faber, SEM, belden, A.G., Luis, PAG., & McInsosh, R. (2023). Variabilidad relacionada con la edad en la participación en la red durante la música
escuchando. Neurociencia en red, Publicación anticipada. https://doi.org/10.1162/netn_a_00333.
Age-related variability in network
engagement during music listening
Author List: faber, S.1,2, belden, A.G.3, Luis, P.3, & McIntosh, A.R.2
1universidad de toronto
2Simon Fraser University
3Northeastern University
1
2
3
4
5
6
7
8
Abstracto
9
Listening to music is an enjoyable behaviour that engages multiple networks of brain regions.
10
Tal como, the act of music listening may offer a way to interrogate network activity, y para
11
examine the reconfigurations of brain networks that have been observed in healthy aging. El
12
present study is an exploratory examination of brain network dynamics during music listening in
13
healthy older and younger adults. Network measures were extracted and analyzed together with
14
behavioural data using a combination of hidden Markov modelling and partial least squares. Nosotros
15
found age- and preference-related differences in fMRI data collected during music listening in
16
healthy younger and older adults. Both age groups showed higher occupancy (the proportion of
17
time a network was active) in a temporal-mesolimbic network while listening to self-selected
18
música. Activity in this network was strongly positively correlated with liking and familiarity
19
ratings in younger adults, but less so in older adults. Además, older adults showed a higher
20
degree of correlation between liking and familiarity ratings consistent with past behavioural
1
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
/
/
t
.
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
norte
mi
norte
_
a
_
0
0
3
3
3
pag
d
/
.
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
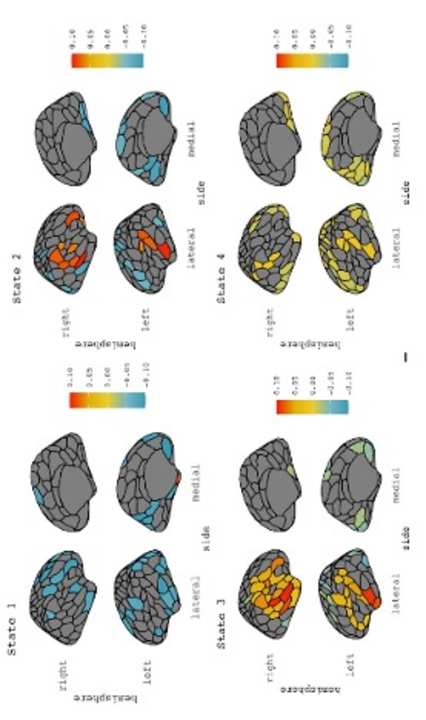
metro
b
mi
r
2
0
2
3
21
work on age-related dedifferentiation. We conclude that, while older adults do show network and
22
behaviour patterns consistent with dedifferentiation, activity in the temporal-mesolimbic network
23
