Levi, P.T., Chopra, S., Pang, J.C., Holmes, A., Gajwani, M., Sassenberg, T.A., DeYoung, C.G. & Fornito, A. (2023). The effect
of using group-averaged or individualized brain parcellations when investigating connectome dysfunction in psychosis.
Netzwerkneurowissenschaften, Advance publication. https://doi.org/10.1162/netn_a_00329.
The effect of using group-averaged or individualized brain
parcellations when investigating connectome dysfunction in
psychosis
Short title: Individualized parcellation and dysconnectivity in psychosis
Priscila T. Levi1, Sidhant Chopra2, James C. Pang1, Alexander Holmes1, Mehul Gajwani1,
Tyler A. Sassenberg3, Colin G. DeYoung3, Alex Fornito1
1. Turner Institute for Brain and Mental Health, Monash University, Melbourne, Australia
2. Abteilung für Psychologie, Yale Universität, New Haven, USA
3. Abteilung für Psychologie, University of Minnesota, Minnesota, USA
1
2
3
4
5
6
7
8
9
10
11
Abstrakt
12
13
14
15
16
17
18
19
20
21
22
23
Functional magnetic resonance imaging (fMRT) is widely used to investigate functional
coupling (FC) disturbances in a range of clinical disorders. Most analyses performed to date
have used group-based parcellations for defining regions of interest (ROIs), in which a single
parcellation is applied to each brain. This approach neglects individual differences in brain
functional organization and may inaccurately delineate the true borders of functional regions.
These inaccuracies could inflate or under-estimate group differences in case-control analyses.
Wir
investigated how
individual differences
in brain organization
influence group
comparisons of FC using psychosis as a case-study, drawing on fMRI data in 121 early
psychosis patients and 57 Kontrollen. We defined FC networks using either a group-based
parcellation or an individually-tailored variant of the same parcellation. Individualized
parcellations yielded more functionally homogeneous ROIs than group-based parcellations.
At individual connections level, case-control FC differences were widespread, but the group-
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
/
T
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
D
Ö
ich
/
.
/
T
/
1
0
1
1
6
2
N
e
N
_
A
_
0
0
3
2
9
2
1
4
2
0
1
4
N
e
N
_
A
_
0
0
3
2
9
P
D
.
T
/
F
B
j
G
u
e
S
T
T
Ö
N
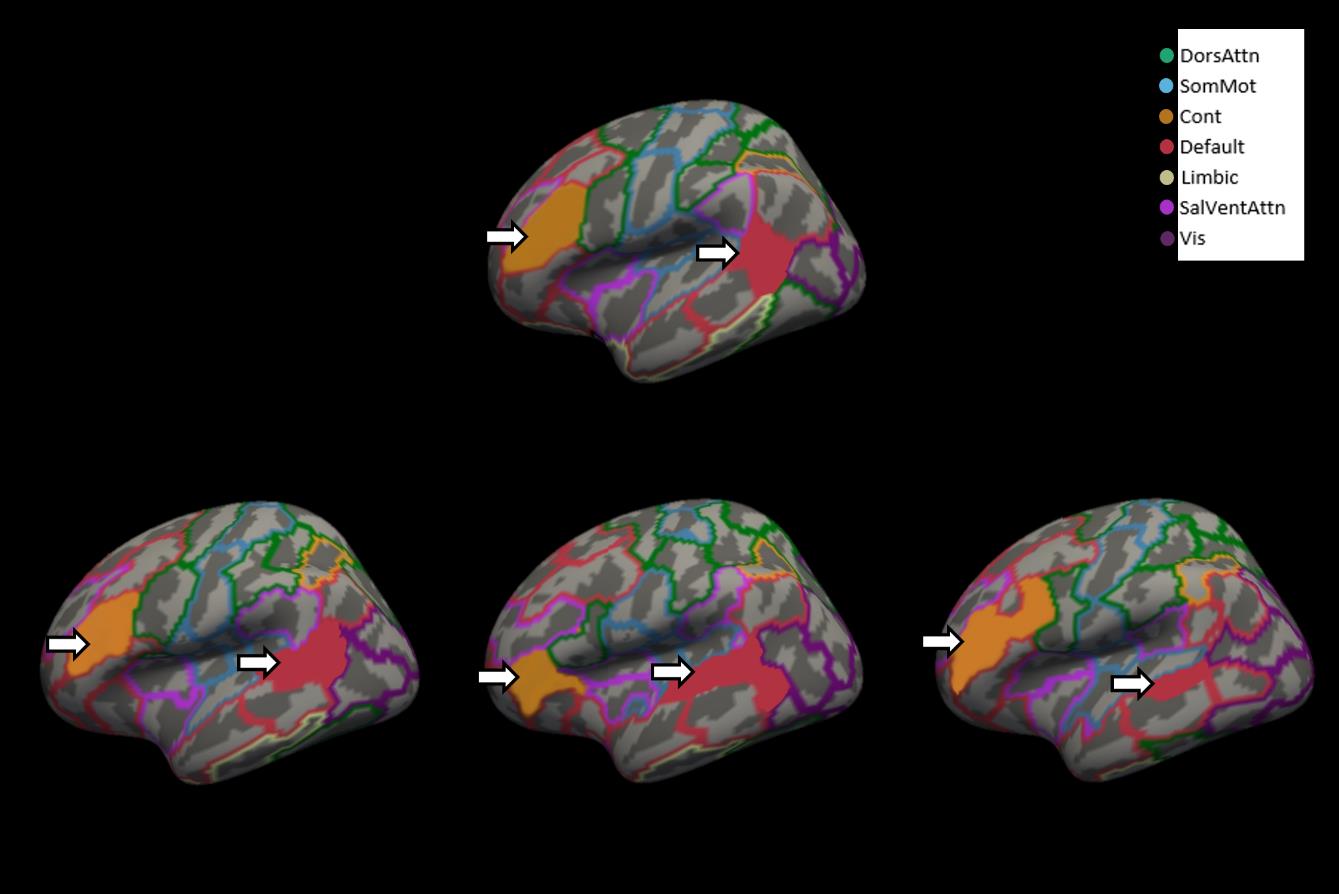
0
7
S
e
P
e
M
B
e
R
2
0
2
3
24
25
26
27
28
based parcellation identified approximately 9% connections as dysfunctional than the
individualized parcellation. When considering differences at the level of functional networks,
the results from both parcellations converged. Our results suggest that a substantial fraction
of dysconnectivity previously observed in psychosis may be driven by the parcellation
method, rather than a pathophysiological process related to psychosis.
29
Author summary
Functional magnetic resonance imaging is widely used to map how brain network
dysfunction is affected by diverse diseases. A fundamental step in this work involved
defining specific brain regions, which act as network nodes in the analysis. Most research to
date has used a one-size-fits all approach, defining such regions on a template brain that is
then applied to individual people, which neglects the potential for variability in regional
borders and brain organization. Hier, we show that using an individualized approach to
region definition results in more valid area definitions and more conservative estimates of
brain network dysfunction in people with psychosis, indicating that at least some of the group
differences reported in the extant literature may be due to differences in regional definitions
rather than a consequence of the illness itself.
30
31
32
33
34
35
36
37
38
39
40
41
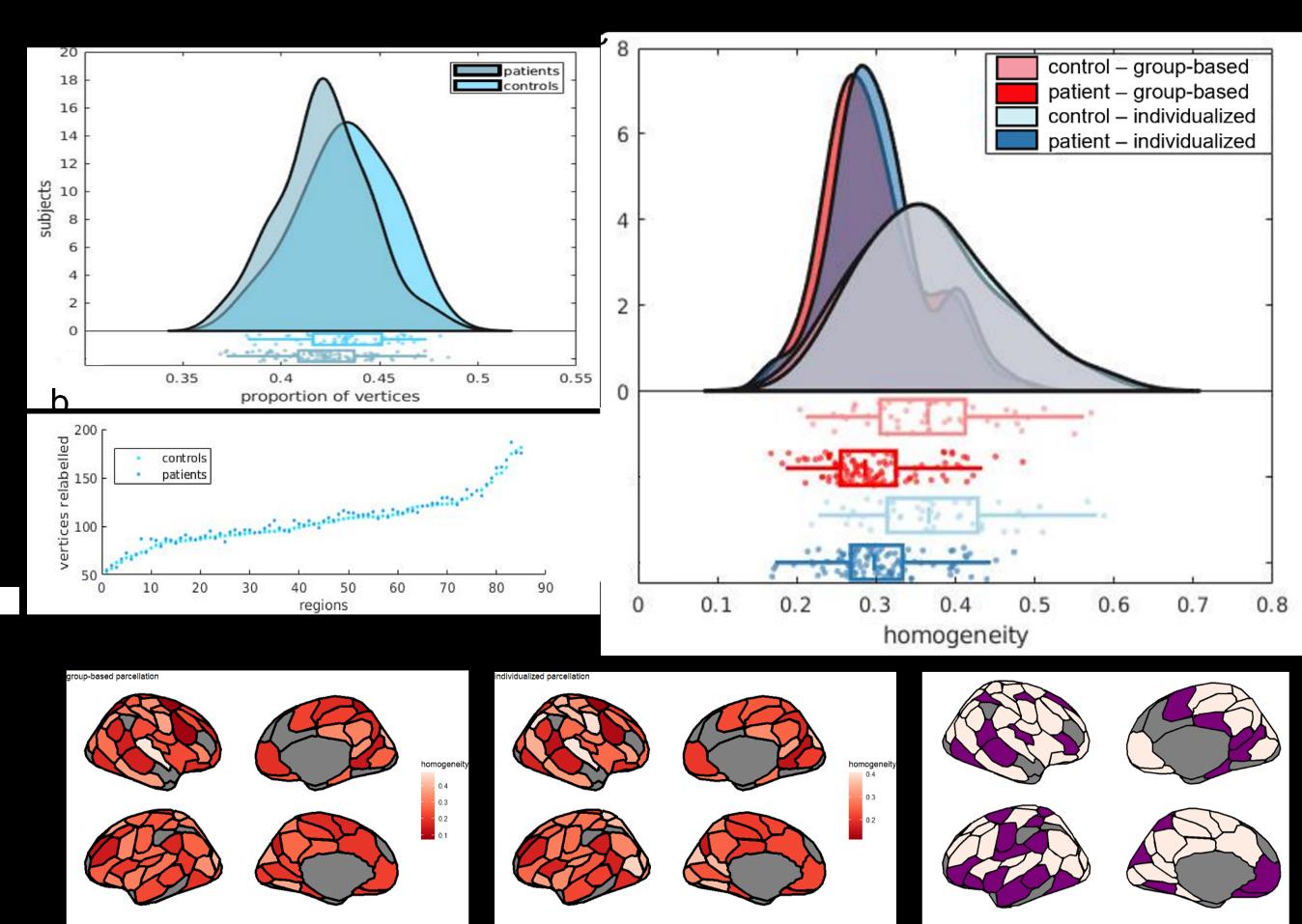
42
43
44
45
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
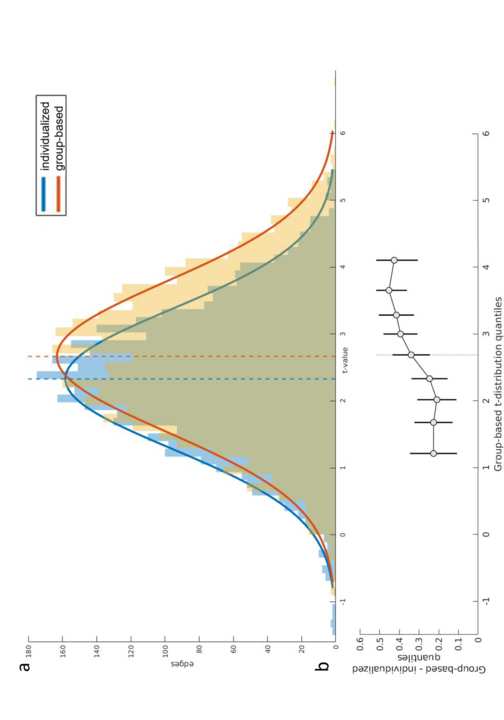
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
/
T
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
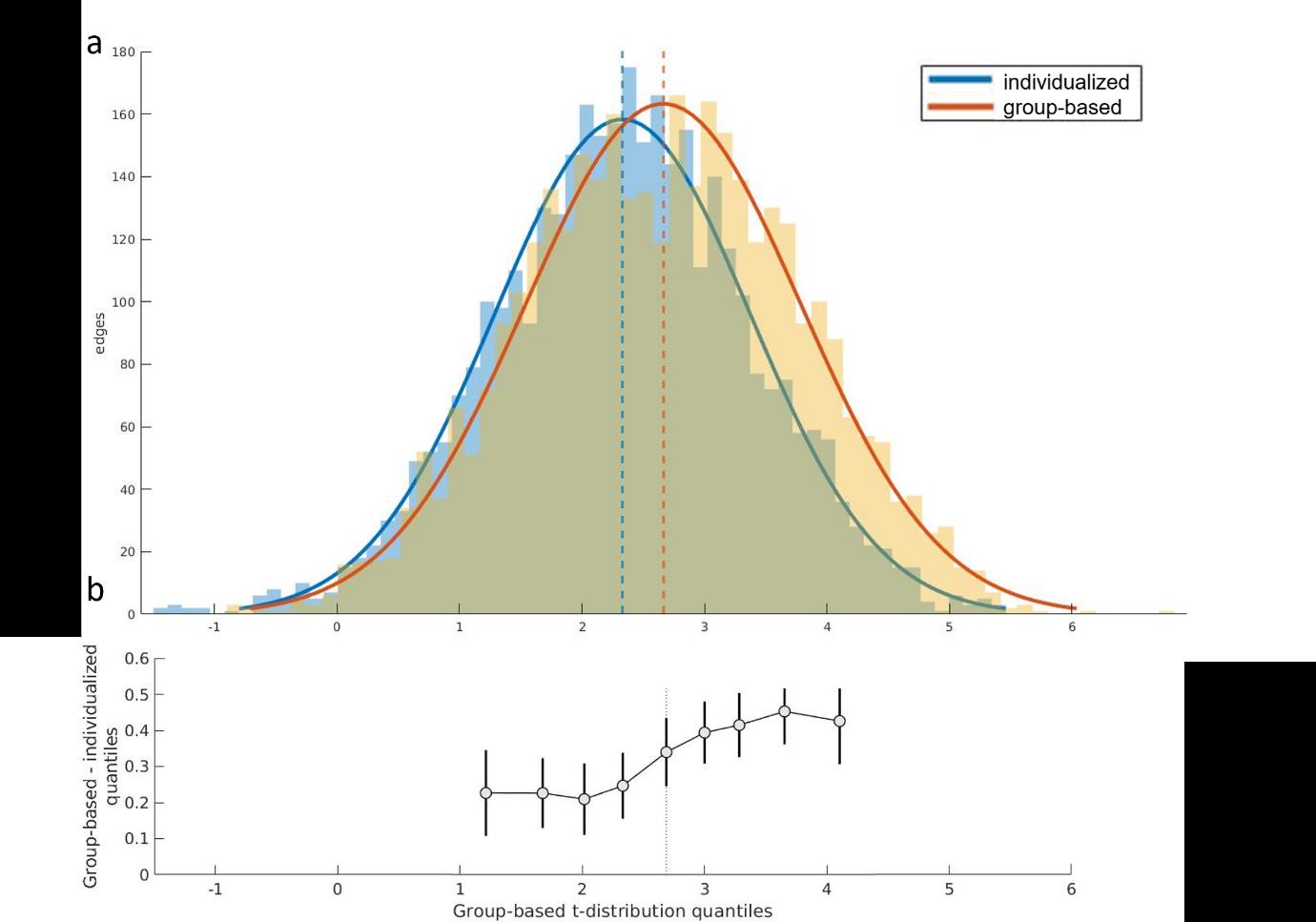
F
/
D
Ö
ich
/
/
T
.
