performed the same recon-
struction method on these simulated signals that was
used for the experimental data. The generation of simu-
lated BOLD signals and subsequent reconstructions intro-
duces significant noise in the single trial data (compared
with directly plotting the neural field activations for a
trial as in Figure 2), spatial smoothing, and temporal fil-
tering (due to the hemodynamic response function).
Moreover, the purely spatial reconstruction collapses
activation from the separate fields for the two colors into
a single representation.
1984
Journal of Cognitive Neuroscience
Volume 29, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
o
6
c
8
n
9
_
6
a
/
_
j
0
o
1
c
1
n
8
0
_
a
p
_
d
0
1
b
1
y
8
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 4. Two-dimensional
reconstructions of spatial
activation patterns in the
R1 condition (A), R2-neutral
condition (B), and R2-valid
condition (C). In each panel,
the stimulus displays for
different phases of the trial are
shown at the top. The first row
shows reconstructions from
the fMRI study (Sprague et al.,
2016); the second row shows
reconstructions from neural
model simulations.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
o
6
c
8
n
9
_
6
a
/
_
j
0
o
1
c
1
n
8
0
_
a
p
_
d
0
1
b
1
y
8
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
again following the presentation of the retro-cue, repro-
ducing this key signature of the retro-cue effect in the
experimental data.
The fidelity of target representation in the simulation
results also rises at the end of the trial. This is an effect
of the response cue given at this time, which is realized
in the same way as the retro-cue in the neural model. A
rise in fidelity at the end of the trial can also be observed
in the experimental results for the R1 and R2-neutral con-
dition, although weaker than in the simulation. At least
for the R2-neutral condition, we would expect that the
color cue at the end of the trial should have a similar ef-
fect as a cue presented earlier. However, it is possible
that, in the experimental data, there is interference in
the reconstruction from motor planning activity for re-
sponse generation, which is not captured in the model.
We analyze the simulation result that a color cue can in-
crease fidelity even when only a single item is held in
working memory below. We further note that variability
of fidelity within each condition was lower in our simula-
tions than in the experimental data. To fully capture the
experimental results, the model would likely have to in-
corporate additional sources of variability such as fluctu-
ations in attention across trials.
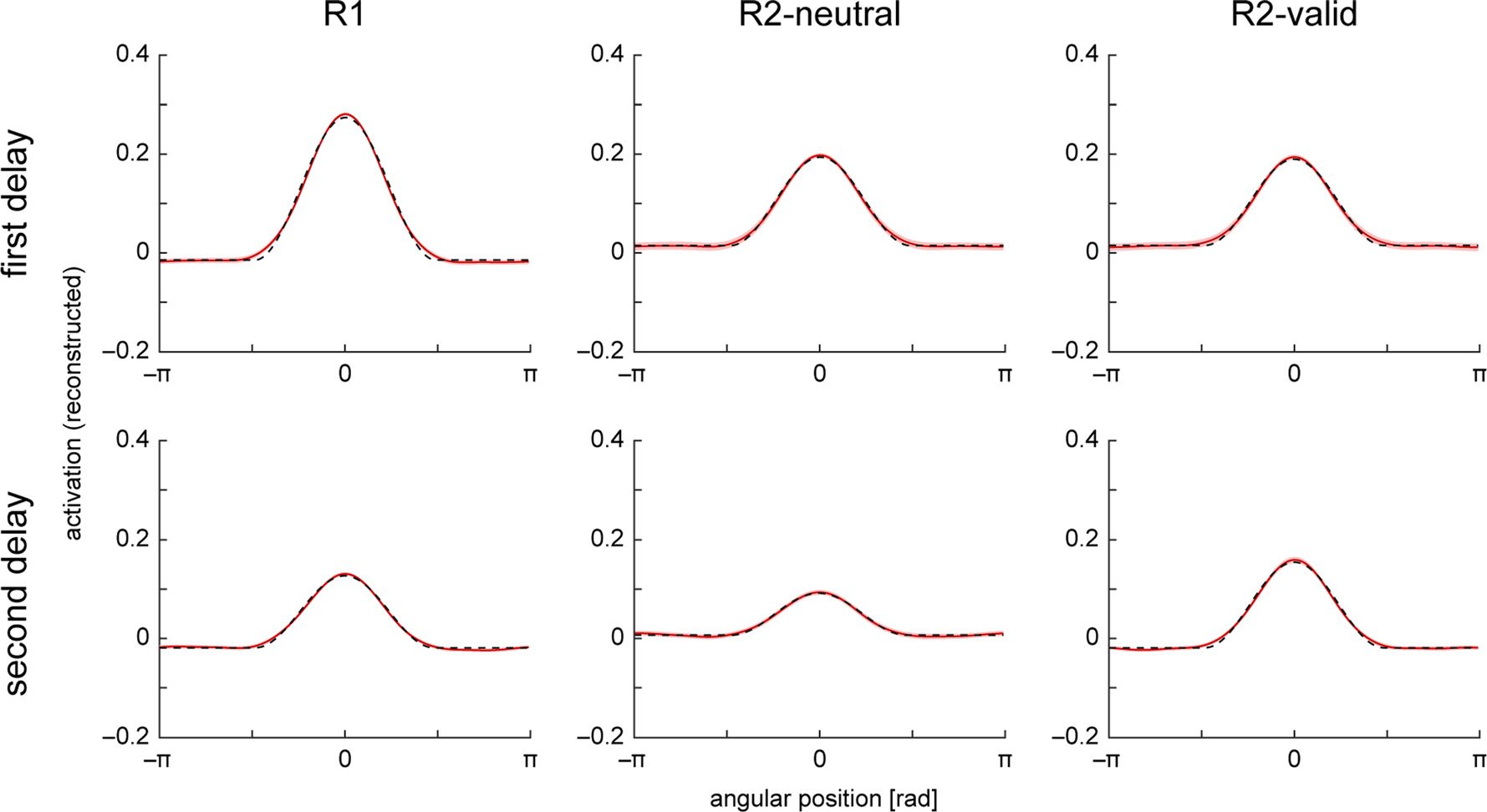
Following the analysis in the fMRI study, we fit the ac-
tivation profiles in all conditions with a Gaussian-like
function with variable baseline, separately for the first
(6.75 and 9 sec) and second memory period (15.75 and
18 sec). The mean activation profiles and function fits are
shown in Figure 6. We used a resampling procedure to
determine significance of differences in the fitted param-
eters between conditions and delay periods. For the first
memory period, we found a significantly higher ampli-
tude of the central activation peak in the R1 condition
compared with both R2-neutral and R2-valid conditions
( p < .001). Conversely, the baseline of the fit was higher
Schneegans and Bays
1985
Figure 5. Time course of representational fidelity for target location in different task conditions for the experimental results (adapted from Sprague
et al., 2016) and model simulations. Shaded areas indicate 95% confidence interval based on resampling.