is relatively robust to dedifferentiation. These findings may help explain how music listening
24
remains meaningful and rewarding in old age.
25
26
Palabras clave: Music, Aging, Computational Neuroscience
27
Fondo
28
Brain function changes with age across multiple spatial scales. The brain can be thought of as a
29
series of overlapping functional networks where each network is a collection of brain regions
30
that act in concert over time. With age, regions that were once nodes in densely-connected
31
functional networks may become disconnected while regions in previously distinct functional
32
networks may become more connected (Grady et al., 2016), though whether this reconfiguration
33
of functional network boundaries is adaptive or maladaptive remains unclear. In healthy older
34
adultos, networks that were once well-defined and responded preferentially to a particular
35
stimulus or set of conditions begin to activate (or to fail to deactivate) less discerningly in a
36
process known as dedifferentiation (Grady et al., 2012; Rieck et al., 2017).
37
38
In music listening, there is behavioural evidence of age-related perceptual changes that may
39
serve as a behavioural counterpart to the dedifferentiation seen in network brain dynamics.
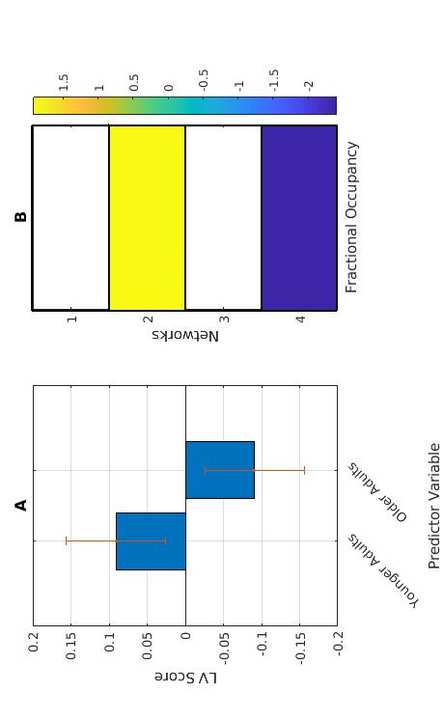
40
Music is reported as more broadly pleasant with age (a positivity effect, Bones & Plack, 2015;
41
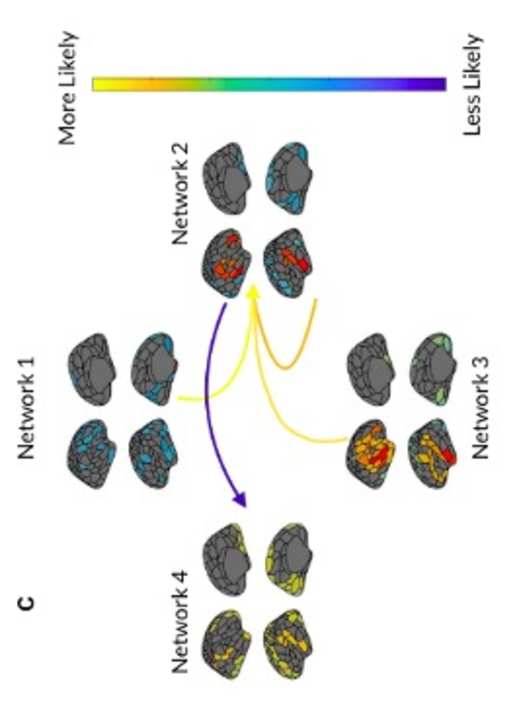
Groarke & Hogan, 2019; Laukka & Juslin, 2007; Lima & Castro, 2011), and perceptual features
42
also become less distinct with age, with higher correlations observed between perceived arousal
2
yo
D
oh
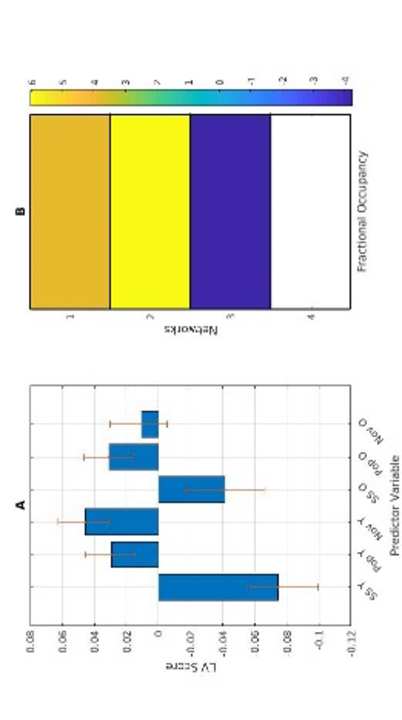
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
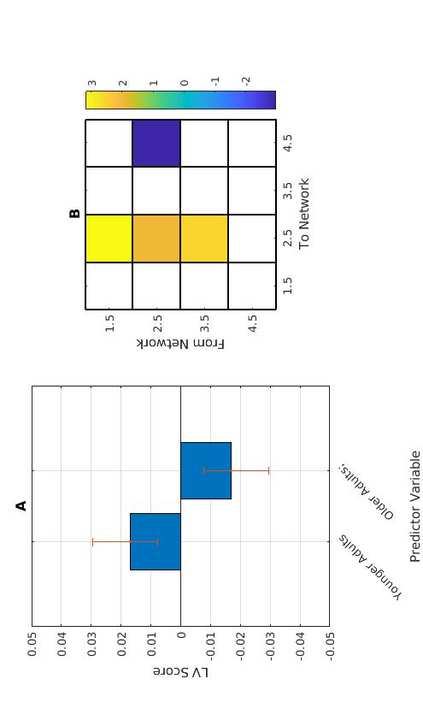
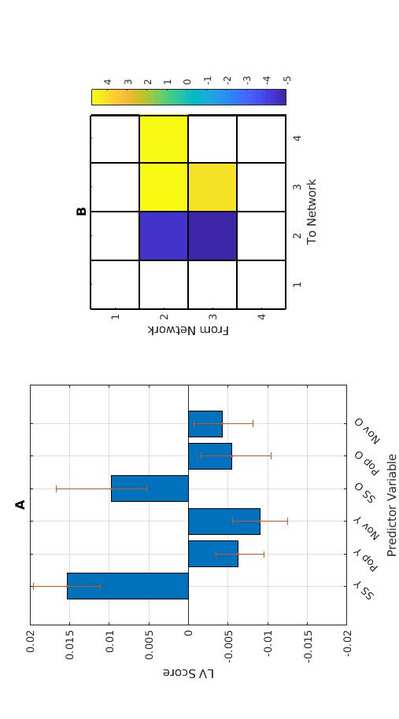
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
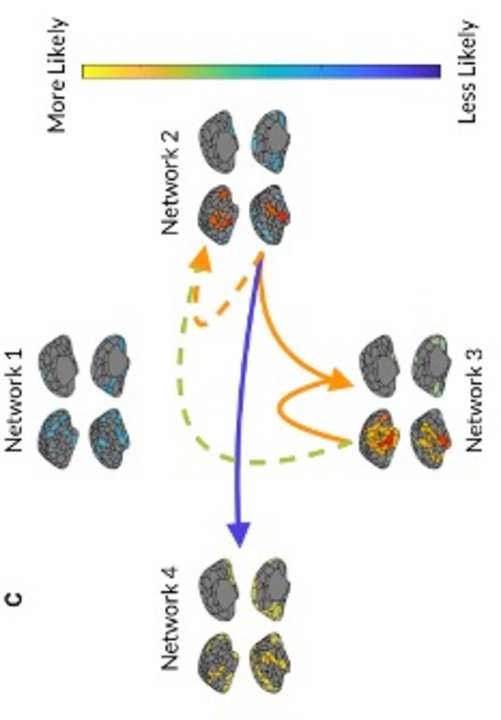
F
/
d
oh
i
/
t
.
/
/
1
0
1
1
6
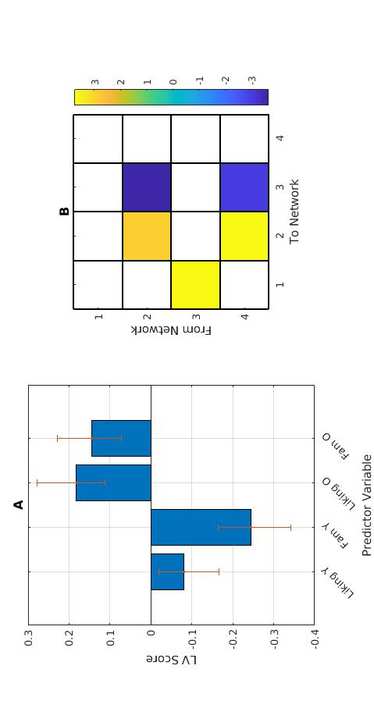
2
norte
mi
norte
_
a
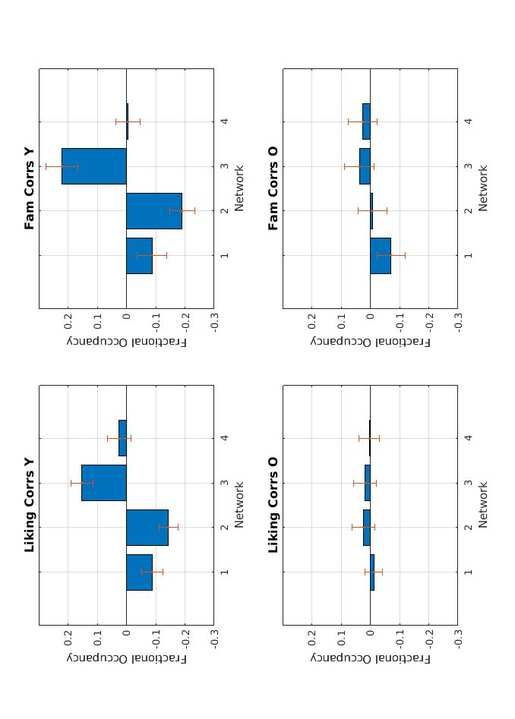
_
0
0
3
3
3
2
1
5
5
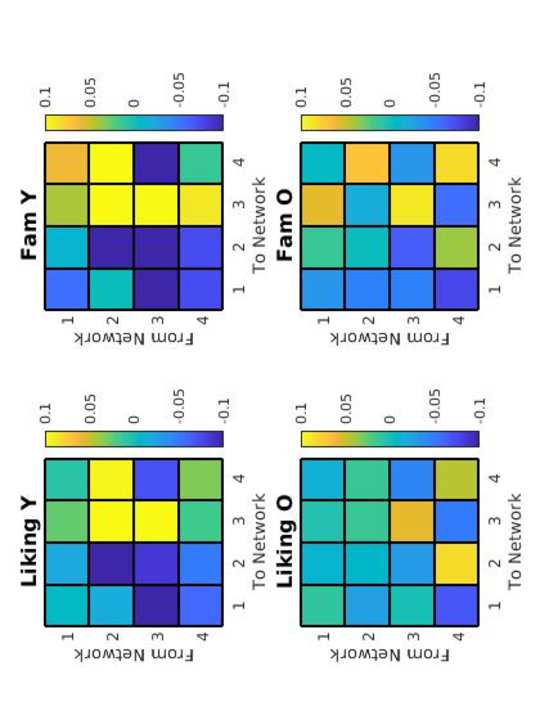
2
0
2
norte
mi
norte
_
a
_
0
0
3
3
3
pag
d
.
