TMS of the FEF Interferes with Spatial Conflict
Lara Bardi1, Ryota Kanai2, Daniela Mapelli1, and Vincent Walsh2
抽象的
■ In the Simon task, a conflict arises because irrelevant spatial
information competes for response selection either facilitating
or interfering with performance. Responses are faster when
stimulus and response position correspond than when they
不要. The FEFs, which have long been characterized for their
role in oculomotor control, are also involved in the control of
visuospatial attention when eye movements are not required.
This study was aimed at investigating whether the FEFs con-
tribute to spatial conflict. Double-pulse TMS was applied to
the FEF of either left or right hemisphere during the execution
of a Simon task at different time windows after the onset of
the visual stimulus. A suppression of the Simon effect was ob-
served after stimulation of the FEF for stimuli appearing in the
contralateral hemifield when TMS was applied to the left hemi-
sphere after stimulus onset (0–40 and 40–80 msec). A reduction
of the correspondence effect was observed after right FEF TMS
for stimuli presented in the left visual hemifield when stim-
ulation was delivered in the 80–120 msec range after stimulus
onset. These outcomes indicate that the FEF play a critical
role in encoding spatial attribute of a stimulus for response
priming, which is the prerequisite for response conflict in the
Simon task. 而且, our finding that the left FEF have a
dominant role during spatial conflict extends the idea of the
left-hemisphere lateralization of the motor network in action
selection by suggesting that the FEF may constitute part of this
网络. ■
介绍
The visual environment continuously presents us with
multiple opportunities for action, and a conflict often
arises between sensorimotor processes and different pos-
sible responses. In many situations, irrelevant information
may interfere with response selection, and this effect of
interference is particularly robust when the irrelevant in-
formation is strongly associated with a dimension of the
response.
In the Simon task, a conflict arises because irrelevant
spatial information competes with relevant nonspatial in-
formation for response selection. 这里, participants are
required to respond to a nonspatial attribute of lateralized
targets (例如, color or shape) while ignoring their spatial
位置. 例如, participants may be asked to press
the left key when a red circle appears and the right key
when a green circle appears. Although stimulus location
is irrelevant to the task, RTs are faster when stimulus and
response positions correspond than when they do not
(鲁 & Proctor, 1995; Umiltà & Nicoletti, 1990; 西蒙 &
Rudell, 1967).
To explain mechanisms underlying conflict, dual-route
型号 (德容, 梁, & Lauber, 1994; Kornblum,
Hasbroucq, & Osman, 1990) postulate two parallel routes
of information processing: spatial information automati-
cally activates its spatially corresponding response via a
fast direct route, whereas the task-relevant stimulus code
1University of Padua, 2伦敦大学学院
activates the required response via a slow controlled in-
direct route ( Wascher, Schatz, Kuder, & Verleger, 2001;
Kornblum et al., 1990). The direct route may be estab-
lished by long-term associations between perceptual
and motor processes, which may depend on the synaptic
consolidation of stimulus–response (S-R) 协会
widely overlearned during lifetime (Tagliabue, Zorzi,
Umiltà, & Bassignani, 2000; 科恩, Dunbar, & 麦克莱兰,
1990). When the direct route (carrying the response linked
to the spatial position of the stimulus) and the indirect
一 (holding the task demands) generate the same re-
sponse codes (IE。, in the corresponding condition [C]),
RTs are speeded up, whereas when the two routes gener-
ate different response codes (IE。, in the noncorresponding
状况 [NC]), a conflict must be resolved, 导致
slower responses.
The “attention shift” hypothesis maintains that the irrele-
vant spatial code of the stimulus is generated by the direc-
tion of the attention shift implemented before response
选择. When the peripheral stimulus is presented, atten-
tion covertly shifts toward its position (Rubichi, Nicoletti,
Iani, & Umiltà, 1997; Nicoletti & Umiltà, 1994). 根据
to the premotor theory of attention, shifting attention is
accomplished by the generation of the motor program for
the corresponding saccade regardless of whether the sac-
cade is subsequently executed or not (Umiltà, Riggio,
Dascola, & Rizzolatti, 1991; Rizzolatti, Riggio, Dascola,
& Umiltà, 1987). The alternative hypothesis, known as
the “referential coding” hypothesis (Hommel, 1993; 看
also Hommel, 2011), states that the irrelevant spatial code
© 2012 麻省理工学院
认知神经科学杂志 24:6, PP. 1305–1313
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
4
/
6
2
4
1
/
3
6
0
/
5
1
1
3
9
0
4
5
3
/
8
1
2
7
6
7
哦
8
C
0
n
2
_
5
A
/
_
j
0
哦
0
C
2
n
2
3
_
A
p
_
d
0
0
乙
2
y
2
G
3
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
t
.
F
/
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
is generated as a result of the comparison between the
location of the target stimulus and the location of an in-
tentionally defined reference stimulus (例如, the fixation
观点). Both the attention shifting and the referential cod-
ing accounts of the Simon effect have linked the process-
ing of the spatial code to attention, to some extent at least
(for a recent discussion of these hypotheses, see also
Van der Lubbe & Abrahamse, 2011).
A neural correlate for encoding spatial coordinates of
stimulus and response during conflict may be attributed
to cerebral areas devoted to the control of visuospatial at-
tention and visuomotor integration. The posterior parietal
cortex (个人电脑), as part of the dorsal visual stream, is thought
to be involved in both visuospatial attention and trans-
formation of sensory information into codes for action
(Kravitz, Saleem, 贝克, & Mishkin, 2011; Goodale &
Milner, 1992; Rizzolatti et al., 1987). Recent studies sug-
gested that PPC plays a critical role in processing irrelevant
spatial information for response priming during conflict.
TMS of PPC, applied after stimulus onset, causes a suppres-
sion of the Simon effect as if no stimulus spatial code inter-
fered with performance (Schiff, Bardi, Basso, & Mapelli,
2011; Rusconi, Turatto, & Umiltà, 2007; Stürmer, Redlich,
Irlbacher, & Brandt, 2007). On the basis of these results,
PPC has been identified as the main neural substrate of
visuomotor transmission within the direct route.