/
1
0
1
1
6
2
N
e
N
_
A
_
0
0
3
2
9
2
1
4
2
0
1
4
N
e
N
_
A
_
0
0
3
2
9
P
D
T
/
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
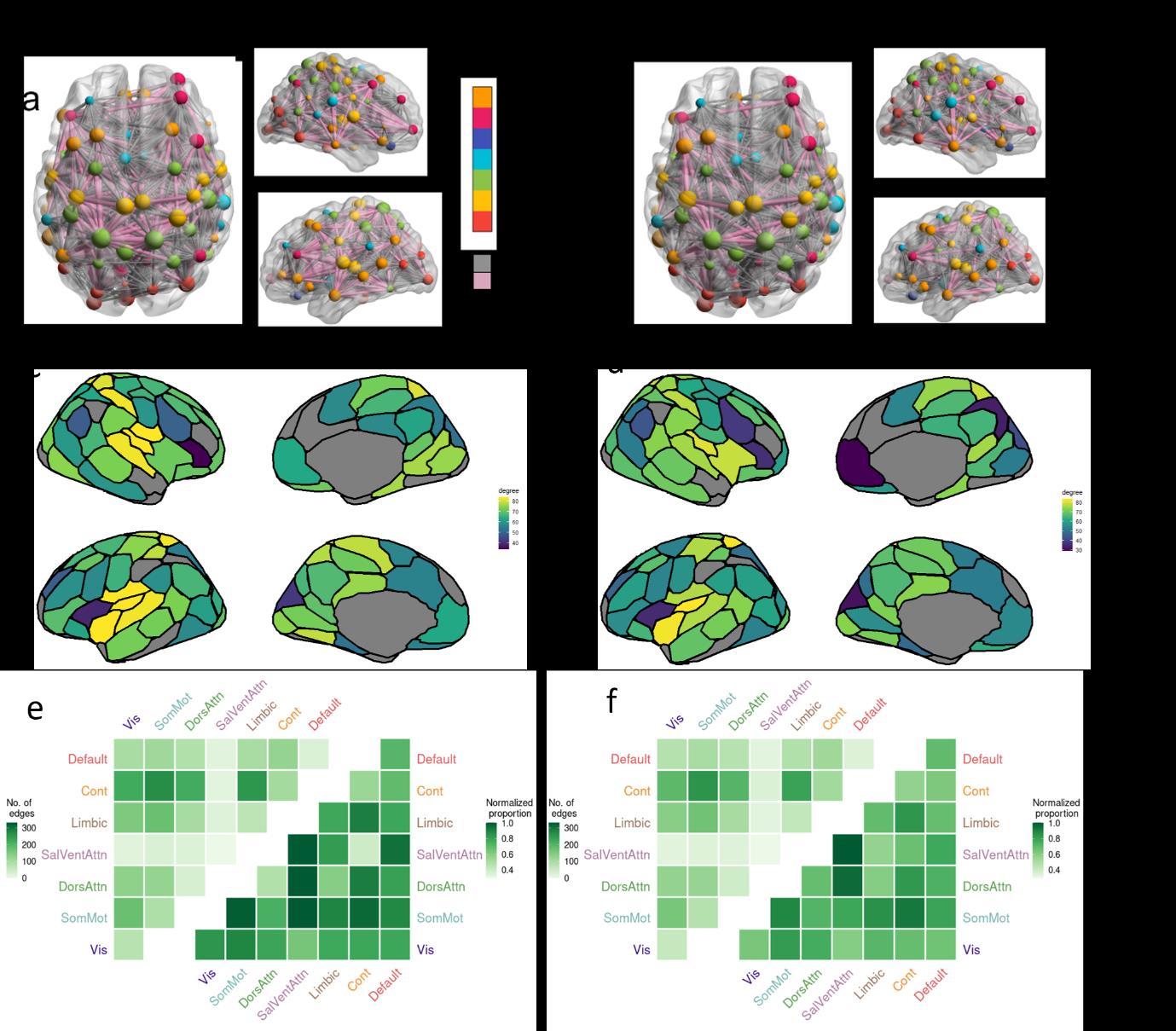
B
e
R
2
0
2
3
46
47
48
49
50
51
52
53
54
55
56
57
58
59
60
61
62
63
64
65
66
67
68
Einführung
Psychosis is a neuropsychiatric condition that has long been thought to arise from
aberrant neural connectivity, or dysconnectivity, between neuronal populations (Andreasen et
al., 1998; Baker et al., 2019; Fornito et al., 2012; Nogovitsyn et al., 2022). Solch
dysconnectivity is often studied using a network-based approach (Fornito et al., 2016), mit
the brains of individuals being modelled as a collection of nodes, representing discrete brain
Regionen, connected by edges, representing inter-regional structural connectivity or functional
coupling (FC). This approach has revealed extensive FC disruptions in psychosis patients,
which are often characterized by a global decrease in FC upon which is superimposed more
network-specific increases and decreases (Argyelan et al., 2014; Baker et al., 2019; Chopra et
al., 2021; Fornito et al., 2012; Hummer et al., 2020; T. Li et al., 2017; Narr & Leaver, 2015;
Nogovitsyn et al., 2022; Tu et al., 2013). Jedoch, the reported findings have been
inconsistent, with reports of increased and decreased FC sometimes found within the same
network in different samples (Lynall et al., 2010; Moran et al., 2013; Whitfield-Gabrieli et
al., 2009; Woodward et al., 2011).
Some of these inconsistencies may be explained by methodological differences in
defining the nodes (brain regions of interest – ROIs) of the constructed brain networks, welche
is a fundamental step in network analysis that could affect the validity and interpretation of
subsequent results (Fornito et al., 2010, 2016; Zalesky, Fornito, Harding, et al., 2010). Jede
node should ideally represent a functionally specialized area with homogenous activity
(Eickhoff, Polizist, et al., 2018; Eickhoff, Yeo, et al., 2018), but there is no consensus on
the optimal way of parcellating the brain, meaning that investigators must rely on various
heuristic methods (Eickhoff, Polizist, et al., 2018; Eickhoff, Yeo, et al., 2018).
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
D
Ö
ich
/
.
T
/
/
1
0
1
1
6
2
N
e
N
_
A
_
0
0
3
2
9
2
1
4
2
0
1
4
N
e
N
_
A
_
0
0
3
2
9
P
D
T
/
.
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
69
70
71
72
73
74
75
76
77
78
79
80
81
82
83
84
85
86
87
88
89
90
91
92
93
The vast majority of studies in patients with psychosis have used a one-size-fits-all,
group-based approach in defining distinct ROIs. A parcellation using this approach is often
defined in a standardized coordinate space based on a sample average and then mapped to
individual participants via a spatial normalization procedure (Eickhoff, Yeo, et al., 2018).
This approach fails to consider interindividual variability in functional and anatomical brain
organization (Amunts et al., 2005; Mueller et al., 2013). Investigation of such variability with
resting-state fMRI (rsfMRI) has shown that, although most cortical areas can indeed be
robustly identified in every individual, their sizes and shapes vary across the population,
especially when using more fine-grained parcellation methods (Gordon et al., 2017).
Außerdem, the topographical locations of specific areas tend to shift between individuals,
sometimes across anatomical landmarks such as sulci and gyri (Gordon et al., 2017), welche
are often used as reference points in many standard parcellations (Fornito et al., 2016).
To better accommodate this individual variability, approaches have been developed to
derive individualized parcellations at either the level of canonical functional networks (S. Li
et al., 2016; Yeo et al., 2011) or cortical regions (Gordon et al., 2017; Kong et al., 2021).
These approaches have revealed that individual variability can considerably impact network
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
D
Ö
ich
/
/
.
T
/
1
0
1
1
6
2
N
e
N
_
A
_
0
0
3
2
9
2
1
4
2
0
1
4
N
e
N
_
A
_
0
0
3
2
9
P
D
T
/
.
Analysen. Zum Beispiel, regions assigned to one network in individual parcellations are often
assigned to a different network in the group average (Bijsterbosch et al., 2018), which could
impact FC analysis. The use of individually-tailored parcellations yields more functionally
homogeneous regions (Chong et al., 2017; Kong et al., 2021), and can improve predictions of
behaviour from FC (Kong et al., 2019). In der Tat, in healthy samples, individual differences in
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
the locations of functional regions, as represented by individualized parcellation, affect
predictions of fluid intelligence (Kong et al., 2019), life satisfaction (Bijsterbosch et al.,
2018), participant sex (Salehi et al., 2018), and performance in reading and working memory
tasks (Kong et al., 2021). Darüber hinaus, some estimates indicate that up to 62% of variance in
94
95
96
97
98
99
network edge strength (d.h., FC values) can be explained by the spatial variability of defined
Regionen (Bijsterbosch et al., 2018). These findings suggest that clinically important
relationships may be masked when using a group-based parcellation. Andererseits, diese
approaches present several challenges, such as establishing a correspondence between similar
regions in different people and accounting for differences in region size.
A particularly salient point in clinical studies, such as those of schizophrenia, is that
100
standard brain atlases have been derived from healthy participants, which may not adequately
101
capture the characteristic properties in the brain organization of patients (Glasser et al., 2016;
102
Schaefer et al., 2018). Patient-specific individual variability in functional organization can
103
influence the results of brain network analyses. In der Tat, one study has found that slight
104
displacements of a seed region in the thalamus can lead to significant differences in disorder-
105
related dysconnectivity (Welsh et al., 2010), emphasizing the importance of a valid and
106
consistent node definition.