t
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
43
and valence in older adults (Vieillard et al., 2012). This blurring of the lines between the
44
perceived pleasantness and dimensions of a musical signal might indicate underlying network
45
cambios, but do not seem to affect the music listening experience negatively.
46
47
Musical sounds are complex stimuli that, using building blocks of timbre, tono, pitch, ritmo,
48
melody, and harmony, can engender expectancy and surprise to make us laugh, cry, dance, sing,
49
and reminisce. As musical stimuli are complex and hierarchically organized, brain responses to
50
music are likewise complex and hierarchical, with many temporally-dependent overlapping
51
procesos. Features extracted from musical signals stimulate activity in multiple brain regions
52
(Alluri et al., 2012; Burunat et al., 2017; Williams et al., 2022), and networks, including the
53
default mode network (DMN; Wilkins et al., 2014; Koelsch et al., 2022; Taruffi et al., 2017) y
54
reward networks (Fasano et al., 2022).
55
56
Multivariate statistical modelling tools provide us with a unique opportunity to observe and
57
describe whole-brain network activity in a data-driven way. Working in network space, dónde
58
the smallest unit of measurement is a network, allows us to examine the shifting patterns of brain
59
activity that accompany music, which has the potential to add nuance that cannot be seen when
60
looking at isolated regions of interest. This approach may also be of value in understanding the
61
neural foundation of age-related perceptual changes, and may shed light on why music is so
62
salient in clinical populations (Cuddy & Duffin, 2005; Leggieri et al., 2017; Särkämö et al.,
63
2014; Thaut et al., 2020, Matziorinis & Koelsch, 2022).
64
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
/
.
t
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
norte
mi
norte
_
a
_
0
0
3
3
3
pag
d
/
t
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
65
Where older adults show network reconfigurations compared to younger cohorts in rest and
66
during cognitive tasks, what can music reveal about the aging brain? In the present exploratory
67
estudiar, we studied age differences in network-level dynamics during familiar and novel music
68
listening in a cohort of healthy younger and older adults. We aim to demonstrate age-related
69
changes in network dynamics using a novel analysis paradigm comprising hidden Markov
70
modelling and partial least squares analyses.
71
72
Métodos
73
Networks were estimated using hidden Markov modelling (HMM) and analyses were completed
74
using partial least squares (PLS). We chose HMM rather than a seed-based or canonical network
75
análisis (see Bressler & menón, 2010) in an effort to base our analyses on data-driven patterns
76
as much as possible. A substantial advantage of HMM is that it derives networks from patterns in
77
the original data without the constraints of canonical network boundaries or specified time
78
windows.
79
80
A brief outline of data collection is included here. For a detailed description of participant
81
recruitment, study protocol, and data acquisition, please see Quinci et al. (2022) and Belden et al.
82
(2023).
83
Participantes
84
Participants were right-handed, cognitively healthy younger (norte = 44, 11 machos, edad media =
85
19.24, DE = 1.92) and older (norte = 27, 13 machos, edad media = 67.34, DE = 8.27) adults with normal
86
hearing established via audiogram. Inclusion criteria included normal hearing, successful
87
completion of MRI screening, and a minimum age of 18 for younger adults and 50 for older
4
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
.
t
/
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
norte
mi
norte
_
a
_
0
0
3
3
3
pag
d
t
/
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
88
adultos. Exclusion criteria comprised medication changes 6 weeks prior to screening, a history of
89
any medical condition that could impair cognition, a history of chemotherapy in the preceding 10
90
años, or any medical condition requiring medical treatment within three months of screening.
91
Data from two younger participants were excluded following data collection due to problems
92
with the ratings apparatus. Ethics approval was granted by the Northeastern University
93
Institutional Review Board and all research was conducted consistent with the Declaration of
94
Helsinki.
95
Procedimiento
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
/
t
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
t
/
.
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
norte
mi
norte
_
a
_
0
0
3
3
3
pag
d
t
/
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
96
Prior to data collection, participants completed a screening call with researchers to confirm their
97
eligibility for the study, and to collect a list of six songs that are familiar and well-liked by the
98
partícipe. Following screening, eligible participants completed a battery of neuropsychological
99
pruebas, structural and functional MRI scans, and a blood draw. The present study focuses on the
100
fMRI data; other aspects of the results are in preparation and will be described in separate
101
reports.
102
Adquisición de datos
103
All scans took place at Northeastern University. Functional scans were acquired with a Siemens
104
Magnetom 3T scanner with a 64-channel head coil. The total scan time for task data was 11.4
105
minutes with continuous acquisition at a fast TR of 475 ms over 1440 volumes. A resting state
106
scan was also performed with these parameters, and findings will be reported in a future
107
manuscript. T1 images were captured, but will not be discussed in detail in this manuscript.
108
5
109
Task fMRI consisted of a block of resting state followed by music presentation (24 excerpts,
110
each played for 20 artículos de segunda clase). Musical excerpts were either familiar and well-liked self-selected
111
música (6/24), or experimenter-selected music chosen to be popular or possibly recognizable
112
(10/24), or novel including excerpts purpose-composed for research purposes (8/24). Estímulos
113
were presented randomly and following each 20 second musical excerpt, participants were asked
114
to rate their familiarity and liking of the excerpt for two seconds each, using 4-point Likert
115
escamas.