in the R2-neutral and R2-valid conditions than in the R1
condition ( p < .001). No other significant differences
between the fitted parameters for different conditions
were observed for the first memory period. These find-
ings fully reproduce the observed effects of set size in
the fMRI study.
The findings can be explained from the evolution of
activation in the neural model and the effects of the re-
construction procedure. The target representation in the
R1 condition is stronger due to the color cue that is pre-
sented right after stimulus offset in this condition. This
raises activation in the whole field for the cued color
and, in particular, strengthens the existing activation
peak in this field, yielding higher activation for the cued
item’s location in the reconstruction (in which the fields
for different colors are collapsed). In the neural model,
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
o
6
c
8
n
9
_
6
a
/
_
j
0
o
1
c
1
n
8
0
_
a
p
_
d
0
1
b
1
y
8
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 6. One-dimensional reconstructions over angular space for different task conditions in neural model simulations. Mean activation profiles
across trials are shown separately for the first (top row) and second delay period (bottom row) of the task. Red line shows reconstructions,
with shaded red area indicating 95% confidence interval based on resampling. Dashed black lines show fitted functions.
1986
Journal of Cognitive Neuroscience
Volume 29, Number 12
the single sustained activation peak during the memory
period in the R1 condition is also stronger than the cor-
responding peaks in the R2 conditions due to lack of
mutual inhibition between peaks. The inhibitory inter-
actions implement a form of normalization of total work-
ing memory activity in the model. This aspect is consistent
with continuous resource models of working memory
capacity (Bays, 2014; Bays & Husain, 2008), although we
note that the neural field model also incorporates aspects
of discrete representations linking them to slot models
(Johnson et al., 2014).
In the R2 conditions, the additional sustained peaks for
the nontarget item in each trial are reflected in the acti-
vation profiles. Nontarget positions are offset from the
target position by ±60°, ±120°, or 180° (with some addi-
tional random variations), so that the corresponding ac-
tivation peaks are distributed evenly in orientation space
when the reconstructions are aligned on the target posi-
tion. They contribute therefore to a higher baseline acti-
vation in the fitted function. Because the fit amplitude is
determined as height of the central activation peak over
this baseline, the increased baseline itself also contrib-
utes to a reduced amplitude estimate in the reconstruc-
tions for the R2 condition.
These effects in the model were confirmed by directly
applying the function fits to the field output (summed
over both fields of the model), taken at the midpoint
of the first delay period. As in the reconstructions, we
found a significantly higher amplitude, but lower base-
line, in the R1 condition compared with both R2-neutral
and R2-valid conditions (all ps < .001). The higher base-
line component in the R2 conditions was greatly dimin-
ished when applying the fits only to the field containing
the target item in each trial, confirming that it primarily
reflects contributions from nontarget items. The higher
amplitude for the R1 condition was still preserved in this
case, showing that the activation for the target item is
stronger in this condition.
During the second memory period, the same set size
effects in the reconstructions were observed when com-
paring R1 and R2-neutral condition, with higher ampli-
tude ( p < .001) and lower baseline ( p < .001) in the
R1 condition. In the R2-valid condition, however, target
representation amplitude following the informative retro-
cue is higher than in both the R2-neutral ( p < .001)
and R1 conditions ( p < .001). These results reproduce
analogous findings in the fMRI study. One effect from
that study that we did not observe in the simulation re-
sults was an increased baseline activation in the R2-valid
condition. In fact, we found baseline values in the R2-
valid condition that were significantly lower than in the
R2-neutral condition ( p < .001) and not significantly dif-
ferent from the R1 condition ( p = .95). No further sig-
nificant differences in parameters across conditions
were observed for the second memory periods.
In the neural model, the effects of the retro-cue on
peak amplitude in the second memory period are ex-
plained by the same mechanisms as the set size effects
in the first memory period. The color cue input tran-
siently strengthens the activation peak for the cued item
and, by suppressing the sustained peak for the noncued
item, removes inhibition from that peak for the second
memory period. The color cue also globally raises the
activation level (and, to some extent, the output signal
derived from it) for the whole field of the matching
color. This may, in principle, provide an explanation for
the experimentally observed increase in fit baseline for
the R2-valid condition compared with the R2-neutral
condition. However, in the present model implementa-
tion, this effect is outweighed by the decrease in baseline
that is brought about by the disappearance of the sus-
tained activation peak for the noncued item.
We again tested these explanations by fitting the field
output directly, now at the midpoint of the second delay
period. We found the same effects as in the reconstruc-
tions when comparing R1 and R2-neutral conditions (all
ps < .001). For the R2-valid condition, both amplitude
and baseline were intermediate between R1 and R2-
neutral condition ( p < .001 for all comparisons). Fit
amplitude in this condition did spike during the pre-
sentation of the retro-cue but then quickly diminished
again in the neural model. In the reconstruction, this
transient spike is carried over to the second delay period
due to the low-pass filter properties of the hemodynamic
response function.