107
One strategy to develop individualized parcellations is to adjust the borders of a
108
group-based template for each individual participant according to pre-defined functional
109
Kriterien. For instance, Chong et al. (Chong et al., 2017) developed a Bayesian algorithm
110
(called Group Prior Individualized Parcellation – GPIP) that uses a group-based template as a
111
prior to find an optimal corresponding parcellation on individual brains using individual FC
112
Daten. The group-based prior ensures that the same regions are mapped in each individual,
113
while updates to the individualized prior account for variability in the shape and size of each
114
parcellated region. Chong et. al. (Chong et al., 2017) have shown that this method yields
115
parcellated regions with increased intra-regional functional homogeneity and reduced
116
variance in connectivity strength between individuals (Chong et al., 2017). Hier, we used this
117
approach to compare FC disruptions observed in people with early psychosis using analyses
118
that rely on either a group-based or individualized parcellation. The parcellation algorithm
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
D
Ö
ich
/
T
/
/
.
1
0
1
1
6
2
N
e
N
_
A
_
0
0
3
2
9
2
1
4
2
0
1
4
N
e
N
_
A
_
0
0
3
2
9
P
D
.
/
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
119
(Chong et al., 2017) allowed us to match all brain regions across participants while
120
accounting for individual variability. Our analyses were conducted using the high-quality,
121
open-access data provided by the Human Connectome Project – Early Psychosis (Glasser et
122
al., 2013; HCP Early Psychosis 1.1 Data Release: Reference Manual HUMAN Connectome
123
PROJECT for Early Psychosis, 2021) (HCP-EP) resource. We tested two competing
124
hypotheses of how individual variability contributes to apparent FC disruptions in psychosis.
125
Under one hypothesis, a failure to consider individual variability may lead to erroneous
126
regional parcellations, adding noise to the analyses and reducing statistical power for
127
detecting valid group differences. In this case, we expect to see fewer differences between
128
patients and controls when using the group-based parcellation compared to individualized
129
parcellation. Alternativ, FC differences between groups may be largely driven by
130
variations in the underlying organization of each individual’s brain, rather than reflecting
131
specific differences in FC. In this case, we expect to see more differences using the group-
132
based parcellation.
133
134
135
136
137
138
139
140
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
D
Ö
ich
/
/
T
/
.
1
0
1
1
6
2
N
e
N
_
A
_
0
0
3
2
9
2
1
4
2
0
1
4
N
e
N
_
A
_
0
0
3
2
9
P
D
/
.
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
141
142
Ergebnisse
Hier, we present results obtained using group-level cortical parcellations provided by
143
Schaefer et al. (Schaefer et al., 2018) as the basis for our analysis, focusing on the 100-region
144
parcellation (s100). To ensure that our results are robust to the number of regions, Wir
145
repeated our analysis using the 200-region variant (s200) and after applying Global Signal
146
Regression (GSR). Results obtained using the s200 atlas, and results for both atlases after
147
GSR, can be found in the Supplementary Materials and are largely consistent with the
148
primary results reported in the following sections.
149
Spatial and functional properties of group-based vs individualized parcellation
150
Figur 1 shows examples of individualized parcellations generated for three
151
individuals compared with the original group-based s100 atlas. The individualized
152
parcellation algorithm preserved the same regions for every individual but shifted their
153
borders and changed their shapes and sizes to accommodate for individualized variations in
154
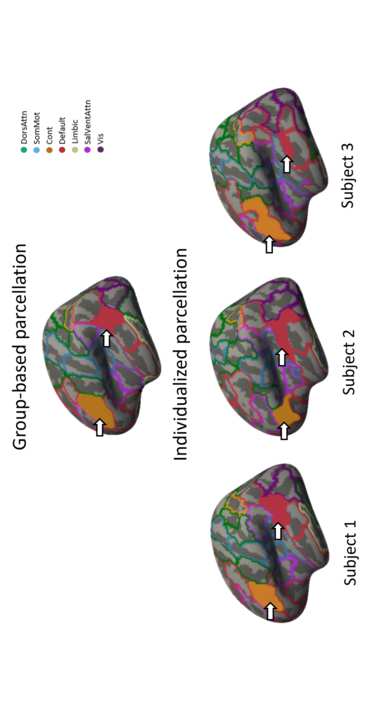
brain organization. In der Tat, on average, 42.56% (𝑆𝐷 = 2.37) of vertices were reallocated to a
155
different region as a result of the individualized parcellation algorithm, highlighting the
156
considerable variability of cortical functional organization between individuals. Figure 2a
157
shows the proportion of vertices that were relabelled in controls 𝑀(𝑆𝐷) = 43.28% (2.34)
158
and in patients 𝑀(𝑆𝐷) = 42.20% (2.31). The difference between the two groups was small
159
but statistically significant, following permutation testing ( 𝑝 = 0.004, 𝐻𝑒𝑑𝑔𝑒𝑠′𝑠 𝑔 =
160
0.465). Jedoch, at a regional level (figure 2b), no parcel showed significant differences in
161
the number of vertices relabelled between patients and controls (d.h., all 𝑝𝐹𝐷𝑅 >
162
0.05, corrected with the Benjamini and Hochberg method).
163
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
D
Ö
ich
/
T
/
/
.
1
0
1
1
6
2
N
e
N
_
A
_
0
0
3
2
9
2
1
4
2
0
1
4
N
e
N
_
A
_
0
0
3
2
9
P
D
.
/
T
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
D
Ö
ich
/
.
/
T
/
1
0
1
1
6
2
N
e
N
_
A
_
0
0
3
2
9
2
1
4
2
0
1
4
N
e
N
_
A
_
0
0
3
2
9
P
D
.
T
/
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
164
165
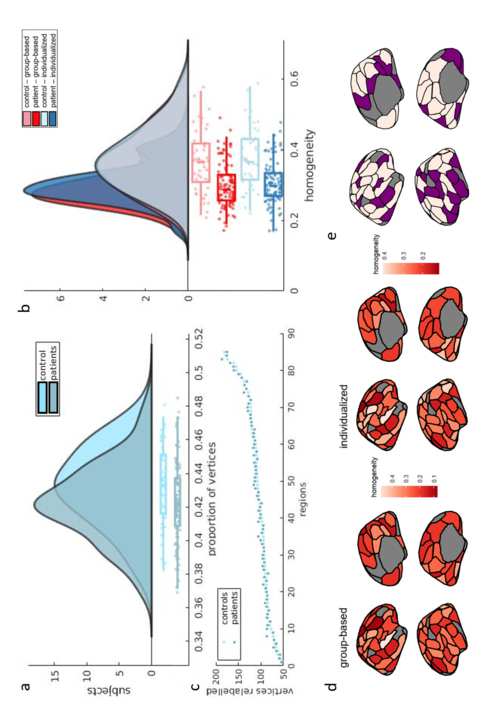
Figur 1. Differences in parcel boundaries between group-based and individualized
166
parcellation. The images show different parcellations overlayed on the inflated fsaverage5
167
template surface of the left hemisphere, mit 20,484 Eckpunkte. The top image shows the
168
group-based parcellation, which was used as a starting point for the individualized
169
parcellation algorithm. Colors correspond to the seven canonical functional networks that are
170
used to group parcels in the atlas (Yeo et al., 2011). The bottom three images show
171
individualized parcellations for three different subjects after 20 iterations of the GPIP
172
Algorithmus. The region shaded in orange corresponds to region 1 in the lateral prefrontal
173
cortex of the control network for all parcellations. The region shaded in red corresponds to
174
region 1 in the parietal lobe of the default mode network. The same regions are present in all
175
individuals, but their locations, sizes and shapes show considerable variability. DorsAttn –
176
dorsal attention network; SomMot – somatomotor network; Cont – control network; Default
177
– default mode network; Limbic – limbic network; SalVentAttn – salience/ventral attention
178
Netzwerk; Vis – visual network.
179
We next compared the average functional homogeneity of the group-based and individualized
180
parcellations. Functional homogeneity was measured out of sample, on functional scans from
181
run 2 with parcellations generated for scans from run 1. In controls, the mean homogeneity
182
War 0.364 (𝑆𝐷 = 0.09), Und 0.372 (𝑆𝐷 = 0.08) for the group-based and individualized
183
parcellations, jeweils. In patients, the mean homogeneity was 0.297 (𝑆𝐷 = 0.06) Und
184
0.304 (𝑆𝐷 = 0.06) for the group-based and individualized parcellations, jeweils (figure
185
2C). A two-way mixed ANOVA revealed that mean homogeneity was higher for the
186
individualized parcellation (𝐹(149) = 54.81, 𝑝 < 0.0001) and higher in controls compared 187 to patients (𝐹(149) = 30.91, 𝑝 < 0.0001), with no interaction between parcellation type and 188 diagnostic group (𝐹(149) = 0, 𝑝 = 0.898). Post-hoc analysis showed that individualized 189 parcellation resulted in greater homogeneity scores in patients (𝑡(103) = 5.64, 𝑝 < 0.0001) 190 and controls (𝑡(46) = 2.90, 𝑝 = 0.006). When comparing homogeneity scores for individual 191 parcels (figure 2d, e), 55 out of 85 regions showed significant differences in homogeneity 192 between parcellation approaches (i.e., 𝑝𝐹𝐷𝑅 < 0.05, corrected with the Benjamini and 193 Hochberg method). Moreover, both methods showed high reliability of homogeneity 194 estimates, as measured with the intraclass correlation coefficient (McGraw & Wong, 1996) l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . t / / e d u n e n a r t i c e - p d l f / d o i / . / t / 1 0 1 1 6 2 n e n _ a _ 0 0 3 2 9 2 1 4 2 0 1 4 n e n _ a _ 0 0 3 2 9 p d t / . 195 (𝑟𝑔𝑟𝑜𝑢𝑝−𝑏𝑎𝑠𝑒𝑑 = 0.842, 𝑝 < 0.0001 𝑎𝑛𝑑 𝑟𝑖𝑛𝑑𝑖𝑣𝑖𝑑𝑢𝑎𝑙𝑖𝑧𝑒𝑑 = 0.862, 𝑝 < 0.0001). To quantify 196 functional distinctions between parcels, we computed the mean Pearson’s correlation of 197 activity between each pair of vertices that were not allocated to the same region. We found 198 that the individualized parcellation (𝑀𝑐𝑜𝑟𝑟(𝑆𝐷) = 0.100 (0.066)) delineates parcels that are 199 slightly more functionally distinct than those in the group-based parcellation (𝑀𝑐𝑜𝑟𝑟(𝑆𝐷) = f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 200 0.102 (0.066)). Although small, the difference was statically significant (𝑡(165) = 201 14.0, 𝑝 < 0.001). 202 Homogeneity scores results were similar for s200 atlas with and without GSR 203 (Supplementary Materials figures 2 and 3). For the s100 atlas with GSR, differences in 204 homogeneity between groups and parcellation approach were similar to the main results. 205 However, there was a significant interaction effect between parcellation type and diagnosis 206 (𝐹(148) = 4.68, 𝑝 = 0.032) (See Supplementary Materials figure 1), such that homogeneity 207 scores in patients were more impacted by individualized parcellation than in controls. This 208 result suggests that, at this particular resolution, parcellation type may differentially affect FC 209 estimates in patients and controls only following the application of GSR. The reasons for this 210 sensitivity to parcellation scale and GSR are unclear. l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . / t / e d u n e n a r t i c e - p d l f / d o i / . / / t 1 0 1 1 6 2 n e n _ a _ 0 0 3 2 9 2 1 4 2 0 1 4 n e n _ a _ 0 0 3 2 9 p d t . / f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 211 212 Figure 2 – Spatial and functional properties of group-based vs individualized 213 parcellations. Panel a shows the proportion of vertices relabelled by the individualized 214 parcellations for controls (𝑀(𝑆𝐷) = 0.433(0.023)) and for patients (𝑀(𝑆𝐷) = 215 0.422(0.023)). Panel b shows the average number of vertices relabelled in every parcel for 216 patients and controls. Panel c shows the distribution of homogeneity scores per subject. 217 Controls produced more homogenous parcels in both individualized (𝑀(𝑆𝐷) = 218 0.372(0.08)) and group-based parcellations (𝑀(𝑆𝐷) = 0.364(0.09)) than patients 219 (𝑖𝑛𝑑𝑖𝑣𝑖𝑑𝑢𝑎𝑙𝑖𝑧𝑒𝑑 𝑀(𝑆𝐷) = 0.304(0.06)), (𝑔𝑟𝑜𝑢𝑝 − 𝑏𝑎𝑠𝑒𝑑 𝑀(𝑆𝐷) = 0.297(0.06)). Panel 220 d shows homogeneity scores for every parcel for group-based and individualized parcellation. 221 Light colored parcels in e represent parcels showing significant difference in homogeneity 222 scores, between parcellation approaches, for 𝑝𝐹𝐷𝑅 < 0.05. Homogeneity is displayed in 223 inflated surfaces with the group-based parcellation. 224 225 Unthresholded edge-level group differences in FC 226 Following exclusion of regions with poor signal (see Methods) the final networks 227 examined comprised 85 regions. The FC matrices resulting from both parcellation methods 228 were positively correlated, with correlations ranging between 0.679 and 0.898 (median = 229 0.794) across participants (Supplementary Materials figure 4a), indicating that the results 230 obtained with individualized and group parcellations are generally similar, although far from 231 identical. 232 Figure 3a shows the distribution of 𝑡-statistics across edges, comparing FC between 233 patients and controls estimated using either the group-based or individualized parcellation. 