The Formation of Symmetrical Gestalts Is
Task-Independent, but Can Be Enhanced
by Active Regularity Discrimination
Alexis D. J. Makin1, Giulia Rampone1, Amie Morris1, and Marco Bertamini1,2
抽象的
■ The brain can organize elements into perceptually mean-
ingful gestalts. Visual symmetry is a useful tool to study gestalt
形成, and we know that there are symmetry-sensitive re-
gions in the extrastriate cortex. 然而, it is unclear whether
symmetrical gestalt formation happens automatically, 什么-
ever the participant’s current task is. Does the visual brain
always organize and interpret the retinal image when possible,
or only when necessary? 为了测试这个, we recorded an ERP
called the sustained posterior negativity (SPN). SPN amplitude
increases with the proportion of symmetry in symmetry +
noise displays. We compared the SPN across five tasks with
different cognitive and perceptual demands. Contrary to our
预测, the SPN was the same across four of the five tasks
but selectively enhanced during active regularity discrimina-
的. 此外, during regularity discrimination, the SPN
was present on hit trials and false alarm trials but absent on
miss and correct rejection trials. We conclude that gestalt for-
mation is automatic and task-independent, although it occa-
sionally fails on miss trials. 然而, it can be enhanced by
attention to visual regularity. ■
介绍
The visual brain must organize and interpret the retinal
图像. We know many rules that govern perceptual orga-
nization (Wagemans et al., 2012; Hoffman, 1998), 但是
circumstances under which perceptual organization hap-
pens are unclear. Does the visual brain always organize
and interpret the image to the fullest extent possible,
or does it often leave aspects of the image uninterpreted
(瘫痪 & 罗尔夫塞玛, 2000; Mack & Rock, 1998;
Treisman & Gelade, 1980)?
This research program requires an objective way to
measure whether perceptual organization has happened
或不. An ERP component called the sustained posterior
negativity (SPN) provides this. Amplitude is more negative
at posterior electrodes when participants view regular or
symmetrical patterns where corresponding elements can
be grouped into an organized whole (马金, Wilton,
Pecchinenda, & Bertamini, 2012; Jacobsen & Höfel,
2003). fMRI has identified symmetry-related BOLD re-
sponses in the extrastriate cortex ( Van Meel, Baeck,
Gillebert, Wagemans, & Op de Beeck, 2019; Keefe et al.,
2018; Kohler, 克拉克, Yakovleva, 刘, & Norcia, 2016;
Sasaki, Vanduffel, Knutsen, Tyler, & Tootell, 2005; Tyler
等人。, 2005), and this extrastriate activity probably gener-
ates the SPN (Makin et al., 2016). We can therefore posit
that if an SPN is recorded at the scalp, then perceptual
1University of Liverpool, 2University of Padova
organization has happened in brain (Bertamini, Silvanto,
Norcia, 马金, & Wagemans, 2018).
The SPN is often similar when participants attend to
stimulus regularity or to a secondary stimulus dimension
such as color (马金, Rampone, & Bertamini, 2015; Höfel
& Jacobsen, 2007). This suggests symmetrical gestalts are
formed automatically, whatever the current task is. 如何-
曾经, Rampone, 马金, and Bertamini (2014) found that
the SPN was abolished when participants read super-
imposed negative words, and the BOLD response to sym-
metry is sometimes reduced during color discrimination
(Keefe et al., 2018; Sasaki et al., 2005). 此外, 任务
manipulations could be more consequential when sym-
metrically arranged elements are embedded among
randomly arranged noise elements (Gheorghiu, 王国,
Remkes, 李, & Rainville, 2016; Palumbo, Bertamini, &
马金, 2015; 巴洛 & Reeves, 1979). Unlike perfect sym-
metry, such intermediate symmetries may not be extracted
automatically in all tasks.
We measured the SPN at 20%, 40%, 60%, 80%, 和
100% symmetry (as difference from the waveform elicited
由 0% symmetry, random pattern). This variable is
termed PSYMM (Palumbo et al., 2015). There were five
任务, all of which involved a binary judgment (数字 1).
In the regularity task, participants judged whether patterns
had some regularity or no regularity. In the color task, par-
ticipants judged whether elements were light green or
dark green. In the sound/color task they judged the con-
gruence of the relationship between element color and
© 2019 麻省理工学院. Published under a
Creative Commons Attribution 4.0 国际的 (抄送 4.0) 执照.
认知神经科学杂志 32:2, PP. 353–366
https://doi.org/10.1162/jocn_a_01485
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
哦
H
w
t
n
t
p
哦
:
A
/
d
/
e
d
米
我
F
r
t
哦
p
米
r
C
H
.
p
s
我
我
d
v
我
r
e
e
r
C
t
C
.
米
H
A
我
e
r
d
.
你
C
哦
哦
米
C
n
/
j
A
哦
r
t
C
我
C
n
e
/
–
A
p
r
d
t
我
3
2
C
我
2
e
3
–
5
p
3
d
F
1
9
/
3
3
1
2
3
/
1
2
5
/
3
哦
5
C
3
n
/
_
A
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
哦
乙
C
y
n
G
_
你
A
e
_
s
0
t
1
哦
4
n
8
0
5
8
.
p
S
d
e
F
p
e
乙
米
y
乙
e
G
r
你
2
e
0
s
2
t
3
/
j
.
/
F
t
哦
n
0
5
中号
A
y
2
0
2
1
数字 1. Stimuli and
procedure. (A) Example dark
and light green pattern stimuli
(rows), with increasing PSYMM
(columns). These pattern types
were used in the regularity,
颜色, sound/color, 和
direction/color tasks. (乙)
Example pattern stimuli used in
the distribution task. (C) 审判
structure in the regularity task.
The other tasks were similar
except the response screen
prompted a different binary
判断 (light green…dark
绿色的, congruent…
incongruent, or uniform…
outside). (D) Example stimulus
with downward triangle from in
the direction/color task.
the pitch of a simultaneous sound (congruent = high
pitch and light green or low pitch and dark green, 然而
incongruent = high pitch and dark green or low pitch and
light green). 同样地, in the direction/color task, 他们
judged the relationship between element color and direc-
tion of a central triangle. 最后, in the distribution task,
participants judged whether elements were spread uni-
formly or concentrated around the periphery. The tasks
had different cognitive and perceptual requirements,
which could vary in the extent to which they take compu-
tational resources away from symmetrical gestalt forma-
的. 然而, all secondary tasks were relatively easy
and involved judgments about the pattern itself (相当
than another object elsewhere on the screen, 这是
beyond the scope of this study).
Three predictions were preregistered (https://aspredicted.
org/yy5gs.pdf). 第一的, we predicted the SPN to scale with
PSYMM in all tasks. This scaling was already found by
Palumbo et al. (2015), and it is strongly predicted by
psychophysics (巴洛 & Reeves, 1979) and theoretical
工作 (van der Helm, 2010). 第二, we predicted the
SPN to scale with how much the task resembled regularity
歧视: regularity = distribution > color = direction/
color > sound/color. 毕竟, in the distribution task,
participants attended to the arrangement of elements,
whereas in the sound/color task they were not even at-
tending exclusively to the visual modality. 第三, we pre-
dicted that the effect of task would be most pronounced
at intermediate levels of PSYMM, 例如 40% 或者 60%.
方法
参加者
有 26 participants in each task and 130 参与-
ipants in total (规律性: age 18–28 years, eight men,
seven left-handed; 颜色: age 18–51 years, four men,
two left-handed; sound/color: age 18–24 years, 三
354
认知神经科学杂志
体积 32, 数字 2
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
哦
H
w
t
n
t
p
哦
:
A
/
d
/
e
d
米
我
F
r
t
哦
p
米
r
C
H
.
p
s
我
我
d
v
我
r
e
e
r
C
t
C
.
米
H
A
我
e
r
d
.
你
C
哦
哦
米
C
n
/
j
A
哦
r
t
C
我
C
n
e
/
–
A
p
r
d
t
我
3
2
C
我
2
e
3
–
5
p
3
d
F
1
9
/
3
3
1
2
3
/
1
2
5
/
3
哦
5
C
3
n
/
_
A
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
哦
乙
C
y
n
G
_
你
A
e
_
s
0
t
1
哦
4
n
8
0
5
8
.
p
S
d
e
F
p
e
乙
米
y
乙
e
G
r
你
2
e
0
s
2
t
3
/
j
/
F
t
.
哦
n
0
5
中号
A
y
2
0
2
1
男人, six left-handed; direction/color: age 18–28 years,
five men, five left-handed; 分配: age 18–37 years,
four men, four left-handed). All had normal or corrected-
to-normal vision. The experiment had local ethics com-
mittee approval and was conducted in accordance with
the Declaration of Helsinki (2008).
the computationally heavy stimulus generation algorithm
on every trial. 相同 600 images were used in regu-
larity and color tasks, 和另一个 600 images were used
in sound/color and direction/color tasks. A third set of
images was required for the distribution task.
Apparatus
EEG was recorded using a BioSemi Active-2 system, 和
64 scalp electrodes arranged according to the interna-
tional 10–20 system. Band-pass filters were set at 0.16
和 100 赫兹. Horizontal EOG and vertical EOG were mon-
itored online for unwanted blinks and eye movements. 在
the regularity and color tasks, stimuli were presented on
A 40 × 30 cm CRT monitor with 60-Hz refresh rate.
Participants were seated 100 cm away from the monitor.
For the other three tasks, our apparatus was upgraded,
and stimuli were presented on a 51 × 29 cm LCD mon-
itor and participants were seated 57 cm away. Stimuli
sizes were consistent in terms of degrees of visual angle.
A chin rest was used for head stabilization in all tasks.
The experiment was programmed in Python using open
source PsychoPy libraries (Peirce, 2007).
