Responding with Restraint: What Are the
Neurocognitive Mechanisms?
Sara Jahfari1,2, Cathy M. Stinear3, Mike Claffey1,
Frederick Verbruggen4, and Adam R. Aron1
抽象的
■ An important aspect of cognitive control is the ability to re-
spond with restraint. 这里, we modeled this experimentally by
measuring the degree of response slowing that occurs when peo-
ple respond to an imperative stimulus in a context where they
might suddenly need to stop the initiated response compared
with a context in which they do not need to stop. We refer to
the RT slowing that occurs as the “response delay effect.” We con-
jectured that this response delay effect could relate to one or
more neurocognitive mechanism(s): partial response suppression
(IE。, “active braking”), prolonged decision time, and slower re-
sponse facilitation. These accounts make different predictions
about motor system excitability and brain activation. To test which
neurocognitive mechanisms underlie the response delay effect,
we performed two studies with TMS and we reanalyzed fMRI
数据. The results suggest that the response delay effect is at least
partly explained by active braking, possibly involving a mechanism
that is similar to that used to stop responses completely. 这些
results further our understanding of how people respond with re-
straint by pointing to proactive recruitment of a neurocognitive
mechanism heretofore associated with outright stopping. ■
介绍
Many situations in life call for us to respond with restraint.
Even as we satisfy an urge by making a movement, 我们
can make the movement in a controlled fashion. 对于前-
充足, one eats oneʼs food carefully rather than wolfing
it down to avoid indigestion. To take another example,
one can speak slowly and deliberately when diplomacy is
需要的, although the ideas may be fast and furious. Ex-
perimentally, this form of control may be examined by
measuring the degree of response slowing that occurs
when people respond to an imperative (去) stimulus in
a context where they might suddenly need to stop the
initiated response compared with a context in which they
do not need to stop. Several behavioral paradigms have
been used to examine this question (Verbruggen & Logan,
2009乙; Zandbelt et al., 2008; Vink et al., 2005; 德容,
Coles, & Logan, 1995). 这里, we used the “conditional
stop signal task” (De Jong et al., 1995; Figure 1A). 在这个
范例, participants initiate a choice response on each
trial and prepare to stop themselves when a stop signal
发生. One responding finger is designated as “critical”
and another as “noncritical.” When the stop signal occurs,
participants must try to stop when the initiated response
is critical, but they can ignore the stop signal when the
response is noncritical. This manipulation leads to signif-
icantly slower responses on critical compared with non-
1加州大学, 圣地亚哥, 2University of Amsterdam,
荷兰人, 3University of Auckland, 新西兰, 4Ghent
大学, 比利时
critical go trials (Aron, 贝伦斯, 史密斯, Frank, & Poldrack,
2007; De Jong et al., 1995), which we refer to here as the
“response delay effect.”
We tested three hypotheses about the neurocognitive
mechanisms underlying the response delay effect (桌子 1).
The first hypothesis is that the response slowing is ex-
plained by an active braking mechanism that can pro-
actively suppress the initiated response without canceling
it completely. Such proactive response suppression (IE。,
active braking) should be reflected in reduced excitability
of motor representations that might have to be stopped.
具体来说, the braking hypothesis predicts that if the
stopping rule says “stop only if an index finger response
has been initiated,” then the index finger motor repre-
sentation will show reduced excitability compared with
when the rule says “stop only if a little finger response
has been initiated.” Furthermore, the braking hypothe-
sis predicts that an excitability reduction of the (批判的)
response that may need to be stopped may even be ob-
served before the go stimulus occurs and not just after-
病房. This is because the stopping rules are known at the
beginning of the experiment, and thus participants may
maintain “suppression” of the critical response through-
out the whole experiment or at least in anticipation of
having to respond.
The second hypothesis for the response delay effect is
that the duration of stimulus categorization and response
selection stages is prolonged (we refer to this as the “pro-
longed decision stage” account). One source of this pro-
longation could relate to the increased cognitive load
© 2009 麻省理工学院
认知神经科学杂志 22:7, PP. 1479–1492
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
2
我
7
e
–
1
p
4
d
7
F
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
哦
7
C
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
3
0
0
7
7
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
.
t
.
/
F
.
哦
n
1
8
中号
A
y
2
0
2
1
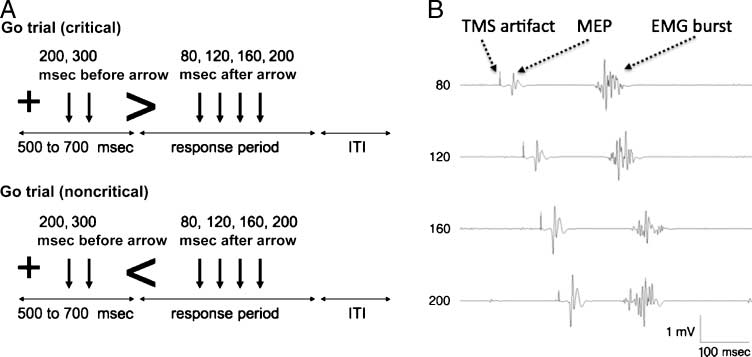
数字 1. Task and example
motor evoked potential (MEP)
Traces. (A) Schematic design
of go trials (when no stop signal
is given) in the conditional
stop task. 在这个例子中,
participants must stop if a stop
signal follows a rightward arrow
(“critical” direction). 这
horizontal lines represent
时间. The cross indicates the
beginning of each trial. 这是
followed by an arrow stimulus
indicating that a response
should be made with the right
button (little finger) or left
button (index finger). 这
vertical arrows indicate the
times of TMS delivery—those
delivered before the go stimulus
are the baseline condition. A TMS stimulus was delivered only once per trial and on some trials not at all. (乙) Representative EMG traces from one
主题. On each trial, a brief TMS artifact is visible as well as an MEP and an EMG burst (muscle activity). Note that the electromechanical delay is the
interval between the onset of EMG burst and the subsequent button press. Note that the MEP increases with time of stimulation, also see Figure 3.
that participants maintain when they expect a stop signal
to occur on critical trials. This may influence the effi-
ciency of information processing and could slow down
responses on go-critical compared with noncritical trials
(see Verbruggen & Logan, 2009乙). Another source of the
prolongation of the decision stage could relate to an in-
creased response threshold for critical trials. The response
threshold determines the amount of information that is
required to select a response; if it is increased on critical
试验, RTs will increase compared with noncritical trials
(Verbruggen & Logan, 2009乙).
The third hypothesis is that the execution of the motor
response is prolonged because of a slower build up of
facilitation in the corticomotor system rather than an ac-
tive inhibitory process. This slowing (perhaps better de-
scribed as “hesitancy” or “caution”) could be reflected in
an increased delay between the initiation of the response
and the actual button press. The major difference with
the braking hypothesis is that the facilitation hypothesis
assumes that no inhibition of motor output is involved.
The second and the third hypotheses (IE。, prolonged
decision stage and slower response facilitation) differ from
the braking hypothesis in predicting that the difference
between critical and noncritical trials will be reflected in
differences in motor excitability during the later stages of
stimulus categorization and response selection only—
whereas the braking hypothesis predicts that an excitability
reduction of the response that may need to be stopped may
be observed even before the go stimulus occurs (桌子 1).
Evaluating the predictions of these three hypotheses
requires a technique that can measure the state of spe-
cific motor representations with high temporal resolu-
的. 这里, we used TMS of the primary motor cortex,
using surface electromyography to record evoked poten-
tials from intrinsic muscles of the hand. In Experiment 1,
we delivered TMS to the left primary motor cortex at
specific time points while participants performed the
conditional stop signal task using index and little fingers
of the right hand (Figure 1A). We delivered TMS either
200 或者 300 msec before the go choice stimulus (基线),
80 和 120 msec after the go stimulus (“early”), 和 160
和 200 msec after the go stimulus (“late”). We expected
that the early time points would correspond to a pre-
response initiation period because 80 msec and possibly
桌子 1. Possible Neurocognitive Mechanisms Underlying the Response Delay Effect
Mechanism
Active braking
Motor Evoked Potential
Electromechanical Delay
Activation of
“Stopping” Regions
Go critical < go noncritical (before/after go stimulus) Go critical > go noncritical
Go critical > go noncritical
Prolonged decision stage
Go critical < go noncritical No difference No differencea (after go stimulus) Slower response facilitation Go critical < go noncritical Go critical > go noncritical
No difference
(after go stimulus)
Different mechanisms make different predictions for MEP, electromechanical delay, and functional MRI data.
aDepending which regions are activated, this could be compatible with more than one mechanism.
1480
认知神经科学杂志
体积 22, 数字 7
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
2
我
7
e
–
1
p
4
d
7
F
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
哦
7
C
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
3
0
0
7
7
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
.
t
F
/
.
.
哦
n
1
8
中号
A
y
2
0
2
1
甚至 120 msec is too early for visual information to be
categorized to determine response selection in a choice
RT task, and we expected that the later time points would
correspond with the response initiation period.
We measured motor-evoked potentials (MEPs) 从
first dorsal interosseous (FDI) muscle of the right hand—
an index of corticomotor excitability for the index finger
response representation. For each participant, the index
finger was critical for one half of the experiment and non-
critical for the other half (with the little finger in the oppo-
site pattern). This let us always record electromyography
from the index finger while comparing the effects on this
finger of the conditional rule (index finger is critical, 指数
finger is noncritical).