Correlations between Reconstructions and
Response Errors
Having demonstrated that the neural model can repro-
duce both behavioral and reconstruction results, we
now analyze interactions between trial-to-trial report
performance and reconstruction quality. We divided the
trials of each block within each condition into two groups
with recall error lower and higher than median recall
error in that block, respectively. We then generated re-
constructions and function fits for each group and each
condition. We found a significantly higher amplitude of
the target representation in the low-error group com-
pared with the high-error group for the R2-neutral and
R2-valid condition in the second delay period (R2-neutral:
mean amplitude 0.058 vs. 0.033, p < .001; R2-valid: mean
amplitude 0.095 vs. 0.076, p = .012). Similar findings
were reported in the experimental study, although a sig-
nificant effect of error rate on fit amplitude was observed
only in the R2-valid condition for the second delay period
and in the R2-neutral condition for the first memory
period. These results were interpreted as evidence that
the reconstruction quality reflects the quality of the
working memory representations.
In the neural simulations, these results can be ex-
plained by the fact that the high-error group will most
likely contain all trials in which the activation peak for
the target item collapsed during the delay period as a
Schneegans and Bays
1987
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
o
6
c
8
n
9
_
6
a
/
_
j
0
o
1
c
1
n
8
0
_
a
p
_
d
0
1
b
1
y
8
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
result of random noise, as well as those trials in which
there was significant drift in peak position. For trials in
which the activation peak collapsed, the reconstruction
will either show no peak or a peak at a new random
location (which can form in response to the retro-cue).
Both will contribute only to baseline activation in the
aligned reconstructions. Trials with high drift of the acti-
vation peak will contribute to a central peak in the recon-
structions, but this peak will be smeared out when trials
are aligned with respect to the stimulus position. Overall,
peak amplitude in the aligned reconstructions will then
be lower in high-error versus low-error trials.
It should be noted here that the retro-cue in the neural
model cannot retroactively restore memory precision.
Although the cue does strengthen the sustained activa-
tion peak for the cued item, any drift in peak position
that occurred before the cue will still be reflected in peak
position after the cue. Also, if the activation peak col-
lapsed before the retro cue was given, the spatially un-
informative cue can only induce a new peak at a random
location. Thus, the interactions between report perfor-
mance and reconstruction quality and the observed in-
crease in reconstruction quality after the retro-cue do not
provide any evidence that a retro-cue actually restores
representational precision. The retro-cue may merely re-
duce further loss of precision after the cue is presented.
Components of the Retro-cue Effect
Using the neural model, we can selectively assess the
contribution of different components of the neural repre-
sentation to the reconstruction quality and its restoration
following the retro-cue. Although we cannot tell from
simulation results alone whether the components iden-
tified here also play a role in the biological neural system,
we can make predictions regarding their signatures in
reconstructions that could be tested with fMRI data.
As a first component in the full reconstruction, we ana-
lyze the effect of the stimulus presentation in the absence
of any working memory processes. To this end, we set
the weight cexc of excitatory interactions in the neural
model to zero. With this modification, the neural popu-
lation representation in the model still forms peaks of
activation in response to external stimulation, but these
peaks collapse as soon as the stimuli disappear (i.e., the
model does not show any working memory capabilities;
Figure 7A).
The time course of reconstruction fidelity for this model
in the R2-valid condition is shown in Figure 7B (the time
courses for the R1 and R2-neutral conditions are virtually
identical in this modified model). During the first half of
the trial, the time course of fidelity is similar to those ob-
served in the full model for the R2-valid and R2-neutral con-
dition, but then fidelity decreases and even becomes
negative during the second half of the trial. This reflects
the shape of the canonical hemodynamic response func-
tion, which likewise takes negative values about 12 sec
after a pulse of activation. These results suggest that a
large part of the early reconstructions are dominated by
a sensory response (and possibly attention effects, which
we do not capture as a separate component in the model),
but it also shows that a working memory component is
necessary to account for the positive reconstruction fidelity
in the latter part of the trial. The simulation results also
confirm that the spatially nonspecific color cue input,
which is provided after the first half of the delay period,
has no effect on reconstruction fidelity in the absence of
sustained activity.
In a second model variant, we assess how removing
irrelevant items from working memory contributes to
the increase of reconstruction fidelity following an infor-
mative retro-cue. In the original model, the retro-cue is
implemented as a positive homogeneous input to the
neural field corresponding to the cue color, which in-
creases activation levels in this field and leads to the ex-
tinction of the activation peak for the uncued item
through global inhibitory interactions. As an alternative,
we now implement the retro-cue as an inhibitory input
to the neural field corresponding to the uncued color
(with amplitude cforget), which extinguishes any activa-
tion peaks in this field without directly strengthening
activation for the cued item (Figure 7C).
Figure 7D shows the time course of reconstruction
fidelity of this model variant in the R2-valid condition.
Removal of the noncued item leads to an increased fidel-
ity in the second half of the trial compared with the
R2-neutral condition (compare to dashed green line).
However, the rise in fidelity following the retro-cue is
weaker and occurs with longer delay than in the original
model. The removal of an irrelevant item thus contributes
to the retro-cue effect in the model but does not fully
account for it.
To test the direct contribution of the cue input to re-
construction quality in the model, we return to the orig-
inal model variant but modify the task. Instead of
presenting two colored stimuli in the beginning, we only
present a single one, which is always the cued item and
response target (Figure 7E). This means that the retro-
cue does not provide any additional information for
the task. Nonetheless, we find that the retro-cue signifi-
cantly increases the reconstruction quality in the model
(Figure 7F) in a manner very similar to the retro-cue effect
in the original task (dashed blue line). Note that overall
reconstruction quality is higher in this condition due to
the absence of the nontarget item.
