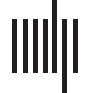RESEARCH ARTICLE
Task-Induced Functional Connectivity of Picture
Naming in Healthy Aging: The Impacts of Age
and Task Complexity
开放访问
杂志
Perrine Ferré1, Julien Jarret1
, Simona Maria Brambati1,2, Pierre Bellec1,2
, and Yves Joanette1
1Centre de recherche de l’Institut universitaire de gériatrie de Montréal, 蒙特利尔, Québec, 加拿大
2Université de Montréal, Québec, 加拿大
关键词: 功能磁共振成像, 功能连接, healthy aging, picture naming, 语言, task-induced
功能连接
抽象的
The topological organization of the brain, governed by the capacity of brain regions to
synchronize their activity, allows for cost-effective performance during everyday cognitive
活动. Functional connectivity is an fMRI method deemed task-specific and demand-
dependent. Although the brain undergoes significant changes during healthy aging,
conceptual knowledge and word-production accuracy are generally preserved. 这
exploration of task-induced functional connectivity patterns during active picture naming may
thus provide additional information about healthy functional cerebral mechanisms that are
specifically adapted to the cognitive activity at hand. The goal of this study is to assess and
describe age-related differences in functional connectivity during an overt picture-naming
任务, as well as to compare age-related differences under complex task demand, defined by
lexical frequency. Results suggest both age-specific and task-specific mechanisms. 在里面
context of preserved behavioral performance in a picture-naming task, older adults show a
complex array of differences in functional connectivity architecture, including both increases
and decreases. In brief, there is increased segregation and specialization of regions that are
classically assigned to naming processes. Results also expand on previous word-production
studies and suggest that motor regions are particularly subject to age-related differences. 这
study also provides the first indication that intrinsic task demand, as manipulated by lexical
频率, interacts little with the relationship between age and functional connectivity.
一起, these findings confirm the value of task-induced functional connectivity analysis in
revealing the brain organization that subserves task performance during healthy aging.
介绍
The aging of the population worldwide creates both immense opportunities and numerous
challenges regarding health and wellness. This critical demographic change is compelling
neuroscientists to engage in studies of cognition in aging, so they can better understand the
constituents of cognitive health, which is central to quality of life (WHO, 2018). 虽然
many cognitive abilities typically decline with healthy aging, conceptual knowledge is long
preserved (Ben-David, Erel, Goy, & 施耐德, 2015; Goulet, Ska, & Kahn, 1994; Salthouse,
2014). Its impairment may be an early marker of major or mild neurocognitive disorders
(Blackwell et al., 2004; 磨坊主, 罗杰斯, Siddarth, & 小的, 2005; Reilly, Peelle, Antonucci, &
Grossman, 2011). The assessment of conceptual knowledge classically includes picture
引文: Ferré, P。, Jarret, J。, Brambati,
S. M。, Bellec, P。, & Joanette, 是. (2020).
Task-induced functional connectivity of
picture naming in healthy aging: 这
impacts of age and task complexity.
Neurobiology of Language, 1(2)
161–184. https://doi.org/10.1162/
nol_a_00007
DOI:
https://doi.org/10.1162/nol_a_00007
支持信息:
https://doi.org/10.1162/nol_a_00007
已收到: 30 九月 2019
公认: 04 行进 2020
利益争夺: 作者有
声明不存在竞争利益
存在.
通讯作者:
Perrine Ferré
perrine.ferre@umontreal.ca
处理编辑器:
Jonathan Peelle
版权: © 2020 马萨诸塞州
Institute of Technology. 已发表
under a Creative Commons Attribution
4.0 国际的 (抄送 4.0) 执照.
麻省理工学院出版社
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
naming, a sensitive word-production test at the core of many clinical assessment tools (例如,
intraoperative language mapping, differential diagnosis; Moritz-Gasser, Herbet, & Duffau, 2013).
Although older individuals often complain about proper name retrieval (Condret-Santi et al.,
2013), they often obtain accuracy scores similar to those of younger adults when naming pictures
of common names. 然而, this typically occurs at the expense of longer response times (Baciu et al.,
2016; Hoyau et al., 2017; Wierenga et al., 2008) in possible relation with a general slowing in
information processing (Feyereisen, Demaeght, & Samson, 1998). The dissociation between time
and accuracy makes this task particularly interesting, as it suggests that older adults engage in
adaptive mechanisms to perform it adequately. This study thus aims to explore the specific neuro-
functional processes that underlie picture naming, a preserved cognitive ability.
The field of cognitive neuroscience of aging (Cabeza, Nyberg, & 公园, 2016) has revealed a
dynamic interplay between declines in structural and functional resources and plasticity phe-
nomena that support cognitive performance. Consistent patterns of enhanced recruitment with
older age, including greater prefrontal bilateral activation, have been found across many cog-
nitive tasks using task-induced blood oxygen level–dependent (大胆的) amplitude activation dif-
ferences (Cabeza, 2002; 戴维斯, 丹尼斯, Daselaar, Fleck, & Cabeza, 2008). Such manifestations
of functional brain adaptation potentially relate to greater demands for cognitive-control pro-
cesses in older adults (公园 & Reuter-Lorenz, 2009; Schneider-Garces et al., 2010). 什么时候
age-related differences such as decreased specificity and more diffuse brain activity are not as-
sociated with performance, a dedifferentiation phenomenon is invoked (Cabeza et al., 2018).
Although such phenomena have generally been explored using abilities known to decline with
年龄, less is known about the functional organization that sustains a well-preserved ability.
Older individuals may exhibit longer latencies and phonological imprecision (Feyereisen,
1997; Goulet et al., 1994; Shafto, James, Abrams, & Tyler, 2017). Some complain about
word-finding difficulties, but mostly so for proper names and with no relation to objective cog-
nitive difficulty (Condret-Santi et al., 2013). 然而, 一般, their accuracy is preserved,
sometimes even improved, when a simple common names task is used, where accuracy and not
response time is scored, and when there are few time constraints (LaBarge, 爱德华兹, &
Knesevich, 1986; Salthouse, 2014; Schmitter-Edgecombe, Vesneski, & 琼斯, 2000;
Verhaegen & Poncelet, 2013; Wierenga et al., 2008). 全面的, naming abilities for common
names appear behaviorally preserved when task demand is controlled, at least until 65 年
年龄 (Salthouse, 2014). Picture naming typically involves an extensive neural network reflect-
ing every cognitive step required to produce a known word. Although many interindividual var-
iations exist, the core areas have been described in young adults (Duffau, Moritz-Gasser, &
Mandonnet, 2014; Friederici & Gierhan, 2013; Price, 2012; Sarubbo et al., 2016). The occipital
cortex and the middle and inferior posterior temporal cortex are involved in the semantic pro-
cessing of visual attributes, along with the orbital inferior frontal gyrus (IFG) for multimodal pro-
cessing. 一起, the opercular IFG, premotor cortex, insula, and inferior parietal and superior
temporal cortex are in charge of phonological processing and articulation. These core areas,
although segregated, do not operate in isolation. 尤其, age-related neurofunctional re-
organization will often happen at the scale of networks, if not the whole brain, by changing ac-
tivity interactions between regions (Perry et al., 2015; Tomasi & Volkow, 2012; Tsvetanov et al.,
2016). 因此, 功能连接 (FC) is a tool of choice to study neurofunctional mecha-
nisms associated with aging.
FC in healthy aging has been studied mainly using resting-state and associated networks,
such as the default mode network (DMN) (Sala-Llonch, Bartrés-Faz, & Junqué, 2015). 在-
tegrity of DMN connectivity appears to be particularly affected by age and related to perfor-
曼斯 (Mak et al., 2017). Generally speaking, a decrease in within-network FC is reported,
Neurobiology of Language
162
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
along with increased FC between functionally related regions (Mak et al., 2017; Sala-Llonch et al.,
2015). This pattern appears in line with previous activation studies that demonstrated decreased
deactivation of the DMN during task performance in older individuals (Persson, Pudas, Nilsson,
& Nyberg, 2014). Examples of increased brain FC with regions belonging to the same functional
network but typically not recruited by younger adults have also been detected using lexicose-
mantic tasks (阿加瓦尔, Stamatakis, Geva, & Warburton, 2016; Hoyau et al., 2018; La et al.,
2016; Marsolais, Perlbarg, Benali, & Joanette, 2014; 穆勒, Mérillat, & Jäncke, 2016). 一些
studies of word production have reported BOLD signal increases in parietal, frontal, or temporal
regions with age (例如, Hoyau et al., 2017; 莫尼耶, Stamatakis, & Tyler, 2014), as well as contra-
lateral recruitment (例如, La et al., 2016; Meinzer et al., 2012). Along with an impact of age, effects
of behavioral performance and task demand are also expected: A recent meta-analysis focusing on
semantic cognition and aging (Hoffman & Morcom, 2018) concluded that augmented activation
was principally observed when older adults performed worse than younger adults, 例如,
during tasks drawing on executive functions.
然而, some authors have recently argued in favor of state-dependent differences in re-
gional functional connectivity with age (坎贝尔 & Schacter, 2016; Greene, 高, Scheinost,
& Constable, 2018; Samu et al., 2017). In regard to lexical knowledge, recent evidence points
toward distinct FC patterns for different tasks (例如, synonyms, antonyms, picture naming) 在
老年人 (Ferré et al., 2019; Varangis, Razlighi, Habeck, Fisher, & Stern, 2019).
It is important to note that a task paradigm allows for direct manipulation of the task require-
ments to characterize neural mechanisms that underlie brain-behavior associations and poten-
tial strategic mechanisms (Crowell et al., 2019; Finn et al., 2017; Greene et al., 2018; Persson,
Lustig, 纳尔逊, & Reuter-Lorenz, 2007; Steffener et al., 2014). Prior studies of aging and cog-
nition have concluded that domain-general control processes become more involved when
general cognitive load increases, to which older adults appear particularly sensitive
(Campbell et al., 2016; 公园 & Reuter-Lorenz, 2009; Peelle, 2019; 王, Dew, & Cabeza,
2015). 然而, little is known about the impact of manipulating task requirements through psy-
chometric criteria (Cole, 史密斯, & 贝克曼, 2010). 例如, lexical frequency influ-
ences brain activity and performance during naming (例如, Basso et al., 2013; 伯克,
MacKay, Worthley, & Wade, 1991). 的确, low-frequency words trigger more tip-of-the-
tongue states than high-frequency words (Burke et al., 1991), and older adults have more dif-
ficulty producing the correct name for low-frequency items (Au et al., 1995; LaGrone &
Spieler, 2006; Rogalski, Peelle, & Reilly, 2011). Lexical frequency thus offers another way
to manipulate the requirements of a picture-naming task, using intrinsic task characteristics.
The general aim of this study is to characterize possible age-related differences in task-
induced FC, using a picture-naming task, while considering the impact of lexical frequency
as an indicator of task difficulty.
The first goal was to assess and describe age-related differences in FC of the regions acti-
vated by the picture-naming task as well as in the DMN during word production. On the basis
of previous reports of increased activity during task performance, older adults were expected
to show higher FC in the regions activated by the task, in relation to greater reliance on lifelong
accumulated semantic knowledge (Agarwal et al., 2016; Hoyau et al., 2018; Marsolais et al.,
2014; Tran et al., 2018) or domain-general control mechanisms (Cabeza et al., 2018; Geerligs,
Maurits, Renken, & Lorist, 2014; La et al., 2016). Decreases in FC within DMN regions, 沿着
with increases in the rest of the brain, are also expected during task performance, in line with
most previous FC studies of aging (Andrews-Hanna et al., 2007; Damoiseaux et al., 2008;
Geerligs et al., 2014).
Neurobiology of Language
163
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
The second goal of the study was to describe the differential impact of lexical frequency on FC
as a function of age. Differences between conditions were expected to be larger for older adults
and to indicate enhanced connectivity, as a result of more demanding psycholinguistic process-
ing and as observed in the context of greater task demand (Avelar-Pereira, Bäckman, Wåhlin,
Nyberg, & Salami, 2017; 坎贝尔 & Schacter, 2016; Dixon et al., 2017; Dubois & Adolphs,
2016; Geerligs, 鲁比诺夫, & Henson, 2015; Grady, Sarraf, Saverino, & 坎贝尔, 2016).
By gathering such a task-specific set of information, this study offers a baseline for further
exploration of adaptive mechanisms subserving the preservation of cognition throughout life.
METHOD
Population
Seventy-two participants (38 young adults, 34 老年人) gave their informed consent to
participate in this study, in accordance with local ethics committee guidelines. 参加者
were native French speakers and right-handed, and were free of neurological disorders and
a history of drug or alcohol dependency, major depression, or moderate to severe auditory
or visual disorders. Adults 65-years-old or older underwent a cognitive and hearing screening.
The Montreal Cognitive Assessment (Nasreddine et al., 2005) was used to test for mild cogni-
tive impairment, with a standard cutoff score of 26. A pure-tone test was used to ensure that all
older participants had hearing acuity within ISO standard 7029:2000. 此外, 广泛的
screening questionnaires were used to exclude participants with MRI contraindications. 这
final sample, after quality control of fMRI preprocessing, 包括 37 young adults and 31 较老的
adults (桌子 1).
Task Design
Participants were asked to complete an overt object naming task during fMRI data acquisition:
The Boston Naming Test (BNT) (古德格拉斯, 卡普兰, & Weintraub, 1983). The BNT is among
the most widely used picture-naming tasks in both clinical and experimental settings. 它是
therefore considered to provide a basis for knowledge development. Participants were asked
桌子 1. Demographic and behavioral characteristics
Age group
Mean age (标清)
性别 (male/female)
Mean years of education (标清)
[min–max]
Mean accuracy (标清) [min–max]
BNT
BNT – easy condition
BNT – hard condition
18–35 (N= 37)
26 (5.2)
61–80 (N= 31)
70 (5.5)
21/16
12/19
14 (3.3) [9–20]
14 (3.4) [11–21]
45 (7.7) [24–57]
27 (2.9) [23–30]
18 (5.1) [7–27]
48 (5.5) [34–55]
28 (2.2) [26–30]
17 (3.9) [9–25]
BNT – reaction time (seconds)
1284 (189.3) [959–1815]
1423 (208.1) [1001–1906]
MoCA
–
28 (1.2) [26–30]
BNT, Boston Naming Test; MoCA, Montreal Cognitive Assessment.