116
Data pre-processing
117
Functional MRI data were pre-processed using the TVB-UKBB pipeline detailed by Frazier-
118
Logue et al. (2022). T1 images were registered to the Montreal Neurological Institute T1
119
template. Functional data pre-processing was done using a pipeline using the FMRIB Software
120
Library (FSL; Woolrich et al., 2009), including the fMRI Expert Analysis Tool (FEAT, versión
121
6.0). Within the pipeline, pre-processing of functional data comprised gradient echo fieldmap
122
distortion correction, motion correction using MCFLIRT, and independent component analysis
123
(ICA) artifact classification using MELODIC and FIX.
124
125
We assembled an ICA training set for non-cerebral artifact detection. ICA reports from 16
126
participants per age group were visually inspected for noisy vs. clean components and manually
127
anotado. Subsequent participants’ ICA reports were cleaned using this training set. El
128
processed datasets were down-sampled to 220 regions of interest using the Schaefer-Tian 220
129
parcellation, which provides ample spatial resolution of auditory regions and subcortical
130
estructuras (Schaefer et al., 2017, Tian et al., 2020). Regional time series data were normalized to
6
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
.
/
t
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
norte
mi
norte
_
a
_
0
0
3
3
3
pag
d
.
/
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
131
control for between-subject amplitude differences and exported to MatLab (MathWorks, 2019)
132
for Hidden Markov Model estimation and analysis.
133
Network Estimation
134
To estimate networks, we used the HMM-MAR Toolbox (Vidaurre et al., 2017, 2018). El
135
estimation uses ROI time series data and calculates the K networks that best describe the entire
136
conjunto de datos. It then allocates each time window to the single best-fitting network within the original
137
time series. HMM, as a dimensionality reduction technique, returns states (hereafter referred to
138
as networks) that can be used to observe how networks interact over time.
139
140
The output from HMM is a time series showing the most prominent network at each timepoint.
141
From this timeseries, it is possible to calculate fractional occupancy and state-wise transitional
142
probabilidad (Vidaurre et al., 2017). Fractional occupancy is the proportion of the total number of
143
timepoints each network was occupied during a time series task, and shows a particular
144
network’s prominence during target time windows. Transitional probability shows the most
145
likely patterns of steps from one network to another. De este modo, both are related measures, but contain
146
different information about how the networks interact.
147
148
We estimated HMMs with variable K values between 3 y 20. We found the estimations with 4
149
y 7 states to provide the most optimal model-derived free energy values (see Vidaurre et al.,
150
2017; Vidaurre et al., 2018). Partial least squares analyses showed statistically significant effects
151
for both estimations with comparable effect sizes (see Fasano et al., 2022). We further
152
interrogated the spatial properties of the states in each estimation by computing a dot product of
153
the normalized state means, finding that the spatial properties of the states in the estimation with
7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
/
t
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
.
t
/
/
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
norte
mi
norte
_
a
_
0
0
3
3
3
pag
d
.
/
t
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
154
7 states were well-represented in the estimation with 4 estados. We ultimately chose the 4 estado
155
estimation as it provided a single state with activity in temporal and mesolimbic regions together.
156
Temporal and mesolimbic region activity has been previously related to auditory reward
157
(Salimpoor et al., 2011, Fasano et al., 2020), including prior analyses of subsets of the present
158
datos (Belden et al., 2023, Quinci et al., 2022).
159
160
The K networks identified by the HMM estimation are shown in Figure 1 (cortical regions only)
161
and the regions of interest are detailed in Table 1. Where this analysis did not use canonical
162
network-based seeds, we assigned anatomical labels to the networks based on the taxonomy of
163
functional brain networks consistent with the wider network literature (Uddin et al, 2019). El
164
functional properties of these states will be addressed in the discussion.
165
166
Cifra 1: Mean activity plots returned from HMM analysis. Colours represent relative activity of
167
the states and all have been normalized within-state. See Table 1 for subcortical regions not
168
displayed here.
169
Estado
Main Regions
Red
1
2
3
Bilateral middle-frontal and left temporal regions.
Subcortical regions include the bilateral temporal
pole, left nucleus accumbens, and right hippocampal
body
Medial frontoparietal network
Bilateral temporal and frontal regions
Temporal network
Bilateral temporal and mesolimbic regions
Subcortical regions include the left globus pallidus,
left hippocampal body, right putamen, y correcto
hippocampal tail
Temporal mesolimbic
network
8
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
t
/
/
mi
d
tu
norte
mi
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
d
oh
i
/
t
/
/
.
1
0
1
1
6
2
norte
mi
norte
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
norte
mi
norte
_
a
_
0
0
3
3
3
pag
d
.
t
/
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
7
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
4
Bilateral superior frontal and middle parietal regions Frontoparietal network
170
Mesa 1: Regions of interest and network labels from HMM analysis. Network labels are based
171
on the work of Uddin et al. (2019).
172
PLS
173
We used partial least squares (PLS) to analyze between- and within-group differences on the
174
HMM-extracted measures. PLS is a multivariate analysis technique that uses singular value
175
decomposition to quantify the relationship(s) between data matrices and experimental features, en
176
este caso, fractional occupancy and transitional probability measures. In these analyses, we used
177
mean-centred PLS to analyze group and task differences using the HMM-extracted measures and
178
the within-subject relation of the measure to participant liking and familiarity ratings. A
179
emphasize group main effects, we performed mean-centred analyses subtracting the overall
180
grand mean from the group means. To focus on task main effects and task by group interactions,
181
secondary mean centred analyses were performed, subtracting the group mean from the task
182
mean within each group (es decir., rendering the group main effect zero).
183
184
PLS analysis returns mutually-orthogonal latent variables (LVs) that describe group and/or task
185
efectos. Each LV’s statistical significance and reliability are calculated via permutation testing
186
and bootstrap estimation, respectively with a statistical threshold of p <.05. The reliability and
187
strength of the group or task effects is depicted through the confidence interval estimation of LV
188
scores for all participants, where LV scores are the dot-product of subject data and LV weights.
189
LV weights themselves are evaluated for reliability through bootstrap ratios of the weight
190
divided by its estimated standard error, which can be interpreted as a z-score for the
191
corresponding confidence interval (see McIntosh & Lobaugh, 2004).
9
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
192
193
Results
194
Prior to HMM decomposition, we tested for sex differences using mean-centred PLS on each
195
participant’s average functional connectivity matrix from the music listening task. No significant
196
sex-related differences were found. Following these analyses, we ran additional PLS analyses to
197
test for sex effects in fractional occupancy and transitional probability, returning no significant
198
effects. Data were subsequently pooled together for the remainder of the analysis.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
199
Fractional Occupancy
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
200
We extracted average fractional occupancy for each participant, and fractional occupancy for
f
/
d
o
i
/
.