Stimuli
Example patterns are shown in Figure 1. 有 160
dots in each pattern. Patterns had four axes of symmetry,
but the number of dots arranged around the axes varied
从 0% (random) 到 100% (perfect symmetry). 这
PSYMM variable increased in 20% increments. 更远
constraints prevented the elements from overlapping or
falling at the center of the pattern. Elements were light
绿色的 (PsychoPy RGB −1, 0.5, −1; 50 Cd/m2) or dark
绿色的 (RGB −1, 0.2, −1; 25 Cd/m2). The background
disk was midgray (RGB 0, 0, 0; 40 Cd/m2) 和其余的
of the screen was black (RGB −1 −1 −1; 0.15 Cd/m2).
Note that PsychoPy RGB coordinates range from −1 to 1.
The gray background disk was 7.7° diameter. The cen-
tral circular area where the elements could land was 7.16°
diameter. Individual dot element diameter was 0.43°, 但
luminance was modulated with a Gaussian mask with a
standard deviation of 1/6, so the visible dot-like element
was approximately 0.21° diameter.
For the distribution task (Figure 1B), the 7.16° diame-
ter zone where the elements fell was implicitly divided
into a central disk and peripheral ring zones of equal
区域. In the uniform condition, there were 80 元素
(50%) in each zone. In the peripheral condition, 那里
是 32 (20%) elements in the central disk zone and
128 (80%) in the outer ring zone.
For all tasks, we generated 600 different stimuli in ad-
vance and saved them as .PNG files. 相同 600 im-
ages were used for all participants within a task, 但在
a different randomized order. This approach allowed
for rapid stimulus presentation without having to engage
程序
All trials began with a fixation baseline (1.5 秒), followed
by pattern presentation (1.5 秒). After these, 参与者
entered their binary judgment. In the regularity task, 他们
classified each image as having “some regularity” or “no
regularity” (see example in Figure 1C). In the color task,
they classified images as “light” or “dark” (referring to the
shade of green). In the sound/color task, participants saw
patterns and simultaneously heard a low (200 赫兹) or high
(800 赫兹) pitched beep. The task was to classify the rela-
tionship between element color and auditory pitch as
either congruent or incongruent (congruent referred to
either low pitched and dark green, or high pitched and
light green, whereas incongruent referred to either low
pitched and light green, or high pitched and dark green).
The direction/color task was cognitively similar to the
sound/color task. Participants classified the relationship
between element color and direction of an up- or down-
ward pointing small central triangle (Figure 5D) 作为骗子-
gruent or incongruent (congruent referred to either a
downward triangle and dark green, or upward triangle
and light green, whereas incongruent referred to either
downward triangle and light green, or upward triangle
and dark green). Participants found the distinction be-
tween congruent and incongruent trials intuitive after
some practice (see Walker, 2012, for discussion of such
sensory correspondences). 最后, the distribution task
required participants to judge whether all dots were
spaced uniformly around the gray disk (uniform) 或dis-
proportionately around the periphery (outside).
The choice of tasks was partly motivated by previous
research and partly by the need for internal coherence
across the study. The color task had been used in previ-
ous work on automatic SPNs with 100% symmetry, and it
allowed us to use same stimuli as the regularity task. 这
congruence tasks were designed to be like the color task,
but further require another judgment about one addi-
tional perceptual dimension, either in the same or differ-
ent sensory modality (例如, sound or triangle direction).
The distribution task was chosen because it focused
attention on an aspect of spatial arrangement, 但不是
on the symmetry itself. We wanted to test whether this
would be sufficient to elicit SPNs with maximum am-
plitude (following similar experiments by Keefe et al.,
2018).
In all five tasks, the judgments were entered after the
stimulus had disappeared. Blue words reminded par-
ticipants of the correct response mapping (Figure 1C).
Participants entered their judgments with the A or L keys
on a computer keyboard. There was no requirement to
Makin et al.
355
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
哦
H
w
t
n
t
p
哦
:
A
/
d
/
e
d
米
我
F
r
t
哦
p
米
r
C
H
.
p
s
我
我
d
v
我
r
e
e
r
C
t
C
.
米
H
A
我
e
r
d
.
你
C
哦
哦
米
C
n
/
j
A
哦
r
t
C
我
C
n
e
/
–
A
p
r
d
t
我
3
2
C
我
2
e
3
–
5
p
3
d
F
1
9
/
3
3
1
2
3
/
1
2
5
/
3
哦
5
C
3
n
/
_
A
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
哦
乙
C
y
n
G
_
你
A
e
_
s
0
t
1
哦
4
n
8
0
5
8
.
p
S
d
e
F
p
e
乙
米
y
乙
e
G
r
你
2
e
0
s
2
t
3
/
j
F
t
.
/
哦
n
0
5
中号
A
y
2
0
2
1
respond quickly. Participants attempted to maintain cen-
tral fixation during baseline and presentation intervals
and blink during the unspeeded response interval be-
tween trials.
In all tasks, there were 600 trials in total. The configu-
ration of the elements in 300 trials was random. 那里
是 60 trials at each of the five levels of PSYMM.
PSYMM was fully crossed with other factors in the design.
All tasks were thus designed so an ideal observer would
enter each binary judgment an equal number of times at
each level of PSYMM.
ERP Analysis
Continuous EEG data were processed offline using
EEGLAB 13.4.4b (Delorme & Makeig, 2004) in MATLAB
2014乙. Preprocessing conventions were chosen for con-
sistency with previous work. Data were first referenced to
the scalp average, low-pass filtered at 25 赫兹, 向下-
sampled to 128 Hz to reduce file size, and then broken
into epochs from −0.5 to +1.5 sec around stimulus on-
放, with a −200 msec prestimulus baseline. Blink and
other large artifacts were removed from the epoched
data with independent components analysis (ICA; 荣格
等人。, 2000). In the regularity task, an average of 9.77
components were removed per participant (min = 4,
max = 17). In the color task, average component re-
moval was 10 (min 3, max 18). In the sound/color task,
它是 9.42 (min 3, max 17); in direction/color task, 它
曾是 10.19 (min 0, max 17); and in the distribution task,
它是 10.27 (min 1, max 18). After ICA, trials were re-
jected if amplitude were more extreme than ±100 μV
at any electrode. Trial exclusion rate was similar across
PSYMM levels, although there was some variation be-
tween tasks (regularity task, 2–3%; 颜色, 4–5%;
sound/color, 4–5%; direction/color, 9–10%; 分配,
6–8%).
Supplementary Material 1 (土井:10.17605/OSF.IO/
WV6XB) examines the consequences of different prepro-
cessing conventions using the regularity task data as an
例子. 除其他事项外, we found that the grand-
average ERPs were very similar when different trial ex-
clusion limits were used and when EEG data were not
cleaned with ICA. We note that ICA cleaning reduced trial
exclusion by approximately 13.4%, without much altering
the shape of the grand-average ERPs.
The SPN at each level of PSYMM was defined as the dif-
ference from random waves at a bilateral posterior elec-
trode cluster [PO7 O1 O2 PO8], 从 300 到 1000 毫秒
poststimulus onset. The left [PO7 O1] and right [O2
PO8] pairs from this cluster were used when comparing
SPN across hemispheres. These electrodes were chosen
a priori.
We then analyzed global field power (GFP) as a func-
tion of PSYMM in each task. GFP was defined as the stan-
dard deviation of amplitudes across all 64 electrodes of
the topographic difference map. If GFP effects parallel
SPN effects, we can be more confident that the latter
were not problematically dependent on our a priori elec-
trode choice.
We conducted two further analyses of the same ERP
data to visualize and confirm the observed effects of
PSYMM and task. 第一的, we used mass univariate analysis
with the hierarchical linear modeling for EEG toolbox in
MATLAB (LIMO; Pernet, Latinus, Nichols, & Rousselet,
2015; Pernet, Chauveau, Gaspar, & Rousselet, 2011).
This applied a pairwise multilevel comparison to all elec-
trodes and time points from −200 to +1000 毫秒. 每个
analysis compared the given PSYMM condition to the
random condition. We used an alpha level of p < .01,
but no spatiotemporal corrections for family-wise error
rate. Second, we used the Randomization Graphical
User Interface toolbox (RAGU; Koenig, Kottlow, Stein,
& Melie-García, 2011) for two purposes. First, we used
RAGU to assess whether topographies differed with
PSYMM. The topographic ANOVA (TANOVA) procedure
in RAGU uses randomization statistics to ascertain
whether observed topographic differences are likely to
occur due to chance. Second, we used RAGU to visualize
the magnitude of differences between topographic maps
on the first two dimensions from PCA.
Null Hypothesis Confirmation
p Values from ANOVA give the probability of obtaining
the observed data given the null hypothesis (pD|H0)
and not the probability of the null hypothesis being true
given the observed data (pH0|D). This becomes a prob-
lem when theoretically interpreting a null result. We
therefore used Bayesian alternatives to null hypothesis
significance testing to supplement theoretically interest-
ing nonsignificant effects (Masson, 2011). This provides
estimates of pH0|D and allowed us to statistically con-
firm null results.
Open Science Policy
All codes for experimental presentation, stimulus, and
EEG and behavioral analysis are freely available on Open
Science Framework (doi:10.17605/OSF.IO/ WV6XB), along
with preprocessed EEG data and Supplementary Anal-
yses 1 and 2. We are happy for other researchers to
reanalyze our data or reuse our experiments or stimuli
for any purpose.
RESULTS
Behavioral Results
Behavioral results from the five tasks are shown in
Figure 2. Participants gave the correct response on nearly
every trial in the color task and in the two congruence
tasks (P correct near 100%). In the distribution task, P
correct was not at ceiling, and there was a tendency to
356
Journal of Cognitive Neuroscience
Volume 32, Number 2
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
2
e
3
-
5
p
3
d
f
1
9
/
3
3
1
2
3
/
1
2
5
/
3
o
5
c
3
n
/
_
a
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
8
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
t
f
.
o
n
0
5
M
a
y
2
0
2
1
was some regularity, P correct increased with PSYMM,
F(1.676, 41.888) = 436.223, p < .001, partial η2 = .946.
