Temporal Organization of Episodic and Experience-near
Semantic Autobiographical Memories: Neural Correlates
and Context-dependent Connectivity
Alice Teghil1,2
, Alessia Bonavita1,2, Federica Procida1,
Federico Giove2,3, and Maddalena Boccia1,2
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
抽象的
■ Autobiographical memory includes a representation of per-
sonal life events with a unique spatiotemporal context (episodic
autobiographical memory) and factual self-knowledge (个人的
语义学). Whereas “experience-far” personal semantics have
undergone complete abstraction, “experience-near” personal
semantics are still linked to a spatiotemporal context. The repre-
sentation of one’s own past involves an autobiographical knowl-
edge base, in the form of a personal timeline, along which
autobiographical information is temporally organized into differ-
ent lifetime periods. Commonalities and differences between
brain networks supporting this temporal organization for auto-
biographical information with different contextual specificity,
然而, have not been investigated to date. 这里, we used
task-based fMRI to assess neural substrates of temporal ordering
along the personal timeline for real autobiographical episodic
and experience-near personal semantic memories. Within a
distributed network, the left calcarine cortex was more strongly
activated for episodic autobiographical memory than personal
语义学, whereas the left ventromedial pFC and right posterior
cingulate cortex (PCC), angular gyrus (AG), and anterior middle
temporal gyrus (aMTG) showed stronger activation for personal
semantics than episodic autobiographical memory. 发现
were confirmed by analyses in independently derived ROIs.
Generalized psychophysiological interaction analyses between
the same regions showed that, during personal semantics com-
pared with episodic autobiographical memory, memory category
modulated activity in the left PCC and right PCC, AG, and aMTG.
Findings provide insights on how personal events and facts are
represented in the timescale of years, suggesting that the tempo-
ral organization of autobiographical memory exploits properties
of situation models developed within posteromedial, lateral pari-
etal, and medial prefrontal regions. ■
介绍
Autobiographical memory, including memory for personal
life events and factual knowledge about oneself (Palombo,
Sheldon, & 莱文, 2018; Renoult, 戴维森, Palombo,
Moscovitch, & 莱文, 2012), plays a key role in the
construction and maintenance of a stable and coherent
representation of the self across time (Prebble, Addis, &
Tippett, 2013; Wilson & Ross, 2003). In line with the sem-
inal distinction proposed by Tulving (1972), an episodic
and a semantic component can be identified within the
autobiographical memory domain (Renoult et al., 2016;
克莱因 & Gangi, 2010; 莱文, Svoboda, 干草, Winocur, &
Moscovitch, 2002). Episodic autobiographical memory
(EAM) involves a representation of specific events from
one’s own past, which occurred at a particular time and
place and are thus linked to a unique spatiotemporal con-
文本. Semantic components of autobiographical memory
(often referred to as “personal semantics” [聚苯乙烯]) involve
1Sapienza University of Rome, 2IRCCS Fondazione Santa Lucia,
罗马, 意大利, 3Museo Storico della Fisica e Centro Studi e
Ricerche Enrico Fermi, 罗马, 意大利
factual knowledge related to the self; this kind of memory
includes autobiographical facts (names of relatives and
朋友们, information such as the address of one’s first
房子), self-knowledge (knowledge of personality traits,
角色, 例如. “I am a researcher” or “I am shy”, and personal
信仰), and repeated events (Grilli & Verfaellie, 2014;
Renoult et al., 2012). The most influential cognitive
models on the organization of autobiographical memory
propose that the representation of the personal past
involves an autobiographical knowledge base, corre-
sponding to an organizing representation of the content
and structure of one’s own life (D’Argembeau, 2020;
康威 & Pleydell-Pearce, 2000). The autobiographical
knowledge base thus provides a sort of personal timeline,
along which autobiographical information is temporally
organized in spatial terms (D’Argembeau, 2020; Arzy,
Adi-Japha, & Blanke, 2009; Arzy, Molnar-Szakacs, &
Blanke, 2008). The autobiographical knowledge base
allows the representation of personal past information at
different levels of specificity, from lifetime periods to gen-
eral events to specific events, thus supporting the repre-
sentation of both episodic autobiographical and personal
© 2022 麻省理工学院. Published under a
Creative Commons Attribution 4.0 国际的 (抄送 4.0) 执照.
认知神经科学杂志 34:12, PP. 2256–2274
https://doi.org/10.1162/jocn_a_01906
semantic information (D’Argembeau, 2020; 康威 &
Pleydell-Pearce, 2000).
Previous research has shown that different types of PS
may be conceived as more episodic or semantic in nature,
depending on the degree to which they are linked to spe-
cific episodes (Renoult et al., 2012; Cabeza & St Jacques,
2007). Pieces of information such as “I go to my brother’s
for dinner every Thursday” involve conceptual knowledge
about the self that is derived from repeated individual epi-
sodes; 反过来, pieces of PS knowledge such as “I have
a brother” are more abstract in nature, as they are not
related to specific time or place information (Sheldon,
Peters, & Renoult, 2020). Specific categories of PS, 这样的
as autobiographical facts, are thus considered particularly
“experience-near” because they are more strongly linked
to a spatiotemporal context compared with general
semantic knowledge that is devoid of contextual features
(Grilli & Verfaellie, 2014, 2016). Supporting this distinc-
的, lesions to medial temporal regions impair the
retrieval of not only specific personal episodes but also
experience-near autobiographical facts (Grilli & Verfaellie,
2014).
Neuroimaging studies have provided evidence that
autobiographical memory relies overall on a broad neural
网络, involving the posterior cingulate cortex (PCC),
medial and lateral temporal regions, the posterior–inferior
parietal cortex, and the anterior cingulate/ventromedial
pFC (vmPFC; Teghil, Bonavita, Guariglia, & Boccia, 2021;
Svoboda, McKinnon, & 莱文, 2006). Brain networks spe-
cifically supporting the temporal organization of autobio-
graphical memories along the personal timeline, 然而,
have not been systematically investigated.
St Jacques, 鲁宾, LaBar, and Cabeza (2008) 报道
stronger activation of the left dorsolateral pFC, 正确的
parahippocampal gyrus, the posterior midline cortex,
and the cuneus when participants discriminated the order
of real-life events close in time; temporal order discrimina-
tion of events more distant in time, 反而, activated the
right dorsolateral pFC and the fusiform gyrus (St Jacques
等人。, 2008). 还, when participants were asked to decide
which of two personal life events came before the other, 或者
which of two hypothetic future life events would happen
before the other, activation was found in the bilateral
intraparietal sulcus, dorsolateral pFC, anterior insula, ACC,
and precuneus (pCu) and visual cortex (D’Argembeau,
Jeunehomme, Majerus, Bastin, & Salmon, 2015). 两个都
studies investigated temporal order processing of auto-
information in the range of 1 week
biographical
(D’Argembeau et al., 2015) or a single day (St Jacques
等人。, 2008) and thus do not allow to draw inference
on whether the same brain networks may also support
the organization of autobiographical knowledge in differ-
ent lifetime periods along one’s own personal timeline.
尤其, along a somewhat different line of research, A
few studies assessed brain correlates of individuals’ ability
to “project” themselves at different time points (现在,
8 years in the past, 或者 8 years in the future) along a
spatialized representation of their life (a “mental time-
line”) and to judge whether past- or future-related stimuli
occurred/should occur before or after the imagined self-
location in time (Arzy, Collette, Ionta, Fornari, & Blanke,
2009; Arzy et al., 2008). Stimuli presented in these studies
were either pictures of the participant’s face and of a
famous person’s face modified to demonstrate different
年龄 (Arzy, Collette, 等人。, 2009), or labels referring to
common personal life events (例如, “first child”) or nonper-
sonal events (例如, “hurricane Katrina”; Arzy, Collette, 等人。,
2009; Arzy et al., 2008).
全面的, which brain networks support the ordinal orga-
nization of autobiographical information along different
lifetime periods on one’s own personal timeline is still
未知. 而且, previous studies assessed mental
travel across common personal life events, nonpersonal
事件, or famous people (Arzy, Collette, 等人。, 2009; Arzy
等人。, 2008) or asked participants to discriminate the order
of personal episodes (D’Argembeau et al., 2015; St Jacques
等人。, 2008), whereas possible differences and similarities
between brain networks and neural dynamics involved in
the temporal organization of different types of personal
记忆 (EAM vs. 聚苯乙烯) have not been tested to date.
The present study had thus two main goals. 第一的, 我们
investigated brain regions supporting the temporal (ordi-
纳尔) organization of autobiographical knowledge into dif-
ferent lifetime periods along the personal timeline, 两个都
for EAMs and experience-near PS (autobiographical facts)
hereafter called “enPS”. To this purpose, we developed a
novel fMRI paradigm, in which participants were pre-
sented with labels corresponding to unique events from
their own lives (EAMs) or names of personally known indi-
个人 (enPS) belonging to different lifetime periods and
were asked to judge their relative chronological order. 在一个
previous behavioral study assessing temporal ordering of
autobiographical memories, a spatiotemporal interfer-
ence effect was reported, for which participants were
more accurate in judging the order of EAMs when the
response direction was compatible with a sagittal mental
timeline (IE。, future in front; Teghil, 马克, & Boccia,
2021). 因此, as a secondary aim, we also tested possible
brain networks supporting such a spatiotemporal compat-
ibility effect. 第二, we assessed context-dependent con-
nectivity within the autobiographical memory network
during temporal ordering of EAMs and enPS using gener-
alized psychophysiological interaction (gPPI). To assess
the convergent validity of our procedure, we further cor-
related differences in brain activation associated with tem-
poral ordering of EAMs and enPS within regions of the
autobiographical memory network with scores on the
Survey of Autobiographical Memory (SAM; Palombo,
威廉姆斯, Abdi, & 莱文, 2013), a previously validated
measure of trait mnemonics in everyday life, 管理的
outside the scanner.
Based on previous literature, we hypothesized that a
common network could support overall the temporal
organization of autobiographical information along
Teghil et al.
2257
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
lifetime periods. This network should involve not only the
medial temporal lobe and anterior temporal and medial
prefrontal regions but also posterior and sensory regions
that have been overall proposed to support the represen-
tation of the personal past at multiple levels of specificity
(D’Argembeau, 2020). Within this network, 然而, 我们
expected differences in brain activation supporting the
temporal ordering of autobiographical memories with dif-
ferent degrees of specificity, 因此, we hypothesized
that distinct nodes may play a different contribution to
the representation of the temporal order of EAMs and
enPS. 一方面, sensory regions such as the striate
and extrastriate cortex, as well as the hippocampus (HC;
D’Argembeau et al., 2015; St Jacques et al., 2008), 可能
be more strongly involved in supporting the chrono-
logical order of EAMs, which entail the integration of
fine-grained sensory details. 另一方面, 多-
modal integration brain regions such as the anterior
temporal lobe, which have been associated to higher-
order features of the representation of the personal time-
线 (D’Argembeau, 2020), may be more strongly involved
in enPS. 而且, we expected that judging the relative
order of EAM versus enPS should be associated to specific
context-dependent connectivity patterns within the
autobiographical memory network. Previous studies on
effective connectivity between brain regions supporting
autobiographical memory (McCormick, Barry, Jafarian,
巴恩斯, & Maguire, 2020; Nawa & Ando, 2020) highlighted
a key role of the vmPFC in driving memory retrieval, 和
this region has been implicated in the integration of, 作为
well as in the switching between, different levels of
autobiographical knowledge (D’Argembeau, 2020). 我们
thus speculated that connectivity patterns of the vmPFC
may discriminate between the EAM and enPS conditions.
方法
参加者
Thirty-one healthy young volunteers (mean age = 26.87
年, 标准差= 2.63; 20 女性) took part in the study. All par-
ticipants were right-handed and had normal or corrected-
to-normal vision.