/
/
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
201
each participant for each category of musical excerpt (self-selected, experimenter-selected
202
popular, and experimenter-selected novel) and used PLS to observe differences in fractional
203
occupancy across age groups and stimuli categories. Mean-centred PLS analysis returned one
204
significant LV (p = .024) showing an age effect, with younger adults showing higher fractional
205
occupancy in the temporal network (network/state 2), and older adults showing higher fractional
206
occupancy in the frontoparietal network (network/state 4).
207
208
Figure 2: Age-related differences in fractional occupancy (FO). (A) PLS contrasts between age
209
groups in music listening. Error bars were calculated using bootstrap resampling and reflect the
210
95% confidence interval. The contrasts show an age effect on FO (B), with the higher FO in
211
network 2 in younger adults, and higher FO in network 4 in older adults. The colour scale
212
represents the bootstrap ratio for each network.
213
10
214
When divided into stimulus categories and analyzed for task main effects and task-by-group
215
interactions, mean-centred PLS analysis returned one significant LV (p < .01, Figure 2) showing
216
an effect of self-selected music vs experimenter-selected music on fractional occupancy in the
217
temporal-mesolimbic network (network 3). Fractional occupancy is higher for this network while
218
listening to self-selected music (music that is highly familiar and well-liked) in both younger and
219
older adults. Fractional occupancy for the temporal network (network 2) is higher when listening
220
to experimenter-selected music. Both effects are qualitatively more reliable in younger adults
221
based on confidence intervals (Figure 3A).
222
223
Figure 3: (A) PLS contrasts between age groups in stimuli category and fractional occupancy
224
(FO). Error bars were calculated using bootstrap resampling and reflect the 95% confidence
225
interval. The contrasts show a stimulus-type effect on FO in both age groups (B), with the higher
226
FO in network 3 in both groups during self-selected music listening (SS Y and SS O), and higher
227
FO in network 2 during experimenter-selected music listening (Pop and Nov delineating popular
228
and novel excerpts respectively).
229
Transitional Probability
230
We next examined the transitional probability matrices for differences in network interaction on
231
average and between the different stimulus categories. Important to note: the data being analyzed
232
is the directional likelihood of transitioning from each network to each other network. Rather
233
than looking at networks by themselves, these results show the link or edge that connects each
234
network to each other network.
235
11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
.
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
236
The averaged transitional probability mean-centred PLS returned one significant LV (p < .001),
237
showing a contrast between younger and older adults, with younger adults more likely than older
238
adults to transition into the temporal network (network 2) from other networks, and less likely
239
than older adults to transition to the frontoparietal network (network 4) from the temporal
240
network (network 2) In examining network persistence (the likelihood of staying in a network),
241
younger adults were more likely to stay in the temporal network when listening to experimenter-
242
selected music (Figure 4).
243
244
Figure 4: (A) PLS contrasts between age groups and transitional probability (TP). Error bars
245
were calculated using bootstrap resampling and reflect the 95% confidence interval. The
246
contrasts show an age effect on TP in both age groups (B), with younger adults more likely to
247
transition into network 2 from networks 1, 2, and 3 than older adults; and less likely to transition
248
to network 4 from network 2 than older adults (C). The colour scale represents the bootstrap
249
ratio for each network.
250
251
When divided into stimulus categories and analyzed for task main effects and task-by-group
252
interactions, both groups were more likely to transition from the temporal network to the
253
temporal-mesolimbic and frontoparietal networks during self-selected music listening. In
254
experimenter-selected music, both groups were most likely to transition from the temporal-
255
mesolimbic network to the temporal network, but this effect was more pronounced in younger
256
adults. In examining network persistence (the likelihood of staying in a network), all participants
257
were more likely to stay in the temporal-mesolimbic network when listening to self-selected
258
music and more likely to stay in the temporal network when listening to experimenter-selected
12
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
.
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
259
music. When analyzed within age, older participants did not show a significant network
260
persistence pattern in the temporal network during experimenter-selected music (Figure 5).
261
262
Figure 5: (A) PLS contrasts between age groups in stimulus category and transitional
263
probabilities. SS refers to self-selected music, Pop and Nov refers to popular and novel
264
experimenter-selected music. Error bars were calculated using bootstrap resampling and reflect
265
the 95% confidence interval. The contrasts (B) show a stimulus-type effect on transitional
266
probability (TP), illustrated with the TP magnitude in panel C. Panel C shows the between-
267
network TP with solid lines representing self-selected music and dashed lines representing
268
experimenter-selected music.
269
Effects of liking and familiarity on brain measures
270
We next analyzed the network fractional occupancy and transitional probability matrices with
271
participants’ liking and familiarity ratings. We correlated liking and familiarity ratings for each
272
excerpt with fractional occupancy for each participant. Initial mean-centred PLS analysis
273
returned no significant LVs. Following this analysis, we ran the PLS centred to the overall grand
274
mean to allow for a full factorial analysis: group main effect, task main effect and group-by-task
275
interactions.
276
277
The results from the full factorial PLS returned one significant LV (p < .001) showing the
278
contrast between age groups. In younger adults, the temporal-mesolimbic network featured
279
prominently, showing a greater positive correlation than other networks with both liking and
280
familiarity. Older adults showed a more ambiguous correlation between liking and familiarity
281
and fractional occupancy in the temporal network (Figure 7).
13
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
/
/
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
282
283
Figure 6:(A) PLS contrasts between age groups in stimulus category and fractional occupancy.
284
Error bars were calculated using bootstrap resampling and reflect the 95% confidence interval.
285
The contrasts show an age effect on correlations between liking and familiarity (Fam) and
286
network fractional occupancy (B), illustrated with the relevant magnitude in panel C.
287
288
We next vectorized the excerpt-wise transitional probability matrices for each participant, and
289
correlated them with each participant’s piece-wise liking and familiarity ratings, returning two
290
transitional probability -correlation matrices per participant: liking*transitional probability and
291
familiarity*transitional probability.
292
293
A full factorial PLS consistent with the above analysis returned one significant LV (p < 0.001)
294
showing an age effect. Younger adults’ liking and familiarity ratings were more strongly
295
positively correlated with the likelihood of transitioning to the temporal-mesolimbic network
296
from the temporal and frontoparietal networks. Younger adults’ ratings were more strongly
297
negatively correlated with persistence in the temporal-mesolimbic network, and the likelihood of
298
transitioning from the temporal-mesolimbic network to the medial frontoparietal network.