234 Both distributions have predominantly positive values, consistent with evidence of 235 widespread hypoconnectivity in patients compared to controls. The distribution for the group- 236 based approach is shifted further to the right, indicating that larger group differences are 237 detected with this method, on average. The difference in the means of the distributions was 238 statistically significant, as calculated with a Wilcoxon signed-rank test (𝑍 = 24.053 𝑝 < 239 0.0001). Figure 4 of the Supplementary Materials shows that most FC edges were positively 240 valued; as such, the significant FC reductions observed in patients result from patients having 241 lower positive FC rather than patients having stronger negative FC. Given the higher l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . / / t e d u n e n a r t i c e - p d l f / d o i / . / t / 1 0 1 1 6 2 n e n _ a _ 0 0 3 2 9 2 1 4 2 0 1 4 n e n _ a _ 0 0 3 2 9 p d t / . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 242 functional homogeneity of the individualized parcellation, this result suggests that the group- 243 based parcellation overstates FC differences between patients and controls. Similar results 244 were obtained when looking at the effect size of the differences in edge strength between 245 patients and controls (Supplementary Materials figure 4), with the group-based parcellation 246 yielding higher effect size estimates than individualized parcellation, on average (𝑝 < 247 0.0001). 248 The 𝑡-matrices obtained using the group-based and individualized parcellations were 249 positively correlated (𝑟 = 0.76, 𝑝 < 0.0001), suggesting that the two approaches show 250 largely similar between-group FC differences. The effects of parcellation type were 251 consistent across the full extent of the 𝑡-distributions, as indicated by the shift function, 252 which compares differences between distributions at each decile. This analysis showed a 253 significantly higher value in every decile of the group-based parcellation, when compared to 254 the individualized parcellation, with the 95% CI never crossing zero (figure 3b). There was, 255 however, a more pronounced effect of parcellation type on edges associated with larger case- 256 control differences in FC relative to those with smaller case-control differences, as can be 257 seen by the greater shift observed in the right tail of the distribution relative to the left (figure 258 3b). This result implies that variations in parcellation type are more likely to influence the 259 edges that are significantly different between patients and controls. Furthermore, results 260 obtained using the s200 parcellations are in agreement with results obtained from the s100 261 parcellation (see Supplementary Materials figure 2). Following GSR, at both parcellation 262 scales, the mean t-values were similar, but the t-distribution for the individualized 263 parcellation was narrower than for the group-based parcellation. The shift function showed 264 that significant differences between parcellation approaches were mainly for edges with 265 positive t-values (see Supplementary Materials figures 1 and 3). l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . / t / e d u n e n a r t i c e - p d l f / d o i / / / . t 1 0 1 1 6 2 n e n _ a _ 0 0 3 2 9 2 1 4 2 0 1 4 n e n _ a _ 0 0 3 2 9 p d t / . f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . / t / e d u n e n a r t i c e - p d l 266 267 Figure 3 – Edge-specific case-control differences in FC depend on parcellation type. a 268 Distributions of 𝑡-values quantifying FC differences between patients and controls at each 269 edge and for each parcellation type. A positive t-value indicates a greater FC value in controls 270 than in patients. For reference, a p-value = 0.05 corresponds to a t-value = 1.65 uncorrected, 271 and t = 4.31 Bonferroni corrected. b Shift function (Rousselet et al., 2017) for the two t- 272 distributions. Each circle represents the difference between the borders of each decile of both 273 distributions as a function of the deciles in the group-based distribution. The bars represent 274 the 95% boot-strap confidence interval associated with the difference. 275 Thresholded edge-level group differences in FC 276 We used the Network Based Statistic (NBS) for inference on the edge-specific 𝑡-statistics 277 (Zalesky, Fornito, & Bullmore, 2010). The NBS identified a single connected component 278 with significant FC differences between patients and controls using both the group-based f / d o i / . / / t 1 0 1 1 6 2 n e n _ a _ 0 0 3 2 9 2 1 4 2 0 1 4 n e n _ a _ 0 0 3 2 9 p d . / t f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 279 (𝑝 < 0.0001) and individualized parcellations (𝑝 < 0.0001), for all primary test statistics 280 thresholds tested. Out of 3,570 possible connections, for a primary threshold equivalent to a 281 p-value = 0.05, the group-based and individualized parcellations resulted in components 282 comprising 2,877 edges and 2,672 edges respectively (figure 4a-b). Thus, the group-based 283 approach implicated approximately 7.7% more dysconnected edges. The binary edge 284 matrices defining these components were moderately correlated (𝑟𝑝ℎ𝑖 = 0.548, 𝑝 < 0.0001) 285 and both components had a total of 571 edges that differed from each other. There was also 286 some variation in the regional affiliation of the edges. For example, figure 4c-d show that the 287 insula has a high dysconnectivity degree in both group-based and individualized 288 parcellations, but that the former approach implicates more insula sub-regions. Furthermore, 289 the right medial prefrontal cortex shows a low degree in the individualized parcellation but 290 not in the group-based parcellation. The NBS was repeated with a primary test statistics 291 threshold equivalent to p-values = 0.01 and 0.001. For 𝑝 = 0.01, the component for 292 individualized parcellation comprised 1,786 edges and for group-based parcellation, 2,120. 293 For 𝑝 = 0.001, the component for individualized parcellation comprised 775 edges and for 294 group-based, 1,257 edges. Note that for all edges in these NBS networks, patients showed 295 reduced FC compared to controls. 296 297 Effects of variations in parcel size 298 A challenge of using individualized parcellations is that the ROIs can vary in size 299 across individuals, which may bias estimates of FC differences between groups. We therefore 300 examined changes in parcel size resulting from the individualization algorithm, as quantified 301 by the number of vertices in each parcel. On average, parcels changed by 50.7 (SD = 45.2) 302 vertices across patients and 52.0 (SD = 45.3) across controls, with no significant difference l D o w n o a d e d f r o m h t t p : / / d i r e c t . m i t . / / t e d u n e n a r t i c e - p d l f / d o i / / t / . 1 0 1 1 6 2 n e n _ a _ 0 0 3 2 9 2 1 4 2 0 1 4 n e n _ a _ 0 0 3 2 9 p d / . t f b y g u e s t t o n 0 7 S e p e m b e r 2 0 2 3 303 between the two groups, according to permutation testing (𝑝 = 0.104) (Supplementary 304 Materials figure 8a). There was also no significant difference in size difference between 305 patients and controls for any of the parcels, when corrected for multiple comparisons 306 following permutation statistics (i.e., all 𝑝𝐹𝐷𝑅 > 0.05). Four parcels had different sizes
307
between patients and controls, without correction for multiple comparisons (visual network
308
parcel 9 of the left hemisphere, 𝑝 = 0.023; somatomotor network parcel 1 of the left
309
hemisphere, 𝑝 = 0.026; limbic network parcel 1 in the orbital frontal cortex of the left
310
hemisphere, 𝑝 = 0.039, limbic network parcel 1 in the orbital frontal cortex of the right
311
hemisphere, 𝑝 = 0.048). We next correlated the differences in parcel size in individualized
312
parcellation between patients and controls with differences in node degree within the NBS
313
network and mean edge dysconnectivity, given by the mean 𝑡-value of edges attached to each
314
node for the case-control comparison (Supplementary Materials figure 8b-c). Neither
315
correlation was significant (𝑟 = 0.148, 𝑝𝑠𝑝𝑖𝑛 = 0.104 and 𝑟 = 0.133, 𝑝𝑠𝑝𝑖𝑛 = 0.127,
316
jeweils), suggesting that parcel size did not impact FC differences between patients and
317
controls in the individualized parcellation.
318
Network-level group differences in FC
319
Having demonstrated that the choice of a parcellation strategy can influence both
320
edge- and region-level inferences about FC disruptions in psychosis, we next examined
321
whether parcellation type affects the specific networks that are considered to be
322
dysfunctional. We therefore examined the proportion of edges within the NBS network that
323
fell within and between each of 7 canonical functional networks (Thomas Yeo et al., 2011).
324
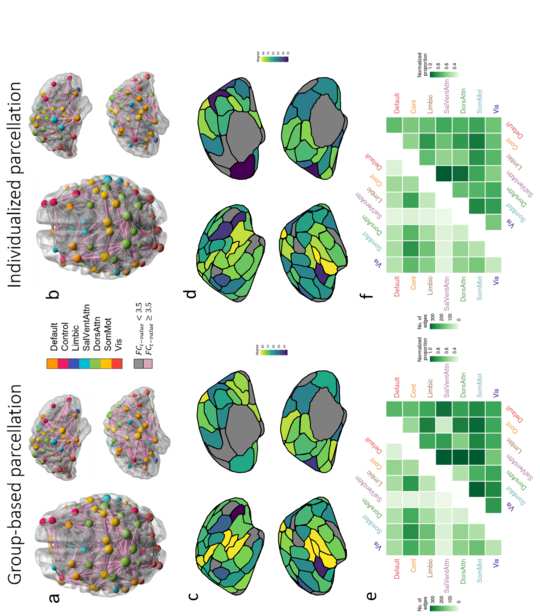
Considering the raw number of affected edges across both parcellation approaches, Die
325
control network was the most impacted in patients with psychosis, with over 1,100
326
dysconnected edges, particularly those linking the control and somatomotor networks (figure
327
4e-f). By comparison, normalized counts, which is adjusted for the total number of possible
l
D
Ö
w
N
Ö
A
D
e
D
F
R
Ö
M
H
T
T
P
:
/
/
D
ich
R
e
C
T
.
M
ich
T
.
T
/
/
e
D
u
N
e
N
A
R
T
ich
C
e
–
P
D
l
F
/
D
Ö
ich
/
/
T
.
/
1
0
1
1
6
2
N
e
N
_
A
_
0
0
3
2
9
2
1
4
2
0
1
4
N
e
N
_
A
_
0
0
3
2
9
P
D
.
T
/
F
B
j
G
u
e
S
T
T
Ö
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
328
edges within or between pairs of networks, suggested a more equal and widespread
329
distribution of FC disruptions across networks. Both the raw count (𝑟 = 0.983, 𝑝 <
330
0.0001 ) and normalized matrices (𝑟 = 0.802, 𝑝 < 0.0001) were strongly correlated across
331
the two parcellation methods. These findings indicate that while parcellation method can
332
influence the specific edges that are identified as dysconnected, these edges generally fall
333
within or between the same canonical networks.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
t
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
334
335
Figure 4 – Edge-level regional and network-level case-control FC differences according
336
to parcellation type. Panels a and b show the specific edges comprising the NBS
337
components obtained with the group-based and individualized parcellations, respectively,
338
with nodes colored according to network affiliation and sized by degree. Edges are sized by
339
strength of dysconnectivity. Edges associated with a t-value < 3.5 are represented by grey
340
lines and those associated with a t-value ≥ 3.5 are represented in pink. The images were
341
created using the software BrainNet Viewer (Xia et al., 2013). Panels a, c, and e result from
342
the group-based parcellation. Panels c and d show the degree of each region in the NBS
343
component for the group and individualized parcellations, respectively. The left most triangle
344
of each matrix in panels e and f shows the total number of NBS component edges (raw
345
counts) falling within and between seven canonical networks. The right most triangles show
346
the same data normalized for network size, i.e. the total number of possible connection within
347
or between networks (normalized counts). DorsAttn – dorsal attention network; SomMot –
348
somatomotor network; Cont – control network; Default – default mode network; Limbic –
349
limbic network; SalVentAttn – salience/ventral attention network; Vis – visual network.