Sample size was defined a priori using G*Power ( Ver-
锡安 3.1.9.6; Faul, Erdfelder, Lang, & Buchner, 2007) 到
achieve a statistical power higher than 95%, considering
an alpha of .05. The effect size (ηp
2 = .23) was derived from
a previous study (Teghil, 马克, & Boccia, 2021). The total
sample size resulting from the power analysis was 29; 骗局-
sidering possible dropouts, we finally enrolled 31 individ-
乌尔斯. This sample is in line with previous fMRI studies
investigating memory for realistic materials (Bromis,
Raykov, Wickens, Roseboom, & Bird, 2022; Raykov, Keidel,
Oakhill, & Bird, 2021). None of the participants had a cur-
rent or previous history of neurological or psychiatric dis-
orders, nor alcohol or drug abuse, as assessed during an
informal interview before testing. The study was designed
in accordance with the principles of the Declaration
of Helsinki and was approved by the Ethical Committee
of IRCCS Fondazione Santa Lucia, 罗马 (Prot.
CE/PROG.824). Written informed consent was obtained
from all individual participants included in the study.
Stimuli Collection
Stimuli to be presented during fMRI were collected out-
side the scanner (mean distance between stimuli collec-
tion and scanning was 5.29 天, 标准差= 6.50), using an
adapted version of the Autobiographical Fluency Task by
Dritschel, 威廉姆斯, Baddeley, and Nimmo-Smith (1992).
For each of five life periods (5–11 years, 11–14 years,
14–19 years, >19 years excluding the last 12 月,
and last year), we asked participants to report personal
事件 (EAMs) and names of friends, 教师, 学校-
伙伴, or colleagues (enPS) corresponding to those
periods, as many as possible. Participants were asked to
provide a personalized label that was meaningful to them
and that allowed them to unambiguously identify the spe-
cific EAM or enPS. They were instructed to report only
events that occurred at a specific time and place and to
provide names of persons that were not associated to
more than one life period (例如, they were asked to avoid
reporting names of siblings or other life-long family mem-
bers). Ninety seconds were given for each combination of
period and memory category (EAM or enPS; Dritschel
等人。, 1992). After task completion, participants were
asked to report when the events occurred (for EAMs)
and when they first met the persons they named (为了
enPS). The whole procedure lasted ∼30 min.
平均而言, participants reported a total number of
35.63 (标准差= 7.75) EAMs and 50.13 (标准差= 12.53) enPS.
The first two items reported for each period and memory
category were used in the fMRI task, assigning them ran-
domly to the compatible and noncompatible conditions
(见下文). Different labels were presented across condi-
系统蒸发散 (compatible and noncompatible) to avoid spurious
effects because of item repetition. 然而, items from
the same periods were presented in different conditions,
thus allowing to control for effects of age/remoteness
between EAMs and enPS.
fMRI Task
We used a factorial 2 × 2 设计, with the factors memory
类别 (EAM vs. enPS) and task condition (遗憾-
ble vs. noncompatible with the mental timeline). 因为
of the multifactorial design, which was aimed to test the
effect of the main conditions rather than the effect of one
condition upon the other (IE。, cross-domain inter-
行动), the fMRI task was developed as a block design.
During fMRI, labels corresponding to EAMs and enPS col-
lected before scanning (see above) were presented one
at a time, in an unbroken sequential manner, in four seri-
ally balanced sequences (one for each memory category
Teghil et al.
2258
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
and task condition), in which each stimulus preceded
and followed every other stimulus the same number of
次 (Aguirre, 2007; Nonyane & Theobald, 2007).
In each trial, participants were instructed to decide
whether the currently presented stimulus preceded or
followed the previously shown stimulus in chronological
命令 (IE。, along their personal timeline; 数字 1).
Responses were provided using a two-button MRI-
compatible keypad. In the compatible condition, partici-
pants were asked to press the “backward” button if the
current stimulus temporally preceded the previous one
and the “forward” button if it followed the previous one;
this mapping was reversed in the noncompatible condi-
的 ( backward/forward responses for EAMs/enPS
following/preceding the previous one; Teghil, 马克, &
Boccia, 2021).
Four runs were acquired for each participant. To ensure
that instructions were properly understood at the begin-
ning of each scan, task conditions (compatible and
数字 1. Example of stimuli presented during the fMRI task. 这
task was conceived as a 1-back task; in each trial, participants had to
decide whether the currently presented personal event (episodic
autobiographical memory condition; (A) or personally known person
(experience-near personal semantics condition; (乙) followed or
preceded the previously presented one in chronological order along
one’s own personal timeline (stimuli showed in (A) corresponds to
actual labels provided by one of the authors during preliminary testing
of the fMRI paradigm).
noncompatible) were presented across runs; written
instructions were presented at the beginning of each
跑步. The order of the four runs was counterbalanced
across participants. Within each run, labels referring to
EAMs and enPS were presented in different blocks, follow-
ing an ABBA order. Ten blocks were presented in each run.
Each block lasted 25,000 毫秒. In each block, 10 labels
were presented for 2000 毫秒, followed by a fixation point
(500 毫秒). A fixation point was also presented during
interblock intervals, lasting 15,000 毫秒. Stimuli were gen-
erated using E-Prime 3.0 (Psychology Software Tools) 和
projected on a translucent screen that participants saw
through a mirror; accuracy and RTs were collected.
Immediately before fMRI, outside the scanner, partici-
pants performed a short familiarization session with the
same structure of the experimental task (labels referring
to standard EAMs and enPS, 例如, “first job” and “work-
mate”, were presented during this phase, for a total
number of 10 trials for each memory category and task
状况).
SAM (Palombo et al., 2013)
The SAM is a self-report questionnaire, assessing episodic
autobiographical (eight items), personal and general
semantic memory (six items), spatial memory (six items),
and future thinking (six items). Items are rated on a
5-point Likert scale (从 1 = strongly disagree to 5 =
strongly agree). The SAM has been validated and used
in different behavioral and neuroimaging studies on auto-
biographical memory (例如, Fan, Abdi, & 莱文, 2021;
Petrican, Palombo, Sheldon, & 莱文, 2020; Sheldon,
Farb, Palombo, & 莱文, 2016; Palombo et al., 2013). Scor-
ing was performed for each participant according to the
procedure by Palombo et al. (2013).
Image Acquisition
MR images were collected using a high-performance 3 时间
scanner (Siemens MAGNETOM Prisma) equipped with
a 32-channel head coil. Functional, whole-brain MR
images were acquired with a T2 -weighted gradient-echo
EPI sequence, a multiband factor of 4, and an isotropic
voxel size of 2.4 mm3 (60 slices, field of view = 208 ×
208 mm2, repetition time [TR] = 1100 毫秒, echo time
[TE] = 30 毫秒, flip angle = 65°, no in-plane acceleration;
徐等人。, 2013; Feinberg et al., 2010; Moeller et al., 2010).
In each run, we acquired 370 fMRI volumes, including six
dummy scans before each run, which were discarded.
Two spin-echo EPI volumes with phase encoding in
opposite direction, no multiband acceleration, 和
same geometrical and sampling properties of functional
runs were acquired for field mapping (= 80 毫秒,
TR = 7000 毫秒).
T1-weighted structural images were acquired on each
subject using an MPRAGE (magnetization-prepared rapid
gradient-echo) sequence with perspective motion
Teghil et al.
2259
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
correction and selective reacquisition of data corrupted by
motion based on interleaved 3-D EPI navigators (Tisdall
等人。, 2012; 赫斯, Tisdall, Andronesi, Meintjes, & van der
Kouwe, 2011). Volumetric imaging included 176 slices,
isotropic resolution = 1 mm3, TR = 2500 毫秒, =
2 毫秒, inversion time = 1070 毫秒, flip angle = 8°.
The MRI acquisition included other scans not used for
the present study.
Behavioral Analyses
Behavioral data were analyzed using SPSS (IBM SPSS Sta-
tistics 20). 第一的, 2 × 2 within-subject ANOVAs were per-
formed on accuracy and RTs in the temporal ordering task,
with the factors Memory Category (EAM vs. enPS) and Task
Condition (compatible vs. noncompatible). 还, we tested
whether the temporal distance between presented items
differently affected temporal ordering of EAMs and enPS.
We categorized each stimulus as coming from the same
lifetime period of the previous one, or as being separated
from the previous one by one, 二, 三, or four lifetime
periods. Because two stimuli from each period were pre-
sented in each carryover sequence and the first stimulus of
the sequence could not be judged (see the fMRI task
description reported above), the maximum number of
correct responses was different between levels of the fac-
tor temporal distance (零, 一, 二, 三, or four life-
time periods). 因此, for each level of temporal distance,
we calculated an accuracy score as the proportion of cor-
rect responses. We then performed a 2 × 5 within-subject
ANOVA on accuracy in the task with the factors Memory
类别 (EAM vs. enPS) and Temporal Distance (无论
the to-be-judged stimulus belonged to the same lifetime
period of the previous one, or it was separated from the
previous one by one, 二, 三, or four lifetime periods).
Analysis of Imaging Data
Image preprocessing and analysis were performed using
SPM12 (https://www.fil.ion.ucl.Ac.uk/spm). A field map
was computed from the spin-echo EPI images acquired
with opposite encoding polarity (Holland, Kuperman, &
戴尔, 2010). All fMRI images were simultaneously cor-
rected for head movements and B0 distortion, 包括
motion × field interaction (realignment and unwarping;
安德森, Hutton, Ashburner, 车工, & 弗里斯顿, 2001)
using the first volume as reference. After slice-timing cor-
反应, the images of each participant were coregistered
onto the respective T1-weighted image and normalized to
the standard MNI-152 template using the T1 image as a
来源 (voxel size: 2.4 × 2.4 × 2.4 mm3). Images were
finally smoothed using a 6-mm FWHM isotropic Gaussian
kernel.
Functional images were analyzed for each participant
separately on a voxel-by-voxel basis, according to the gen-
eral linear model. Neural responses during EAM and enPS
blocks were modeled as boxcar functions, convolved with
a canonical hemodynamic response function, and used as
separate predictors in the general linear model (one for
each experimental condition). Interblock intervals were
also modeled in relation to the nature of the previous
block (EAM-rest or enPS-rest) and treated as baseline.
Whole-brain Analyses
To identify brain regions generally supporting the tempo-
ral organization of autobiographical information, 没有
considering the specific memory category or task condi-
的, we first computed an omnibus F-contrast comparing
all experimental conditions against the baseline; 仅有的
positive activations were tested (the t-contrast map was
used as an inclusive mask thresholded at p < .5). The
resulting statistical parametric map was thresholded using
p < .05 family-wise error (FWE) and a cluster size k > 30
voxels. For each subject and region, a regional estimate of
the amplitude of the hemodynamic response in each
experimental condition was calculated entering a spatial
average (across all voxels in the region) of the prepro-
cessed time series into the individual general linear
型号. 然后, regional hemodynamic responses were
analyzed using 2 × 2 ANOVAs, with Memory Category
and Task Condition as independent variables. 下列的
Bonferroni’s procedure, alpha level for these ANOVAs
was set at p < .002.
In a separate voxel-wise analysis, we investigated brain
regions more strongly involved in supporting the tempo-
ral ordering of autobiographical information depending
on the memory category (EAM or enPS). Thus, contrast
maps resulting from the contrasts EAM–enPS and enPS–
EAM at the first-level analysis were entered into second-
level random effects analyses, and statistical inference for
each contrast was derived using a one-sample t test. The
resulting statistical parametrical maps were thresholded
using p < .05 FWE at the peak level and a cluster size
k > 30 voxels.
ROI Analyses
We further investigated activation specifically related to
the temporal sequencing of EAM and enPS in relation
to one another within the brain network generally sup-
porting autobiographical memory. The same steps
described above for voxel-wise analyses comparing EAM
and enPS were performed in a series of independent and
theoretically motivated ROIs. Ten spherical ROIs (10-毫米
radius) were centered on peak coordinates derived from
a previous activation likelihood estimation meta-analysis
on autobiographical memory (Teghil, Bonavita, 等人。,
2021). The following ROIs were derived: 左和右
angular gyrus (AG), left and right anterior middle tempo-
ral gyrus (aMTG), left and right HC, left and right PCC,
right posterior cerebellum (pCer), and left vmPFC (岑-
ter coordinates of each ROI are reported in Table 1).