299
Transitioning from the frontoparietal network to the temporal network was more positively
300
correlated with ratings in older adults, and more negatively correlated with ratings in younger
301
adults (Figure 7).
302
303
Figure 7:(A) PLS contrasts between age groups in stimulus category and transitional
304
probabilities. Error bars were calculated using bootstrap resampling and reflect the 95%
14
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
.
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
305
confidence interval. The contrasts (B) show an age effect on correlations between liking and
306
familiarity (Fam) and network transitional probability, illustrated with the relevant magnitude in
307
panel C. The colour scale represents the bootstrap ratio for each network.
308
Within-age mean-centred PLS analyses did not return any significant LVs.
309
Liking and familiarity behavioural ratings
310
Finally, we examined the ratings themselves. Mean-centred PLS showed older adults rated
311
excerpts as significantly less familiar than younger adults (p < .01). However, they did not
312
significantly differ in liking ratings. Mean-centred PLS also showed older adults’ liking and
313
familiarity data were significantly more highly correlated than younger adults (r = 0.57 for older
314
adults and r = 0.43 for younger adults, PLS p < .01).
315
Discussion
316
Music listening engages multiple brain networks that may reorganize in multiple ways as we age.
317
While there are well-documented effects of music listening on auditory and reward networks and
318
auditory-motor networks, less is known about how music listening may encourage persistence
319
within networks, or transitions between networks. Treating data-driven brain networks as units of
320
analysis, we detailed age-related similarities and differences in network occupancy and between-
321
network transitional probabilities during music listening. The two most commonly-featured
322
networks in these analyses were the temporal and temporal-mesolimbic networks. Activity in
323
temporal-mesolimbic regions overlaps with auditory-reward network activity (see Wang et al.,
324
2020), while temporal regions are firmly affiliated with auditory processing (Belfi & Loui,
325
2019).
15
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
/
/
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
326
327
Both younger and older adults showed the highest fractional occupancy in the temporal-
328
mesolimbic network while listening to self-selected music compared to experimenter-selected
329
music. These stimuli were selected by participants to be familiar and well-liked, and auditory-
330
reward network activation for preferred music has been well-documented in prior studies
331
(Salimpoor et al., 2011, Fasano et al., 2020), including on a subset of these data (Quinci et al.,
332
2022). This network was active for experimenter-selected music as well, though to a lesser extent
333
than self-selected music, particularly in younger adults.
334
335
When looking at the transitional probability matrices, self-selected music was again linked to
336
persistence in the temporal-mesolimbic network and a greater probability of transition to this
337
network from the temporal network in both age groups. Experimenter-selected music was linked
338
to higher persistence in the temporal network and a greater probability of transition to the
339
temporal network from the temporal-mesolimbic network in both age groups, indicating that
340
music listening employs a distributed network of frontal and temporal regions; but to engage
341
mesolimbic structures, a degree of liking and familiarity is needed.
342
343
However, when analyzed separately, group differences were more obvious. Older subjects
344
showed an increased likelihood of persistence in the temporal network during experimenter-
345
selected music, but this effect was less reliable than in younger adults. Older adults also showed
346
an increased likelihood of transitioning to the temporal-mesolimbic network from the medial
347
frontoparietal network in self-selected music. This network shares many regions with the default
348
mode network (DMN; Uddin et al., 2019). The DMN is implicated in listening to liked (Wilkins
16
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
.
/
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
349
et al., 2014; Pereira et al., 2011) and timbrally rich music (Alluri et al., 2012), and is less
350
attenuated during cognitive tasks with age (Rieck et al., 2017). One possible explanation is that
351
older adults are less likely to transition from the medial frontoparietal network to the temporal
352
network during music listening than younger adults, instead remaining in the medial
353
frontoparietal network until transitioning to the temporal-mesolimbic network while a younger
354
adult may transition from the medial frontotemporal network to the temporal network.
355
356
The older adult transitional probability matrices showed more transitions to the temporal-
357
mesolimbic network during experimenter-selected music, which could indicate an age-related
358
shift in between-network dynamics. Former pathways (in this case, the likelihood of transitioning
359
from an auditory reward network to an auditory perception network during unfamiliar music, or
360
staying in an auditory perception network during unfamiliar music) reconfigure in favour of
361
consistency across multiple types of music involving the temporal mesolimbic network. This is
362
consistent with earlier findings that network functional specificity declines in favour of a more
363
standard set of responses to multiple stimuli types (Rieck et al., 2020).
364
365
In younger adults, liking and familiarity ratings were correlated with fractional occupancy in the
366
temporal and temporal mesolimbic networks, with the temporal network most occupied when
367
familiarity and liking are low and the temporal mesolimbic network most occupied when
368
familiarity and liking are high. In older adults, correlations between fractional occupancy and
369
liking and familiarity ratings are more ambiguous, indicating a reconfiguration of network
370
engagement related to aging. Correlations between ratings and transitional probabilities were
371
consistent with this pattern: younger adults’ likelihood of transitioning into the temporal and
17
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
.
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
372
temporal-mesolimbic networks were more strongly correlated with liking and familiarity than
373
older adults who showed a more diffuse pattern.
374
375
Older adults showed high fractional occupancy in the temporal-mesolimbic network during all
376
music types. This difference could be because older adults show less differentiation between
377
liking and familiarity during novel music listening. If familiarity is lower among older adults, but
378
liking is consistent with younger adults, it is possible that older adults would engage a different
379
network response to music that is unfamiliar but liked. Liking and familiarity are more
380
positively correlated in older adults than younger adults, consistent with earlier findings on age-
381
related blunting of emotional intensity and liking (where stimuli are consistently rated as less
382
extremely pleasant and unpleasant. See Baird et al., 2020; Groarke & Hogan, 2019; Laukka &
383
Juslin, 2007).
384
385
While these results offer a promising look into capturing age-related changes in network-level
386
dynamics in naturalistic behaviours, there are several areas for further inquiry. To more fully
387
examine age, future studies could include a more continuous range of participants, particularly
388
those in middle adulthood to disambiguate age and cohort effects. While this study did not focus
389
on music and memory, future work could include a measure of music-related memory (see
390
Jakubowski & Eerola, 2022) to disambiguate group differences due to memory and lived
391
experience. The methods presented here were in effort to identify networks most relevant to this
392
dataset in a data-driven way. This approach, while advantageous in presenting nuanced
393
fluctuations in network membership, may prove challenging to reconcile with the canonical
394
network literature. Future work could employ both canonical and data-driven methods to directly
18
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
t
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
395
examine network membership and behaviour in an effort to link both methodological
396
approaches.
