The Timing of Anterior Temporal Lobe Involvement
in Semantic Processing
丽贝卡·L. Jackson, Matthew A. Lambon Ralph, and Gorana Pobric
抽象的
■ Despite indications that regions within the anterior tempo-
ral lobe (ATL) might make a crucial contribution to pan-modal
语义表示, to date there have been no investiga-
tions of when during semantic processing the ATL plays a crit-
ical role. To test the timing of the ATL involvement in semantic
加工, we studied the effect of double-pulse TMS on
behavioral responses in semantic and difficulty-matched con-
trol tasks. Chronometric TMS was delivered over the left ATL
(10 mm from the tip of the temporal pole along the middle
temporal gyrus). During each trial, two pulses of TMS (40 毫秒
apart) were delivered either at baseline (before stimulus pre-
sentation) or at one of the experimental time points 100, 250,
400, 和 800 msec poststimulus onset. A significant disruption
to performance was identified from 400 msec on the semantic
task but not on the control assessment. Our results not only
reinforce the key role of the left ATL in semantic representa-
tion but also indicate that its contribution is especially impor-
tant around 400 msec poststimulus onset. 一起, these facts
suggest that the ATL may be one of the neural sources of the
N400 ERP component. ■
介绍
The anterior temporal lobe (ATL) appears to make a crit-
ical contribution to pan-modal semantic representation
(Lambon Ralph, 2014; Lambon Ralph, 智者, 琼斯, &
Mayberry, 2010; 帕特森, Nestor, & 罗杰斯, 2007), 但
little is known about the timing within this area. Neuro-
心理学, functional neuroimaging, and repetitive TMS
experiments have identified a three-part network respon-
sible for multimodal semantic representation and con-
控制, including the pFC, temporoparietal region, 和
bilateral ATL (Noonan, Jefferies, Visser, & Lambon Ralph,
2013; Visser, Jefferies, Embleton, & Lambon Ralph, 2012;
Visser & Lambon Ralph, 2011; Whitney, 柯克, O’Sullivan,
Lambon Ralph, & Jefferies, 2011; Binney, Embleton,
Jefferies, 派克, & Lambon Ralph, 2010; Hoffman,
Jefferies, & Lambon Ralph, 2010; Binder, Desai, 格雷夫斯,
& Conant, 2009; Pobric, Jefferies, & Lambon Ralph,
2007; Jefferies & Lambon Ralph, 2006; 瓦格纳, Pare-
Blagoev, 克拉克, & Poldrack, 2001). 相比之下, the tempo-
ral dynamics of processing within the ATL have not been
studied comprehensively. The temporal resolution of
fMRI studies is poor, and although electrophysiological
techniques have good temporal resolution, spatial resolu-
tion is sacrificed ( Walsh & Cowey, 2000). A long history
of EEG and MEG studies have related semantic process-
ing across a variety of modalities to a negative ERP, 这
N400, 成立 250 到 550 msec after stimulus onset (Kutas
& Federmeier, 2011乙). Although somewhat inconsistent,
attempts to localize the N400 have identified a number of
曼彻斯特大学
sources within the semantic network, including areas in
the temporal lobe (Kutas & Federmeier, 2011乙; Helenius,
Salmelin, Service, & Connolly, 1998; 麦卡锡, 高贵,
Bentin, & Spencer, 1995). Beyond N400-focused studies,
one MEG investigation showed converging auditory-
related and visually related activity for a semantic judg-
ment task within the ATL around 400 毫秒 (Marinkovic
等人。, 2003). 然而, other studies have suggested
much earlier semantic influences in reading and visual
object recognition (刘, Agam, Madsen, & Kreiman,
2009; Pulvermuller, Shtyrov, & Hauk, 2009; Bar et al., 2006;
Halgren et al., 2002). 最后, a recent MEG investigation
found that these differential timing effects may be related
to the precision of semantic activation required, with ear-
lier (120 毫秒) synchronization between anterior and pos-
terior temporal regions for domain-level picture–name
verification and later (260 毫秒) for basic level decisions
(克拉克, 泰勒, & Tyler, 2011).
In contrast to electrophysiological approaches (在哪里
data are correlational and therefore could be epiphenom-
enal), TMS assesses the necessity of a specific brain area
for a task (Silvanto & Pascual-Leone, 2012; Sandrini, Umilta,
& Rusconi, 2011乙). This may be performed offline (在一个
rest period before the task) or online (during processing
within a trial; Sandrini, Umilta, & Rusconi, 2011A; Pascual-
莱昂内, Walsh, & Rothwell, 2000). Although the effects of
online TMS are more subtle, they allow increased temporal
specificity ( Walsh & Cowey, 2000). One form of online
TMS involves varying the time at which the pulse is deliv-
ered during the trial. 像这样, chronometric TMS (cTMS)
assesses when neural activity within a specific brain area is
critical for a task (Silvanto & Pascual-Leone, 2012; Sandrini
© 2015 根据知识共享署名发布 3.0
Unported (抄送 3.0) 执照
认知神经科学杂志 27:7, PP. 1388–1396
土井:10.1162/jocn_a_00788
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
7
/
7
2
7
1
/
3
7
8
/
8
1
1
3
9
8
4
8
9
/
2
1
9
7
6
8
哦
2
C
9
n
2
_
0
A
/
_
j
0
哦
0
C
7
n
8
8
_
A
p
_
d
0
0
乙
7
y
8
G
8
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
t
.
F
/
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
等人。, 2011乙). 因此, the time at which there is a direct rela-
tionship between the activity of a specific region and a
particular behavior may be identified (Silvanto & Pascual-
莱昂内, 2012; Duncan, Pattamadilok, & Devlin, 2010).
Concurrent imaging suggests that the disruption caused
by a TMS pulse has an effect on activity lasting around
10–30 msec ( Walsh & Cowey, 2000; Ilmoniemi et al.,
1997). 相似地, the temporal specificity of cTMS is
considered to be around 10–20 msec ( Walsh & Cowey,
2000). This specificity has allowed detection of behavioral
effects in time windows separated on the order of 10 毫秒.
This feature of cTMS allows processing to be tracked in
detail over time, either within one area or in different re-
祇翁 (Schuhmann, Schiller, Goebel, & Sack, 2012; Pitcher,
Walsh, Yovel, & Duchaine, 2007; Jahanshahi & Dirnberger,
1999; Schulter, 拉什沃斯, 米尔斯, & Passingham, 1999; Terao
等人。, 1998).
This study investigated the timing of ATL involvement
in semantic processing by applying TMS in a chronomet-
ric fashion. Double-pulse TMS was applied to the left ATL
during a synonym judgment task and a nonsemantic
control task (to assess for any nonspecific effects of
ATL cTMS and to test if the cTMS effect is specific to
semantic processing). This allowed identification of the
time at which the ATL is crucial for semantic processing,
hypothesized to be around 400 msec from stimulus
onset. The ATL may also be active at other time points,
例如, by contributing to early top–down process-
英, but it is unclear whether this activation is vital for
semantic processing.
方法
参加者
Fifteen healthy, 本国的, English-speaking volunteers with
normal or corrected-to-normal vision (seven women; 意思是
age = 24.39 年, 标准差= 5.98 年) completed the exper-
iment, which was approved by the local ethics board.