We predicted that participants would respond more
slowly on go-critical than go-noncritical trials—the re-
sponse delay effect. The braking hypothesis predicts that
the response delay effect would have its counterpart in
reduced MEPs at both the early preinitiation time points
(80 或者 120 毫秒) and late time points (160 或者 200 毫秒).
Showing that MEPs are reduced, relative to baseline, 在
early time points would provide support for the hypoth-
esis that participants can proactively brake a response ten-
dency even before they know which response they might
actually have to make and even before a stop signal oc-
curs. An alternative outcome is that there is no difference
in MEPs for critical and noncritical conditions before re-
sponse initiation (IE。, 在 80 或者 120 毫秒), but instead there
is a difference at the late time points only (IE。, 160 或者
200 毫秒). This would be consistent with the prolonged
decision stage and slower motor facilitation hypotheses
as well as with a modified version of the active braking
假设. This modified version predicts that an active
braking mechanism operates only when the critical re-
sponse is being initiated to restrain it in anticipation of a
possible stop. This would be like starting to stop an in-
cipient motor tendency only when that tendency has been
triggered.
Another way to distinguish between the three hypoth-
eses is to examine the electromechanical delay. This re-
fers to the interval between the onset of the EMG burst
and the button press on a particular trial. Prior research
showed an elongation of the electromechanical delay when
suppression of motor output occurred (Coxon, Stinear, &
Byblow, 2007). The braking hypothesis and the slower
motor facilitation hypothesis predict that the response
delay effect would have its counterpart in a prolonged
electromechanical delay for critical responses compared
with noncritical responses. 相比之下, the prolonged
decision stage hypothesis predicts similar electromechani-
cal delays for critical and noncritical responses because,
by definition, the electromechanical delay is postdecision.
因此, the electromechanical delay is associated with dif-
ferent predictions for the three candidate mechanisms un-
derlying the response delay effect (桌子 1).
As the reader will discover below, the results for Ex-
periment 1 are most compatible with a modified version
of the braking hypothesis rather than with a purely pro-
longed decision stage and/or slower motor facilitation ac-
计数. An additional feature of this experiment was a
general “MEP suppression” (IE。, MEPs were at below
baseline levels for responding and nonresponding fingers
at the early time points). To help interpret this finding
更好的, we performed Experiment 2, in which TMS was
delivered at the same time points during a choice RT task
as for Experiment 1, but without the presence of stop
signals. This allowed us to examine if MEP suppression
effects found in Experiment 1 might relate to the exigen-
cies of response selection itself rather than to the possi-
bility that a stop is required.
In Experiment 3, we used neuroimaging to further dis-
tinguish the hypotheses. If active braking uses a response
suppression mechanism that has something in common
with outright stopping, then brain regions that are critical
for stopping, such as the right inferior frontal gyrus, 这
pre-SMA, and the subthalamic nucleus region (reviewed in
Chambers, Garavan, & Bellgrove, 2009; Aron, Durston, 等人。,
2007), should also be active during braking (桌子 1). 我们
examined this prediction by performing a reanalysis of
previously published fMRI data acquired with the condi-
tional stop signal task (Aron, 贝伦斯, 等人。, 2007). We ex-
amined whether brain regions important for stopping are
activated more for go-critical than go-noncritical trials.
EXPERIMENT 1: TMS STUDY WITH THE
CONDITIONAL STOP SIGNAL TASK
方法
参加者
Thirteen young adults participated (five males and three
left-handed; 年龄, 米= 20 年, range = 18–24 years). 全部
participants provided written consent in accordance with in-
ternal review board guidelines of the University of California
at San Diego, completed a TMS safety screen questionnaire,
and had no contraindications to TMS. One participant did
not have reliable MEPs and was excluded from further
分析.
EMG Recordings
Participants were seated comfortably in front of an iMac
desktop computer (Apple Corporation, Cupertino, CA).
They responded with index and little fingers of the right
手, which was placed flat on the table, palm down. 这
index finger movement was a lateral abduction to the left
to depress a key whose surface was perpendicular to the
table surface. This movement maximally activated the FDI
muscle while minimizing activation of other finger mus-
克莱斯. The little finger movement was flexion downward
against a key whose surface was horizontal relative to the
table surface. This movement maximally activated the ab-
ductor digiti minimi (ADM) 肌肉 (of the “little” finger).
Surface EMG recordings were made via 10-mm-diameter
Jahfari et al.
1481
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
2
我
7
e
–
1
p
4
d
7
F
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
哦
7
C
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
3
0
0
7
7
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
.
t
.
/
.
F
哦
n
1
8
中号
A
y
2
0
2
1
Ag–AgCl hydrogel electrodes (Medical Supplies, 公司, 新的-
bury Park, CA) placed over the FDI and abductor digiti minimi
(little finger) 肌肉. A ground electrode was placed over
the lateral epicondyle of the right elbow. The EMG signal
was amplified using a Grass QP511 Quad AC Amplifier System
Grass amplifier (Grass Technologies, West Warwick, RI), 和
a band-pass filter between 30 赫兹和 1 kHz and a notch filter
在 60 赫兹. Data were sampled at 2 kHz using a CED Micro
1401 mk II acquisition system and displayed and recorded
to disk using CED Signal v4 (Cambridge Electronic Design,
剑桥, 英国). MEP analysis was performed using custom
software in Matlab R2007a (The MathWorks, Natick, 嘛).
TMS
We used a MagStim 200-2 系统 (Magstim, Whitland, 英国)
with a figure-of-eight coil (7-cm diameter) to deliver a sin-
gle test stimulus during task performance. To locate the
representation of the FDI in the left primary motor cor-
tex, the coil was initially located at a point 5 cm lateral
和 2 cm anterior of the vertex. The coil was incrementally
repositioned while administering single stimuli to locate
the position that produced the largest, reliable MEPs in
right FDI. This location was marked on a snug-fitting cap
worn by the participant to ensure the consistent place-
ment of the coil through the experiment. Resting motor
threshold was determined by finding the lowest stimulus
intensity that produced MEPs of at least 0.05 mV ampli-
tude on at least 5 的 10 试验 (Rossini et al., 1994). 下一个,
the participantʼs maximum MEP size was determined by in-
creasing stimulus intensity in 5% increments, starting at
resting motor threshold, until MEP amplitude no longer
increased with increasing stimulus intensity. Test stimulus
intensity was set to produce an MEP amplitude that was
approximately half of the participantʼs maximum MEP
振幅. This ensured that the test stimulus intensity
was on the ascending limb of the individualʼs stimulus–
response curve, so that both increases and decreases in cor-
ticomotor excitability could be detected (Devanne, Lavoie,
& Capaday, 1997).
Task and Procedure
Before TMS preparation, participants completed two prac-
tice blocks to familiarize them with the task. 参加者
subsequently performed a total of six blocks, 的 96 试验,
with each block containing 24 stop and 72 go trials (576
trials total). Before each block, instructions on the com-
puter screen indicated the critical direction for the stop
任务, which changed after three blocks. For seven partici-
pants, the left response (index finger) was the critical re-
sponse in the first three blocks, and the right response
(little finger) was the critical response in the last three
blocks. The order of the mapping rules was reversed for
the other participants. By reversing the critical rule half
way through the experiment, we could compare MEPs from
the right index finger for critical and noncritical conditions.
Instructions emphasized that participants should do
their best to respond as quickly as possible while also
doing their best to stop the response when an auditory
stop signal occurred, but only if the initiated response
was in the critical response. If the subject initiated a re-
sponse on a noncritical trial and a stop signal occurred,
the subject was to ignore the stop signal. On each trial, A
white fixation cross was displayed on a black computer
screen followed by a left- or a right-pointing arrow stimu-
字 (Figure 1A). The time between the fixation cross and
arrow stimulus ranged from 500 到 700 毫秒 (steps of
100 毫秒, 米= 600 毫秒).
In every four trials, there was one stop trial and three go
试验, and the number of leftward- and rightward-pointing
arrows was equal. The delay between the go stimulus (这
箭) and the stop signal, 那是, the stop signal delay
(SSD), was sampled from four different step-up and step-
down staircases to ensure convergence to P (inhibit) 的
50% by the end of the experiment. If a stop signal from
a particular staircase was presented for the critical direc-
tion and the subject responded, then the SSD for that
staircase was reduced by 50 msec on a subsequent stop
审判; if the subject did not respond (IE。, 成功地
stopped), then the SSD was increased by 50 毫秒. SSD
values for noncritical trials were yoked to the values for
critical trials. The four staircases started with SSD values
的 100, 150, 200, 和 250 毫秒, 分别.
In each block of 96 试验, TMS was delivered on 66 试验.
We included no-TMS trials to estimate go-critical RT, 去-
noncritical RT, and stop signal RT (SSRT) uncontaminated
by possible effects of the TMS on response emission RT
and accuracy (Ziemann, Tergau, Netz, & Hömberg, 1997;
Pascual-Leone et al., 1992). SSRT is an index of the speed
with which someone stops a motor response (见下文).