A growing body of anatomical and physiological evi-
dence suggests, 然而, that space representation and
visuomotor transformation involve other brain areas than
PPC within a distributed fronto-parietal cortical circuit. A
prominent role within this network has recently been
assigned to the FEF (科尔贝塔 & 舒尔曼, 2002), 哪个
have been shown to contribute to transformation of visual
signals into saccade commands. This area has long been
characterized for its role in the control of eye movements
(例如, Olk, 张, Kingstone, & Ro, 2006; 摩尔 & Farah,
2001; Gaymard, Ploner, Rivaud, Vermersch, & Pierrot-
Deseilligny, 1998) and has also been shown to play a role
in the allocation of attention in space, regardless of whether
movements are executed or not (例如, Gosbras & Paus,
2002; Collin, Cowey, Latto, & Marzi, 1982; Collin & Cowey,
1980).
In recent studies, TMS was applied to interfere tempo-
rarily with the activity of the FEF while participants were
engaged in a cuing task where a central cue predicted the
location of the upcoming target and the gaze was main-
tained at fixation. The results revealed that FEF TMS affects
the covert shift of attention with the control of either bilat-
eral or contralateral visual hemifield (例如, 史密斯, Jackson,
& Rorden, 2005; Grosbras & Paus, 2002). 例如,
Smith et al. (2005) showed that briefly disrupting the left
FEF eliminated the slow RTs associated with the invalid
strategic cues when the target appeared in the right visual
场地.
而且, there is evidence that TMS over the FEF of
the right hemisphere disrupts performance during con-
junction visual search tasks that require the covert alloca-
tion of attention to the target. In their study, Muggleton,
Juan, Cowey, and Walsh (2003) found that performance
was worse when the right FEF was stimulated at the same
time as the target display was presented. 有趣的是,
although many functions of the FEF and PPC seem to
重叠, the contribution of these areas to visual search
has been dissociated in the time course of their involve-
蒙特 (OʼShea, Muggleton, Cowey, & Walsh, 2006). Kalla,
Muggleton, Juan, Cowey, and Walsh (2008) reported that
TMS over the right FEF disrupted performance (caused a
reduction in d-prime scores) in an earlier period than the
right PPC in the same conjunction search task. The FEF
TMS effect occurred in a 0–40 msec range after stimulus
onset and the same effect was obtained by TMS over PPC
在 120 和 160 毫秒, a timing similar to that obtained
in previous studies ( Walsh, Ashbridge, & Cowey, 1998;
Ashbridge, Walsh, & Cowey, 1997).
The activity of the FEF in tasks in which eye movements
are not required has been interpreted in terms of the pre-
motor theory of attention (Rizzolatti et al., 1987). 然而,
this interpretation has also been challenged by more recent
findings supporting the idea that stimulus encoding and
response preparation can be decoupled in the FEF ( Juan
等人。, 2008; Juan, Shorter-Jacobi, & 沙尔, 2004; Sato &
沙尔, 2003; 汤普森, Bichot, & 沙尔, 1997). 在里面
macaque brain, two overlapping neuronal populations
have been identified, one population of visual neurons that
code visual attention and a second population of motor
neurons that code saccadic movements. Sato and Schall
(2003) recorded the activity of single neurons in the FEF
of macaque monkeys during the execution of a search task
with prosaccade or antisaccade responses. Two types of
neurons were distinguished responding differently to the
position of the target and to the endpoint of saccade.
此外, visual analysis and saccade preparation have
been dissociated in time in human FEF as indicated by
two distinct time points of TMS delivery that resulted in ele-
vated saccade latencies in a visually instructed prosaccade/
antisaccade task ( Juan et al., 2008).
On the basis of the reported functions of the FEF in
both orienting attention and transforming visual signals
into motor commands, a question arises concerning a pos-
sible role of the FEF in encoding irrelevant spatial infor-
mation for response priming in situations of conflict. 这
role of the FEF in spatial S-R conflict has never been inves-
tigated, 然而. Only one study (Rusconi, Bueti, Walsh, &
Butterworth, 2011) tested the effect of FEF TMS in the S-R
compatibility effect induced in numerical tasks (这
Spatial–Numerical Association of Response Codes [SNARC]
影响). Results revealed that the right FEF TMS after stimu-
lus onset interferes with contralateral orienting attention
in the representational space of numbers (IE。, the men-
tal number line) abolishing the compatibility effect for
small numbers (referring to the left portion of the mental
number line).
This study aimed at investigating the role of the FEF
in spatial conflict. To address this issue, we temporarily
1306
认知神经科学杂志
体积 24, 数字 6
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
4
/
6
2
4
1
/
3
6
0
/
5
1
1
3
9
0
4
5
3
/
8
1
2
7
6
7
哦
8
C
0
n
2
_
5
A
/
_
j
0
哦
0
C
2
n
2
3
_
A
p
_
d
0
0
乙
2
y
2
G
3
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
t
F
.
/
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
interfered with activity of the left and right FEF by applying
double-pulse TMS while participants were performing a
standard Simon task with lateralized stimuli. We predicted
that if the FEF play a role in automatic encoding of the
irrelevant stimulus spatial code, temporary interference
with the neural activity in this area should reduce the im-
pact of direct route processing on performance, thus reduc-
ing the correspondence effect. FEF TMS during the Simon
task was thus expected to interfere with the processing of
the irrelevant spatial attribute of the target, at least for stim-
uli appearing in the contralateral visual hemifield. 而且,
to evaluate the timing of the involvement of the FEF, 我们
applied magnetic stimulation at various time intervals after
stimulus onset. On the basis of previous data on visual
search tasks (例如, OʼShea et al., 2006), we predicted that
TMS would interfere in the Simon task in an early timing
after the onset of the visual stimulus.
方法
参加者
Fourteen healthy undergraduate students of University
College London (all right-handed; mean age = 23 年,
标准差= 2 年) with normal or corrected-to-normal visual
acuity took part in this experiment. All participants were
checked for TMS exclusion criteria ( Wassermann, 1998)
and gave their written informed consent before participa-
的. The local ethics committee approved the procedure.
The participants were reimbursed with cash for partici-
pating in the experiment. All participants were naive as to
the purpose of the study.