Neurobiology of Language
164
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
to name the pictures they saw on the screen aloud, as fast as possible. Word-production pro-
cesses occur rapidly, typically within 600 ms of seeing an object (Indefrey & Levelt, 2004).
Pilot acquisitions with both young and healthy older adults occurred before settling on the
final task paradigm.
Each of the 60 BNT stimuli was presented for 1,500 多发性硬化症, and participants had an additional
1,500 ms to give their answer before the next trial. A long allowable response time was pre-
ferred because it is only after 700 ms that neurofunctional response to the tip-of-the-tongue
phenomenon is typically observed (Shafto & Tyler, 2014); 而且, the pilot acquisitions sug-
gested the same thing. Many previous FC studies used resting-state and offline performance
措施 (Sala-Llonch et al., 2015), but those methods are suspected of reflecting the actual
cognitive processes during the task less closely (坎贝尔 & Schacter, 2016; Tran et al., 2018).
因此, an overt naming task was preferred in this study. An interstimulus interval of 350 多发性硬化症
separated the stimuli. A fixation cross indicated the end of the trial. The task was composed
的 12 blocks lasting 17.5 s, with five images each to be named. The blocks were separated by
rest epochs (fixation cross) lasting 17.5 s, for a total of 145 volumes. Half of the blocks had a
low frequency (high requirement) 等级. Linguistic complexity was defined by the lexical fre-
quency obtained from the French lexical database lexique3 (http://lexique.org/). To validate
the assumption that a frequent item should generally be named successfully, each item’s cat-
egorization was compared with the success rate of the test item from the French norms: A word
designated as frequent systematically triggered 80% 成功 (罗伯茨 & Doucet, 2011).
Behavioral Analysis
Accuracy (number of correct responses given in the maximum time window) 和回应
次 (RT) for all naming trials were first analyzed to determine whether task performance
was similar across age groups. Three trained raters independently scored the accuracy results
and reached a consensus. Age groups were compared for their total score as well as each
frequency level using two independent sample t tests. An analysis of variance (ANOVA) 经测试
the relationship between age group and accuracy or RT for each frequency level. No devia-
tions from homoscedasticity or normality were observed. Three individuals were excluded
from the behavioral analysis because of technical issues resulting in missing data (例如, 和-
plugged microphone, inaudible answer). 桌子 1 presents all the behavioral results.
fMRI Scanning and Data Processing
MRI images were acquired with a 32-channel head coil and a 3T SIEMENS TrioTim magnetic
resonance imaging system. Participants were instructed to limit their movement to the extent
可能的, and a practice session was held before the scanning session to ensure they knew how
to limit their movement. Foam rubber pads within the head coil also restricted head movement.
Earplugs reduced scanner noise. A microphone was oriented toward the mouth to allow vocal
记录, using MR Confon (https://www.crsltd.com/tools-for-functional-imaging/audio-for-
fmri/mr-confon/). When necessary, vision was corrected using MRI-175 compatible lenses that
matched the distance prescription used by the participant. The task stimuli were presented using
DMDX presentation software (Forster & Forster, 2003).
Anatomical images (T1) were acquired with a Multi echo multi planar rapid gradient echo
pulse sequence and a Generalized autocalibrating partial parallel acquisition (GRAPPA) 交流电-
celeration factor using the following parameters: field of view (FoV) = 256.0 mm2, 矩阵大小=
256 × 256, 176 slices covering whole brain, 1 mm isotropic voxel size, echo time/repetition time
(TE/TR) = 1.64/253 多发性硬化症, flip angle = 7.0°.
Neurobiology of Language
165
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
Functional images (T2*) were acquired with an echo-planar imaging pulse sequence and a
GRAPPA acceleration factor using the following parameters: FoV = 220 × 220 毫米, 矩阵大小=
74 × 74, 50 ascending slices covering the whole brain, 3 mm isotropic voxel size, TR/TE =
3,000/20 多发性硬化症, flip angle = 90°. T2* image acquisition was oriented −30° from the anterior
commissure–posterior commissure line (to reduce the signal loss from the anterior temporal
lobes). The first five volumes of each run were automatically discarded during acquisition.
预处理
Preprocessing of structural and BOLD functional metrics was done using SPM12 (http://万维网.
fil.ion.ucl.ac.uk/spm/) and CONN toolboxes (v.18.b; www.nitrc.org/projects/conn) imple-
mented in Matlab 2015b (https://www.mathworks.com/). Functional images were first coregis-
tered and realigned to account for minor head motion using the CONN preprocessing
pipeline.
Using the VBM12 toolbox in SPM12 (http://dbm.neuro.uni-jena.de/wordpress/vbm/), 核-
gistered structural (T1) images were segmented into gray matter, white matter, and cerebrospi-
nal fluid, with a sampling of 1.5 mm × 1.5 mm × 1.5 mm using trilinear interpolation. 后
segmentation, we performed image normalization using Diffeomorphic Anatomical
Registration through Exponentiated Lie (DARTEL; Ashburner, 2007) to create a custom tem-
plate. The flow field images obtained during the DARTEL template creation were used to warp
all realigned functional images and the coregistered structural images into the Montreal
Neurological Institute (MNI) 空间. Images were modulated by multiplying the Jacobian de-
formation parameters defined during normalization to preserve the total amount of original
gray matter before normalization (Ashburner & 弗里斯顿, 2005; Zhu et al., 2013). 下一个, the mod-
ulated/warped images were smoothed with a 6 mm full-width at half-maximum (FWHM) iso-
tropic Gaussian kernel and normalized into the MNI space.
Functional images were coregistered to the structural T1 images obtained during the pre-
vious realignment step using CONN options. Smoothed normalized images were entered in
the CONN toolbox.
A motion-censoring procedure was applied to remove unwanted motion, 生理,
and other artifactual effects from the BOLD signal. An ART-based functional outlier detection
method was used, as implemented in the CONN toolbox (Mazaika, Whitfield, & 库珀,
2005). The threshold was established using the maximum voxel displacement with a scrub-
bing criterion established at 0.9 mm scan-to-scan head motion or global signal intensity 5 标清
above the mean signal for the session (Mazaika, Hoeft, Glover, & Reiss, 2009; Whitfield-
Gabrieli & Nieto-Castanon, 2012). A dummy variable represented each outlier in the first-level
denoising step. The mean number of invalid scans was 4.84, 或者 3.34% of the total number of
volumes. Three participants who presented more than 25% outliers out of their total volumes
were excluded from further FC analysis.
An anatomical component-based noise correction method was also applied (Behzadi,
Restom, Liau, & 刘, 2007), regressing the white matter and cerebrospinal fluid from the
BOLD signal. This method has proven useful to improve the specificity of connectivity mea-
surements (Muschelli et al., 2014), and it partially reduced the effect of vascular health on FC
措施 (Geerligs, Tsvetanov, & Henson, 2017).
The six realignment parameters (with their first temporal derivatives) and the task effect
(BOLD time series orthogonalization to task effects) were also included as regressors.
Neurobiology of Language
166
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
最后, a high-pass filter (<0.01 Hz) was applied after nuisance regression, to filter signals
related to physiological or motion artifacts (Muschelli et al., 2014; Satterthwaite et al., 2013). A
high-pass filter has been demonstrated to produce stronger, more reliable age effects than a
bandpass filter (Geerligs et al., 2017).
Quality assessment was performed before and after denoising, by visually inspecting the
overlay of functional and structural individual realigned images, the overlay of the functional
images and the MNI template, as well as the BOLD functional time points movie and distri-
bution of the scrubbed volumes across time. One participant was excluded from further FC
analysis because of an isolated but massive amplitude movement. No other anomalies were
noted.
In total, 4 participants were excluded of the initial 72. The remaining sample was com-
posed of 68 individuals: 37 young adults and 31 older adults.
Functional connectivity fMRI analysis
Regions of interest (ROIs) were defined based on the BOLD signal characterizing the execution
of the naming task (the main effect of naming contrast). This method reduces the number of
observations and guides the FC data analysis. Reduction allows for easier and clearer interpre-
tation and is deemed a simple yet powerful method (Fjell & Walhovd, 2016).
Task-induced ROI definition. A data-driven approach using a task- and sample-specific template
was favored over canonical resting-state networks because the latter may be less sensitive to
task-related changes in network connectivity (Crowell et al., 2019; Davis, Stanley, Moscovitch,
& Cabeza, 2017). For example, several of the networks found in the left perisylvian language
regions during active language tasks (e.g., left inferior frontal, posterior temporal, and inferior
parietal cortices) may appear as a single network of correlated regions using a resting state
(Jackson, Hoffman, Pobric, & Lambon Ralph, 2015; Liljeström, Stevenson, Kujala, &
Salmelin, 2015; Tran et al., 2018). Thus, seed ROIs were based on the whole brain statistical
parametric map of the task main effect for the whole sample. However, to reduce the number of
observations and limit redundancy, we selected the activation peaks that best summarized the
whole brain activation maps in the whole brain for the whole sample.
First, voxel-wise T-maps were constructed for each subject using a task > rest contrast (任务
main effect). Second-level group analyses were then performed to test for significant differ-
ences across groups. Activation peaks of the union of the task main effect maps (t tests) in both
age groups were determined. A sphere was centered on each peak with a radius of 7 mm using
MARSBAR (http://marsbar.sourceforge.net). The Euclidean distance between the centroid’s co-
ordinates in the standard MNI space was also calculated, to verify that the distance between
peaks was at least twice the width of the smoothing kernel (IE。, 12 毫米).
第二, we grouped the activation peaks according to responses during the task and se-
lected the most relevant from each grouping. A one-sample t tests was computed for all partici-
pants using all activation peaks during the task condition. An ROI-to-ROI hierarchical
clustering algorithm sorted all regions and grouped ROIs showing the most similar time series,
using a network-based statistic (NBS; 扎莱斯基, 假如, & 布莫尔, 2010) with a false discov-
ery rate connection-level threshold of p < .0001 at the analysis level. The ROI with the largest
size (i.e., the number of suprathreshold connections between this seed and all other ROIs) in
each cluster was selected for further seed-to-voxel analysis.
To better compare these results with the literature, one additional seed was selected to rep-
resent the DMN. Age-related differences in DMN connectivity have indeed been consistently
Neurobiology of Language
167
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
o
_
a
_
0
0
0
0
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Naming functional connectivity in aging
reported, especially in the posterior cingulate cortex (PCC; Andrews-Hanna et al., 2007; Chan,
Alhazmi, Park, Savalia, & Wig, 2017; Geerligs et al., 2014; Kong et al., 2018; Tomasi &
Volkow, 2010). In sum, a total of six ROIs were selected for further seed-to-voxel FC analysis.
Functional connectivity statistics. The spatial topography of seed-to-voxel FC was examined
using the selected ROI spheres defined as seeds in previous steps.
Seed-to-voxel connectivity was measured at the first level using an haemodynamic response
function-weighted general linear model (which deweights the initial scans within each block).
The Fisher transform of the Pearson correlation between each seed time series and all other
voxels was calculated.
Group-level analysis in CONN implemented repeated-measures analyses using the ReML
estimation of covariance components and evaluated through F-statistical parameter maps.
Correction for multiple comparisons was done using a combined voxel-level height threshold
( p < .001 uncorrected) and a cluster extent threshold ( p < .05 family-wise error [FWE]
corrected).
Connectivity maps were automatically labeled by CONN using the anatomy toolbox v2.0
(Eickhoff et al., 2005) and manually checked using the AAL, Tzourio-Mazoyer, and Brodmann
atlases.
The main contrasts of interest were the differences between age groups in absolute connec-
tivity during the task condition—ignoring the fixation epochs—as tested by a two-sample t test
for each ROI. Results from a simple linear association in FC studies can lead to misleading
interpretations (Ferreira et al., 2016; Geerligs & Henson, 2016; Song et al., 2012). For exam-
ple, “increases” in FC may reflect both an increase in the magnitude of a positive correlation or
a loss of anticorrelation. To distinguish between processes and support interpretation, within-
group mean connectivity maps during the task were thus extracted.
The second aim of the study was to investigate how the age effect on FC varies as a function
of task demand. The condition (low vs. high frequency) by group (older vs. younger adults)
interaction was tested. A 2 × 2 mixed ANOVA between age group and frequency level was
computed for each ROI.
We were interested in the main effect of condition. The simple main effect of complex-
ity in each group (paired t test) and mean within-group FC at each frequency level were
explored for every significant interaction to support description and interpretation of the
interaction.
RESULTS
Behavioral Results
Mean behavioral performance metrics (accuracy and RTs) were first compared between
groups using an ANOVA. Interaction effects were tested by comparing accuracy scores and
RTs in each lexical frequency condition for younger and older individuals. The hypothesis
regarding both accuracy and reaction times were verified, and although frequency did interact
with RT, no main effect of frequency on accuracy scores was observed.
As expected, younger (mean = 44.67, SD = 7.69) and older (mean = 43.50, SD = 7.93)
adults did not differ significantly in their total BNT accuracy scores (F[1,63] = 0.005, p =
0.946), neither in the high-frequency (F[1,63] = 1.13, p = 0.292) nor in the low-frequency
(F[1,63] = 0.89, p = 0.347) condition.