的 66 trials on which TMS was delivered, 60 were go
trials and 6 were stop trials. On stop trials, the magnetic
stimulus could only occur in the baseline (prestimulus) pe-
里约德, whereas on go trials it could occur in the baseline or
poststimulus period (Figure 1A). For stop trials, TMS was
delivered on 3 trials at 200 msec before the arrow presen-
站 (IE。, 基线, b1) 和另一个 3 trials at 300 毫秒
before the arrow presentation (IE。, b2). For go trials, TMS
was delivered on 6 trials for b1 and 6 trials for b2. Baseline
MEPs were used to normalize the poststimulus MEPs (看
analysis below). On go trials, 12 magnetic stimuli were de-
livered at each of 80, 120, 160, 和 200 msec after the go
(箭) 刺激. On all trials on which test stimuli were
delivered in the block, we balanced the number of test
stimuli for critical and noncritical directions. We note, 和
respect to the baseline stimulus, that TMS was delivered
在 6 stop trials and 12 go trials per block. 所以, 这
probability of a stop given a TMS stimulus was 0.33.
分析
Behavioral data. As TMS can speed or prolong RT (Ziemann
等人。, 1997; Pascual-Leone et al., 1992), and because the
1482
认知神经科学杂志
体积 22, 数字 7
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
2
我
7
e
–
1
p
4
d
7
F
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
哦
7
C
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
3
0
0
7
7
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
.
F
.
.
/
t
哦
n
1
8
中号
A
y
2
0
2
1
probability of stop signal was slightly different for TMS
and no-TMS trials, we separately computed key behavioral
indices on trials on which TMS was and was not present.
We calculated error rates and median RTs on critical and
noncritical go trials for the index finger and computed the
response delay effect by subtracting go-noncritical RT from
go-critical RT. We also computed the speed of stopping
for those blocks where the index finger was critical. 我们
estimated SSRT using the so-called “integration method”
(Verbruggen & Logan, 2009A; Logan & 考恩, 1984). 在
添加, based on some preliminary data (Greenhouse,
Verbruggen, & Aron, unpublished observations) we exam-
ined whether, across subjects, the response delay effect
predicted SSRT.
TMS data. All MEPs from all trials were inspected. 第一的,
trials were rejected from further analysis if there was an
overlap between the MEP and the onset of voluntary
EMG activity, or if the MEP amplitude was smaller than
0.05 mV. 第二, trials were sorted by arrow stimulus
方向 (批判的, noncritical), whether the MEP was col-
lected from the responding finger (index responding,
index not responding) and stimulation time (基线
[b1 + b2] / 2; 80, 120, 160, 和 200 毫秒). 第三, the MEPs
recorded were trimmed by removing those trials where
the MEPs were more than 3 SD from the mean under
each condition. 平均而言, 5.3% of the trials (标准差=
3.4%) were rejected for each participant. 最后, MEPs
were normalized in each condition by dividing the aver-
age MEP amplitude under a given condition by the mean
baseline MEP amplitude. The MEP data reported here are
only from the FDI muscle. Representative MEPs for this
肌肉, across the four time points, are show in Figure 1B.
Inspection of the little finger (ADM muscle) data showed
that MEPs were smaller and less reliable than those from
FDI. This was probably due to the stimulation site being
optimized for the FDI muscle. The fact that the critical
direction was switched after Block 3 meant that we could
examine FDI MEPs when the responding finger was the
index finger or when the responding finger was the little
finger (and within these conditions, when the index finger
was critical and when it was not). For the statistical analysis,
we examined FDI MEPs for the two early time points, 和
a repeated measures ANOVA that included test stimulus
间隔 (80, 120), 规则 (index finger is critical, index finger
is noncritical), and responding finger (index responding, 在-
dex not responding). We also examined FDI MEPs for the
two later time points with a repeated measures ANOVA,
which included test stimulus interval (160, 200), critical di-
反应 (批判的, noncritical), and responding finger (指数
responding, index not responding).
We also computed root mean square EMG activity in
这 100 msec preceding the TMS stimuli for each condi-
tion to establish if the muscle of interest (FDI) was “quiet”
at the time of MEP recording and to establish if there were
systematic pre-TMS differences in muscle activity for con-
ditions of interest. 最后, we also computed the electro-
mechanical delay for go-critical and noncritical trials. EMG
burst onset was determined as follows. For each trial, 这
standard deviation of the EMG trace was established for
a “quiet” period of the trial. A threshold was computed,
which was three times the magnitude of the standard de-
viation of the quiet period. An algorithm then determined
when it was during the 150 msec before the button press
that the electromyograph rose above the threshold. A
researcher, blind to condition, then reviewed (and ad-
justed if necessary) the estimated EMG burst onset. 这
electromechanical delay refers to the interval between
the onset of the EMG burst and the button press on a par-
ticular trial. A recent study showed that an elongation
of electromechanical delay can be produced by a stop-
ping process (Coxon et al., 2007). 具体来说, 这些
authors demonstrated a significantly increased electro-
mechanical delay in a responding muscle when an alterna-
tive muscle was stopped, relative to when the alternative
muscle did not need to be stopped. Elongation of the elec-
tromechanical delay for go-critical trials could relate to
active braking via suppression of motor output (Coxon
等人。, 2007).
结果
Behavior
桌子 2 shows behavioral data for all trials, in both the
TMS and the no-TMS conditions. Although TMS is some-
times shown to influence RT (例如, Ziemann et al., 1997;
Pascual-Leone et al., 1992), here we found minimal differ-
ences between TMS and no-TMS trials: critical RT, t(11) =
0.5, p = .6; noncritical RT, t(11) = 0.7, p = .5. 这
was probably because the TMS stimuli were delivered
200 msec or more before the motor response. 所以,
we report statistical results for all trials. Three participants
were left-handed, but an analysis of MEP amplitude
showed that handedness was not a factor, F(1, 10) =
0.27, p = .61; 所以, 全部 12 participants were analyzed
一起.
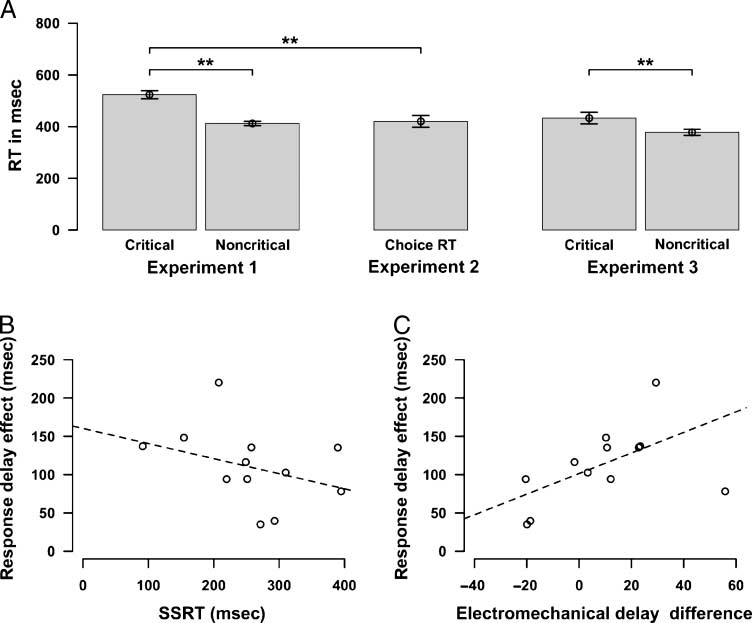
There was a reliable response delay effect as partici-
pants responded significantly more slowly on critical than
noncritical trials, t(11) = 7.7, p < .001 (Figure 2A). Par-
ticipants made very few errors of omission or discrim-
ination on go trials (combined errors, M = 1.1%, SD =
1.2%). The number of combined errors was very similar
for go-critical and noncritical trials (go-critical trial, M =
0.7, SD = 0.8; go-noncritical trial, M = 0.4, SD = 0.9),
t(11) = 0.9, p = .4. The average SSD at which stop sig-
nals was delivered was 228.6 msec (SD = 78.7 msec). SSRT
was estimated at 257.5 msec (SD = 86.8 msec). Impor-
tantly, we found that across participants, the response
delay effect was correlated with SSRT; i.e., participants
who showed a larger response delay effect stopped more
quickly (faster SSRT), t(10) = 2.08, p < .05, robust regres-
sion (Figure 2B).
Jahfari et al.
1483
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
7
e
-
1
p
4
d
7
f
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
o
7
c
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
0
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
f
t
/
.
o
n
1
8
M
a
y
2
0
2
1
Table 2. Behavioral Data for TMS Experiments 1 and 2
Corticomotor Excitability
TMS
No TMS
All Trials
Experiment 1
Clinical trials
Median RT (msec) 522.0 (55.9) 531.9 (73.9) 523.7 (53.9)
Errors (%)
1.1 (1.2)
0.8 (1.5)
0.7 (0.8)
SSRT (msec)
Mean SSD (msec)
Noncritical trials
257.5 (86.8)
228.6 (78.7)
Median RT (msec) 411.8 (29.4) 416.2 (37.7) 412.3 (29.5)
Errors (%)
0.9 (1.9)
0.0 (0.0)
0.4 (0.9)
Overall
Response delay
effect (msec)
Experiment 2
110.2 (52.0) 115.6 (66.9) 111.3 (50.3)
Median RT (msec) 418.9 (63.5) 428.1 (65.3) 420.2 (63.9)
Errors (%)
0.7 (0.5)
1.4 (1.5)
0.8 (0.5)
Critical and noncritical RTs refer to trials without stop signals. Values in
parentheses are SDs.