Apparatus and Stimuli
The participants were seated in a dimly lit and soundproof
room with their head held by a fixed head-and-chin rest at a
distance of 60 cm from a 17-in. monitor. 本实验
was programmed with Matlab Psychtoolbox (Brainard,
1997). The target stimulus was either a red or green square
subtending 1.5° × 1.5° (width × height) of visual angle
presented for 200 msec on a black background.
Procedure and Experimental Design
Each participant was tested in one experimental session
(1 天) lasting approximately 1.5 小时. Participants completed
a block of 20 practice trials followed by the experimental
blocks. A trial started with the presentation of a central fixa-
tion point (a circle subtending 0.5° of visual angle) lasting
400 毫秒. Then a stimulus was presented for 200 msec at a
visual angle of 4.5° either to the left or to the right of the
central fixation point. The intertrial interval was 3000 毫秒.
After an incorrect response, a tone (600 赫兹) was delivered
to provide feedback on the wrong answer. 参加者
were instructed to maintain their gaze on the fixation point
during the whole experimental session and were asked to
keep their eyes on the central point. Participants were in-
structed to respond, as quickly and as accurately as possi-
布莱, by pressing the left key (“F”) on the keyboard with the
index finger of the left hand when the target square was red
and the right key (“J”) with the index finger of the right
hand when the target square was green (数字 1). 这
maximum RT was 1200 毫秒.
The design involved within-participant manipulation of
sites of stimulation, S-R correspondence, and time intervals
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
4
/
6
2
4
1
/
3
6
0
/
5
1
1
3
9
0
4
5
3
/
8
1
2
7
6
7
哦
8
C
0
n
2
_
5
A
/
_
j
0
哦
0
C
2
n
2
3
_
A
p
_
d
0
0
乙
2
y
2
G
3
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
F
t
/
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
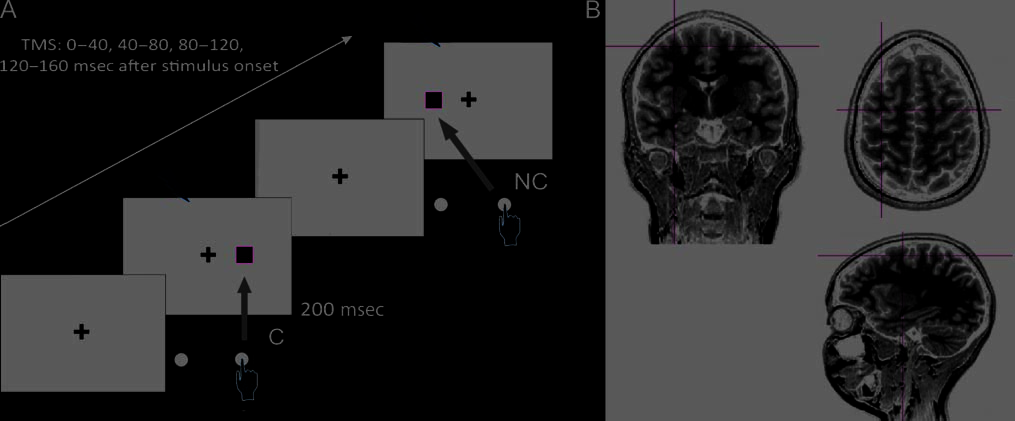
数字 1. A schematic representation of the trial sequence in the Simon task. (A) Paired-pulse TMS was randomly delivered in each trial at
different time intervals after the onset of the visual stimulus. (乙) The left FEF stimulation site is shown in coronal, sagittal, and transverse
sections of a participant MRI. The targeted location was identified for each participant by applying a normalization procedure. The left
FEF site corresponded to the Montreal Neurological Institute standard coordinates −32, −2, 57 and the right FEF to +32, −2, 57.
Bardi et al.
1307
between the onset of the target and the application of the
double-pulse TMS (SOA). Participants performed a baseline
condition without TMS of 64 trials followed by 12 experi-
mental blocks, four for each stimulation site (right FEF, 左边
FEF, and control site). Each block was composed of 64 试验
in which stimulus position (left or right), response position
(left or right), and TMS timing (0–40, 40–80, 80–120, 120–
180 毫秒) were fully crossed to produce the same num-
ber of trials for each possible combination presented in a
random order. A total of 256 trials were given for each
stimulation site. The order of the site of stimulation was
counterbalanced across participants.
Localization of Brain Targets for TMS Stimulation
Right and left FEF were localized by using the Brainsight
frameless stereotaxy system (Rogue Research, 蒙特利尔,
加拿大). The stimulation sites were identified on each
subjectʼs T1-weighted MRI scan and were then coregis-
tered with scalp coordinates over which TMS was delivered
(数字 1). The location of each subjectʼs right and left
FEF was determined according to anatomical landmarks.
Stimulation was applied over the posterior middle frontal
gyrus, just rostral of the junction of the precentral sulcus
and the superior frontal sulcus (Blanke et al., 2000). 我们
also checked the site of stimulation by referencing it to
each individual motor hand area (Ro, Cheifet, Ingle, Shoup,
& Rafal, 1999). With this method, 一般, TMS was ap-
plied 5 cm lateral of the sagittal midline and 3–4 cm rostral
of each subjectʼs motor hand area. This site corresponds
well with scalp coordinates used in other TMS studies of
the FEF (Kalla et al., 2008; Muggleton et al., 2003; Müri,
赫斯, & Meienberg, 1991). After normalization of the MRI
images to the Montreal Neurological Institute series aver-
年龄 (埃文斯, 柯林斯, & Holmes, 1996), mean coordinates
for the sites stimulated were ±32, −2, 57. These coordi-
nates correspond well with mean Talairach coordinates
for the FEF derived from a review of PET imaging studies
(Paus, 1996). As a control site, the point on the scalp cor-
responding to CPz in the 10–20 international system for
electrode placement was chosen. TMS over a control
site was chosen for the nonspecific effects of TMS, 这样的
as somatosensory and acoustic artifacts.