This result can be explained by interaction effects in
the neural model. The color cue is emulated as a homo-
geneous positive input to the neural field. In the absence
of any sustained activation, this input would simply raise
activation equally over the whole field, which would not
produce any change in the fidelity measure (as demon-
strated in the model variant without working memory
states; Figure 7B). When a sustained peak of activation
is present, activation initially also rises homogeneously;
1988
Journal of Cognitive Neuroscience
Volume 29, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
o
6
c
8
n
9
_
6
a
/
_
j
0
o
1
c
1
n
8
0
_
a
p
_
d
0
1
b
1
y
8
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
o
6
c
8
n
9
_
6
a
/
_
j
0
o
1
c
1
n
8
0
_
a
p
_
d
0
1
b
1
y
8
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 7. Evolution of activation patterns and fidelity time courses in model simulations for different model and task variants based on the original
R2-valid condition. (A, B) Neural model without lateral excitatory interactions. (C, D) Model variant with color cue realized through inhibition of
noncued items. (E, F) Task variant with only the target item presented in the sample array. (G, H) Model variant with weak (unreliable) retro-cue
input. The field activations are shown for the case of a valid retro-cue; fidelity plots are shown for both valid (blue) and invalid (red) retro-cue.
The dashed lines in the fidelity plots show the mean fidelity from the original R2-valid (blue) and R2-neutral condition (green) for comparison.
Shaded areas in fidelity plots indicate 95% confidence interval based on resampling.
Schneegans and Bays
1989
but as it does, the field output that drives lateral inter-
actions (computed from the activation via a sigmoid
function, Equation 2) increases most strongly at the
edges of the existing peak. The peak expands further
under the influence of local excitation and consequently
suppresses activation in the remaining field (as well as
the second, noncued field) through global inhibition
(Figure 7E). Overall, the concentration of activation at
the location of the existing peak is increased, leading to
the increase in measured reconstruction fidelity.
We note that, for this simulation, we assumed that the
cue would have the same effect as in the original task de-
spite being uninformative. Human observers would likely
pay less attention to the cue or ignore it entirely under
these conditions, leading to diminished effects on work-
ing memory representations. We did not observe any
benefit of the retro-cue on behavioral performance in
the model (compared with simulations with a single tar-
get stimulus without retro-cue; mean response error in
both cases 0.41°, p = .91). This is due to the fact that
behavioral cueing effects in the model are based on bias-
ing the competition between different working memory
items, which does not occur in the case of a single item.
Generalization of the Model to
Unreliable Retro-cues
The experiment we emulated with the neural model
(Sprague et al., 2016) exclusively uses retro-cues with
100% reliability, but many studies on retro-cues use
lower reliabilities, such that participants have an incen-
tive to retain uncued items in memory as well (Souza &
Oberauer, 2016). Here, we test whether the neural model
can be adapted to this scenario. We make a single change
to the implementation, namely reducing the strength of
the homogeneous boost input that emulates the color
cue (amplitude 2.5 compared with 17.5 in the original
model; the amplitude for the definitive response cue at
the end of the trial is left unchanged). With this setting,
the input transiently strengthens the sustained activation
for the cued item but does not increase its activation
levels to such a degree that the peak for the noncued
item will be extinguished (Figure 7G).
We tested the model in trials with either a valid
(matching the target color) or invalid retro-cue (match-
ing the nontarget color) provided after the first half of
the delay period and an unambiguous response cue indi-
cating the target at the end. The fidelity time course for
these simulations is shown in Figure 7H. The valid retro-
cue causes a small rise in fidelity, whereas the invalid
retro-cue causes a faster drop in the representational fidel-
ity of the target compared with the original R2-neutral
condition (dashed green line). Nonetheless, even with an
invalid retro-cue the fidelity remains above zero through-
out the trial and recovers when the response cue is given,
indicating that the uncued item is retained in working
memory in a majority of trials.
We found a significant effect of cue validity on re-
sponse errors, with response error lower than in the
original R2-neutral condition for valid retro-cues (mean
error 0.74°) and higher for invalid retro-cues (1.46°; sig-
nificantly different based on resampling procedure, p <
.001). This qualitatively reproduces the most prominent
effect of retro-cueing in the literature (see, e.g., Pertzov,
Bays, Joseph, & Husain, 2013). The retro-cue effect on
performance was driven by a lower rate of peak col-
lapse in the valid than in the invalid retro-cue condition
(14.0% vs. 43.5%). We did not find a significant differ-
ence in precision for those trials where the peak was
sustained (mean response error 0.41° for both condi-
tions, p = .96), likely due to the fact that the separate
fields for red and blue item do not allow any local inter-
actions between the two activation peaks in the model.
Behavioral studies on retro-cue effects have consistently
shown a benefit for the probability of reporting the cor-
rect item (rather than guessing or reporting the feature
of nontarget item), with only some of the studies show-
ing a precision benefit (see Souza & Oberauer, 2016, for
a review). We note that our model does not produce
any swap errors, which contribute significantly to non-
target responses in behavioral studies. These might
potentially be addressed by extending the model with
an additional purely spatial memory representation that
does not capture binding information (Matthey, Bays, &
Dayan, 2015).