Mean resting motor threshold was 42.3% (SD = 3.7%),
mean test stimulus intensity was 50.4% (SD = 4.4%),
and mean baseline MEP amplitude in FDI was 1.25 mV
(SD = 0.35 mV). The main effects of time point (80,
120, 160, or 200 msec) and rule (index finger is critical,
index finger is noncritical) and the interaction between
the two were significant (all ps < .05). However, our
starting hypotheses make different predictions for early
and late time points. Therefore, in the following analyses,
we will analyze the data for early and late time points sep-
arately. To test the braking hypothesis, specifically the
idea that participants might proactively suppress the “crit-
ical” response representation even before the response
is initiated, we examined FDI MEPs for the two early time
points. We used a repeated measures ANOVA, which
included test stimulus interval (80 and 120 msec), rule
(index finger is critical, index finger is noncritical), and
responding finger (index responding, index not respond-
ing). There was a main effect of interval—FDI MEPs de-
creased significantly from 80 to 120 msec, F(1, 11) = 6.4,
p < .05 (Figure 3A). There were no further main effects or
interactions. Inspection of the pattern in Figure 3A shows
that FDI MEP amplitude was reduced below baseline at
the 80- and 120-msec time points. To formally test this
“MEP suppression,” we collapsed the normalized MEP data
across responding finger and across rule condition and
Figure 2. Behavioral and
electromechanical delay results.
(A) Median RT for the three
experiments. In Experiments 1
and 2, the values are for
index finger responses. In
Experiment 3, critical and
noncritical were index or
middle fingers for subjects
(counterbalanced across
subjects). Error bars are SEM.
These are all trials without
stop signals. (B) Negative
correlation between the
response delay effect (i.e.,
RT difference between
go-critical and noncritical trials)
and the SSRT. (C) Positive
correlation between the
response delay effect and
the electromechanical delay
difference between go-critical
and noncritical trials. Regression
lines and p values were
computed with the use of
robust regression by iteratively
reweighted least squares to
prevent the influence of
outliers.
1484
Journal of Cognitive Neuroscience
Volume 22, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
7
e
-
1
p
4
d
7
f
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
o
7
c
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
0
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
t
.
.
f
.
o
n
1
8
M
a
y
2
0
2
1
We performed a pre-TMS EMG validation to make sure
that the FDI was equivalently “quiet” across conditions. We
analyzed these data with a repeated measures ANOVA, in-
cluding test stimulus interval (80, 120, 160, and 200 msec),
rule (current response is critical, noncritical), and respond-
ing finger (index responding, index not responding), with
root mean square electromyography as the dependent var-
iable. There were no significant main effects or interactions.
Overall, the FDI muscle was “at rest” before magnetic stim-
ulation (root mean square, M = 0.6 μV, SD = 0.1 μV).
Finally, we found a nonsignificant trend for the electro-
mechanical delay to be longer for go-critical than noncritical
trials (critical: M = 135.5 msec, SD = 29.7 msec; noncriti-
cal: M = 128.6 msec, SD = 15.1 msec; p = .090 Wilcoxon
test, one-tailed). Interestingly, the electromechanical delay
difference between these trials types was strongly corre-
lated with the response delay effect, t(10) = 8.4, p <
.001, robust regression (Figure 2C).
A key result of this TMS experiment was that the differ-
ence in the excitability of the FDI representation, be-
tween critical and noncritical conditions, only emerges
at the 160- and 200-msec time points. This was contrary
to the prediction of the braking hypothesis that a differ-
ence between these conditions may be observable before
the response is initiated by primary motor cortex (Table 1).
The finding of a later difference in excitability is consistent
with the three possible accounts: a modified active braking
account in which the braking mechanism operates when
the critical response is initiated as well as the prolonged
decision stage and slower motor facilitation accounts. How-
ever, other aspects of the data speak against these latter
two accounts as explaining all of the response delay effect.
First, there was a significant correlation between the re-
sponse delay effect and the SSRT—those participants with
a longer response delay effect stopped more quickly. This
suggests that a process related to the increased slowing
could also be related to the faster stopping. It is unlikely
that slower motor facilitation would explain this because
slower facilitation should not alter the speed of the stop-
ping mechanism, as going and stopping are thought to be
independent (Verbruggen & Logan, 2009a). In the current
experiment, the SSD was adjusted dynamically—so that if
a subject facilitated their motor response more slowly,
then the SSD would be adjusted for that and the SSRT es-
timate would not be influenced. Therefore, the response
delay effect/SSRT correlation speaks against the slower
motor facilitation account; instead, the correlation be-
tween response delay effect and SSRT could be explained
by either active braking or prolonged decision stage ac-
counts. If active braking operates via partial response sup-
pression, then starting this process in advance of the stop
signal would produce a response delay effect and would
also enable faster stopping. Similarly, if a prolonged deci-
sion stage relates to increased cognitive load on go-critical
trials (i.e., monitoring both a stop goal and a go goal or
paying more attention to the occurrence of a stop signal),
then this could produce a response delay effect as well as
Jahfari et al.
1485
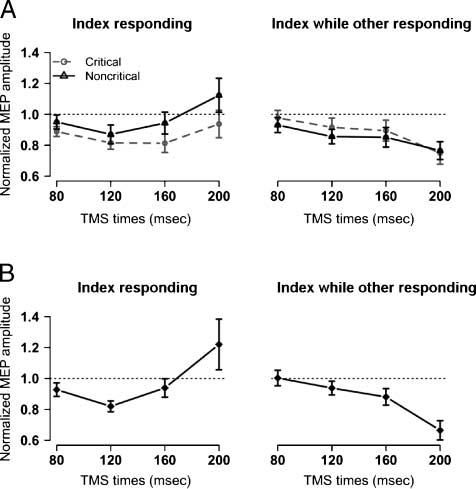
Figure 3. Corticomotor excitability for TMS Experiments 1 and 2.
Normalized mean MEP amplitudes are shown for the FDI (index finger)
muscle after stimulus presentation. These were computed by dividing
the MEPs for each subject in each condition by the baseline MEPs
for that subject. The dotted line represents the size of the MEP
amplitude at baseline. (A) Experiment 1: conditional stop signal task.
(B) Experiment 2: choice RT task (no stop signals). The left side of
each panel shows MEPs from the FDI muscle when the stimulus
indicates a response with the index finger; the right side shows
mean MEP amplitudes, from the FDI muscle, when the stimulus
indicates a response with the little finger (ADM).
tested whether the MEP was different from “1.” The MEP
was significantly suppressed at 80 msec, t(11) = −1.9, p <
.05, one-tailed, and at 120 msec, t(11) = −3.0, p < .01,
one-tailed.
Second, we analyzed FDI MEPs for the two late time
points. As can be seen in Figure 3A, the MEP data show
that a response initiation stage is evident at 160 msec and
later. We performed a repeated measures ANOVA with
test stimulus interval (160 and 200 msec), rule (index fin-
ger is critical, index finger is noncritical), and responding
finger (index responding, index not responding). There was
a main effect of rule—FDI MEP amplitudes were smaller
in the critical than noncritical condition, F(1, 11) = 5.6,
p < .05, and an interaction between rule and test stimulus
interval, F(1, 11) = 6.5, p < .05—indicating that the excit-
ability rose more slowly under the critical condition. More-
over, there was a main effect of responding finger—FDI
MEP amplitudes were greater when the index was the re-
sponding finger than when it was not, F(1, 11) = 8.0,
p < .05, and there was an interaction between responding
finger and time, F(1, 11) = 13.9, p < .05—such that the
difference in excitability between responding and non-
responding fingers was larger for the 200-msec time point
than for the 160-msec time point.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
7
e
-
1
p
4
d
7
f
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
o
7
c
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
0
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
/
t
f
.
o
n
1
8
M
a
y
2
0
2
1
faster stopping (because the stop goal is already active or
because the stop signal is detected more quickly). Thus,
the significant correlation between the response delay ef-
fect and SSRT is inconsistent with the slower motor fa-
cilitation account but is consistent with the prolonged
decision stage account and the braking account (Table 1).
Second, there was a significant correlation between the
response delay effect and the electromechanical delay
difference for go-critical versus noncritical trials. This is
compatible with both active braking and slower motor
facilitation accounts, but not with a prolonged decision
stage account. The electromechanical delay reflects the
delay between the EMG burst and the button press and
thus corresponds to a stage after a decision about the
response has been made, and this could reflect active
braking. For example, Coxon et al. (2007) demonstrated
a significantly increased electromechanical delay in a re-
sponding muscle when an alternative muscle was stopped,
relative to when the alternative muscle did not need to
be stopped. Thus, elongation of the electromechanical
delay for go-critical versus noncritical trials could relate
to active braking via suppression of motor output. How-
ever, such elongation could also relate to slower motor
facilitation. Thus, the significant correlation between the
response delay effect and the electromechanical delay dif-
ference is inconsistent with the prolonged decision stage
account but is consistent with the slower motor facilita-
tion account and the braking account (Table 1).