350
351
352
353
354
355
356
357
358
359
360
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
361
362
Discussion
Several studies have reported functional brain dysconnectivity in psychosis. A
363
fundamental step in such analyses involves defining a priori ROIs to serve as nodes in the
364
network analysis, which are typically derived from standard parcellation atlases generated
365
from a population or group average template. Here, we asked whether the failure of such an
366
approach to account for individual differences in brain functional organization can bias
367
estimates of case-control differences in FC. Standard methods could either result in an under-
368
estimation of the extent of network dysfunction (due to noisy FC estimation caused by
369
inaccurate ROI delineations) or an inflated estimate of the dysfunction (due to FC differences
370
being attributable to ROI misalignment), compared to when accounting for individual
371
differences in functional organization of the brain. Our findings indicate that group-based
372
parcellations might inflate estimates of FC differences in psychosis, especially at the edge-
373
level. Moreover, the use of individualized parcellations, while yielding a generally consistent
374
pattern of findings, leads to some different conclusions about the specific edges and regions
375
most affected by the disorder, although inferences at the network level were robust to
376
parcellation variations. Together, our findings suggest that the use of individualized
377
parcellations can impact findings of brain dysconnectivity in psychosis and, by extension,
378
other disorders.
379
Individualized parcellations yield more functionally homogeneous regions
380
The individualized parcellations resulted in nearly half (over 40%) of vertices being
381
assigned to regions that differed from the group-based atlas, as per prior work (Chong et al.,
382
2017). This finding reiterates how group-based parcellations can result in a substantial
383
misspecification of regional borders in individuals and highlights the high degree of variance
384
present in the topographical organization of functional areas. Despite the high percentage of
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
.
t
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
385
vertices relabelled, FC matrices generated by both parcellations were highly correlated,
386
indicating the overall FC patterns seen with group-based parcellation are maintained with the
387
individualized parcellation. Note that with GPIP, correspondence between regions is
388
determined based on similarity in FC profiles rather than spatial location. As such,
389
corresponding regions can shift their spatial location from person to person (see Figure 1).
390
The higher functional homogeneity of the individualized parcellations supports its
391
improved validity, although the increment was small (2.4%), which is consistent with past
392
reports (Kong et al., 2021; Y. Li et al., 2022), increased homogeneity was seen in the
393
majority of parcels. Regional homogeneity was also marginally (2.3%) higher in controls
394
compared to patients. This differential improvement in homogeneity was expected, as the
395
starting point for the GPIP algorithm was the Schaefer atlas (Schaefer et al., 2018), which
396
was derived from a sample of people with no psychiatric disorders. Defining an initial group
397
atlas in patients would better account for differences in cortical functional organization
398
caused by psychosis. However, it would complicate comparisons between groups because of
399
the requirement to have consistently defined nodes in both patients and controls, which is one
400
of the challenges of using individualized parcellation. The interaction effect between
401
diagnosis and parcellation approach was not significant in most cases (apart from s100 with
402
GSR). This result indicates that individualized parcellations led to a similar improvement in
403
patients and controls. Since most case-control studies use data obtained in healthy individuals
404
to establish a normative benchmark for measures acquired in patients (Chopra et al., 2021;
405
Nabulsi et al., 2020; Nogovitsyn et al., 2022), we relied on the Schaefer parcellation in our
406
analysis. Future work could develop methods to better capture variations in functional
407
organization associated with psychosis.
408
Individualized parcellations lead to more conservative estimates of case-control FC
409
differences
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
.
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
410
Widespread decreases in FC in patients with psychosis were identified using both
411
parcellation approaches, highlighting that the dominant effect of both parcellations is
412
generally similar. However, the magnitude of the differences in FC was greater in the group-
413
based parcellation compared to individualized parcellation. Notably, the shift function
414
analysis indicated that differences between the two parcellation approaches were greater for
415
edges associated with large case-control differences. These edges are precisely the ones that
416
are most likely to be declared as statistically significant following the application of some
417
thresholding procedure. Accordingly, comparison of NBS results revealed a 7.7% reduction
418
in the size of the dysfunctional component identified using the group-based parcellation.
419
Given the higher functional homogeneity, and thus validity, of the individualized
420
parcellation, these results support the hypothesis that at least part of the group differences
421
identified in past studies in psychosis samples do not reflect actual differences in inter-
422
regional FC but instead result from inaccurate ROI boundaries caused by a failure to account
423
for individual differences in functional organization. These findings imply that individualized
424
parcellations can yield different estimates of FC differences in case-control studies, especially
425
when investigating FC changes at an edge-, or node-level.
426
Parcellation type affects FC differences in edges and regions, but not networks
427
While widespread decreases in FC were apparent in patients with psychosis using both
428
parcellation methods, the specific edges affected varied considerably. The NBS components
429
of both group-based and individualized parcellations showed differences in 571 edges (i.e.,
430
19.8% of the total identified with the group-based parcellation). Examining the regions most
431
affected by quantifying the node degrees of the NBS components resulted in broadly similar
432
patterns, but there were some notable differences in location. For example, the right medial
433
frontal region accounts for 1.7% of dysconnectivity in the group-based and 2.3% in the
434
individualized parcellation. The left insula accounts for a slightly smaller percentage (6.5%)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
/
t
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
435
of dysconnectivity in the group-based than in the individualized parcellation (6.7%). These
436
findings suggest that conclusions about the specific edges and regions affected by psychosis
437
can vary depending on the parcellation method used. In contrast, inferences at the network
438
level were largely consistent across the two parcellation approaches, indicating that coarse-
439
grained localizations of FC differences are robust to this methodological choice. This could
440
be attributed to network-level inference effectively reducing the dimensionality of the
441
analysis, minimizing the nuances of more fine-grained individual variations. Therefore,
442
studies looking at group differences in FC at a coarse, network level might not be impacted
443
by the use of individualized vs group-based parcellations.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
444
445
Limitations
446
To minimize the computational cost, we used fsaverage5, a surface mesh with a
447
relatively low number of vertices. Since GPIP parameters depend on the number of vertices
448
of the mesh, future work could investigate the impact of different surface mesh resolutions
449
and whether the differences observed here apply at different mesh resolution.
450
To facilitate comparison between subjects, the individualized parcellation algorithm
451
maintains the same number of regions for every subject and uses a parcellation derived in
452
healthy individuals as a starting point. This approach may mask differences in cortical
453
organization in patients, where regions may merge or split, resulting in a different number of
454
ROIs. However, generating separate parcellations in each group complicates comparisons
455
between groups. Resolving this challenge remains an open problem for the field. Moreover,
456
we only looked at cortical regions, due to the lack of methods available for individual
457
parcellation of subcortical structures.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
458
A proportion of patients in our sample were medicated, and recent evidence has
459
shown that anti-psychotic medication can impact FC, even after only 3 months of use
460
(Chopra et al., 2021). However, given that most samples examined in past research are also
461
medicated, our sample is directly comparable to the broader literature. Similarly, the study
462
included more patients than controls and future work could benefit from a balanced sample
463
size. We also emphasize that this study is not focused on identifying the specific nature of FC
464
disturbances associated with psychosis but instead concentrates on how parcellation type
465
affects FC differences in the same patients. In this context, medication exposure was constant
466
across our main contrast of interest (parcellation type), meaning that it cannot explain the
467
differences that we focus on here. The same reasoning applies to the clinical heterogeneity of
468
the patient sample, which comprised people diagnosed with both affective and non-affective
469
psychoses. Future work could use individualized parcellations to delineate FC differences
470
more precisely between distinct patient subgroups.
471
We have focused here on how the use of individualized vs group-based parcellations
472
affects group differences in FC. A separate question concerns whether parcellation type also
473
affects correlations with behavioural or clinical variables. Several studies have shown that
474
individualized parcellations yield FC estimates that are marginally more correlated with
475
various forms of behaviour, including psychopathological ratings (Bijsterbosch et al., 2018;
476
Kong et al., 2019, 2021). A useful direction for future work could involve investigating
477
whether individualized parcellation improves prediction of clinically meaningful outcomes.
478
479
Conclusion
480
Our findings indicate that traditional reliance on group-based parcellations may inflate case-
481
control differences in FC at a fine-grained level. The use of individualized parcellations can
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
t
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
482
yield a more conservative understanding of brain network disruptions in psychotic and
483
possibly other disorders. However, it does not greatly impact case-control differences in
484
network level analyses.
485
486
487
488
489
490
491
492
493
494
495
496
497
498
499
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
t
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
500
Methods
501
Study participants
502
All data for this study were collected as part of the Human Connectome Project –
503
Early Psychosis (HCP-EP) study, which is an open-access collection aiming to generate high-
504
quality imaging data in early psychosis patients and healthy controls (HCP Early Psychosis
505
1.1 Data Release: Reference Manual HUMAN Connectome PROJECT for Early Psychosis,
506
2021). This study includes high-resolution structural and functional Magnetic Resonance
507
Image (MRI) data from 121 patients with early psychosis (74 males) and 57 healthy
508
individuals (37 males). Demographic information is provided in Table 1. Data collection by
509
HCP-EP has been approved by the Partners Healthcare Human Research Committee/IRB,
510
and comply with the regulations set forth by the Declaration of Helsinki (Lewandowski et al.,
511
2020).
512
The patient group was comprised of outpatients with psychosis, meeting criteria for
513
affective or non-affective psychosis, according to the DSM-5, who were within the first five
514
years of onset of symptoms. Patients were recruited by four clinical programs: Beth Israel
515
Deaconess Medical Center (BMH) – Massachusetts Mental Health Center (BIDMC-MMHC),
516
Prevention of and Recovery from Early Psychosis (PREP) Program; Indiana University
517
Psychotic Disorders Program, Prevention and Recovery for Early Psychosis (PARC); the
518
McLean Hospital, McLean On Track; and Massachusetts General Hospital, First Episode and
519
Early Psychosis Program (FEPP) (HCP Early Psychosis 1.1 Data Release: Reference Manual
520
HUMAN Connectome PROJECT for Early Psychosis, 2021). Imaging took place in three of
521
these sites.
522
The control group included volunteers that did not present with anxiety disorders
523
and/or psychotic disorders, had no first-degree relative with schizophrenia spectrum disorder,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
524
were not taking psychiatric medication at the time of the study, and had never been
525
hospitalized for psychiatric reasons. All participants were aged between 16 and 35 years old
526
(mean = 23, SD = ±3.9) at the time of the study (Table 1). A total of 11 subjects were
527
excluded due to poor data quality, as detailed below, leaving a final sample of 55 (36 male)
528
controls and 112 (67 male) patients.
529
Table 1. Demographic details
Age
Sex
Control
AP
NAP
24.7 (4.1)
24.2 (4.3)
22.1 (3.3)
36M; 19F
7M; 19F
60M; 26F
Antipsychotic
--
1.5 (0 – 54)
11.5 (0 – 56)
exposure (months)
NIH cognition
113.5(8.8)
108.9 (7.8)
98.2 (13.0)
PANSS total score
UI
Scan site
BMH
McLean
--
23
26
6
40.7 (12.6)
48.8 (16.7)
7
9
10
48
30
8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
t
/
.
530
AP – affective psychosis; NAP – non-affective psychosis; PANSS – Positive and Negative
531
Syndrome Scale;
IU – Indiana University; BMH – Beth Israel Deaconess Medical
532
Center; Cont – healthy controls; F – females; M – males; age is given as mean (SD) in years
533
at the time of their first interview; antipsychotic exposure is given as median (range) in
534
months at the time of their first interview; PANSS total score is given as mean (SD); NIH
535
cognition is given as the mean (SD) of cognitive composite score, unadjusted for age,
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
536
assessed by the NIH Toolbox.
537
Data Acquisition
538
The participants recruited from four locations were scanned at three sites: BMH;
539
Indiana University; and McLean Hospital, using Siemens MAGNETOM Prisma 3T scanners.
540
The acquisition parameters between the three sites were harmonized and followed the widely
541
used HCP protocol (Demro et al., 2021; HCP Early Psychosis 1.1 Data Release: Reference
542
Manual HUMAN Connectome PROJECT for Early Psychosis, 2021). The project collected
543
whole brain T1-weighted MRI (T1w), T2-weighted MRI (T2w), diffusion MRI, spin echo
544
field maps with Anterior to Posterior (AP) and Posterior to Anterior (PA) phase encoding
545
(PE) directions - and four resting-state functional MRI (rsfMRI) sessions. The current study
546
uses the T1w and T2w images, the spin echo field maps, and the first two runs of the rsfMRI
547
scans. A 32-channel head coil was used at BMH and Indiana University. A 64-channel head
548
and neck coil, with neck channels turned off was used at McLean Hospital. Real-time image
549
reconstruction and processing was performed for quality control and scans with detectable
550
problems were repeated (HCP Early Psychosis 1.1 Data Release: Reference Manual
551
HUMAN Connectome PROJECT for Early Psychosis, 2021).