DISCUSSION
We have presented a neurodynamic model of retro-cue
effects in visual working memory, and we have demon-
strated that this model can reproduce behavioral and re-
construction results from an fMRI study, which were
interpreted as supporting a latent working memory state
(Sprague et al., 2016). The study had found an increase in
reconstruction quality after a retro-cue, as well as cor-
relations between reconstruction quality and response
error. The authors had argued that these findings provide
evidence for a working memory state that is not detect-
able in the fMRI signal but that can be reactivated by the
retro-cue. They proposed that this was likely an activity-
silent state, encoded, for example, in changes in synaptic
connectivity or sustained subthreshold changes in mem-
brane potential (Stokes, 2015). The neural model pre-
sented here does not feature such a latent memory
state, relying entirely on sustained neural activity for
working memory representations (Wang, 2001). The fact
that the model can nonetheless reproduce all key exper-
imental findings refutes the argument for an activity-
silent working memory state. Such a state may still exist,
but it is not necessary to explain any of the experimental
findings from the fMRI study.
We aimed to keep the neural model as simple as pos-
sible, making only minimal assumptions about the neural
architecture beyond the basic mechanism of sustained
1990
Journal of Cognitive Neuroscience
Volume 29, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
o
6
c
8
n
9
_
6
a
/
_
j
0
o
1
c
1
n
8
0
_
a
p
_
d
0
1
b
1
y
8
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
activity through self-excitation. In particular, we em-
ployed a continuous approximation of the discrete spik-
ing activity in biological neural systems. Previous models
of sustained memory activity using continuous firing
rates have produced results very similar to more detailed
spiking models (Compte et al., 2000; Camperi & Wang,
1998), so we believe that this simplification does not
qualitatively alter model behavior. As a further simplifica-
tion, we did not separately model individual cortical areas
or distinguish between sensory and working memory
representations. The simulation results suggest that a
large portion of the fMRI signal in the experimental study
is driven directly by the visual stimuli (which may comprise
both purely sensory activity and working memory encod-
ing), but a sustained working memory component is still
necessary to fully explain the signal. Spatial attention may
also contribute to the fMRI signal and the observed retro-
cue effect, given that previous behavioral studies have
found that a color cue can enhance spatial attention for a
memorized item location even if the location is not task
relevant (Theeuwes, Kramer, & Irwin, 2011). However,
neither the experimental study nor the model presented
here investigates the role of attention in this task.
One key assumption we did make in the model is that
the working memory representation captures the bind-
ing between colors and locations (Treisman, 1996). Bind-
ing information is essential to solve the behavioral task
(reporting the location of the cued item, rather than
randomly reporting the location of either stimulus) but
is not reflected in the reconstructions used by Sprague
et al. (2016). Following previous neural field models of
feature binding in visual working memory (Schneegans
et al., 2016; Johnson, Spencer, & Schöner, 2008), we
employ a population representation with conjunctive
coding. This kind of representation has recently been
shown to account for behavioral signatures of binding
failures in working memory tasks (Schneegans & Bays,
2017; Matthey et al., 2015). Some of the previous imple-
mentations have combined the conjunctive coding with
separate feature-specific representation for more efficient
representation of individual feature values (Schneegans
et al., 2016; Matthey et al., 2015). We omitted such a
mechanism here for the sake of simplicity, but we note
that some of the neural representations for working
memory and attention that contribute to the fMRI signal
are likely to be purely spatial and only indirectly interact
with representations of stimulus color.
The proposed model constitutes to our knowledge the
first neural process model of retro-cue effects. The neu-
rodynamic mechanism employed here integrates several
hypotheses about the mechanisms underlying retro-cue
effects (Souza & Oberauer, 2016). The cue directly in-
creases activation of the sustained activation peak that
reflects the cued item, making use of the conjunctive
coding to increase activation in the spatial representation
in response to a color cue. This realizes a form of refresh-
ing of the cued item (Rerko & Oberauer, 2013; Chun &
Johnson, 2011). Increasing the activation of one working
memory item reduces activation for other items by
means of lateral inhibition in the model, consistent with
resource models of working memory (Bays, 2014; van
den Berg, Shin, Chou, George, & Ma, 2012; Bays, Catalao,
& Husain, 2009; Bays & Husain, 2008). For fully reliable
retro-cues, this leads to the complete extinction of other
working memory items, consistent with the idea that retro-
cues act through removal of irrelevant items (Souza et al.,
2014; Kuo, Stokes, & Nobre, 2012). The model also qualita-
tively captures effects of less reliable cues, which decrease
the chance of forgetting the cued item (through noise-
induced collapse of the sustained activation peak) at the
cost of an increased chance of forgetting noncued items.
All performance effects of the retro-cue in the present
model are due to protection of the cued item from decay
after the retro-cue. This is consistent with previous find-
ings from delayed estimation tasks showing a continuous
decrease of response precision with retention interval
duration that can be alleviated by a valid retro-cue (Pertzov
et al., 2013). This explanation has been questioned by
Souza and Oberauer (2016) based on contradictory find-
ings from Gressmann and Janczyk (2016), but these latter
findings were based on a change detection task, which
may not provide sufficient sensitivity to detect changes in
memory precision. Souza and Oberauer (2016) also point
to findings that retro-cues increase performance compared
with no-cue trials with shorter delay period (matched to
the time before the retro-cue; Makovski, Sussman, & Jiang,
2008). A possible cause for this effect is that the retro-cue
triggers the retrieval of the cued item without the per-
ceptual interference that may be created by elements of
the response display (such as probe stimuli in change de-
tection tasks or a color wheel in delayed estimation tasks;
Souza, Rerko, & Oberauer, 2016; Makovski, Watson,
Koutstaal, & Jiang, 2010). Accounting for such effects is
beyond the scope of the present model.
The retro-cue mechanism in the neural model accounts
for the restoration of reconstruction quality for the target
item’s location that was observed in the fMRI study. Sprague
et al. (2016) interpreted this effect of the retro-cue as evi-
dence for an activity-silent working memory state based
on the following reasoning: By the time the retro-cue was
given, the working memory representation of the memo-
rized location as measured by fMRI reconstruction quality
was already degraded to a certain degree; the retro-cue
did not provide any spatial information, so it could not have
improved the representation of the target’s location; thus,
the improved reconstruction quality after the retro-cue must
have been caused by the reactivation of an activity-silent
working memory representation (realized, e.g., through
changes in synaptic connectivity) that was not detectable
in the fMRI data before the retro-cue.
