Why You Think Milan is Larger than Modena:
Neural Correlates of the Recognition Heuristic
Kirsten G. Volz1, Lael J. Schooler2, Ricarda I. Schubotz1,
Markus Raab3, Gerd Gigerenzer2, 和D. Yves von Cramon1
抽象的
& When ranking two alternatives by some criteria and only one
of the alternatives is recognized, participants overwhelmingly
adopt the strategy, termed the recognition heuristic (RH), 的
choosing the recognized alternative. Understanding the neural
correlates underlying decisions that follow the RH could help
determine whether people make judgments about the RH’s
applicability or simply choose the recognized alternative. 我们
measured brain activity by using functional magnetic resonance
imaging while participants indicated which of two cities they
thought was larger (实验 1) or which city they recognized
(实验 2). In Experiment 1, increased activation was ob-
served within the anterior frontomedian cortex (aFMC), precu-
neus, and retrosplenial cortex when participants followed the RH
compared to when they did not. 实验 2 revealed that RH
decisional processes cannot be reduced to recognition memory
流程. As the aFMC has previously been associated with self-
referential judgments, we conclude that RH decisional processes
involve an assessment about the applicability of the RH. &
介绍
We often need to rank two alternatives by some criteri-
在, 例如, which road to a city is faster. In one
experiment done to explore how people do such rank-
英格斯, Goldstein and Gigerenzer (2002) presented U.S.
students with pairs of large U.S. cities and with pairs of
large German cities. The task was to infer which city in
each pair had the larger population. The students
performed equally well on the German and U.S. 城市
对. How can this be that the students performed as
well with German cities as with the American cities,
when they knew almost nothing about the German
城市? Goldstein and Gigerenzer found that people
reliably employ a remarkably effective decision strategy
they dubbed the recognition heuristic (RH), which can
be stated as follows: ‘‘If one of two objects is recognized
and the other is not, then infer that the recognized
object has the higher value with respect to the criterion’’
(p. 76). According to Goldstein and Gigerenzer, 这
students’
imperfect recognition of the German cities
allowed them to frequently apply the RH, which led
them to pick the recognized German cities, which tend
to be larger than those that are unrecognized. 这
students could not use this heuristic when comparing
我们. 城市, 尽管, because they recognized all of them.
The RH works only in domains, or environments, 在
which recognition is correlated—in either direction—
1马克斯·普朗克人类认知和脑科学研究所,
德国, 2Max Planck Institute for Human Development, 格尔-
许多, 3University of Flensburg, 德国
with the inaccessible criterion being predicted, 对于前任-
充足, between city name recognition and population
尺寸. To the extent that the RH functions well by
exploiting characteristics of an environment, it is eco-
logically rational with respect to that particular environ-
蒙特. Goldstein and Gigerenzer (2002) argue the
effectiveness of the RH in the city judgment task de-
pends on three correlations: Population size is reflected
by how often a city is mentioned in the media, which in
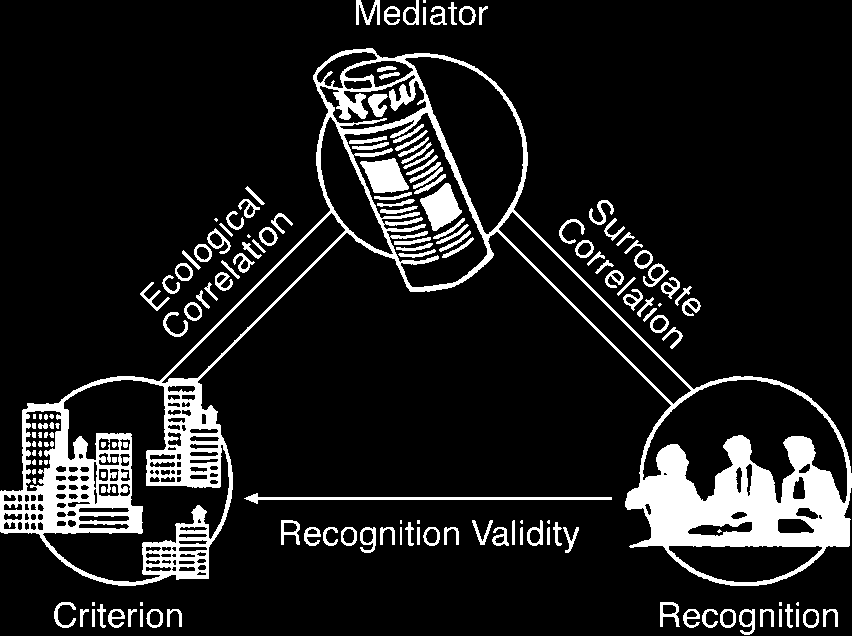
turn influences the probability of recognition (数字 1).
因此, recognition information correlates with and can
be used to infer the inaccessible criterion. 因此,
the effective use of the RH depends on the outcome
of two processes. The first yields whether the alterna-
tives are recognized and so whether the RH can be
applied. The second process assesses whether the RH
should be applied, and so is essentially a judgment
about its ecological rationality. The judgment of eco-
logical rationality could, 例如, depend on an
assessment of whether the chain of correlations linking
the criterion to recognition through the mediator (例如,
the frequency of city name in the media) justifies fol-
lowing the RH.
然而, it is not necessarily the case that subjects have to
judge the ecological rationality of the RH before apply-
理解它. 反而, subjects could simply choose the recog-
nized alternative without considering why recognition
should be predictive of population size. Such a strategy
would be successful here because recognition is so
highly correlated with how often a stimulus occurs in
环境.
D 2006 麻省理工学院
认知神经科学杂志 18:11, PP. 1924–1936
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
t
w
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
n
t
我
C
/
A
e
r
–
p
t
d
我
C
1
我
8
e
1
–
1
p
1
d
9
F
2
/
4
1
8
1
/
9
1
3
5
1
6
/
1
1
7
9
2
哦
4
C
/
n
1
2
7
0
5
0
5
6
9
1
8
8
7
/
1
j
1
哦
1
C
9
n
2
.
4
2
p
0
d
0
6
乙
.
y
1
G
8
你
.
e
1
s
1
t
.
哦
1
n
9
0
2
8
4
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
.
.
.
F
哦
n
1
8
中号
A
y
2
0
2
1
why there is only little support of a sharp distinction of
labor within the MTL. Besides these cortices, 顶叶
areas have consistently been shown to be crucial for
remembering, particularly for episodic retrieval process-
英语 (瓦格纳, Shannon, Kahn, & 巴克纳, 2005). 为了
例子, (lateral and medial) parietal areas have been
shown to increase their activity to recognized old items
as well as to erroneously recognized new items and to
correlate with the subjective experience of remember-
英. 因此, if RH-based decisional processes in the
present study were not only premised on recognition
information but rather depend on additional judgments
of ecological rationality, we expected regions beyond
MTL regions and parietal areas to be activated when
participants decided according to the RH.
因此, by using functional magnetic resonance imaging
(功能磁共振成像) and an adapted version of the two-alternative
forced-choice task used by Goldstein and Gigerenzer
(2002), we contrasted the neural correlates of decisions
based on the RH to those in which the heuristic was not
applied. 然而, finding activation in areas besides those
regions known to reflect recognition memory processes
would not definitively rule out an alternative interpreta-
的, 即, that those areas reflect decisional process-
es that simply opt for recognized alternatives. 为了这
原因, we ran the same experiment again with a
different group of participants and with the instruction
to simply choose the recognized city in each pair
(实验 2). The test of city recognition and the
paired comparison test were run as a between-subjects
design to avoid potential carryover effects. In Experi-
蒙特 2, participants were asked to execute the task
‘‘Which city do you know?’’ instead of ‘‘Which city is
larger?’’ By contrasting trials in which participants were
instructed to choose the recognized city with trials in
which they were instructed to choose the unrecognized
城市, we built a contrast parallel to the RH contrast from
实验 1. 在这样做, we could compare the acti-
vation patterns of both experiments. 在某种程度上
the RH-based decisional processes (1) can be adequately
explained by participants simply choosing the recog-
nized alternative and (2) do not depend on a judgment
of the ecological rationality, the activation patterns of
the two contrasts should not differ with respect to the
critical activations.
方法
实验 1
参加者
Eighteen (10 女性, mean age 25.6 年, 标清 3.4,
range 20–32 years) right-handed, healthy volunteers
participated in the fMRI experiment. Informed consent
was obtained prior to the experiment from each partic-
ipant according to the Declaration of Helsinki. Experi-
mental standards were approved by the local ethics
Volz et al.