Materials
The synonym judgment task was adapted from previous
ATL offline rTMS, neuropsychological, and fMRI studies
(Lambon Ralph, Ehsan, 贝克, & 罗杰斯, 2012; Binney
等人。, 2010; Jefferies, 帕特森, 琼斯, & Lambon Ralph,
2009; Pobric et al., 2007). This paradigm has proved to be
a sensitive probe of semantic processing and has gen-
erated convergent cross-methodology evidence for the
selective role of the ATL in semantic processing. 这些
studies have not provided information, 然而, 关于
the time course of this ATL semantic computation (这
aim of the current study). As dual-pulse TMS has a small
effect size, we picked a task known to be sensitive to
semantic impairment and tried to maximize the number
of trials per condition. The TMS task was developed in a
series of steps to meet these design goals but to maintain
a minimal level of accuracy and thus relatively stable item
RT. In the first step, we selected 300 low-imageability
(target words mean imageability = 265.95, 标准差= 59.30,
using Bird, Franklin, & Howard’s, 2001, ratings) 和
low-frequency (target words mean CELEX frequency =
9.85, 标准差= 5.35) words because past studies have shown
them to be more sensitive to ATL rTMS, to the mild
semantic impairment in patients with unilateral ATL
resection, and to lead to poorer performance in SD
患者 (Hoffman, 琼斯, & Ralph, 2013; Lambon Ralph
等人。, 2012; Hoffman & Lambon Ralph, 2011; Jefferies
等人。, 2009; Lambon Ralph, Pobric, & Jefferies, 2009;
Pobric, Lambon Ralph, & Jefferies, 2009). In the second
step, we screened these items in a behavioral pilot of
eight postgraduate students and staff (six women; 意思是
age = 27 年). Only items that reached 75% 准确性
were selected for further consideration, as is commonly
performed in psycholinguistic studies (例如, 吉布森,
匹安多糖, & Fedorenko, 2011; Aoshima, Phillips, &
Weinberg, 2004; 吉布森 & Warren, 2004). 因此,
each task contained 200 experimental and 55 practice tri-
作为. A third and final screening of the items was necessary:
The TMS experiment employed predominantly under-
研究生 (seven women; mean age = 24.39 年)
who displayed a different speed–accuracy trade off on
the two tasks (they were around 10% faster on the syn-
onym task yet less accurate than the participants in the
second item screening step). 因此, the least accu-
rate trials were again screened at the 75% accuracy level
to remove the most errorful items and to maintain a low
level of noise within the associated RTs. Seventeen items
in the synonym judgment task and 23 in the control judg-
ment task had a mean accuracy below 75%. These items
were removed from the analyses.
Semantic Task
In each semantic trial, participants were presented with
three words: a probe (例如, rhythm), a target synonym
(例如, cadence), and an unrelated foil (例如, 网络).
Participants were asked to select which word was most
related in meaning to the probe. The two options were
matched within trials for frequency (foil mean 10.01,
标准差= 11.15, t(199) = −0.22, p > .05), imageability (foil
意思是 268.78, 标准差= 58.00, t(199) = −1.51, p > .05), 和
part of speech. The RT and accuracy was examined in a
pilot study (mean RT = 1265.4 毫秒, mean accuracy =
94.4%).
Intertask and Intratask Controls
The number judgment task was designed to match the
synonym test in overall difficulty (RTs and accuracy: pilot
学习, mean RT = 1295.38, mean accuracy = 96.2%).
Participants were asked to choose which three-digit num-
ber was closest to the probe in terms of numerical value
(例如, probe, 391; 目标, 379; foil, 377). This intertask
Jackson, Lambon Ralph, and Pobric
1389
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
7
/
7
2
7
1
/
3
7
8
/
8
1
1
3
9
8
4
8
9
/
2
1
9
7
6
8
哦
2
C
9
n
2
_
0
A
/
_
j
0
哦
0
C
7
n
8
8
_
A
p
_
d
0
0
乙
7
y
8
G
8
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
.
t
F
/
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
comparison provides, 所以, an important assessment
for the selectivity of semantic processing in the ATL and
also for any nonspecific effects of ATL cTMS. 下列的
previous rTMS examinations of ATL function (例如, Chiou,
Sowman, Etchell, & Rich, 2013; Pobric et al., 2007), 我们
expected a slowing of semantic decision times but no
effect on the control task—which would rule out any
alternative explanation of the semantic data in terms of
nonspecific effects of ATL cTMS. 此外, a further
potential advantage of cTMS over offline rTMS is that
cTMS can provide an “intratask control” if decision times
are slowed at some but not all of the probed time points
(例如, Duncan et al., 2010). 如上所述, we hypothe-
sized that cTMS might have its greatest effect on the
semantic task around 400 msec poststimulus presen-
站, but not at its onset. If this timing pattern was
coupled with no effect of TMS on the control task at
any time point, then the intertask and intratask data
would provide evidence for both task and time selectivity
of semantic processing in the ATL.
程序
A PC running ePrime (Psychology Software Tools, 公司,
Pittsburgh, PA) was used to present the items and record
participant’s responses. The participants completed two
sessions (one for each task) 至少 1 week apart. 这
order of sessions was counterbalanced across participants.
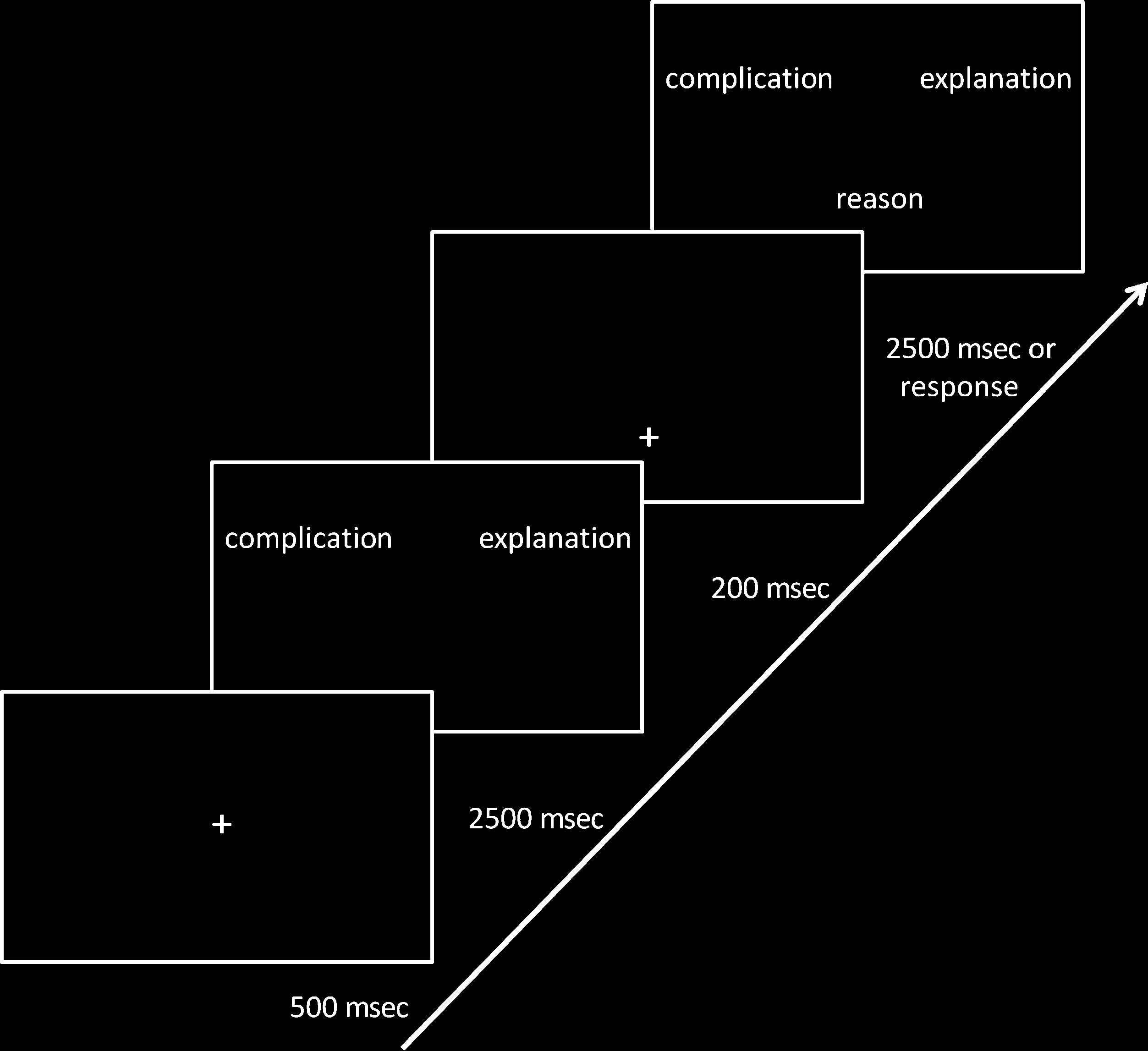
The order of trials was randomized. At the start of
every trial, a fixation point was presented in the middle
of the screen for 500 毫秒. Then the target and foil
choices appeared at the top of the screen and remained
为了 2500 毫秒. These items were replaced by a fixation
cross at the bottom of the screen, which remained for
200 msec before the probe item appeared in its place.