One-sample t tests in these ROIs were performed for
Teghil et al.
2260
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
桌子 1. ROIs
Region
AGl
AGr
aMTGl
aMTGr
HCl
HCr
PCCl
PCCr
pCer
vmPFCl
X
−46
52
−58
58
−24
26
−8
10
26
−4
y
−68
−68
−4
−4
−26
−14
−54
−50
−80
50
z
32
32
−22
−18
−16
−18
14
32
−34
−6
For each ROI, center MNI coordinates are reported.
the t-contrast comparing the two memory categories
(EAM and enPS) by applying a Bonferroni-corrected
threshold of p < .005 (0.05/10). ROI analyses were per-
formed using the MarsBaR toolbox (https://marsbar
.sourceforge.net). Finally, two-tailed Pearson correlation
coefficients were calculated between BOLD signal change
between the two memory categories, which reflects neu-
ral activity unique of ordering enPS compared with order-
ing EAMs, within the ROIs and participants’ scores on the
SAM, entering total SAM score and scores on the Epi-
sodic, Semantic, Spatial, and Future Thinking subscales
in the correlation matrix (since SAM scores were not
available for six participants, correlation analyses were
performed on n = 25).
gPPI Analysis
Context-dependent interactions between brain regions
involved in the autobiographical memory network were
assessed using a gPPI approach (McLaren, Ries, Xu, &
Johnson, 2012), as implemented in the CONN Toolbox
(v. 20b; www.nitrc.org/projects/conn, RRID:SCR_009550;
Whitfield-Gabrieli & Nieto-Castanon, 2012). PPI is one of
the more simple and intuitive methods to test the interac-
tion between a physiological variable and an experimental
factor (Friston, 2011), allowing to assess whether and how
brain regions interact depending on the specific experi-
mental condition. In brief, neural responses in a target
region are modeled in terms of the interaction between
a psychological process and the neural signal from a
source region, thus allowing to assess whether the func-
tional connectivity between a source and a target region
is modulated by the experimental condition. In PPI, the
interaction can be interpreted as a change, following an
experimental manipulation, in the effective connectivity
between a brain region, expressing a significant interac-
tion, and a seed region (Friston, 2011). In block designs,
the PPI can be interpreted as the difference in the regres-
sion slopes when regressing activity in one brain region
on another one, under the two experimental contexts;
thus, the PPI corresponds to the change in effective con-
nectivity, given the experimental condition (Di, Zhang, &
Biswal, 2021).
BOLD signal in each target region was modeled as a
combination of (1) the effect of the two experimental con-
ditions (EAM and enPS), (2) the time course in the source
region, and (3) PPI terms corresponding to the product of
the first two regressors, expressing the interaction
between activation in each experimental condition and
the neural signal in the source region, according to
McLaren et al. (2012). gPPI analyses were performed
between the 10 theoretically motivated ROIs described
in the previous step. All of these regions have been found
to be intrinsically connected (default mode network
[DMN]) and have been recently hypothesized to be key
nodes of a larger brain network involved in transmodal
high-level processes (e.g., declarative memory, autobio-
graphical memory, verbal semantics, and cognitive con-
trol; Margulies et al., 2016). Also, the pCer is functionally
connected to the autobiographical memory network both
Figure 2. Regions involved in temporal sequencing of autobiographical information without considering memory category and compatibility with
the mental timeline (see Table 2 for labels and results).
Teghil et al.
2261
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
o
c
n
_
a
_
0
1
9
0
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 2. Results of the 2 × 2 ANOVAs Performed in Brain Regions Generally Supporting the Temporal Sequencing of Autobiographical Information along the Autobiographical
Knowledge Base
Memory Category
Task Condition
Memory Category × Task Condition
Label
LPC
pHC
PCC
vmPFC
aIns
IFG
SMA
midFG
LFC
preCG
CC
BG
Cerebellum
Vermis
Hemisphere
F(1, 30)
p
R
L
R
L
R
L
L
R
L
R
R
L
R
R
L
R
L
L
R
L
L
2.74
8.33
8.22
5.34
94.72
22.64
65.09
2.10
2.29
8.94
4.70
5.27
3.62
4.30
0.49
6.34
.11
.007 (EAM > enPS)
.008 (EAM > enPS)
.028 (EAM > enPS)
<.001 (enPS > EAM)
<.001 (enPS > EAM)
<.001 (enPS > EAM)
.16
.14
.006 (enPS > EAM)
.038 (enPS > EAM)
.029 (enPS > EAM)
.067
.047 (enPS > EAM)
.49
.017 (EAM > enPS)
17.11
<.001 (EAM > enPS)
6.91
0.09
0.73
6.20
.013 (enPS > EAM)
.76
.40
.019 (enPS > EAM)
η p
2
.08
.22
.22
.15
.76
.43
.68
.07
.07
.23
.14
.15
.11
.13
.02
.17
.36
.19
.003
.02
.17
F(1, 30)
p
0.01
0.21
2.50
0.01
0.64
0.40
2.72
3.82
4.40
2.25
4.42
4.27
1.93
2.77
2.60
<0.001 0.08 0.76 1.06 0.39 0.91 .96 .65 .12 .94 .43 .53 .11 .06 .045 (NC > C)
.14
.044 (NC > C)
.048 (NC > C)
.18
.11
.12
.98
.78
.39
.31
.54
.35
η p
2
<.001
.01
.08
<.001
.02
.01
.08
.11
.13
.07
.13
.12
.06
.08
.08
<.001
.003
.02
.03
.01
.03
F(1, 30)
0.01
0.08
0.31
0.04
0.01
0.51
0.10
0.16
<0.001
1.72
0.03
0.22
0.19
0.01
0.88
0.08
0.12
0.16
0.06
0.01
0.02
p
.92
.78
.58
.85
.92
.48
.76
.69
.99
.20
.86
.65
.66
.94
.36
.78
.73
.69
.81
.93
.90
η p
2
<.001
.003
.01
.001
<.001
.02
.003
.01
<.001
.05
.001
.01
.01
<.001
.03
.003
.004
.01
.002
<.001
.001
Results surviving Bonferroni correction ( p < .002) are reported in bold, together with the direction of the observed effect. R = right; L = left; LPC = lateral parietal cortex; pHC = posterior portion of the
HC; aINS = anterior insula; IFG = inferior frontal gyrus; midFG = middle frontal gyrus; LFC = lateral frontal cortex; preCG = precentral gyrus.
T
e
g
h
i
l
e
t
a
l
.
2
2
6
2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
o
c
n
_
a
_
0
1
9
0
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
during rest and autobiographical memory retrieval (Addis,
Moloney, Tippett, Roberts, & Hach, 2016). ROI-to-ROI
parametric maps were thresholded using threshold-free
cluster enhancement analyses (Smith & Nichols, 2009)
with an FWE-corrected threshold of p < .05 at the con-
nection level.
RESULTS
Behavioral Results
The 2 × 2 ANOVA on accuracy showed a trend toward a
main effect of Memory Category (EAM: M = 73.16, SD =
2.19; enPS: M = 74.93, SD = 2.33) F(1, 30) = 4.03, p =
.054, ηp
2 = .12, whereas the effect of Task Condition
was significant (compatible: M = 76.89, SD = 2.70;
noncompatible: M = 71.21, SD = 2.27), F(1, 30) = 6.25,
p = .018, ηp
2 = .17. There was no significant interaction
effect, F(1, 30) = 0.32, p = .57, ηp
2 = .01. Concerning
RTs, there was a significant main effect of Memory Cate-
gory, with participants being faster to respond to enPS
(M = 1079.76, SD = 130.55) than to EAMs (M =
1131.58, SD = 148.72), F(1, 30) = 18.96, p < .001, ηp
2 =
.39. The effect of Task Condition was also significant,
F(1, 30) = 8.16, p = .008, ηp
2 = .21, with faster responses
in the compatible (M = 1077.93, SD = 27.09) than in the
noncompatible condition (M = 1133.41, SD = 26.82). The
interaction effect was not significant, F(1, 30) < .001,
p = .1, η2
p < .001.
Concerning the effects of the temporal separation
between presented EAMs and enPS, we found a trend
toward a significant main effect of Memory Category,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
o
c
n
_
a
_
0
1
9
0
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Brain activations more strongly associated with temporal ordering along lifetime periods for EAM compared with enPS (shown in red to
yellow) and for enPS compared with EAM (shown in blue to green). (A) Axial view. (B) Sagittal view.
Teghil et al.
2263
F(1, 30) = 3.96, p = .056, ηp
2 = .12, with participants more
accurate in ordering enPS (M = .84, SD = .025) than EAMs
(M = .823, SD = .024). There was a significant effect of
Temporal Distance, F(1, 1.7) = 43.03, Greenhouse–Geisser
corrected, p < .001, ηp
2 = .59. Post hoc analyses applying
Bonferroni’s correction showed that all comparisons were
significant, with the proportion of correct responses
increasing with increasing temporal distance between
consecutive stimuli (same lifetime period: M = .72,
SD = .02; 1 lifetime period distance: M = .79, SD = .03;
2 lifetime periods distance: M = .85, SD = .03; 3 lifetime
periods distance: M = .88, SD = .03; 4 lifetime periods
distance: M = .91, SD = .3). The interaction between
Memory Category and Temporal Distance was not signifi-
cant, F(1, 2.73) = 0.41, Greenhouse–Geisser corrected,
p = .73, ηp
2 = .01.
Whole-brain Analyses
As a first step, we investigated brain networks generally
supporting the temporal sequencing of autobiographical
information along the autobiographical knowledge base.
We thus performed an omnibus F-contrast comparing all
experimental conditions versus the baseline, identifying
brain regions generally involved in ordinal processing of
autobiographical information independently from the
memory category or task condition. A broad network
was highlighted, spanning both hemispheres (Figure 2).
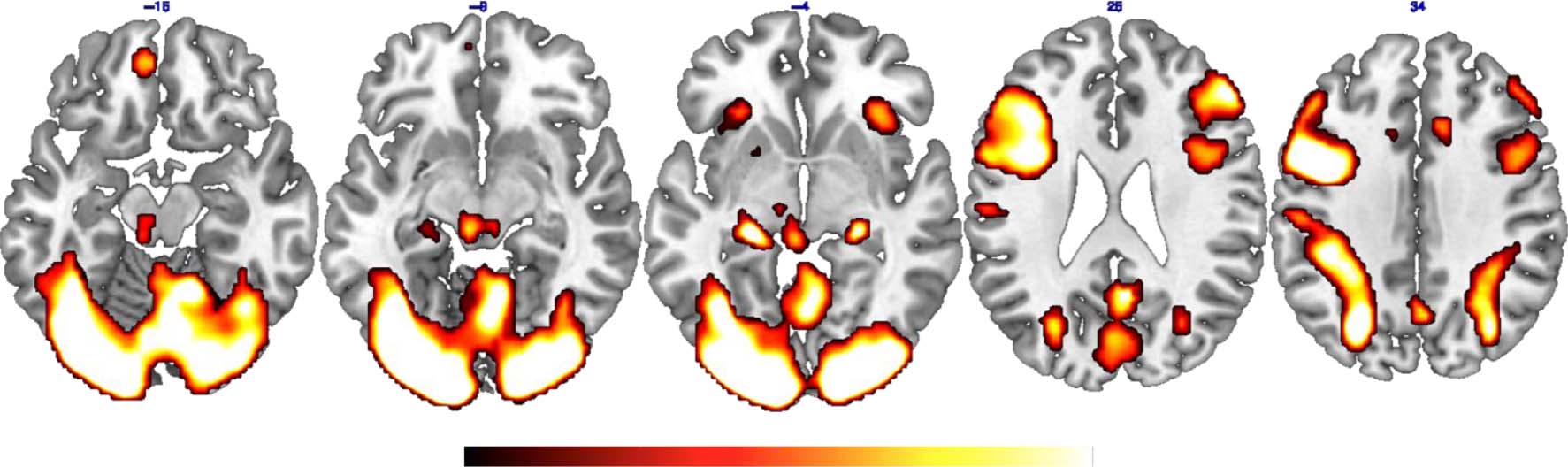
Specifically, in the occipital lobe, we found activation span-
ning the bilateral calcarine cortex (CC); the posteromedial
cortex (pCu) was also activated. Activation was also found
in the bilateral HC (posterior portion), in lateral parietal
regions, in both hemispheres, and in the left precentral
gyrus. The bilateral anterior insula was also activated,
spanning the inferior frontal gyrus in the right hemi-
sphere. In the frontal lobes, further activations were found
corresponding to the right middle frontal gyrus, bilateral
SMA, and left vmPFC. The bilateral cerebellum and the
BG in the left hemisphere were also activated. Results of
the 2 × 2 ANOVAs with the factors Memory Category and
Task Condition in each region showed that the bilateral
pCu and the left vmPFC were more activated during the
ordering of enPS compared with EAMs, whereas the left
CC was more strongly involved in the EAM than in the
enPS condition. No other significant effect was detected.