Neurobiology of Language
168
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
o
_
a
_
0
0
0
0
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Naming functional connectivity in aging
There was a significant difference in the overall mean RT of younger (mean = 1284.24, SD =
189.37) and older (mean = 1417.34, SD = 208.41) adults (F[1,63] = 5.74, p = 0.005). The inter-
action between age group and word frequency was significant on RT (F[1,63]= 5.74, p=0.005),
and when tested individually, interaction between age and RT is significant for the low frequency
(F[1,63]= 10.16, p=0.002) but not the high frequency (F[1,63]= 3.037 p=0.086) condition, show-
ing that older adults were significantly slower to answer in the more difficult condition.
Identification of the Task-Induced Regions of Interest
The thresholded map of the union of both age groups in the naming versus fixation contrast
was used to determine peaks of activation in the task-related network. As expected, the nam-
ing contrast activated an extensive bilateral occipital-temporal-parietal-frontal circuit in both
younger and older adults. The maps of the two age groups were, for the most part, similar
(Figure 1). The only significant difference was found in the older > younger contrast in the
right inferior cerebellum lobule VIII (MNI: 27 −54 −46). This cluster was encompassed in a
more massive occipital cluster in the union activation maps, so it was not selected as a distinct
seed in FC analysis.
The task-induced statistical parametric map was composed of 13 clusters. The ROI-to-ROI
analysis revealed that these clusters belonged to five distinct functional clusters: A visual occip-
ital cluster, a sensorimotor cluster, a left frontoparietal cluster, a frontal anterior cluster, and a
posterior superior temporal gyrus (STG) cluster. The seeds that presented the best spatial overlap
and the most similar functional response to each canonical network were the left occipital, 左边
premotor cortex, left orbital IFG, left opercular IFG, and left posterior superior temporal gyrus.
MNI coordinates for the task-induced activation peaks, along with the NBS functional hi-
erarchical clustering statistics, are presented in Table 2.
One additional ROI was selected in the PCC from among the CONN network ROI list to
represent the DMN (MNI coordinates: x = 1 y = −61 z = 38). 数字 1 illustrates the method
and results of selecting the seed ROIs.
Age-Related Differences in Functional Connectivity
The main goal of this study was to assess and describe age-related differences in task-induced
FC in the regions activated during the naming task.
The effect of age on FC topography during task completion was investigated using a seed-
to-voxel FC for each selected ROI for the task and the PCC (t test between two independent
样品). The mean within-group FC was also computed to better characterize the differences
in topological FC organization. Results are reported with a voxel-level height threshold (p <
.001 uncorrected) and a cluster extent threshold (p < .05 FWE corrected) and illustrated in
Figure 2. The mean and group difference maps for all other activation peaks are available
in Supporting Information 1.
Based on previous reports of enhanced differences in FC architecture during a task, older
adults were expected to show significant differences in the integration of both the task and
default mode networks: decreased FC within the DMN, but increased FC within regions out-
side the DMN, as well as increased FC in the task-activated regions with semantic and/or ex-
ecutive control regions. The results only partially validated the initial expectations.
The task-induced FC architecture presented many similarities between age groups. Although
small, significant clusters of age-related differences were nonetheless present for each ROI of the
task and the DMN. Both increases and decreases were observed with aging.
Neurobiology of Language
169
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
o
_
a
_
0
0
0
0
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Naming functional connectivity in aging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
o
_
a
_
0
0
0
0
7
p
d
/
.
l
Figure 1. Task-induced seed definition: Task-induced activation maps of younger and older adults, ROI functional hierarchical clustering,
and selected seeds. Above: activation clusters in the main contrast of interest (task > 休息) for both younger (黄色) 和更老的 (in cyan)
adults. Overlaps (绿色) are observed between younger and older adults in the task contrast. Slice numbers are indicated on the sagittal axis.
以下: Functional hierarchical clustering of the ROI spheres for the task-induced activation peaks. 这 13 activation peaks belonged to five
functional clusters. The most relevant activation peaks are indicated in bright yellow. 例如, the left STG shows the largest number of
connections with all other ROIs. Each colored connection (magenta, 蓝色的, 绿色的, orange) represent the distinct clusters, whereas the width and
opacity are proportional to stats, with the most opaque and greatest width representing the highest connection between regions in each group-
英. ROI = region of interest; STG = superior temporal gyrus; premotor = premotor cortex; PMA = primary motor area; occ = occipital lobe;
orIFG = orbital inferior frontal gyrus; opIFG = opercular IFG; MTG = middle temporal gyrus; SPL = superior parietal lobule. L = left; R = right.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Sensorimotor regions within the frontal lobe also exhibited FC increases. In older adults, 这
left primary motor cortex showed an age-related enlargement of ipsilateral connectivity sur-
rounding frontoparietal motor areas (dorsolateral prefrontal cortex, middle frontal gyrus, 和
caudate nucleus).
Anterior frontal regions (IE。, the orbital IFG) mainly exhibited age-related FC increases in
portions of the left dorsal striatum (caudate and putamen).
相比之下, there were decreases in the coupling between canonical anterior and posterior
language regions: the left posterior STG ( Wernicke’s area) with the temporal-frontal junction
(temporal pole, inferior frontal orbital cortex, frontal pole, and pars triangularis); as well as the
Neurobiology of Language
170
Naming functional connectivity in aging
桌子 2. Task-induced seed definition
Activation Analysis
NBS Hierarchical Clustering Analysis
MNI coordinates of
peak activation (毫米)
z
y
X
−12
−72
−40
Anatomical region
Left occipital lobe
Right occipital lobe
Left posterior superior temporal
gyrus
27
−60
Right posterior superior temporal
50
gyrus
Right middle temporal gyrus
Left opercular inferior frontal
gyrus
Left superior parietal lobule
Left orbital inferior frontal
gyrus
54
−38
−22
−30
−80
−39
−34
−24
6
−66
26
Right orbital inferior frontal
33
27
gyrus
Right insula
Left primary motor area
Right primary motor area
Left premotor cortex
34
−44
50
−60
18
−12
−4
2
Cluster
尺寸
7118
6920
65
70
126
94
159
336
81
49
249
242
133
T peak
价值
9
7
12
9
10
10
8
11
10
9
12
11
8
F(56)
35.1
35.8
38.8
39.0
30.7
30.1
14.0
38.8
32.2
18.0
54.9
47.3
27.3
尺寸
9
Visual
Posterior superior
temporal gyrus
Fronto-parietal
Frontal anterior
Sensorimotor
7
12
9
10
10
8
11
10
9
12
11
8
−10
6
8
−2
30
36
2
−2
8
36
39
21
Bold characters show the activation peaks selected as seeds, the most functionally relevant to the corresponding canonical functional network.
left opIFG (Broca’s area) with the posterior inferior temporal regions (fusiform gyrus). The left
visual occipital regions also showed decreased interaction with the occipital pole and superior
frontal regions.
As anticipated, the DMN showed marked age-related differences in FC. There was de-
creased coupling between the PCC and frontal portions of the DMN (medial prefrontal cortex,
bilateral paracingulate cortex, bilateral middle and superior frontal gyrus, bilateral superior
parietal lobule). 反过来, connectivity increased with nearby posterior regions, 大多数
which do not correspond to the traditional description of the DMN (angular gyrus, supramar-
ginal gyrus, superior lateral occipital cortex, superior parietal lobule).
数字 2 and Supplementary Material 1, illustrate a general trend, across both DMN and
task-induced ROIs, for reduced long-range and increased short-range connectivity in older
adults. 例如, the left superior parietal lobule seed mainly exhibited age-related in-
creases in FC with surrounding parietal and medial regions (dorsal striatum) but decreases
with distant frontal and temporal areas. 相似地, the PCC showed increased coupling with
posterior parietal regions but reduced coupling with frontal (medial and lateral) regions in
老年人.
Neurobiology of Language
171
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
These patterns of differences in the regions activated by the task tend to confirm that young
and older adults rely on different neurofunctional and cognitive processes to perform the
word-production task.
The Impact of Lexical Frequency on Age-Related Differences in FC
The final goal of this study was to test whether there was a group difference when manipulating
lexical frequency as an example of a task-specific requirement in a naming task (a condition-
by-group interaction). Within-group statistical maps of between-condition contrasts and mean
FC within age group for each lexical frequency level were computed to support the description
and interpretation of the interaction.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
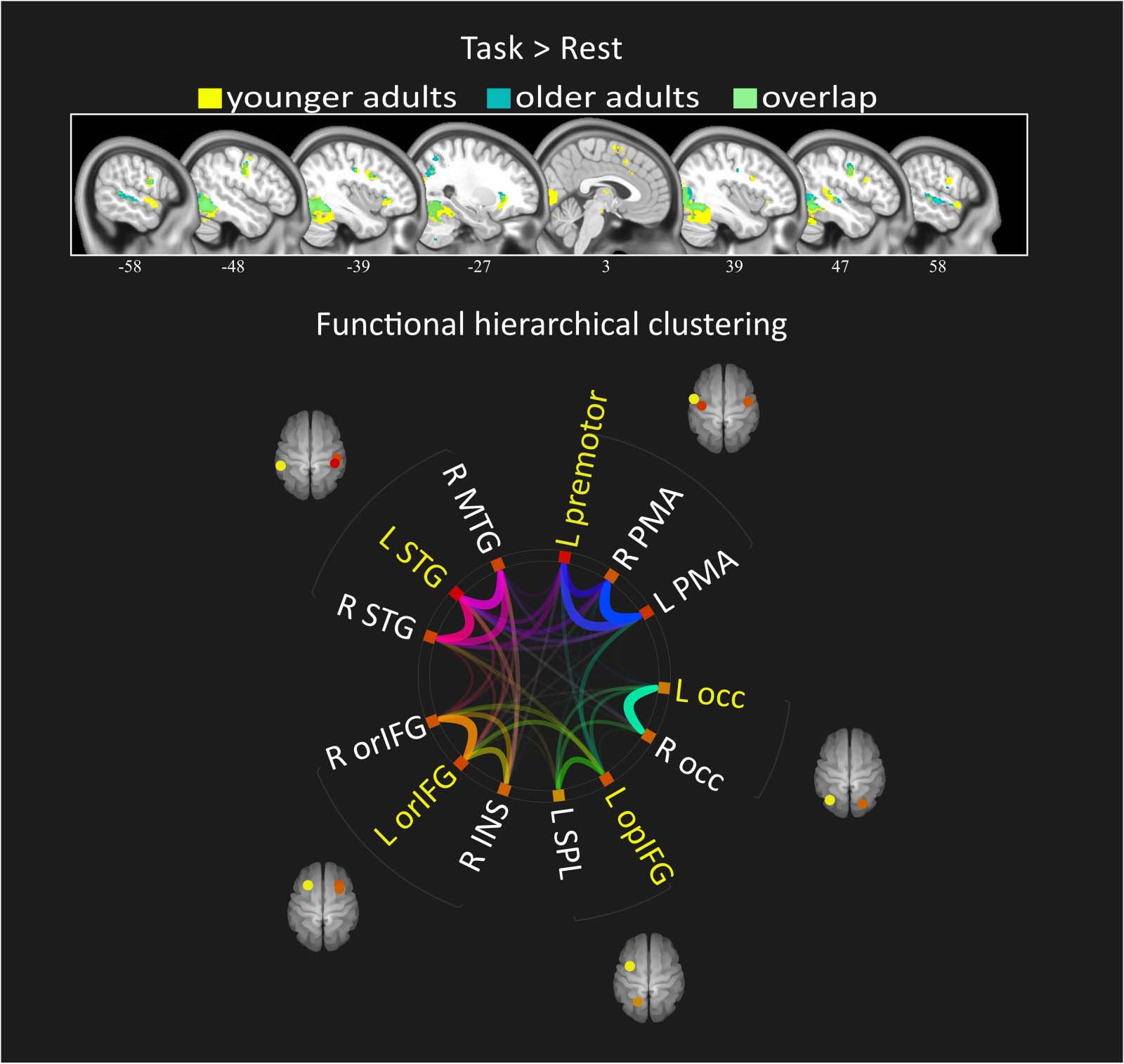
数字 2. Age-related differences in functional connectivity. Spatial maps show the seed-based con-
nectivity of six most representative ROIs activated by the task, in addition to the default mode net-
工作. For each ROI (yellow sphere) the mean functional connectivity maps are presented for the older
团体, the younger group, and the difference between age groups (OA > YA). 例如, 较老的
adults showed decreased coupling between the left posterior STG and the anterior IFG-insula region,
compared with younger adults. ROI = region of interest; OA = older adult; YA = young adult; IFG =
inferior frontal gyrus; opIFG = opercular inferior frontal gyrus; orIFG = orbital inferior frontal gyrus;
occ = occipital lobe; STG = superior temporal gyrus; PCC = posterior cingulate cortex. L = left.
Neurobiology of Language
172
Naming functional connectivity in aging
The manipulation of lexical frequency yielded a significant age-by-condition interaction for
the left occipital ROI only. Contradicting the initial expectation, there was no systematic ev-
idence of greater differences between conditions in FC in older adults than in young adults.
A simple main-effects analysis indicated double dissociations in each significant effect. 作为
预期的, older adults showed increased FC with lower lexical frequency (the more challeng-
ing condition). Young adults, on the contrary, showed anti-correlated FC between each pair of
regions in the same condition (数字 3).
The left occipital ROI showed a significant interaction in coupling with the right posterior
inferior temporal gyrus/fusiform gyrus and the right superior cerebellum. As can be seen in
Supplementary Figure 1, the right insula, which was not selected as one of the most represen-
tative seeds, also exhibited an increase in FC with the right inferior cerebellum (I and VI). 为了
后者, although the age-by-condition interaction was significant, it did not reach statistical
significance at the within-group level (main effect of condition in each group).