We note that the idea that subjects slow go-critical re-
sponses to increase the probability of successful stopping
resembles the idea that subjects slow responses to in-
crease the probability of a correct response in a choice task
(i.e., the speed/accuracy trade-off; Rinkenauer, Osman, Ulrich,
Muller-Gethmann, & Mattes, 2004; Howell & Kreidler, 1963).
In this experiment, we did not observe increased accuracy
on go-critical than noncritical trials (but see Verbruggen
& Logan, 2009b); however, this may have related to a very
slow error rate overall. We assume that the similarity be-
tween slowing in anticipation of a stop signal and slowing
to prevent an erroneous response exists because in both
situations, subjects prolong decision and nondecisional
(motor-related) stages to prevent fast responses.
An interesting but unexpected finding from Experi-
ment 1 was a significant suppression of FDI MEPs com-
pared with baseline at the 120-msec time point, both
when the index was the responding and the nonrespond-
ing finger. Possibly, participants suppress all motor output
when a stimulus is detected (or even before it is detected)
in the conditional stop task and maintain suppression for
critical trials and release suppression for noncritical trials
after stimulus categorization and response selection. Al-
ternatively, the general suppression could be due to the
exigencies of response selection and thus would be unre-
lated to the requirement to stop occasionally. To examine
this question further, we performed a second experiment
in which we again used TMS to probe motor cortex excit-
ability during a choice RT task, but this time without stop
signals. These were different participants who knew noth-
ing about the requirement to stop in Experiment 1. Our
objective here was to assess whether the general suppres-
sion at the early time points was due to the requirement to
stop occasionally or whether it was due to response selec-
tion itself.
EXPERIMENT 2: TMS STUDY WITH A CHOICE
RT TASK
Methods
Participants
Eight young adults participated (four males, all right-
handed; age M = 21.8 years, range = 19–34 years). All
participants provided written consent in accordance with
the internal review board guidelines of the University of
California at San Diego, completed a TMS safety screen
questionnaire, and had no contraindications to TMS. These
were different participants from Experiment 1.
EMG Recordings and TMS
The same methods were used as in Experiment 1 above.
Task and Procedure
Experiment 2 comprised a total of six blocks, with each
containing 72 trials. Apart from the number of trials per
block and the absence of stop instructions or stop sig-
nals, every aspect of task procedure and TMS recording
was the same as for Experiment 1 (Figure 1A). Partici-
pants were instructed to respond as fast as possible while
maintaining accuracy, with a left or a right keypress after
the arrow was presented (again using index and little fin-
gers of the right hand). Of the 72 trials in each block, mag-
netic stimuli were delivered before the arrow on 12 trials
(baseline) and after the arrow on 48 trials. There were
12 trials with no magnetic stimuli. As in Experiment 1, 6
magnetic stimuli were given at 200 msec (b1) and 6 at
300 msec (b2) before the arrow presentation to record
baseline MEPs. After the go stimulus, 48 magnetic stimuli
were given at the same four time intervals as in Experi-
ment 1. These stimuli were equally distributed over the
four time points for each arrow direction.
Analysis
Behavioral data. Median RT and percentage omission/
discrimination errors were computed for the index finger.
TMS data. Trials were rejected using the same criteria
as for Experiment 1. Trials were then sorted by respond-
ing finger (index responding, index not responding) and
TMS stimulus time (baseline [b1 + b2] / 2; 80, 120, 160,
and 200 msec). Data were trimmed and normalized as for
1486
Journal of Cognitive Neuroscience
Volume 22, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
7
e
-
1
p
4
d
7
f
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
o
7
c
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
0
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
f
t
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Experiment 1. On average 7.0% of all the trials per subject
were rejected (SD = 3.7%). As for Experiment 1, we per-
formed separate ANOVAs for the early and late time
points. In addition, we validated our results by computing
pre-TMS electromyography in the 100 msec preceding the
magnetic stimuli for each condition. The pretrigger elec-
tromyography was analyzed as for Experiment 1.
Results
Behavior
A comparison of RT between TMS (M = 418.9, SD =
63.5) and no-TMS trials (M = 428.1, SD = 65.3) revealed
no significant differences, t(7) = −1.8, p = .1; thus, we
collapsed RT over all trials. We found that median choice
RT in this experiment was significantly faster than critical
go RT in Experiment 1 (Experiment 1: M = 523.7, SD =
53.9; Experiment 2: M = 420.2, SD = 63.9), t(18) = 3.9,
p < .01, independent samples t test, but not significantly
different from noncritical go RT in Experiment 1 (Experi-
ment 1: M = 412.3, SD = 29.5; Experiment 2: M = 420.2,
SD = 63.9), t(18) = −0.4, p = .7 (Figure 2A). Thus, par-
ticipants responded to “pure go” trials in this experiment
with a similar latency to noncritical go trials in Experi-
ment 1. Again, omission/discrimination errors on go trials
were few (M = 0.8%, SD = 0.5%).
Corticomotor Excitability
Mean resting motor threshold was 41.2% (SD = 5.8%), mean
test stimulus intensity was 46.9% (SD = 7.5%), and mean
baseline FDI MEP amplitude was 1.1 mV (SD = 0.3 mV).
Overall, the pattern of results from Experiment 2 closely
resembled the findings from Experiment 1 (compare Fig-
ure 3A with Figure 3B). For the ANOVA for the early time
points (80 and 120 msec), there was a main effect of time,
F(1, 7) = 10.6, p < .05—more MEP suppression at 120
than 80 msec, and a main effect of finger, F(1, 7) = 17.4,
p < .01—excitability was less when the FDI was the re-
sponding finger than when it was not.
A key planned analysis was to examine whether signif-
icant below-baseline MEP suppression occurred in this
experiment. Collapsing across responding finger, we exam-
ined whether normalized MEP amplitudes were different
from “1.” As in Experiment 1, there was a significant “MEP
suppression” at 120 msec, t(7) = −3.55, p < .01, one-tailed.
Yet, this effect was not observed at 80 msec, t(7) = −0.81,
p = .44, one-tailed.
For the ANOVA for the late time points (160 and 200 msec),
there was a main effect of finger, F(1, 7) = 7.52, p < .05—
where FDI MEP amplitude was greater when it was respond-
ing than nonresponding, and an interaction between re-
sponding finger and a main effect of time, F(1, 7) = 7.2,
p < .05—the facilitation of FDI MEPs increased from 160
to 200 msec to a greater extent when the finger was re-
sponding than when it was not.
For the validation analysis of pre-TMS electromyogra-
phy, ANOVA was performed with test stimulus interval
(80, 120, 160, and 200 msec) and responding finger (in-
dex responding, index not responding). There were no
significant main effects or interactions. Overall, the FDI
muscle was “at rest” before the magnetic stimulation (M =
0.9 μV, SD = 0.4 μV).
The results of Experiment 2 suggest that the general
suppression at 120 msec in Experiment 1 was due to
the requirement to select responses and not to the re-
quirement to stop occasionally. Based on prior research
(Duque & Ivry, 2009; Boulinguez, Jaffard, Granjon, &
Benraiss, 2008; Davranche et al., 2007; Hasbroucq et al.,
1999; Hasbroucq, Kaneko, Akamatsu, & Possamai, 1997)
and as we argue in the General discussion section, it is
likely that this MEP suppression is due to the imposition
of an inhibitory process when selecting response. This
process of inhibiting the corticospinal pathway could be-
gin at the fixation period of the trial or some time before
stimulus onset, perhaps to prevent premature responses
(as the above authors have argued), or it could be ap-
plied around the time of response initiation itself, con-
sistent with neurophysiological models proposing that
response initiation is preceded by suppression of com-
petitor motor programs (Mink, 1996). As we only included
a baseline at 200 or 300 msec pre-go-stimulus, we cannot
judge when the MEP suppression began.
We now return to the question of the neurocognitive
mechanisms underlying the response delay effect, ob-
served in Experiment 1. We noted that this behavioral
effect could be explained by a prolonged decision stage
as well as slower motor facilitation, but it was most com-
patible with a modified version of active braking. Another
way to elucidate between these accounts is to use func-
tional MRI to examine activation for go-critical versus
noncritical trials. If active braking occurs via a (partial)
stopping mechanism, then brain regions that are key for
outright stopping may be activated more for go-critical
than noncritical trials (Table 1).
EXPERIMENT 3: REANALYSIS OF fMRI DATA
FROM THE CONDITIONAL STOP SIGNAL TASK
Methods
Participants
Fifteen right-handed young adults participated in the fMRI
study (10 males; age, M = 28.1 years). All were free of
neurological or psychiatric history and gave informed con-
sent according to the institutional review board protocol
of the University of California at Los Angeles.
Task and Procedure
The conditional stop signal task was highly similar to the
one used in Experiment 1. Full details are provided by Aron,
Behrens, et al. (2007). In brief, for the go task, participants
Jahfari et al.
1487
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
7
e
-
1
p
4
d
7
f
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
o
7
c
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
0
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
.
/
.