Results are summarized in Table 2.
Next, we investigated brain networks more strongly
involved in the temporal organization of EAMs and enPS.
Because no significant interaction effect between mem-
ory category and task condition was found either at the
behavioral or neural level in the analyses mentioned
above, only the factor memory category was considered
in following analyses.
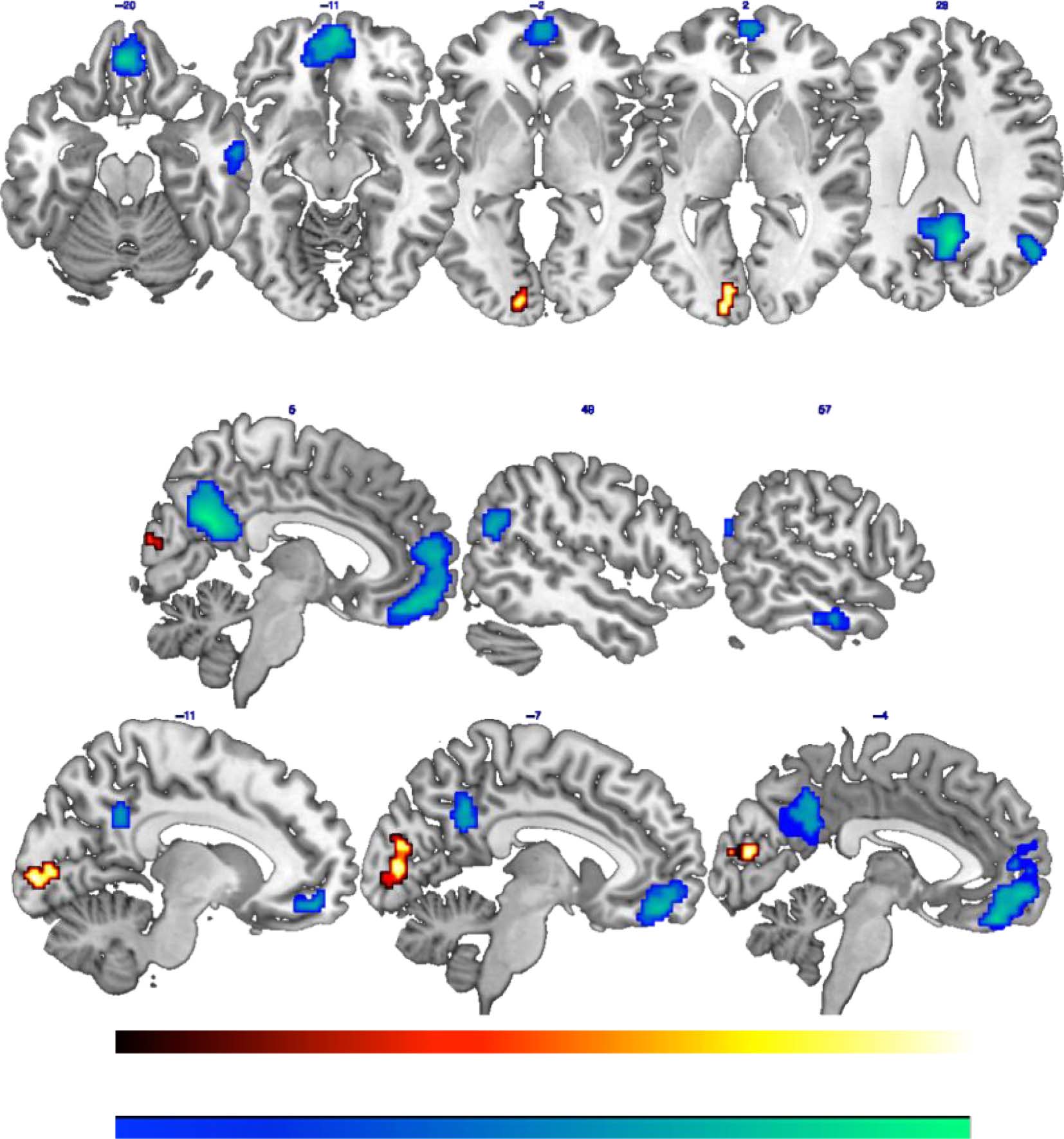
The contrast EAM > enPS highlighted a cluster in the
left CC that was more strongly activated during ordering
of EAMs than of enPS (见图 3 和表 3). Concern-
ing the reverse contrast (enPS > EAM), results of the
whole-brain analyses showed stronger activation during
ordering of enPS than EAMs in a set of brain regions includ-
ing the right pCu/PCC and the left vmPFC on the medial
brain surface and the AG and aMTG of the right hemi-
sphere on the lateral surface (见图 3 和表 3).
ROI Analyses
Further analyses were performed to investigate differ-
ences in brain activation for temporal ordering of EAM
and enPS within the core brain network supporting
桌子 3. Brain Regions More Strongly Activated during Temporal Ordering of EAM and enPS Compared with the Other Condition
Region
Hemisphere
X
y
EAM > enPS
CC
enPS > EAM
pCu/PCC
vmPFC
AG
aMTG
L
右
右
右
L = Left hemisphere; R = Right hemisphere.
−8
−4
−11
6
−6
−1
1
1
52
61
−86
−83
−93
−54
−52
44
58
51
−66
−9
z
4
14
0
26
38
−20
−8
−15
24
−22
时间
p-FWE
Voxels
7.16
7.09
7.03
11.72
7.90
9.99
9.84
9.62
8.46
7.66
.003
.004
.005
.000
.000
.000
.000
.000
.000
.001
113
494
623
81
46
Teghil et al.
2264
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
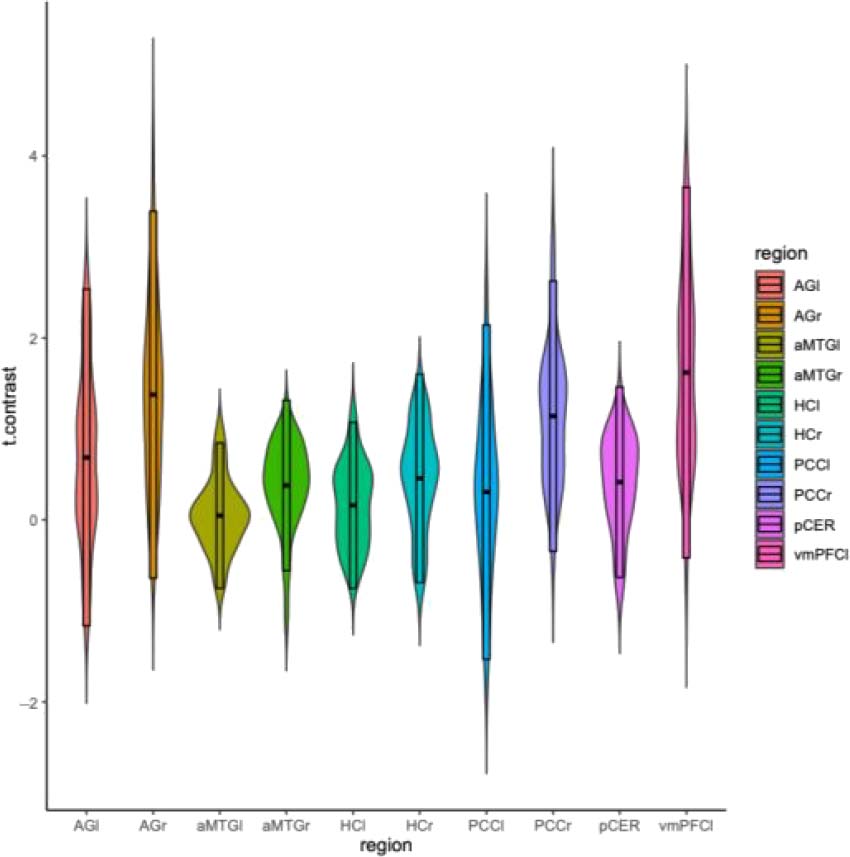
autobiographical memory (bilateral AG, aMTG, HC, 和
PCC; right pCer; left vmPFC; Teghil, Bonavita, 等人。,
2021). A significant stronger activation in enPS com-
pared with EAM was found in the left and right AG, 正确的
HC, right PCC, right aMTG,
left vmPFC, and pCer
(Figure 4A).
Correlation analyses with self-report measures of
everyday memory (Palombo et al., 2013) 表明
total SAM scores and scores on the Episodic subscale
correlated positively with the portion of activation spe-
cifically associated to enPS compared with EAM in the
right AG (SAM total: r = .44, p = .029; SAM Episodic:
r = .58, p = .003) and in the left aMTG ROIs (SAM total:
r = .47, p = .019; SAM Episodic: r = .42, p = .035).
Activation difference in the right HC ROI was also signif-
icantly positively correlated with the SAM total (r = .47,
p = .019), Episodic (r = .47, p = .018), and Semantic
(r = .54, p = .005) subscales. A significant positive cor-
relation was also found between differences in BOLD
signal in the right PCC ROI and SAM Episodic scores
(r = .43, p = .030; Figures 4B and 5). The full correla-
tion matrix is reported in Table 4. No significant correla-
tion was found between BOLD signal extracted in the
ROIs and scores on the Spatial and Future Thinking sub-
scales. This was an exploratory analysis, and findings
should be considered with caution, because not all cor-
relations would survive a stringent correction for multi-
ple comparison. Results of the correlations, 然而,
show an association between differences in task-related
brain activation in response to the temporal ordering of
different categories of autobiographical information in
key nodes of the autobiographical memory network
(Teghil, Bonavita, 等人。, 2021) and individual variations
in trait mnemonics. 而且, no correlation was
observed between differences in task-related brain acti-
vation and scores on the Spatial and Future Thinking
subscales, in line with findings from the original article
by Palombo et al. (2013) that these subscales could be
more reliably separated from those assessing episodic
and semantic memory. These results suggest that differ-
ences in brain activation between the two memory catego-
ries evoked by our paradigm could be reliably associated
to autobiographical memory processes.
数字 4. (A) Violin plots of t statistics for the contrast enPS > EAM for the
10 ROIs. Significant results of the one-sample t tests are marked with an
asterisk ( p < .005; the results of the one-sample t tests on the reverse
contrast are not shown because they yield to symmetrical results). (B)
Correlation plot showing the association between participants’ scores on the
SAM and activation difference between enPS and EAM in the ROIs. Only
significant correlations are shown ( p < .05, two-tailed). l = left hemisphere;
r = right hemisphere; SAM.T = SAM total; SAM.Ep = SAM Episodic Memory
subscale; SAM.Sem = SAM Semantic Memory subscale; SAM.Sp = SAM
Spatial Memory subscale; SAM.FT = SAM Future Thinking subscale.
Figure 5. Scatter plot showing significant associations between SAM scores and activation differences between enPS and EAM in the ROIs.
Teghil et al.