There was a significant increase in P correct with every
increment of PSYMM (although performance was already
near ceiling at 80%). At 40% PSYMM, performance was
around chance level (P correct = 55.8%; not significantly
greater than 50%, t(25) = 1.825, p = .08).
Sustained Posterior Negativity
The SPN is best represented as a difference from 0% sym-
metry (random) wave, as in the second row of Figure 3.
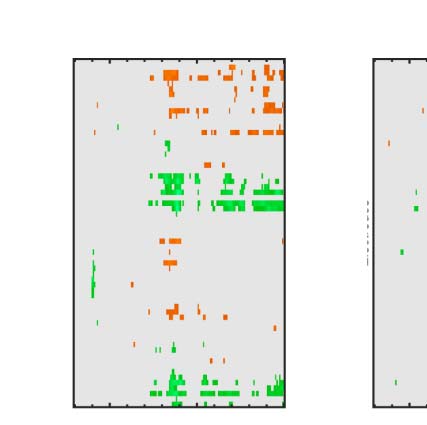
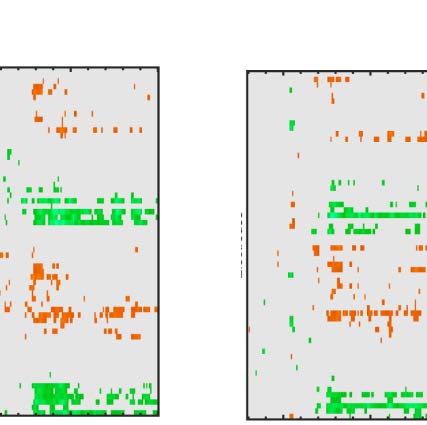
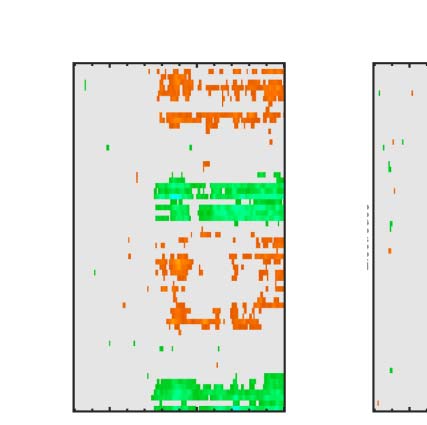
There was a strong effect of PSYMM on SPN amplitude in
all five tasks. However, the SPN was selectively enhanced
in the regularity task (Figure 3, first column). These ef-
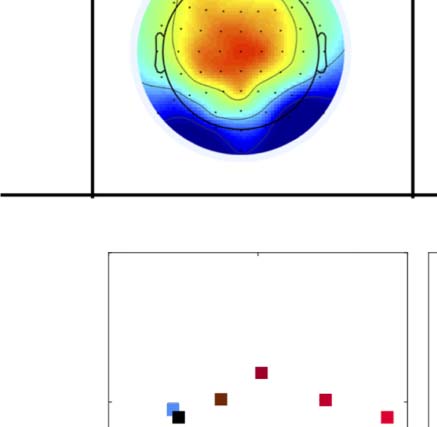
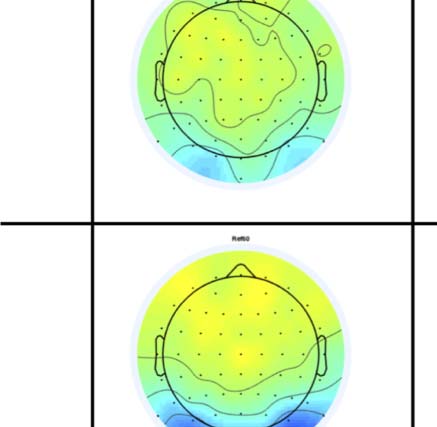
fects can also be seen in the topographic difference maps
in Figure 4, where the SPN appears as blue at posterior
electrodes.
For statistical analysis, SPN amplitude in the 300–
1000 msec interval was examined with mixed ANOVA
(5 PSYMM × 5 Task). Mean SPN amplitudes are shown
in Figure 5C, with 95% confidence intervals. There were
main effects of PSYMM, F(3.174, 396.812) = 123.961, p <
.001, partial η2 = .498, and Task, F(4,125) = 8.851, p <
.001, partial η2 = .221, and a PSYMM × Task inter-
action, F(12.698, 396.812) = 3.219, p < .001, partial
η2 = .093.
Figure 2. Behavioral results. Bars represent mean percent correct
judgments in each task at different levels of PSYMM. Error bars = ±1 SEM.
give the wrong answer more often at higher levels of
PSYMM, F(1.823, 45.573) = 14.847, p < .001, partial
η2 = .373.
In the regularity task, participants judged most random
trials correctly (by reporting “no regularity”). When there
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
2
e
3
-
5
p
3
d
f
1
9
/
3
3
1
2
3
/
1
2
5
/
3
o
5
c
3
n
/
_
a
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
8
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
/
.
f
o
n
0
5
M
a
y
2
0
2
1
Figure 3. ERP results. The five tasks are arranged in columns. The first row shows grand-averaged ERPs from bilateral posterior electrodes
cluster [PO7 O1 O2 PO8]. The second row show the SPN for each level of PSYMM as a difference from the random wave. The third row shows GFP of
the topographic difference maps for each level of PSYMM.
Makin et al.
357
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
2
e
3
-
5
p
3
d
f
1
9
/
3
3
1
2
3
/
1
2
5
/
3
o
5
c
3
n
/
_
a
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
8
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
0
5
M
a
y
2
0
2
1
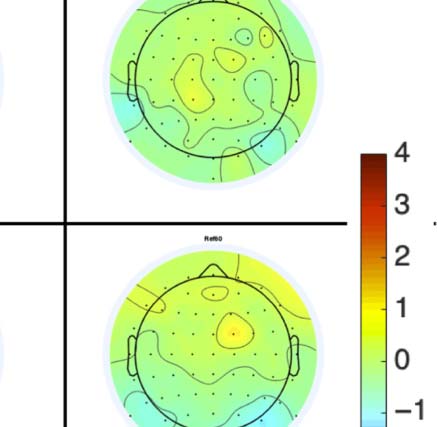
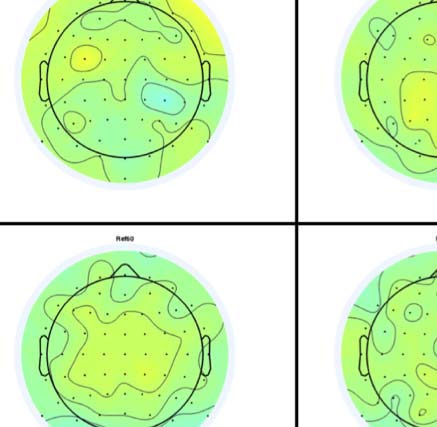
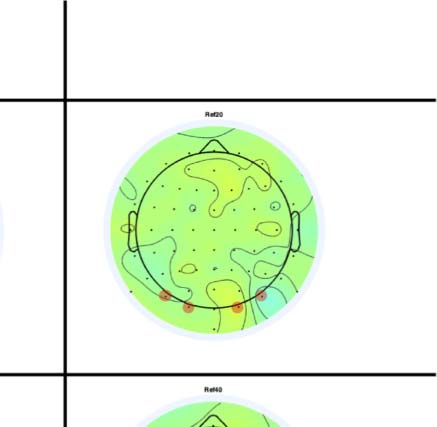
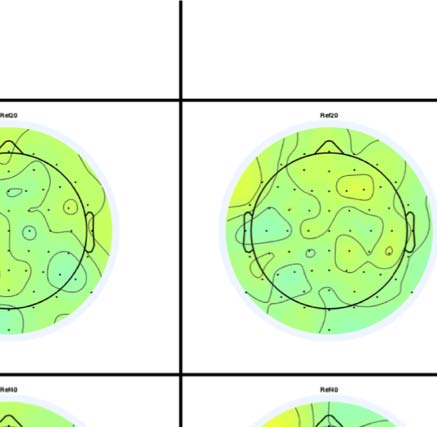
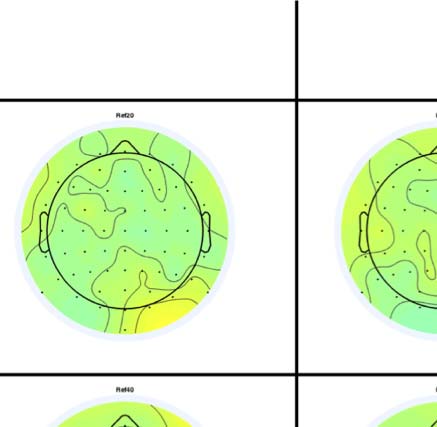
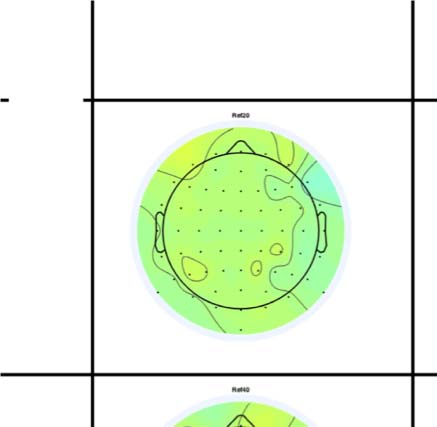
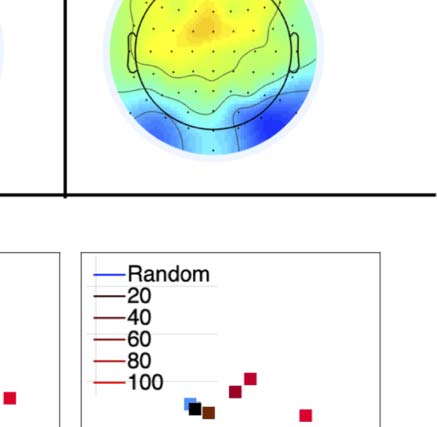
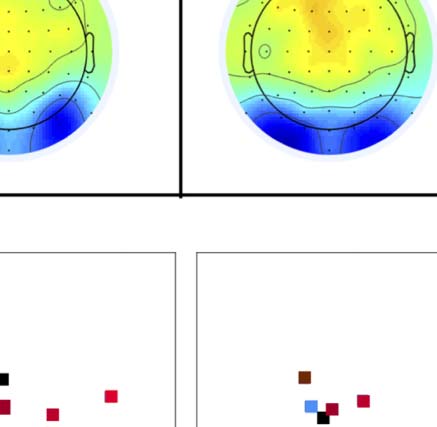
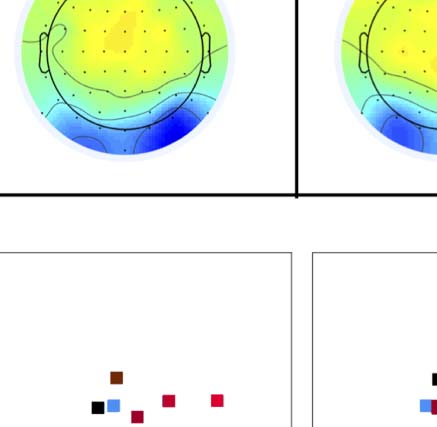
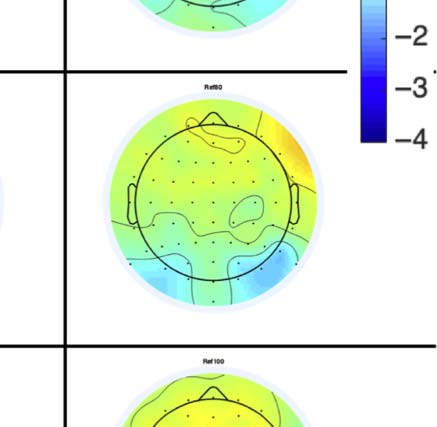
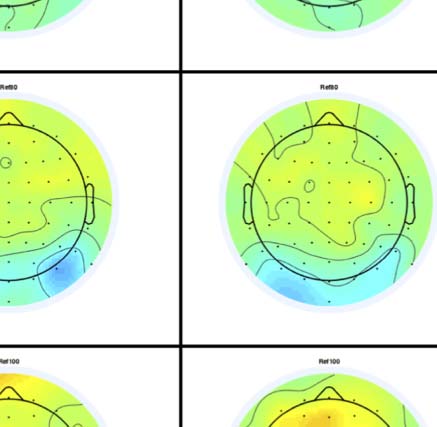
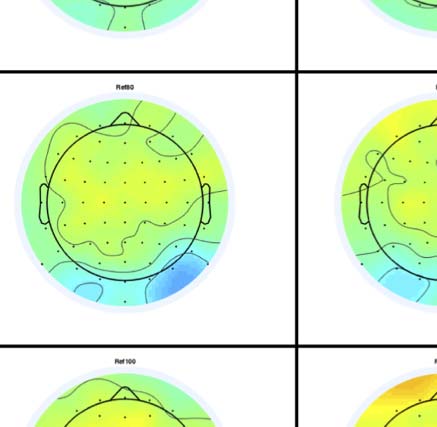
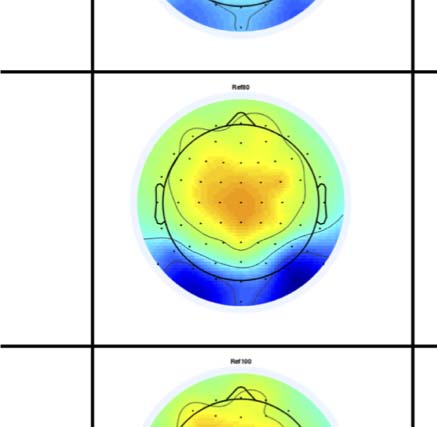
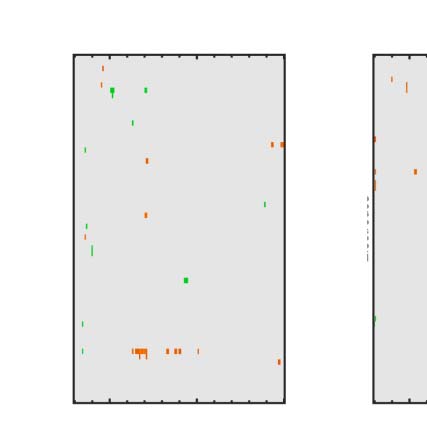
Figure 4. SPN topoplots. (A) Topographic difference maps with increasing PSYMM (rows) in the five tasks (columns). Electrodes used for SPN
analysis are highlighted in red top right. (B) Topographies from each condition are shown as points arranged on MDS plots. These represent the first
two dimensions from PCA. If data points are close together in the square area, then the topographies are similar and vice versa. The five MDS
plots are from the five tasks, corresponding to columns in A. (C) Schematic topographies representing the spatial distribution of the first principal