1925
数字 1. The ecological rationality of the recognition heuristic
(RH), 那是, the ability to exploit the structure of the information
in the environment: An inaccessible criterion (例如, 人口
size of a certain city) is reflected but not revealed by a mediator
variable (例如, the number of times a city is mentioned in the
媒体), and the mediator influences the probability of recognition.
The recognition information is then used to infer the criterion.
This inference is successful only if recognition is correlated with
the criterion. Adapted from Goldstein and Gigerenzer (2002).
Understanding the neural correlates underlying the
use of the RH could help determine whether RH-based
decisional processes depend on additional judgments of
ecological rationality. Such judgments of ecological ra-
tionality should draw on brain areas beyond those
known to reflect recognition memory processes.
Some have concluded from behavioral studies that
declarative memory (episodic memory [事件] 和
semantic memory [facts]; 比照. Squire, 1994) is supported
by at least two distinct processes: The assessment of
what has been termed item familiarity and the recollec-
tion of the context in which an item was encountered
前 (Yonelinas et al., 2002). Neuropsychological as
well as neuroscientific evidence support the proposed
dual-process framework in that recollection and famil-
iarity draw on distinct neural mechanisms: Whereas the
hippocampus is considered to play a crucial role in
recollection (例如, Aggleton et al., 2005; Ranganath
等人。, 2003; Yonelinas et al., 2002; Aggleton & Shaw,
1996; for an overview, see Rugg & Yonelinas, 2003),
adjacent medial temporal lobe (MTL) 地区 (perirhinal
cortex) are hypothesized to reflect familiarity-based
认出 (Yonelinas et al., 2002). 然而, a recent review
on the MTL outlines that the available data may not
support such a simple dichotomy (Squire, Stark, &
克拉克, 2004). 相当, the authors suggest that the various
anatomical components of the MTL all signal informa-
tion that is significant to successful recognition memory
表现: The hippocampal formation provides a
conjunctive recognition signal, whereas adjacent corti-
ces provide stimulus-specific (认出) signals. 两个都
components are considered necessary for intact recog-
nition memory performance and hence may account for
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
t
w
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
n
t
我
C
/
A
e
r
–
p
t
d
我
C
1
我
8
e
1
–
1
p
1
d
9
F
2
/
4
1
8
1
/
9
1
3
5
1
6
/
1
1
7
9
2
哦
4
C
/
n
1
2
7
0
5
0
5
6
9
1
8
8
7
/
1
j
1
哦
1
C
9
n
2
.
4
2
p
0
d
0
6
乙
.
y
1
G
8
你
.
e
1
s
1
t
.
哦
1
n
9
0
2
8
4
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
F
.
.
.
.
.
哦
n
1
8
中号
A
y
2
0
2
1
committee of the University of Leipzig. Data were han-
dled anonymously.
Stimuli, 任务, and Experimental Session
Stimuli consisted of two concurrently presented words,
one on the right and one on the left side of the screen
(horizontal visual angle, 118; vertical visual angle, 1.78).
Participants had their index fingers on a left and a right
response button, spatially corresponding to the stimulus
locations on the screen. Within each trial a cue was
presented for 500 msec signaling that the next trial was
about to start, followed by a fixation cross presented for
500 毫秒. 随后, word pairs were presented for
a maximum of 4 sec during which participants’ response
was recorded. As soon as participants indicated their
choice by a button press, the stimuli disappeared and
were replaced by a fixation cross. No performance
feedback was delivered whatsoever. The participants’
task was to indicate which city in each pair had the
larger population.
An experimental session consisted of 140 实验-
tal trials and 30 null events, in which no stimulus was
presented and so the blood oxygenation level depen-
凹痕 (大胆的) response was allowed to return to a
baseline state. All trials lasted for 8 sec each (IE。, 四
scans of repetition time [TR] 2 秒). To allow for
measurements to be taken at numerous time points
along the BOLD signal curve, the onset of each stimulus
presentation relative to the beginning of the first of the
four scans was varied randomly in four time steps (0,
500, 1000, 和 1500 毫秒). The purpose of this proce-
dure was to enhance the temporal resolution of the
image acquisition (Birn, 考克斯, & Bandettini, 2002; 米津,
Maccotta, Ollinger, 彼得森, & 巴克纳, 2000).
After the fMRI session, a recognition test was admin-
istered in which participants had to indicate whether
or not they knew each city from before the experi-
mental session. It was emphasized to the participants
that they should declare as recognized only those cities
that they had heard of before the functional session.
These data were used to determine individual trial types,
那是, whether both, none, or only one of the cities
were recognized. 因此, we could individually de-
termine RR (recognized–recognized), RU (recognized–
unrecognized), and UU (unrecognized–unrecognized)
试验. 此外, the data of the recognition test were
also used to assess in which RU trials participants
decided in favor of the recognized city, 那是, 这
application of the RH. After the recognition test, 参与-
ipants were asked to fill out a questionnaire asking
wherefrom they knew most of the cities; 随后,
they were debriefed and thanked.
Our experimental design called for the city compari-
son city task to always come first, followed by the
recognition test, rather than counterbalancing the task
命令. Having the recognition test before the city task
could have biased participants towards using the RH
by making salient to the participants that we (这
experimenters) were interested in whether or not they
recognized the stimuli. 此外, by keeping a fixed
task order, we were able to reliably measure the hemo-
dynamic response elicited by recognition judgments that
are presumed to underlie the RH. 那是, 判断
about whether the city can be recognized from their
daily life, and so can be used to make inferences about
the cities. Had the recognition task come first, 这
participants would have had to judge not only whether
they recognized the city, but also whether the source of
the recognition was just from the experiment or possibly
from elsewhere. The additional demands of this discrim-
ination task means that the recognition judgments from
the two task orders could draw on somewhat different
brain structures and so involve brain structures that
would not otherwise be involved in the application of
the RH. 尽管如此, we expect that in the recognition
任务, participants could reliably report whether they
saw the city from the experiment or from elsewhere.
那是, they would rarely miscategorize as recognized a
city that they had only encountered in the experiment.
Support for this come from the studies of Pohl (2006)
and Goldstein and Gigerenzer (2002). Neither study
found any difference in the recognition rates that de-
pended on the order of the two tasks.
Pilot Study for Stimulus Material Preparation
A key design goal of the present study was that each city
in the experiment should only be seen once, so that par-
ticipant’s recognition judgments would not be contami-
nated by their experience within the experiment. 为了这
原因, the city task always preceded the recognition test.
To construct the trials so that subjects would be faced
with approximately equal proportions of RR, RU, 和
UU trials, we ran a behavioral pilot study to obtain aver-
age recognition rates for 400 cities necessary to make
140 experimental trials. In this pilot study, 管理的
as a paper-and-pencil task, 60 students from Berlin and
Flensburg were asked to answer the following question:
Please indicate if you have heard the name of this city be-
fore. Students answered this question for 400 cities from
following countries: 阿根廷, 巴西, 加拿大, 中国,
法国, 大不列颠, Holland, 印度, 伊拉克, 意大利, 日本,
波兰, Portugal, 俄罗斯, 西班牙, 南非, 瑞典,
瑞士, 和美国. The cities were
blocked by country and both the order of the country
blocks and the cities within each country block were ran-
domized. As a complete counterbalancing of the country
blocks would result in 20! (= 2.4329E+18) different or-
德斯, to keep things simple, one ordering of stimuli was
used for all participants. Countries with a larger number
of cities potentially known by a German population had
a higher proportion of cities on the list than countries
that we expected to be less familiar (例如, 美国
1926
认知神经科学杂志
体积 18, 数字 11
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
t
w
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
n
t
我
C
/
A
e
r
–
p
t
d
我
C
1
我
8
e
1
–
1
p
1
d
9
F
2
/
4
1
8
1
/
9
1
3
5
1
6
/
1
1
7
9
2
哦
4
C
/
n
1
2
7
0
5
0
5
6
9
1
8
8
7
/
1
j
1
哦
1
C
9
n
2
.
4
2
p
0
d
0
6
乙
.
y
1
G
8
你
.
e
1
s
1
t
.
哦
1
n
9
0
2
8
4
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
.
.
.
F
哦
n
1
8
中号
A
y
2
0
2
1
与. 中国). Based on the pilot study England, 法国,
Holland, 意大利, 加拿大, 西班牙, and the United States were
the only countries with approximately equal proportions
of recognized and unrecognized cities.
For the paired comparison task a new set of stimuli
was generated for each subject as follows. Pairs were
generated for each country separately. From those cities
that had yet to be included in the experiment, all possi-
ble pairs of cities from the same country were generated.