2265
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
o
c
n
_
a
_
0
1
9
0
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 4. Pearson’s Correlation Coefficients between Scores on the SAM and Activation Difference between enPS and EAM in the ROIs
SAM Total
SAM Ep
SAM Sem SAM Sp
SAM FT
AGl
SAM total
1
.78***
.72***
.60**
.55**
SAM Ep
.78***
1
.45*
SAM Sem
.72***
.45*
1
SAM Sp
SAM FT
.60**
.55**
AGl
AGr
HCl
HCr
PCCl
PCCr
aMTGl
aMTGr
vmPFCl
pCer
.30
.44*
.12
.47*
.26
.26
.47*
.28
.05
.40*
.38
.14
.28
.58**
.34
.47*
.31
.43*
.42*
.32
.23
.38
.38
.22
.21
.03
.08
.54**
.14
−.01
.33
.28
−.05
.22
.38
.38
1
.02
.30
.32
.14
.30
.28
.11
.23
.16
.15
.27
.14
.22
.02
1
.05
.17
−.19
.04
.03
.11
.22
.07
−.23
.18
AGr
.44*
.58**
.03
.32
.17
.30
.28
.21
.30
.05
1
.51**
.51**
1
.38
.12
.41*
.29
.54**
.29
.47*
.74***
.34
.32
.52**
.49*
.23
.24
.45*
.48*
HCl
.12
.34
.08
.14
−.19
.38
.34
1
HCr
.47*
.47*
.54**
.30
.04
.12
.32
.53**
.53**
1
.46*
.50*
.45*
.29
.42*
.19
.60**
.64**
.21
.35
.13
.10
PCCl
PCCr
aMTGl
aMTGr
vmPFCl
pCer
.26
.31
.14
.28
.03
.41*
.52**
.46*
.29
1
.33
.31
.33
.53**
.38
.26
.43*
−.01
.11
.11
.29
.49*
.50*
.42*
.33
1
.47*
.42*
.33
.23
.22
.54**
.23
.45*
.19
.31
.22
.22
1
.63**
.35
.39
.45*
.13
.48*
.28
.32
.28
.16
.07
.29
.24
.60**
.64**
.33
.63**
.45*
1
.34
.19
.05
.23
−.05
.15
−.23
.47*
.45*
.21
.13
.53**
.35
.13
.34
1
.40*
.38
.22
.27
.18
.74***
.48*
.35
.10
.38
.39
.48*
.19
.45*
.45*
1
SAM Ep = SAM Episodic memory subscale; SAM Sem = SAM Semantic Memory subscale; SAM Sp = SAM Spatial Memory subscale; SAM FT = SAM Future Thinking subscale; l = left hemisphere; r = right
hemisphere.
* p < .05.
** p < .01.
*** p < .001.
T
e
g
h
i
l
e
t
a
l
.
2
2
6
6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
o
c
n
_
a
_
0
1
9
0
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Schematic illustration
of the results of the gPPI
analyses. Significant effects are
represented by arrows,
identifying seed-to-target
direction. Seeds showing
significant PPIs with target
regions are identified by darker
colors. ROIs showing a stronger
activation during enPS
compared with EAM are
highlighted by red edges.
Table 5. Results of the gPPI Analysis for the Contrast enPS > EAM
Analysis Unit
Cluster 1/1
Connection PCCl–PCCr
Connection PCCl–AGr
Connection PCCr–aMTGr
Connection vmPFCl–PCCl
Connection AGr–PCCr
Connection AGr–aMTGr
Statistic
TFCE = 27.13
时间(30) = 4.20
时间(30) = 1.99
时间(30) = 0.79
时间(30) = 0.75
时间(30) = 0.71
时间(30) = 0.01
TFCE = threshold-free cluster enhancement; l = left hemisphere; r = right hemisphere.
p-unc
.001829
p-FWE
.004000
gPPI Analysis
讨论
Context-dependent connectivity between the 10 ROIs
described above was investigated using gPPI. 因此, 我们
assessed whether the functional connectivity between
each couple of regions within the brain network involved
in autobiographical memory from previous literature
(Teghil, Bonavita, 等人。, 2021) was affected by the experi-
mental condition (temporal ordering of EAMs or enPS).
For all seeds, functional connectivity was stronger during
the enPS than the EAM condition. In detail, 我们发现,
during enPS compared with EAM, the BOLD time course
in the right PCC and in the right AG was predicted by the
PPI in the left PCC. 反过来, activity in the right PCC and in
the right aMTG was predicted by the interaction between
the experimental condition and the time course in the
right AG. Activity in the right PCC was also significantly
predicted by the PPI in the aMTG. 最后, neural activity
in the left PCC was predicted by the interaction between
the experimental condition and the time course in the left
vmPFC (见图 6 和表 5).
The present study investigated neural substrates of the
ordinal representation of different formats of autobio-
graphical information along one’s own personal timeline.
Both local regional effects and network-level interactions
were assessed.
第一的, voxel-wise analyses highlighted a distributed net-
work commonly involved in supporting the chronological
organization of autobiographical knowledge. Within this
网络, different brain regions were preferentially
involved in ordering EAMs and enPS along one’s own per-
sonal timeline. Concerning EAMs, we found a single clus-
ter in the left CC that was significantly more activated in
this condition. Different fMRI studies have reported activa-
tion of occipital areas during tasks relying on EAM ( Viard
等人。, 2011; Daselaar et al., 2008; Cabeza et al., 2004). 这样的
an activation has been associated with the construction of
a visual scene in service of episodic autobiographical
remembering (Cabeza & St Jacques, 2007), also in line
with neuropsychological evidence that lesions to the
Teghil et al.
2267
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
visual cortex may lead to autobiographical memory impair-
评论 (Cabeza & St Jacques, 2007; Greenberg & 鲁宾,
2003). It has been proposed that the construction of a
visual scene, including its spatial layout, is an integral part
of reliving and characterizes specifically episodic remem-
bering (鲁宾 & Umanath, 2015). The recollection of com-
plex episodic details, 然而, involves both a memory
construction and a memory elaboration phase and
extends across several seconds (Daselaar et al., 2008;
康威 & Pleydell-Pearce, 2000). Stronger activation of
the left CC during EAM in the present study could thus
reflect a partial reactivation of visual features of EAMs dur-
ing the temporal ordering task. Activation of occipital
地区, together with that of lateral parietal regions and
the pCu, has been also reported when participants
decided which of two recently experienced events pre-
ceded the other, suggesting that these regions may sup-
port EAM sequencing through a spatialized representation
时间的 (D’Argembeau et al., 2015). Although the present
study does not allow to disentangle between these two
可能性, our results suggest that activation of regions
of the ventral visual stream may specifically characterize
temporal sequencing along lifetime periods of specific
事件, compared with that of PS information that,
although experience-near, do not possess the same level
of contextual specificity (Grilli & Verfaellie, 2014). 更远
studies will be needed to understand the specific role of
visual areas in supporting the temporal organization of
EAMs along lifetime periods, as well as the role of factors
such as the vividness of these memories.
Voxel-wise analyses also highlighted a set of brain
regions more strongly activated during temporal sequenc-
ing of enPS compared with EAM and involving the left
vmPFC, the right pCu/posterior cingulate, 和右边的
AG and aMTG.
Medial prefrontal regions interact with lateral temporal
and posterior parietal structures during the organization
of personal past experiences in thematically related clus-
特尔斯 (Demblon, Bahri, & D’Argembeau, 2016) 因此
have been proposed to be pivotal for the integration of
specific autobiographical elements within higher-order
autobiographical knowledge (D’Argembeau, 2020). 还,
the vmPFC represents temporal positions within sche-
matic sequences (Hsieh & Ranganath, 2015) 并且是
sensitive to the temporal organization of narratives seg-
ments within familiar scripts, over the order of multiple
minutes (Baldassano, Hasson, & Norman, 2018). 更远
supporting a possible role of this region in orientation in
时间, activation of the vmPFC was found in a previous
study when participants judged which of two common life
events was temporally closer to them (Peer, Salomon,
Goldberg, Blanke, & Arzy, 2015). These findings have been
recently extended to the domain of autobiographical
memory because, in individuals with highly superior
autobiographical memory, the left vmPFC shows
increased pattern distinctness for memories more distant
in time (Santangelo, Pedale, Macrì, & Campolongo, 2020),
supporting the possibility that this region contributes to
the representation of the temporal structure of one’s
own past.
Concerning the cluster of activation found in the
pCu/posterior cingulate, lesions to the right PCC impair
the ability to establish the relation between one’s own
position and a larger spatial context (Aguirre & D’Esposito,
1999). 的确, the retrosplenial complex has been shown
to code for imagined facing direction and location during
memory retrieval, allowing to situate one’s own imagined
position and heading direction with respect to a repre-
sentation of the local environment based on memory
(Marchette, Vass, Ryan, & 爱泼斯坦, 2014). 而且, 在
a recent fMRI study in which participants compared the
time elapsed between personal events at different time-
scales (小时, 天, week, 月), the PCC/pCu showed a
sensitivity for the temporal timescale, with a maximum
preference for longer timescales (IE。, 月; Monsa,
Peer, & Arzy, 2020). Because of its preference for longer
timescales, it is not surprising that the PCC/pCu was
more activated during temporal sequencing along life-
time periods for enPS than for EAMs; 的确, 回忆
for personally known persons span across timescales
inevitably longer than those of memories for specific
事件 (which span a few hours).
The AG has been implicated in egocentric spatial com-
假设 (Boccia, Nemmi, & Guariglia, 2014), and disrup-
tion of neural activity in this region reduces the tendency
to report personal memories in a first-person perspective
(Bonnici, Cheke, 绿色的, FitzGerald, & Simons, 2018).
尤其, activation of the right pCu, together with the ipsi-
lateral AG, has been recently shown to carry information
about the temporal distance between previously experi-
enced movie frames and their context (Foudil, Kwok, &
Macaluso, 2020), suggesting that these two regions may
participate in the temporal organization of cluster of
events along long timescales. 因此, present findings of
activation of the right retrosplenial complex together with
the right AG suggest that the PCC may be involved in the
representation of the order of autobiographical informa-
tion along the personal timeline, contributing to its orga-
nization also along the different lifetime periods (Monsa
等人。, 2020), and are overall in line with evidence that
movement in mental time is conceived in a spatialized
方式 (Bender & Beller, 2014).
Concerning activation found in the aMTG, 前部
temporal regions are commonly activated during process-
ing of PS information (Renoult et al., 2012; Svoboda et al.,
2006), and the MTG has been particularly implicated in
representing general events and knowledge about signifi-
cant persons and autobiographical facts (Martinelli,
Sperduti, & Piolino, 2013). 因此, present results are con-
sistent with proposals that the anterior temporal lobe
contributes specifically to the representation of higher-
order aspects of autobiographical memory, 例如
organization of autobiographical knowledge into lifetime
periods and general events (D’Argembeau, 2020).
Teghil et al.
2268
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Results of the analyses in a series of theoretically moti-
vated ROIs consistently involved in autobiographical
记忆 (Teghil, Bonavita, 等人。, 2021) further confirmed
findings from the voxel-wise analyses. 的确, we found
significantly stronger activation during ordering of enPS
compared with EAMs in the bilateral AG; right HC, PCC,
aMTG, and pCer; and left vmPFC.
The pCer shows intrinsic connectivity with the right AG,
MTG, and medial frontal cortex and, during the retrieval of
EAMs, interacts with the right PCC and MTG (Addis et al.,
2016), all regions found to be activated in the present
学习. 而且, disruption of activity in the right pCer
impairs semantic integration and retrieval (Gatti, Vecchi,
& Mazzoni, 2021; Gatti, Van Vugt, & Vecchi, 2020), in line
with a possible role of this region in the flexible use of
stored knowledge. 因此, although the contribution of
the right pCer to autobiographical memory is still unclear,
our results suggest that this region may contribute to the
temporal organization of enPS.