讨论
This study proposed a structured exploration of the FC characteristics revealed in healthy older
adults during picture naming, a cognitive ability that is central to human functioning and widely
used in clinical cognitive assessments. Most previous FC studies used the resting state to exam-
ine cognitive activity. 然而, it was felt that task-induced FC is better adapted to describe
possible changes in FC that are specific to the cognitive activity. By using seed-based FC during
picture naming, while regressing the task effect and carefully controlling for major confounds,
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
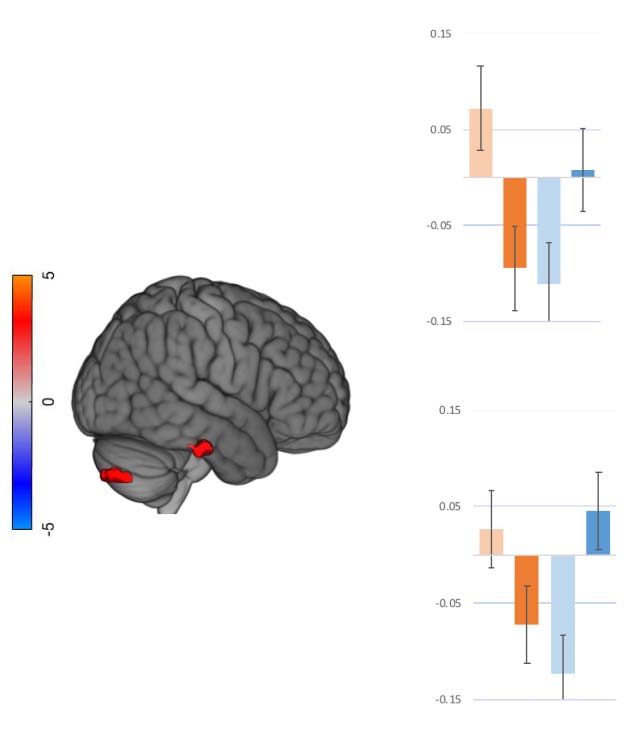
数字 3.
Interaction between age and lexical frequency: Spatial maps and mean FC values of
interaction between age and lexical frequency on task-induced FC. Spatial maps (on the left) 展示
the significant interaction effect, FWE thresholded. Bars and whiskers on the right denote FC effect
size for each group (younger and older adults) in each condition (high and low lexical frequency).
Across-group differences in FC effect sizes are given above the bars. FC = functional connectivity;
FWE = family-wise error; occ = occipital lobe; ITG = ; L = left; R = right.
Neurobiology of Language
173
Naming functional connectivity in aging
this study confirms some of the initial hypotheses but also expands upon prior knowledge sur-
rounding word production in aging.
In brief, the results show that, although the brain topography of picture naming is very similar
in younger and older adults, the FC architecture underlying naming exhibits significant differ-
恩塞斯. In older adults, there is less coupling of the traditionally described core structures of
language acquisition (例如, Broca’s and Wernicke’s areas), along with enhanced FC in regions
involved in both semantic retrieval and motor control. This study also provides the first evi-
dence that intrinsic task psychometric characteristics (IE。, lexical frequency) do not interact
much with the age-FC relationship. 一起, the findings confirm the mutual interest of
task-induced FC over a resting-state paradigm to study brain organization in healthy aging.
A Common Naming Activation Network
By choosing a sample- and task-specific template, this study proposed a neurofunctional ex-
ploration as similar as possible to the actual cognitive activity (Dickie et al., 2017; Geerligs
等人。, 2017; Salehi et al., 2018; Salehi, Karbasi, Scheinost, & Constable, 2017). As anticipated,
the brain activation analysis yielded a large fronto-temporo-occipital network of higher activity
during naming. This statistical parametric map is in line with the most prominent model of
word processing (Hickok & Poeppel, 2007). Typical ventral semantic and dorsal phonological
streams were revealed by brain activation studies (Baciu et al., 2016; Price, 2012) and brain
stimulation mapping (Duffau et al., 2014) in younger adults, and these findings were replicated
in studies of FC in aging (Hoyau et al., 2018; Indefrey, 2011). 而且, the activation peaks
were spatially and functionally coherent with the canonical functional networks that could be
expected to be active during a naming task, such as the visual, sensorimotor, salience, 和
anterior and posterior language networks.
The task-related network involved the same regions across age groups, except for the right
inferior cerebellum, which was more active in older than in younger adults. It is now recog-
nized that the cerebellum is engaged in the processing of complex cognitive material (Bernard
& Seidler, 2014; Keren-Happuch, 陈, Ho, & 德斯蒙德, 2014; Stoodley, Valera, &
Schmahmann, 2012). 尤其, the right inferior cerebellum lobule VIII has been described
as the “sensorimotor cerebellum” because it is involved in overt motor processing (articula-
的) and phonological storage (陈 & 德斯蒙德, 2005). This may suggest increased phono-
logical activity in older adults.
Some studies of word production previously reported increases in BOLD signals in parietal,
frontal, or temporal regions with age (例如, Hoyau et al., 2017; Meunier et al., 2014), 也
contralateral recruitment (例如, La et al., 2016; Meinzer et al., 2012), to support performance. A
recent meta-analysis focusing on semantic cognition and aging (Hoffman & Morcom, 2018)
concluded that increased activation was principally observed when older adults performed
worse than younger ones, 例如, during tasks drawing on executive functions. 虽然
examples of difficulties during naming are common in the literature (Feyereisen, 1997; Goulet
等人。, 1994; Shafto et al., 2017), mean accuracy is generally preserved in our study, in line with
previous reports that examined accuracy with untimed tasks (LaBarge et al., 1986; Salthouse,
2014; Schmitter-Edgecombe et al., 2000; Verhaegen & Poncelet, 2013; Wierenga et al., 2008).
In the context of preserved cognitive performance, 然后, the absence of such “compensatory” ac-
tivity is not surprising. It should be mentioned that word production is not systematically preserved
with age (伯克 & Shafto, 2004) 和, although the causes are still under study (Facal, Juncos-
Rabadán, Rodríguez, & Pereiro, 2012; 施瓦茨 & 弗雷泽, 2005; Shafto, 伯克, Stamatakis,
Tam, & Tyler, 2007), the impact of task demand will require further investigation.
Neurobiology of Language
174
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
Age-Related Functional Connectivity Differences
The main question raised in this study was whether there were differences in the FC architec-
ture during word production between younger and older healthy adults. 全面的, a common
FC architecture is observed across age groups during the task. Small yet significant, and po-
tentially functionally meaningful, differences were observed: Both increases and decreases
were seen in task-activated regions as well as in the DMN.
第一的, there is a general tendency for older adults to reduce long-range and increase short-
range connectivity. This pattern may play a functional role and reflect the need for more local
processing of neuronal information with age, at the expense of long-distance connectivity (看
Sala-Llonch et al., 2015, for a review), in line with the idea that functional organization be-
comes more segregated with age (Cao et al., 2014; 费雷拉 & Busatto, 2013; Tomasi &
Volkow, 2012). 或者, this pattern may be an unfortunate consequence of head move-
蒙特 (力量, 施拉加尔, & 彼得森, 2015; Van Dijk, 肥皂, & 巴克纳, 2012). 虽然
overt naming is deemed to be similar to natural/clinical conditions, it does induce consider-
able task-related motion. Even though many precautionary and corrective measures were taken
during the acquisition and preprocessing steps, some task-related movement remains. No stan-
dard procedure has yet been shown to unequivocally eliminate the confounding factor of
movement (巴克纳, 克里宁, & 杨, 2013); 然而, scrubbing—as used in this study—is
an efficient method of reducing related artifacts (力量, 巴恩斯, 斯奈德, 施拉加尔, &
彼得森, 2012; Yan et al., 2013). Further investigation in various task contexts and a better
comprehension of the many structural, molecular, or physiological changes that occur in
healthy aging will be required to determine which interpretation is most valid.
Functional Connectivity Differences in Task-Activated Regions
Earlier studies of FC in aging generally reported expansion of the system involved in word
processing at a younger age (Agarwal et al., 2016; Hoyau et al., 2018; La et al., 2016;
Marsolais et al., 2014). Our results suggest that older adults use similar circuits but also rely
on multiple different mechanisms during task performance.
第一的, the connectivity of the task-activated regions decreases along a fronto-temporo-
occipital pathway (IFG and posterior STG, posterior occipital, and superior frontal gyrus), rem-
iniscent of the semantic ventral stream traditionally described in younger adults (Duffau, 2015;
Hickok & Poeppel, 2007). Although frontotemporal connections have long been considered
crucial for word production, findings emerging from brain surgery, stimulation, and multi-
modal imagery in young adults suggest that their role is multimodal rather than specific
(Binder, Desai, 格雷夫斯, & Conant, 2009; Chao, Haxby, & 马丁, 1999; Duffau, 2015; Etard
等人。, 2000; Simons, Koutstaal, 王子, 瓦格纳, & Schacter, 2003; Tyler, 张, Devereux,
& 克拉克, 2013). Inferior-frontal to posterior-temporal connections—which show less coacti-
vation in older adults—are, 例如, involved during tasks that require high semantic con-
控制 (Duffau, 2015). This finding is in line with accumulated evidence that older adults rely less
on executive control—and more on semantic retrieval—when performing a word production
任务, as demonstrated using univariate activation analysis (Ansado, Marsolais, Methqal, Alary,
& Joanette, 2013; Baciu et al., 2016; Diaz, Rizio, & Zhuang, 2016; Hoyau et al., 2017;
Lacombe, Jolicoeur, Grimault, Pineault, & Joubert, 2015; Marsolais et al., 2014; Methqal,
Marsolais, Wilson, Monchi, & Joanette, 2019) and dynamic FC (Hoyau et al., 2018).
一起, these findings suggest that older adults depend less than younger adults on seman-
tic control abilities to perform the picture-naming task and possibly rely on automatized pro-
cesses instead.
Neurobiology of Language
175
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
A decrease in frontoparietal coupling (right insula, supramarginal gyrus, and parietal
operculum) is in line with earlier assertions that increased FC in domain-general systems
is not required for everyday language functioning (坎贝尔 & Tyler, 2018) and contrasts
with the general expectation that decreased resource allocation is a characteristic of aging
(公园 & Reuter-Lorenz, 2009). The insula and the supramarginal gyrus are structurally con-
nected (Ghaziri et al., 2017) and are part of control networks that guide directed attention
(Dosenbach et al., 2007; Menon & Uddin, 2010; Seeley et al., 2007; Zabelina & Andrews-
Hanna, 2016). Their deactivation during speech was previously associated with top-down
regulation of auditory attention, like that induced in a noisy environment (Elmer, 迈耶,
Marrama, & Jäncke, 2011), possibly to maximize the somatosensory feedback when speech
production becomes error-prone (Golfinopoulos et al., 2011; Seghier et al., 2015).
最后, another striking and unanticipated age-related feature is the tendency for in-
creased coupling of functional brain activity within and between motor regions responsible
for higher-level phonological processing—for example, significant increases in surrounding
ipsilateral activity in the frontal motor regions in charge of articulation control and initia-
的, such as the left dorsolateral frontal cortex and middle frontal gyrus (Duffau, 2015;
Guenther, 2016; 明智的, Greene, Büchel, & 斯科特, 1999). The pattern is coherent with pre-
viously described indirect connectivity along the dorsal phonological stream between the
primary motor and the opercula IFG (Broca’s area) via the premotor cortex (马古利斯,
Böttger, Watanabe, & Gorgolewski, 2013). The primary motor cortex also shows increased
coupling with the caudate nucleus, which plays a role in the control of articulation
(Argyropoulos, Tremblay, & 小的, 2013; Duffau et al., 2014; Guenther, 2016; 马古利斯
等人。, 2013; Tremblay, Deschamps, & Gracco, 2016). In summation, more functional re-
organization and less segregation can be seen along the dorsal phonological stream with
increased age (Agarwal et al., 2016; Diaz et al., 2016; 马丁斯, Simard, & Monchi, 2014;
Muller et al., 2016; Sörös, Bose, Sokoloff, Graham, & 愚蠢的, 2011), in line with the hypoth-
esized weakening of the links between phonological and lexical representations (伯克
等人。, 1991).
Functional Connectivity Differences in the DMN
In accordance with previous reports, the DMN showed an age-related increase with other pa-
rietal regions during the task (Chan et al., 2017; Geerligs et al., 2014; Grady et al., 2016; Mak
等人。, 2017; Spreng, Stevens, Viviano, & Schacter, 2017). This kind of mechanism has been
suggested to indicate a broad dedifferentiation of neural activity in later life (例如, 坎贝尔,
Grady, 的, & Hasher, 2012). 或者, the interaction between the DMN and task com-
ponents may indicate active strategic cognitive processes. According to the default-executive
coupling hypothesis of aging (DECHA) 模型 (车工 & Spreng, 2015), aging is characterized
by a semanticization process: As cognitive control resources decline, cognitive behavior be-
comes more and more influenced by past experiences and knowledge. The DMN therefore
becomes increasingly engaged with task components to support task performance. 虽然
Turner and Spreng specifically described increases in DMN-prefrontal lateral coupling, 他们
used cognitive tasks that rely partly on executive processes. 实际上, the increase in DMN ac-
tivity has also been demonstrated with other regions (Damoiseaux et al., 2008; Sambataro
等人。, 2010) and the DECHA might therefore be extended to any region involved in the task
in relation to the posterior DMN. The posterior DMN has been specifically described as a key
actor in semantic retrieval processes, 事实上, the DMN and the semantic system share an
overlapping functional network (Binder et al., 2009; Bonnelle et al., 2012; Krieger-Redwood
等人。, 2016).
Neurobiology of Language
176
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
The Impact of Lexical Frequency Manipulation
Behavioral analysis showed that there is an interaction between age, 频率, and RT, 哪个
suggests that lower frequency does make the cognitive activity more demanding for older
adults, even though they still manage to accomplish the task. Lexical frequency did not influ-
ence accuracy scores in either age group. More segregated frequency levels may have allowed
for a stronger task-demand effect (Moberg, 费拉罗, & Petros, 2000), and more robust conclu-
sions will require the manipulation of task demands through more than two levels.