For each pair, the expected proportion of participants
who would recognize both, just one or neither city was
estimated based on the recognition rates of the Flensburg
and Berlin participants. 拿, 例如, the English
cities of Sheffield and Poole, with respective recognition
的比率 .95 和 .1, which we refer to, 分别, 作为
a and b. By making the strong assumption that these
recognition judgments are independent, we can estimate
that the probability that the average participant will rec-
ognize both cites is .095 (= a (西德:1) 乙), that the probabil-
ity that only one alternative will be recognized is .86
(= a[1 (西德:2) 乙] + [1 (西德:2) A] (西德:1) 乙) and that the probability that
neither city will be recognized is .045 (= [1 (西德:2) A][1 (西德:2) 乙]).
下一个, the city pair that would do the most to equalize the
proportion of RR, RU, and UU trials was selected. 这
process was repeated for the remaining cities.
After the functional session, we compared the recog-
nition rates of all 280 cities between the Berlin and
Flensburg participants (60) on the one hand and the
Leipzig participants (18) on the other and found recog-
nition values were highly correlated (r = .91; p < .0001).
Furthermore, the distribution of RR, RU, and UU trials
was not significantly different from an equal distribu-
tion (Kolmogorov–Smirnov test: Z = 1.10; p = .18), as
was intended.
Experiment 2
Participants
Fourteen (8 women, mean age 25.8 years, SD 2.5, range
22–30 years) right-handed, healthy volunteers participated
in the fMRI experiment. Informed consent was obtained
prior to the experiment from each participant according
to the Declaration of Helsinki. Experimental standards
were approved by the local ethics committee of the Uni-
versity of Leipzig. Data were handled anonymously.
Stimuli, Task, and Experimental Session
Experiment 2 differed from Experiment 1 in the follow-
ing points: In the functional session, participants had to
perform two tasks that were indicated by different color
cues; in addition, the city pairs were also presented in
the color corresponding to the indicated task. In one
task (respond–recognize), participants had to indicate
which city they recognized by pressing with their index
finger that key spatially corresponding to the stimulus
locations on the screen. In those cases when partici-
pants recognized both cities, they were instructed to
indicate this with their right middle finger, whereas in
cases when they recognized none of the cities, they were
instructed to indicate this with their left middle finger.
The mappings for the middle fingers were counterbal-
anced between participants. In another task (respond–
unrecognized) that was signaled by a color cue, partic-
ipants were required, in case they recognized only one
city, to indicate which one they did not recognize. The
response had to be made with the index fingers spatially
corresponding to the stimulus locations on the screen of
the unrecognized city. In case both or none of the cities
were recognized, the assignment for the keys stayed the
same as in the respond–recognize task, that is, right and
left middle fingers. Across the entire experiment, the
two tasks were never performed on the same stimuli.
Respond–recognize and respond–unrecognized trials
were intermixed and presented in random order. One
quarter of the trials assigned as RU trials were catego-
rized as such. As in Experiment 1, the actual number of
RU trials was individually different depending on the
knowledge of the participants. Parallel to Experiment 1,
one functional session consisted of 140 experimental
trials and 30 null events lasting for 8 sec each. In
addition, the procedure of the oversampling was kept
identical to that of Experiment 1.
MRI Scanning Procedure
Imaging
Imaging was performed on a 3T scanner (Siemens TRIO,
Erlangen, Germany). Twenty-two axial slices (4 mm
thickness, 20% spacing, field of view [FOV] 19.2 cm,
data matrix of 64 (cid:1) 64 voxels, and in-plane resolution of
3 mm (cid:1) 3 mm) parallel to the bicommissural plane (AC–
PC) covering the whole brain were acquired using
a single-shot echo-planar imaging (EPI) sequence (TR
2 sec, echo time [TE] 30 msec, flip angle 908). One
functional run with 872 time points was run with each
time point sampling over the 22 slices. Prior to func-
tional runs, 22 anatomical T1-weighted modified driven
equilibrium Fourier transform (MDEFT; Norris, 2000;
Ugurbil et al., 1993) images (data matrix 256 (cid:1) 256,
TR 1.3 sec, TE 10 msec) were acquired as well as 22
T1-weighted EPI images with the same spatial orienta-
tion as the functional data. The latter were used to co-
register the functional scans with previously acquired
high-resolution full-brain 3-D brain scans.
Data Analysis
The MRI data were processed by using the software
package LIPSIA (Lohmann et al., 2001). Functional data
were motion-corrected off-line with the Siemens motion-
correction protocol. To correct for the temporal offset
Volz et al.
1927
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
t
w
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
n
t
i
c
/
a
e
r
-
p
t
d
i
c
1
l
8
e
1
-
1
p
1
d
9
f
2
/
4
1
8
1
/
9
1
3
5
1
6
/
1
1
7
9
2
o
4
c
/
n
1
2
7
0
5
0
5
6
9
1
8
8
7
/
1
j
1
o
1
c
9
n
2
.
4
2
p
0
d
0
6
b
.
y
1
g
8
u
.
e
1
s
1
t
.
o
1
n
9
0
2
8
4
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
f
.
.
.
o
n
1
8
M
a
y
2
0
2
1
between the slices acquired in one scan, a cubic spline
interpolation was applied. A temporal high-pass filter
with a cutoff frequency of 1/160 Hz was used for base-
line correction of the signal and a spatial Gaussian filter
with 5.65-mm full width half maximum (FWHM) was ap-
plied. The anatomical slices were coregistered with
the high-resolution full-brain scan that resided in the
stereotactic coordinate system and then transformed by
linear scaling to a standard size (Talairach & Tournoux,
1988). The transformation parameters obtained from
this step were subsequently applied to the preprocessed
functional slices so that the functional slices were also
registered into the stereotactic space. This linear nor-
malization process was improved by a subsequent pro-
cessing step that performed an additional nonlinear
normalization known as ‘‘demon matching.’’ In this
type of nonlinear normalization, an anatomical 3-D data
set (i.e., the model) is deformed such that it matches
another 3-D anatomical data set (i.e., the source) that
serves as a fixed reference image (Thirion, 1998). The
voxel size was interpolated during the coregistration
from 3 (cid:1) 3 (cid:1) 4 mm to 3 (cid:1) 3 (cid:1) 3 mm. The statistical
evaluation was based on a least-squares estimation
using the general
linear model for serially autocorre-
lated observations (random effects model; Friston, Frith,
Turner, & Frackowiak, 1995; Worsely & Friston, 1995).
The general linear regression performs a ‘‘precoloring’’
of the data; that is,
it applies a temporal Gaussian
smoothing with a user-specified kernel width given by
the parameter FWHM. The smoothing imposes a tem-
poral autocorrelation that determines the degrees of
freedom. An event-related design was implemented;
that is, the hemodynamic response function was mod-
eled by means of the experimental conditions for each
stimulus (event = onset of stimulus presentation). The
design matrix was generated using a synthetic hemody-
namic response function and its first and second deriv-
ative (Friston et al., 1998) and a response delay of 6 sec.
The model equation, including the observation data, the
design matrix, and the error term, was convolved with
a Gaussian kernel of dispersion of 4 sec FWHM to deal
with the temporal autocorrelation (Worsley & Friston,
Table 1. Behavioral Results of Experiment 1
1995). Contrast images, that is, estimates of the raw score
differences between specified conditions, were gener-
ated for each subject. The single-subject contrast images
entered into a second-level random effects analysis for
each of the contrasts. The group analysis consisted of a
one-sample t test across the contrast images of all sub-
jects that indicated whether observed differences be-
tween conditions were significantly different from zero.
Subsequently, t values were transformed into Z scores.
Group statistical parametric maps were thresholded at
Z > 3.09 ( p = .001, uncorrected). Only clusters of at
least six connected voxels (IE。, 162 mm3) were reported
to ensure an overall imagewise false-positive rate of 5%
(Forman et al., 1995). This nonarbitrary voxel cluster size
was determined by using the program AlphaSim (afni.
nimh.nih.gov/afni/doc/manual/AlphaSim by Ward, 2000).
结果
实验 1
Behavioral Results
Reaction times (RT) of all RR, RU, and UU trials as well
as of correctly answered trials are shown in Table 1. 在
average, participants were faced with 55 (± 13) RR, 46
(± 5) RU, 和 39 (± 15) UU trials during an experimen-
tal session. A repeated measures analysis of variance of
all trials as well as of correctly answered trials revealed
the three experimental conditions to differ significantly
with regard to RT: [all trials: F(2,16) = 21.93; p = .001;
correctly answered trials: F(2,16) = 23.76; p = .001].
