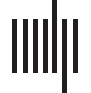研究文章
Canonical Sentence Processing and the Inferior
Frontal Cortex: Is There a Connection?
开放访问
杂志
Nicholas Riccardi1
, Chris Rorden1,2
, Julius Fridriksson2,3
, and Rutvik H. Desai1,2
1心理学系, 南卡罗来纳大学, 哥伦比亚, sc
2Institute for Mind and Brain, 南卡罗来纳大学, 哥伦比亚, sc
3Department of Communication Sciences and Disorders, 南卡罗来纳大学, 哥伦比亚, sc
关键词: 中风, Broca’s area, 句子理解, anterior temporal lobe, lesion-symptom
映射, 连接性
抽象的
The role of left inferior frontal cortex (LIFC) in canonical sentence comprehension is
有争议的. Many studies have found involvement of LIFC in sentence production or complex
句子理解, but negative or mixed results are often found in comprehension
of simple or canonical sentences. We used voxel-, region-, and connectivity-based lesion
symptom mapping ( VLSM, RLSM, CLSM) in left-hemisphere chronic stroke survivors to
investigate canonical sentence comprehension while controlling for lexical-semantic,
executive, and phonological processes. We investigated how damage and disrupted white
matter connectivity of LIFC and two other language-related regions, the left anterior temporal
叶 (LATL) and posterior temporal-inferior parietal area (LpT-iP), affected sentence
理解. VLSM and RLSM revealed that LIFC damage was not associated with
canonical sentence comprehension measured by a sensibility judgment task. LIFC damage was
associated instead with impairments in a lexical semantic similarity judgment task with high
semantic/executive demands. Damage to the LpT-iP, specifically posterior middle temporal
回 (PMTG), predicted worse sentence comprehension after controlling for visual lexical
使用权, semantic knowledge, and auditory-verbal short-term memory (STM), but not auditory
single-word comprehension, suggesting pMTG is vital for auditory language comprehension.
CLSM revealed that disruption of left-lateralized white-matter connections from LIFC to LATL
and LpT-iP was associated with worse sentence comprehension, controlling for performance
in tasks related to lexical access, auditory word comprehension, and auditory-verbal STM.
然而, the LIFC connections were accounted for by the lexical semantic similarity judgment
任务, which had high semantic/executive demands. This suggests that LIFC connectivity is
relevant to canonical sentence comprehension when task-related semantic/executive demands
are high.
介绍
Comprehending a spoken sentence is a complex process that requires coordination of multiple
cognitive resources, such as phonological, executive, 词汇的, 句法的, and semantic opera-
系统蒸发散. Reflecting this complexity, studies demonstrate that numerous distributed brain areas
functionally contribute to sentence comprehension (Dronkers et al., 2004; 弗里德里克, 2012;
哈科特 & Indrey, 2014; Walenski et al., 2019). Of these areas, the contribution of the left
inferior frontal cortex (LIFC) in sentence comprehension remains particularly controversial.
引文: Riccardi, N。, Rorden, C。,
Fridriksson, J。, & 德赛, 右. H. (2022).
Canonical sentence processing and
the inferior frontal cortex: Is there
a connection? 神经生物学
语言, 3(2), 318–344. https://doi.org
/10.1162/nol_a_00067
DOI:
https://doi.org/10.1162/nol_a_00067
已收到: 1 四月 2021
公认: 21 一月 2022
利益争夺: 作者有
声明不存在竞争利益
存在.
通讯作者:
Nicholas Riccardi
riccardn@email.sc.edu
处理编辑器:
Liina Pylkkanen
版权: © 2022
麻省理工学院
在知识共享下发布
归因 4.0 国际的
(抄送 4.0) 执照
麻省理工学院出版社
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
While many studies have found involvement of LIFC in sentence production and in complex
句子理解, negative or mixed results are often found in the comprehension of
simple or canonical sentences. A more complete understanding of the role of LIFC in canonical
sentence comprehension would help inform neuroanatomical models of language processing.