component. Data points near the left of MDS plots in B are more like the left schematic topography (i.e., more positive at posterior electrodes)
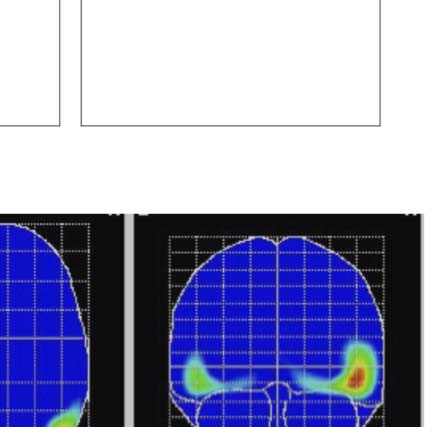
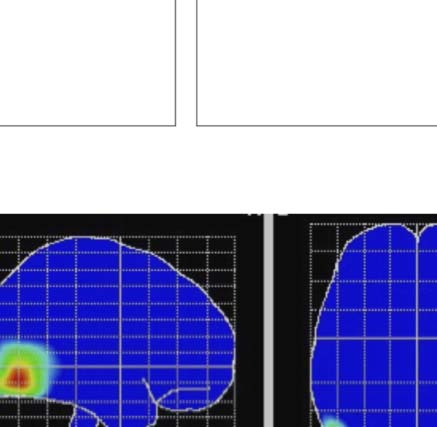
and data points near the right are more like the right schematic topography (i.e., more negative at posterior electrodes). (D) LORETA-estimated
anatomical sources of the SPN, based on grand-averaged 100% difference map, collapsed across tasks.
358
Journal of Cognitive Neuroscience
Volume 32, Number 2
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
2
e
3
-
5
p
3
d
f
1
9
/
3
3
1
2
3
/
1
2
5
/
3
o
5
c
3
n
/
_
a
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
8
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
0
5
M
a
y
2
0
2
1
Figure 5. ERP results. (A–C) P1 peak, N1 peak, and SPN (as difference from random) as a function of PSYMM in each task. In A and B, error
bars = ±1 SEM. In C, error bars = 95% confidence intervals (so there is an SPN when error bars do not cross zero).
The PSYMM × Task interaction was followed up with
sub-ANOVAs. When regularity task was removed from the
analysis, the SPN results were characterized by a sig-
nificant main effect of PSYMM, F(3.098, 309.803) =
78.998, p < .001, partial η2 = .441, but no main effect
of Task, F < 1, ns, pH0|D = 0.999, and no PSYMM ×
Task interaction, F(9.294, 309.803) = 1.316, p = .226,
pH0|D > 0.999. 同时, the regularity task differed
from each of the others when pairs of tasks were ana-
lyzed, both in terms of main effects and interactions:
smallest effect, F(3.037, 151.856) = 4.461, p = .005, par-
tial η2 = .082. The regularity task thus statistically stands
out from the other four.
In all our tasks, PSYMM was crossed with another factor,
such as element color, congruence, or distribution. 这
SPN never interacted with these factors (see Supplemen-
tary Material 2, 土井:10.17605/OSF.IO/ WV6XB). 对于在-
姿态, PSYMM did not interact with Greenshade,
although this generated an independent ERP at posterior
central electrodes. This is consistent with previous work,
where the extrastriate symmetry response was indepen-
dent of low-level luminance and color properties, 只要
as contrast was far above threshold (Martinovic, Jennings,
马金, Bertamini, & Angelescu, 2018).
We predicted that the effect of Task would be larger at
intermediate levels of PSYMM, 例如 40% 或者 60%.
然而, our results were not consistent with this hy-
pothesis (Figure 5C). 反而, the SPN enhancement in
the regularity task was equivalent at all levels of PSYMM
多于 20%. 的确, 当。。。的时候 20% PSYMM condition was
排除在分析之外, there was a main effect of
任务, F(4, 125) = 11.368, partial η2 = .267, 但不是
PSYMM × Task interaction, F(9.907, 309.607) = 1.743,
p = .071, pH0|D > 0.999.
There are several ways to describe the magnitude of
the task effect. Amplitude was around 0.75–1.61 μV larger
in the regularity task than in the average of the other four
(depending on PSYMM; Figure 5C). 在里面 100% PSYMM
状况, this constituted an average of 60% SPN in-
crease when participants attended to regularity. 这
could equally be described as an average of 36% SPN de-
crease when participants attended to something other
than regularity.
最后, we note that, in the regularity task, there was a
significant SPN at 40% PSYMM, t(25) = −4.675, p < .001.
In the color task and distribution task, the first significant
SPNs were at 60% (t(25) = −2.952, p = .007, t(25) =
−3.350, p = .003), whereas in the sound/color task and
direction/color task, the first significant SPNs were at 80%
(t(25) = −4.603, p < .001; t(25) = −4.298, p < .001).