As mentioned in the Introduction, the right HC has also
been implicated in representing PS information, 例如
repeated events (Addis, McIntosh, Moscovitch, Crawley,
& McAndrews, 2004). It has been proposed that the
involvement of medial temporal lobe structures in PS
depends on the extent to which these memories embed a
spatiotemporal context: more experience-near enPS,
although not to the same degree of EAMs, entail some spa-
tiotemporal information and thus rely on the medial-
temporal lobe; 反过来, more experience-far PS, 哪个
have undergone complete abstraction, map on cortical
networks involved in general semantic memory (Grilli &
Verfaellie, 2016). 这里, we tested the temporal organiza-
tion of enPS asking participants to retrieve the relative
order of names of people associated with different lifetime
periods; this type of autobiographical facts embeds a tem-
poral context by definition. Our results are thus compati-
ble with the possibility that a continuum exists between
episodic and semantic autobiographical memory, 和他们的
phenomenological features depend on the differential
weighting of different component processes (Renoult
等人。, 2012, 2016). This interpretation is in line with the
significant correlation we found between activity specifi-
cally associated with enPS in the right AG and PCC and
the left aMTG with the Episodic subscale of the SAM; 还,
enPS-specific activity in the right HC was correlated with
scores on both the Episodic and Semantic subscales.
Our final set of analyses assessed context-dependent
connectivity between the abovementioned ROIs using
gPPI (McLaren et al., 2012). The results showed that differ-
ent regions were more strongly connected during enPS
compared with EAM (数字 6). More in detail, we found
increased connectivity during the enPS compared with the
EAM condition between the vmPFC and the ipsilateral
PCC. 还, the connectivity between this latter region
and the right PCC and AG was increased during ordering
of enPS compared with EAMs. During the same condition,
increased connectivity was also observed between the
right AG and the ipsilateral PCC and aMTG. Results of
the gPPI analysis are thus consistent with those of the
voxel-wise and ROIs analyses and suggest overall that the
left vmPFC, the right PCC/pCu, and the right AG and aMTG
are key nodes in the network supporting temporal
sequencing of autobiographical information.
尤其, the set of brain regions commonly highlighted
by the present analyses considerably overlaps with the
“core” subsystem of the DMN, including the medial pFC,
the posteromedial cortex, the AG, and the aMTG
(Andrews-Hanna, Smallwood, & Spreng, 2014; Andrews-
Hanna, Reidler, 墓, Poulin, & 巴克纳, 2010). 在
line with recent suggestions that regions of the DMN
support different forms of complex thought characterized
by various degrees of abstraction (Smallwood et al.,
2021), including episodic and semantic memory (爱尔兰语 &
Vatansever, 2020), this network has been shown to be
involved during tasks requiring the application of previ-
ously learned schematic knowledge ( Vatansever, Menon,
& Stamatakis, 2017), of which enPS is an example. Specif-
ically, the left vmPFC, the AG, the anterior temporal
cortex, and the PCC have been strongly implicated in the
processing of the so-called “schemas,” conceived as high-
level knowledge structures abstracted across multiple
经历 (Gilboa & Marlatte, 2017). The sensitivity to
chronological order has been recently theorized to be
among the defining features of schemas, specifically in sit-
uations in which temporal order is relevant to guide
行为 (戈什 & Gilboa, 2014). More in detail, 一起
with the other nodes of the DMN, the mPFC, retrosplenial
cortex, and AG have been proposed to allow the instanti-
ation of particular types of schemas, the so-called “situa-
tion models,” that represent different kinds of relations,
including temporal ones, within a specific context, sup-
porting a wide range of complex cognitive functions
(Ranganath & Ritchey, 2012). Within this network, 这
AG and pCu/PCC may specifically support the orientation
within space and time (Ranganath & Ritchey, 2012), in line
with evidence that these regions are involved in spatial
导航 (Teghil, Bonavita, 等人。, 2021; Ekstrom,
Huffman, & Starrett, 2017; Spreng, 三月, & Kim, 2009;
Aguirre & D’Esposito, 1999) and with studies suggesting
that temporal relations between lifetime periods may be
represented in a spatialized manner (Teghil, 马克, &
Boccia, 2021; Miles, Nind, & Macrae, 2010; Arzy, Adi-Japha,
& Blanke, 2009). Present results thus suggest overall that
the temporal organization of autobiographical informa-
tion into different lifetime periods exploits situation
models developed within core DMN regions.
Our results may appear somewhat at odd with previous
文学, showing that brain regions often associated with
episodic autobiographical retrieval (see Boccia, Teghil, &
Guariglia, 2019, for a recent meta-analysis) were more
activated during temporal sequencing of enPS than of
EAMs. It is important to point out that here we aimed to
investigate temporal sequencing along one’s own per-
sonal timeline at the boundary between episodic and
Teghil et al.
2269
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
semantic autobiographical memory. 的确, the results of
behavioral analyses on accuracy in the ordering task as
a function of temporal distance between stimuli are con-
sistent with the possibility that both EAMs and enPS have a
specific temporal organization along the personal time-
线. 因此, ROI analyses showed that differences
in activation between memory categories in the autobio-
graphical memory network were correlated with scores
on the Episodic subscale of the SAM, supporting the pos-
sibility that these regions contribute to the temporal orga-
nization of enPS by processing contextual information.
Based on these results, future fMRI studies are warranted
to test the effect of temporal distance on regional activa-
的, using event-related designs.
One may also wonder whether the enPS labels triggered
the retrieval of specific events related to the cued persons.
If this were the case, slower RTs to enPS than to EAMs
should be expected, because enPS would entail recollect-
ing both the identity of cued persons and contextual
features of EAMs. 然而, 此情况并非如此, 因为
we observed faster RT to enPS than EAMs and no evidence
for significance differences between performances in the
EAM and enPS condition. 因此, present results may be
especially important to understand neural mechanisms
supporting the representation of temporal information
for enPS, which has been significantly less investigated
compared with EAM. Previous fMRI studies investigating
PS mainly required participants to retrieve repeated
事件 (例如, Levine et al., 2004) or to provide judgments
on statements concerning self-knowledge or self-traits
(例如, Araujo, 卡普兰, Damasio, & Damasio, 2014; Maguire
& Frith, 2003). To our knowledge, none of these studies
assessed the temporal organization of this personal knowl-
edge along different lifetime periods. Further studies,
using PS characterized by variable contextual specificity
(例如, different types of autobiographical facts, 例如
one’s job, street address, ETC。, as well as different concep-
tual categories of PS information) are thus needed to
understand whether this temporal organization is com-
mon to other types of enPS knowledge.
最后, temporal sequencing of enPS in our study
yielded to stronger activation compared with EAM in dif-
ferent brain regions, whereas ordering of EAMs entailed
stronger than enPS activation only of a single cluster. 这
fits well with findings of a recent fMRI study, reporting that
during the recalling of fictional talk show conversations
involving three fixed hosts and nine different celebrities,
the identity of the (repeated) hosts was strongly reinstated
in regions of the posteromedial network, whereas that of
这 (独特的) celebrities was not (Bromis et al., 2022). 我们的
findings that different regions of the DMN are activated
more strongly during temporal ordering of personally
known individuals than of single experienced events are
thus in line with the possibility that repeated elements
are more strongly represented at the neural level, 更远
suggesting that this may be also true for autobiographical
记忆. 与此相类似, core regions of the DMN such as
the vmPFC and the PCC have been also reported to be sen-
sitive to the presence of social information (Tso,
Rutherford, Fang, Angstadt, & 泰勒, 2018). 因此, 未来
studies specifically manipulating social significance are
warranted to understand whether and how the degree of
social information embedded within EAMs and enPS
affects the strength of their neural representation along
one’s own personal timeline.
全面的, present findings are consistent with the theo-
retical proposal that autobiographical knowledge is orga-
nized along lifetime periods according to temporal and
thematic relations (康威 & Pleydell-Pearce, 2000) 和
suggest that such an organization may be supported by
temporal properties of situation models. From a broader
看法, these results are in line with recent theoreti-
cal accounts highlighting the privileged role of regions of
the DMN in the gathering of multiple sources of informa-
tion across the cortex and thus in allowing a continuum of
abstraction from concrete, sensory-detailed representa-
tions typically involved in EAM to more abstract semantic
知识 (Smallwood et al., 2021; 爱尔兰语 & Vatansever,
2020). This conceptualization of PS would fit well with evi-
dence that more experience-near types of autobiographi-
cal knowledge possess at least partial spatial and temporal
特征 (Grilli & Verfaellie, 2014, 2016) and would provide
a biological substrate to proposals that enPS information
represents an intermediate entity between episodic and
semantic memory (Renoult et al., 2012).
致谢
This study was supported by funding from Sapienza University of
Rome to A. 时间. (Avvio alla Ricerca, 2021; 不. AR22117A4293B7F5).
Multiband EPI sequence and reconstruction software was
received from the University of Minnesota Center for Magnetic
Resonance Research. MPRAGE navigated sequence and recon-
struction software was received from the Massachusetts General
Hospital, Athinoula A. Martinos Center for Biomedical Imaging.
We would like to thank Nika Zahedi and the Levine Lab for shar-
ing scoring procedures for the Survey of Autobiographical
记忆.
Reprint requests should be sent to Alice Teghil, Department of
心理学, Sapienza University of Rome, Via dei Marsi, 78,
00185 罗马, 意大利, or via e-mail: alice.teghil@uniroma1.it, 或者
Maddalena Boccia, 心理学系, Sapienza Univer-
sity of Rome, Via dei Marsi, 78, 00185 罗马, 意大利, or e-mail:
maddalena.boccia@uniroma1.it.
Data Availability Statement
The datasets generated during and/or analysed during the
current study are available from the corresponding author
on reasonable request.
Author Contributions
Alice Teghil: 概念化; 形式分析; 资金
acquisition; 调查; 方法; Project adminis-
翻译; 可视化; Writing–original draft; Writing–
review & 编辑. Alessia Bonavita: 调查;
Writing–review & 编辑. Federica Procida: 调查;
Teghil et al.
2270
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Writing–review & 编辑. Federico Giove: Formal analy-
姐姐; 方法; 软件; Writing–review & 编辑.
Maddalena Boccia: 概念化; 形式分析;
方法; 项目管理; 监督; Visual-
化; Writing–review & 编辑.
Funding Information
Alice Teghil, Facoltà di Medicina e Psicologiaa, Sapienza Uni-
versità di Roma (https://dx.doi.org/10.13039/100010143),
grant number: Avvio alla Ricerca, AR22117A4293B7F5.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 到 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN ) 在这段时期
were M(一个)/米= .407, 瓦(oman)/米= .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Fulvio, JoCN, 34:1, PP. 1–3). 最后, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance. 这
authors of this article report its proportions of citations by
gender category to be as follows: M/M = .485; W/M = .235;
M/ W = .147; W/ W = .132.
参考
Addis, D. R。, McIntosh, A. R。, Moscovitch, M。, Crawley, A. P。, &
McAndrews, 中号. 磷. (2004). Characterizing spatial and temporal
features of autobiographical memory retrieval networks: A
partial least squares approach. Neuroimage, 23, 1460–1471.
https://doi.org/10.1016/j.neuroimage.2004.08.007, 考研:
15589110
Addis, D. R。, Moloney, 乙. E., Tippett, L. J。, 罗伯茨, 磷. R。, &
Hach, S. (2016). Characterizing cerebellar activity during
autobiographical memory retrieval: ALE and functional
connectivity investigations. Neuropsychologia, 90, 80–93.
https://doi.org/10.1016/j.neuropsychologia.2016.05.025,
考研: 27235570
Aguirre, G. K. (2007). Continuous carry-over designs for fMRI.
Neuroimage, 35, 1480–1494. https://doi.org/10.1016/j
.neuroimage.2007.02.005, 考研: 17376705
Aguirre, G. K., & D’Esposito, 中号. (1999). Topographical
disorientation: A synthesis and taxonomy. Brain, 122,
1613–1628. https://doi.org/10.1093/brain/122.9.1613,
考研: 10468502
安德森, J. L。, Hutton, C。, Ashburner, J。, 车工, R。, & 弗里斯顿,
K. (2001). Modeling geometric deformations in EPI time
系列. Neuroimage, 13, 903–991. https://doi.org/10.1006
/nimg.2001.0746, 考研: 11304086
Andrews-Hanna, J. R。, Reidler, J. S。, 墓, J。, Poulin, R。, &
巴克纳, 右. L. (2010). Functional-anatomic fractionation
of the brain’s default network. 神经元, 65, 550–562.
https://doi.org/10.1016/j.neuron.2010.02.005, 考研:
20188659
Andrews-Hanna, J. R。, Smallwood, J。, & Spreng, 右. 氮. (2014).