.
f
o
n
1
8
M
a
y
2
0
2
1
responded as fast as possible with a left or right keypress
(using index and middle fingers of the right-hand) to arrows
pointing left or right. For the stop task (25% of trials), partic-
ipants attempted to stop the response when a stop signal
was sounded after a particular SSD, but only if the arrow
was pointing in the critical direction: for half the participants,
this was leftward pointing; for the other half, rightward
pointing. There were 32 stop trials and 96 go trials per scan
(128 trials total). Each subject performed three scans. In
every 4 trials, there was 1 stop trial and 3 go trials, and
the number of leftward and rightward pointing arrows
was equal. The SSD value for the stop trial was sampled
from one of the four staircases in turn. Null events were in-
terposed between every stop or go trial. The duration of
null time ranged between 0.5 and 4 sec (M = 1 sec).
Behavioral Data Analysis
This was similar to Experiment 1.
fMRI Acquisition and Processing
Full details are provided by Aron, Behrens, et al. (2007).
In brief, images were acquired using a 3-T Siemens Allegra
MRI scanner at the Ahmanson-Lovelace Brain Mapping
Center at the University of California at Los Angeles. Each
scanning run acquired 166 functional T2*-weighted echo-
planar images (4 mm slice thickness, 33 slices, repetition
time = 2 sec, echo time = 30 msec, flip angle = 90°, ma-
trix 64 × 64, field of view = 200, in-plane resolution =
3.125 mm). The first two volumes in each run were dis-
carded to allow for T1 equilibrium effects. In addition, a
high-resolution structural scan (MP-RAGE) was acquired
for registration: acquisition parameters, repetition time =
2.3, echo time = 2.1, field of view = 256, matrix = 192 ×
192, sagittal plane, slice thickness = 1 mm, 160 slices. Data
were preprocessed using the FMRIB software library (www.
fmrib.ox.ac.uk/fsl), including realignment, spatial smooth-
ing, temporal filtering, and registration steps.
fMRI Model Fitting
Three different models were fit for the analysis here, with
slightly different regressors. These were as follows: (a) the
basic model, which included go-critical, go-noncritical, suc-
cessful stop, and unsuccessful stop trials and a nuisance
event consisting of go trials on which participants did
not respond or made errors of discrimination (see Aron,
Behrens, et al., 2007); (b) the basic model with binned
RT, which was the same as the basic model except that sep-
arate regressors were created for fast, medium, and slow
RT in go-critical and go-noncritical conditions (in each con-
dition, for each scan, the go RTs were split into three
roughly equally sized bins); and (c) the parametric model,
which was the same as the basic model but with two extra
regressors, go-critical parametric and go-noncritical para-
metric, which added RT as a covariate for each trial type.
fMRI Statistical Analysis
We performed three kinds of analyses. First, an anatomi-
cally defined ROI approach to test whether regions of the
brain known to be critical for stopping would be activated
more for go-critical than noncritical and especially whether
this would interact with RT. Our ROIs were the right in-
ferior frontal gyrus, the pre-SMA, and the subthalamic nu-
cleus region based on a prior study (Aron, Behrens, et al.,
2007) and on other literature pointing to these as critical
“nodes” for behavioral stopping (reviewed in Chambers
et al., 2009; Aron, Durston, et al., 2007). Using the results
from the basic model with binned RT, we extracted the
mean activity for each subject for each level of the rule
factor (current response is critical, noncritical) and the
RT factor (fast, medium, and slow) and performed ANOVA
for each of the three ROIs. Second, we performed whole-
brain voxel-based analysis using the basic model. For each
subject and for each of three scans, we computed the
contrast: go critical–go noncritical. Third, we performed
whole-brain voxel-based analysis using the parametric mod-
el. For each subject and for each of three scans, we com-
puted the contrast: go-critical parametric–go-noncritical
parametric. Analysis was carried out using the fMRI Expert
Analysis Tool Version 5.1, part of the FMRIBʼs Software Li-
brary (www.fmrib.ox.ac.uk/fsl). Higher level analysis (one-
sample t test) was carried out using ordinary least squares
simple mixed effects. For the whole-brain analyses, z statis-
tic images were thresholded using clusters determined by
z > 2.3 and a (corrected) cluster threshold of p = .05
(using Gaussian random field theory).
结果
Behavior
As reported by Aron, 贝伦斯, 等人. (2007), 参与者
responded significantly more slowly on go-critical than
noncritical trials (median go-critical RT = 433 毫秒, 标准差=
85 毫秒; median go-noncritical RT = 378 毫秒, 标准差=
44 毫秒), t(14) = 5.7, p < .0001. SSRT was estimated at
266 msec (SD = 53 msec). As for Experiment 1, the SSRT
and the response delay effect were negatively correlated
(r = −.26) so that subjects who responded more slowly
on critical than noncritical trials also stopped more
quickly, but this was not statistically significant here.
fMRI
Go-critical versus noncritical activation for three ROIs.
For the right inferior frontal gyrus, there was a significant
main effect of rule (activation was greater when current
response was critical than noncritical), F(1, 14) = 17.5,
1488
Journal of Cognitive Neuroscience
Volume 22, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
7
e
-
1
p
4
d
7
f
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
o
7
c
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
3
0
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
.
.
f
.
t
o
n
1
8
M
a
y
2
0
2
1
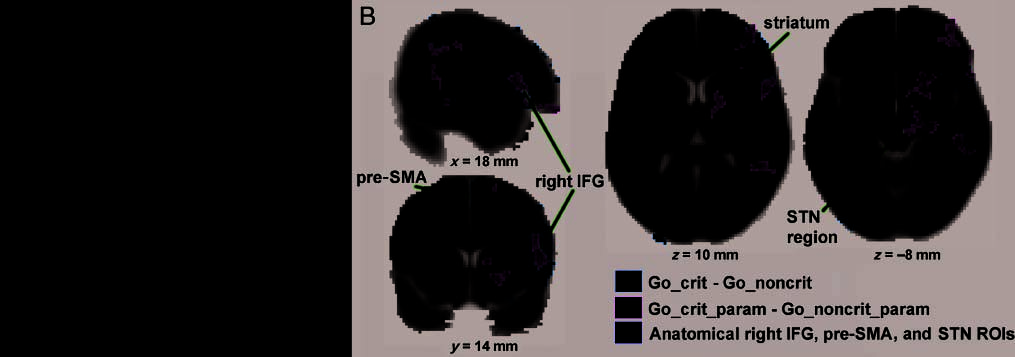
p < .001, and a significant interaction between rule and
RT (the activation difference between go-critical and non-
critical was greatest for the slower RTs), F(2, 28) = 3.54,
p < .05 (Figure 4A). For the pre-SMA, there was a signif-
icant main effect of rule (go-critical greater than noncriti-
cal), F(1, 14) = 10.4, p < .01, but no interaction between
rule and RT. For the subthalamic nucleus region, there
were no significant effects. As an auxiliary analysis, in light
of the findings of Experiment 2 that an inhibitory process
may be recruited as part of response selection on go trials
(even without the potential need to stop), we examined
whether go-noncritical activation was above baseline. Sig-
nificant activation was not present in any of the ROIs (all
ps > .26), nor was there activation at the voxel level, 小的
volume correction for multiple comparisons in the ana-
tomically defined right inferior frontal gyrus, pre-SMA,
and subthalamic nucleus regions.
Whole-brain voxel-based analysis. Go-critical trials acti-
vated a large focus of right lateral pFC significantly more
than go-noncritical trials (z > 2.3, whole-brain cluster cor-
rected). 重要的, this included the right inferior frontal
gyrus (max Z = 4.43, [52 20 4]; Figure 4B), overlapping
with regions of the inferior frontal gyrus that we have
previously shown to be activated by outright stopping in
these same subjects (Aron, 贝伦斯, 等人。, 2007). 那里
was also significant activation of the right pre-SMA (max
Z = 3.45, [10 6 72]), as well as the right superior parietal
cortex (max Z = 4.49, [42 −44 48]) and the right mid-
dle temporal gyrus (max Z = 4.04, [58 −26 −2]). At a
whole-brain-corrected threshold, no subcortical activation
was evident. For the parametric contrast, a significantly
stronger relationship between activation and RT was ob-
served for go-critical than for noncritical trials in several
right hemisphere regions including the inferior frontal
gyrus and the DLPF cortex (max Z = 3.68, [50 16 24]),
the striatum (max Z = 3.53, [14 14 6]), and the parietal
cortex (max Z = 3.4, [60 −46 30]) (all z > 2.3, whole-brain
corrected; Figure 4B). There was also activation in the mid-
brain subthalamic nucleus region; 然而, without using
high-resolution methods, it is difficult to locate this with
confidence.
GENERAL DISCUSSION
We examined the neurocognitive mechanisms that under-
lie the response delay effect that is observed when peo-
ple anticipate they might need to stop. In a TMS study,
we found that corticomotor excitability was lower and in-
creased more slowly for go-critical versus noncritical trials
从大约 160 毫秒. We also found that those partici-
pants with a larger response delay effect were able to
stop their responses more quickly and that those with
a larger response delay effect had a bigger difference in
electromechanical delay for go-critical than for noncritical
试验. In a reanalysis of fMRI data, we found that a pre-
frontal region (the right inferior frontal gyrus) that is nec-
essary to outright suppress a motor response was more
activated for go-critical trial versus noncritical trial, 更多的-
over in proportion to the degree of RT slowing.