Therefore, attention to regularity made the critical dif-
ference between the presence and absence of an SPN
at 40% PSYMM.
P1 and N1 Peaks
The top row in Figure 3 shows grand-averaged ERP waves
in each task. Among other things, it can be seen that P1
was reduced in the distribution task, whereas the N1 was
enhanced in the regularity task. To explore these effects,
we obtained P1 and N1 peak amplitude during the 130–
180 msec and 180–250 msec windows (Figure 5A and B).1
There was only a borderline effect of PSYMM on P1 peak
amplitude, F(4.339, 542.377) = 2.291, p = .053, partial
η2 = .018, pH0|D > 0.999. 然而, there was a main
effect of Task, F(4, 125) = 3.764, p = .006, partial η2 =
.107 (Figure 5A). 反过来, N1 peak amplitude in-
creased with PSYMM, F(4.194, 524.249) = 17.878, p <
.001, partial η2 = .125, and differed between tasks, F(4,
125) = 5.796, p < .001, partial η2 = .156 (Figure 5B).
Source Localization
We used low-resolution electromagnetic tomography
(LORETA) to estimate the anatomical location of SPN
generators (Pascual-Marqui, Michel, & Lehmann, 1994).
Makin et al.
359
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
2
e
3
-
5
p
3
d
f
1
9
/
3
3
1
2
3
/
1
2
5
/
3
o
5
c
3
n
/
_
a
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
8
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
.
t
o
n
0
5
M
a
y
2
0
2
1
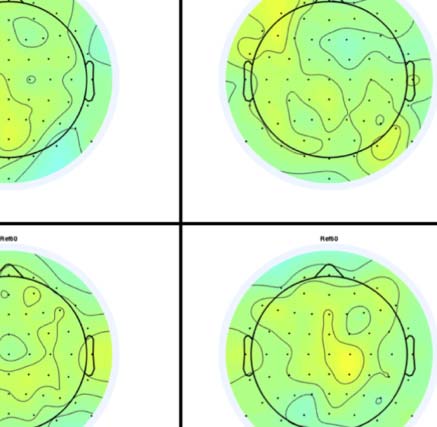
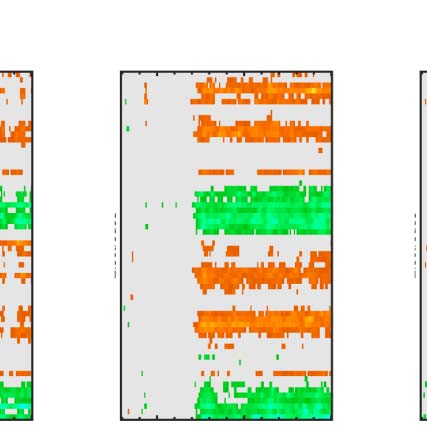
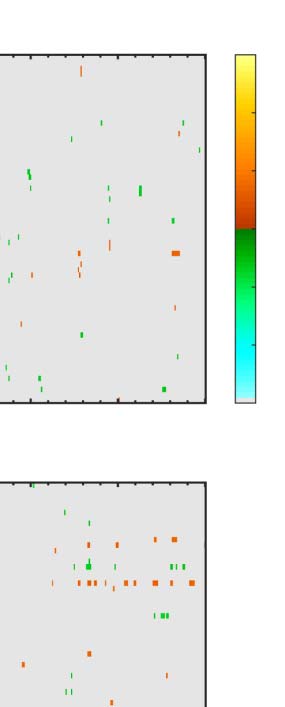
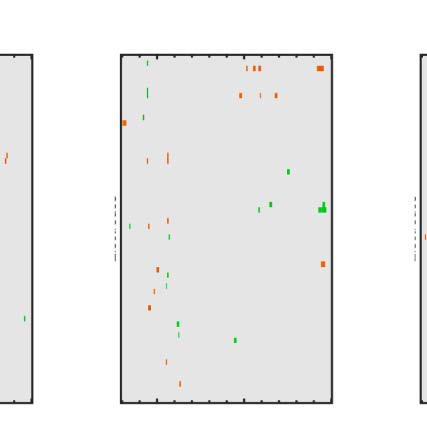
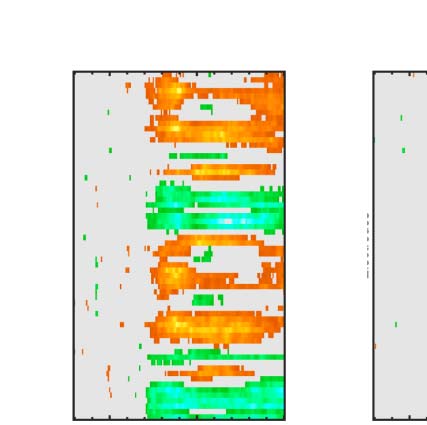
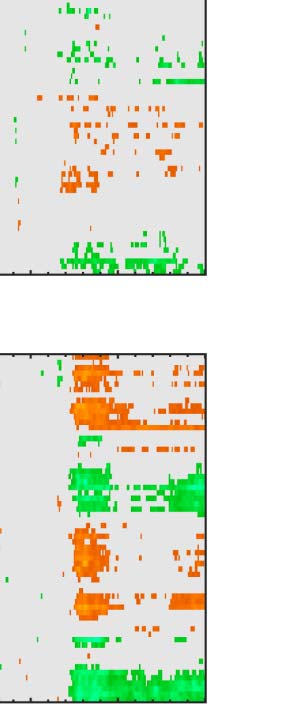
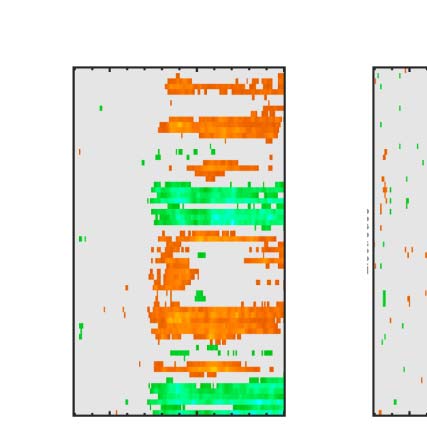
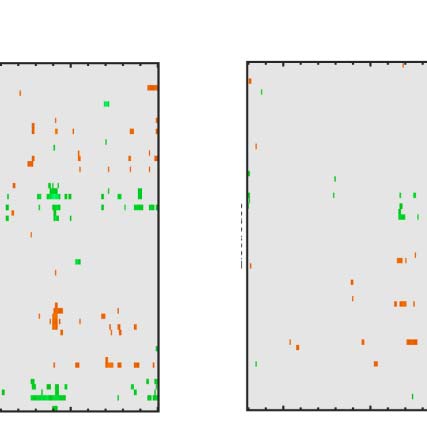
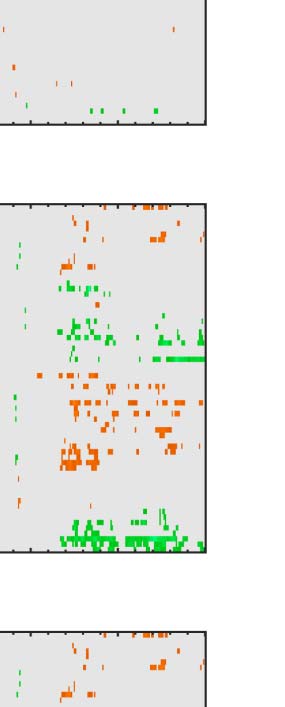
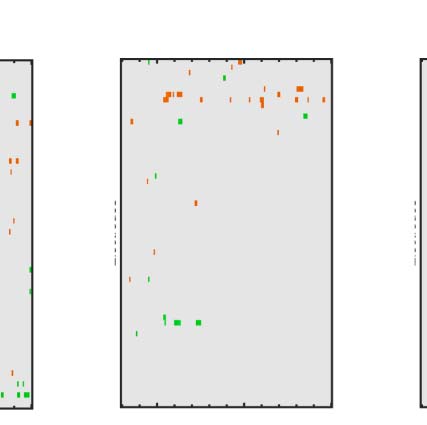
Figure 6. Mass univariate analysis. Each panel shows output from a task (columns) and PSYMM (rows) condition. Within each panel, a row
represents 1 of the 64 electrodes in the BioSemi montage, and the x-axis represents time. The color scale indicates direction and magnitude of
the statistical test, and non-significant effects are masked gray. We used an alpha level of p < .01, the 1% false positive rate resulted in a few isolated
colors throughout. The SPN appears as cold colors at bilateral posterior electrodes (indicated by arrows in lower right).
To obtain the strongest SPN signal, we used the grand-
averaged 100% PSYMM SPN, collapsed across all tasks
(n = 130). As expected, LORETA found sources in the bilat-
eral extrastriate cortex, with some right hemisphere bias
(Figure 4D). This is consistent with fMRI investigations,
which have consistently found symmetry activations in these
brain areas (Van Meel et al., 2019; Sasaki et al., 2005). It is
also consistent with more sophisticated EEG source analysis
reported in an earlier work (Kohler et al., 2016). The right
lateralization of the SPN was also confirmed statistically
(Supplementary Material 2, doi:10.17605/OSF.IO/ WV6XB).
Global Field Power
GFP was defined as standard deviation of 64 amplitudes
across a topographic difference map. We obtained GFP at
360
Journal of Cognitive Neuroscience
Volume 32, Number 2
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
2
e
3
-
5
p
3
d
f
1
9
/
3
3
1
2
3
/
1
2
5
/
3
o
5
c
3
n
/
_
a
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
8
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
.