TMS Parameters
A Magstim Super Rapid2 machine (Magstim Company,
Dyfed, 英国) was used to deliver two magnetic
pulses at a frequency of 25 Hz during the task over the
right FEF, the left FEF, and CPz. A fixed level of 60% 的
the maximum machine output was employed. This level
was selected as it has previously proved effective in TMS
studies of FEF (例如, Kalla et al., 2008). A series of figure-
of-eight TMS coils (70 毫米) was used to apply stimula-
tion over the cortical sites of interest. Coils were cooled
before use to prevent overheating during a trial block
and were replaced after two blocks. For all the stimulation
站点, the experimenter clamped the coil with the handle
running in an anterior–posterior direction.
Control Experiment
Twelve healthy undergraduate students of University Col-
lege London (all right-handed; mean age = 23 年, 标准差=
1 年) took part in this experiment. Apparatus, 刺激,
and procedure were the same as in the main study. 每个
participant was tested in one experimental session (1 天)
lasting approximately 1 小时. Participants completed a prac-
tice block followed by the experimental blocks. The de-
sign involved the same within-participant manipulation
of factors as in the main experiment except for the num-
ber of TMS sites. Two sites were stimulated: the inter-
hemispheric control site used in the main experiment (这
point on the scalp corresponding to CPz in the 10–20 EEG
系统) and a left-lateralized sensorimotor (SM) 控制
site. For the SM site, the TMS coil was positioned at 2 厘米
posterior at the same laterality from each individual motor
hot spot. A similar procedure was adopted in previous
学习 (例如, Schluter, 拉什沃斯, Passingham, & 米尔斯,
1998). The TMS of this site did not induce a motor twitch
in the contralateral hand.
结果
Statistical analyses were performed on the magnitude of
the Simon effect (mean RTs for noncorresponding trials
minus mean RTs for corresponding trials) calculated as a
function of stimulus position (IE。, for left- and right-side
stimuli separately). The Simon effect for left-side stimuli
was obtained by subtracting left-side responses to left-side
刺激 (C) from right-side responses to left-side stimuli
(NC). The Simon effect for right-side stimuli was calculated
by subtracting right-side responses to right-side stimuli
(C) from left-side responses to right-side stimuli (NC). A
repeated-measures ANOVA was performed on the magni-
tude of the Simon effect with TMS Site (left FEF, right FEF,
CPz), TMS Timing Condition (0–40, 40–80, 80–120, 和
120–160 msec) and Stimulus Position (左边, 正确的) as within-
participants factors. Pairwise comparisons were then per-
formed to evaluate significant interactions.
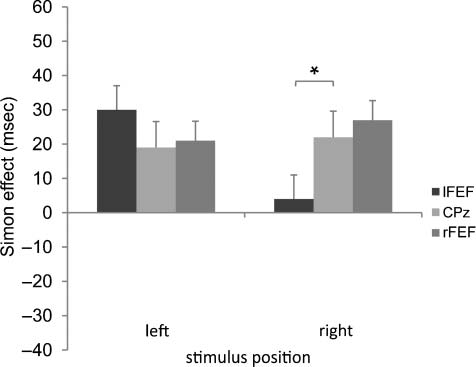
Results showed a significant interaction of Site × Stimu-
lus position [F(2, 26) = 4.74, p < .05, ηp
2 = .27]. Left FEF
TMS reduced the Simon effect for right-side stimuli com-
pared with the control site ( p < .05; Figure 2). The Simon
effect for right-side stimuli was 4 msec following TMS over
the left FEF (C, 411 msec; NC, 415 msec), whereas it was
27 msec following TMS over the control site (C, 394 msec;
NC, 421 msec).
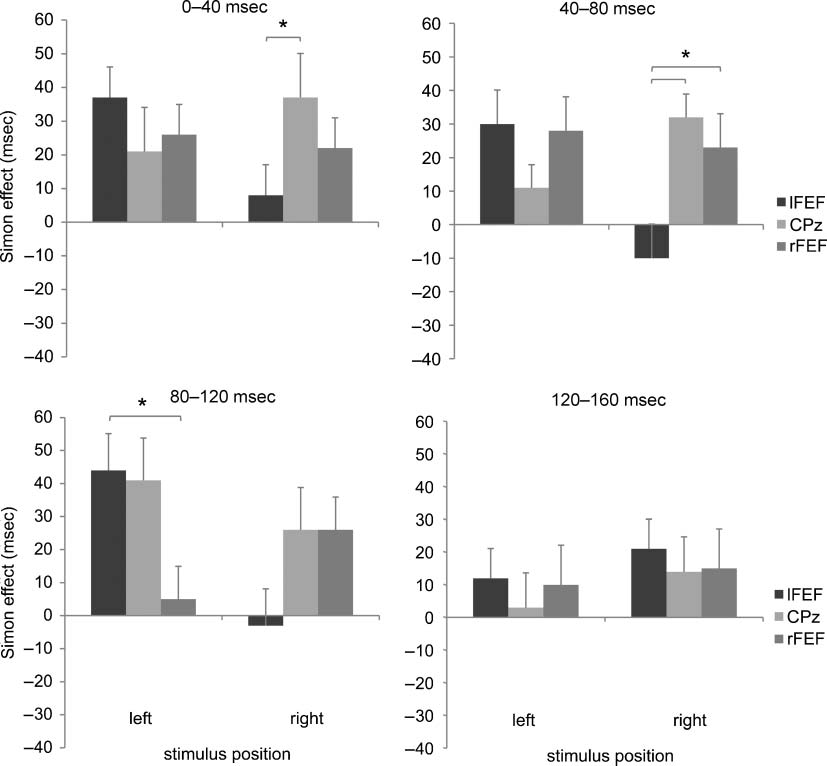
Furthermore, the effect of double-pulse TMS over the
left FEF was better understood by the Site × TMS Timing ×
Stimulus Position interaction [F(6, 78) = 2.538, p < .05,
η p
2 = .16]. This outcome revealed that the TMS inter-
ference had different effects across time windows. TMS
1308
Journal of Cognitive Neuroscience
Volume 24, Number 6
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
6
2
4
1
/
3
6
0
/
5
1
1
3
9
0
4
5
3
/
8
1
2
7
6
7
o
8
c
0
n
2
_
5
a
/
_
j
0
o
0
c
2
n
2
3
_
a
p
_
d
0
0
b
2
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
reduced the Simon effect for right-side stimuli, compared
with the control site, when applied over the left FEF in the
0–40 and 40–80 msec range after stimulus onset. In the first
time window, the Simon effect was reduced by TMS over
the left FEF (8 msec; C, 402 msec; NC, 410 msec) com-
pared with the control site (37 msec, p < .05; C, 379 msec;
NC, 416 msec). When TMS was applied at 40–80 msec,
the Simon effect was inverted (−10 msec; C, 416; NC,
406) as compared with the control site (32 msec; C, 383;
NC, 415). In the 80–120 msec time window, there was a
significant difference in the magnitude of the Simon effect
for left-side stimuli between the left and right FEF TMS
( p < .05). This effect was primarily driven by a reduction
of the Simon effect after right FEF TMS. The Simon effect
for left-side stimuli amounted to 5 msec after right FEF
TMS (C, 414 msec; NC, 419 msec) and to 44 msec after
the left FEF TMS (C, 402 msec; NC, 446 msec). No differ-
ence was detected between the Simon effect for left-side
stimuli and the Simon effect for the control site (41 msec;
C, 400 msec; NC, 441 msec; Figure 3).