We argue that there are several problems with this line of
reasoning. First, although the color cue does not provide
any spatial information by itself, it does allow the selection
of a single memorized location if the working memory
Schneegans and Bays
1991
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
o
6
c
8
n
9
_
6
a
/
_
j
0
o
1
c
1
n
8
0
_
a
p
_
d
0
1
b
1
y
8
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
representation provides information about the binding of
colors to locations, as we have argued above. This allows, at
the very least, the selective removal of uncued items from
working memory. We have demonstrated in model simula-
tions that the selective removal of uncued items produces
an increase in reconstruction quality for the cued item
using the fidelity measure proposed by Sprague et al.
(2016). The reconstruction method does not attempt to
estimate memorized color information and thereby loses
part of the information that is necessary to explain the
effect of the color cue on the spatial representation.
Second, even an entirely uninformative cue can pro-
duce an increase in reconstruction quality through the
effects of neural interactions in conjunction with sus-
tained activation states. We have shown in the simu-
lations that, even when only a single item is held in
working memory, a color cue for that item produces a
rise in the fidelity of its reconstruction. The same color
cue does not affect fidelity in the absence of sustained
working memory activity. In this latter case, the cue only
produces a homogeneous rise in activation. But in the
presence of a sustained activation peak, the elementary
connection pattern of local excitation and surround inhi-
bition in the model transforms this homogeneous input
into a specific increase of activation for the existing peak.
In a more complex neural architecture with multiple inter-
connected representations, the combination of a non-
specific input with a sustained activation state may even
have much more dramatic effects. Such effects have been
employed to drive complex state transitions in neuro-
dynamic models (Schneegans et al., 2016; Richter et al., 2014).
The results of other reconstruction studies may still
pose a challenge for the present model. In several exper-
iments, decoding quality was found to fall to chance level
for memorized items outside the focus of attention but
could be restored at a later time following informative
cues or unspecific stimulation ( Wolff et al., 2017; Rose
et al., 2016). In the neural model, once sustained activity
has ceased and the activation peak has collapsed, it can-
not be restored by unspecific external stimulation. It is
possible, however, that the experimental studies merely
failed to detect the signatures of weak neural activity that
was sustained throughout the memory period. This
might occur because the relationship between firing rate
and BOLD/EEG signal is less straightforward than the
direct linear relationship assumed here (e.g., Logothetis,
Pauls, Augath, Trinath, & Oeltermann, 2001). If this is the
case, mechanisms similar to the ones described in this
study may explain how weak sustained activity can be
amplified and become detectable again in response to
unspecific input to the neural system.
Acknowledgments
We thank Thomas C. Sprague, Edward F. Ester, and John T.
Serences for making data and analysis code from their study avail-
able online. This work was supported by the Wellcome Trust.
Reprint requests should be sent to Sebastian Schneegans,
Department of Psychology, University of Cambridge, Downing
Street, Cambridge CB2 3EB, UK, or via e-mail: ss2361@cam.ac.uk.
REFERENCES
Amari, S. (1977). Dynamics of pattern formation in lateral-
inhibition type neural fields. Biological Cybernetics, 27,
77–87.
Barak, O., & Tsodyks, M. (2014). Working models of working
memory. Current Opinion in Neurobiology, 25, 20–24.
Bays, P. M. (2014). Noise in neural populations accounts for
errors in working memory. Journal of Neuroscience, 34,
3632–3645.
Bays, P. M. (2015). Spikes not slots: Noise in neural populations
limits working memory. Trends in Cognitive Sciences, 19,
431–438.
Bays, P. M., Catalao, R. F., & Husain, M. (2009). The precision
of visual working memory is set by allocation of a shared
resource. Journal of Vision, 9, 7.1–7.11.
Bays, P. M., & Husain, M. (2008). Dynamic shifts of limited
working memory resources in human vision. Science, 321,
851–854.
Burak, Y., & Fiete, I. R. (2012). Fundamental limits on persistent
activity in networks of noisy neurons. Proceedings of the
National Academy of Sciences, U.S.A., 109, 17645–17650.
Camperi, M., & Wang, X.-J. (1998). A model of visuospatial
working memory in prefrontal cortex: Recurrent network and
cellular bistability. Journal of Computational Neuroscience,
5, 383–405.
Chaudhuri, R., & Fiete, I. (2016). Computational principles of
memory. Nature Neuroscience, 19, 394–403.
Chun, M. M., & Johnson, M. K. (2011). Memory: Enduring traces
of perceptual and reflective attention. Neuron, 72, 520–535.
Compte, A., Brunel, N., Goldman-Rakic, P. S., & Wang, X.-J.
(2000). Synaptic mechanisms and network dynamics
underlying spatial working memory in a cortical network
model. Cerebral Cortex, 10, 910–923.
Courtney, S. M., Ungerleider, L. G., Keil, K., & Haxby, J. V.
(1997). Transient and sustained activity in a distributed
neural system for human working memory. Nature, 386,
608–611.
Edin, F., Macoveanu, J., Olesen, P., Tegnér, J., & Klingberg, T.
(2007). Stronger synaptic connectivity as a mechanism
behind development of working memory-related brain
activity during childhood. Journal of Cognitive Neuroscience,
19, 750–760.
Emrich, S. M., Riggall, A. C., LaRocque, J. J., & Postle, B. R.
(2013). Distributed patterns of activity in sensory cortex
reflect the precision of multiple items maintained in visual
short-term memory. Journal of Neuroscience, 33, 6516–6523.