The target and foil item returned at the same time. 全部
three items remained on screen for 2500 msec or until
the participant’s button-press response (见图 1).
They were instructed to respond as quickly and accurate-
ly as possible. Displaying the foil and target before the
probe allowed us to measure RT and to compute stimu-
lation times against a single stimulus event (presentation
of the probe item) rather than the complexities involved
in timing from dual presentation of the target and foil
项目.
设计
The experiment employed a 2 × 5 repeated-measures
design with Task (semantic vs. 控制) and TMS time
(−40 msec vs. 100 msec vs. 250 msec vs. 400 msec vs.
800 毫秒) as the within-participant factors. The −40 msec
time point was employed as a baseline. This is superior to
comparison against a no-TMS condition, because TMS
can produce generalized alerting effects (Pobric et al.,
2007; Dräger, Breitenstein, Helmke, Kamping, & Knecht,
2004). Interleaving trials without TMS stimulation has also
been shown to affect the online TMS results and our own
pilot study found slowing in the trials immediately after
no TMS trials, perhaps reflecting increased attention to
the stimulation following its absence (Kapoula, 哪个,
Coubard, Daunys, & Orssaud, 2005). 因此, 我们
数字 1. The structure and
timing of a trial. 同时
任务, a target and a foil item
appeared after a fixation
叉. Participants had time
to read these items before
presentation of the probe
item in place of a fixation
叉. The participants were
then required to indicate
which item was closer
numerically or semantically.
1390
认知神经科学杂志
体积 27, 数字 7
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
7
/
7
2
7
1
/
3
7
8
/
8
1
1
3
9
8
4
8
9
/
2
1
9
7
6
8
哦
2
C
9
n
2
_
0
A
/
_
j
0
哦
0
C
7
n
8
8
_
A
p
_
d
0
0
乙
7
y
8
G
8
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
t
/
F
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
adopted two types of control/baseline: (A) comparison to
stimulation at −40 msec when semantic processing of the
probe item had not yet started and (乙) an RT-matched non-
semantic control task used to detect potential nonsemantic
variation of decision times after stimulation at different
SOAs. In both sessions, participants completed a practice
的 55 试验. They then completed 200 trials with TMS deliv-
ered at five different time points. These trials were split into
blocks of 40, allowing participants to take a break in
之间.
Stimulation Parameters
TMS was delivered using a Magstim Rapid2 stimulator
(Magstim Co., Whitland, 英国) and a figure-of-eight coil
with a diameter of 50 毫米. Stimulation was performed
在 100% of the individual’s motor threshold, 测量的
before the start of each session. The resting motor
threshold of the relaxed contralateral abductor pollicis
brevis muscle was measured as the lowest stimulation in-
tensity able to cause a visible twitch in the muscle 5 出去
的 10 次 (Sandrini et al., 2011乙). Motor thresholds
ranged from 40% 到 66% of stimulator output (mean =
57.2, 标准差= 7.06).
Double-pulse TMS was used as the effects of two
pulses summate while maintaining temporal specificity
(Pitcher et al., 2007). This inhibitory effect has been dem-
onstrated in a variety of domains including language
加工 (Sliwinska, Khadilkar, Campbell-Ratcliffe,
Quevenco, & Devlin, 2012; Duncan et al., 2010; O’Shea,
Johansen-Berg, Trief, Gobel, & 拉什沃斯, 2007; Pitcher
等人。, 2007; Juan & Walsh, 2003). Two TMS pulses were
delivered 40 msec apart in each trial. This gap is the smal-
lest possible with the TMS equipment used and has been
consistently employed in double pulse studies (Sliwinska
等人。, 2012; O’Shea et al., 2007; Pitcher et al., 2007; Juan
& Walsh, 2003). The two pulses were applied at −40 and
0 毫秒, 100 和 140 毫秒, 250 和 290 毫秒, 400 和
440 毫秒, 和 800 和 840 msec following presentation
of the probe number or word. Participants received stim-
ulation in every trial, but the stimulation time was ran-
domized. These stimulation times were designed to
sample a full range of potentially important times at
which crucial semantic processing might occur in the
ATL, including the hypothesized time of 400 msec as well
as earlier and later time points.
Selection of TMS Site
A Phillips MR Achieva scanner (Phillips Electronics,
阿姆斯特丹, 荷兰人) was used to acquire high-
resolution T1-weighted anatomical images of each par-
ticipant. The scan had an in-plane resolution of 1 毫米
with a slice thickness of 1.8 毫米. The acquisition matrix
曾是 256 × 256 voxels. Full head coverage was maintained,
causing the number of slices acquired to vary depending
on head size.
An Ascension Minibird magnetic tracking system
(Ascension Technology Co., 伯灵顿, VT) was used
to coregister the participant’s scalp and T1-weighted
MRI scan on MRIreg (www.MRIcro.com/mrireg.html).
The individual’s ATL coordinates were determined by
measuring 10 mm posterior along the middle temporal
gyrus from the tip of the left temporal pole. The average
MNI coordinates were [−50, 12, −29].
Every effort was taken to minimize the potential dis-
comfort of stimulating the ATL. Following the procedures
developed in our previous studies (Pobric, Jefferies, &
Lambon Ralph, 2010A; Pobric et al., 2007), coil orienta-
tion was manipulated for maximum comfort and stimulus
intensity was reduced if the participant considered the
stimulation unpleasant. 如上所述, nonspecific
effects of online TMS (such as muscle twitches) on RTs
were evaluated using the difficulty-matched nonsemantic
control task as well as intratask comparisons (不同的
time points during the semantic task).
分析
A composite RT–accuracy measure was used as the pri-
mary indicator of overall performance, because it allows
for speed–accuracy trade-off variation across participants.
The measure is commonly used in experimental psychol-
ogy for this reason and is also useful in TMS studies
在哪里, across participants, the effects of TMS can be
found in RTs, 错误, 或两者 (卡塔内奥, Vecchi, Pascual-
莱昂内, & Silvanto, 2009; Chambers, Stokes, & Mattingley,
2004; Townsend & Ashby, 1983). Following the stan-
dard method, the composite measure was computed as
RT/accuracy (Townsend & Ashby, 1983). For each partici-
pant and each time point, trials with RTs more than 2 SDs
from the mean were considered outliers and removed,
causing a loss of 4.42% of the remaining semantic and
4.4% of the control trials.