552
Structural MRI acquisition parameters
553
Acquisition parameters followed HCP standards. T1w images were obtained using a
554
magnetization-prepared rapid gradient-echo (MPRAGE), with 0.8 mm isotropic spatial
555
resolution echo time (TE) = 2.22 ms, repetition time (TR) = 2400 ms, and field of view (FoV)
556
= 256 mm. T2w images were acquired following a 3D-SPACE sequence, with 0.8 mm
557
isotropic spatial resolution, TE = 563 ms, TR = 33200 ms, and FoV = 256 mm (HCP Early
558
Psychosis 1.1 Data Release: Reference Manual HUMAN Connectome PROJECT for Early
559
Psychosis, 2021).
560
Functional MRI acquisition parameters
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
561
The present study mainly utilized the first rsfMRI run (with anterior to posterior phase
562
encoding). The second run (with posterior to anterior phase encoding) was used to validate
563
the parcellation with out-of-sample analysis of within-parcel homogeneity. Scans were
564
acquired for a length of 6.5 minutes, resulting in a total of 420 volumes; the first 10 volumes
565
were removed prior to the dataset release. Images have an isotropic spatial resolution of 2
566
mm, TE = 37 ms, TR = 800 ms, and FoV = 208 mm. A multi-band acceleration factor of 8
567
was used to improve spatial and temporal resolution (HCP Early Psychosis 1.1 Data Release:
568
Reference Manual HUMAN Connectome PROJECT for Early Psychosis, 2021).
569
Structural and Functional Image Analysis
570
Raw Image Quality Control
571
All analyses were done on the MASSIVE high-performance computing facility
572
(Goscinski et al., 2014).
573
Raw structural and functional images were first visually inspected for large artefacts
574
and distortions. Images were then put through an automated quality control pipeline
575
(MRIQC) (Esteban et al., 2017) which computes 15 image quality metrics for each scan with
576
the purposes of identifying outliers warranting closer inspection. At this stage, three subjects
577
were excluded for missing or unusable structural images.
578
Head motion is a major source of noise in fMRI signals. Its effects remain present
579
even after volume realignment and can introduce systematic bias in case-control studies when
580
not strictly controlled (Parkes et al., 2018; Power et al., 2012). Head motion during the fMRI
581
scan was estimated using frame-wise displacement (FD), which is a summary measure of the
582
movement of the head from one volume to the next (Parkes et al., 2018). For each scan, FD
583
was calculated according to the method described by Jenkinson et al. (Jenkinson et al., 2002)
584
and the resulting trace was band-pass filtered and down sampled to account for the high
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
.
t
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
585
sampling rate of the multiband fMRI acquisition (Power et al., 2019). Subjects were excluded
586
if they met at least one of the following stringent exclusion criteria: scans had a mean filtered
587
FD greater than 0.25 mm; more than 20% of frames were displaced by more than 0.2 mm; or
588
any frame was displaced by more than 5 mm. These criteria have previously been shown to
589
effectively mitigate motion-related contamination in fMRI connectivity analyses (Parkes et
590
al., 2018). In total, 11 subjects (2 controls) were excluded for excessive head movement in
591
the scanner.
592
Image Preprocessing
593
T1w images were processed using FreeSurfer version 6.0.1 (Dale et al., 1999) to
594
generate cortical surface models for each participant. Surfaces were visually examined for
595
inaccuracies and distortions. The fMRI data were processed according to the Minimal
596
Preprocessing Pipeline for HCP data (Glasser et al., 2013). The pipeline adapts steps from
597
FMRIB Software Library (FSL) and FreeSurfer to account for greater spatial and temporal
598
resolution and HCP-like distortions resulting from acquisition choices such as multiband
599
acceleration (Glasser et al., 2013). Briefly, images were skull stripped by the brain extraction
600
tool (BET) (Smith, 2002) of FSL, which removes non-brain matter from the image. Skull
601
stripped T1w, T2w, and fMRI were aligned using FMRIB’s Linear Image Registration Tool
602
(FLIRT) (Jenkinson et al., 2002; Jenkinson & Smith, 2001). Spin Echo EPI field maps with
603
opposite phase encoding directions were used to estimate spatial distortion caused by
604
magnetic field inhomogeneities, with corrections applied using FSL’s “topup” (Andersson et
605
al., 2003) and FLIRT. This process was fine-tuned and optimized using FreeSurfer’s
606
BBRegister (Greve & Fischl, 2009). Furthermore, bias field correction was performed on
607
structural images to remove gradients of voxel intensity differences, following the HCP
608
pipeline (Glasser et al., 2013). The fMRI volumes were realigned to the first volume for each
609
participant using FLIRT. The fMRI data were then co-registered to their structural image, and
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
t
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
610
the structural image was non-linearly normalized into standard Montreal Neurological
611
Institute (MNI) ICBM152 space (Grabner et al., 2006) using FLIRT and FMRIB’s nonlinear
612
image registration tool (FNIRT) (Andersson et al., 2010). The resulting transform was then
613
applied to the functional data.
614
fMRI Denoising
615
The functional data were denoised using Independent Component Analysis (ICA)-
616
based X-noiseifier (FIX), which decomposes the data into spatially independent components
617
and uses machine learning to label each resulting component as either signal or noise
618
(Griffanti et al., 2014; Salimi-Khorshidi et al., 2014). The preprocessed fMRI timeseries were
619
then regressed against the estimated noise component signals and the residuals were retained
620
for further analysis. Component decomposition was performed using Multivariate
621
Exploratory Linear Optimized Decomposition into Independent Components (MELODIC)
622
(Griffanti et al., 2014; Salimi-Khorshidi et al., 2014). HCP’s training set – HCP_hp2000,
623
which includes pre-trained weights to classify independent components, was used as the
624
training set for the algorithm. A temporal high-pass filter (2000s Full Width Half Maximum)
625
was applied to remove low-frequency signal drifts, as recommended by the HCP
626
preprocessing guideline (Glasser et al., 2013). Following HCP’s guidelines (Glasser et al.,
627
2013), a lenient threshold component labelling in FIX was used (th=10), regressing out the
628
noise components while controlling for the signal components. The accuracy of the labels
629
was manually verified. The analyses were repeated after applying Global Signal Regression
630
(GSR), which removes widespread signal fluctuations associated with respiratory variations
631
(Aquino et al., 2020; Power et al., 2017) (see Supplemental Material).
632
Surface Registration
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
633
The processed images in MNI volume space were resampled to each individual’s
634
cortical surface, as generated by FreeSurfer, and then registered to the fsaverage5 template
635
using a surface-based registration algorithm (Dale et al., 1999; Fischl, 2012). fsaverage5 is a
636
standard template generated by FreeSurfer, the resulting surface mesh comprises a total of
637
20,484 vertices.
638
Parcellations
639
We used group parcellations provided by Schaefer et al. (Schaefer et al., 2018) as the
640
basis for our analysis, as this parcellation is widely used and has shown superior functional
641
homogeneity compared to other leading approaches (Schaefer et al., 2018). Our study
642
focused on the 100-region parcellation, organized into 7 networks (s100) but we repeated the
643
analyses using the 200-region variant to check the robustness of the results (see
644
Supplementary Materials). Regions were screened for low BOLD signal intensity, with a
645
method adapted from Brown et. al. (Brown et al., 2019). Specifically, we found the elbow of
646
the BOLD signal distribution, given by the largest decrease in pair-wise differences of the
647
mean BOLD signal of each region. This was used as a cut-off for signal dropout and regions
648
with lower signal than the cut-off were considered to have signal dropout. Regions that were
649
found to have signal dropout in over 5% of subjects were excluded before analysis. For the
650
s100 atlas, 15 regions were excluded; for the s200 atlas, 16 regions were excluded from
651
further analysis.
652
To derive individually-tailored parcellations, we used the Group Prior Individualized
653
Parcellation (GPIP) model (Chong et al., 2017), which relies on a Bayesian formulation with
654
two priors: one based on group FC and one that drives individualized parcel boundaries. The
655
former uses a group sparsity constraint to represent FC between parcels, which allows the
656
model to maintain comparability between subjects. The latter uses a Markov Random Field in
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
t
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
657
the form of a Potts model to label the set of parcels and maximize the FC homogeneity within
658
each parcel based on individual data. This model allows for comparability between subjects,
659
as it maintains the same areas and labels for every individual while capturing the variability
660
in the shape and size of each parcel to best estimate each subject’s functional regions.
661
Individualized parcel borders were optimised across 20 iterations, starting with the group-
662
based Schaefer atlas and iteratively alternating between updating individual borders and the
663
group FC prior. Further details are provided in Chong et al. (Chong et al., 2017). The
664
algorithm was applied to patients and controls together.
665
For both group-based and individualized parcellations, mean timeseries were
666
extracted for each region in the s100 and s200 atlases using each individual’s spatially
667
normalized and denoised functional data. Product-moment correlations were then estimated
668
for every pair of regional time series to generate FC matrices. We only consider cortical areas
669
here as, to our knowledge, methods for developing individualized parcellations for
670
subcortical and cerebellar regions have not yet been developed.
671
Parcellation homogeneity and variability
672
To investigate the differences in parcels between the two parcellation approaches, we
673
computed how many vertices were reassigned to a different parcel after applying GPIP. We
674
then compared the number of vertices relabelled between patients and controls at a ROI and
675
whole-brain levels. All between-group statistical analyses were evaluated using permutation-
676
based inference, with 5000 permutations, unless otherwise indicated. Statistically significant
677
effects for ROI-level analysis were identified using an FDR-corrected (Benjamini &
678
Hochberg, 1995) threshold of 𝑝𝐹𝐷𝑅 < 0.05, two-tailed.
679
We compared the within-parcel functional homogeneity of the group-based and
680
individualized parcellations as per prior work (Chong et al., 2017; Schaefer et al., 2018). We
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
.
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
681
calculated the average FC between all pairs of vertices in a given parcel 𝑖, denoted 𝐹𝐶𝑖. Then,
682
parcellation homogeneity 𝐻 was normalised by parcel size as follows:
683
𝐻 =
𝑛
𝑖=1
∑ 𝐹𝐶𝑖 × 𝑁𝑉𝑖
𝑛
∑ 𝑁𝑉𝑖
𝑖=1
684
where 𝑛 is the total number of parcels in the parcellation and 𝑁𝑉 is the number of vertices in
685
the 𝑖𝑡ℎ parcel. This analysis was done out of sample, on functional scans from the second run
686
(PE=PA) with parcellations generated for scans from the first run (PE=AP).
687
To measure intra-subject reliability, we also computed homogeneity scores in the first
688
run and compared these results between parcellation approaches, using the intraclass
689
correlation coefficient.
690
Case-control differences in inter-regional functional coupling
691
We assessed how parcellation type influences FC differences between patients with
692
psychosis and healthy controls in three ways. First, we examined the distribution of
693
unthresholded t-statistics obtained at each edge using a general linear model to quantify mean
694
differences between patients and controls groups. This and all subsequent analyses are
695
controlled for age, sex, test site, and mean FD. The contrast was specified such that a larger t-
696
statistic indicated lower FC in patients, compared to controls. To compare the similarity of
697
the symmetric t-matrices, we vectorized their upper triangles and computed their Spearman
698
correlation. The effect of parcellation type was evaluated using a shift function test on these
699
distributions (Rousselet et al., 2017) to evaluate whether differences between parcellations
700
were restricted to specific quantiles of the 𝑡-statistic distributions (rather than just comparing
701
the means of these distributions). The shift function computes the difference in value of the 9
702
deciles of the distributions. For inference, it computes the 95% CI associated with each decile
703
difference, based on a bootstrap estimation of the standard error of each decile, controlling
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
t
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
704
for multiple comparisons, via the Hochberg’s method. This analysis thus allowed us to
705
determine whether parcellation type preferentially affected results for edges that showed
706
small, moderate, or large case-control differences.
707
Second, we compared thresholded results obtained with the Network Based Statistic
708
(NBS) (Zalesky, Fornito, & Bullmore, 2010). NBS is an adaptation of cluster-based statistics
709
for network data. A primary threshold of 𝑝 = 0.05, uncorrected, was applied to the matrix of
710
𝑡-statistics obtained using the general linear model described above. Results were repeated
711
with a threshold p = 0.01 and 0.001. The sizes of the connected components of the resulting
712
network (in terms of number of edges) were then estimated. In this context, the connected
713
components represent sets of nodes through which a path can be found via supra-threshold
714
edges. The group labels (patients and controls) were permuted 5000 times and the previous
715
steps were repeated. At each step, the size of the largest connected component was retained,
716
resulting in an empirical distribution of maximal component sizes under the null hypothesis.