/
f
o
n
0
5
M
a
y
2
0
2
1
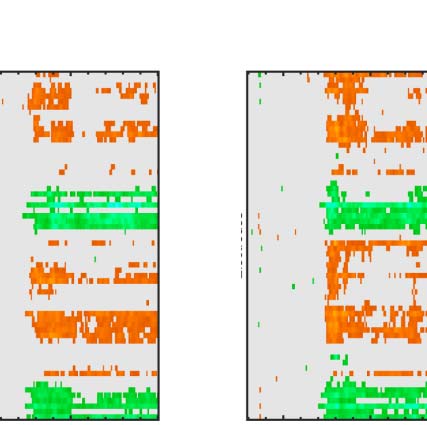
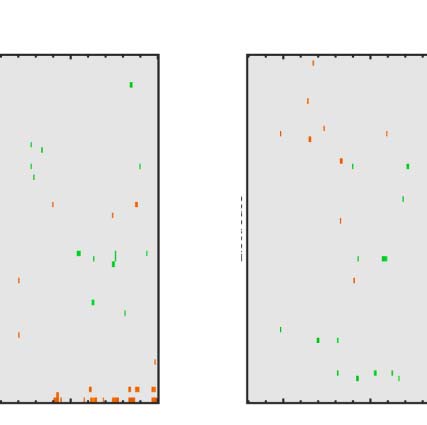
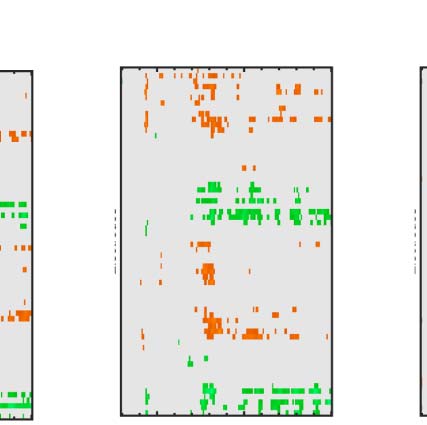
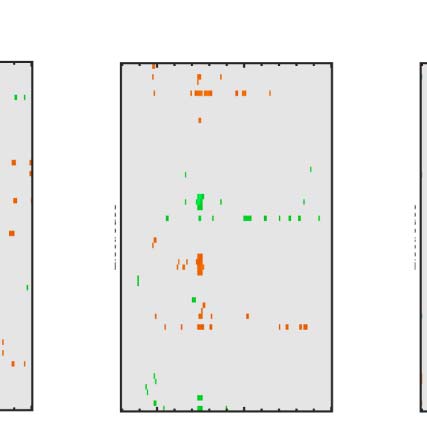
Figure 7. Regularity task ERPs from correct and incorrect trials. Grand-averaged ERPs (left) and SPN difference waves (right). Note the SPN-like wave
on the incorrect random trials (false alarms, top row) and SPN scaling with PSYMM on the correct (hit) trials thereafter (rows 2–4).
each time point. The third row of Figure 3 shows that GFP
effects paralleled the SPN effects. GFP increased dramatically
at around 200 msec and scaled with PSYMM thereafter. Most
importantly, GFP was stronger in the regularity task than the
other four tasks (see Supplementary Material 2, doi:10.17605/
OSF.IO/ WV6XB). The similarity between GFP and SPN
results demonstrates that the SPN results were not prob-
lematically dependent on a priori electrode choice.
Makin et al.
361
Topographic Analysis
Next, we examined SPN topography using TANOVA
(Koenig et al., 2011) and found a strong effect of PSYMM
( p < .001). This reconfirms that the topographies changed
with PSYMM. More interestingly, Figure 4B shows multidi-
mensional scaling (MDS) plots from each task. Here the
grand-averaged topographies are represented as points,
with x- and y-axes representing the first two dimensions
from PCA. If data points are close together in this 2-D
space, then the corresponding topographies are similar. If
data points are far apart, the corresponding topographies
are disparate (see Koenig et al., 2011, for details). The sche-
matic topographies in Figure 4C are not data but represent
the distribution of the first principal component. Data
points near the left are more like the left schematic topog-
raphy (i.e., more positive at posterior electrodes), and data
points near the right are more like the right schematic to-
pography (i.e., more negative at posterior electrodes). As
expected, topographies from each level of PSYMM are
spread along the first dimension (x-axis), with far less vari-
ability along the second dimension ( y-axis). Furthermore,
it can be seen that the topographies are uniquely spread
out across the x-axis in the regularity task.
Mass Univariate Analysis
The SPN results were also examined and visualized with
mass univariate analysis (Pernet et al., 2011, 2015). Multi-
level pairwise comparisons contrasted each PSYMM level
to random at each electrode and time point (Figure 6).
For example, in the lower row, each point represents a
t score from a pairwise comparison between 100% sym-
metry and random at a particular time point and from a
particular electrode. The SPN appears as green/blue at
posterior electrodes (see arrows at the bottom right).
Again, we see the SPN increased with PSYMM (rows)
and was selectively enhanced in the regularity task (left
column).
SPN from Correct and Incorrect Trials of the
Regularity Task
In the regularity task, participants did not always give
the correct answer at intermediate levels of PSYMM
(Figure 2). We assume participants applied an internal
decision threshold to the analogue symmetry signal to
make a binary judgment (no regularity or some regular-
ity). If the analogue signal exceeded the internal thresh-
old, they reported some regularity (Kohler, Cottereau,
& Norcia, 2018; Palumbo et al., 2015).
In the 20% trials, the symmetry signal rarely exceeded
threshold, so these patterns were nearly always judged
incorrectly as having “no regularity” (miss). Conversely,
In the 80% and 100% trials, the symmetry signal nearly
always exceeded threshold, so these patterns were nearly
always judged correctly as having “some regularity” (hit).
Likewise, we can distinguish between random trials
where participants correctly reported “no regularity”
(correct rejection) and random trials where they incor-
rectly reported “some regularity” (false alarm).
First, we consider the distinction between correction
rejection and false alarm trials, where the patterns were
random. In the false alarm trials, there was a low-amplitude
SPN wave (Figure 7 top row). It is possible that images
that generated false alarm SPN had some undesigned
symmetrical-looking features, which might be discernable
through trial-by-trial image analysis. Furthermore, fea-
tures of the trial sequence, such as many random patterns
in a row, could have led to occasional false alarm SPNs.2
Next, we consider the distinction between hit and miss
trials, where PSYMM was > 0 (数字 7, rows 2–4). 那里
was a clear SPN on hit trials but not the miss trials.
有趣的是, the P1 and N1 peaks were very similar on
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
哦
H
w
t
n
t
p
哦
:
A
/
d
/
e
d
米
我
F
r
t
哦
p
米
r
C
H
.
p
s
我
我
d
v
我
r
e
e
r
C
t
C
.
米
H
A
我
e
r
d
.
你
C
哦
哦
米
C
n
/
j
A
哦
r
t
C
我
C
n
e
/
–
A
p
r
d
t
我
3
2
C
我
2
e
3
–
5
p
3
d
F
1
9
/
3
3
1
2
3
/
1
2
5
/
3
哦
5
C
3
n
/
_
A
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
哦
乙
C
y
n
G
_
你
A
e
_
s
0
t
1
哦
4
n
8
0
5
8
.
p
S
d
e
F
p
e
乙
米
y
乙
e
G
r
你
2
e
0
s
2
t
3
/
j
.
/
t
F
哦
n
0
5
中号
A
y
2
0
2
1
数字 8. Regularity task ERPs on correct and incorrect trials. (A) Peak P1 amplitude as a function of PSYMM on correct (绿色的) and incorrect (红色的)
试验. (乙) Peak N1 amplitude as a function of PSYMM on correct and incorrect trials. (C) SPN amplitude as a function of PSYMM on correct and
incorrect trials. In A and B, error bars = ±1 SEM. 在C中, error bars = 95% confidence interval. *Difference between correct and incorrect p < .05.
***Difference between correct and incorrect p < .001.
362
Journal of Cognitive Neuroscience
Volume 32, Number 2
hit and miss trials. This suggests that our participants had
not merely closed their eyes and thus guessed incorrectly
or fixated elsewhere on the screen throughout the trial
on miss trials.
Separate repeated-measures ANOVAs confirmed these im-
pressions (2 judgment [correct, incorrect] × 4 PSYMM [ran-
dom, 20%, 40%, 60%]). For P1 and N1 peaks (Figure 8A and
B) there were no main effects or interactions: largest ef-
fect, F(1.966, 49.158) = 1.086, p = .345, pH0|D = 0.934.
In contrast, for the SPN, there was a Judgment × PSYMM
interaction, F(2.208, 55.200) = 7.878, p = .001, partial
η2 = .240, because there was a main effect of PSYMM on
the correct trials, F(3, 75) = 16.981, p < .001, partial η2 =
.404, but not on the incorrect trials, F(1.965, 49.127) =
1.024, p = .366, pH0|D = 0.936 (Figure 8C). Three condi-
tions produced a significant SPN (Figure 8C). These were
the incorrect (false alarm) random trials, t(25) = −2.453,
p = .021; correct (hit) at 40% trials, t(25) = −4.441, p <
.001; and correct (hit) 60% trials, t(25) = −8.463, p <
.001.3 These conditions also generated significant dif-
ferences between correct and incorrect trials (see stars
on Figure 8C).
DISCUSSION
As expected, we found an SPN response to symmetry,
which increased with PSYMM (Palumbo et al., 2015).
Surprisingly, the SPN was similar in all tasks except
the regularity task, where it was selectively enhanced.
Indeed, at 40% PSYMM, attention to regularity made
the difference between the presence and absence of
an SPN.
It is remarkable that the four non-regularity-related
tasks produced such similar SPN waves. By design, the
tasks were different in several respects. In the distribu-
tion task, participants attended to arrangement of the
elements, so we predicted large SPNs here. At the other
extreme, in the sound/color task, participants partly at-
tended to the auditory modality, so we predicted small
SPNs here. However, the expected task differences were
not found.