Although exploratory, our results show a few regions that are significantly mediated by in-
trinsic task requirement according to age. With greater task demand, integration increases in
老年人, whereas it decreases in young adults. Previous studies with other types of task
load interpreted this pattern as an adaptive form of compensation, beneficial to performance
(例如, Crowell et al., 2019; Nagels et al., 2012; 车工 & Spreng, 2015). Those studies, 和
others that presented age-related compensation patterns (Cabeza, 2002; Campbell et al.,
2012; Grady et al., 2016; Morcom & Henson, 2018; 公园 & Reuter-Lorenz, 2009; Rajah &
D’Esposito, 2005; 车工 & Spreng, 2015), advocated for the notion that there are domain-
general mechanisms, expressed mostly through increased activity in prefrontal regions. 这
hypothesis builds upon the underlying assumption that older adults find just about any task
more difficult than young adults, because of a general cognitive decline (Craik & Byrd,
1982). 然而, domain-general upregulation was mostly reported using tasks that represent
a strong cognitive challenge for older adults (Hoffman & Morcom, 2018). To our knowledge,
only one other study previously explored the impact of linguistic criteria on verbal fluency
(Marsolais et al., 2014); it showed an interaction between age and increased FC in posterior
地区. Using an intrinsic—linguistic—task demand, rather than general cognitive load, 我们的
findings are also in line with the concept of task-specific mechanisms (坎贝尔 & Schacter,
2016; 坎贝尔 & Tyler, 2018; Hearne, Cocchi, 扎莱斯基, & Mattingley, 2017; Peelle, 2019;
Samu et al., 2017). Campbell and Tyler (2018) suggested that domain-general networks should
not be considered as stable, singular mechanisms. 因此, neurofunctional patterns invoked
during a task may instead reflect processes that are specific to that task.
In this study, age differences between conditions are small in both size and number.
此外, the FC of the DMN was not significantly affected by lower lexical frequency,
contrary to what had been reported previously in studies that manipulated executive task de-
要求 (Persson et al., 2007; Steffener, Habeck, & Stern, 2012). 反而, the interaction was
significant in regions that are functionally meaningful and specialized for the task: the left oc-
cipital cortex and fusiform gyrus, which together encode visual representations of objects
(Mahon et al., 2007; 马丁 & Chao, 2001); the fusiform gyrus, the cerebellum, and the insula,
which were previously reported to be active during the processing of low-frequency words
(Basso et al., 2013); 和, as mentioned earlier, the right inferior cerebellum lobule VIII, 哪个
is also related to phonological production (陈 & 德斯蒙德, 2005).
Greater insight into domain-general and task-specific mechanisms will be gained by large
data set studies that manipulate various cognitive states, such as those initiated by projects
such as Reference Ability Neural Network study (Stern et al., 2014) and Cambridge Centre
for Ageing and Neuroscience, or CamCAN (Taylor et al., 2015).
最后, some general methodological considerations should be raised regarding the diffi-
culty of interpreting the neurobiological meaning of linear FC changes in group comparisons.
Increased FC could either indicate decreased network adaptability to task demands (Avelar-
Pereira et al., 2017; 坎贝尔 & Schacter, 2016; Dixon et al., 2017; Dubois & Adolphs, 2016;
Geerligs et al., 2015; Grady et al., 2016) or reflect optimization for efficient (IE。, 小的)
Neurobiology of Language
177
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
network updates, reducing processing demands and supporting behavioral performance
(Schultz & Cole, 2016; Shine et al., 2016). Although this study was not designed to rule on
these options, the latter fits the current findings better: Considering the preservation of perfor-
曼斯, and the tendency for the core naming network to shrink and for the dorsal phonolog-
ical stream to suffer more from the impact of age, FC topography differences could indicate an
efficient strategy relying on a more experienced, automatized, and specialized circuit in older
adults. 或者, interindividual heterogeneity should not be underestimated: The intricate
patterns emerging from averaged differences between groups could reflect more diverse brain
physiology and functioning among older people, as has been reported frequently (例如, Amiri
等人。, 2014; Dong et al., 2012; Raz et al., 2005; Zuo et al., 2010). 此外, the task effect
was regressed in this study, because this technique had previously been demonstrated to in-
crease the reliability of task FC estimates (Cao et al., 2014; Cole, Bassett, 力量, Braver, &
彼得森, 2014). This choice may have reduced some of the true task-related effects on
FC but was felt to be preferable considering the overt nature of the naming task and knowing
that participant movement can have large effects on FC estimates (Mowinckel, Espeseth, &
Westlye, 2012).
结论
This study was designed to characterize the possible age-related reorganization of functional
connectivity associated with a word-naming task, while manipulating task demand using the
lexical frequency of the stimuli.
Word production is a basic yet essential cognitive activity central to human everyday function-
英. This study, like others before it, emphasizes that vocabulary knowledge increases throughout
the life span, into the later stages of life, even though word-production processes slow down.
在此背景下, the task-induced functional architecture of older adults, compared with
younger adults, is characterized by greater reliance on regions in charge of motor control
地区, along with lesser inclusion of regions classically associated with word production, 这样的
as the IFG and the posterior STG. Earlier descriptions of the core word-production network
were based mostly on samples of young adults and used resting-state paradigms. This study,
using a sample- and task-specific design, suggests that older adults have a more efficient strat-
egy, relying on a more experienced, specialized, and segregated, naming pathway.
This study also reinforces the notion that task-specific rather than domain-general mecha-
nisms apply in older adults. Although additional involvement of frontal and bilateral executive
regions was previously suggested to support the cognitive performance of older adults in every
domain, it appears that those findings cannot be generalized to all cognitive states or types of
task demand. 反而, older adults rely on highly specialized regions to perform more complex
linguistic processes.
Considering the current limitations affecting the interpretation of FC changes with age, fur-
ther exploration is definitely required to disentangle the complex interplay between preserved
performance and neurofunctional behavior, using various tasks and stimuli.
资金信息
Yves Joanette, Institute of Aging (http://dx.doi.org/10.13039/501100000026), 奖项ID: 191272.
Simona Maria Brambati, Canadian Network for Research and Innovation in Machining
技术, Natural Sciences and Engineering Research Council of Canada (http://dx.doi.
org/10.13039/501100002790).
Neurobiology of Language
178
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
作者贡献
Julien Jarret: 数据管理: 平等的. Simona Maria Brambati: 概念化: 配套;
方法: 配套; Writing — Review & Editing: 配套. Pierre Bellec:
方法: 配套; 监督: 配套; Writing — Review & Editing: 配套.
Yves Joanette: 概念化: 配套; 资金获取: 带领; 项目管理:
平等的; 资源: 带领; 监督: 配套; Writing — Review & Editing: 配套.
参考
阿加瓦尔, S。, Stamatakis, 乙. A。, Geva, S。, & Warburton, 乙. A. (2016).
Dominant hemisphere functional networks compensate for struc-
tural connectivity loss to preserve phonological retrieval with ag-
英. Brain and Behavior, 6, e00495. https://doi.org/10.1002/
brb3.495
Amiri, M。, Pouliot, P。, Bonnéry, C。, Leclerc, P.-O., Desjardins, M。,
Lesage, F。, & Joanette, 是. (2014). An exploration of the effect of
hemodynamic changes due to normal aging on the fNIRS re-
sponse to semantic processing of words. Frontiers in
Neurology, 5, 249. https://doi.org/10.3389/fneur.2014.00249
Andrews-Hanna, J. R。, 斯奈德, A. Z。, Vincent, J. L。, Lustig, C。, Head,
D ., Raichle, 中号. E., & 巴克纳, 右. L. (2007). Disruption of large-
scale brain systems in advanced aging. 神经元, 56, 924–935.
https://doi.org/10.1016/j.neuron.2007.10.038
Ansado, J。, Marsolais, Y。, Methqal, 我。, Alary, F。, & Joanette, 是. (2013).
The adaptive aging brain: Evidence from the preservation of com-
munication abilities with age. 欧洲神经科学杂志,
37, 1887–1895. https://doi.org/10.1111/ejn.12252
Argyropoulos, G. P。, Tremblay, P。, & 小的, S. L. (2013). The neo-
striatum and response selection in overt sentence production: 一个
fMRI study. 神经影像, 82, 53–60.
Ashburner, J. (2007). A fast diffeomorphic image registration algo-
rithm. 神经影像, 38, 95–113.
Ashburner, J。, & 弗里斯顿, K. J. (2005). Unified segmentation.
神经影像, 26, 839–851.
Au, R。, Joung, P。, 尼古拉斯, M。, Obler, L. K., Kass, R。, & 阿尔伯特,
中号. L. (1995). Naming ability across the adult life span. 老化,
Neuropsychology, 和认知, 2, 300–311. https://doi.org/
10.1080/13825589508256605
Avelar-Pereira, B., Bäckman, L。, Wåhlin, A。, Nyberg, L。, & Salami,
A. (2017). Age-related differences in dynamic interactions among
default mode, frontoparietal control, and dorsal attention networks
during resting-state and interference resolution. Frontiers in Aging
神经科学, 9, 152. https://doi.org/10.3389/fnagi.2017.00152
Baciu, M。, Boudiaf, N。, Cousin, E., Perrone-Bertolotti, M。, Pichat,
C。, Fournet, N。, … Krainik, A. (2016). Functional MRI evidence
for the decline of word retrieval and generation during normal
老化. 年龄 (多德雷赫特, 荷兰), 38, 3. https://doi.org/
10.1007/s11357-015-9857-y
Basso, G。, Magon, S。, Reggiani, F。, Capasso, R。, 莫尼托拉, G。,
哪个, F. J。, & Miceli, G. (2013). Distinguishable neurofunctional
effects of task practice and item practice in picture naming: A
BOLD fMRI study in healthy subjects. Brain and Language,
126(3), 302–313. https://doi.org/10.1016/j.bandl.2013.07.002
Behzadi, Y。, Restom, K., Liau, J。, & 刘, 时间. 时间. (2007). A component
based noise correction method (CompCor) for BOLD and perfu-
sion based fMRI. 神经影像, 37, 90–101.
Ben-David, 乙. M。, Erel, H。, Goy, H。, & 施耐德, 乙. A. (2015).
“Older is always better”: Age-related differences in vocabulary
scores across 16 年. Psychology and Aging, 30, 856–862.
https://doi.org/10.1037/pag0000051
Bernard, J. A。, & Seidler, 右. D. (2014). Moving forward: Age effects
on the cerebellum underlie cognitive and motor declines.
Neuroscience and Biobehavioral Reviews, 42, 193–207. https://
doi.org/10.1016/j.neubiorev.2014.02.011
Binder, J. R。, Desai, 右. H。, 格雷夫斯, 瓦. W., & Conant, L. L. (2009).
Where is the semantic system? A critical review and meta-analysis
的 120 functional neuroimaging studies. 大脑皮层, 19,
2767–2796. https://doi.org/10.1093/cercor/bhp055
布莱克威尔, A. D ., Sahakian, 乙. J。, Vesey, R。, Semple, J. M。, Robbins,
时间. W., & Hodges, J. 右. (2004). Detecting dementia: Novel neuro-
psychological markers of preclinical Alzheimer’s disease.
Dementia and Geriatric Cognitive Disorders, 17, 42–48.
Bonnelle, 五、, Ham, T。, 水蛭, R。, Kinnunen, K., Mehta, M。,
Greenwood, R。, & 锋利的, D. (2012). Salience network integrity
predicts default mode network function after traumatic brain in-
陪审团. 美国国家科学院院刊
美国, 109, 4690–4695. https://doi.org/
10.1073/pnas.1113455109
巴克纳, 右. L。, 克里宁, F. M。, & 杨, 乙. 时间. 时间. (2013). Opportunities
and limitations of intrinsic functional connectivity MRI. 自然
神经科学, 16, 832–837.
伯克, D. M。, MacKay, D. G。, Worthley, J. S。, & Wade, 乙. (1991). 在
the tip of the tongue: What causes word finding failures in young
and older adults? 记忆与语言杂志, 30, 542–579.
伯克, D. M。, & Shafto, 中号. A. (2004). Aging and language produc-
的. Current Directions in Psychological Science, 13, 21–24.
Cabeza, 右. (2002). Hemispheric asymmetry reduction in older
adults: The HAROLD model. Psychology and Aging, 17, 85–100.
Cabeza, R。, 阿尔伯特, M。, Belleville, S。, Craik, F. 我. M。, Duarte, A。,
Grady, C. L。, … Rajah, 中号. 氮. (2018). Maintenance, reserve
and compensation: The cognitive neuroscience of healthy age-
英. 自然评论神经科学, 19, 701–710. https://土井.
org/10.1038/s41583-018-0068-2
Cabeza, R。, Nyberg, L。, & 公园, D. C. (2016). Cognitive neurosci-
ence of aging: Linking cognitive and cerebral aging. 纽约:
牛津大学出版社. https://doi.org/10.1093/acprof:oso/
9780195156744.001.0001
坎贝尔, K. L。, Grady, C. L。, 的, C。, & Hasher, L. (2012). Age differ-
ences in the frontoparietal cognitive control network: Implications
for distractibility. Neuropsychologia, 50, 2212–2223. https://土井.
org/10.1016/J.NEUROPSYCHOLOGIA.2012.05.025
坎贝尔, K. L。, Samu, D ., 戴维斯, S. W., Geerligs, L。, Mustafa, A。,
Tyler, L. K., & Cambridge Centre for Aging and Neuroscience.
(2016). Robust resilience of the frontotemporal syntax system
to aging. 神经科学杂志, 36, 5214–5227. https://土井.
org/10.1523/JNEUROSCI.4561-15.2016
坎贝尔, K. L。, & Schacter, D. L. (2016). Aging and the resting state:
Is cognition obsolete? 语言, Cognition and Neuroscience, 32,
661–668. https://doi.org/10.1016/j.bbi.2017.04.008
坎贝尔, K. L。, & Tyler, L. K. (2018). Language-related domain-
specific and domain-general systems in the human brain.