717
The fraction of null values that exceeded the observed component sizes corresponds to a
718
family-wise corrected 𝑝-value for each component. The test was repeated with different
719
FWER corrected p-values = 0.05, 0.01, and 0.001, all resulting in the same connected
720
component. By performing inference at the level of connected components rather than
721
individual edges, the NBS results in greater statistical power than traditional mass univariate
722
thresholding methods (Zalesky, Fornito, & Bullmore, 2010). This analysis was repeated for
723
each parcellation type (i.e., group-based and individualized) and scale (i.e., s100 and s200).
724
Differences between significant component sizes observed using the two parcellation
725
methods were then estimated and evaluated with respect to the differences between null
726
component sizes estimated for the two approaches.
727
We calculated changes in parcel size between parcellation approaches for patients and
728
controls. We compared parcel size difference with a two-sample t-test between patients and
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
.
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
729
controls. To understand how parcel size impacted FC measures, we calculated the Spearman
730
rho’s correlation between the t-values for parcel size and mean dysconnectivity per parcel
731
and degree of dysconnectivity. p-values were calculated with a spin permutation test, with
732
5000 permutations (Alexander-Bloch et al., 2018).
733
Finally, we examined how parcellation type affects case-control differences at the
734
level of 7 canonical networks. We considered the control network; the default mode network;
735
the dorsal attention network; the limbic network; the salience/ventral attention network; the
736
somatomotor network; and the visual network using the seven Yeo network assignments
737
associated with the s100 and s200 atlases (Yeo et al., 2011). Specifically, we quantified the
738
number of edges in the significant NBS component that fell within and between these seven
739
networks. We examined both raw edge counts and counts normalized for the size of each
740
network/network pair and quantified the correlation between the resulting network-level
741
matrices obtained for each parcellation type.
742
Code used for analysis and
image generation can be found on-line at
743
https://github.com/NSBLab/individualised_parc_psychosis and code
for
individualized
744
parcellation can be acquired on-line at https://neuroimageusc.github.io/GPIP
745
746
747
748
749
750
751
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
.
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
752
Reference
753
Alexander-Bloch, A. F., Shou, H., Liu, S., Satterthwaite, T. D., Glahn, D. C., Shinohara, R. T., Vandekar,
754
755
756
S. N., & Raznahan, A. (2018). On testing for spatial correspondence between maps of human
brain structure and function. NeuroImage, 178, 540–551.
https://doi.org/10.1016/j.neuroimage.2018.05.070
757
Amunts, K., Kedo, O., Kindler, M., Pieperhoff, P., Mohlberg, H., Shah, N. J., Habel, U., Schneider, F., &
758
759
760
Zilles, K. (2005). Cytoarchitectonic mapping of the human amygdala, hippocampal region and
entorhinal cortex: Intersubject variability and probability maps. Anatomy and Embryology,
210(5–6), 343–352. https://doi.org/10.1007/s00429-005-0025-5
761
Andersson, J. L. R., Jenkinson, M., & Smith, S. (2010). FMRIB Analysis Group Technical Reports.
762
https://www.fmrib.ox.ac.uk/datasets/techrep/
763
Andersson, J. L. R., Skare, S., & Ashburner, J. (2003). How to correct susceptibility distortions in spin-
764
765
echo echo-planar images: Application to diffusion tensor imaging. NeuroImage, 20(2), 870–
888. https://doi.org/10.1016/S1053-8119(03)00336-7
766
Andreasen, N. C., Paradiso, S., & O’Leary, D. S. (1998). “Cognitive dysmetria” as an integrative theory
767
768
of schizophrenia: A dysfunction in cortical-subcortical-cerebellar circuitry? Schizophrenia
Bulletin, 24(2), 203–218. https://doi.org/10.1093/oxfordjournals.schbul.a033321
769
Aquino, K. M., Fulcher, B. D., Parkes, L., Sabaroedin, K., & Fornito, A. (2020). Identifying and
770
771
772
removing widespread signal deflections from fMRI data: Rethinking the global signal
regression problem. NeuroImage, 212, 116614.
https://doi.org/10.1016/j.neuroimage.2020.116614
773
Argyelan, M., Ikuta, T., Derosse, P., Braga, R. J., Burdick, K. E., John, M., Kingsley, P. B., Malhotra, A.
774
775
776
K., & Szeszko, P. R. (2014). Resting-state fMRI connectivity impairment in schizophrenia and
bipolar disorder. Schizophrenia Bulletin, 40(1), 100–110.
https://doi.org/10.1093/schbul/sbt092
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
t
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
777
Baker, J. T., Dillon, D. G., Patrick, L. M., Roffman, J. L., Brady, R. O., Pizzagalli, D. A., Öngür, D., &
778
779
780
Holmes, A. J. (2019). Functional connectomics of affective and psychotic pathology.
Proceedings of the National Academy of Sciences, 116(18), 9050–9059.
https://doi.org/10.1073/pnas.1820780116
781
Benjamini, Y., & Hochberg, Y. (1995). Controlling the False Discovery Rate: A Practical and Powerful
782
783
Approach to Multiple Testing. Journal of the Royal Statistical Society. Series B
(Methodological), 57(1), 289–300.
784
Bijsterbosch, J. D., Woolrich, M. W., Glasser, M. F., Robinson, E. C., Beckmann, C. F., Van Essen, D. C.,
785
786
Harrison, S. J., & Smith, S. M. (2018). The relationship between spatial configuration and
functional connectivity of brain regions. https://doi.org/10.7554/eLife.32992.001
787
Brown, J. A., Deng, J., Neuhaus, J., Sible, I. J., Sias, A. C., Lee, S. E., Kornak, J., Marx, G. A., Karydas, A.
788
789
790
791
M., Spina, S., Grinberg, L. T., Coppola, G., Geschwind, D. H., Kramer, J. H., Gorno-Tempini, M.
L., Miller, B. L., Rosen, H. J., & Seeley, W. W. (2019). Patient-Tailored, Connectivity-Based
Forecasts of Spreading Brain Atrophy. Neuron, 104(5), 856-868.e5.
https://doi.org/10.1016/J.NEURON.2019.08.037
792
Chong, M., Bhushan, C., Joshi, A. A., Choi, S., Haldar, J. P., Shattuck, D. W., Spreng, R. N., & Leahy, R.
793
794
M. (2017). Individual parcellation of resting fMRI with a group functional connectivity prior.
NeuroImage, 156, 87–100. https://doi.org/10.1016/j.neuroimage.2017.04.054
795
Chopra, S., Francey, S. M., O’Donoghue, B., Sabaroedin, K., Arnatkeviciute, A., Cropley, V., Nelson, B.,
796
797
798
799
800
Graham, J., Baldwin, L., Tahtalian, S., Yuen, H. P., Allott, K., Alvarez-Jimenez, M., Harrigan, S.,
Pantelis, C., Wood, S. J., McGorry, P., & Fornito, A. (2021). Functional Connectivity in
Antipsychotic-Treated and Antipsychotic-Naive Patients with First-Episode Psychosis and
Low Risk of Self-harm or Aggression: A Secondary Analysis of a Randomized Clinical Trial.
JAMA Psychiatry, 78(9), 994–1004. https://doi.org/10.1001/jamapsychiatry.2021.1422
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
801
Dale, A. M., Fischl, B., & Sereno, M. I. (1999). Cortical Surface-Based Analysis: I. Segmentation and
802
803
Surface Reconstruction. NeuroImage, 9(2), 179–194.
https://doi.org/10.1006/NIMG.1998.0395
804
Demro, C., Mueller, B. A., Kent, J. S., Burton, P. C., Olman, C. A., Schallmo, M. P., Lim, K. O., &
805
806
Sponheim, S. R. (2021). The psychosis human connectome project: An overview.
NeuroImage, 241. https://doi.org/10.1016/j.neuroimage.2021.118439
807
Eickhoff, S. B., Constable, R. T., & Yeo, B. T. T. (2018). Topographic organization of the cerebral
808
809
cortex and brain cartography. NeuroImage, 170, 332–347.
https://doi.org/10.1016/j.neuroimage.2017.02.018
810
Eickhoff, S. B., Yeo, B. T. T., & Genon, S. (2018). Imaging-based parcellations of the human brain.
811
Nature Reviews Neuroscience, 19(11), 672–686. https://doi.org/10.1038/s41583-018-0071-7
812
Esteban, O., Birman, D., Schaer, M., Koyejo, O. O., Poldrack, R. A., & Gorgolewski, K. J. (2017).
813
814
MRIQC: Advancing the automatic prediction of image quality in MRI from unseen sites. PLOS
ONE, 12(9), e0184661. https://doi.org/10.1371/JOURNAL.PONE.0184661
815
Fischl, B. (2012). FreeSurfer. NeuroImage, 62(2), 774–781.
816
https://doi.org/10.1016/j.neuroimage.2012.01.021
817
Fornito, A., Zalesky, A., & Bullmore, E. T. (2010). Network scaling effects in graph analytic studies of
818
819
human resting-state FMRI data. Frontiers in Systems Neuroscience, 4, 22.
https://doi.org/10.3389/fnsys.2010.00022
820
Fornito, A., Zalesky, A., & Bullmore, E. T. (2016). Fundamentals of Brain Network Analysis. Academic
821
Press. https://doi.org/10.1016/B978-0-12-407908-3.09999-4
822
Fornito, A., Zalesky, A., Pantelis, C., & Bullmore, E. T. (2012). Schizophrenia, neuroimaging and
823
824
connectomics. NeuroImage, 62(4), 2296–2314.
https://doi.org/10.1016/j.neuroimage.2011.12.090
825
Glasser, M. F., Coalson, T. S., Robinson, E. C., Hacker, C. D., Harwell, J., Yacoub, E., Ugurbil, K.,
826
Andersson, J., Beckmann, C. F., Jenkinson, M., Smith, S. M., & Van Essen, D. C. (2016). A
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
827
828
multi-modal parcellation of human cerebral cortex. Nature, 536(7615), 171–178.
https://doi.org/10.1038/nature18933
829
Glasser, M. F., Sotiropoulos, S. N., Wilson, J. A., Coalson, T. S., Fischl, B., Andersson, J. L., Xu, J.,
830
831
832
Jbabdi, S., Webster, M., Polimeni, J. R., Van Essen, D. C., & Jenkinson, M. (2013). The minimal
preprocessing pipelines for the Human Connectome Project. NeuroImage, 80, 105–124.
https://doi.org/10.1016/j.neuroimage.2013.04.127
833
Gordon, E. M., Laumann, T. O., Gilmore, A. W., Newbold, D. J., Greene, D. J., Berg, J. J., Ortega, M.,
834
835
836
837
Hoyt-Drazen, C., Gratton, C., Sun, H., Hampton, J. M., Coalson, R. S., Nguyen, A. L.,
McDermott, K. B., Shimony, J. S., Snyder, A. Z., Schlaggar, B. L., Petersen, S. E., Nelson, S. M.,
& Dosenbach, N. U. F. (2017). Precision Functional Mapping of Individual Human Brains.
Neuron, 95(4), 791-807.e7. https://doi.org/10.1016/j.neuron.2017.07.011
838
Goscinski, W. J., McIntosh, P., Felzmann, U., Maksimenko, A., Hall, C., Gureyev, T., Thompson, D.,
839
840
841
842
843
Janke, A., Galloway, G., Killeen, N., Raniga, P., Kaluza, O., Ng, A., Poudel, G., Barnes, D.,
Nguyen, T., Bonnington, P., & Egan, G. (2014). The multi-modal Australian ScienceS Imaging
and Visualization Environment (MASSIVE) high performance computing infrastructure:
Applications in neuroscience and neuroinformatics research. Frontiers in Neuroinformatics,
8. https://www.frontiersin.org/articles/10.3389/fninf.2014.00030
844
Grabner, G., Janke, A. L., Budge, M. M., Smith, D., Pruessner, J., & Collins, D. L. (2006). Symmetric
845
846
847
848
atlasing and model based segmentation: An application to the hippocampus in older adults.
Medical Image Computing and Computer-Assisted Intervention : MICCAI ... International
Conference on Medical Image Computing and Computer-Assisted Intervention, 9(Pt 2), 58–
66. https://doi.org/10.1007/11866763_8
849
Greve, D. N., & Fischl, B. (2009). Accurate and robust brain image alignment using boundary-based
850
851
registration. NeuroImage, 48(1), 63–72.
https://doi.org/10.1016/J.NEUROIMAGE.2009.06.060
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
.