The same basic pattern of results was confirmed with
GFP, TANOVA, and mass univariate analysis. We thus con-
clude that there is a default, task-independent parametric
response to symmetry in the extrastriate cortex. How-
ever, this default response can be upregulated when
necessary, for example, when the task is to find sym-
metry in random noise.
The apparent automaticity of the symmetry detection,
at least during nondemanding secondary tasks, is poten-
tially adaptive. Symmetry is a visual cue used to find ob-
jects against a visually cluttered background (Machilsen,
Pauwels, & Wagemans, 2009). Perhaps symmetry detec-
tion mechanisms are nearly always online because it is
nearly always important to parse the visual scene?
Hemispatial neglect caused by right dorsal stream
lesions can impair vertical symmetry discrimination be-
cause patients ignore the left side of the stimulus. How-
ever, symmetry still aids figure-ground segmentation in
these patients (Driver, Baylis, & Rafal, 1992). This again
suggests symmetry can be processed automatically (and
unconsciously) in the ventral stream. This is consistent
with our results, where symmetry was processed auto-
matically in all tasks.
Generalizability of Conclusions
Of course, we only explored four nonregularity tasks
here. We cannot assume the same results would be ob-
tained across all conceivable secondary tasks. For in-
stance, SPN might be greatly reduced if the secondary
task were very demanding and used more attentional re-
sources, as in the experiments by Mack and Rock (1998).
However, we note that SPN was nearly identical in our
easy color task, in the more difficult distribution task
(where performance was not a ceiling), and in our con-
gruence tasks where information had to be integrated
across two perceptual dimensions. This suggests that
the SPN is robust to a wide range of task manipulations,
although the boundaries of task independence remain to
be established.
Likewise, our experiments only tested automaticity of
symmetry perception, not the automaticity of other kinds
of perceptual organization. The SPN can also be gener-
ated by other gestalts such as glass patterns (Makin
et al., 2016) and perhaps by recognizable real objects
(Martinovic, Mordal, & Wuerger, 2011). This indirectly
suggests that our results may generalize beyond sym-
metry. However, automaticity is unlikely to be a uni-
versal law of perceptual organization, which involves a
heterogenous set of more or less automatic subprocesses
(Barbot, Liu, Kimchi, & Carrasco, 2018).
There is probably an important distinction between
the less and more computationally demanding types of
perceptual organization. Perhaps only the less demand-
ing types happen automatically across secondary tasks.
Extraction of symmetry from the image is an example
of this. In contrast, more demanding types of perceptual
organization are probably not automatic and often fail
during secondary tasks. Examples include the formation
of allocentric, view invariant object representations
(Makin et al., 2015), and formation of an object represen-
tations from temporally separated images (Rampone,
Makin, Tatlidil, & Bertamini, 2019).
SPN and Binary Judgments
There was a systematic relationship between recorded
brain responses and behavior in our regularity task.
When the pattern was random, participants usually gave
the correct answer and reported “no regularity” (a cor-
rect rejection). However, they occasionally gave an in-
correct answer and reported “some regularity” (a false
alarm). There was a weak SPN on these false alarm trials.
Makin et al.
363
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
2
e
3
-
5
p
3
d
f
1
9
/
3
3
1
2
3
/
1
2
5
/
3
o
5
c
3
n
/
_
a
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
8
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
f
t
o
n
0
5
M
a
y
2
0
2
1
When the pattern was symmetrical, participants sometimes
correctly reported “some regularity” (a hit) or incorrectly re-
ported “no regularity” (a miss). There was a parametric SPN
on the hit trials, but no SPN on miss trials. On miss trials, it
seems that perceptual organization failed, and no symmetry
representation was generated in the extrastriate cortex,
despite availability the symmetry in the distal stimulus.
This result is interesting because P1 and N1 components
of the visual evoked potential (VEP) were present on miss
trials, so participants were likely processing the stimulus
during the early part of the trial (and not, for example,
closing their eyes or fixating elsewhere throughout).
Although the extrastriate symmetry response some-
times failed, we do not think this means it should be con-
sidered as nonautomatic. After all, even an automatic
perceptual response will not be elicited by the stimulus
with 100% reliability.
An ERP that is present on both hit and false alarm trials,
but absent on both miss and correct rejection trials,
would be typically associated with binary decision-
making or behavioral responses rather than perception.
However, we argue that the SPN is a measure of percep-
tual organization, not decision-making or behavior. First,
SPN amplitude increased with PSYMM on hit trials, al-
though the behavioral response was identical. Second,
the false alarm SPN was much weaker than the 60% hit
SPN, even though the response was again identical.
Third, SPN amplitude was related to PSYMM in a linear
fashion, even though the frequency of “some regularity”
responses nearly plateaued at 80%. All these observations
are explicable if we assume that participants applied an
internal threshold to the analogue symmetry signal and
reported “some regularity” if the symmetry signal ex-
ceeded threshold. The SPN is generated by the analogue
symmetry signal, not the decision-making processes or
behavior (see Kohler et al., 2018, for further analysis of
decision-making in symmetry perception).
Why did we observe a false alarm SPN on some ran-
dom trials in the regularity task? There are two obvious
explanations. First, our stimulus construction algorithm
may have occasionally introduced some regularity into
the random patterns by chance. In this case, the term
“false alarm” is a misnomer: The visual system detected
slight regularity present in the stimulus. Second, internal
neural fluctuations may have led to super-threshold sym-
metry activations on some occasions. These are genuine
false alarms. The relative contribution of each effect to
the observed “false alarm” SPN is uncertain. However,
neither phenomena fundamentally challenges our claim
that the SPN is a measure of automatic perceptual
organization.
Finally, we note that the participants in our regularity
task were searching for intermediate symmetry in noisy
displays, and the weak symmetry signal was often near
the threshold of discriminability. These task parameters
are likely to maximize responsiveness to any residual
regularity in randomly generated images. This could be
tested in future work: The false alarm SPN on random tri-
als might disappear if the task required mere discrimina-
tion of random from 100% symmetry, with no ambiguous
intermediate levels of PSYMM.
VEP and Task
Analysis of P1 and N1 peaks was also interesting in other
ways. N1 was much larger in the regularity task than the
others. Previous work has found that visual processes at
the N1 latency mediate spatial integration (Shpaner,
Molholm, Forde, & Foxe, 2013). Maybe spatial integration
was more pronounced in the regularity task, where it
was required? Furthermore, N1 increased with PSYMM.
This replicates previous research where later SPN effects
are already weakly present at N1 (Makin, Rampone,
Pecchinenda, & Bertamini, 2013). Further work is needed
to clarify early visual processing stages in gestalt for-
mation, which happen at the N1 latency. Conversely, P1
was not modulated by PSYMM, presumably because this
peak indexes early visual onset detection, before the
extrastriate symmetry response begins.
Right Lateralization of the SPN
The SPN was slightly stronger over the right hemisphere
than the left, particularly in the regularity and color tasks,
and source localization also found a predominantly right
hemisphere symmetry activation. Although there is no
clear explanation for why this right lateralization should
be stronger in some tasks than others, it has been ob-
served previously (Bertamini & Makin, 2014). Moreover,
converging evidence comes from three other methodol-
ogies. First, TMS disruption of the right lateral occipital
complex selectively impairs regularity discrimination
(Bona, Herbert, Toneatto, Silvanto, & Cattaneo, 2014).
Second, regularity presented to the right hemisphere is
discriminated more quickly (Verma, Van der Haegen, &
Brysbaert, 2013). Third, regularity can be more reliably
decoded from right lateral occipital complex multivoxel
patterns ( Van Meel et al., 2019). We conclude that the
extrastriate symmetry response is weakly right-lateralized,
although there is certainly a symmetry response in the left
hemisphere too (Wright, Makin, & Bertamini, 2017).
Conclusions
When does the visual brain expend computational re-
sources to compare correspondence across locations
and extract symmetry from the image? We found a de-
fault parametric response to symmetry in the extrastriate
cortex, which was similar across five very different tasks.
This suggests the brain always engages some perceptual
organization to detect symmetry. However, this default
symmetry response was selectively enhanced when regu-
larity was task relevant. We conclude that symmetrical
gestalt formation is (1) largely task independent, (2) can
364
Journal of Cognitive Neuroscience
Volume 32, Number 2
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
2
e
3
-
5
p
3
d
f
1
9
/
3
3
1
2
3
/
1
2
5
/
3
o
5
c
3
n
/
_
a
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
8
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
t
f
o
n
0
5
M
a
y
2
0
2
1
be enhanced by attention to symmetry, but (3) fails on a
minority of trials, so there is no extrastriate activation and
symmetry is missed.
Acknowledgments
This project was part funded by an Economic and Social Research
Council grant award to Alexis D. J. Makin (ES/S014691/1). We
would like to thank project students Zaynah Ahmed, Luke
Bennett, Eleanor Dickens, Alice Newton-Fenner, and Kasia Gmaj
for helping with data collection.
Reprint request should be sent to Alexis D. J. Makin, Depart-
ment of Psychological Sciences, Eleanor Rathbone Building,
University of Liverpool, Liverpool, L69 7ZA, United Kingdom,
or via e-mail: alexis.makin@liverpool.ac.uk.
Notes
1.
It is unclear why the VEP was delayed by approximately 33 msec
in these experiments. This was at least partly due to timing dif-
ferences between older and newer apparatus. However, we are
not theoretically interpreting P1 or N1 latency, just amplitude.
2. After the regularity task, our participants filled in the Oxford–
Liverpool Inventory of Feelings and Experiences (O-LIFE) ques-
tionnaire, which measures psychotic symptoms in the normal
population (Mason & Claridge, 2006). There was no hint of a cor-
relation between the amplitude of false alarm SPN and scores of
the unusual experiences subscale, which measures hallucination
proneness (r = .04). Future researchers interested in clinically
relevant individual differences in vision should be aware of this
null result.