The effects of double-pulse TMS at different time
points on the composite performance was assessed using
A 2 × 5 within-subject ANOVA with the repeated-measures
factors Task (synonym judgment, number judgment) 和
TMS time (−40, 100, 250, 400, 800 毫秒). To assess the
time points at which there was a greater effect of TMS on
the semantic task, the ANOVA was repeated for two sub-
sets of time points. The change in performance caused by
TMS at time points implicated in these ANOVAs was com-
puted by subtracting the individual’s mean composite
score at these time points from their mean composite
score at the baseline time point on the each task. 这
allowed comparison of the effect of TMS at each time
point on the two different tasks.
结果
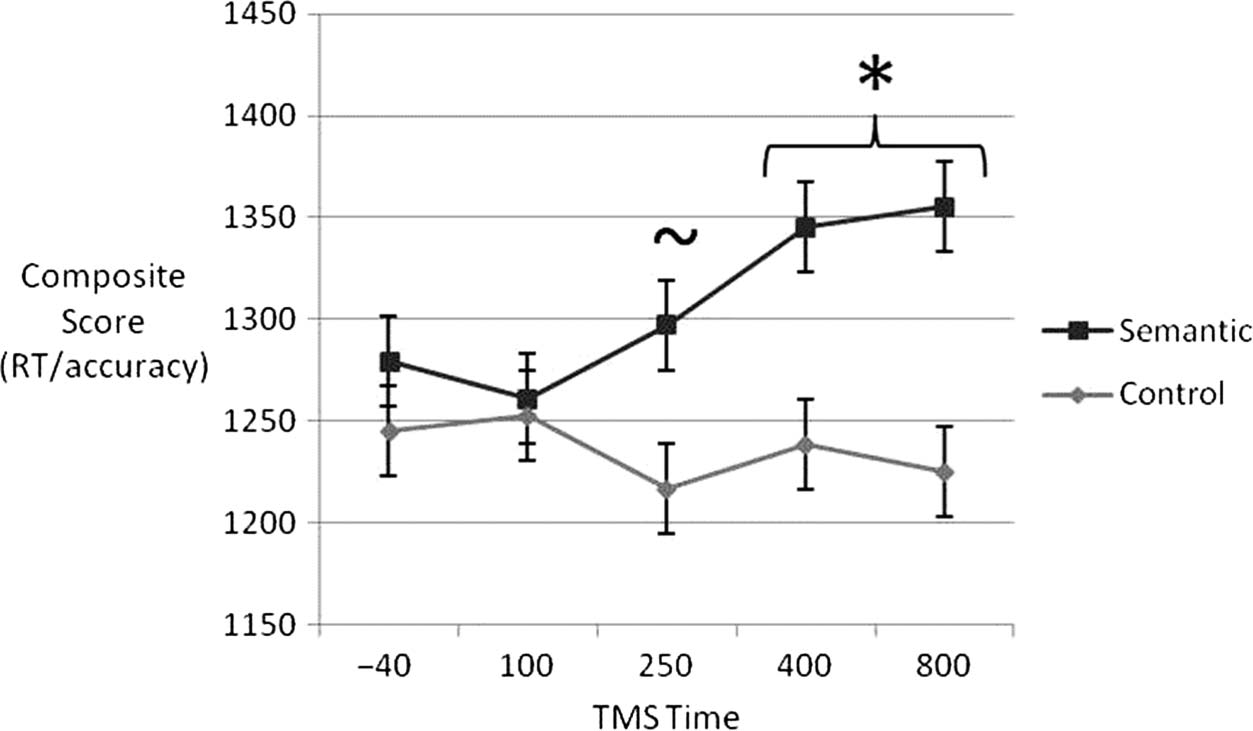
The results for the two tasks at each time point are
shown in Figure 2, with a clear effect of cTMS arising
Jackson, Lambon Ralph, and Pobric
1391
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
7
/
7
2
7
1
/
3
7
8
/
8
1
1
3
9
8
4
8
9
/
2
1
9
7
6
8
哦
2
C
9
n
2
_
0
A
/
_
j
0
哦
0
C
7
n
8
8
_
A
p
_
d
0
0
乙
7
y
8
G
8
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
F
/
.
t
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
数字 2. Composite score
(RT/accuracy) for the semantic
and control tasks at each
different TMS time point.
Error bars denote SEM,
corrected for a within-
participant design (Loftus &
Masson, 1994). A significant
TMS effect was found for
the later time points (denoted
by the asterisk). A borderline,
weak effect was detected at
250 毫秒 (see main text).
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
7
/
7
2
7
1
/
3
7
8
/
8
1
1
3
9
8
4
8
9
/
2
1
9
7
6
8
哦
2
C
9
n
2
_
0
A
/
_
j
0
哦
0
C
7
n
8
8
_
A
p
_
d
0
0
乙
7
y
8
G
8
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
F
.
t
/
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
for the semantic task but no effect at any time point for
the control task. A 2 ( 任务; control vs. semantic) × 5
(Stimulation time point; −40 vs. 100 与. 250 与. 400 与.
800 毫秒) repeated-measures ANOVA found no signifi-
cant main effects, 但, 关键地, 有一个互动
of Task and TMS time point, F(4, 56) = 2.6, p = .046, par-
tial η2 = .156. The nature of this TMS interaction was con-
firmed in the following way. 正如预期的那样, given the pilot
data and careful RT matching of the tasks for general dif-
ficulty, performance was not significantly different be-
tween the two tasks at the −40 msec baseline time
观点, t(14) = −.632, p = .538, two-tailed; 因此, this time
point provided a suitable baseline reference to compare
changes in performance across the other time points. 乙酰胆碱-
cordingly, the relative effect of cTMS at each time point was
extracted by subtracting the baseline (−40 msec) compos-
ite score from the composite score at each experimental
time point. The two tasks were then compared directly
by computing direction-specific, one-tailed t tests as we ex-
pected ATL TMS to slow semantic decision times, as found
in numerous previous ATL offline rTMS studies (例如, Chiou
等人。, 2013; Pobric et al., 2007). As is clear from Figure 2,
this analysis confirmed that there was significant slowing of
the semantic over control task at 400 毫秒, t(14) = −2.101,
p = .027, 和 800 毫秒, t(14) = −2.038, p = .030. 差异-
ference at 250 msec was of borderline significance ( p = .15).
There were no significant effects at any of the other time
点 ( p > .05).
A second additional analysis confirmed these same
结果. We used an interaction contrast to confirm the
significant effects of cTMS at 400 和 800 msec on the
semantic task alone. 具体来说, we averaged the data
across these two time points for each task and contrasted
them against the combined data for the earlier time
点. 再次, following the pattern shown in Figure 2,
there was no change in performance on the control task
between the earlier and later time points, t(14) = .224,
p = .826, whereas there was a significant slowing for
the semantic task, t(14) = 9.536, p < .001. We also for-
mally compared the early-to-late changes in performance
across the two tasks and confirmed that the early-to-late
slowing on the semantic task was significantly greater
than the null effect on the control task, t(14) = 4.891,
p < .001.