同样地, the three experimental conditions differed
significantly with regard to the rate of correct responses,
F(2,16) = 15.83; p = .001. Kolmogorov–Smirnov tests
indicate that the means of the subjects’ RTs as well as
the rate of correct responses were normally distributed
(all Z values > .478). As the standard errors for all
conditions were less 2.7%, it is apparent that perform-
ance in the RR and RU condition differed significantly
from the chance level of 50%, whereas performance in
the UU condition was at 50.1%. Regarding the use of the
RH, participants applied the heuristic in 84% 的
Condition
RR
UU
RU
RU(右)
RU(U)
RTall
2818
3024
2804
2781
3042
SE(RTall)
RTcorr
SE(RTcorr)
% 正确的
SE(%corr)
92
108
89
90
116
2815
3074
2767
2735
3064
92
116
84
80
159
63.2
50.1
69.2
73.5
44.2
1.9
1.9
1.5
1.5
3.8
Reaction times (RT) of all experimental trials as well as for correctly answered trials and the rates of correct responses (each with standard error
[SE]) are shown for condition RR (both cities recognized), RU (one city recognized), and UU (no city recognized). The last two rows show RTs and
rates of correct responses for RU trials in which participants decided in favor of the recognized alternative (RU(右)) and in favor of the unrecognized
选择 (RU(U)).
1928
认知神经科学杂志
体积 18, 数字 11
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
t
w
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
n
t
我
C
/
A
e
r
–
p
t
d
我
C
1
我
8
e
1
–
1
p
1
d
9
F
2
/
4
1
8
1
/
9
1
3
5
1
6
/
1
1
7
9
2
哦
4
C
/
n
1
2
7
0
5
0
5
6
9
1
8
8
7
/
1
j
1
哦
1
C
9
n
2
.
4
2
p
0
d
0
6
乙
.
y
1
G
8
你
.
e
1
s
1
t
.
哦
1
n
9
0
2
8
4
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
.
F
.
.
哦
n
1
8
中号
A
y
2
0
2
1
案例. Whether reliance on the RH pays is indicated by
the strength of the relationship between recognition
and criterion, which is referred to as recognition validity
in a specific environment or data set. In the present
实验, the recognition validity averaged .63, 哪个
is calculated as the proportion of times a recognized city
was indeed larger than an unrecognized city. 然而, as no
performance feedback was given, participants could not
determine the recognition validity. 合在一起, 这
present results are consistent with previous results
(戈德斯坦 & Gigerenzer, 2002).
After the experimental session, participants were
asked wherefrom they knew most of the cities. 它是
revealed that more than half of the recognized cities
were known from the media, such as newspapers, 消息
on television or the Internet, and novels (63%). As other
important sources, participants indicated schooling
(17%) and holiday (10%).
MRI Results
一般来说, to control for the RT differences between the
状况 (比照. Behavioral Results), we included RT
parameters for each condition as covariates. This covari-
ation analysis is capable of removing extraneous variabil-
ity that derives from RT differences between conditions.
To test for the specific neural correlates of RH-based
decisions we investigated the hemodynamic response
elicited by all RU trials in which the RH was applied as in
contrast to all those (RU) trials in which this was not the
案件 (all trials of each condition were included regard-
less of correctness so as to reliably estimate the beta
价值观). The analysis revealed a significantly higher
hemodynamic activity for decisions based on the RH
within the anterior frontomedian cortex (aFMC) 和
precuneus bilaterally extending into the retrosplenial
cortex (数字 2, 桌子 2). No brain region showed
significant activation for the inverse contrast. 结果
remained even if the number of RH trials was randomly
and individually adjusted to match the number of non-
RH trials (数字 2, 桌子 2). 例如, if a participant
chose in a specific number of RU trials, 说 20 试验, 这
unrecognized alternative, we would have included in
该模型 20 randomly chosen RU trials in which the
participant selected the recognized alternative.
To investigate which brain areas were involved with
successful recognition, RR trials were contrasted with
UU trials. Significant activation was revealed bilaterally
within the midportion of the parahippocampal gyrus,
the ventral striatum extending into the caudate nucleus,
the retrosplenial cortex extending into the medial pari-
etal cortex, the intraparietal sulcus, within the posterior
frontomedian cortex (pFMC), ventromedial prefrontal
cortex (VMPFC), and left inferior prefrontal cortex
(LIPFC; 数字 2, 桌子 2).
To identify regions commonly activated by RH-based
decisional processes and recognition memory processes,
we calculated a conjunction analysis, 那是, a test for a
logical AND (Nichols, Brett, 安德森, Wager, & 波琳娜,
2005). A positive conjunction test implies those regions
that are commonly activated across the two kinds of
认知过程. According to Nichols et al. (2005) A
correct test for a logical AND requires that all the
comparisons in the conjunction are individually signifi-
cant at a specified alpha rate. 因此,
if a voxel
showed a Z score less than the critical Z value of 3.09
then the associated voxel in the other Z map was set to
零. 那是, if either the map of the RH contrast or the
map of the recognition–memory contrast contained a
零, then the conjunction was false. The resulting
conjunction map reveals the intersection of the two
statistical maps thresholded at a specific alpha rate and
hence reliably displays those regions where there is an
effect in both maps. In the present study, results of the
conjunction analysis revealed significant activation only
within two areas: within the retrosplenial cortex and
precuneus (数字 2, 桌子 2). This finding suggests the
retrosplenial cortex activation and precuneus activation
but not aFMC activation during RH-based decisions to
reflect recognition and retrieval processes.
To test whether medial parietal cortex activation can
indeed be considered as an index of the strength of the
recognition signal, we calculated a parametric analysis
(post hoc) by including a covariate reflecting the average
recognition signal per trial, 那是, the mean recognition
value of the two presented cities. The recognition value
for each city in turn was calculated as the percentage of
participants in Experiment 1 that recognized it. When test-
ing for the effect of an increasing recognition strength,
the same activation pattern as for the recognition mem-
ory contrast was revealed: activation revealed bilaterally
within the midportion of the parahippocampal gyrus,
ventral striatum, retrosplenial cortex extending into the
medial parietal cortex,
intraparietal sulcus bilaterally,
within the pFMC, VMPFC, and LIPFC. 因此, medial
parietal cortex activation that was revealed when partic-
ipants applied the RH as compared when they did not is
taken to reflect the strength of the recognition signal.
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
t
w
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
n
t
我
C
/
A
e
r
–
p
t
d
我
C
1
我
8
e
1
–
1
p
1
d
9
F
2
/
4
1
8
1
/
9
1
3
5
1
6
/
1
1
7
9
2
哦
4
C
/
n
1
2
7
0
5
0
5
6
9
1
8
8
7
/
1
j
1
哦
1
C
9
n
2
.
4
2
p
0
d
0
6
乙
.
y
1
G
8
你
.
e
1
s
1
t
.
哦
1
n
9
0
2
8
4
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
F
.
.
.
哦
n
1
8
中号
A
y
2
0
2
1
实验 2
Behavioral Results
Behavioral results of Experiment 2 are shown in Table 3:
RT of the four experimental conditions (trials in which
participants recognized both cities, 两者都不, or one
city and had to choose the recognized city or the un-
recognized city) differed significantly in that partici-
pants were faster on trials in which they recognized
neither of the two presented cities, F(3,11) = 2.98;
p = .04 (桌子 3); a Kolmogorov–Smirnov test indicates
that the means of the subjects RTs were normally dis-
tributed (all Z values > .527). Regarding the RU trials,
participants were only 156 msec faster when choosing
Volz et al.
1929
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
t
w
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
n
t
我
C
/
A
e
r
–
p
t
d
我
C
1
我
8
e
1
–
1
p
1
d
9
F
2
/
4
1
8
1
/
9
1
3
5
1
6
/
1
1
7
9
2
哦
4
C
/
n
1
2
7
0
5
0
5
6
9
1
8
8
7
/
1
j
1
哦
1
C
9
n
2
.
4
2
p
0
d
0
6
乙
.
y
1
G
8
你
.
e
1
s
1
t
.
哦
1
n
9
0
2
8
4
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
.
F
.
.
哦
n
1
8
中号
A
y
2
0
2
1
数字 2. fMRI results of Experiments 1 和 2. Top: Results are shown for the effects of decisional processes based on the RH in
实验 1 (对比: RU trials in which the RH was applied > RU trials in which the RH was not applied) and effects of decisional
processes solely based on recognition in Experiment 2 (对比: RU trials in which the recognized alternative was chosen > RU trials
in which the unrecognized alternative was chosen). 中间: The effects of recognition memory processes are shown for both experiments
(contrasts: RR trials > UU trials). Bottom: 左边, results of the conjunction analysis of the two pictured contrasts in Experiment 1; 正确的,
result of Contrast 1 when trial numbers were adjusted.