The LIFC has traditionally been associated with language production, but a growing num-
ber of studies report its involvement in comprehension (德赛 & Riccardi, 2021; Fadiga et al.,
2009; Rogalsky & 希科克, 2011). In regards to sentence processing specifically, results of neu-
roimaging studies of the LIFC, here defined as Brodmann Areas (BA) 44 和 45, 已经
inconsistent (see Kemmerer, 2021). A meta-analysis of 53 neuroimaging studies comparing
sentence listening or reading to control conditions found that the inferior frontal gyrus pars
opercularis (IFGoper) and pars triangularis (IFGtri) were only activated in 13 和 23 学习,
分别 (哈科特 & Indrey, 2014). 的确, multiple neuroimaging studies have found
that reading or listening to sentences passively does not activate the LIFC compared to word
列表 (Humphries et al., 2006; Mazoyer et al., 1993; Rogalsky & 希科克, 2009). 然而, 一个
additional subanalysis within the Hagoort and Indefrey (2014) investigation revealed that
semantic or syntactic demands (such as violation detection) during sentential processing reli-
ably activated areas in the LIFC. Multiple neuroimaging studies have found LIFC activation for
simple phrases and canonical word orders during tasks such as semantic/syntactic violation
detection or meaningfulness judgment (Graessner et al., 2021; Schell et al., 2017; Zaccarella
& 弗里德里克, 2015).
These findings suggest that LIFC may be involved in the comprehension of simple phrases
and canonical sentences, especially when there are task-related demands requiring attention
to meaning or form. This predicts that damage to the LIFC should be associated with
canonical/simple sentence comprehension impairments when measured by tasks that orient
attention to semantic/syntactic error detection or meaningfulness. 然而, neuropsychological
evidence supporting this prediction is relatively scarce, as briefly reviewed below. While consid-
ering these studies it is important to note that, even if noncanonical sentence comprehension is
significantly more associated with LIFC disfunction than canonical comprehension, 它不是
follow that the LIFC does not also functionally contribute to canonical comprehension.
Two studies of patients with gliomas in the LIFC (Kinno et al., 2009, 2014) 发现, com-
pared to healthy controls, patients were impaired at two-argument active canonical sentence
comprehension as measured by a sentence-picture matching task. They used voxel-based
lesion-symptom mapping ( VLSM) to demonstrate that these sentence impairments were signif-
icantly associated with damage to portions of the LIFC. 威尔逊等. (2016) 发现, com-
pared to healthy controls, patients with primary progressive aphasia (PPA) were less accurate
and responded more slowly to canonical sentences, especially for longer sentences, as mea-
sured by a sentence-picture matching task. Voxel-based morphometry ( VBM) implicated LIFC
and surrounding frontal cortices in the accuracy impairments. 然而, the VBM analysis of
comprehension accuracy in that study grouped canonical and noncanonical accuracies
一起, and LIFC atrophy was significantly associated with slower response times for nonca-
nonical sentences specifically, controlling for canonical performance. In a VLSM study of
patients with left-hemisphere stroke, Magnusdottir et al. (2013) found that damage to white
matter underlying LIFC was associated with worse sentence-picture matching performance
for canonical sentences, in addition to damage to posterior temporal regions. Caramazza
等人. (2005) 证明了这一点, in a group of patients with Broca’s aphasia and confirmed
damage to LIFC, 17 在......之外 38 patients were unable to perform significantly better than chance
accuracy on a sentence-picture matching task of active canonical reversible sentences. 如何-
曾经, patient performance was highly variable, and the lack of an explicit anatomical lesion-
语言的神经生物学
319
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
deficit association analysis makes it impossible to ascribe these impairments to LIFC damage
specifically instead of other areas commonly damaged in patients diagnosed with Broca’s apha-
是 (例如, insula, 颞上回 (STG); Dronkers et al., 2007; Fridriksson et al., 2015).