3. This analysis should be treated with some caution, because
the number of trials at 20% correct and 60% incorrect con-
ditions was low (average 12 and 14). This is not considered
enough to generate reliable ERP waves (see noise in the pre-
stimulus baseline). However, this did not obscure systematic
effects, and the P1 and N1 were similar.
REFERENCES
Barbot, A., Liu, S., Kimchi, R., & Carrasco, M. (2018). Attention
enhances apparent perceptual organization. Psychonomic
Bulletin & Review, 25, 1824–1832.
Barlow, H. B., & Reeves, B. C. (1979). Versatility and absolute
efficiency of detecting mirror symmetry in random dot
displays. Vision Research, 19, 783–793.
Bertamini, M., & Makin, A. D. J. (2014). Brain activity in
response to visual symmetry. Symmetry, 6, 975–996.
Bertamini, M., Silvanto, J., Norcia, A. M., Makin, A. D. J., &
Wagemans, J. (2018). The neural basis of visual symmetry
and its role in mid- and high-level visual processing. Annals
of the New York Academy of Sciences, 1426, 111–126.
Bona, S., Herbert, A., Toneatto, C., Silvanto, J., & Cattaneo, Z.
(2014). The causal role of the lateral occipital complex in
visual mirror symmetry detection and grouping: An fMRI-
guided TMS study. Cortex, 51, 46–55.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21.
Höfel, L., & Jacobsen, T. (2007). Electrophysiological indices
of processing aesthetics: Spontaneous or intentional
processes? International Journal of Psychophysiology, 65,
20–31.
Hoffman, D. (1998). Visual intelligence: How we create what
we see. New York: W.W. Norton & Company.
Jacobsen, T., & Höfel, L. (2003). Descriptive and evaluative
judgment processes: Behavioral and electrophysiological
indices of processing symmetry and aesthetics. Cognitive,
Affective & Behavioral Neuroscience, 3, 289–299.
Jung, T. P., Makeig, S., Humphries, C., Lee, T. W., McKeown, M. J.,
Iragui, V., et al. (2000). Removing electroencephalographic
artifacts by blind source separation. Psychophysiology, 37,
163–178.
Keefe, B. D., Gouws, A. D., Sheldon, A. A., Vernon, R. J. W.,
Lawrence, S. J. D., McKeefry, D. J., et al. (2018). Emergence
of symmetry selectivity in the visual areas of the human
brain: fMRI responses to symmetry presented in both
frontoparallel and slanted planes. Human Brain Mapping,
39, 3813–3826.
Koenig, T., Kottlow, M., Stein, M., & Melie-García, L. (2011).
RAGU: A free tool for the analysis of EEG and MEG event-
related scalp field data using global randomization statistics.
Computational Intelligence and Neuroscience, 2011,
938925.
Kohler, P. J., Clarke, A., Yakovleva, A., Liu, Y., & Norcia, A. M.
(2016). Representation of maximally regular textures in
human visual cortex. Journal of Neuroscience, 36, 714–729.
Kohler, P. J., Cottereau, B. R., & Norcia, A. M. (2018). Dynamics
of perceptual decisions about symmetry in visual cortex.
Neuroimage, 167, 316–330.
Lamme, V. A., & Roelfsema, P. R. (2000). The distinct modes of
vision offered by feedforward and recurrent processing.
Trends in Neurosciences, 23, 571–579.
Machilsen, B., Pauwels, M., & Wagemans, J. (2009). The role of
vertical mirror symmetry in visual shape detection. Journal of
Vision, 9, 1–11.
Mack, A., & Rock, I. (1998). Inattentional blindness.
Cambridge, MA: MIT Press.
Makin, A. D., Rampone, G., & Bertamini, M. (2015). Conditions
for view invariance in the neural response to symmetry.
Psychophysiology, 52, 532–543.
Makin, A. D., Rampone, G., Pecchinenda, A., & Bertamini, M.
(2013). Electrophysiological responses to visuospatial
regularity. Psychophysiology, 50, 1045–1055.
Makin, A. D., Wilton, M. M., Pecchinenda, A., & Bertamini, M.
(2012). Symmetry perception and affective responses: A
combined EEG/EMG study. Neuropsychologia, 50, 3250–3261.
Makin, A. D., Wright, D., Rampone, G., Palumbo, L., Guest, M.,
Sheehan, R., et al. (2016). An electrophysiological index of
perceptual goodness. Cerebral Cortex, 26, 4416–4434.
Martinovic, J., Jennings, B. J., Makin, A. D. J., Bertamini, M., &
Angelescu, I. (2018). Symmetry perception for patterns
defined by color and luminance. Journal of Vision, 18, 4.
Martinovic, J., Mordal, J., & Wuerger, S. M. (2011). Event-related
potentials reveal an early advantage for luminance contours
in the processing of objects. Journal of Vision, 11. https://
doi.org/110.1167/11.7.1.
Mason, O., & Claridge, G. (2006). The Oxford-liverpool
inventory of feelings and experiences (O-LIFE): Further
description and extended norms. Schizophrenia Research,
82, 203–211.
Driver, J., Baylis, G. C., & Rafal, R. D. (1992). Preserved figure-
Masson, M. E. (2011). A tutorial on a practical Bayesian
ground segregation and symmetry perception in visual
neglect. Nature, 360, 73–75.
Gheorghiu, E., Kingdom, F. A. A., Remkes, A., Li, H.-C. O., &
Rainville, S. (2016). The role of color and attention-to-color in
mirror-symmetry perception. Scientific Reports, 6, 29287.
alternative to null-hypothesis significance testing. Behavior
Research Methods, 43, 679–690.
Palumbo, L., Bertamini, M., & Makin, A. (2015). Scaling of the
extrastriate neural response to symmetry. Vision Research,
117, 1–8.
Makin et al.
365
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
2
e
3
-
5
p
3
d
f
1
9
/
3
3
1
2
3
/
1
2
5
/
3
o
5
c
3
n
/
_
a
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
8
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
f
/
t
o
n
0
5
M
a
y
2
0
2
1
Pascual-Marqui, R. D., Michel, C. M., & Lehmann, D. (1994).
Low resolution electromagnetic tomography: A new method
for localizing electrical activity in the brain. International
Journal of Psychophysiology, 18, 49–65.
Peirce, J. W. (2007). PsychoPy–Psychophysics software in
Python. Journal of Neuroscience Methods, 162, 8–13.
Pernet, C. R., Chauveau, N., Gaspar, C., & Rousselet, G. A.
(2011). LIMO EEG: A toolbox for hierarchical linear modeling
of electroencephalographic data. Computational
Intelligence and Neuroscience, 2011, 831409.
Pernet, C. R., Latinus, M., Nichols, T. E., & Rousselet, G. A. (2015).
Cluster-based computational methods for mass univariate
analyses of event-related brain potentials/fields: A simulation
study. Journal of Neuroscience Methods, 250, 85–93.
Rampone, G., Makin, A. D. J., & Bertamini, M. (2014).
Electrophysiological analysis of the affective congruence
between pattern regularity and word valence.
Neuropsychologia, 58, 107–117.
Rampone, G., Makin, A. D. J., Tatlidil, S., & Bertamini, M.
(2019). Representation of symmetry in the extrastriate visual
cortex from temporal integration of parts: An EEG/ERP
study. Neuroimage, 193, 214–230.
Sasaki, Y., Vanduffel, W., Knutsen, T., Tyler, C. W., & Tootell, R.
(2005). Symmetry activates extrastriate visual cortex in
human and nonhuman primates. Proceedings of the
National Academy of Sciences, U.S.A., 102, 3159–3163.
Shpaner, M., Molholm, S., Forde, E., & Foxe, J. J. (2013).
Disambiguating the roles of area V1 and the lateral occipital
complex (LOC) in contour integration. Neuroimage, 69,
146–156.
Treisman, A. M., & Gelade, G. (1980). A feature-integration
theory of attention. Cognitive Psychology, 12, 97–136.
Tyler, C. W., Baseler, H. A., Kontsevich, L. L., Likova, L. T., Wade,
A. R., & Wandell, B. A. (2005). Predominantly extra-
retinotopic cortical response to pattern symmetry.
Neuroimage, 24, 306–314.
van der Helm, P. A. (2010). Weber-Fechner behavior in
symmetry perception? Attention Perception & Psychophysics,
72, 1854–1864.
Van Meel, C., Baeck, A., Gillebert, C. R., Wagemans, J., &
Op de Beeck, H. P. (2019). The representation of symmetry
in multi-voxel response patterns and functional connectivity
throughout the ventral visual stream. Neuroimage, 191,
216–224.
Verma, A., Van der Haegen, L., & Brysbaert, M. (2013).
Symmetry detection in typically and atypically speech
lateralized individuals: A visual half-field study.
Neuropsychologia, 51, 2611–2619.
Wagemans, J., Elder, J. H., Kubovy, M., Palmer, S. E.,
Peterson, M. A., Singh, M., et al. (2012). A century of
Gestalt psychology in visual perception: I. Perceptual
grouping and figure–ground organization. Psychological
Bulletin, 138, 1172–1217.
Walker, P. (2012). Cross-sensory correspondences and cross
talk between dimensions of connotative meaning: Visual
angularity is hard, high-pitched, and bright. Attention
Perception & Psychophysics, 74, 1792–1809.
Wright, D., Makin, A. D. J., & Bertamini, M. (2017).
Electrophysiological responses to symmetry presented in the
left or in the right visual hemifield. Cortex, 86, 93–108.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
n
t
p
o
:
a
/
d
/
e
d
m
i
f
r
t
o
p
m
r
c
h
.
p
s
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
2
c
l
2
e
3
-
5
p
3
d
f
1
9
/
3
3
1
2
3
/
1
2
5
/
3
o
5
c
3
n
/
_
a
1
_
8
0
6
1
1
4
2
8
2
5
7
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
4
n
8
0
5
8
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
t
f
o
n
0
5
M
a
y
2
0
2
1
366
Journal of Cognitive Neuroscience
Volume 32, Number 2