1930
认知神经科学杂志
体积 18, 数字 11
桌子 2. 实验 1
桌子 3. Behavioral Results of Experiment 2
RT
3114
3015
3121
3277
SE (RT)
69
86
111
70
Area
X
y
z
Z
Condition
RR
UU
RU(右)
RU(U)
RH-based decisional processes
aFMC
Retrosplenial cortex
Medial parietal cortex
(西德:2)5
54
6 4.6
4 (西德:2)48
15 3.8
(西德:2)17 (西德:2)62
24 3.9
10 (西德:2)62
27 4.1
Recognition memory processes
Midportion of the parahippocampal
(西德:2)23 (西德:2)36 (西德:2)6 4.4
gyrus
28 (西德:2)35 (西德:2)6 4.4
Precuneus extending into retrosplenial
(西德:2)11 (西德:2)54
15 4.4
4 (西德:2)53
12 4.7
(西德:2)5
36 (西德:2)9 4.2
MRI Results
cortex
VMPFC
Ventral striatum extending into the
caudate nucleus
pFMC
Intraparietal sulcus
Posterior parietal cortex
LIPFC
(西德:2)11
7
(西德:2)2
6
3
9 4.7
9 4.1
30
39 3.8
(西德:2)29 (西德:2)65
42 4.8
40 (西德:2)68
39 4.0
(西德:2)29
36 (西德:2)3 4.7
Conjunction analysis of the contrasts RH-based decisional
processes and recognition memory processes
Retrosplenial cortex
Precuneus
7 (西德:2)53
15 4.2
10 (西德:2)60
24 4.0
(西德:2)20 (西德:2)57
24 3.6
Reaction times (RT) of the following conditions are shown: partici-
pants recognized both cities (RR), no city (UU), participants recognized
one city and had to choose the recognized alternative (RUR) or had to
choose the unrecognized alternative (RUU).
the recognized alternative as compared to choosing the
unrecognized alternative, t(13) = (西德:2)1.94; p = .08.
一般来说, to control for the RT differences between the
状况, we included RT parameters for each condi-
tion as in Experiment 1. Parallel to the heuristic contrast
in Experiment 1, we compared trials in which partici-
pants recognized one alternative and chose the recog-
nized one with trials in which they recognized one
alternative and chose the unrecognized one. A signifi-
cantly higher hemodynamic activity was elicited within
the left inferior occipitotemporal cortex and inferior
frontal sulcus when participants chose the recognized
城市 (数字 2, 桌子 4).
桌子 4. 实验 2
Area
X
y
z
Z
Parametric analysis of increasing recognition strength
Decisional processes based on recognition only
Midportion of the parahippocampal
(西德:2)20 (西德:2)33 (西德:2)6 4.4
Inferior occipitotemporal cortex
(西德:2)44 (西德:2)59
28 (西德:2)36 (西德:2)6 4.5
Inferior frontal sulcus
(西德:2)44
36
0
9
3.8
3.6
Retrosplenial cortex extending into
(西德:2)5 (西德:2)54
21 4.7
7 (西德:2)53
12 4.9
Recognition memory processes
(西德:2)5
34 (西德:2)9 4.4
Midportion of the parahippocampal
(西德:2)29 (西德:2)38
(西德:2)6
(西德:2)11
7
(西德:2)2
6
4
9 4.5
3 4.0
30
39 3.9
gyrus
Precuneus extending into
retrosplenial cortex
25 (西德:2)36 (西德:2)11
(西德:2)11 (西德:2)59
15
(西德:2)8
(西德:2)2
(西德:2)11
(西德:2)29
39
7
60
28
(西德:2)9
3
24
(西德:2)8
4.4
3.6
4.0
4.0
4.7
4.2
4.4
Posterior parietal cortex/intraparietal
(西德:2)29 (西德:2)65
42 4.7
VMPFC
sulcus
LIPFC
小脑
37 (西德:2)66
39 4.4
(西德:2)29
39 (西德:2)6 4.7
10 (西德:2)86 (西德:2)18 4.2
Ventral striatum extending into the
caudate nucleus
aFMC (布罗德曼区 9)
LIPFC
aFMC = anterior frontomedian cortex; VMPFC = ventromedial pre-
frontal cortex; pFMC = posterior frontomedian cortex; LIPFC = left
inferior prefrontal cortex.
VMPFC = ventromedial prefrontal cortex; aFMC = anterior frontome-
dian cortex; LIPFC = left inferior prefrontal cortex.
Anatomical specification, Talairach coordinates (X, y, z) and maximal
Z scores of significantly activated voxels are shown for each contrast
calculated in Experiment 1.
Anatomical specification, Talairach coordinates (X, y, z), and maximal
Z scores of significantly activated voxels are shown for each contrast
calculated in Experiment 2.
Volz et al.
1931
gyrus
precuneus
VMPFC
Caudate nucleus
pFMC
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
t
w
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
n
t
我
C
/
A
e
r
–
p
t
d
我
C
1
我
8
e
1
–
1
p
1
d
9
F
2
/
4
1
8
1
/
9
1
3
5
1
6
/
1
1
7
9
2
哦
4
C
/
n
1
2
7
0
5
0
5
6
9
1
8
8
7
/
1
j
1
哦
1
C
9
n
2
.
4
2
p
0
d
0
6
乙
.
y
1
G
8
你
.
e
1
s
1
t
.
哦
1
n
9
0
2
8
4
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
F
.
.
.
.
哦
n
1
8
中号
A
y
2
0
2
1
To investigate which brain areas were involved with
successful recognition we contrasted RR trials with UU
trials parallel to Experiment 1. Significant activation was
observed bilaterally within the midportion of the para-
hippocampal gyrus, right precuneus extending into
retrosplenial cortex, bilaterally within the ventral stria-
tum extending into caudate nucleus, LIPFC, aFMC
(布罗德曼区 9), and VMPFC (数字 2, 桌子 4).
A conjunction analysis between these two contrasts
revealed no area to be commonly activated.
To identify overlap in activation patterns for success-
ful recognition memory processes in Experiments 1 和
2, we calculated a test for the logical AND (Nichols et al.,
2005). Regions that revealed to be commonly activated
by recognition memory processes in both experiments
were bilaterally the midportion of the parahippocampal
gyrus, precuneus extending into the retrosplenial cor-
tex, ventral striatum bilaterally, and LIPFC. Activation
within the MTL and medial parietal cortex have repeat-
edly been associated with recognition memory process-
英语 (例如, Henson, 2005; Wagner et al., 2005), and LIPFC
activation with controlled semantic processing (例如,
金子, Balota, Kirchhoff, & 巴克纳, 2005). 因此, 这
activation obtained in these experiments suggests that
recognition memory processes are engaged both when
assessing which city was larger (实验 1) 什么时候
assessing which city was recognized (实验 2).
讨论
The present study investigated the neural correlates of de-
cisional processes that are based on the RH. Simply stated,
the RH claims that when making a judgment about two
项目, a person recognizing only one of the alternatives
will infer the recognized one has the higher criterion value.
The effective use of the RH depends on the outcome of
two processes: whether the alternatives are recognized
and a judgment about the ecological rationality of the
RH, 那是, whether the chain of correlations linking the
(inaccessible) criterion to recognition through a mediator
variable justifies following the RH. 因此, RH-based
decisional processes are special
in that they feature a
judgment of ecological rationality compared to decisional
processes that solely rely on recognition information.
In the following we will discuss the medial parietal
activation and the frontomedian activation that was
found to be specific to the application of the RH.
随后, we will discuss why there was no fronto-
median activation when the RH was not applied.
Activation within Medial Parietal Areas Reflect
Recognition Processes When Decisions Follow
the RH
Generalizing from anatomical studies in monkeys, 这
precuneus and retrosplenial cortex are upstream to the
medial temporal memory system (Kobayashi & Amaral,
2003). 因此, medial parietal areas are directly or indi-
rectly connected to the MTL. The retrosplenial cortex is
special in that its afferent connections are dominated by
MTL projections. Based on this finding, Kobayashi and
Amaral (2003) suggested parietal midline structures as
the major pathways through which the MTL influences
cortical information processing in the service of declar-
ative memory. This assumption is further supported by
a phenomenon referred to as ‘‘retrosplenial amnesia.’’
Damage to the retrosplenial cortex has been associ-
ated with the loss of verbal episodic memory as well
as with a loss for spatial relations (Maeshima et al., 2001;
Valenstein et al., 1987). Imaging studies repeatedly re-
ported activation within parietal midline structures that
extend from the retrosplenial cortex and posterior cin-
gulate to the precuneus for episodic memory retrieval
流程 (Krause et al., 1999; Maguire, Frith, & 莫里斯,
1999; Squire et al., 1992): Activation within medial pa-
rietal areas, specifically within the precuneus, 已经
shown to be sensitive to successful retrieval (old/new
effects; Henson, Rugg, Shallice, Josephs, & Dolan, 1999;
Fletcher et al., 1995), autobiographical memory retrieval
(Addis, McIntosh, Moscovitch, Crawley, & McAndrews,
2004; Gilboa, Winocur, Grady, Hevenor, & Moscovitch,
2004), retrieval orientation (Dobbins, 米, 瓦格纳,
& Schacter, 2003), and for the subjective perception
that information is old (perceived recognition; Kahn,
Davachi, & 瓦格纳, 2004; Wheeler & 巴克纳, 2003).