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
852
Griffanti, L., Salimi-Khorshidi, G., Beckmann, C. F., Auerbach, E. J., Douaud, G., Sexton, C. E., Zsoldos,
853
854
855
856
E., Ebmeier, K. P., Filippini, N., Mackay, C. E., Moeller, S., Xu, J., Yacoub, E., Baselli, G.,
Ugurbil, K., Miller, K. L., & Smith, S. M. (2014). ICA-based artefact removal and accelerated
fMRI acquisition for improved resting state network imaging. NeuroImage, 95, 232–247.
https://doi.org/10.1016/J.NEUROIMAGE.2014.03.034
857
HCP Early Psychosis 1.1 Data Release: Reference Manual HUMAN Connectome PROJECT for Early
858
Psychosis. (2021). https://github.com/pnlbwh
859
Hummer, T. A., Yung, M. G., Goñi, J., Conroy, S. K., Francis, M. M., Mehdiyoun, N. F., & Breier, A.
860
861
(2020). Functional network connectivity in early-stage schizophrenia. Schizophrenia
Research, 218, 107–115. https://doi.org/10.1016/j.schres.2020.01.023
862
Jenkinson, M., Bannister, P., Brady, M., & Smith, S. (2002). Improved Optimization for the Robust
863
864
and Accurate Linear Registration and Motion Correction of Brain Images. NeuroImage, 17(2),
825–841. https://doi.org/10.1006/NIMG.2002.1132
865
Jenkinson, M., & Smith, S. (2001). A global optimisation method for robust affine registration of
866
867
brain images. Medical Image Analysis, 5(2), 143–156. https://doi.org/10.1016/S1361-
8415(01)00036-6
868
Kong, R., Li, J., Orban, C., Sabuncu, M. R., Liu, H., Schaefer, A., Sun, N., Zuo, X. N., Holmes, A. J.,
869
870
871
Eickhoff, S. B., & Yeo, B. T. T. (2019). Spatial Topography of Individual-Specific Cortical
Networks Predicts Human Cognition, Personality, and Emotion. Cerebral Cortex, 29(6), 2533–
2551. https://doi.org/10.1093/cercor/bhy123
872
Kong, R., Yang, Q., Gordon, E., Xue, A., Yan, X., Orban, C., Zuo, X.-N., Spreng, N., Ge, T., Holmes, A.,
873
874
875
Eickhoff, S., & Yeo, B. T. T. (2021). Individual-Specific Areal-Level Parcellations Improve
Functional Connectivity Prediction of Behavior. Cerebral Cortex (New York, N.Y.: 1991),
31(10), 4477–4500. https://doi.org/10.1093/cercor/bhab101
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
t
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
876
Lewandowski, K. E., Bouix, S., Ongur, D., & Shenton, M. E. (2020). Neuroprogression across the Early
877
878
Course of Psychosis. Journal of Psychiatry and Brain Science, 5, e200002.
https://doi.org/10.20900/jpbs.20200002
879
Li, S., Chen, S., Yue, C., & Caffo, B. (2016). A Parcellation Based Nonparametric Algorithm for
880
881
Independent Component Analysis with Application to fMRI Data. Frontiers in Neuroscience,
10. https://www.frontiersin.org/articles/10.3389/fnins.2016.00015
882
Li, T., Wang, Q., Zhang, J., Rolls, E. T., Yang, W., Palaniyappan, L., Zhang, L., Cheng, W., Yao, Y., Liu, Z.,
883
884
885
886
Gong, X., Luo, Q., Tang, Y., Crow, T. J., Broome, M. R., Xu, K., Li, C., Wang, J., Liu, Z., … Feng, J.
(2017). Brain-Wide Analysis of Functional Connectivity in First-Episode and Chronic Stages of
Schizophrenia. Schizophrenia Bulletin, 43(2), 436–448.
https://doi.org/10.1093/schbul/sbw099
887
Li, Y., Liu, A., Fu, X., Mckeown, M. J., Wang, Z. J., & Chen, X. (2022). Atlas-guided parcellation:
888
889
Individualized functionally-homogenous parcellation in cerebral cortex. Computers in Biology
and Medicine, 150, 106078. https://doi.org/10.1016/j.compbiomed.2022.106078
890
Lynall, M. E., Bassett, D. S., Kerwin, R., McKenna, P. J., Kitzbichler, M., Muller, U., & Bullmore, E.
891
892
(2010). Functional connectivity and brain networks in schizophrenia. Journal of
Neuroscience, 30(28), 9477–9487. https://doi.org/10.1523/JNEUROSCI.0333-10.2010
893
McGraw, K. O., & Wong, S. P. (1996). Forming inferences about some intraclass correlation
894
coefficients. Psychological Methods, 1, 30–46. https://doi.org/10.1037/1082-989X.1.1.30
895
Moran, L. V., Tagamets, M. A., Sampath, H., O’Donnell, A., Stein, E. A., Kochunov, P., & Hong, L. E.
896
897
898
(2013). Disruption of anterior insula modulation of large-scale brain networks in
schizophrenia. Biological Psychiatry, 74(6), 467–474.
https://doi.org/10.1016/j.biopsych.2013.02.029
899
Mueller, S., Wang, D., Fox, M. D., Yeo, B. T. T., Sepulcre, J., Sabuncu, M. R., Shafee, R., Lu, J., & Liu, H.
900
901
(2013). Individual Variability in Functional Connectivity Architecture of the Human Brain.
Neuron, 77(3), 586–595. https://doi.org/10.1016/j.neuron.2012.12.028
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
902
Nabulsi, L., McPhilemy, G., Kilmartin, L., Whittaker, J. R., Martyn, F. M., Hallahan, B., McDonald, C.,
903
904
905
906
Murphy, K., & Cannon, D. M. (2020). Frontolimbic, Frontoparietal, and Default Mode
Involvement in Functional Dysconnectivity in Psychotic Bipolar Disorder. Biological
Psychiatry. Cognitive Neuroscience and Neuroimaging, 5(2), 140–151.
https://doi.org/10.1016/j.bpsc.2019.10.015
907
Narr, K. L., & Leaver, A. M. (2015). Connectome and schizophrenia. Current Opinion in Psychiatry,
908
28(3), 229–235. https://doi.org/10.1097/YCO.0000000000000157
909
Nogovitsyn, N., Metzak, P. D., Casseb, R. F., Souza, R., Harris, J. K., Prati, L. M., Zamyadi, M., Bray, S.
910
911
912
913
L., Lebel, C., Hassel, S., Strother, S., Goldstein, B. I., Wang, J., Kennedy, S. H., MacQueen, G.
M., & Addington, J. (2022). Cerebello-limbic functional connectivity patterns in youth at
clinical high risk for psychosis. Schizophrenia Research, 240, 220–227.
https://doi.org/10.1016/j.schres.2021.12.041
914
Parkes, L., Fulcher, B., Yücel, M., & Fornito, A. (2018). An evaluation of the efficacy, reliability, and
915
916
sensitivity of motion correction strategies for resting-state functional MRI. NeuroImage, 171,
415–436. https://doi.org/10.1016/J.NEUROIMAGE.2017.12.073
917
Power, J. D., Barnes, K. A., Snyder, A. Z., Schlaggar, B. L., & Petersen, S. E. (2012). Spurious but
918
919
systematic correlations in functional connectivity MRI networks arise from subject motion.
Neuroimage, 59(3), 2142. https://doi.org/10.1016/J.NEUROIMAGE.2011.10.018
920
Power, J. D., Lynch, C. J., Silver, B. M., Dubin, M. J., Martin, A., & Jones, R. M. (2019). Distinctions
921
922
among real and apparent respiratory motions in human fMRI data. NeuroImage, 201,
116041. https://doi.org/10.1016/j.neuroimage.2019.116041
923
Power, J. D., Plitt, M., Laumann, T. O., & Martin, A. (2017). Sources and implications of whole-brain
924
925
fMRI signals in humans. NeuroImage, 146, 609–625.
https://doi.org/10.1016/j.neuroimage.2016.09.038
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
.
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
926
Rousselet, G. A., Pernet, C. R., & Wilcox, R. R. (2017). Beyond differences in means: Robust graphical
927
928
methods to compare two groups in neuroscience (p. 121079). bioRxiv.
https://doi.org/10.1101/121079
929
Salehi, M., Karbasi, A., Shen, X., Scheinost, D., & Constable, R. T. (2018). An exemplar-based
930
931
approach to individualized parcellation reveals the need for sex specific functional networks.
NeuroImage, 170, 54–67. https://doi.org/10.1016/j.neuroimage.2017.08.068
932
Salimi-Khorshidi, G., Douaud, G., Beckmann, C. F., Glasser, M. F., Griffanti, L., & Smith, S. M. (2014).
933
934
935
Automatic denoising of functional MRI data: Combining independent component analysis
and hierarchical fusion of classifiers. NeuroImage, 90, 449–468.
https://doi.org/10.1016/J.NEUROIMAGE.2013.11.046
936
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X.-N., Holmes, A. J., Eickhoff, S. B., & Yeo,
937
938
939
B. T. T. (2018). Local-Global Parcellation of the Human Cerebral Cortex from Intrinsic
Functional Connectivity MRI. Cerebral Cortex, 28(9), 3095–3114.
https://doi.org/10.1093/cercor/bhx179
940
Smith, S. M. (2002). Fast robust automated brain extraction. Human Brain Mapping, 17(3), 143–155.
941
https://doi.org/10.1002/HBM.10062
942
Thomas Yeo, B. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D., Hollinshead, M.,
943
944
945
946
Roffman, J. L., Smoller, J. W., Zöllei, L., Polimeni, J. R., Fischl, B., Liu, H., & Buckner, R. L.
(2011). The organization of the human cerebral cortex estimated by intrinsic functional
connectivity. Journal of Neurophysiology, 106(3), 1125–1165.
https://doi.org/10.1152/jn.00338.2011
947
Tu, P. C., Lee, Y. C., Chen, Y. S., Li, C. T., & Su, T. P. (2013). Schizophrenia and the brain’s control
948
949
950
network: Aberrant within- and between-network connectivity of the frontoparietal network
in schizophrenia. Schizophrenia Research, 147(2–3), 339–347.
https://doi.org/10.1016/j.schres.2013.04.011
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
.
/
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
951
Welsh, R. C., Chen, A. C., & Taylor, S. F. (2010). Low-frequency BOLD fluctuations demonstrate
952
953
altered thalamocortical connectivity in schizophrenia. Schizophrenia Bulletin, 36(4), 713–
722. https://doi.org/10.1093/schbul/sbn145
954
Whitfield-Gabrieli, S., Thermenos, H. W., Milanovic, S., Tsuang, M. T., Faraone, S. V., Mccarley, R. W.,
955
956
957
958
Shenton, M. E., Green, A. I., Nieto-Castanon, A., Laviolette, P., Wojcik, J., Gabrieli, J. D. E.,
Seidman, L. J., & Raichle, M. E. (2009). Hyperactivity and hyperconnectivity of the default
network in schizophrenia and in first-degree relatives of persons with schizophrenia.
www.pnas.org/cgi/content/full/
959
Woodward, N. D., Rogers, B., & Heckers, S. (2011). Functional resting-state networks are
960
961
differentially affected in schizophrenia. Schizophrenia Research, 130(1–3), 86–93.
https://doi.org/10.1016/j.schres.2011.03.010
962
Xia, M., Wang, J., & He, Y. (2013). BrainNet Viewer: A Network Visualization Tool for Human Brain
963
Connectomics. PLOS ONE, 8(7), e68910. https://doi.org/10.1371/journal.pone.0068910
964
Yeo, B. T. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D., Hollinshead, M., Roffman, J. L.,
965
966
967
Smoller, J. W., Zöllei, L., Polimeni, J. R., Fisch, B., Liu, H., & Buckner, R. L. (2011). The
organization of the human cerebral cortex estimated by intrinsic functional connectivity.
Journal of Neurophysiology, 106(3), 1125–1165. https://doi.org/10.1152/jn.00338.2011
968
Zalesky, A., Fornito, A., & Bullmore, E. T. (2010). Network-based statistic: Identifying differences in
969
970
brain networks. NeuroImage, 53(4), 1197–1207.
https://doi.org/10.1016/J.NEUROIMAGE.2010.06.041
971
Zalesky, A., Fornito, A., Harding, I. H., Cocchi, L., Yücel, M., Pantelis, C., & Bullmore, E. T. (2010).
Whole-brain anatomical networks: Does the choice of nodes matter? NeuroImage, 50(3),
970–983. https://doi.org/10.1016/j.neuroimage.2009.12.027
972
973
974
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
t
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure1
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
t
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
/
t
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
/
/
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
2
9
2
1
4
2
0
1
4
n
e
n
_
a
_
0
0
3
2
9
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3