Neurobiology of Language
179
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
Current Opinion in Behavioral Sciences, 21, 132–137. https://
doi.org/10.1016/j.cobeha.2018.04.008
曹, M。, 王, J. H。, Dai, Z. J。, 曹, X. Y。, Jiang, L. L。, Fan, F. M。,
… He, 是. (2014). Topological organization of the human brain
functional connectome across the lifespan. Developmental
Cognitive Neuroscience, 7, 76–93. https://doi.org/10.1016/j.
dcn.2013.11.004
Chan, 中号. Y。, Alhazmi, F. H。, 公园, D. C。, Savalia, 氮. K., & 假发,
G. S. (2017). Resting-state network topology differentiates task
signals across the adult life span. 神经科学杂志, 37,
2734–2745. https://doi.org/10.1523/JNEUROSCI.2406-16.2017
Chao, L. L。, Haxby, J. 五、, & 马丁, A. (1999). Attribute-based neu-
ral substrates in temporal cortex for perceiving and knowing
about objects. 自然神经科学, 2, 913–919.
陈, S. H. A。, & 德斯蒙德, J. 乙. (2005). Temporal dynamics of
cerebro-cerebellar network recruitment during a cognitive task.
Neuropsychologia, 43, 1227–1237.
Cole, 中号. W., Bassett, D. S。, 力量, J. D ., Braver, 时间. S。, & 彼得森,
S. 乙. (2014). Intrinsic and task-evoked network architectures of
the human brain. 神经元, 83, 238–251. https://doi.org/
10.1016/j.neuron.2014.05.014
Cole, D. M。, 史密斯, S. M。, & 贝克曼, C. F. (2010). Advances and
pitfalls in the analysis and interpretation of resting-state FMRI
数据. Frontiers in Systems Neuroscience, 4, 8. https://doi.org/
10.3389/fnsys.2010.00008
Condret-Santi, 五、, Barbeau, 乙. J。, Matharan, F。, Le Goff, M。,
Dartigues, J.-F., & Amieva, H. (2013). Prevalence of word re-
trieval complaint and prediction of dementia in a population-
based study of elderly subjects. Dementia and Geriatric
Cognitive Disorders, 35(5–6), 313–321. https://doi.org/10.1159/
000342594
Craik, F. 我. M。, & Byrd, 中号. (1982). Aging and cognitive deficits. 在
F. 我. 中号. Craik & S. Trehub (编辑。), Aging and cognitive processes
(PP. 191–211). 波士顿, 嘛: Springer US. https://doi.org/10.
1007/978-1-4684-4178-9_11
Crowell, C. A。, 戴维斯, S. W., Beynel, L。, Deng, L。, Lakhlani, D .,
Hilbig, S. A。, … Cabeza, 右. (2019). Older adults benefit from
more widespread brain network integration during working
记忆. BioRxiv, 642447. https://doi.org/10.1101/642447
Damoiseaux, J. S。, 贝克曼, C. F。, Arigita, 乙. J. S。, Barkhof, F。,
Scheltens, P。, 斯塔姆, C. J。, … Rombouts, S. A. 右. 乙. (2008).
Reduced resting-state brain activity in the “default network” in
normal aging. 大脑皮层, 18, 1856–1864. https://doi.org/
10.1093/cercor/bhm207
戴维斯, S. W., 丹尼斯, 氮. A。, Daselaar, S. M。, Fleck, 中号. S。, &
Cabeza, 右. (2008). Qué pasa? The posterior-anterior shift in ag-
英. 大脑皮层, 18, 1201–1209.
戴维斯, S. W., 斯坦利, 中号. L。, Moscovitch, M。, & Cabeza, 右. (2017).
Resting-state networks do not determine cognitive function net-
作品: A commentary on Campbell and Schacter (2016).
语言, Cognition and Neuroscience, 32, 669–673.
Diaz, 中号. T。, Rizio, A. A。, & Zhuang, J. (2016). The neural language
systems that support healthy aging: Integrating function, struc-
真实, and behavior. Language and Linguistics Compass, 10,
314–334. https://doi.org/10.1111/lnc3.12199
Dickie, D. A。, Shenkin, S. D ., Anblagan, D ., 李, J。, Blesa Cabez, M。,
罗德里格斯, D ., … Wardlaw, J. 中号. (2017). Whole brain magnetic res-
onance image atlases: A systematic review of existing atlases and
caveats for use in population imaging. 神经信息学前沿,
11, 1. https://doi.org/10.3389/fninf.2017.00001
狄克逊, 中号. L。, Andrews-Hanna, J. R。, Spreng, 右. N。, Irving, Z. C。,
米尔斯, C。, Girn, M。, & Christoff, K. (2017). Interactions between
the default network and dorsal attention network vary across
default subsystems, 时间, and cognitive states. 神经影像, 147,
632–649. https://doi.org/10.1016/J.NEUROIMAGE.2016.12.073
Dong, L。, 沉, Y。, Lei, X。, Luo, C。, 李, Q.-W., 吴, W.-Y., … Li,
C.-B. (2012). The heterogeneity of aging brain: Altered functional
connectivity in default mode network in older adults during ver-
bal fluency tests. Chinese Medical Journal, 125, 604–610. https://
doi.org/10.3760/cma.j.issn.0366-6999.2012.04.009
Dosenbach, 氮. U. F。, Fair, D. A。, 米津, F. M。, 科恩, A. L。,
Wenger, K. K., Dosenbach, 右. A. T。, … Raichle, 中号. 乙. (2007).
Distinct brain networks for adaptive and stable task control in hu-
芒斯. 美国国家科学院院刊, 104,
11073–11078.
Dubois, J。, & Adolphs, 右. (2016). Building a science of individual
differences from fMRI. 认知科学的趋势, 20, 425–443.
https://doi.org/10.1016/j.tics.2016.03.014
Duffau, H. (2015). Stimulation mapping of white matter tracts to
study brain functional connectivity. Nature Reviews Neurology,
11, 255–265. https://doi.org/10.1038/nrneurol.2015.51
Duffau, H。, Moritz-Gasser, S。, & Mandonnet, 乙. (2014). A re-
examination of neural basis of language processing: Proposal of
a dynamic hodotopical model from data provided by brain stim-
ulation mapping during picture naming. Brain and Language,
131, 1–10. https://doi.org/10.1016/j.bandl.2013.05.011
Eickhoff, S. B., Stephan, K. E., Mohlberg, H。, Grefkes, C。, Fink, G. R。,
Amunts, K., & Zilles, K. (2005). A new SPM toolbox for combining
probabilistic cytoarchitectonic maps and functional imaging data.
神经影像, 25, 1325–1335.
Elmer, S。, 迈耶, M。, Marrama, L。, & Jäncke, L. (2011). Intensive
language training and attention modulate the involvement of
fronto-parietal regions during a non-verbal auditory discrimina-
tion task. 欧洲神经科学杂志, 34, 165–175.
Etard, 奥。, Mellet, E., Papathanassiou, D ., Benali, K., Houdé, 奥。,
Mazoyer, B., & Tzourio-Mazoyer, 氮. (2000). Picture naming
without Broca’s and Wernicke’s area. NeuroReport, 11, 617–622.
https://doi.org/10.1097/00001756-200002280-00036
Facal, D ., Juncos-Rabadán, 奥。, Rodríguez, 中号. S。, & Pereiro, A. X.
(2012). Tip-of-the-tongue in aging: Influence of vocabulary,
working memory and processing speed. Aging Clinical and
Experimental Research, 24, 647–656.
Ferré, P。, Benhajali, Y。, Steffener, J。, Stern, Y。, Joanette, Y。, & Bellec,
磷. (2019). Resting-state and vocabulary tasks distinctively inform
on age-related differences in the functional brain connectome.
语言, Cognition and Neuroscience, 34, 949–972. https://
doi.org/10.1080/23273798.2019.1608072
费雷拉, L. K., & Busatto, G. F. (2013). Resting-state functional
connectivity in normal brain aging. Neuroscience and
Biobehavioral Reviews, 37, 384–400. https://doi.org/10.1016/
j.neubiorev.2013.01.017
费雷拉, L. K., Regina, A. C. B., Kovacevic, N。, 马丁, 中号. da G. M。,
Santos, 磷. P。, Carneiro, C. de G., … Busatto, G. F. (2016). 老化
effects on whole-brain functional connectivity in adults free of
cognitive and psychiatric disorders. 大脑皮层, 26, 3851–3865.
https://doi.org/10.1093/cercor/bhv190
Feyereisen, 磷. (1997). A meta-analytic procedure shows an age-
related decline in picture naming: Comments on Goulet, Ska, 和
Kahn (1994). Journal of Speech, 语言, and Hearing Research,
40, 1328–1333. https://doi.org/10.1044/jslhr.4006.1328
Feyereisen, P。, Demaeght, N。, & Samson, D. (1998). Why do picture
naming latencies increase with age: General slowing, greater sensi-
tivity to interference, or task-specific deficits? Experimental Aging
研究, 24(1), 21–51. https://doi.org/10.1080/036107398244346
芬恩, 乙. S。, Scheinost, D ., 芬恩, D. M。, 沉, X。, Papademetris,
X。, & Constable, 右. 时间. (2017). Can brain state be manipulated
Neurobiology of Language
180
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
to emphasize individual differences in functional connectivity?
神经影像, 160, 140–151. https://doi.org/10.1016/J.NEUROIMAGE.
2017.03.064
Fjell, A. M。, & Walhovd, K. 乙. (2016). Multi-modal imaging of the
aging brain. 在R中. Cabeza, L. Nyberg, & D. C. 公园 (编辑。),
Cognitive neuroscience of aging: Linking cognitive and cerebral
老化 ( 卷. 2, PP. 359–379). 纽约: 牛津大学出版社.
Forster, K. 我。, & Forster, J. C. (2003). DMDX: A Windows display
program with millisecond accuracy. Behavior Research
方法, Instruments, and Computers, 35, 116–124.
Friederici, A. D ., & Gierhan, S. 中号. 乙. (2013). The language network.
神经生物学的当前观点, 23, 250–254. https://doi.org/
10.1016/j.conb.2012.10.002
Geerligs, L。, & Henson, 右. 氮. (2016). Functional connectivity and
structural covariance between regions of interest can be mea-
sured more accurately using multivariate distance correlation.
神经影像, 135, 16–31.
Geerligs, L。, Maurits, 氮. M。, Renken, 右. J。, & Lorist, 中号. 中号. (2014).
Reduced specificity of functional connectivity in the aging brain
during task performance. 人脑图谱, 35, 319–330.
https://doi.org/10.1002/hbm.22175
Geerligs, L。, 鲁比诺夫, M。, & Henson, 右. 氮. (2015). State and trait com-
ponents of functional connectivity: Individual differences vary with
mental state. 神经科学杂志, 35, 13949–13961.
Geerligs, L。, Tsvetanov, K. A。, & Henson, 右. 氮. (2017). 挑战
in measuring individual differences in functional connectivity
using fMRI: The case of healthy aging. 人脑图谱,
38, 4125–4156. https://doi.org/10.1002/hbm.23653
Ghaziri, J。, Tucholka, A。, Girard, G。, Houde, J.-C., Boucher, 奥。,
吉尔伯特, G。, … Nguyen, D. K. (2017). The corticocortical struc-
tural connectivity of the human insula. 大脑皮层, 27,
1216–1228. https://doi.org/10.1093/cercor/bhv308
Golfinopoulos, E., Tourville, J. A。, Bohland, J. W., 戈什, S. S。,
Nieto-Castanon, A。, & Guenther, F. H. (2011). fMRI investigation
of unexpected somatosensory feedback perturbation during
speech. 神经影像, 55, 1324–1338. https://doi.org/10.1016/J.
NEUROIMAGE.2010.12.065
古德格拉斯, H。, 卡普兰, E., & Weintraub, S. (1983). Boston naming
测试. 费城, PA: Lea & Febiger.
Goulet, P。, Ska, B., & Kahn, H. J. (1994). Is there a decline in picture
naming with advancing age? Journal of Speech and Hearing
研究, 37, 629–644. https://doi.org/10.1044/jshr.3703.629
Grady, C。, Sarraf, S。, Saverino, C。, & 坎贝尔, K. (2016). Age differ-
ences in the functional interactions among the default, frontoparietal
控制, and dorsal attention networks. Neurobiology of Aging, 41,
159–172. https://doi.org/10.1016/j.neurobiolaging.2016.02.020
Greene, A. S。, 高, S。, Scheinost, D ., & Constable, 右. 时间. (2018).
Task-induced brain state manipulation improves prediction of in-
dividual traits. Nature Communications, 9, 2807. https://doi.org/
10.1038/s41467-018-04920-3
Guenther, F. H. (2016). Neural models of motor speech control.
In G. Hickok & S. L. 小的 (编辑。), Neurobiology of language
(PP. 725–740). 伦敦: 学术出版社. https://doi.org/10.
1016/ B978-0-12-407794-2.00058-4
Hearne, L. J。, Cocchi, L。, 扎莱斯基, A。, & Mattingley, J. 乙. (2017).
Reconfiguration of brain network architectures between resting-
state and complexity-dependent cognitive reasoning. 杂志
神经科学, 37, 8399–8411. https://doi.org/10.1523/
JNEUROSCI.0485-17.2017
Hickok, G。, & Poeppel, D. (2007). The cortical organization of
speech processing. 自然, 8, 393–402.
Hoffman, P。, & Morcom, A. 中号. (2018). Age-related changes in the
neural networks supporting semantic cognition: A meta-analysis
的 47 functional neuroimaging studies. Neuroscience and
Biobehavioral Reviews, 84, 134–150. https://doi.org/10.1016/J.