Accuracy and RT scores were also analyzed separately
(see Table 1). Accuracy was relatively stable across condi-
tions and tasks. The ANOVA on these data demonstrated
no significant results, although numerically the greatest
TMS effects were apparent at the 400 and 800 msec time
points in the semantic task. RT changes mirrored those
in the composite score, and the ANOVA on these data
confirmed the same interaction between Task and TMS
Table 1. Mean RT (msec) and Accuracy per TMS Time Point
TMS Time (msec)
RT (SD)
Accuracy (SD)
RT (SD)
Accuracy (SD)
Semantic Task
Control Task
−40
100
250
400
800
1177.31 (231.58)
1150.57 (261.07)
1178.56 (310.07)
1206.56 (291.9)
1234.15 (310.09)
.924 (.06)
.919 (.08)
.907 (.04)
.895 (.08)
.917 (.05)
1149.58 (297.16)
1141.11 (288.99)
1108.46 (292.72)
1125.34 (327.71)
1145.72 (303.90)
.922 (.05)
.911 (.05)
.906 (.06)
.912 (.08)
.939 (.04)
1392
Journal of Cognitive Neuroscience
Volume 27, Number 7
time, F(4, 56) = 2.556, p = .049, partial η2 = .154. We
explored this interaction in the same way as described
above for the composite score. The same pattern emerged,
with a significant slowing in the semantic compared to
control task at 400 msec, t(14) = −2.694, p = .009, and
800 msec, t(14) = −2.109, p = .026. As before, the TMS
effect at these time points was not significantly different
to the −40 msec baseline for the control task, t(14) =
−.127, p = .901, but was for the semantic task, t(14) =
−2.50, p = .025. Again, this difference between the two
tasks was significant, t(14) = 2.528, p = .024.
DISCUSSION
cTMS was used to elucidate the time at which the left ATL
is crucial for semantic processing. TMS had a significant
effect on semantic performance at 400 and 800 msec
poststimulus onset. No significant effects of TMS were
observed at any point during the control task. These
results add to the convergent evidence for a critical
role of the ATL in semantic processing (from neuro-
psychological, offline TMS, and neuroimaging studies:
Visser et al., 2012; Visser & Lambon Ralph, 2011; Binney
et al., 2010; Patterson et al., 2007; Pobric et al., 2007) and
reveal the temporal dynamics of this processing for the
first time.
MEG studies demonstrate two stages of processing in
semantic tasks: early processing within sensory areas and
then a large degree of interactivity between higher-order
pan-modal areas (Marinkovic et al., 2003; Halgren et al.,
2002; Dale et al., 2000). A translational phase between
modality-specific and pan-modal processing is thought
to start around 230 msec poststimulus presentation
(Marinkovic et al., 2003). ATL involvement has been iden-
tified within the second pan-modal stage, peaking around
400 msec (Shimotake et al., 2014; Marinkovic et al., 2003;
Halgren et al., 2002). The current study not only provides
convergent evidence for these hypotheses but goes fur-
ther to demonstrate that ATL activity at this time point is
critical for semantic processing, which neuroimaging
studies alone cannot establish. Although this has only
been demonstrated here in a single modality using visu-
ally presented abstract words, the evidence of a more
general necessity from rTMS and the demonstration of
a multimodal processing stage support the likelihood
that the critical role of the ATL occurs at a similar time
for different modalities. Theories that limit the role
of the ATL to social entities (e.g., Ross & Olson, 2010;
Olson, Ploaker, & Ezzyat, 2007; Moll, Zahn, de Oliveira-
Souza, Krueger, & Grafman, 2005), unique entities (e.g.,
Tranel, 2009), or combinatorial processes (Lau, Phillips,
& Poeppel, 2008; Hickok & Poeppel, 2007) cannot easily
explain the current and prior rTMS studies that used a
range of stimuli including single basic-level, nonsocial
concepts and found significant effects of TMS in all cases
(see Lambon Ralph, 2014, for a more detailed review and
discussion).
Our results indicate that there was a contribution from
the left ATL to semantic processing (as probed by syno-
nym judgements), which began to emerge at 250 msec
and became statistically reliable from 400 msec after stim-
ulus presentation. This fits the timing of the N400 in elec-
trophysiological studies, suggesting the ATL as one
potential source. Source localization of the EEG signal
in previous studies has generated conflicting results about
the N400 source, although a number of studies have
implicated a distributed frontotemporal network (Kutas
& Federmeier, 2011b; Lau et al., 2008; Van Petten & Luka,
2006). In addition, intracortical recordings have localized
an N400-like ERP to areas including anterior fusiform
gyrus and the temporal pole (McCarthy et al., 1995; Nobre
& McCarthy, 1995; Halgren, Baudena, Heit, Clarke, &
Marinkovic, 1994; Nobre, Allison, & McCarthy, 1994), and
MEG studies indicate a pan-modal role for the ATL at this
time point as well (Maess, Herrmann, Hahne, Nakamura, &
Friederici, 2006; Marinkovic et al., 2003; Halgren et al.,
2002). Finally, damage to left or right ATL can result in a
loss of the N400 (Kotz, Opitz, & Friederici, 2007).
One interpretation of the N400 is that it reflects semantic
access regardless of input modality (Kutas & Federmeier,
2011b; Kutas & Federmeier, 2000). This implies that the
areas involved are responsible for pan-modal processing
(Kutas & Federmeier, 2011b; Holcomb & Anderson,
1993). Within the hub-and-spoke semantic model, the ATL
hub is responsible for pan-modal representation, whereas
the “spokes” represent modality-specific information
(Pobric, Jefferies, & Lambon Ralph, 2010b; Patterson
et al., 2007). Thus, the modality-invariant processing within
the ATL could be reflected in the N400, a notion consistent
with the current cTMS findings.
Instead of viewing the N400 as representing semantic
processing per se, some researchers have suggested that
it is an index of violations of expectation, reflecting a pro-
cess whereby words or other constituent parts are inte-
grated into a context (Brown & Hagoort, 1993; Kutas &
Hillyard, 1980). This violation requires semantic and lex-
ical access, however, and so it is unclear that these expla-
nations are mutually exclusive. The left ATL may be one
of the N400 sources responsible for access and integra-
tion of pan-modal semantic representations. To assess
whether the ATL is a source of the N400, future studies
could assess the effect of ATL TMS in eradicating the
N400 component. The left ATL seems to remain involved
in the semantic task at 800 msec after stimulus presenta-
tion. This extends beyond the time of the N400, although
studies of the N400 typically use simpler stimuli and sin-
gle items (for a review, see Kutas & Federmeier, 2011a).
This longer involvement of the left ATL in semantic pro-
cessing might reflect continued semantic processing of
the items and comparison between them.
In keeping with most TMS explorations of higher cog-
nition, we observed an effect on RT and the combined
efficiency measure, rather than on accuracy alone (e.g.,
Sliwinska et al., 2012; Ishibashi, Lambon Ralph, Saito, &
Jackson, Lambon Ralph, and Pobric
1393
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
7
2
7
1
/
3
7
8
/
8
1
1
3
9
8
4
8
9
/
2
1
9
7
6
8
o
2
c
9
n
2
_
0
a
/
_
j
0
o
0
c
7
n
8
8
_
a
p
_
d
0
0
b
7
y
8
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Pobric, 2011; Duncan et al., 2010; Hoffman et al., 2010;
Stoeckel, Gough, Watkins, & Devlin, 2009; Gough, Nobre,
& Devlin, 2005; Dräger et al., 2004; Devlin, Matthews, &
Rushworth, 2003). Two possible interpretations of this
outcome are possible. One explanation is that double-
pulse TMS does not stop processing in the target region
but instead “injects noise” into the local computation
undertaken by that area, resulting in inefficient rather
than deficient processing. The second is that TMS might
halt local computation, but this is transient in nature and
limited in coverage (e.g., like a small lesion in some neuro-
psychological patients—which can generate changes in
RTs rather than accuracy). The second possibility is con-
sistent with the facts that TMS disruption is transient
(around 10–20 msec) and relatively focal (around 20 mm).