Figure 2. The magnitude of the Simon effect (noncorresponding
minus corresponding trials) is shown separately for stimuli presented
in the left and right hemifield. Left FEF TMS resulted in a suppression
of the Simon effect for right-side stimuli.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
6
2
4
1
/
3
6
0
/
5
1
1
3
9
0
4
5
3
/
8
1
2
7
6
7
o
8
c
0
n
2
_
5
a
/
_
j
0
o
0
c
2
n
2
3
_
a
p
_
d
0
0
b
2
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 3. The magnitude of the Simon effect (noncorresponding–corresponding trials) is depicted separately for stimuli presented in the left
and right visual hemifields. Double-pulse TMS caused a suppression of the Simon effect for right-side stimuli when applied in the 0–40 time
windows after stimulus onset as compared with the control site. A reduction of the Simon effect has been observed for left-side stimuli when
TMS was applied in the 80–120 msec timing over the right FEF.
Bardi et al.
1309
Control Study
DISCUSSION
Results revealed that temporary interfering with activity
of the FEF modulates the magnitude of spatial conflict
(i.e., the Simon effect). The left FEF TMS caused a sup-
pression of the Simon effect for stimuli appearing in the
contralateral visual hemifield. This effect emerged when
TMS was applied early after stimulus onset (0–40, 40–
80 msec). Because the position of the TMS coil could
provide an additional spatial cue, one might argue that
this might have interfered with S-R correspondence. To
address this issue, we conducted a control experiment
in which an additional left-lateralized site was stimulated
to test possible nonspecific effects from left-lateralized
position of the TMS coil.
As in the main experiment, a repeated-measures ANOVA
was performed on the magnitude of the Simon effect with
TMS Site (left SM, CPz), TMS Timing Condition (0–40, 40–
80, 80–120, and 120–160 msec) and Stimulus Position
(left, right) as within-participants factors. Results revealed
no sources of variance. For CPz TMS, the Simon effect
amounted to 26 msec for left and right FEF TMS (C, 401 msec;
NC, 427 msec) and to 50 msec for right side stimuli (C,
388 msec; NC, 438 msec). For SM TMS, the Simon effect
amounted to 35 for the left-side stimuli (C, 392 msec;
NC, 427 msec) and to 42 for right-side stimuli (C, 389 msec;
NC, 431 msec). This outcome revealed that the TMS of
the left SM did not affect the magnitude of the Simon
effect. This result allows us to rule out a possible confound
of spatial cue introduced by the position of the coil over
one hemisphere.
Eye Monitoring
In this study, participants were instructed to maintain
fixation at the center of the screen throughout each
trial. To verify that TMS delivery over the FEF did not
induce involuntary gaze shifts during the task, eye posi-
tion was monitored in five additional participants in the
same experimental conditions as in the main experi-
ment. Participantsʼ gaze behavior was recorded using a
Tobii T120 eye-tracking screen run via the Tobii Studio
package. The eyes were monitored from the onset of
the fixation cross until the end of the stimulus presen-
tation. A gaze movement was determined as a shift of
at least 1.5° of visual angle from the central point. Each
participant underwent two blocks of task (120 trials per
block), in which paired-pulse TMS was applied over the
left FEF and CPz (0–40, 40–80, 80–120 msec SOA),
respectively. The rate for eye movements in valid trials
(after excluding eye blinks) was 0.6% for left FEF stim-
ulation and 0.7% for CPz stimulation. This outcome
is in line with data from previous studies that failed
to trigger eye movements by TMS of the frontal cor-
tex even when high stimulation intensities were used
(Müri et al., 1991; Wessel, Kcempf, Klostermann, & Moser,
1991).
Previous studies suggested that the FEF play an impor-
tant role in encoding stimulus position which may be
independent of the execution of eye movements. No
studies had investigated the contribution of the FEF in
a spatial conflict task, however. In the Simon task, a
conflict arises because irrelevant spatial information com-
petes for response selection. In the corresponding condi-
tion, responses are speeded up because stimulus spatial
code automatically activates a response code that overlaps
with the correct response code. In the noncorresponding
condition, spatial information competes with manual re-
sponse preparation generated on the basis of task instruc-
tions. The results of this study support the hypothesis
that the FEF contribute to the encoding of irrelevant spa-
tial information, which is the precondition of response
conflict in the Simon task. In line with our predictions,
paired-pulse TMS applied over the FEF caused a suppres-
sion of the Simon effect for visual stimuli presented in
the hemifield contralateral to the site of stimulation. This
effect was evident when TMS was applied to the left FEF in
the first three time windows (0–40, 40–80, 80–120 msec),
but this value was significant only in the earlier timings
(0–40, 40–80 msec) when compared with the vertex
stimulation. In contrast, a reduction of the Simon effect
was observed for right FEF TMS in the late 80–120 msec
time window.
Previous studied reported similar involvement of PPC in
the Simon task (Schiff et al., 2011; Rusconi et al., 2007).