One explanation for the response delay effect is an active
braking mechanism. This could involve partial response
suppression, perhaps using the same brain mechanism that
is used to stop a response outright. 因此, the partic-
ipant may prepare to partially inhibit a particular effector
according to the critical rule, and this could potentially
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
2
我
7
e
–
1
p
4
d
7
F
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
哦
7
C
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
3
0
0
7
7
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
.
F
.
.
t
/
哦
n
1
8
中号
A
y
2
0
2
1
数字 4. Responding with restraint activates brain regions important for outright stopping. (A) Activation of the right inferior frontal gyrus (IFG),
anatomically defined, increases with slower RT in the go-critical but not noncritical condition. Mean activation is extracted from the ROI for
each participant for each trial type (in critical and noncritical conditions and at three levels of RT). (乙) Whole-brain analyses at the voxel level.
Go-critical trials activate several brain regions more than noncritial trials, including the right inferior frontal gyrus (IFG) and the pre-SMA (n = 15,
voxel threshold z > 2.3, whole-brain cluster corrected). 而且, the correlation between activation and RT is significantly greater for go-critical
than noncritical trials in several right hemisphere regions including the right IFG, the striatum, the parietal cortex, and a midbrain region in
the vicinity of the subthalamic nucleus (n = 15, voxel threshold z > 2.3, whole-brain cluster corrected).
Jahfari et al.
1489
occur even before the go stimulus is displayed. 然而,
the results were inconsistent with this prediction: 那里
was no effect of rule (critical versus noncritical) at the early
time points of 80 和 120 毫秒. 反而, 我们发现
the difference between these conditions emerged at the
later time points of 160 和 200 毫秒 (IE。, after stimulus
categorization and response selection and during response
initiation). Although this late effect could be explained by
an active braking mechanism that restrains the critical re-
sponse once it is initiated, it can also be partly explained
by a prolonged decision stage account and by slower mo-
tor facilitation (桌子 1). We argued that the correlation
between the response delay effect and the SSRT speaks
against the slower motor facilitation account, whereas the
correlation between the response delay effect and the
electromechanical delay difference speaks against the pro-
longed decision stage account. 因此, the data from Ex-
periment 1 point most clearly toward a modified version
of active braking as a mechanism to explain the response
delay effect. Notwithstanding this, it is likely that the dif-
ferent underlying mechanisms are all in play in different
participants to differing degrees. In an earlier study that ex-
amined a different form of response slowing in the context
of stop signals, mathematical modeling showed that the
slowing was accounted for by variance in both decision
and nondecision (response initiation) 阶段 (Verbruggen
& Logan, 2009乙).
Active braking could occur through a partially activated
stopping mechanism. This predicts that brain regions crit-
ical for outright stopping will be activated in relation to
the response delay effect. In Experiment 3, 我们发现
this was indeed the case. For the anatomically defined
ROI analysis, we observed significantly greater activation
for go-critical than noncritical trials in the right inferior
frontal gyrus and the pre-SMA (two regions that are criti-
cal for outright stopping, see Chambers et al., 2009), 和
we observed that activation increased with increasing RT
more for go-critical than noncritical trials in the right in-
ferior frontal gyrus. The whole-brain voxel-level analysis
confirmed these observations with greater spatial resolu-
的. 尤其, for the parametric analysis, a signifi-
cantly stronger relationship between activation and RT
was observed for go-critical than for noncritical trials in
the right inferior frontal gyrus. 然而, additional re-
gions of the right hemisphere were also implicated, 在-
cluding the DLPF cortex, the striatum, and the parietal
cortex as well as a midbrain region.
This pattern of fMRI data is compatible with both ac-
tive braking and prolonged decision stage accounts. 在
go-critical trials, participants have to maintain the goal to
stop and the goal to go at the same time, whereas on go-
noncritical trials, they only have a goal to go. The increased
load, which partly must include the working memory mon-
itoring of the conditional rule, would be expected to acti-
vate the DLPF cortex, the head of the caudate, 和
parietal cortex, according to a well-established working
memory circuitry (穆勒 & 骑士, 2006; Wager & 史密斯,
2003; Braver et al., 1997; Petrides, 1994), as we observed
这里. 此外, the added task load could also explain
the increased activation of the right inferior frontal gyrus.
Activation of this region has been reported for sustained
注意力, working memory manipulation, and dual task re-
quirements (例如, McNab et al., 2008; Wager & 史密斯, 2003;
Coull, Frackowiak, & Frith, 1998). 因此, although the right
inferior frontal gyrus is critical for behavioral stopping, 作为
shown by lesion studies, activation of this region cannot
be taken as definitive evidence that it plays a causal role
in active braking. Future research using loss-of-function
approaches may be able to elucidate if the right inferior
frontal gyrus plays a necessary role in active braking, 和
imaging with higher temporal and spatial resolution may
be able to parse the different functions of the right inferior
frontal gyrus with respect to working memory load, 他们的-
tained attention, and proactive inhibitory control.
This study also provided interesting information about
response selection. 实验 2, which had no stop sig-
nals, clarified that the decision phase, which occurs before
response initiation, is associated with a reduction of cortico-
motor excitability (beneath baseline levels). Two aspects of
the wider literature suggest that this MEP suppression is re-
lated to active inhibition of the corticospinal tract via in-
creased gamma-aminobutyric acid (GABA)-ergic activity in
初级运动皮层. 第一的, MEP suppression has been pre-
viously reported for a pre-go-stimulus period—in which
it may help prevent premature responding (Duque &
Ivry, 2009; Boulinguez et al., 2008; Davranche et al., 2007;
Hasbroucq et al., 1997, 1999). Such studies have also shown
that the corticospinal excitability reduction relates to in-
creases in short interval intracortical inhibition or cortical
silent period—both indices of GABA-ergic inhibition in
M1. 第二, our data from Experiments 1 和 2 indicate
that the selection of the responding finger representation
within primary motor cortex does not occur until at least
120 msec after the presentation of the go stimulus. After this
time point, FDI MEP amplitudes are facilitated when the in-
dex finger is about to respond and further suppressed when
the little finger is about to respond. The strong suppression
of FDI MEP amplitude before activation of the little finger
is in line with previous studies of selective finger movement
(Beck et al., 2008; Stinear & Byblow, 2003). 尤其, 它
has been shown that intracortical inhibition can be re-
cruited above resting levels, and this contributes to the sup-
pression of MEP amplitude in nonmoving fingers (Stinear
& Byblow, 2003; Liepert, Classen, 科恩, & Hallett, 1998).
Our findings, in conjunction with the published literature,
所以, suggest that the preinitiation phase in this study
was accompanied by active inhibition of all possible re-
sponse representations. Following this, the initiation phase
was accompanied by an even greater inhibition of the
nonselected motor representation, possibly by continued/
increased GABA-ergic activity in primary motor cortex. 它
is possible that the putative inhibition in anticipation of
response initiation/selection is generated by a premotor
cortex circuit rather than the putative prefrontal/basal
1490
认知神经科学杂志
体积 22, 数字 7
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
2
我
7
e
–
1
p
4
d
7
F
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
哦
7
C
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
3
0
0
7
7
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
/
F
.
.
.
t
哦
n
1
8
中号
A
y
2
0
2
1
ganglia circuit important for outright stopping (or active
braking). Consistent with this, we did not observe signifi-
cant activation above baseline levels for go-noncritical trials
for the right inferior frontal gyrus, nor did we observe acti-
vation of the subthalamic nucleus region—and yet neuro-
physiological models propose that response initiation is
preceded by suppression of competitor motor programs
via the subthalamic nucleus (Mink, 1996). Future research
is required to further explore the neural correlates of puta-
tive inhibitory processes operating in anticipation of or at
the same time as the response initiation.
总之, we examined the neurocognitive mecha-
nisms underlying the response slowing that is observed in
anticipation of a possible stop. We suggest that this is an ex-
perimental model for the kind of real-world control that is
evident when people respond with restraint. We attained
results for two phases: a preresponse initiation phase and
a response initiation phase. The preresponse initiation pe-
riod of the task was associated with a global reduction of
corticomotor excitability (beneath baseline levels), 和
we speculate that this relates to GABA-ergic inhibition of
primary motor cortical representations in this early phase.
Once the response is selected, the corticomotor excitability
of the responding fingerʼs representation increases sharply,
whereas that of the nonresponding finger is reduced even
更远, consistent with continued application of a GABA-
ergic inhibitory mechanism. During the response initia-
tion phase, we observed a different pattern, which likely
explains the response delay effect. A difference in the rate
of corticomotor facilitation for critical and noncritical trials
emerged after 160 毫秒. Corticomotor excitability rose
more slowly, under the critical condition. This was compat-
ible with a modified version of the active braking hypoth-
esis as well as with prolonged decision stage and slower
motor facilitation accounts. 然而, other features of
the data pointed more clearly to active braking as a mecha-
nism underlying the response delay effect. 区别
between go-critical and noncritical trials was also reflected
in increased activation of a prefrontal region (the right in-
ferior frontal gyrus) that is required to outright suppress a
motor response. Although the imaging results cannot dis-
tinguish between active braking and prolonged decision
阶段 (via increased cognitive load) 账户, when taken
together with the TMS findings, we suggest that at least part
of the response delay effect is explained by active braking
and that this is probably reflected in activation of regions
of the brain important for outright stopping. 全面的, the re-
sults further our understanding of cognitive control by sug-
gesting that a neurocognitive system heretofore associated
with outright stopping is proactively recruited to enable
people to respond with restraint.