397
398
These observations could illustrate the broader pattern of the network dynamics of music
399
listening, and the age-related reorganization of these networks. For older adults, the temporal
400
network becomes less finely tuned to liking and familiarity, while the temporal mesolimbic
401
network remains active. There are several exciting implications of these findings. The first is in
402
studying naturalistic behaviours in “network space”: investigating the behaviours and
403
interactions of networks as behaviour unfolds. The need to understand the brain as a complex,
404
dynamic system, one that is continually adapting to its surroundings, has been the topic of much
405
discussion (see McIntosh & Jirsa, 2019; Calhoun et al., 2014). The brain is more than a
406
collection of regions and its emergent properties can be captured in fascinating detail using
407
music. Though the methods presented here are not unique to music, we also hope to present
408
music as a viable stimulus to interrogate higher cognition.
409
410
In the same way that the brain is not merely a collection of regions, music is more than a simple
411
collection of notes. It is ubiquitous in the human experience (Savage, 2019; Cross & Morley,
412
2010) but has yet to experience its renaissance in cognitive neuroscience. There are good reasons
413
for this: music data contain many layers of information from the content of the signal itself to the
414
content of the memories or the quality of movement it generates in the listener. However, the
415
scientific potential of music is too beguiling to ignore. Here is a stimulus that, unlike rest, has a
416
rich, externally-measurable temporal structure that, unlike traditional task paradigms, does not
417
require extensive training or fortitude to endure. It combines the best of both worlds with the
19
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
.
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
418
added benefit of being accessible to clinical populations in ways that other tasks, especially those
419
reliant on language, are not.
420
421
By examining music’s network properties, we present a data-driven methodological framework
422
for future hypothesis-driven studies of musical behaviour while offering an alternative to
423
traditional paradigms that is externally measurable, ecologically valid, and accessible to those
424
with cognitive decline or who are non-verbal.
425
References
426
Alluri, V., Toiviainen, P., Jääskeläinen, I. P., Glerean, E., Sams, M., & Brattico, E. (2012).
427
Large-scale brain networks emerge from dynamic processing of musical timbre, key and rhythm.
428
Neuroimage, 59(4), 3677-3689.
429
430
Baird, A., Brancatisano, O., Gelding, R., & Thompson, W. F. (2020). Music evoked
431
autobiographical memories in people with behavioural variant frontotemporal dementia.
432
Memory, 28(3), 323-336.
433
434
Belden, A., Quinci, M. A., Geddes, M., Donovan, N., Hanser, S. B., & Loui, P. (2023).
435
Functional Organization of Auditory and Reward Systems in Healthy Aging. bioRxiv.
436
437
Bones, O., & Plack, C. J. (2015). Losing the music: aging affects the perception and subcortical
438
neural representation of musical harmony. Journal of Neuroscience, 35(9), 4071-4080.
439
20
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
440
Belfi, A. M., & Loui, P. (2020). Musical anhedonia and rewards of music listening: Current
441
advances and a proposed model. Annals of the New York Academy of Sciences, 1464(1), 99-114.
442
443
Bressler, S. L., & Menon, V. (2010). Large-scale brain networks in cognition: emerging methods
444
and principles. Trends in cognitive sciences, 14(6), 277-290.
445
446
Burunat, I., Tsatsishvili, V., Brattico, E., & Toiviainen, P. (2017). Coupling of action-perception
447
brain networks during musical pulse processing: evidence from region-of-interest-based
448
independent component analysis. Frontiers in Human Neuroscience, 11, 230.
449
450
Calhoun, V. D., Miller, R., Pearlson, G., & Adalı, T. (2014). The chronnectome: time-varying
451
connectivity networks as the next frontier in fMRI data discovery. Neuron, 84(2), 262-274.
452
453
Cross, I., & Morley, I. R. M. (2010). The evolution of music: Theories, definitions and the nature
454
of the evidence. In Malloch, S. & Trevarthen, C. (Eds.), Communicative Musicality (pp.61-82).
455
Oxford University Press.
456
457
Cuddy, L. L., & Duffin, J. (2005). Music, memory, and Alzheimer’s disease: is music
458
recognition spared in dementia, and how can it be assessed?. Medical hypotheses, 64(2), 229-
459
235.
460
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
.
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
21
461
Fasano, M. C., Cabral, J., Stevner, A., Vuust, P., Cantou, P., Brattico, E., & Kringelbach, M. L.
462
(2023). The early adolescent brain on music: Analysis of functional dynamics reveals
463
engagement of orbitofrontal cortex reward system. Human Brain Mapping, 44(2), 429-446.
464
465
Frazier-Logue, N., Wang, J., Wang, Z., Sodums, D., Khosla, A., Samson, A. D., ... & Shen, K.
466
(2022). A robust modular automated neuroimaging pipeline for model inputs to
467
TheVirtualBrain. Frontiers in Neuroinformatics, 16, 883223.
468
469
Grady, C. (2012). The cognitive neuroscience of ageing. Nature Reviews Neuroscience, 13(7),
470
491-505.
471
472
Grady, C., Sarraf, S., Saverino, C., & Campbell, K. (2016). Age differences in the functional
473
interactions among the default, frontoparietal control, and dorsal attention networks.
474
Neurobiology of aging, 41, 159-172.
475
476
Groarke, J. M., & Hogan, M. J. (2019). Listening to self-chosen music regulates induced
477
negative affect for both younger and older adults. Plos one, 14(6), e0218017.
478
479
Jakubowski, K., & Eerola, T. (2022). Music evokes fewer but more positive autobiographical
480
memories than emotionally matched sound and word cues. Journal of Applied Research in
481
Memory and Cognition, 11(2), 272.
482
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
/
/
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
22
483
Koelsch, S., Andrews‐ Hanna, J. R., & Skouras, S. (2022). Tormenting thoughts: The posterior
484
cingulate sulcus of the default mode network regulates valence of thoughts and activity in the
485
brain's pain network during music listening. Human brain mapping, 43(2), 773-786.
486
487
Laukka, P., & Juslin, P. N. (2007). Similar patterns of age-related differences in emotion
488
recognition from speech and music. Motivation and Emotion, 31(3), 182-191.
489
490
Leggieri, M., Thaut, M. H., Fornazzari, L., Schweizer, T. A., Barfett, J., Munoz, D. G., &
491
Fischer, C. E. (2019). Music intervention approaches for Alzheimer’s disease: A review of the
492
literature. Frontiers in neuroscience, 13, 132.
493
494
Lima, C. F., & Castro, S. L. (2011). Emotion recognition in music changes across the adult life
495
span. Cognition and Emotion, 25(4), 585-598.