The precise mechanism by which TMS affects behavior
is not well understood. Both hypotheses have been
proposed, before; that is, TMS might inhibit activity (the
traditional “virtual lesion” view) or inject noise into the
system (Miniussi, Ruzzoli, & Walsh, n.d.; Sandrini et al.,
2011a; Harris, Clifford, & Miniussi, 2008; Walsh & Cowey,
2000). In this context, it is perhaps worth noting that
even the effect resulting from partial removal of neural
tissue can be considered in both ways: for example, partial
removal of units or connections in computational models
of semantic memory generates a combination of reduced
flow of activation and noisy processing (e.g., Schapiro,
McClelland, Welbourne, Rogers, & Lambon Ralph, 2013;
Lambon Ralph, Lowe, & Rogers, 2007). Even if one con-
ceives dual-pulse TMS as injecting transient noise into local
processing, then the main conclusions from this study
hold: namely, the left ATL is directly implicated in semantic
cognition and its involvement (at least in verbal tasks such
as synonym judgment) is maximally important 400 msec
after stimulus onset.
Finally, we note that, although we only obtained a
borderline, minimal effect of cTMS at 250 msec, some
neuroimaging studies have implicated this and earlier
time points. Superordinate semantic differences have
been shown in the phase-locking of the ATL from 120 msec
and in activity level from 170 msec (Clarke et al., 2011).
Similarly, intracortical electrode recordings have shown
category-sensitive responses in inferior anterior temporal
areas less than 200 msec poststimulus presentation (Liu
et al., 2009). There are at least three possible explanations.
First, electrophysiological data reflect changes in activity
level, whereas TMS might elucidate the time at which an
area becomes necessary (Walsh & Cowey, 2000). Thus, it
may be that the left ATL starts to become involved before
250 msec, but its contribution does not become crucial
until around 400 msec. A second possibility is that the early
activity may be critical in certain conditions, such as more
general semantic (domain/category) distinctions than
those probed in this cTMS study (Clarke et al., 2011) or
in tasks using impoverished stimuli that may promote
greater top–down processing (e.g., Bar et al., 2006). Finally,
an alternative possibility is that there is an earlier critical
phase of ATL involvement but that this is much smaller
and transient in nature, making it much harder to detect
with cTMS (the borderline effect observed at 250 msec is
consistent with this hypothesis).
Acknowledgments
R. L. J. was supported by an EPSRC-funded studentship. The
research was supported by an MRC program grant to M. A. L. R.
(MR/J004146/1).
Reprint requests should be sent to Dr. Gorana Pobric or Prof.
Matthew A. Lambon Ralph, Neuroscience and Aphasia Research
Unit, School of Psychological Sciences (Zochonis Building),
University of Manchester, Brunswick Street, Manchester, M13
9PL, United Kingdom, or via e-mail: gorana.pobric@manchester.
ac.uk, matt.lambon-ralph@manchester.ac.uk.
REFERENCES
Aoshima, S., Phillips, C., & Weinberg, A. (2004). Processing
filler-gap dependencies in a head-final language. Journal
of Memory and Language, 51, 23–54.
Bar, M., Kassam, K. S., Ghuman, A. S., Boshyan, J., Schmidt,
A. M., Dale, A. M., et al. (2006). Top–down facilitation of
visual recognition. Proceedings of the National Academy
of Sciences, U.S.A., 103, 449–454.
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L. (2009).
Where is the semantic system? A critical review and
meta-analysis of 120 functional neuroimaging studies.
Cerebral Cortex, 19, 2767–2796.
Binney, R. J., Embleton, K. V., Jefferies, E., Parker, G. J. M., &
Lambon Ralph, M. A. (2010). The ventral and inferolateral
aspects of the anterior temporal lobe are crucial in semantic
memory: Evidence from a novel direct comparison of
distortion-corrected fMRI, rTMS, and semantic dementia.
Cerebral Cortex, 20, 2728–2738.
Bird, H., Franklin, S., & Howard, D. (2001). Age of acquisition
and imageability ratings for a large set of words, including
verbs and function words. Behavior Research Methods
Instruments & Computers, 33, 73–79.
Brown, C., & Hagoort, P. (1993). The processing nature of the
N400—Evidence from masked priming. Journal of Cognitive
Neuroscience, 5, 34–44.
Cattaneo, Z., Vecchi, T., Pascual-Leone, A., & Silvanto, J. (2009).
Contrasting early visual cortical activation states causally
involved in visual imagery and short-term memory.
European Journal of Neuroscience, 30, 1393–1400.
Chambers, C. D., Stokes, M. G., & Mattingley, J. B. (2004).
Modality-specific control of strategic spatial attention in
parietal cortex. Neuron, 44, 925–930.
Chiou, R., Sowman, P. F., Etchell, A. C., & Rich, A. N. (2013).
A conceptual lemon: Theta burst stimulation to the left
anterior temporal lobe untangles object representation and
its canonical color. Journal of Cognitive Neuroscience, 26,
1066–1074.
Clarke, A., Taylor, K. I., & Tyler, L. K. (2011). The evolution of
meaning: Spatio-temporal dynamics of visual object recognition.
Journal of Cognitive Neuroscience, 23, 1887–1899.
Dale, A. M., Liu, A. K., Fischl, B. R., Buckner, R. L., Belliveau,
J. W., Lewine, J. D., et al. (2000). Dynamic statistical
parametric mapping: Combining fMRI and MEG for high-
resolution imaging of cortical activity. Neuron, 26, 55–67.
Devlin, J. T., Matthews, P. M., & Rushworth, M. F. S. (2003).
Semantic processing in the left inferior prefrontal cortex:
A combined functional magnetic resonance imaging and
1394
Journal of Cognitive Neuroscience
Volume 27, Number 7
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
7
2
7
1
/
3
7
8
/
8
1
1
3
9
8
4
8
9
/
2
1
9
7
6
8
o
2
c
9
n
2
_
0
a
/
_
j
0
o
0
c
7
n
8
8
_
a
p
_
d
0
0
b
7
y
8
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
transcranial magnetic stimulation study. Journal of Cognitive
Neuroscience, 15, 71–84.
Dräger, B., Breitenstein, C., Helmke, U., Kamping, S., & Knecht,
S. (2004). Specific and nonspecific effects of transcranial
magnetic stimulation on picture-word verification. European
Journal of Neuroscience, 20, 1681–1687.
Duncan, K. J., Pattamadilok, C., & Devlin, J. T. (2010).
Investigating occipito-temporal contributions to reading with
TMS. Journal of Cognitive Neuroscience, 22, 739–750.
Gibson, E., Piantadosi, S., & Fedorenko, K. (2011). Using
mechanical turk to obtain and analyze English acceptability
judgments. Language and Linguistics Compass, 5, 509–524.
Gibson, E., & Warren, T. (2004). Reading-time evidence for
intermediate linguistic structure in long-distance
dependencies. Syntax, 7, 55–78.
Gough, P. M., Nobre, A. C., & Devlin, J. T. (2005). Dissociating
linguistic processes in the left inferior frontal cortex with
transcranial magnetic stimulation. Journal of Neuroscience,
25, 8010–8016.
Halgren, E., Baudena, P., Heit, G., Clarke, M., & Marinkovic, K.
(1994). Spatiotemporal stages in face and word-processing.
1. Depth recorded potentials in the human occipital and
parietal lobes. Journal of Physiology-Paris, 88, 1–50.