Schiff et al. (2011) applied single-pulse TMS over the left
and right PPC and found a suppression of the Simon effect
when the stimulation was delivered in a 130–160 msec
range after stimulus onset. The present outcome suggests
that both FEF and PPC may be seen as components of
visuomotor processing through the direct route during
spatial conflict (De Jong et al., 1994). Interestingly, the con-
tribution of these areas to spatial conflict may be disso-
ciated in the time course of their involvement revealing
an early role of the FEF and a late role of PPC. A similar
dissociation was previously found in visual search tasks
(OʼShea et al., 2006; OʼShea, Muggleton, Cowey, & Walsh,
2004).
Our results can well accommodate both with a premotor
theory of attention and with the view that visual analysis
and saccade preparation are dissociated in human FEF
( Juan et al., 2008). In the Simon task, the target may auto-
matically activate a saccade preparation that competes with
endogenously generated manual responses. When the
TMS is applied over the FEF, the saccade preparation is
disrupted, so that the spatially corresponding response
receives neither facilitation nor interference from the com-
peting action. Our data support the idea that TMS specifi-
cally interfered with the stimulus spatial code, which
probably represents saccade parameters and interacts with
the selection of the manual response. Nevertheless, be-
cause the effect of FEF TMS in our task is evident along
1310
Journal of Cognitive Neuroscience
Volume 24, Number 6
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
6
2
4
1
/
3
6
0
/
5
1
1
3
9
0
4
5
3
/
8
1
2
7
6
7
o
8
c
0
n
2
_
5
a
/
_
j
0
o
0
c
2
n
2
3
_
a
p
_
d
0
0
b
2
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
different timings after stimulus onset, we cannot exclude
that TMS had interfered with both visual and motor neu-
rons in the FEF being both related to the automatic activa-
tion of the corresponding response in the Simon task. In
fact, a reduction of the Simon effect in specific conditions
may be obtained by preventing the shift of spatial attention
toward the position of the peripheral target (Nicoletti &
Umiltà, 1994) or inducing changes in the locus of attention
in paradigms with peripheral and central spatial precues
(see Van der Lubbe & Abrahamse, 2011).
The predominant role of the left hemisphere in the
Simon task, compared with the right-hemisphere domi-
nance found in SNARC task (Rusconi et al., 2011) provides
new insight into differences in neural circuits that may
underlie the Simon and SNARC effects (Rusconi, Umiltà,
& Galfano, 2006; Mapelli, Rusconi, & Umiltà, 2003). These
effects of compatibility share some important characteris-
tics: The fact that RTs are affected by task-irrelevant infor-
mation (stimulus position or magnitude) and the fact that
the information influencing response selection is spatial
in nature (perceptual or representational). However,
some important differences must be considered as the
processing of a stimulus appearing in the peripheral field
and the processing of central symbolic cues bearing a spa-
tial meaning (the number) could follow different paths
before converging and gaining access to the primary mo-
tor cortex for response execution (Rusconi et al., 2007).
In the Simon task, the spatial irrelevant information pro-
vided by stimulus position can be directly extracted and
has direct access to motor selection, whereas position
on the mental number line is not as directly accessible
to the viewers as the physical position of a stimulus. Dif-
ferences between the Simon effect and the SNARC effect
also manifest themselves in their behavioral patterns. For
instance, several studies reported that the size of the
Simon effect, with standard horizontal S-R mapping, de-
creases with longer RTs (Vallesi, Mapelli, Schiff, Amodio,
& Umiltà, 2005; Rubichi et al., 1997; Hommel, 1993),
whereas the size of SNARC effect is stable across differ-
ent RT ranges (e.g., Mapelli, Rusconi, & Umiltà, 2003).
According to Wascher and colleagues (2001), the time
course of the Simon and the SNARC effects (decreasing
vs. stable/increasing) reflects the presence or absence of
the automatic activation of the response. According to
these authors, only with spatial–anatomical S-R mapping
and visual stimuli, spatial parameters are processed within
privileged visuomotor pathways, automatically activating
the correspondent response. Spatial–anatomical S-R map-
ping refers to the overlap between stimulus and natural
hand position.
Our results, along with data from Rusconi et al. (2007),
support the view that cognitive mechanisms subtending
the Simon and the SNARC effects may be instantiated in
different neural circuits even if they may still converge
to a common area where the conflict is resolved. The
access to a representational space and the activation of
the response code in the SNARC effect seems to follow
a different temporal dynamics, which may be reflected in
the activation of different neural circuits.
Beside the need of further investigation, we speculate
that the automatic activation of the corresponding re-
sponse and the resulting time course of the Simon effect
with horizontal S-R mapping may be due in part to an
early activation of the left FEF, which may exert their in-
fluence by a direct connection with the left-lateralized
system for action selection in right-handed participants
(Rushworth, Johansen-Berg, Gobel, & Devlin, 2003;
Schluter, Krams, Rushworth, & Passingham, 2001).
The early involvement of the FEF of the left hemisphere
compared with the right hemisphere during spatial conflict
may be attributed to the dominance of the left hemisphere
in motor attention and action selection processes in right-
handed participants (Rushworth et al., 2003; Schluter et al.,
2001). In the Simon task, spatial information affects perfor-
mance at the response selection stage, and visuomotor
processing through the direct (automatic) pathway has
been proposed to be more efficient for the dominant
hand in right-handed participants (Spironelli, Tagliabue, &
Umiltà, 2009; Rubichi & Nicoletti, 2006).
In summary, this study provides the first evidence of the
involvement of the FEF in a conflict task. Our findings also
provide insight into the hemispheric asymmetries of the
FEF, which have been explored very little compared with
the parietal lobes. Regions in the left hemisphere such as
the SMA and the premotor cortex are important for atten-
tion to action and action selection (Rushworth et al., 2003;
Schluter et al., 2001). Our findings extend the idea of the
left-hemisphere lateralization of the motor network in action
selection by suggesting that the left FEF may constitute part
of the action selection network in the left hemisphere.
Acknowledgments
The authors are deeply indebted to Luca Semenzato for his sup-
port in the set-up of the experiments. We also thank Sami Schiff
for his invaluable comments on the results of this study.
Reprint requests should be sent to Lara Bardi, Department of
General Psychology, University of Padua, Via Venezia 8, 35131
Padua, Italy, or via e-mail: lara.bardi@unipd.it.