另一方面, many studies have found no relationship between LIFC damage and
canonical sentence comprehension. In a seminal VLSM study, Dronkers et al. (2004) 成立
no relationship between damage to IFGoper or IFGtri and sentence-picture matching for
canonical sentences. Newhart et al. (2012) also found no association between LIFC damage
and canonical sentences using sentence-picture matching and sentence reenactment, 和
LIFC damage being associated instead with auditory-verbal and working memory deficits as
measured by forward and backward digit span. Also using sentence-picture matching,
Rogalsky et al. (2018) found no association between LIFC damage and canonical sentence
理解, even in a subset of patients (N= 11) with relatively focal lesions to LIFC.
反而, these studies find evidence either that damage to posterior temporal areas is related
to worse sentence comprehension (例如, Kristinsson et al., 2020; Thothathiri et al., 2012), 或者
that LIFC damage is only associated with impairments for complex syntactic structures. 拍摄
一起, these studies suggest that the LIFC does not functionally contribute to canonical sen-
tence comprehension, with its role instead being dependent on either production demands or
syntactic complexity.
These mixed findings raise questions about the functional involvement of LIFC during com-
prehension of simple phrases and sentences. One consideration is the relative lack of task
variety used in patient studies for measuring canonical comprehension. A large majority of
those studies use sentence-picture matching. In that task, an incompatible picture trial for
canonical sentences is typically created either by switching the places of the subject and
目的 (for reversible sentences), or by replacing the subject or object with a different
entity/item than what is described in the sentence (for nonreversible). Sentence-picture match-
ing for simple sentences therefore focuses mainly on aspects of comprehension related to
thematic role assignment (例如, who is the agent and who is the patient) or single word under-
常设, especially related to nouns (例如, whether the item/entity in the picture matches the
word in the sentence). Other aspects of sentence comprehension, such as those related to the
comprehension of verbs and their compatibility with the nouns in the sentence, are not nec-
essarily measured. Using sentence comprehension tasks with demands that differ from
sentence-picture matching may provide novel insights about the functional contribution (或者
lack thereof ) of LIFC to canonical sentence comprehension.
Another consideration is that most patient studies of canonical sentence comprehension
have used traditional lesion overlap methods, such as VLSM, to examine the relationship
between behavior and brain damage. A limitation of these methods is that they can only detect
areas of overlapping necrosis/gliosis. 或者, white matter damage/disconnection has
been demonstrated to have wide-ranging consequences outside of the necrotic area and
can lead to functional disruption of cortical regions that are spared by the lesion (Bonilha
等人。, 2014; Bonilha & Fridriksson, 2009; Bonilha, Rorden, & Fridriksson, 2014; Catani
等人。, 2012; Catani & ffytche, 2005; Fridriksson et al., 2007). VLSM also requires a number
of patients to have overlapping damage within a given area of interest in order to detect behav-
ioral associations, a limitation that may lead to false negatives or inconsistent results between
studies depending on sample size and etiology of the brain damage (例如, glioma, PPA, 中间
cerebral artery stroke). Connectivity-based lesion-symptom mapping (CLSM; Gleichgerrcht
等人。, 2017) can complement traditional VLSM methods by detecting effects of disrupted white
matter connectivity resulting from damage anywhere along white matter tracts that connect
two grey matter regions. This provides two advantages. 第一的, it can detect effects associated
语言的神经生物学
320
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
with disconnection between two grey matter areas even if those areas are spared by the lesion.
第二, because it is senstive to damage located anywhere along a given white matter tract
(even if the specific areas of damage do not overlap between patients), it does not necessarily
require the same amount of lesion overlap as VLSM. Using complementary connectivity-based
measures in addition to traditional VLSM may reveal effects associated with LIFC disconnec-
tion from the larger language comprehension network that cannot be detected using VLSM in
隔离.