Engel, T. A., & Wang, X.-J. (2011). Same or different? A neural
circuit mechanism of similarity-based pattern match decision
making. Journal of Neuroscience, 31, 6982–6996.
Erickson, M. A., Maramara, L. A., & Lisman, J. (2010). A single
brief burst induces glur1-dependent associative short-term
potentiation: A potential mechanism for short-term memory.
Journal of Cognitive Neuroscience, 22, 2530–2540.
Eriksson, J., Vogel, E. K., Lansner, A., Bergström, F., & Nyberg,
L. (2015). Neurocognitive architecture of working memory.
Neuron, 88, 33–46.
Funahashi, S., Bruce, C. J., & Goldman-Rakic, P. S. (1989).
Mnemonic coding of visual space in the monkey’s
dorsolateral prefrontal cortex. Journal of Neurophysiology,
61, 331–349.
Fuster, J. M., & Alexander, G. E. (1971). Neuron activity related
to short-term memory. Science, 173, 652–654.
1992
Journal of Cognitive Neuroscience
Volume 29, Number 12
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
o
6
c
8
n
9
_
6
a
/
_
j
0
o
1
c
1
n
8
0
_
a
p
_
d
0
1
b
1
y
8
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Gressmann, M., & Janczyk, M. (2016). The (un)clear effects of
invalid retro-cues. Frontiers in Psychology, 7, 244.
Harrison, S. A., & Tong, F. (2009). Decoding reveals the
contents of visual working memory in early visual areas.
Nature, 458, 632–635.
Johnson, J. S., Simmering, V. R., & Buss, A. T. (2014). Beyond
slots and resources: Grounding cognitive concepts in
neural dynamics. Attention, Perception, & Psychophysics,
76, 1630–1654.
Johnson, J. S., Spencer, J. P., & Schöner, G. (2008). Moving to
higher ground: The dynamic field theory and the dynamics
of visual cognition. New Ideas in Psychology, 26, 227–251.
Johnson, J. S., Spencer, J. P., & Schöner, G. (2009). A layered
neural architecture for the consolidation, maintenance, and
updating of representations in visual working memory. Brain
Research, 1299, 17–32.
Kuo, B.-C., Stokes, M. G., & Nobre, A. C. (2012). Attention
modulates maintenance of representations in visual short-term
memory. Journal of Cognitive Neuroscience, 24, 51–60.
LaRocque, J. J., Lewis-Peacock, J. A., Drysdale, A. T., Oberauer,
K., & Postle, B. R. (2013). Decoding attended information in
short-term memory: An EEG study. Journal of Cognitive
Neuroscience, 25, 127–142.
Lewis-Peacock, J. A., Drysdale, A. T., Oberauer, K., & Postle, B. R.
(2012). Neural evidence for a distinction between short-term
memory and the focus of attention. Journal of Cognitive
Neuroscience, 24, 61–79.
Linden, D. E., Bittner, R. A., Muckli, L., Waltz, J. A., Kriegeskorte,
N., Goebel, R., et al. (2003). Cortical capacity constraints for
visual working memory: Dissociation of fMRI load effects in
a fronto-parietal network. Neuroimage, 20, 1518–1530.
Lindquist, M. A., Loh, J. M., Atlas, L. Y., & Wager, T. D. (2009).
Modeling the hemodynamic response function in fMRI:
Efficiency, bias and mis-modeling. Neuroimage, 45, S187–S198.
Lipinski, J., Schneegans, S., Sandamirskaya, Y., Spencer, J. P., &
Schöner, G. (2012). A neurobehavioral model of flexible
spatial language behaviors. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 38, 1490.
Logothetis, N. K., Pauls, J., Augath, M., Trinath, T., &
Oeltermann, A. (2001). Neurophysiological investigation of
the basis of the fMRI signal. Nature, 412, 150–157.
Makovski, T., Sussman, R., & Jiang, Y. V. (2008). Orienting
attention in visual working memory reduces interference
from memory probes. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 34, 369.
Makovski, T., Watson, L. M., Koutstaal, W., & Jiang, Y. V. (2010).
Method matters: Systematic effects of testing procedure on
visual working memory sensitivity. Journal of Experimental
Psychology: Learning, Memory, and Cognition, 36, 1466.
Matthey, L., Bays, P. M., & Dayan, P. (2015). A probabilistic
palimpsest model of visual short-term memory. PLoS
Computational Biology, 11, e1004003.
Mi, Y., Katkov, M., & Tsodyks, M. (2017). Synaptic correlates
of working memory capacity. Neuron, 93, 323–330.
Mongillo, G., Barak, O., & Tsodyks, M. (2008). Synaptic theory
of working memory. Science, 319, 1543–1546.
Pertzov, Y., Bays, P. M., Joseph, S., & Husain, M. (2013). Rapid
forgetting prevented by retrospective attention cues. Journal
of Experimental Psychology: Human Perception and
Performance, 39, 1224.
Rerko, L., & Oberauer, K. (2013). Focused, unfocused, and
defocused information in working memory. Journal of
Experimental Psychology: Learning, Memory, and
Cognition, 39, 1075.
Richter, M., Lins, J., Schneegans, S., & Schöner, G. (2014). A
neural dynamic architecture resolves phrases about spatial
relations in visual scenes. In S. Wermter, C. Weber, W. Duch,
T. Honkela, O. Koprinkova-Hristova, S. Magg, G. Palm, &
A. E. P. Villa (Eds.), International Conference on Artificial
Neural Networks (pp. 201–208). Cham: Springer.
Riggall, A. C., & Postle, B. R. (2012). The relationship between
working memory storage and elevated activity as measured
with functional magnetic resonance imaging. Journal of
Neuroscience, 32, 12990–12998.
Rose, N. S., LaRocque, J. J., Riggall, A. C., Gosseries, O., Starrett,
M. J., Meyering, E. E., et al. (2016). Reactivation of latent
working memories with transcranial magnetic stimulation.