REFERENCES
Ashbridge, E., Walsh, V., & Cowey, A. (1997). Temporal
aspects of visual search studied by transcranial magnetic
stimulation. Neuropsychologia, 35, 1121–1131.
Blanke, O., Spinelli, L., Thut, G., Michel, C. M., Perrig, S.,
Landis, T., et al. (2000). Location of the human frontal eye
field as defined by electrical cortical stimulation: Anatomical,
functional and electrophysiological characteristics.
NeuroReport, 11, 1907–1913.
Brainard, D. H. (1997). The psychophysics toolbox. Spatial
Vision, 10, 433–436.
Cohen, J. D., Dunbar, K., & McClelland, J. L. (1990). On
the control of automatic processes: A parallel distributed
processing account of the Stroop effect. Psychological
Review, 97, 332–361.
Bardi et al.
1311
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
6
2
4
1
/
3
6
0
/
5
1
1
3
9
0
4
5
3
/
8
1
2
7
6
7
o
8
c
0
n
2
_
5
a
/
_
j
0
o
0
c
2
n
2
3
_
a
p
_
d
0
0
b
2
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Collin, N. G., & Cowey, A. (1980). The effect of ablation of
frontal eye-fields and superior colliculi on visual stability
and movement discrimination in rhesus monkeys.
Experimental Brain Research, 40, 251–260.
Collin, N. G., Cowey, A., Latto, R., & Marzi, C. (1982). The
role of frontal eye-fields and superior colliculi in visual
search and non-visual search in rhesus monkeys.
Behavioral Brain Research, 4, 177–193.
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed
and stimulus-driven attention in the brain. Nature Reviews
Neuroscience, 3, 201–215.
De Jong, J. R., Liang, C. C., & Lauber, E. (1994). Conditional
and unconditional automaticity: A dual-process model
of effects of spatial stimulus–response correspondence.
Journal of Experimental Psychology: Human Perception
and Performance, 20, 731–750.
Evans, A. C., Collins, D. L., & Holmes, C. J. (1996). In
J. C. Mazziotta (Ed.), Brain mapping: The methods
(pp. 343–361). San Diego, CA: Academic Press.
Gaymard, B., Ploner, C. J., Rivaud, S., Vermersch, A. I., &
Pierrot-Deseilligny, C. (1998). Cortical control of saccades.
Experimental Brain Research, 123, 159–163.
Goodale, M. A., & Milner, A. D. (1992). Separate visual pathways
for perception and action. Trends in Neuroscience, 15,
20–25.
Grosbras, M. H., & Paus, T. (2002). Transcranial magnetic
stimulation of the human frontal eye field: Effects on
visual perception and attention. Journal of Cognitive
Neuroscience, 14, 1109–1120.
Hommel, B. (1993). The role of attention for the Simon
effect. Psychological Research, 55, 208–222.
Hommel, B. (2011). The Simon effect as tool and heuristic.
Acta Psychologica, 136, 189–202.
Juan, C. H., Muggleton, N. G., Tzeng, O. J., Hung, D. L.,
Cowey, A., & Walsh, V. (2008). Segregation of visual
selection and saccades in human frontal eye fields.
Cerebral Cortex, 18, 2410–2415.
Juan, C. H., Shorter-Jacobi, S. M., & Schall, J. D. (2004).
Dissociation of spatial attention and saccade preparation.
Proceedings of the National Academy of Sciences, U.S.A.,
101, 15541–15544.
Kalla, R., Muggleton, N. G., Juan, C. H., Cowey, A., & Walsh, V.
(2008). The timing of the involvement of the frontal eye
fields and posterior parietal cortex in visual search.
NeuroReport, 19, 1067–1071.
Kornblum, S., Hasbroucq, T., & Osman, A. (1990).
Dimensional overlap: Cognitive basis for stimulus–response
compatibility—A model and taxonomy. Psychological
Review, 97, 253–270.
Kravitz, D. J., Saleem, K. S., Baker, C. I., & Mishkin, M. (2011).
A new neural framework for visuospatial processing.
Nature Reviews Neuroscience, 12, 217–230.
Lu, C. H., & Proctor, R. W. (1995). The influence of irrelevant
location information on performance—A review of the
Simon and spatial Stroop effects. Psychonomic Bulletin
& Review, 2, 174–207.
Mapelli, D., Rusconi, E., & Umiltà, C. (2003). The SNARC
effect: An instance of the Simon effect? Cognition, 88,
B1–B10.
Moore, T., & Farah, M. (2001). Control of eye movements
and spatial attention. Proceeding of the National Academy
of Sciences, U.S.A., 98, 1273–1276.
Muggleton, N. G., Juan, C. H., Cowey, A., & Walsh, V. (2003).
Human frontal eye fields and visual search. Journal of
Neurophysiology, 89, 3340–3343.
Müri, R. M., Hess, C. W., & Meienberg, O. (1991). Transcranial
stimulation of the human frontal eye field by magnetic
pulses. Experimental Brain Research, 86, 219–223.
Nicoletti, K. R., & Umiltà, C. A. (1994). Attentional shift
produce spatial stimulus codes. Psychological Research,
56, 144–150.
Olk, B., Chang, E., Kingstone, A., & Ro, T. (2006).
Modulation of antisaccades by transcranial magnetic
stimulation of the human frontal eye field. Cerebral
Cortex, 16, 76–82.
OʼShea, J., Muggleton, N. G., Cowey, A., & Walsh, V. (2004).
Timing of target discrimination in human frontal eye fields.
Journal of Cognitive Neuroscience, 16, 1060–1067.
OʼShea, J., Muggleton, N. G., Cowey, A., & Walsh, V. (2006).
On the roles of the human frontal eye fields and parietal
cortex in visual search. Visual Cognition, 14, 934–957.
Paus, T. (1996). Location and function of the human frontal
eye-field: A selective review. Neuropsychologia, 34, 475–483.
Rizzolatti, G., Riggio, L., Dascola, I., & Umiltà, C. (1987).
Reorienting attention across the horizontal and vertical
meridians: Evidence in favor of a premotor theory of
attention. Neuropsychologia, 25, 31–40.