然而, 同时, precuneus activation has also
imagery, 自己-
been found in studies of visuospatial
processing operations, and consciousness (for an over-
看法, see Cavanna & Trimble, 2006; Naghavi & Nyberg,
2005). Given the involvement of the precuneus in such
a wide spectrum of higher order cognitive functions,
the exact nature of the precuneus function has long
been considered elusive, and domain-specific interpre-
tations dominated. By reviewing current knowledge
about the anatomical and cytoarchitectonic structure
imaging data,
of the precuneus as well as functional
Cavanna and Trimble (2006) have suggested a functional
subdivision within the precuneus: an anterior region
that is especially involved in self-centered mental imag-
ery strategies, and a posterior region mainly subserving
successful episodic memory retrieval, irrespective of the
imagery content of the retrieved information.
The precuneus activation in the present study for RH-
based decisional processes occurred in the posterior
part of the precuneus, specifically anterior to the parieto-
occiptial sulcus, and thus is suggested to be associated
with successful retrieval attempts. This interpretation
is in line with the assumption that the precuneus par-
ticularly responds to familiarity confidence (Yonelinas,
Otten, Shaw, & Rugg, 2005). Yonelinas et al. (2005)
reported a positive correlation between posterior pre-
cuneus activation and recognition confidence, 建议-
ing that activation within the posterior precuneus can be
1932
认知神经科学杂志
体积 18, 数字 11
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
t
w
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
n
t
我
C
/
A
e
r
–
p
t
d
我
C
1
我
8
e
1
–
1
p
1
d
9
F
2
/
4
1
8
1
/
9
1
3
5
1
6
/
1
1
7
9
2
哦
4
C
/
n
1
2
7
0
5
0
5
6
9
1
8
8
7
/
1
j
1
哦
1
C
9
n
2
.
4
2
p
0
d
0
6
乙
.
y
1
G
8
你
.
e
1
s
1
t
.
哦
1
n
9
0
2
8
4
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
F
.
.
.
哦
n
1
8
中号
A
y
2
0
2
1
used as proxy for the overall strength of the recognition
signal. Additional support for this comes from our
parametric analysis linking medial posterior parietal
cortex with how likely a city was recognized by our
参与者. 实验 1 revealed higher activation
within the aFMC and precuneus extending into the
retrosplenial cortex when participants responded ac-
cording to the RH (IE。, choosing the recognized city
as larger) as compared to when they did not. 这
activation foci that were elicited by RH-based decisional
流程 (IE。, aFMC, precuneus, and retrosplenial cor-
tex) could be dissociated with respect to their involve-
ment in recognition memory processes: The medial
parietal areas alone were found to be activated both
by the application of the RH and recognition. Accord-
英利, we suggest medial parietal cortex activation to
mainly reflect recognition memory processes during
RH-based decisions, whereas aFMC activation could be
specific to judgmental processes about the RH’s ecolog-
ical rationality.
Activation within the aFMC Reflects Judgments
of Ecological Rationality When Decisions Follow
the RH
Based on Experiment 1, one could entertain the hypoth-
esis that aFMC is doing nothing more than accumulating
a recognition signal from medial parietal areas, implying
that decisions are made strictly on the basis of recogni-
的. This suggests that to the extent that RH-based
decisional processes can adequately be explained by
participants simply choosing the recognized alternative,
aFMC activation should also be elicited by decisional
processes based solely on recognition information. 到
test this hypothesis, we ran Experiment 2, 但反而
asked participants to choose the recognized alternative
in each trial. Contrasting trials (in Experiment 2) 其中
participants chose the recognized alternative with trials
in which they did the opposite revealed no significant
aFMC activation, but instead activation within the left
inferior occipitotemporal cortex. This area has repeat-
edly been found for lexical decisions in the visual do-
for word–pseudoword discrimination;
主要的 (例如,
Fiebach, Friederici, 穆勒, & von Cramon, 2002). 这
finding led to the assumption that occipitotemporal
brain areas could be conceived of as a functionally spe-
cialized word recognition region within the ventral visual
路径. Fiebach and Friederici (2002) replicated the
lexical decision task in the auditory domain and again
found activation within the occipitotemporal cortex for
real words as compared to pseudowords, which led the
authors to suggest this area to be related to modality-
independent aspects of word recognition. 因此,
when participants (in Experiment 2) chose the recog-
nized city, they simply opted for that word whose word
representation form was stored in their lexicon.
These results further bolster the claim that the aFMC
activation observed in Experiment 1 is specific to judg-
ments of ecological rationality and not merely for the
strength of the recognition signal. 最近, the aFMC
has been proposed to reflect self-related processes
or social–cognitive judgments (Northoff & Bempohl,
2004; Ochsner et al., 2004; Gusnard, Akbudak, 舒尔曼,
& Raichle, 2001). 一般来说, such processes can be char-
acterized by the requirement to relate an aspect of the
external world to oneself involving the ranking, scaling,
and evaluating of one’s own priorities and notions in re-
lation to parameters of the external situation (Goldberg
& Podell, 1999). A feature of paradigms investigating
the cerebral correlates of self-related processes is that re-
sponses cannot be scored against an absolute standard;
相当, correct responses are relative and actor cen-
tered, such as what is conceived of as moral (Heekeren,
Wartenburger, 施密特, Schwintowski, & 狂野的召唤者, 2003;
Moll et al., 2002; Greene, Sommerville, Nystrom, Darley,
& 科恩, 2001) or as coherent (Ferstl & von Cramon,
2002) or as beautiful ( Jacobsen, Schubotz, Hofel, &
Cramon, 2006). In the city judgment task in Experi-
蒙特 1, participants had to evaluate the applicability
of the RH, which could be done by inferring a rough
correlation between the (strength of ) recognition and
city population. 因此, judgments of the RH’s ecolog-
ical rationality are largely self-referential, as they depend
on assessing one’s own sense of recognition.
Why Wasn’t There aFMC Activation When
the RH Was Not Followed?
One might expect judgments of ecological rationality
should be required in those trials in which the RH was
not applied. 然而, these trials did not elicit aFMC activa-
的. To investigate whether aFMC activation results
from a relative difference in signal strength between
the two sorts of trials or rather because trials in which
the RH was not applied did not activate the aFMC at all,
we plotted the time course of the mean percentage
signal change in the aFMC. Results revealed less deacti-
vation for trials in which the RH was applied as com-
pared to those trials in which the RH was not applied
(数字 3). The time course of the aFMC supports the
default-mode hypothesis ascribed to the medial prefron-
tal cortex that was put forward by Gusnard et al. (2001).
The finding that the aFMC is among those brain regions
showing the highest baseline metabolic activity and task-
related activation reductions during goal-directed cogni-
tive processing led to the assumption that the medial
prefrontal cortex subserves functions that are essential
to the self-concept: During attention-demanding tasks
the self-referential activity of the aFMC is reduced. 在里面
present study, activation in the aFMC was reduced more
when participants failed to apply the RH, suggesting that
the suspension of the RH required more cognitive effort
Volz et al.
1933
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
t
w
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
n
t
我
C
/
A
e
r
–
p
t
d
我
C
1
我
8
e
1
–
1
p
1
d
9
F
2
/
4
1
8
1
/
9
1
3
5
1
6
/
1
1
7
9
2
哦
4
C
/
n
1
2
7
0
5
0
5
6
9
1
8
8
7
/
1
j
1
哦
1
C
9
n
2
.
4
2
p
0
d
0
6
乙
.
y
1
G
8
你
.
e
1
s
1
t
.
哦
1
n
9
0
2
8
4
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
.
.
.
F
哦
n
1
8
中号
A
y
2
0
2
1
数字 3. Mean percentage
signal changes (MPSCs, IE。,
event-related averages) 和
error bars for the aFMC, (正确的)
precuneus, and retrosplenial
cortex in Experiment 1: Top,
MPSCs are shown for the aFMC
for trials in which the RH was
applied (红线) 并且是
not applied (蓝线). 中间,
MPSCs in the (正确的) precuneus
for all conditions. Bottom,
MPSCs for the retrosplenial
cortex (绿色的, RR trials;
黑色的, UU trials). The stimulus
presentation of the two city
名字, lasting for 4 秒, started
at time point zero.