The default network and self-generated thought: 成分
流程, dynamic control, and clinical relevance. Annals of
the New York Academy of Sciences, 1316, 29–52. https://土井
.org/10.1111/nyas.12360, 考研: 24502540
Araujo, H. F。, 卡普兰, J。, Damasio, H。, & Damasio, A. (2014).
Involvement of cortical midline structures in the processing
of autobiographical information. 同行杂志, 2, e481. https://土井
.org/10.7717/peerj.481, 考研: 25097820
Arzy, S。, Adi-Japha, E., & Blanke, 氧. (2009). The mental time
线: An analogue of the mental number line in the mapping
of life events. Consciousness and Cognition, 18, 781–785.
https://doi.org/10.1016/j.concog.2009.05.007, 考研:
19553141
Arzy, S。, Collette, S。, Ionta, S。, Fornari, E., & Blanke, 氧. (2009).
Subjective mental time: The functional architecture of
projecting the self to past and future. European Journal of
神经科学, 30, 2009–2017. https://doi.org/10.1111/j.1460
-9568.2009.06974.X, 考研: 19912333
Arzy, S。, Molnar-Szakacs, 我。, & Blanke, 氧. (2008). Self in time:
Imagined self-location influences neural activity related to
mental time travel. 神经科学杂志, 28, 6502–6507.
https://doi.org/10.1523/JNEUROSCI.5712-07.2008, 考研:
18562621
Baldassano, C。, Hasson, U。, & Norman, K. A. (2018). Representation
of real-world event schemas during narrative perception.
神经科学杂志, 38, 9689–9699. https://doi.org/10
.1523/JNEUROSCI.0251-18.2018, 考研: 30249790
Bender, A。, & Beller, S. (2014). Mapping spatial frames of
reference onto time: A review of theoretical accounts and
empirical findings. 认识, 132, 342–382. https://doi.org
/10.1016/j.cognition.2014.03.016, 考研: 24873738
Boccia, M。, Nemmi, F。, & Guariglia, C. (2014). Neuropsychology
of environmental navigation in humans: Review and
meta-analysis of fMRI studies in healthy participants.
Neuropsychology Review, 24, 236–251. https://doi.org/10
.1007/s11065-014-9247-8, 考研: 24488500
Boccia, M。, Teghil, A。, & Guariglia, C. (2019). Looking into
recent and remote past: Meta-analytic evidence for cortical
re-organization of episodic autobiographical memories.
Neuroscience and Biobehavioral Reviews, 107, 84–95.
https://doi.org/10.1016/j.neubiorev.2019.09.003, 考研:
24488500
Bonnici, H. M。, Cheke, L. G。, 绿色的, D ., FitzGerald, T。, &
Simons, J. S. (2018). Specifying a causal role for angular gyrus
in autobiographical memory. 神经科学杂志, 38,
10438–10443. https://doi.org/10.1523/JNEUROSCI.1239-18
.2018, 考研: 30355636
Bromis, K., Raykov, 磷. P。, Wickens, L。, Roseboom, W., & Bird,
C. 中号. (2022). The neural representation of events is
dominated by elements that are most reliably present.
认知神经科学杂志, 34, 517–531. https://土井
.org/10.1162/jocn_a_01802, 考研: 34942648
Cabeza, R。, 王子, S. E., Daselaar, S. M。, Greenberg, D. L。,
Budde, M。, Dolcos, F。, 等人. (2004). Brain activity during
episodic retrieval of autobiographical and laboratory
事件: An fMRI study using a novel photo paradigm.
认知神经科学杂志, 16, 1583–1594.
https://doi.org/10.1162/0898929042568578, 考研:
15622612
Cabeza, R。, & St Jacques, 磷. (2007). Functional neuroimaging of
autobiographical memory. 认知科学的趋势, 11,
219–227. https://doi.org/10.1016/j.tics.2007.02.005, 考研:
17382578
康威, 中号. A。, & Pleydell-Pearce, C. 瓦. (2000). The construction
of autobiographical memories in the self-memory system.
心理评论, 107, 261–288. https://doi.org/10.1037
/0033-295x.107.2.261, 考研: 10789197
Teghil et al.
2271
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
D’Argembeau, A. (2020). Zooming in and out on one’s life:
Autobiographical representations at multiple time scales.
认知神经科学杂志, 32, 2037–2055. https://
doi.org/10.1162/jocn_a_01556, 考研: 32163320
D’Argembeau, A。, Jeunehomme, 奥。, Majerus, S。, Bastin, C。, &
Salmon, 乙. (2015). The neural basis of temporal order
processing in past and future thought. 认知杂志
神经科学, 27, 185–197. https://doi.org/10.1162/jocn_a
_00680, 考研: 24960045
Daselaar, S. M。, 米, H. J。, Greenberg, D. L。, Cabeza, R。, LaBar,
K. S。, & 鲁宾, D. C. (2008). The spatiotemporal dynamics
of autobiographical memory: Neural correlates of recall,
emotional intensity, and reliving. 大脑皮层, 18, 217–229.
https://doi.org/10.1093/cercor/bhm048, 考研: 17548799
Demblon, J。, Bahri, 中号. A。, & D’Argembeau, A. (2016). Neural
correlates of event clusters in past and future thoughts: 如何
the brain integrates specific episodes with autobiographical
知识. Neuroimage, 127, 257–266. https://doi.org/10
.1016/j.neuroimage.2015.11.062, 考研: 26658926
从, X。, 张, Z。, & Biswal, 乙. 乙. (2021). Understanding
psychophysiological interaction and its relations to beta
series correlation. Brain Imaging and Behavior, 15,
958–973. https://doi.org/10.1007/s11682-020-00304-8,
考研: 32710336
Dritschel, 乙. H。, 威廉姆斯, J. 中号. G。, Baddeley, A. D ., & Nimmo-
史密斯, 我. (1992). Autobiographical fluency: A method for
the study of personal memory. 记忆 & 认识, 20,
133–140. https://doi.org/10.3758/bf03197162, 考研:
1565011
Ekstrom, A. D ., Huffman, D. J。, & Starrett, 中号. (2017). Interacting
networks of brain regions underlie human spatial navigation:
A review and novel synthesis of the literature. 杂志
Neurophysiology, 118, 3328–3344. https://doi.org/10.1152/jn
.00531.2017, 考研: 28931613
Fan, C. L。, Abdi, H。, & 莱文, 乙. (2021). On the relationship
between trait autobiographical episodic memory and spatial
导航. 记忆 & 认识, 49, 265–275. https://doi.org
/10.3758/s13421-020-01093-7, 考研: 33051816
Faul, F。, Erdfelder, E., Lang, A. G。, & Buchner, A. (2007).
G*Power 3: A flexible statistical power analysis program for
the social, behavioral, and biomedical sciences. Behavior
Research Methods, 39, 175–191. https://doi.org/10.3758
/bf03193146, 考研: 17695343
范伯格, D. A。, Moeller, S。, 史密斯, S. M。, 奥尔巴赫, E.,
Ramanna, S。, Gunther, M。, 等人. (2010). Multiplexed echo
planar imaging for sub-second whole brain fMRI and fast
diffusion imaging. 公共图书馆一号, 5, e15710. https://doi.org/10
.1371/journal.pone.0015710, 考研: 21187930
Foudil, S. A。, Kwok, S. C。, & Macaluso, 乙. (2020). Context-
dependent coding of temporal distance between cinematic
events in the human precuneus. 神经科学杂志,
40, 2129–2138. https://doi.org/10.1523/JNEUROSCI.2296-19
.2020, 考研: 31996453
弗里斯顿, K. J. (2011). Functional and effective connectivity: A
review. Brain Connectivity, 1, 13–36. https://doi.org/10.1089
/brain.2011.0008, 考研: 22432952
Gatti, D ., Van Vugt, F。, & Vecchi, 时间. (2020). A causal role for the
cerebellum in semantic integration: A transcranial magnetic
stimulation study. Scientific Reports, 10, 18139. https://土井
.org/10.1038/s41598-020-75287-z, 考研: 33097802
Gatti, D ., Vecchi, T。, & Mazzoni, G. (2021). Cerebellum and
semantic memory: A TMS study using the DRM paradigm.
Cortex, 135, 78–91. https://doi.org/10.1016/j.cortex.2020.11
.017, 考研: 33360762
戈什, V. E., & Gilboa, A. (2014). What is a memory schema?
A historical perspective on current neuroscience literature.
Neuropsychologia, 53, 104–114. https://doi.org/10.1016/j
.neuropsychologia.2013.11.010, 考研: 24280650
Gilboa, A。, & Marlatte, H. (2017). Neurobiology of schemas and
schema-mediated memory. 认知科学的趋势, 21,
618–631. https://doi.org/10.1016/j.tics.2017.04.013, 考研:
28551107
Greenberg, D. L。, & 鲁宾, D. C. (2003). The neuropsychology
of autobiographical memory. Cortex, 39, 687–728. https://土井
.org/10.1016/s0010-9452(08)70860-8, 考研: 14584549
Grilli, 中号. D ., & Verfaellie, 中号. (2014). Personal semantic memory:
Insights from neuropsychological research on amnesia.
Neuropsychologia, 61, 56–64. https://doi.org/10.1016/j
.neuropsychologia.2014.06.012, 考研: 24949553
Grilli, 中号. D ., & Verfaellie, 中号. (2016). Experience-near but not
experience-far autobiographical facts depend on the medial
temporal lobe for retrieval: Evidence from amnesia.
Neuropsychologia, 81, 180–185. https://doi.org/10.1016/j
.neuropsychologia.2015.12.023, 考研: 26721761
赫斯, A. T。, Tisdall, 中号. D ., Andronesi, 氧. C。, Meintjes, 乙. M。, &
范德库韦, A. J. (2011). Real-time motion and B0
corrected single voxel spectroscopy using volumetric
navigators. Magnetic Resonance in Medicine, 66, 314–323.
https://doi.org/10.1002/mrm.22805, 考研: 21381101
Holland, D ., Kuperman, J. M。, & 戴尔, A. 中号. (2010). Efficient
correction of inhomogeneous static magnetic field-induced
distortion in echo planar imaging. Neuroimage, 50, 175–183.
https://doi.org/10.1016/j.neuroimage.2009.11.044, 考研:
19944768
Hsieh, L. T。, & Ranganath, C. (2015). Cortical and subcortical
contributions to sequence retrieval: Schematic coding of
temporal context in the neocortical recollection network.
Neuroimage, 121, 78–90. https://doi.org/10.1016/j
.neuroimage.2015.07.040, 考研: 26209802
爱尔兰语, M。, & Vatansever, D. (2020). Rethinking the
episodic-semantic distinction from a gradient perspective.
Current Opinion in Behavioral Sciences, 32, 43–49. https://
doi.org/10.1016/j.cobeha.2020.01.016
克莱因, S. B., & Gangi, C. 乙. (2010). The multiplicity of self:
Neuropsychological evidence and its implications for the self
as a construct in psychological research. Annals of the New
York Academy of Sciences, 1191, 1–15. https://doi.org/10
.1111/j.1749-6632.2010.05441.x, 考研: 20392272
莱文, B., Svoboda, E., 干草, J. F。, Winocur, G。, & Moscovitch,
中号. (2002). Aging and autobiographical memory: Dissociating
episodic from semantic retrieval. Psychology and Aging,
17, 677–689. https://doi.org/10.1037/0882-7974.17.4.677,
考研: 12507363
莱文, B., 车工, G. R。, Tisserand, D ., Hevenor, S. J。, Graham,
S. J。, & McIntosh, A. 右. (2004). The functional neuroanatomy
of episodic and semantic autobiographical remembering:
A prospective functional MRI study. 认知杂志
神经科学, 16, 1633–1646. https://doi.org/10.1162
/0898929042568587, 考研: 15601525
Maguire, 乙. A。, & Frith, C. D. (2003). Aging affects the
engagement of the hippocampus during autobiographical
memory retrieval. Brain, 126, 1511–1523. https://doi.org/10
.1093/brain/awg157, 考研: 12805116
Marchette, S. A。, Vass, L. K., Ryan, J。, & 爱泼斯坦, 右. A. (2014).