NEUBIOREV.2017.11.010
Hoyau, E., Boudiaf, N。, Cousin, E., Pichat, C。, Fournet, N。, Krainik,
A。, … Baciu, 中号. (2017). Aging modulates the hemispheric special-
ization during word production. Frontiers in Aging Neuroscience,
9, 125. https://doi.org/10.3389/fnagi.2017.00125
Hoyau, E., Roux-Sibilon, A。, Boudiaf, N。, Pichat, C。, Cousin, E.,
Krainik, A。, … Baciu, 中号. (2018). Aging modulates fronto-temporal
cortical interactions during lexical production. A dynamic causal
modeling study. Brain and Language, 184, 11–19. https://doi.org/
10.1016/j.bandl.2018.06.003
Indefrey, 磷. (2011). The spatial and temporal signatures of word
production components: A critical update. Frontiers in
心理学, 2, 255.
Indefrey, P。, & Levelt, 瓦. J. 中号. (2004). The spatial and temporal
signatures of word production components. 认识, 92,
101–144.
Jackson, 右. L。, Hoffman, P。, Pobric, G。, & Lambon Ralph, 中号. A.
(2015). The nature and neural correlates of semantic association
versus conceptual similarity. 大脑皮层, 25, 4319–4333.
https://doi.org/10.1093/cercor/bhv003
Keren-Happuch, E., 陈, S.-H. A。, Ho, M.-H. R。, & 德斯蒙德, J. 乙.
(2014). A meta-analysis of cerebellar contributions to higher cog-
nition from PET and fMRI studies. 人脑图谱, 35,
593–615.
孔, R。, 李, J。, Orban, C。, 肥皂, 中号. R。, 刘, H。, Schaefer, A。, ……
杨, 乙. 时间. 时间. (2018). Spatial topography of individual-specific
cortical networks predicts human cognition, 性格, 和
情感. 大脑皮层, 29, 2533–2551. https://doi.org/10.1093/
cercor/bhy123
Krieger-Redwood, K., Jefferies, E., Karapanagiotidis, T。, Seymour,
R。, Nunes, A。, Ang, J. 瓦. A。, … Smallwood, J. (2016). 向下
but not out in posterior cingulate cortex: Deactivation yet func-
tional coupling with prefrontal cortex during demanding seman-
tic cognition. 神经影像, 141, 366–377. https://doi.org/
10.1016/j.neuroimage.2016.07.060
这, C。, Garcia-Ramos, C。, Nair, V. A。, Meier, 时间. B., Farrar-Edwards,
D ., Birn, R。, … Prabhakaran, V. (2016). Age-related changes in
BOLD activation pattern in phonemic fluency paradigm: An in-
vestigation of activation, functional connectivity and psycho-
physiological interactions. Frontiers in Aging Neuroscience, 8,
110. https://doi.org/10.3389/fnagi.2016.00110
LaBarge, E., 爱德华兹, D ., & Knesevich, J. 瓦. (1986). Performance
of normal elderly on the Boston Naming Test. Brain and
语言, 27, 380–384. https://doi.org/10.1016/0093-934X(86)
90026-X
Lacombe, J。, Jolicoeur, P。, Grimault, S。, Pineault, J。, & Joubert, S.
(2015). Neural changes associated with semantic processing in
healthy aging despite intact behavioral performance. Brain and
语言, 149, 118–127. https://doi.org/10.1016/j.bandl.
2015.07.003
LaGrone, S。, & Spieler, D. H. (2006). Lexical competition and pho-
nological encoding in young and older speakers. Psychology and
老化, 21, 804–809. https://doi.org/10.1037/0882-7974.
21.4.804
Liljeström, M。, 史蒂文森, C。, Kujala, J。, & Salmelin, 右. (2015).
任务- and stimulus-related cortical networks in language produc-
的: Exploring similarity of MEG- and fMRI-derived functional
连接性. 神经影像, 120, 75–87. https://doi.org/10.1016/
j.neuroimage.2015.07.017
Mahon, 乙. Z。, Milleville, S. C。, Negri, G. A. L。, Rumiati, 右. 我。,
Caramazza, A。, & 马丁, A. (2007). Action-related properties
Neurobiology of Language
181
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
shape object representations in the ventral stream. 神经元, 55,
507–520.
Mak, L. E., Minuzzi, L。, MacQueen, G。, 大厅, G。, 肯尼迪, S. H。, &
Milev, 右. (2017). The default mode network in healthy individ-
乌尔斯: A systematic review and meta-analysis. Brain Connectivity,
7, 25–33. https://doi.org/10.1089/brain.2016.0438
马古利斯, D. S。, Böttger, J。, Watanabe, A。, & Gorgolewski, K. J.
(2013). Visualizing the human connectome. 神经影像, 80,
445–461. https://doi.org/10.1016/j.neuroimage.2013.04.111
Marsolais, Y。, Perlbarg, 五、, Benali, H。, & Joanette, 是. (2014). 年龄-
related changes in functional network connectivity associated
with high levels of verbal fluency performance. Cortex, 58,
123–138. https://doi.org/10.1016/j.cortex.2014.05.007
马丁, A。, & Chao, L. L. (2001). Semantic memory and the brain:
Structure and processes. 神经生物学的当前观点, 11,
194–201.
马丁斯, R。, Simard, F。, & Monchi, 氧. (2014). Differences between
patterns of brain activity associated with semantics and those
linked with phonological processing diminish with age. PLOS
一, 9, e99710. https://doi.org/10.1371/journal.pone.0099710
Mazaika, 磷. K., Hoeft, F。, Glover, G. H。, & Reiss, A. L. (2009).
Methods and software for fMRI analysis of clinical subjects.
神经影像, 47, S58.
Mazaika, 磷. K., Whitfield, S。, & 库珀, J. C. (2005). Detection and
repair of transient artifacts in fMRI data. 神经影像, 26, S36.
Meinzer, M。, Flaisch, T。, Seeds, L。, Harnish, S。, Antonenko, D .,
Witte, 五、, … Crosson, 乙. (2012). Same modulation but different
starting points: Performance modulates age differences in inferior
frontal cortex activity during word-retrieval. PLOS One, 7,
e33631. https://doi.org/10.1371/journal.pone.0033631
Menon, 五、, & Uddin, L. 问. (2010). Saliency, switching, attention and
控制: A network model of insula function. Brain Structure and
Function, 214, 655–667. https://doi.org/10.1007/s00429-010-0262-0
Methqal, 我。, Marsolais, Y。, Wilson, 中号. A。, Monchi, 奥。, & Joanette,
是. (2019). More expertise for a better perspective: Task and strategy-
driven adaptive neurofunctional reorganization for word pro-
duction in high-performing older adults. 老化, Neuropsychology,
和认知, 26, 190–221. https://doi.org/10.1080/13825585.
2017.1423021
莫尼耶, D ., Stamatakis, 乙. A。, & Tyler, L. K. (2014). Age-related
functional reorganization, structural changes, and preserved cog-
尼尼申. Neurobiology of Aging, 35, 42–54. https://doi.org/
10.1016/j.neurobiolaging.2013.07.003
磨坊主, K. J。, 罗杰斯, S. A。, Siddarth, P。, & 小的, G. 瓦. (2005).
Object naming and semantic fluency among individuals with
genetic risk for Alzheimer’s disease. International Journal of
Geriatric Psychiatry, 20, 128–136.
Moberg, M。, 费拉罗, F. R。, & Petros, 时间. V. (2000). Lexical properties
of the Boston Naming Test stimuli: Age differences in word naming
and lexical decision latency. Applied Neuropsychology, 7,
186–188. https://doi.org/10.1207/S15324826AN0703_5
Morcom, A. M。, & Henson, 右. 氮. A. (2018). Increased prefrontal
activity with aging reflects nonspecific neural responses rather
than compensation. 神经科学杂志, 38, 7303–7313.
https://doi.org/10.1523/jneurosci.1701-17.2018
Moritz-Gasser, S。, Herbet, G。, & Duffau, H. (2013). Mapping the
connectivity underlying multimodal (verbal and non-verbal)
semantic processing: A brain electrostimulation study.
Neuropsychologia, 51, 1814–1822. https://doi.org/10.1016/j.
neuropsychologia.2013.06.007
Mowinckel, A. M。, Espeseth, T。, & Westlye, L. 时间. (2012). 网络-
specific effects of age and in-scanner subject motion: A resting-
state fMRI study of 238 healthy adults. 神经影像, 63, 1364–1373.
穆勒, A. M。, Mérillat, S。, & Jäncke, L. (2016). Older but still flu-
耳鼻喉科? Insights from the intrinsically active baseline configuration
of the aging brain using a data driven graph-theoretical ap-
普罗奇. 神经影像, 127, 346–362. https://doi.org/10.1016/j.
neuroimage.2015.12.027
Muschelli, J。, Nebel, 中号. B., Caffo, 乙. S。, Barber, A. D ., Pekar, J. J。, &
Mostofsky, S. H. (2014). Reduction of motion-related artifacts in
resting state fMRI using aCompCor. 神经影像, 96, 22–35.
https://doi.org/10.1016/J.NEUROIMAGE.2014.03.028
Nagels, A。, Kircher, T。, Dietsche, B., Backes, H。, Marquetand, J。, &
Krug, A. (2012). Neural processing of overt word generation in
healthy individuals: The effect of age and word knowledge. Neuro-
图像, 61, 832–840. https://doi.org/10.1016/j.neuroimage.
2012.04.019
Nasreddine, Z. S。, Philips, 氮. A。, Bedirian, 五、, Charbonneau, S。,
Whitehead, 五、, Collin, 我。, … Chertkow, H. (2005). 蒙特利尔
Cognitive Assessment (MoCA). Journal of the American
Geriatrics Society, 53, 695–699.
公园, D. C。, & Reuter-Lorenz, 磷. (2009). The adaptive brain: 老化
and neurocognitive scaffolding. 心理学年度评论,
60, 173–196. https://doi.org/10.1146/annurev.psych.59.
103006.093656
Peelle, J. 乙. (2019). Language and aging. 客栈. 氧. Schiller & G. 我. 的
Zubicaray (编辑。), The Oxford handbook of neurolinguistics.
牛津: 牛津大学出版社. https://doi.org/10.1093/oxfordhb/
9780190672027.013.12
Perry, A。, Wen, W., Lord, A。, Thalamuthu, A。, 罗伯茨, G。,
米切尔, P。, … Breakspear, 中号. (2015). The organisation of the
elderly connectome. 神经影像, 114, 414–426. https://土井.
org/10.1016/j.neuroimage.2015.04.009
Persson, J。, Lustig, C。, 纳尔逊, J. K., & Reuter-Lorenz, 磷. A. (2007).
Age differences in deactivation: A link to cognitive control?
认知神经科学杂志, 19, 1021–1032. https://土井.
org/10.1162/jocn.2007.19.6.1021
Persson, J。, Pudas, S。, Nilsson, L.-G., & Nyberg, L. (2014).
Longitudinal assessment of default-mode brain function in aging.
Neurobiology of Aging, 35, 2107–2117. https://doi.org/10.1016/
J.NEUROBIOLAGING.2014.03.012
力量, J. D ., 巴恩斯, K. A。, 斯奈德, A. Z。, 施拉加尔, 乙. L。, &
彼得森, S. 乙. (2012). Spurious but systematic correlations in
functional connectivity MRI networks arise from subject motion.
神经影像, 59, 2142–2154. https://doi.org/10.1016/j.
neuroimage.2011.10.018
力量, J. D ., 施拉加尔, 乙. L。, & 彼得森, S. 乙. (2015). Recent prog-
ress and outstanding issues in motion correction in resting state
功能磁共振成像. 神经影像, 105, 536–551. https://doi.org/10.1016/j.
neuroimage.2014.10.044
Price, C. J. (2012). A review and synthesis of the first 20 years of
PET and fMRI studies of heard speech, spoken language and
阅读. 神经影像, 62, 816–847. https://doi.org/10.1016/j.
neuroimage.2012.04.062
Rajah, 中号. N。, & D’Esposito, 中号. (2005). Region-specific changes in
prefrontal function with age: A review of PET and fMRI studies on
working and episodic memory. Brain, 128, 1964–1983.
Raz, N。, Lindenberger, U。, Rodrigue, K. M。, 肯尼迪, K. M。, Head,
D ., 威廉森, A。, … Acker, J. D. (2005). Regional brain changes
in aging healthy adults: General trends, individual differences and
modifiers. 大脑皮层, 15, 1676–1689. https://doi.org/
10.1093/cercor/bhi044
Reilly, J。, Peelle, J. E., Antonucci, S. M。, & Grossman, 中号. (2011).
Anomia as a marker of distinct semantic memory impairments
in Alzheimer’s disease and semantic dementia. Neuropsychology,
25, 413.
Neurobiology of Language
182
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
罗伯茨, 磷. M。, & Doucet, 氮. (2011). Résultats du Boston Naming Test
chez des adultes francophones de Québec. Canadian Journal of
Speech-Language Pathology and Audiology, 35, 254–267.
Rogalski, Y。, Peelle, J. E., & Reilly, J. (2011). Effects of perceptual and
contextual enrichment on visual confrontation naming in adult
老化. Journal of Speech, 语言, and Hearing Research, 54,
1349–1360. https://doi.org/10.1044/1092-4388(2011/10-0178)
Sal a-Llonch, R。, Bartrés-Fa z, D. , & Ju nq ué , C .
( 20 15 ).
Reorganization of brain networks in aging: A review of functional
connectivity studies. 心理学前沿, 6, 663. https://土井.
org/10.3389/fpsyg.2015.00663
Salehi, M。, Greene, A. S。, Karbasi, A。, 沉, X。, Scheinost, D ., &
Constable, 右. 时间. (2018). There is no single functional atlas even
for a single individual: Parcellation of the human brain is state
dependent. BioRxiv, 431833.
Salehi, M。, Karbasi, A。, Scheinost, D ., & Constable, 右. 时间. (2017). A
submodular approach to create individualized parcellations of
the human brain. 在米. 德科托, L. 迈尔-海因, A. Franz,
磷. Jannin, D. 柯林斯, & S. Duchesne (编辑。), Medical image com-
puting and computer assisted intervention – MICCAI 2017
(PP. 478–485). 占婆, 瑞士: 施普林格.