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
t
w
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
n
t
我
C
/
A
e
r
–
p
t
d
我
C
1
我
8
e
1
–
1
p
1
d
9
F
2
/
4
1
8
1
/
9
1
3
5
1
6
/
1
1
7
9
2
哦
4
C
/
n
1
2
7
0
5
0
5
6
9
1
8
8
7
/
1
j
1
哦
1
C
9
n
2
.
4
2
p
0
d
0
6
乙
.
y
1
G
8
你
.
e
1
s
1
t
.
哦
1
n
9
0
2
8
4
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
F
.
.
.
.
than judgments in line with the heuristic, suggesting
participants follow the RH by default. This conclusion is
consistent with the RT data: Participants in Experiment 1
took significantly longer to decide against the RH, 那
是, when assessing the unrecognized alternative as the
larger city (F(1,17) = 30.2; p = .0001). The finding that it
takes additional time to suspend the RH has also been
reported by Pachur and Hertwig (in press).
There could be multiple reasons that would lead
participants to go against the RH. 例如, a person
could discount their recognition because they know that
the city is small (Oppenheimer, 2003). 例如, 这
city of Pisa in Italy is probably known by its leaning
tower, so participants may infer that the recognition of
Pisa is not directly indicative of a large population size
(关于 86,000 inhabitants). Another possibility is that
they are not only taking into account whether they
recognize the city or not, but how confident they are
that they recognize it. Evidence for this comes from
the time course for activation in the precuneus and
retrosplenial cortex when the RH was followed and
when it was not. 数字 3 shows activation in the
precuneus and retrosplenial areas for the four kinds of
trials of interest in Experiment 1. Notice that the UU,
RU(右), and RR trials are ordered in terms of the number
of alternatives recognized, in line with the suggestion of
Yonelinas et al. (2005) that the precuneus ref lects
recognition confidence and our parametric analysis of
哦
n
1
8
中号
A
y
2
0
2
1
1934
认知神经科学杂志
体积 18, 数字 11
recognition rates and activation in these regions. 这
surprising result is the paltry activation in medial parietal
areas for the RU(U ) trials where the activation is not
significantly different from the signal of UU trials, 那是,
when neither item was recognized [precuneus: t(17) =
(西德:2)0.42; p = .68; retrosplenial cortex: t(17) = (西德:2)1.5;
p = .17] (数字 3), consistent with the hypothesis that
they failed to follow the RH when recognition confi-
dence is low.
合在一起, the present fMRI studies converge on
the following conclusions: (1) Whereas RH-based deci-
sions do depend on the strength of the recognition
signal, as measured by activation in medial parietal areas,
(2) the processes underlying RH-based decisions go
beyond simply choosing the recognized alternative. (3)
具体来说, RH-based decisional processes are distin-
guished by judgments about the ecological rationality
of the RH that are specific to the aFMC.
致谢
The authors thank Jennifer Kittel for assistance in data collec-
的, Ralph Hertwig for valuable considerations and productive
discussions, and two anonymous reviewers for their very con-
structive and helpful comments on a prior version of the
manuscript.
Reprint requests should be sent to Kirsten G. Volz, Max Planck
Institute for Human Cognitive and Brain Sciences P.O. Box
500 355, D-04303 莱比锡, 德国, or via e-mail: volz@cbs.
mpg.de, www.cbs.mpg.de.
参考
Addis, D. R。, McIntosh, A. R。, Moscovitch, M。, Crawley, A. P。,
& McAndrews, 中号. 磷. (2004). Characterizing spatial and
temporal features of autobiographical memory retrieval
网络: A partial least squares approach. Neuroimage,
23, 1460–1471.
Aggleton, J. P。, 货车, S. D ., Denby, C。, Dix, S。, Mayes, A. R。,
罗伯茨, N。, 等人. (2005). Sparing of the familiarity
component of recognition memory in a patient with
hippocampal pathology. Neuropsychologia, 43, 1810–1823.
Annual report 2002 (PP. 28–29). 莱比锡: Max Planck
Institute for Human Cognitive and Brain Sciences.
Fiebach, C. J。, Friederici, A. D ., 穆勒, K., & von Cramon,
D. 是. (2002). fMRI evidence for dual routes to the mental
lexicon in visual word recognition. 认知杂志
神经科学, 14, 11–23.
弗莱彻, 磷. C。, Frith, C. D ., Grasby, 磷. M。, Shallice, T。,
Frackowiak, 右. S。, & Dolan, 右. J. (1995). Brain systems
for encoding and retrieval of auditory–verbal memory.
An in vivo study in humans. Brain, 118, 401–416.
Forman, S. D ., 科恩, J. D ., Fitzgerald, M。, Eddy, 瓦. F。,
Mintun, 中号. A。, & Noll, D. C. (1995). Improved assessment
of significant activation in functional magnetic resonance
成像 (功能磁共振成像): Use of a cluster-size threshold. Magnetic
Resonance in Medicine, 33, 636–647.
弗里斯顿, K. J。, 弗莱彻, P。, Josephs, 奥。, Holmes, A。, Rugg, 中号. D .,
& 车工, 右. (1998). Event-related fMRI: Characterizing
differential responses. Neuroimage, 7, 30–40.
弗里斯顿, K. J。, Frith, C. D ., 车工, R。, & Frackowiak, 右. S.
(1995). Characterizing evoked hemodynamics with fMRI.
Neuroimage, 2, 157–165.
Gilboa, A。, Winocur, G。, Grady, C. L。, Hevenor, S. J。, &
Moscovitch, 中号. (2004). Remembering our past: Functional
neuroanatomy of recollection of recent and very remote
personal events. 大脑皮层, 14, 1214–1225.
金子, 乙. T。, Balota, D. A。, Kirchhoff, 乙. A。, & 巴克纳, 右. L.
(2005). Common and dissociable activation patterns
associated with controlled semantic and phonological
加工: Evidence from fMRI adaptation. Cerebral
Cortex, 15, 1438–1450.
Goldberg, E., & Podell, K. (1999). Adaptive versus veridical
decision making and the frontal lobes. 意识
和认知, 8, 364–377.
戈德斯坦, D. G。, & Gigerenzer, G. (2002). Models of
ecological rationality: The recognition heuristic.
心理评论, 109, 75–90.
Greene, J. D ., Sommerville, 右. B., Nystrom, L. E., Darley,
J. M。, & 科恩, J. D. (2001). An fMRI investigation of
emotional engagement in moral judgment. 科学, 293,
2105–2108.
Gusnard, D. A。, Akbudak, E., 舒尔曼, G. L。, & Raichle,
中号. 乙. (2001). Medial prefrontal cortex and self-referential
mental activity: Relation to a default mode of brain function.
美国国家科学院院刊, 美国。,
98, 4259–4264.
Heekeren, H. R。, Wartenburger, 我。, 施密特, H。, Schwintowski,
H. P。, & 狂野的召唤者, A. (2003). An fMRI study of simple
ethical decision-making. NeuroReport, 14, 1215–1219.
Aggleton, J. P。, & Shaw, C. (1996). Amnesia and recognition
Henson, 右. (2005). A mini-review of fMRI studies of
记忆: A re-analysis of psychometric data.
Neuropsychologia, 34, 51–62.
Birn, 右. M。, 考克斯, 右. W., & Bandettini, 磷. A. (2002). Detection
versus estimation in event-related fMRI: Choosing the
optimal stimulus timing. Neuroimage, 15, 252–264.
Cavanna, A. E., & Trimble, 中号. 右. (2006). The precuneus:
A review of its functional anatomy and behavioural
correlates. Brain, 129, 564–583.
Dobbins, 我. G。, 米, H. J。, 瓦格纳, A. D ., & Schacter, D. L.
(2003). Memory orientation success: Separable
neurocognitive components underlying episodic
认出. Neuropsychologia, 41, 318–333.
Ferstl, 乙. C。, & Cramon, 是. D. (2002). What does the
frontomedian cortex contribute to language processing:
Coherence or theory of mind? Neuroimage, 17,
1599–1612.
Fiebach, C. J。, & Friederici, A. D. (2002). Neural correlates
of lexicality and word frequency in the auditory lexical
decision task. 在一个. D. Friederici & D. 是. von Cramon (编辑。),
human medial temporal lobe activity associated with
recognition memory. Quarterly Journal of Experimental
心理学, Series B, Comparative and Physiological
心理学, 58, 340–360.
Henson, 右. 氮. A。, Rugg, 中号. D ., Shallice, T。, Josephs, 奥。,
& Dolan, 右. J. (1999). Recollection and familiarity in
recognition memory: An event-related functional magnetic
resonance imaging study. 神经科学杂志, 19,
3962–3972.