Halgren, E., Dhond, R. P., Christensen, N., Van Petten, C.,
Marinkovic, K., Lewine, J. D., et al. (2002). N400-like
magnetoencephalography responses modulated by semantic
context, word frequency, and lexical class in sentences.
Neuroimage, 17, 1101–1116.
Harris, J. A., Clifford, C. W., & Miniussi, C. (2008). The
functional effect of transcranial magnetic stimulation: Signal
suppression or neural noise generation? Journal of Cognitive
Neuroscience, 20, 734–740.
Helenius, P., Salmelin, R., Service, E., & Connolly, J. F. (1998).
Distinct time courses of word and context comprehension in
the left temporal cortex. Brain, 121, 1133–1142.
Hickok, G., & Poeppel, D. (2007). The cortical organization
of speech processing. Nature Reviews Neuroscience, 8,
393–402.
Hoffman, P., Jefferies, E., & Lambon Ralph, M. A. (2010).
Ventrolateral prefrontal cortex plays an executive regulation
role in comprehension of abstract words: Convergent
neuropsychological and repetitive TMS evidence. Journal
of Neuroscience, 30, 15450–15456.
Hoffman, P., Jones, R. W., & Ralph, M. A. L. (2013). Be concrete
to be comprehended: Consistent imageability effects in
semantic dementia for nouns, verbs, synonyms and
associates. Cortex, 49, 1206–1218.
Hoffman, P., & Lambon Ralph, M. A. (2011). Reverse
concreteness effects are not a typical feature of semantic
dementia: Evidence for the hub-and-spoke model of
conceptual representation. Cerebral Cortex, 21, 2103–2112.
Holcomb, P. J., & Anderson, J. E. (1993). Cross-modal
semantic priming—A time-course analysis using
event-related brain potentials. Language and Cognitive
Processes, 8, 379–411.
Ilmoniemi, R. J., Virtanen, J., Ruohonen, J., Karhu, J., Aronen,
H. J., Näätänen, R., et al. (1997). Neuronal responses to
magnetic stimulation reveal cortical reactivity and connectivity.
NeuroReport, 8, 3537–3540.
Ishibashi, R., Lambon Ralph, M. A., Saito, S., & Pobric, G.
(2011). Different roles of lateral anterior temporal lobe
and inferior parietal lobule in coding function and
manipulation tool knowledge: Evidence from an rTMS
study. Neuropsychologia, 49, 1128–1135.
Jahanshahi, M., & Dirnberger, G. (1999). The left dorsolateral
prefrontal cortex and random generation of responses:
Studies with transcranial magnetic stimulation.
Neuropsychologia, 37, 181–190.
Jefferies, E., & Lambon Ralph, M. A. (2006). Semantic
impairment in stroke aphasia versus semantic dementia:
A case-series comparison. Brain, 129, 2132–2147.
Jefferies, E., Patterson, K., Jones, R. W., & Lambon Ralph, M. A.
(2009). Comprehension of concrete and abstract words in
semantic dementia. Neuropsychology, 23, 492–499.
Juan, C. H., & Walsh, V. (2003). Feedback to V1: A reverse
hierarchy in vision. Experimental Brain Research, 150,
259–263.
Kapoula, Z., Yang, Q., Coubard, O., Daunys, G., & Orssaud, C.
(2005). Contextual influence of TMS on the latency of
saccades and vergence. Neuroscience Letters, 376, 87–92.
Kotz, S. A., Opitz, B., & Friederici, A. D. (2007). ERP effects of
meaningful and non-meaningful sound processing in anterior
temporal patients. Restorative Neurology and Neuroscience,
25, 273–284.
Kutas, M., & Federmeier, K. D. (2000). Electrophysiology
reveals semantic memory use in language comprehension.
Trends in Cognitive Sciences, 4, 463–470.
Kutas, M., & Federmeier, K. D. (2011a). Thirty years and
counting: Finding meaning in the N400 component of the
event-related brain potential (ERP). In S. T. Fiske, D. L.
Schacter, & S. E. Taylor (Eds.), Annual review of psychology
(Vol. 62, pp. 621–647). Palo Alto, CA: Annual Review, Inc.
Kutas, M., & Federmeier, K. D. (2011b). Thirty years and
counting: Finding meaning in the N400 component of the
event-related brain potential (ERP). Annual Review of
Psychology, 62, 621–647.
Kutas, M., & Hillyard, S. A. (1980). Reading senseless sentences—
Brain potentials reflect semantic incongruity. Science, 207,
203–205.
Lambon Ralph, M. A. (2014). Neurocognitive insights on
conceptual knowledge and its breakdown. Royal Society
Proceedings B, 369, 20120392.
Lambon Ralph, M. A., Ehsan, S., Baker, G. A., & Rogers, T. T.
(2012). Semantic memory is impaired in patients with
unilateral anterior temporal lobe resection for temporal lobe
epilepsy. Brain, 135, 242–258.
Lambon Ralph, M. A., Lowe, C., & Rogers, T. T. (2007). Neural
basis of category-specific semantic deficits for living things:
Evidence from semantic dementia, HSVE and a neural
network model. Brain, 130, 1127–1137.
Lambon Ralph, M. A., Pobric, G., & Jefferies, E. (2009).
Conceptual knowledge is underpinned by the temporal pole
bilaterally: Convergent evidence from rTMS. Cerebral Cortex,
19, 832–838.
Lambon Ralph, M. A., Sage, K., Jones, R. W., & Mayberry, E. J.
(2010). Coherent concepts are computed in the anterior
temporal lobes. Proceedings of the National Academy of
Sciences, U.S.A., 107, 2717–2722.
Lau, E. F., Phillips, C., & Poeppel, D. (2008). A cortical network
for semantics: (De)constructing the N400. Nature Reviews
Neuroscience, 9, 920–933.
Liu, H. S., Agam, Y., Madsen, J. R., & Kreiman, G. (2009).
Timing, timing, timing: Fast decoding of object information
from intracranial field potentials in human visual cortex.
Neuron, 62, 281–290.
Loftus, G. R., & Masson, M. E. J. (1994). Using confidence
intervals in within-subject designs. Psychonomic Bulletin &
Review, 1, 476–490.
Maess, B., Herrmann, C. S., Hahne, A., Nakamura, A., &
Friederici, A. D. (2006). Localizing the distributed language
network responsible for the N400 measured by MEG during
auditory sentence processing. Brain Research, 1096, 163–172.
Marinkovic, K., Dhond, R. P., Dale, A. M., Glessner, M., Carr, V.,
& Halgren, E. (2003). Spatiotemporal dynamics of modality-
specific and supramodal word processing. Neuron, 38,
487–497.
Jackson, Lambon Ralph, and Pobric
1395
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
7
2
7
1
/
3
7
8
/
8
1
1
3
9
8
4
8
9
/
2
1
9
7
6
8
o
2
c
9
n
2
_
0
a
/
_
j
0
o
0
c
7
n
8
8
_
a
p
_
d
0
0
b
7
y
8
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
McCarthy, G., Nobre, A. C., Bentin, S., & Spencer, D. D.
Sandrini, M., Umilta, C., & Rusconi, E. (2011b). The use of
(1995). Language-related field potentials in the anterior-
medial temporal lobe. 1. Intracranial distribution and neural
generators. Journal of Neuroscience, 15, 1080–1089.
Miniussi, C., Ruzzoli, M., & Walsh, V. (n.d.). The mechanism of
transcranial magnetic stimulation in cognition. Cortex, 46,
128–130.
Moll, J., Zahn, R., de Oliveira-Souza, R., Krueger, F., & Grafman,
J. (2005). The neural basis of human moral cognition. Nature
Reviews Neuroscience, 6, 799–809.