Salthouse, 时间. A. (2014). Quantity and structure of word knowledge
across adulthood. 智力, 46, 122–130. https://doi.org/
10.1016/j.intell.2014.05.009
Sambataro, F。, Murty, V. P。, Callicott, J. H。, Tan, H.-Y., 这, S。,
温伯格, D. R。, & Mattay, V. S. (2010). Age-related alter-
ations in default mode network: Impact on working memory per-
formance. Neurobiology of Aging, 31, 839–852. https://doi.org/
10.1016/j.neurobiolaging.2008.05.022
Samu, D ., 坎贝尔, K. L。, Tsvetanov, K. A。, Shafto, 中号. A。, Cam-CAN,
& Tyler, L. K. (2017). Preserved cognitive functions with age are
determined by domain-dependent shifts in network responsivity.
Nature Communications, 8, 14743. https://doi.org/10.1038/
ncomms14743
Sarubbo, S。, De Benedictis, A。, Merler, S。, Mandonnet, E.,
Barbareschi, M。, Dallabona, M。, … Duffau, H. (2016).
Structural and functional integration between dorsal and ventral
language streams as revealed by blunt dissection and direct elec-
trical stimulation. 人脑图谱, 37, 3858–3872.
https://doi.org/10.1002/hbm.23281
Satterthwaite, 时间. D ., Elliott, 中号. A。, Gerraty, 右. T。, Ruparel, K.,
Loughead, J。, Calkins, 中号. E., … Wolf, D. H. (2013). An improved
framework for confound regression and filtering for control of
motion artifact in the preprocessing of resting-state functional
connectivity data. 神经影像, 64, 240–256. https://doi.org/
10.1016/j.neuroimage.2012.08.052
Schmitter-Edgecombe, M。, Vesneski, M。, & 琼斯, D. 瓦. 右. (2000).
Aging and word-finding: A comparison of spontaneous and con-
strained naming tests. Archives of Clinical Neuropsychology, 15,
479–493. https://doi.org/10.1016/S0887-6177(99)00039-6
Schneider-Garces, 氮. J。, Gordon, 乙. A。, Brumback-Peltz, C. R。,
Shin, E., 李, Y。, Sutton, 乙. P。, … Fabiani, 中号. (2010). Span,
CRUNCH, and beyond: Working memory capacity and the ag-
ing brain. 认知神经科学杂志, 22, 655–669.
https://doi.org/10.1162/jocn.2009.21230
Schultz, D. H。, & Cole, 中号. 瓦. (2016). Higher intelligence is associ-
ated with less task-related brain network reconfiguration. 杂志
of Neuroscience, 36, 8551–8561. https://doi.org/10.1523/
JNEUROSCI.0358-16.2016
施瓦茨, 乙. L。, & 弗雷泽, L. D. (2005). Tip-of-the-tongue states and
老化: Contrasting psycholinguistic and metacognitive perspec-
特维斯. Journal of General Psychology, 132, 377–391. https://土井.
org/10.3200/GENP.132.4.377-391
Seeley, 瓦. W., Menon, 五、, Schatzberg, A. F。, 凯勒, J。, Glover, G. H。,
Kenna, H。, … Greicius, 中号. D. (2007). Dissociable intrinsic con-
nectivity networks for salience processing and executive control.
神经科学杂志, 27, 2349–2356.
Seghier, 中号. L。, Hope, 时间. 中号. H。, Prejawa, S。, Parker Jones, 奥。,
Vitkovitch, M。, & Price, C. J. (2015). A trade-off between somato-
sensory and auditory related brain activity during object naming
but not reading. 神经科学杂志, 35, 4751–4759. https://
doi.org/10.1523/JNEUROSCI.2292-14.2015
Shafto, 中号. A。, 伯克, D. M。, Stamatakis, 乙. A。, Tam, 磷. P。, & Tyler,
L. K. (2007). On the tip-of-the-tongue: Neural correlates of in-
creased word-finding failures in normal aging. 杂志
Cognitive Neuroscience, 19, 2060–2070. https://doi.org/
10.1162/jocn.2007.19.12.2060
Shafto, 中号. A。, James, L. E., Abrams, L。, & Tyler, L. K. (2017). 年龄-
related increases in verbal knowledge are not associated with
word finding problems in the Cam-CAN cohort: What you know
won’t hurt you. The Journals of Gerontology Series B, 72, 100–106.
https://doi.org/10.1093/geronb/gbw074
Shafto, 中号. A。, & Tyler, L. K. (2014). Language in the aging brain:
The network dynamics of cognitive decline and preservation.
科学, 346(6209), 583–588.
Shine, J. M。, Bissett, 磷. G。, 钟, 磷. T。, Koyejo, 奥。, Balsters, J. H。,
Gorgolewski, K. J。, … Poldrack, 右. A. (2016). The dynamics of
functional brain networks: Integrated network states during cog-
nitive task performance. 神经元, 92, 544–554. https://doi.org/
10.1016/j.neuron.2016.09.018
Simons, J. S。, Koutstaal, W., 王子, S。, 瓦格纳, A. D ., & Schacter,
D. L. (2003). Neural mechanisms of visual object priming:
Evidence for perceptual and semantic distinctions in fusiform
cortex. 神经影像, 19, 613–626.
歌曲, J。, Desphande, A. S。, Meier, 时间. B., Tudorascu, D. L。, Vergun,
S。, Nair, V. A。, … Bellec, 磷. (2012). Age-related differences in
test-retest reliability in resting-state brain functional connectivity.
PLOS One, 7, e49847.
Sörös, P。, Bose, A。, Sokoloff, L. G。, Graham, S. J。, & 愚蠢的, D. 时间. (2011).
Age-related changes in the functional neuroanatomy of overt
speech production. Neurobiology of Aging, 32, 1505–1513.
https://doi.org/10.1016/ J.NEUROBIOLAGING.2009.08.015
Spreng, 右. N。, Stevens, 瓦. D ., Viviano, J. D ., & Schacter, D. L.
(2017). Attenuated anticorrelation between the default and dor-
sal attention networks with aging: Evidence from task and rest.
Neurobiology of Aging, 45, 149–160. https://doi.org/10.1016/j.
neurobiolaging.2016.05.020
Steffener, J。, Barulli, D ., Habeck, C。, O’Shea, D ., Razlighi, Q.,
Stern, Y。, … Bosch, 乙. (2014). The role of education and verbal
abilities in altering the effect of age-related gray matter differ-
ences on cognition. PLOS One, 9, e91196. https://doi.org/10.
1371/journal.pone.0091196
Steffener, J。, Habeck, C. G。, & Stern, 是. (2012). Age-related changes
in task related functional network connectivity. PLOS One, 7,
e44421. https://doi.org/10.1371/journal.pone.0044421
Stern, Y。, Habeck, C。, Steffener, J。, Barulli, D ., Gazes, Y。, Razlighi,
Q., … Salthouse, 时间. (2014). The Reference Ability Neural
Network Study: 动机, 设计, and initial feasibility analy-
塞斯. 神经影像, 103, 139–151. https://doi.org/10.1016/j.
neuroimage.2014.09.029
Stoodley, C. J。, Valera, 乙. M。, & Schmahmann, J. D. (2012).
Functional topography of the cerebellum for motor and cognitive
任务: An fMRI study. 神经影像, 59, 1560–1570. https://土井.
org/10.1016/J.NEUROIMAGE.2011.08.065
泰勒, J. R。, 威廉姆斯, N。, Cusack, R。, Auer, T。, Shafto, 中号. A。,
狄克逊, M。, … Henson, 右. 氮. (2015). The Cambridge Centre for
Neurobiology of Language
183
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Naming functional connectivity in aging
Ageing and Neuroscience (Cam-CAN) data repository: 结构性
and functional MRI, 乙二醇, and cognitive data from a cross-
sectional adult lifespan sample. 神经影像, 144, 262–269.
https://doi.org/10.1016/j.neuroimage.2015.09.018
Tomasi, D ., & Volkow, 氮. D. (2010). Functional connectivity den-
sity mapping. 美国国家科学院院刊,
107, 9885–9890. https://doi.org/10.1073/pnas.1001414107
Tomasi, D ., & Volkow, 氮. D. (2012). Aging and functional brain
网络. Molecular Psychiatry, 17, 549–558. https://doi.org/
10.1038/mp.2011.81
特兰, S. M。, 麦格雷戈, K. M。, James, G. A。, Gopinath, K.,
Krishnamurthy, 五、, Krishnamurthy, L. C。, & Crosson, 乙. (2018).
Task-residual functional connectivity of language and attention
网络. Brain and Cognition, 122, 52–58. https://doi.org/
10.1016/J.BANDC.2018.02.003
Tremblay, P。, Deschamps, 我。, & Gracco, V. L. (2016). Neurobiology
of speech production: A motor control perspective. In G. Hickok &
S. L. 小的 (编辑。), Neurobiology of language (PP. 741–750).
伦敦: 学术出版社. https://doi.org/10.1016/ B978-0-12-
407794-2.00059-6
Tsvetanov, K. A。, Henson, 右. 氮. A。, Tyler, L. K., Razi, A。, Geerligs, L。,
Ham, 时间. E., … Cambridge Centre for Ageing and Neuroscience.
(2016). Extrinsic and intrinsic brain network connectivity main-
tains cognition across the lifespan despite accelerated decay of re-
gional brain activation. 神经科学杂志, 36, 3115–3126.
https://doi.org/10.1523/JNEUROSCI.2733-15.2016
车工, G. R。, & Spreng, 右. 氮. (2015). Prefrontal engagement and
reduced default network suppression co-occur and are dynami-
cally coupled in older adults: The default–executive coupling
hypothesis of aging. 认知神经科学杂志, 27,
2462–2476. https://doi.org/10.1162/jocn_a_00869
Tyler, L. K., 张, 时间. 磷. L。, Devereux, 乙. J。, & 克拉克, A. (2013).
Syntactic computations in the language network: Characterizing
dynamic network properties using representational similarity
分析. 心理学前沿, 4, 271.
Van Dijk, K. 右. A。, 肥皂, 中号. R。, & 巴克纳, 右. L. (2012). 这
influence of head motion on intrinsic functional connectivity
MRI. 神经影像, 59, 431–438. https://doi.org/10.1016/j.
neuroimage.2011.07.044
Varangis, E., Razlighi, Q., Habeck, C. G。, Fisher, Z。, & Stern, 是.
(2019). Between-network functional connectivity is modified by
age and cognitive task domain. 认知神经科学杂志,
31, 607–622. https://doi.org/10.1162/jocn_a_01368
Verhaegen, C。, & Poncelet, 中号. (2013). Changes in naming and se-
mantic abilities with aging from 50 到 90 年. Journal of the
International Neuropsychological Society, 19, 119–126. https://
doi.org/10.1017/S1355617712001178
王, W.-C., Dew, 我. 时间. Z。, & Cabeza, 右. (2015). Age-related dif-
ferences in medial temporal lobe involvement during conceptual
fluency. 脑研究, 1612, 48–58. https://doi.org/10.1016/j.
brainres.2014.09.061
Whitfield-Gabrieli, S。, & Nieto-Castanon, A. (2012). Conn: A func-
tional connectivity toolbox for correlated and anticorrelated
brain networks. Brain Connectivity, 2, 125–141. https://doi.org/
10.1089/brain.2012.0073
WHO. (2018). What is healthy ageing? Retrieved February 8, 2018,
from http://www.who.int/ageing/healthy-ageing/en/
Wierenga, C. E., 本杰明, M。, Gopinath, K., Perlstein, 瓦. M。,
Leonard, C. M。, Rothi, L. J. G。, … Crosson, 乙. (2008). Age-related
changes in word retrieval: Role of bilateral frontal and subcorti-
cal networks. Neurobiology of Aging, 29, 436–451. https://土井.
org/10.1016/j.neurobiolaging.2006.10.024
明智的, R。, Greene, J。, Büchel, C。, & 斯科特, S. (1999). Brain regions
involved in articulation. The Lancet, 353(9158), 1057–1061.
https://doi.org/10.1016/S0140-6736(98)07491-1
严, C.-G., 张, B., 凯莉, C。, Colcombe, S。, Craddock, 右. C。,
Di Martino, A。, … Milham, 中号. 磷. (2013). A comprehensive
assessment of regional variation in the impact of head micro-
movements on functional connectomics. 神经影像, 76,
183–201. https://doi.org/10.1016/j.neuroimage.2013.03.004
Zabelina, D. L。, & Andrews-Hanna, J. 右. (2016). Dynamic network
interactions supporting internally-oriented cognition. 当前的
Opinion in Neurobiology, 40, 86–93. https://doi.org/10.1016/J.
CONB.2016.06.014
扎莱斯基, A。, 假如, A。, & 布莫尔, 乙. 时间. (2010). 基于网络
statistic: Identifying differences in brain networks. 神经影像,
53, 1197–1207.
朱, D ., 张, T。, Jiang, X。, 胡, X。, 陈, H。, 哪个, N。, … Liu, 时间.
(2013). Fusing DTI and fMRI data: A survey of methods and ap-
并发症. 神经影像, 102, 184–191. https://doi.org/10.1016/j.
neuroimage.2013.09.071
Zuo, X.-N., 凯莉, C。, Di Martino, A。, Mennes, M。, 马古利斯, D. S。,
Bangaru, S。, … Milham, 中号. 磷. (2010). Growing together and growing
apart: Regional and sex differences in the lifespan developmental tra-
jectories of functional homotopy. 神经科学杂志, 30,
15034–15043. https://doi.org/10.1523/JNEUROSCI.2612-10.2010
Neurobiology of Language
184
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
2
1
6
1
1
8
6
7
6
9
1
n
哦
_
A
_
0
0
0
0
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3