致谢
The authors thank Chris Chambers and Wery van den Wildenberg
for helpful comments, Russ Poldrack for making available the
fMRI data for reanalysis, and Sarah Sheldon for assisting with
data analysis. They are grateful to the reviewers and the journal
editor for constructive input. A. 右. A. is funded by an Alfred P.
Sloan Foundation Fellowship and NIH grant 1R01DA026452-
01A. F. V. is a Postdoctoral Fellow of Research Foundation-
Flanders (FWO-Vlaanderen).
Reprint requests should be sent to Adam R. Aron, Department
心理学系, 加州大学, 圣地亚哥, 拉霍亚, CA
92023, or via e-mail: adamaron@ucsd.edu.
参考
Aron, A. R。, 贝伦斯, 时间. E., 史密斯, S。, Frank, 中号. J。, & Poldrack,
右. A. (2007). Triangulating a cognitive control network
using diffusion-weighted magnetic resonance imaging
(MRI) and functional MRI. 神经科学杂志, 27,
3743–3752.
Aron, A. R。, Durston, S。, Eagle, D. M。, Logan, G. D ., Stinear,
C. M。, & Stuphorn, V. (2007). Converging evidence for a
fronto-basal-ganglia network for inhibitory control of
action and cognition. 神经科学杂志, 27,
11860–11864.
Beck, S。, 理查森, S. P。, Shamim, 乙. A。, 当, N。, Schubert,
M。, & Hallett, 中号. (2008). Short intracortical and surround
inhibition are selectively reduced during movement initiation
in focal hand dystonia. 神经科学杂志, 28,
10363–10369.
Boulinguez, P。, Jaffard, M。, Granjon, L。, & Benraiss, A. (2008).
Warning signals induce automatic EMG activations and
proactive volitional inhibition: Evidence from analysis of error
distribution in simple RT. 神经生理学杂志, 99,
1572–1578.
Braver, 时间. S。, 科恩, J. D ., Nystrom, L. E., Jonides, J。, 史密斯,
乙. E., & Noll, D. C. (1997). A parametric study of prefrontal
cortex involvement in human working memory.
Neuroimage, 5, 49–62.
Chambers, C. D ., Garavan, H。, & Bellgrove, 中号. A. (2009).
Insights into the neural basis of response inhibition from
cognitive and clinical neuroscience. Neuroscience and
Biobehavioral Reviews, 33, 631–646.
Coull, J. T。, Frackowiak, 右. S。, & Frith, C. D. (1998). Monitoring
for target objects: Activation of right frontal and parietal
cortices with increasing time on task. Neuropsychologia,
36, 1325–1334.
Coxon, J. P。, Stinear, C. M。, & Byblow, 瓦. (2007). Selective
inhibition of movement. 神经生理学杂志, 97,
2480–2489.
Davranche, K., Tandonnet, C。, Burle, B., Meynier, C。, Vidal, F。, &
Hasbroucq, 时间. (2007). The dual nature of time preparation:
Neural activation and suppression revealed by transcranial
magnetic stimulation of the motor cortex. European Journal
of Neuroscience, 25, 3766–3774.
德容, R。, Coles, 中号. G。, & Logan, G. D. (1995). Strategies
and mechanisms in nonselective and selective inhibitory
motor control. 实验心理学杂志, 21,
498–511.
Devanne, H。, Lavoie, 乙. A。, & Capaday, C. (1997).
Input-output properties and gain changes in the human
corticospinal pathway. Experimental Brain Research,
114, 329–338.
Duque, J。, & Ivry, 右. 乙. (2009). Role of corticospinal suppression
during motor preparation. 大脑皮层, 19, 2025–2037.
Hasbroucq, T。, Kaneko, H。, Akamatsu, M。, & Possamai, C. A.
(1997). Preparatory inhibition of cortico-spinal excitability:
A transcranial magnetic stimulation study in man. Brain
研究, Cognitive Brain Research, 5, 185–192.
Hasbroucq, T。, Osman, A。, Possamai, C. A。, Burle, B., Carron, S。,
Depy, D ., 等人. (1999). Cortico-spinal inhibition reflects
Jahfari et al.
1491
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
2
我
7
e
–
1
p
4
d
7
F
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
哦
7
C
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
3
0
0
7
7
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
.
.
.
/
F
t
哦
n
1
8
中号
A
y
2
0
2
1
time but not event preparation: Neural mechanisms of
preparation dissociated by transcranial magnetic stimulation.
心理学学报 (阿姆斯特丹), 101, 243–266.
豪厄尔, 瓦. C。, & Kreidler, D. L. (1963). Information-processing
under contradictory instructional sets. 杂志
实验心理学, 65, 39–46.
Liepert, J。, Classen, J。, 科恩, L. G。, & Hallett, 中号. (1998).
Task-dependent changes of intracortical inhibition.
Experimental Brain Research, 118, 421–426.
Logan, G. D ., & 考恩, 瓦. 乙. (1984). On the ability to
inhibit thought and action: A theory of an act of control.
心理评论, 91, 295–327.
McNab, F。, Leroux, G。, Strand, F。, Thorell, L。, 伯格曼, S。, &
Klingberg, 时间. (2008). Common and unique components of
inhibition and working memory: An fMRI, within-subjects
调查. Neuropsychologia, 46, 2668–2682.
Mink, J. 瓦. (1996). The basal ganglia: Focused selection and
inhibition of competing motor programs. Progress in
Neurobiology, 50, 381–425.
穆勒, 氮. G。, & 骑士, 右. 时间. (2006). The functional
neuroanatomy of working memory: Contributions of human
brain lesion studies. 神经科学, 139, 51–58.
Pascual-Leone, A。, Valls-Solé, J。, Wassermann, 乙. M。, Brasil-Neto,
J。, 科恩, L. G。, & Hallett, 中号. (1992). Effects of focal
transcranial magnetic stimulation on simple reaction time
to acoustic, visual and somatosensory stimuli. Brain, 115,
1045–1059.
Petrides, 中号. (1994). Frontal lobes and working memory:
Evidence from investigations of the effects of cortical
excisions in nonhuman primates. 在F中. Boller, & J. Grafman
(编辑。), Handbook of neuropsychology ( 卷. 9, PP. 59–84).
阿姆斯特丹: 爱思唯尔.
Rinkenauer, G。, Osman, A。, Ulrich, R。, Muller-Gethmann, H。, &
Mattes, S. (2004). On the locus of speed-accuracy trade-off in
reaction time: Inferences from the lateralized readiness
潜在的. 实验心理学杂志: General,
133, 261–282.
Rossini, 磷. M。, Barker, A. T。, Berardelli, A。, Caramia, 中号. D .,
Caruso, G。, Cracco, 右. Q., 等人. (1994). Non-invasive
electrical and magnetic stimulation of the brain, spinal
cord and roots: Basic principles and procedures for
routine clinical application. Report of an IFCN committee.
Electroencephalography and Clinical Neurophysiology,
91, 79–92.
Stinear, C. M。, & Byblow, 瓦. D. (2003). Role of intracortical
inhibition in selective hand muscle activation. 杂志
Neurophysiology, 89, 2014–2020.
Verbruggen, F。, & Logan, G. D. (2009A). Models of response
inhibition in the stop-signal and stop-change paradigms.
Neuroscience and Biobehavioral Reviews, 33, 647–661.
Verbruggen, F。, & Logan, G. D. (2009乙). Proactive
adjustments of response strategies in the stop-signal
范例. 实验心理学杂志: 人类
Perception and Performance, 35, 835–854.
Vink, M。, Kahn, 右. S。, Raemaekers, M。, van den Heuvel, M。,
Boersma, M。, & Ramsey, 氮. F. (2005). Function of striatum
beyond inhibition and execution of motor responses.
人脑图谱, 25, 336–344.
Wager, 时间. D ., & 史密斯, 乙. 乙. (2003). Neuroimaging studies of
工作记忆: A meta-analysis. 认知的, Affective &
Behavioral Neuroscience, 3, 255–274.
Zandbelt, B., Van Buuren, M。, Gladwin, 时间. E., Hoogendam, R。,
Kahn, S。, & Vink, 中号. (2008). Brain regions involved in
response inhibition are also activated during anticipation
of inhibition. Paper presented at the Society for
神经科学.
Ziemann, U。, Tergau, F。, Netz, J。, & Hömberg, V. (1997).
Delay in simple reaction time after focal transcranial
magnetic stimulation of the human brain. 脑研究,
744, 32–40.
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
2
我
7
e
–
1
p
4
d
7
F
9
/
1
2
9
2
3
/
8
7
9
/
1
1
6
4
哦
7
C
9
n
/
1
2
0
7
0
7
9
0
0
2
1
1
4
3
0
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
3
0
0
7
7
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
.
F
.
t
/
.
哦
n
1
8
中号
A
y
2
0
2
1
1492
认知神经科学杂志
体积 22, 数字 7