的确, prior studies have raised the possibility that, despite negative VLSM findings
regarding the association between LIFC damage and sentence comprehension, structural or
functional disconnection of LIFC from the larger language network may have behavioral
结果 (den Ouden et al., 2019; Fridriksson et al., 2018; Lukic et al., 2021; 土耳其人
& drungers, 2011). 例如, although Dronkers et al. (2004) did not find a relationship
between LIFC damage and sentence comprehension, a follow-up study in healthy participants
showed that portions of the LIFC display functional and structural connectivity to the posterior
temporal area that was associated with sentence comprehension in the original lesion study
(土耳其人 & drungers, 2011). The current investigation seeks to expand upon that finding by
using CLSM to explicitly examine the behavioral associates of structural disconnection of LIFC.
此外, Fridriksson et al. (2018) 发现, while damage to LIFC was not associated
with sentence comprehension impairments (canonical and noncanonical included together),
white matter disconnection between the IFGoper and IFGtri did predict sentence comprehen-
sion impairments. Den Ouden et al. (2019) found similar results, but specifically for more
complex syntactic structures. The current study uses similar methods, but focuses specifically
on canonical sentences, to provide additional information about the role of LIFC in sentence
理解.
这里, in a retrospective study using data collected as part of a previously existing language
and cognition task battery, we used VLSM, CLSM, and region-based lesion symptom mapping
(RLSM) in a group of unilateral left-hemisphere chronic stroke survivors to investigate compre-
hension of canonical sentences while controlling for related cognitive abilities (例如, 词汇的
语义学, auditory single-word comprehension, auditory-verbal short-term memory (STM)).
Our focus was to investigate how damage and disrupted white matter connectivity of the LIFC
and areas within two other language-related regions, the left anterior temporal lobe (LATL) 和
left posterior temporal-inferior parietal area (LpT-iP), affected canonical sentence comprehen-
sion and related processes. We used an auditory sentence sensibility task on declarative sen-
tences which required participants to determine if a sentence made sense, as opposed to the
commonly used sentence-picture matching task. We included four control tasks with varying
lexical-semantic, executive, and phonological demands to interrogate the LIFC’s involvement
in these processes (for a full description of the demands of each task, see Materials and Pro-
cedure). A visual lexical decision task with low semantic, executive, and phonological
demands was used to control for lexical access. Auditory word comprehension, 有
relatively higher semantic, executive, and phonological demands, was used to control for
auditory single-word comprehension. A visual semantic similarity judgment task, which had
high semantic and executive demands, but low phonological demands, was used to control for
semantic retrieval. The forward digit span task was used to control for auditory-verbal STM.
Several hypotheses regarding the role of LIFC can be tested given this retrospective lan-
guage and cognition battery. 第一的, the merge hypothesis states that the LIFC is involved in
the binding of two or more elements into a hierarchical structure (Zaccarella & 弗里德里克,
2015; Zaccarella et al., 2017), starting with smaller units (例如, the boy) and expanding into
larger structures as the phrase or sentence continues (例如, the boy kicks). This process would
语言的神经生物学
321
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
be important for successful completion of the sentence sensibility task but less so for the other
tasks included here. Merge would be important for the sentence task because to judge the
sensibility of a sentence, a hierarchical structure must be built such that the meaningfulness
of the semantic and grammatical relations among the subject, 动词, and object can be
accepted or rejected. Two of the other tasks (visual lexical decision, visual semantic similarity
判断) have the merge operation at the most basic level (例如, nouns and verbs are pre-
ceded by the and to, 分别), but they only contain this basic two-word structure as
opposed to phrase-level combinations in the sentence sensibility task. If LIFC disruption is
associated with worse sentence comprehension, but not with the other tasks, it could be taken
as evidence that the LIFC functionally contributes to merge or related structure-building pro-
cesses at the phrase or sentence level.
A second hypothesis is that the LIFC contributes to sentence comprehension via executive
task-related demands, especially pertaining to detection of syntactic or semantic violations.
Evidence for this account comes from previously discussed neuroimaging findings suggesting
that LIFC is especially active in sentence comprehension tasks that require special attention to
syntactic or semantic information (哈科特 & Indrey, 2014; Hasson et al., 2006; Love et al.,
2006; Rogalsky & 希科克, 2009). Because the current sentence task explicitly orients attention
to the semantic meaningfulness of the sentences, support for this hypothesis would come from
LIFC being a shared neural substrate for the sentence comprehension task and the semantic
similarity judgment, as that task also requires explicit semantic analysis.