Nobre, A. C., Allison, T., & McCarthy, G. (1994). Word
recognition in the human inferior temporal lobe. Nature,
372, 260–263.
Nobre, A. C., & McCarthy, G. (1995). Language-related field
potentials in the anterior-medial temporal-lobe. 2. Effects of
word type and semantic priming. Journal of Neuroscience,
15, 1090–1098.
Noonan, K. A., Jefferies, E., Visser, M., & Lambon Ralph, M. A.
(2013). Going beyond inferior prefrontal involvement in
semantic control: Evidence for the additional contribution of
dorsal angular gyrus and posterior middle temporal cortex.
Journal of Cognitive Neuroscience, 25, 1824–1850.
Olson, I. R., Ploaker, A., & Ezzyat, Y. (2007). The enigmatic
temporal pole: A review of findings on social and emotional
processing. Brain, 130, 1718–1731.
O’Shea, J., Johansen-Berg, H., Trief, D., Gobel, S., & Rushworth,
M. F. S. (2007). Functionally specific reorganisation in human
premotor reorganization cortex. Neuron, 54, 479–490.
Pascual-Leone, A., Walsh, V., & Rothwell, J. (2000). Transcranial
magnetic stimulation in cognitive neuroscience—Virtual
lesion, chronometry, and functional connectivity. Current
Opinion in Neurobiology, 10, 232–237.
Patterson, K., Nestor, P. J., & Rogers, T. T. (2007). Where do you
know what you know? The representation of semantic
knowledge in the human brain. Nature Reviews Neuroscience,
8, 976–987.
Pitcher, D., Walsh, V., Yovel, G., & Duchaine, B. (2007).
TMS evidence for the involvement of the right occipital
face area in early face processing. Current Biology, 17,
1568–1573.
Pobric, G., Jefferies, E., & Lambon Ralph, M. A. (2007). Anterior
temporal lobes mediate semantic representation: Mimicking
semantic dementia by using rTMS in normal participants.
Proceedings of the National Academy of Sciences, U.S.A.,
104, 20137–20141.
Pobric, G., Jefferies, E., & Lambon Ralph, M. A. (2010a). Amodal
semantic representations depend on both anterior temporal
lobes: Evidence from repetitive transcranial magnetic
stimulation. Neuropsychologia, 48, 1336–1342.
Pobric, G., Jefferies, E., & Lambon Ralph, M. A. (2010b).
Category-specific versus category-general semantic
impairment induced by transcranial magnetic stimulation.
Current Biology, 20, 964–968.
Pobric, G., Lambon Ralph, M. A., & Jefferies, E. (2009). The role
of the anterior temporal lobes in the comprehension of
concrete and abstract words: rTMS evidence. Cortex, 45,
1104–1110.
Pulvermuller, F., Shtyrov, Y., & Hauk, O. (2009). Understanding
in an instant: Neurophysiological evidence for mechanistic
language circuits in the brain. Brain and Language, 110,
81–94.
Ross, L. A., & Olson, I. R. (2010). Social cognition and the
anterior temporal lobes. Neuroimage, 49, 3452–3462.
Sandrini, M., Umilta, C., & Rusconi, E. (2011a). The use of
transcranial magnetic stimulation in cognitive neuroscience:
A new synthesis of methodological issues. Neuroscience and
Biobehavioral Reviews, 35, 516–536.
transcranial magnetic stimulation in cognitive neuroscience:
A new synthesis of methodological issues. Neuroscience and
Behavioural Reviews, 35, 516–536.
Schapiro, A. C., McClelland, J. L., Welbourne, S. R., Rogers, T. T.,
& Lambon Ralph, M. A. (2013). Why bilateral damage is worse
than unilateral damage to the brain. Journal of Cognitive
Neuroscience, 25, 2107–2123.
Schuhmann, T., Schiller, N. O., Goebel, R., & Sack, A. T. (2012).
Speaking of which: Dissecting the neurocognitive network of
language production in picture naming. Cerebral Cortex, 22,
701–709.
Schulter, N. D., Rushworth, M. F., Mills, K. R., & Passingham,
R. E. (1999). Signal-, set-, and movement-related activity in
the human premotor cortex. Neuropsychologia, 37, 233–243.
Shimotake, A., Matsumoto, R., Ueno, T., Kunieda, T., Saito, S.,
Hoffman, P., et al. (2014) Direct exploration of the ventral
anterior temporal lobe in semantic memory: Cortical
stimulation and local field potential evidence from subdural
grid electrodes. Cerebral Cortex. doi: 10.1093/cercor/bhu262.
Silvanto, J., & Pascual-Leone, A. (2012). Why the assessment
of causality in brain-behavior relations requires brain
stimulation. Journal of Cognitive Neuroscience, 24, 775–777.
Sliwinska, M. W., Khadilkar, M., Campbell-Ratcliffe, J.,
Quevenco, F., & Devlin, J. T. (2012). Early and sustained
supramarginal gyrus contributions to phonological
processing. Frontiers in Psychology, 3, 161.
Stoeckel, C., Gough, P. M., Watkins, K. E., & Devlin, J. T. (2009).
Supramarginal gyrus involvement in visual word recognition.
Cortex, 45, 1091–1096.
Terao, Y., Fukuda, H., Ugawa, Y., Hikosaka, O., Hanajima, R.,
Furubayashi, T., et al. (1998). Visaulisation of the information
flow through human occulomotor cortical regions by
transcranial magnetic stimulation. Journal of Neurophysiology,
80, 936–946.
Townsend, J. T., & Ashby, F. G. (1983). Stochastic modelling of
elementary psychological processes. London: Cambridge
University Press.
Tranel, D. (2009). The left temporal pole is important for
retrieving words for unique concrete entities. Aphasiology,
23, 867–884.
Van Petten, C., & Luka, B. J. (2006). Neural localization of
semantic context effects in electromagnetic and
hemodynamic studies. Brain and Language, 97, 279–293.
Visser, M., Jefferies, E., Embleton, K. V., & Lambon Ralph,
M. A. (2012). Both the middle temporal gyrus and the
ventral anterior temporal area are crucial for multimodal
semantic processing: Distortion-corrected fMRI evidence
for a double gradient of information convergence in the
temporal lobes. Journal of Cognitive Neuroscience, 24,
1766–1778.
Visser, M., & Lambon Ralph, M. A. (2011). Differential
contributions of bilateral ventral anterior temporal lobe
and left anterior superior temporal gyrus to semantic
processes. Journal of Cognitive Neuroscience, 23,
3121–3131.
Wagner, A. D., Pare-Blagoev, E. J., Clark, J., & Poldrack, R. A.
(2001). Recovering meaning: Left prefrontal cortex guides
controlled semantic retrieval. Neuron, 31, 329–338.
Walsh, V., & Cowey, A. (2000). Transcranial magnetic
stimulation and cognitive neuroscience. Nature Reviews
Neuroscience, 1, 73–79.
Whitney, C., Kirk, M., O’Sullivan, J., Lambon Ralph, M. A., &
Jefferies, E. (2011). The neural organization of semantic
control: TMS evidence for a distributed network in left
inferior frontal and posterior middle temporal gyrus.
Cerebral Cortex, 21, 1066–1075.
1396
Journal of Cognitive Neuroscience
Volume 27, Number 7
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
7
2
7
1
/
3
7
8
/
8
1
1
3
9
8
4
8
9
/
2
1
9
7
6
8
o
2
c
9
n
2
_
0
a
/
_
j
0
o
0
c
7
n
8
8
_
a
p
_
d
0
0
b
7
y
8
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1