Jacobsen, T。, Schubotz, 右. 我。, Hofel, L。, & Cramon, D. 是.
(2006). Brain correlates of aesthetic judgment of beauty.
Neuroimage, 29, 276–285.
Kahn, 我。, Davachi, L。, & 瓦格纳, A. D. (2004). Functional–
neuroanatomic correlates of recollection: Implications
for models of recognition memory. 杂志
神经科学, 24, 4172–4180.
Kobayashi, Y。, & Amaral, D. G. (2003). Macaque monkey
retrosplenial cortex: 二. Cortical afferents. 杂志
Comparative Neurology, 466, 48–79.
Volz et al.
1935
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
t
w
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
n
t
我
C
/
A
e
r
–
p
t
d
我
C
1
我
8
e
1
–
1
p
1
d
9
F
2
/
4
1
8
1
/
9
1
3
5
1
6
/
1
1
7
9
2
哦
4
C
/
n
1
2
7
0
5
0
5
6
9
1
8
8
7
/
1
j
1
哦
1
C
9
n
2
.
4
2
p
0
d
0
6
乙
.
y
1
G
8
你
.
e
1
s
1
t
.
哦
1
n
9
0
2
8
4
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
.
F
.
.
哦
n
1
8
中号
A
y
2
0
2
1
克劳斯, 乙. J。, 施密特, D ., Mottaghy, F. M。, 泰勒, J。,
Halsband, U。, Herzog, H。, 等人. (1999). Episodic retrieval
activates the precuneus irrespective of the imagery
content of word pair associates. A PET study. Brain,
122, 255–263.
Lohmann, G。, 穆勒, K., Bosch, 五、, Mentzel, H。, Hessler, S。,
陈, L。, 等人. (2001). LIPSIA—A new software system for
the evaluation of functional magnetic resonance imaging
of the human brain. Computerized Medical Imaging and
Graphics, 25, 449–457.
Maeshima, S。, Ozaki, F。, Masuo, 奥。, Yamaga, H。, Okita, R。,
& Moriwaki, H. (2001). Memory impairment and spatial
disorientation following a left retrosplenial lesion. 杂志
of Clinical Neuroscience, 8, 450–451.
Maguire, 乙. A。, Frith, C. D ., & 莫里斯, 右. G. (1999).
The functional neuroanatomy of comprehension and
记忆: The importance of prior knowledge. Brain,
122, 1839–1850.
Pohl, 右. F. (2006). Empirical tests of the recognition
启发式的. Journal of Behavioral Decision Making.
Ranganath, C。, Yonelinas, A. P。, 科恩, 中号. X。, Dy, C. J。,
汤姆, S. M。, & D’Esposito, 中号. (2003). Dissociable
correlates of recollection and familiarity within the
medial temporal lobes. Neuropsychologia, 42, 2–13.
Rugg, 中号. D ., & Yonelinas, A. 磷. (2003). Human recognition
记忆: A cognitive neuroscience perspective. Trends
in Cognitive Sciences, 7, 313–319.
Squire, L. 右. (1994). Declarative and non-declarative
记忆: Multiple brain systems supporting learning and
记忆. 在D中. L. Schacter & 乙. Tulving (编辑。), 记忆
系统 (PP. 203–231). 剑桥: 与新闻界.
Squire, L. R。, Ojemann, J. G。, 米津, F. M。, 彼得森, S. E.,
Videen, 时间. 奥。, & Raichle, 中号. 乙. (1992). Activation of the
hippocampus in normal humans: A functional anatomical
study of memory. 国家科学院院刊
科学系, 美国。, 89, 1837–1818.
米津, F. M。, Maccotta, L。, Ollinger, J. M。, 彼得森, S. E.,
Squire, L. R。, Stark, C. 乙. L。, & 克拉克, 右. 乙. (2004). 这
& 巴克纳, 右. L. (2000). Characterizing the hemodynamic
response: Effects of presentation rate, sampling
procedure, and the possibility of ordering brain activity
based on relative timing. Neuroimage, 11, 735–759.
Moll, J。, de Oliveira-Souza, R。, Eslinger, 磷. J。, Bramati, 我. E.,
medial temporal lobe. Annual Review of Neuroscience,
27, 279–306.
Talairach, P。, & Tournoux, J. (1998). Co-planar stereotactic
atlas of the human brain. Stuttgart: Thieme.
Thirion, J. 磷. (1998). Image matching as a diffusion process:
Moura˜o-Miranda, J。, Andreiuolo, 磷. A。, 等人. (2002).
The neural correlates of moral sensitivity: A functional
magnetic resonance imaging investigation of basic and
moral emotions. The Journal of Neuroscience, 22,
2730–2736.
Naghavi, H. R。, & Nyberg, L. (2005). Common fronto-parietal
activity in attention, 记忆, and consciousness: Shared
demands on integration? Consciousness and Cognition,
14, 390–425.
Nichols, T。, Brett, M。, 安德森, J。, Wager, T。, & 波琳娜,
J. 乙. (2005). Valid conjunction inference with the
minimum statistic. Neuroimage, 25, 653–660.
Northoff, G。, & Bermpohl, F. (2004). Cortical midline
structures and the self. 认知科学的趋势,
8, 102–107.
Norris, D. G. (2000). Reduced power multislice MDEFT
成像. Journal of Magnetic Resonance Imaging, 11,
445–451.
Ochsner, K. N。, Knierim, K., Ludlow, D. H。, Hanelin, J。,
Ramachandran, T。, Glover, G。, 等人. (2004). Reflecting
upon feelings: An fMRI study of neural systems
supporting the attribution of emotion to self and other.
认知神经科学杂志, 16, 1746–1772.
Oppenheimer, D. 中号. (2003). Not so fast! (and not so
frugal!): Rethinking the recognition heuristic. 认识,
90, B1–B9.
Pachur, T。, & Hertwig, 右. (in press). On the psychology
of the recognition heuristic: Retrieval primacy as a
key determinant of its use. 实验杂志
心理学: 学习, 记忆, 和认知.
An analogy with Maxwell’s demons. Medical Image
分析, 2, 243–260.
Ugurbil, K., Garwood, M。, Ellermann, J。, Hendrich, K.,
Hinke, R。, 胡, X. P。, 等人. (1993). Imaging at high
magnetic fields: Initial experiences at 4 时间. Magnetic
Resonance Quarterly, 9, 259–277.
Valenstein, E., Bowers, D ., Verfaellie, M。, Heilman, K. M。,
Day, A。, & 沃森, 右. 时间. (1987). Retrosplenial amnesia.
Brain, 110, 1631–1646.
瓦格纳, A. D ., Shannon, 乙. J。, Kahn, 我。, & 巴克纳, 右. L.
(2005). Parietal lobe contributions to episodic memory
恢复. 认知科学的趋势, 9, 445–453.
Wheeler, 中号. E., & 巴克纳, 右. L. (2003). Functional
dissociation among components of remembering:
控制, perceived oldness, and content. 杂志
神经科学, 23, 3869–3880.
Worsley, K. J。, & 弗里斯顿, K. J. (1995). Analysis of fMRI
time-series revisited–再次. Neuroimage, 2, 173–181.
Ward, 乙. D. (2000). Simultaneous inference for fMRI data.
可用的: afni.nimh.nih.gov/afni/doc/manual/AlphaSim
[状态 2006-01-06].
Yonelinas, A. P。, Otten, L. J。, Shaw, K. N。, & Rugg, 中号. D.
(2005). Separating the brain regions involved in
recollection and familiarity in recognition memory.
The Journal of Neuroscience, 25, 3002–3008.
Yonelinas, A. P。, Kroll, 氮. E., Quamme, J. R。, Lazzara, 中号. M。,
Sauve, 中号. J。, Widaman, K. F。, 等人. (2002). Effects of
extensive temporal lobe damage or mild hypoxia on
recollection and familiarity. 自然神经科学, 5,
1236–1241.
1936
认知神经科学杂志
体积 18, 数字 11
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
t
w
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
n
t
我
C
/
A
e
r
–
p
t
d
我
C
1
我
8
e
1
–
1
p
1
d
9
F
2
/
4
1
8
1
/
9
1
3
5
1
6
/
1
1
7
9
2
哦
4
C
/
n
1
2
7
0
5
0
5
6
9
1
8
8
7
/
1
j
1
哦
1
C
9
n
2
.
4
2
p
0
d
0
6
乙
.
y
1
G
8
你
.
e
1
s
1
t
.
哦
1
n
9
0
2
8
4
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
.
.
.
F
哦
n
1
8
中号
A
y
2
0
2
1