Science, 354, 1136–1139.
Schneegans, S., & Bays, P. M. (2017). Neural architecture for
feature binding in visual working memory. Journal of
Neuroscience, 37, 3913–3925.
Schneegans, S., & Schöner, G. (2008). Dynamic field theory as
a framework for understanding embodied cognition.
In P. Calvo & T. Gomila (Eds.), Handbook of cognitive
science: An embodied approach (pp. 241–271). Amsterdam:
Elsevier.
Schneegans, S., Spencer, J. P., & Schöner, G. (2016). Integrating
“what” and “where”: Visual working memory for objects in
a scene. In G. Schöner & J. P. Spencer (Eds.), Dynamic
thinking: A primer on dynamic field theory (pp. 197–226).
New York: Oxford University Press.
Schneegans, S., Spencer, J. P., Schöner, G., Hwang, S., &
Hollingworth, A. (2014). Dynamic interactions between visual
working memory and saccade target selection. Journal of
Vision, 14, 1–23.
Schutte, A. R., & Spencer, J. P. (2009). Tests of the dynamic field
theory and the spatial precision hypothesis: Capturing a
qualitative developmental transition in spatial working
memory. Journal of Experimental Psychology: Human
Perception and Performance, 35, 1698.
Serences, J. T., Ester, E. F., Vogel, E. K., & Awh, E. (2009).
Stimulus-specific delay activity in human primary visual
cortex. Psychological Science, 20, 207–214.
Simmering, V. R., Spencer, J. P., & Schöner, G. (2006).
Reference-related inhibition produces enhanced position
discrimination and fast repulsion near axes of symmetry.
Attention, Perception, & Psychophysics, 68, 1027–1046.
Souza, A. S., & Oberauer, K. (2016). In search of the focus
of attention in working memory: 13 Years of the retro-cue
effect. Attention, Perception, & Psychophysics, 78,
1839–1860.
Souza, A. S., Rerko, L., & Oberauer, K. (2014). Unloading and
reloading working memory: Attending to one item frees
capacity. Journal of Experimental Psychology: Human
Perception and Performance, 40, 1237.
Souza, A. S., Rerko, L., & Oberauer, K. (2016). Getting
more from visual working memory: Retro-cues enhance
retrieval and protect from visual interference. Journal of
Experimental Psychology: Human Perception and
Performance, 42, 890.
Sprague, T. C., Ester, E. F., & Serences, J. T. (2014).
Reconstructions of information in visual spatial working
memory degrade with memory load. Current Biology, 24,
2174–2180.
Sprague, T. C., Ester, E. F., & Serences, J. T. (2016). Restoring
latent visual working memory representations in human
cortex. Neuron, 91, 694–707.
Stokes, M. G. (2015). “Activity-silent” working memory in
prefrontal cortex: A dynamic coding framework. Trends in
Cognitive Sciences, 19, 394–405.
Tanoue, R. T., & Berryhill, M. E. (2012). The mental wormhole:
Internal attention shifts without regard for distance.
Attention, Perception, & Psychophysics, 74, 1199–1215.
Theeuwes, J., Kramer, A. F., & Irwin, D. E. (2011). Attention on
our mind: The role of spatial attention in visual working
memory. Acta Psychologica, 137, 248–251.
Schneegans and Bays
1993
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
o
6
c
8
n
9
_
6
a
/
_
j
0
o
1
c
1
n
8
0
_
a
p
_
d
0
1
b
1
y
8
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Todd, J. J., & Marois, R. (2004). Capacity limit of visual short-
term memory in human posterior parietal cortex. Nature,
428, 751–754.
neural fields: An integrative cognitive neuroscience
approach. Journal of Mathematical Psychology, 76,
212–235.
Treisman, A. (1996). The binding problem. Current Opinion in
Neurobiology, 6, 171–178.
van den Berg, R., Shin, H., Chou, W.-C., George, R., & Ma,
W. J. (2012). Variability in encoding precision accounts for
visual short-term memory limitations. Proceedings of the
National Academy of Sciences, U.S.A., 109, 8780–8785.
Wang, X.-J. (2001). Synaptic reverberation underlying mnemonic
persistent activity. Trends in Neurosciences, 24, 455–463.
Wei, Z., Wang, X.-J., & Wang, D.-H. (2012). From distributed
resources to limited slots in multiple-item working memory:
A spiking network model with normalization. Journal of
Neuroscience, 32, 11228–11240.
Wijeakumar, S., Ambrose, J. P., Spencer, J. P., & Curtu, R.
(2016). Model-based functional neuroimaging using dynamic
Wimmer, K., Nykamp, D. Q., Constantinidis, C., & Compte, A.
(2014). Bump attractor dynamics in prefrontal cortex explains
behavioral precision in spatial working memory. Nature
Neuroscience, 17, 431–439.
Wolff, M. J., Ding, J., Myers, N. E., & Stokes, M. G. (2015).
Revealing hidden states in visual working memory using
electroencephalography. Frontiers in Systems Neuroscience,
9, 123.
Wolff, M. J., Jochim, J., Akyürek, E. G., & Stokes, M. G. (2017).
Dynamic hidden states underlying working-memory-guided
behavior. Nature Neuroscience, 20, 864–871.
Xu, Y., & Chun, M. M. (2006). Dissociable neural mechanisms
supporting visual short-term memory for objects. Nature,
440, 91–95.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
2
9
/
1
1
9
2
7
/
7
1
1
9
9
7
5
7
3
/
2
1
8
0
7
8
o
6
c
8
n
9
_
6
a
/
_
j
0
o
1
c
1
n
8
0
_
a
p
_
d
0
1
b
1
y
8
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
1994
Journal of Cognitive Neuroscience
Volume 29, Number 12