Ro, T., Cheifet, S., Ingle, H., Shoup, R., & Rafal, R. (1999).
Localization of the human frontal eye fields and motor hand
area with transcranial magnetic stimulation and magnetic
resonance imaging. Neuropsychologia, 37, 225–231.
Rubichi, S., & Nicoletti, R. (2006). The Simon effect and
handedness: Evidence for a dominant-hand attentional
bias in spatial coding. Perception & Psychophysics, 68,
1059–1069.
Rubichi, S., Nicoletti, R., Iani, C., & Umiltà, C. (1997).
The Simon effect occurs relative to the direction of an
attention shift. Journal of Experimental Psychology:
Human Perception and Performance, 23, 1353–1364.
Rusconi, E., Bueti, D., Walsh, V., & Butterworth, B. (2011).
Contribution of the frontal cortex to the spatial
representation of number. Cortex, 47, 2–13.
Rusconi, E., Turatto, M., & Umiltà, C. (2007). Two orienting
mechanisms in posterior parietal lobule: An rTMS study of
the Simon and SNARC effects. Cognitive Neuropsychology,
24, 373–392.
Rusconi, E., Umiltà, C., & Galfano, G. (2006). Breaking ranks:
Space and number may march to the beat of a different
drum. Cortex, 42, 1124–1127.
Rushworth, M. F., Johansen-Berg, H., Gobel, S. M., & Devlin,
J. T. (2003). The left parietal and premotor cortices: Motor
attention and selection. Neuroimage, 20(Suppl. 1), S89–S100.
Sato, T. R., & Schall, J. D. (2003). Effects of stimulus–response
compatibility on neural selection in frontal eye fields.
Neuron, 38, 637–648.
Schiff, S., Bardi, L., Basso, D., & Mapelli, D (2011). Timing
spatial conflict within the parietal cortex. Journal of
Cognitive Neuroscience, 23, 3998–4007.
Schluter, N. D., Krams, M., Rushworth, M. F., & Passingham,
R. E. (2001). Cerebral dominance for action in the human
brain: The selection of actions. Neuropsychologia, 39,
105–113.
Schluter, N. D., Rushworth, M. F., Passingham, R. E., & Mills,
K. R. (1998). Temporary interference in human lateral
premotor cortex suggests dominance for the selection
of movements. A study using transcranial magnetic
stimulation. Brain, 121, 785–799.
Simon, J. R., & Rudell, A. P. (1967). Auditory S-R compatibility:
The effect of an irrelevant cue on information processing.
Journal of Applied Psychology, 51, 300–304.
Smith, D. T., Jackson, S. R., & Rorden, J. C. (2005). Transcranial
magnetic stimulation of the left human frontal eye fields
eliminates the cost of invalid endogenous cues.
Neuropsychologia, 43, 1288–1296.
Spironelli, C., Tagliabue, M., & Umiltà, C. (2009). Response
selection and attention orienting: A computational model
1312
Journal of Cognitive Neuroscience
Volume 24, Number 6
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
6
2
4
1
/
3
6
0
/
5
1
1
3
9
0
4
5
3
/
8
1
2
7
6
7
o
8
c
0
n
2
_
5
a
/
_
j
0
o
0
c
2
n
2
3
_
a
p
_
d
0
0
b
2
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
of Simon effect asymmetries. Experimental Psychology,
56, 274–282.
Stürmer, B., Redlich, M., Irlbacher, K., & Brandt, S. (2007).
Executive control over response priming and conflict:
A transcranial magnetic stimulation study. Experimental
Brain Research, 183, 329–339.
Tagliabue, M., Zorzi, M., Umiltà, C., & Bassignani, F. (2000).
The role of long-term-memory and short-term-memory
links in the Simon effect. Journal of Experimental
Psychology: Human Perception and Performance, 26,
648–670.
Thompson, K. G., Bichot, N. P., & Schall, J. D. (1997).
Dissociation of visual discrimination from saccade
programming in macaque frontal eye field. Journal
of Neurophysiology, 77, 1046–1050.
Umiltà, C. A., & Nicoletti, R. (1990). Spatial S-R compatibility.
In R. W. Proctor & T. G. Reeve (Eds.), Stimulus–response
compatibility: An integrated perspective (pp. 89–116).
Amsterdam: North Holland.
Umiltà, C. A., Riggio, I., Dascola, I., & Rizzolatti, G. (1991).
Differential effects of central and peripheral cues on the
reorienting of spatial attention. European Journal of
Cognitive Psychology, 3, 247–267.
Vallesi, A., Mapelli, D., Schiff, S., Amodio, P., & Umiltà, C.
(2005). Horizontal and vertical Simon effect: Different
underlying mechanisms? Cognition, 96, B33–B43.
Van der Lubbe, R. H. J., & Abrahamse, E. L. (2011). The
premotor theory of attention and the Simon effect.
Acta Psychologica, 136, 259–264.
Walsh, V., Ashbridge, E., & Cowey, A. (1998). Cortical plasticity
in perceptual learning demonstrated by transcranial magnetic
stimulation. Neuropsychologia, 36, 45–49.
Wascher, E., Schatz, U., Kuder, T., & Verleger, R. (2001).
Validity and boundary conditions of automatic response
activation in the Simon task. Journal of Experimental
Psychology: Human Perception and Performance, 27,
731–751.
Wassermann, E. M. (1998). Risk and safety of repetitive
transcranial magnetic stimulation: Report and suggested
guidelines from the International Workshop on the
Safety of Repetitive Transcranial Magnetic Stimulation,
June 5–7, 1996. Electroencephalography and Clinical
Neurophysiology, 10, 1–16.
Wessel, K., Kcempf, D., Klostermann, W., & Moser, A. (1991).
Lack of oculomotor response after transcranial magnetic
stimulation. Neuro-ophthalmology, 11, 199–208.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
4
/
6
2
4
1
/
3
6
0
/
5
1
1
3
9
0
4
5
3
/
8
1
2
7
6
7
o
8
c
0
n
2
_
5
a
/
_
j
0
o
0
c
2
n
2
3
_
a
p
_
d
0
0
b
2
y
2
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Bardi et al.
1313