Anchoring the neural compass: Coding of local spatial
reference frames in human medial parietal lobe. 自然
神经科学, 17, 1598–1606. https://doi.org/10.1038/nn
.3834, 考研: 25282616
马古利斯, D. S。, 戈什, S. S。, Goulas, A。, Falkiewicz, M。,
Huntenburg, J. M。, Langs, G。, 等人. (2016). Situating the
default-mode network along a principal gradient of
macroscale cortical organization. 国家会议录
Academy of Sciences, 美国。, 113, 12574–12579. https://土井
.org/10.1073/pnas.1608282113, 考研: 27791099
Martinelli, P。, Sperduti, M。, & Piolino, 磷. (2013). Neural
substrates of the self-memory system: New insights from a
Teghil et al.
2272
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
meta-analysis. 人脑图谱, 34, 1515–1529.
https://doi.org/10.1002/hbm.22008, 考研: 22359397
McCormick, C。, Barry, D. N。, Jafarian, A。, 巴恩斯, G. R。, &
Maguire, 乙. A. (2020). vmPFC drives hippocampal processing
during autobiographical memory recall regardless of
remoteness. 大脑皮层, 30, 5972–5987. https://doi.org
/10.1093/cercor/bhaa172, 考研: 32572443
McLaren, D. G。, Ries, 中号. L。, 徐, G。, & 约翰逊, S. C. (2012). A
generalized form of context-dependent psychophysiological
互动 (gPPI): A comparison to standard approaches.
Neuroimage, 61, 1277–1286. https://doi.org/10.1016/j
.neuroimage.2012.03.068, 考研: 22484411
Miles, L. K., Nind, L. K., & Macrae, C. 氮. (2010). Moving through
时间. 心理科学, 21, 222–223. https://doi.org/10
.1177/0956797609359333, 考研: 20424050
Moeller, S。, Yacoub, E., Olman, C. A。, 奥尔巴赫, E., Strupp, J。,
哈雷尔, N。, 等人. (2010). Multiband multislice GE-EPI at 7 tesla,
with 16-fold acceleration using partial parallel imaging with
application to high spatial and temporal whole-brain fMRI.
Magnetic Resonance in Medicine, 63, 1144–1153. https://土井
.org/10.1002/mrm.22361, 考研: 20432285
Monsa, R。, Peer, M。, & Arzy, S. (2020). Processing of different
temporal scales in the human brain. 认知杂志
神经科学, 32, 2087–2102. https://doi.org/10.1162/jocn_a
_01615, 考研: 32762522
Nawa, 氮. E., & Ando, H. (2020). Effective connectivity during
autobiographical memory search. Brain and Behavior,
10, e01719. https://doi.org/10.1002/brb3.1719, 考研:
32538553
Nonyane, 乙. A。, & Theobald, C. 中号. (2007). Design sequences for
sensory studies: Achieving balance for carry-over and position
effects. British Journal of Mathematical and Statistical
心理学, 60, 339–349. https://doi.org/10.1348
/000711006X114568, 考研: 17971273
Palombo, D. J。, Sheldon, S。, & 莱文, 乙. (2018). 个人
differences in autobiographical memory. Trends in Cognitive
科学, 22, 583–597. https://doi.org/10.1016/j.tics.2018.04
.007, 考研: 29807853
Palombo, D. J。, 威廉姆斯, L. J。, Abdi, H。, & 莱文, 乙. (2013). 这
Survey of Autobiographical Memory (SAM): A novel measure
of trait mnemonics in everyday life. Cortex, 49, 1526–1540.
https://doi.org/10.1016/j.cortex.2012.08.023, 考研:
23063319
Peer, M。, Salomon, R。, Goldberg, 我。, Blanke, 奥。, & Arzy, S.
(2015). Brain system for mental orientation in space, 时间,
and person. 美国国家科学院院刊
科学, 美国。, 112, 11072–11077. https://doi.org/10.1073
/pnas.1504242112, 考研: 26283353
Petrican, R。, Palombo, D. J。, Sheldon, S。, & 莱文, 乙. (2020).
The neural dynamics of individual differences in episodic
autobiographical memory. eNeuro, 7, ENEURO.0531-
19.2020. https://doi.org/10.1523/ENEURO.0531-19.2020,
考研: 32060035
Prebble, S. C。, Addis, D. R。, & Tippett, L. J. (2013).
Autobiographical memory and sense of self. Psychological
Bulletin, 139, 815–840. https://doi.org/10.1037/a0030146,
考研: 23025923
Ranganath, C。, & Ritchey, 中号. (2012). Two cortical systems for
memory-guided behaviour. 自然评论神经科学, 13,
713–726. https://doi.org/10.1038/nrn3338, 考研: 22992647
Raykov, 磷. P。, Keidel, J. L。, Oakhill, J。, & Bird, C. 中号. (2021).
Activation of person knowledge in medial prefrontal cortex
during the encoding of new lifelike events. 大脑皮层,
31, 3494–3505. https://doi.org/10.1093/cercor/bhab027,
考研: 33866362
科学, 16, 550–558. https://doi.org/10.1016/j.tics.2012.09
.003, 考研: 23040159
Renoult, L。, Tanguay, A。, Beaudry, M。, Tavakoli, P。, Rabipour, S。,
坎贝尔, K., 等人. (2016). Personal semantics: Is it distinct
from episodic and semantic memory? An electrophysiological
study of memory for autobiographical facts and repeated
events in honor of Shlomo Bentin. Neuropsychologia, 83,
242–256. https://doi.org/10.1016/j.neuropsychologia.2015.08
.013, 考研: 26277459
鲁宾, D. C。, & Umanath, S. (2015). Event memory: A theory of
memory for laboratory, autobiographical, and fictional
事件. 心理评论, 122, 1–23. https://doi.org/10
.1037/a0037907, 考研: 25330330
Santangelo, 五、, Pedale, T。, Macrì, S。, & Campolongo, 磷. (2020).
Enhanced cortical specialization to distinguish older and
newer memories in highly superior autobiographical
记忆. Cortex, 129, 476–483. https://doi.org/10.1016/j
.cortex.2020.04.029, 考研: 32599463
Sheldon, S。, Farb, N。, Palombo, D. J。, & 莱文, 乙. (2016).
Intrinsic medial temporal lobe connectivity relates to
individual differences in episodic autobiographical
remembering. Cortex, 74, 206–216. https://doi.org/10.1016/j
.cortex.2015.11.005, 考研: 26691735
Sheldon, S。, Peters, S。, & Renoult, L. (2020). Altering access to
autobiographical episodes with prior semantic knowledge.
Consciousness and Cognition, 86, 103039. https://doi.org/10
.1016/J.CONCOG.2020.103039, 考研: 33220651
Smallwood, J。, Bernhardt, 乙. C。, 水蛭, R。, Bzdok, D ., Jefferies,
E., & 马古利斯, D. S. (2021). The default mode network in
认识: A topographical perspective. 自然评论
神经科学, 22, 503–513. https://doi.org/10.1038/s41583
-021-00474-4, 考研: 34226715
史密斯, S. M。, & Nichols, 时间. 乙. (2009). Threshold-free cluster
enhancement: Addressing problems of smoothing, 临界点
dependence and localisation in cluster inference.
Neuroimage, 44, 83–98. https://doi.org/10.1016/j.neuroimage
.2008.03.061, 考研: 18501637
Spreng, 右. N。, 三月, 右. A。, & Kim, A. S. (2009). The common
neural basis of autobiographical memory, prospection,
导航, 心理理论, and the default mode: A
quantitative meta-analysis. 认知杂志
神经科学, 21, 489–510. https://doi.org/10.1162/jocn.2008
.21029, 考研: 18510452
St Jacques, P。, 鲁宾, D. C。, LaBar, K. S。, & Cabeza, 右. (2008).
The short and long of it: Neural correlates of temporal-order
memory for autobiographical events. 认知杂志
神经科学, 20, 1327–1341. https://doi.org/10.1162/jocn
.2008.20091, 考研: 18284345
Svoboda, E., McKinnon, 中号. C。, & 莱文, 乙. (2006). 这
functional neuroanatomy of autobiographical memory: A
meta-analysis. Neuropsychologia, 44, 2189–2208. https://土井
.org/10.1016/j.neuropsychologia.2006.05.023, 考研:
16806314
Teghil, A。, Bonavita, A。, Guariglia, C。, & Boccia, 中号. (2021).
Commonalities and specificities between environmental
navigation and autobiographical memory: A synthesis and a
theoretical perspective. Neuroscience and Biobehavioral
评论, 127, 928–945. https://doi.org/10.1016/j.neubiorev
.2021.06.012, 考研: 34102149
Teghil, A。, 马克, 我. B., & Boccia, 中号. (2021). Mental
representation of autobiographical memories along the
sagittal mental timeline: Evidence from spatiotemporal
干涉. Psychonomic Bulletin & 审查, 28,
1327–1335. https://doi.org/10.3758/s13423-021-01906-z,
考研: 33782918
Renoult, L。, 戴维森, 磷. S。, Palombo, D. J。, Moscovitch, M。, &
莱文, 乙. (2012). Personal semantics: At the crossroads of
semantic and episodic memory. Trends in Cognitive
Tisdall, 中号. D ., 赫斯, A. T。, Reuter, M。, Meintjes, 乙. M。, 菲舍尔, B.,
& 范德库韦, A. J. (2012). Volumetric navigators for
prospective motion correction and selective reacquisition in
Teghil et al.
2273
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
neuroanatomical MRI. Magnetic Resonance in Medicine,
68, 389–399. https://doi.org/10.1002/mrm.23228, 考研:
22213578
Tso, 我. F。, Rutherford, S。, Fang, Y。, Angstadt, M。, & 泰勒, S. F.
(2018). The “social brain” is highly sensitive to the mere
presence of social information: An automated meta-analysis
and an independent study. 公共图书馆一号, 13, e0196503. https://
doi.org/10.1371/journal.pone.0196503, 考研: 29723244
Tulving, 乙. (1972). Chap. 10. In Organization of Memory
(PP. 381–403). 学术出版社.
Vatansever, D ., Menon, D. K., & Stamatakis, 乙. A. (2017).
Default mode contributions to automated information
加工. 美国国家科学院院刊
科学, 美国。, 114, 12821–12826. https://doi.org/10.1073
/pnas.1710521114, 考研: 29078345
Viard, A。, Chételat, G。, Lebreton, K., Desgranges, B., Landeau,
B., de La Sayette, 五、, 等人. (2011). Mental time travel into the
past and the future in healthy aged adults: An fMRI study.
Brain and Cognition, 75, 1–9. https://doi.org/10.1016/j
.bandc.2010.10.009, 考研: 21093970
Whitfield-Gabrieli, S。, & Nieto-Castanon, A. (2012). Conn:
A functional connectivity toolbox for correlated and
anticorrelated brain networks. Brain Connectivity, 2,
125–141. https://doi.org/10.1089/brain.2012.0073, 考研:
22642651
Wilson, A. E., & Ross, 中号. (2003). The identity function of
autobiographical memory: Time is on our side. 记忆,
11, 137–149. https://doi.org/10.1080/741938210, 考研:
12820827
徐, J。, Moeller, S。, 奥尔巴赫, 乙. J。, Strupp, J。, 史密斯, S. M。,
范伯格, D. A。, 等人. (2013). Evaluation of slice accelerations
using multiband echo planar imaging at 3 时间. Neuroimage, 83,
991–1001. https://doi.org/10.1016/j.neuroimage.2013.07.055,
考研: 23899722
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
3
4
1
2
2
2
5
6
2
0
5
6
4
9
0
/
/
j
哦
C
n
_
A
_
0
1
9
0
6
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Teghil et al.
2274