496
497
Matziorinis, A. M., & Koelsch, S. (2022). The promise of music therapy for Alzheimer's disease:
498
A review. Annals of the New York Academy of Sciences, 1516(1), 11-17.
499
500
McIntosh, A. R., & Jirsa, V. K. (2019). The hidden repertoire of brain dynamics and
501
dysfunction. Network Neuroscience, 3(4), 994-1008.
502
503
McIntosh, A. R., & Lobaugh, N. J. (2004). Partial least squares analysis of neuroimaging data:
504
applications and advances. Neuroimage, 23, S250-S263.
505
23
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
.
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
506
Quinci, M. A., Belden, A., Goutama, V., Gong, D., Hanser, S., Donovan, N. J., ... & Loui, P.
507
(2022). Longitudinal changes in auditory and reward systems following receptive music-based
508
intervention in older adults. Scientific Reports, 12(1), 11517.
509
510
Rieck, J. R., Baracchini, G., Nichol, D., Abdi, H., & Grady, C. L. (2021). Reconfiguration and
511
dedifferentiation of functional networks during cognitive control across the adult lifespan.
512
Neurobiology of Aging, 106, 80-94.
513
514
Rieck, J. R., Rodrigue, K. M., Boylan, M. A., & Kennedy, K. M. (2017). Age-related reduction
515
of BOLD modulation to cognitive difficulty predicts poorer task accuracy and poorer fluid
516
reasoning ability. Neuroimage, 147, 262-271.
517
518
Salimpoor, V. N., Benovoy, M., Larcher, K., Dagher, A., & Zatorre, R. J. (2011). Anatomically
519
distinct dopamine release during anticipation and experience of peak emotion to music. Nature
520
neuroscience, 14(2), 257-262.
521
522
Savage, P. E. (2019). Cultural evolution of music. Palgrave Communications, 5(1), 1-12.
523
524
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X.-N., Holmes, A. J., et al. (2018).
525
Local-Global Parcellation of the Human Cerebral Cortex from Intrinsic Functional Connectivity
526
MRI. Cerebral cortex, 28, 3095. doi:10.1093/CERCOR/BHX179.
527
24
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
528
Särkämö, T., Tervaniemi, M., Laitinen, S., Numminen, A., Kurki, M., Johnson, J. K., &
529
Rantanen, P. (2014). Cognitive, emotional, and social benefits of regular musical activities in
530
early dementia: randomized controlled study. The Gerontologist, 54(4), 634-650.
531
532
Taruffi, L., Pehrs, C., Skouras, S., & Koelsch, S. (2017). Effects of sad and happy music on
533
mind-wandering and the default mode network. Scientific reports, 7(1), 14396.
534
535
Thaut, M. H., Fischer, C. E., Leggieri, M., Vuong, V., Churchill, N. W., Fornazzari, L. R., &
536
Schweizer, T. A. (2020). Neural Basis of Long-term Musical Memory in Cognitively Impaired
537
Older Persons. Alzheimer Disease & Associated Disorders, 34(3), 267-271.
538
539
Tian, Y., Margulies, D. S., Breakspear, M., and Zalesky, A. (2020). Topographic organization of
540
the human subcortex unveiled with functional connectivity gradients. Nature Neuroscience, 23,
541
1421–1432. doi:10.1038/s41593-020-00711-6.
542
543
Uddin, L. Q., Yeo, B. T., & Spreng, R. N. (2019). Towards a universal taxonomy of macro-scale
544
functional human brain networks. Brain topography, 32(6), 926-942.
545
546
Vidaurre, D., Abeysuriya, R., Becker, R., Quinn, A. J., Alfaro-Almagro, F., Smith, S. M., &
547
Woolrich, M. W. (2018). Discovering dynamic brain networks from big data in rest and task.
548
Neuroimage, 180, 646-656.
549
25
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
.
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
550
Vidaurre, D., Smith, S. M., & Woolrich, M. W. (2017). Brain network dynamics are
551
hierarchically organized in time. Proceedings of the National Academy of Sciences, 114(48),
552
12827-12832.
553
554
Vieillard, S., Didierjean, A., & Maquestiaux, F. (2012). Changes in the perception and the
555
psychological structure of musical emotions with advancing age. Experimental Aging Research,
556
38(4), 422-441.
557
558
Wang, D., Belden, A., Hanser, S. B., Geddes, M. R., & Loui, P. (2020). Resting-state
559
connectivity of auditory and reward systems in Alzheimer’s disease and mild cognitive
560
impairment. Frontiers in Human Neuroscience, 14, 280.
561
562
Wilkins, R. W., Hodges, D. A., Laurienti, P. J., Steen, M., & Burdette, J. H. (2014). Network
563
science and the effects of music preference on functional brain connectivity: From Beethoven to
564
Eminem. Scientific reports, 4(1), 1-8.
565
566
Williams, J. A., Margulis, E. H., Nastase, S. A., Chen, J., Hasson, U., Norman, K. A., &
567
Baldassano, C. (2022). High-order areas and auditory cortex both represent the high-level event
568
structure of music. Journal of cognitive neuroscience, 34(4), 699-714.
569
570
Woolrich, M. W., Jbabdi, S., Patenaude, B., Chappell, M., Makni, S., Behrens, T., ... & Smith, S.
571
M. (2009). Bayesian analysis of neuroimaging data in FSL. Neuroimage, 45(1), S173-S186.
26
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
/
t
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
t
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
.
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
t
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
t
/
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
t
/
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
.
/
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
/
/
t
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
t
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
.
t
/
/
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Author Summary
This article explores age-related differences in between-network dynamics during music
listening using fMRI data collected from a sample of healthy younger and older adults. We
estimated brain networks using Hidden Markov Modelling (HMM) and tested for age- and
stimulus-related differences using Partial Least Squares (PLS). HMM returned four functional
connectivity networks, including a bilateral temporal network and a bilateral temporal-
mesolimbic network. We found differences related to age and stimulus with both age groups
spending more time in the temporal-mesolimbic network while listening to familiar, well-liked
music. Younger adults’ activity in this network was positively correlated with liking and
familiarity ratings, but this was not the case for older adults, consistent with past work on age-
related dedifferentiation. We conclude that activity in the temporal-mesolimbic network is robust
to dedifferentiation and discuss how these conclusions and analysis tools can be of use in future
work with clinical populations.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
d
o
i
/
/
/
t
.
1
0
1
1
6
2
n
e
n
_
a
_
0
0
3
3
3
2
1
5
5
2
0
2
n
e
n
_
a
_
0
0
3
3
3
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3