第三, the LIFC may contribute to sentence comprehension through auditory-verbal STM.
Evidence for this hypothesis comes from neuroimaging studies showing that some areas in the
LIFC are activated by both syntactic demands and STM (Matchin et al., 2017; Rogalsky &
希科克, 2011; Rogalsky等。, 2008). Neuropsychological studies also find that LIFC damage
is correlated with reduced digit span and impaired comprehension of sentences (Pettigrew &
Hillis, 2014). To successfully complete the auditory sentence sensibility task, 参与者
must hold the subject, 动词, and object in their STM long enough to make the meaningfulness
判断. In the current study, finding that forward digit span and sentence comprehension
share the LIFC as a common neural substrate would support the auditory-verbal STM
假设.
最后, the LIFC may be involved in a variety of operations that subserve general lexical-
语义处理 (Fiebach et al., 2002; Heim et al., 2009; Kotz et al., 2002, 2010; 拉夫
等人。, 2008), which may contribute to sentence comprehension. The current sentence sensi-
bility task requires participants to understand individual words, as well as conceptual relation-
ships between words, to judge meaningfulness. Finding that LIFC is a common neural substrate
for sentence comprehension and the tasks requiring lexical-semantic search and access (IE。,
auditory word comprehension, visual lexical decision, and visual semantic similarity), 会
support this hypothesis.
材料和方法
参加者
Seventy-five (24 女性) native English-speaking participants with unilateral left-hemisphere
stroke were recruited. Fourteen participants did not complete the sentence sensibility task
due to time constraints on the day of testing or technological malfunction, 离开 61 (17 女性)
participants for the neuroanatomical analyses (见表 1 for demographic information). 过去的
power analysis has demonstrated that samples of ∼50 or greater provide adequate power to
语言的神经生物学
322
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
桌子 1.
Participant demographic information
参与者
M2025
年龄
56
性别
中号
教育
(年)
16
Dominant hand
prior to stroke
正确的
Aphasia type
Broca’s
WAB: AQ
64.6
M2071
M2005
M2036
M2006
M2002
M2069
M2007
M2020
M2141
M2142
M2046
M2014
M2074
M2061
M2078
M2143
M2072
M2144
M2145
M2146
M2075
M2059
M2040
M2076
M2147
M2082
M2079
M2149
M2151
M2152
69
38
54
56
65
77
76
59
79
59
56
62
55
65
54
69
43
65
65
37
41
61
52
75
61
55
59
64
62
61
中号
F
中号
中号
中号
F
中号
F
F
中号
F
中号
中号
中号
中号
F
中号
F
中号
F
F
中号
中号
中号
F
中号
中号
F
中号
中号
16
16
18
12
16
18
12
12
16
16
14
12
16
14
12
16
16
12
12
18
18
16
16
16
16
12
13
18
16
14
正确的
正确的
正确的
正确的
正确的
正确的
正确的
左边
左边
正确的
正确的
正确的
正确的
正确的
左边
正确的
左边
左边
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
Broca’s
Broca’s
Broca’s
Anomic
Broca’s
Anomic
Broca’s
Anomic
没有任何
没有任何
Conduction
Anomic
Broca’s
Wernicke’s
Conduction
没有任何
全球的
没有任何
Anomic
没有任何
Anomic
Broca’s
Broca’s
Conduction
没有任何
Anomic
全球的
没有任何
没有任何
Anomic
63.6
55.2
76.2
83.2
80.4
90.5
26.1
86.2
99.1
99.2
51.5
94
59.4
52.7
88.8
99.2
23.6
99.6
93.2
98.5
94.2
58.2
57.5
72.1
97.3
91.1
25.3
98.6
96.9
93.1
语言的神经生物学
323
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
参与者
M2153
年龄
48
性别
中号
桌子 1.
(持续 )
教育
(年)
14
Dominant hand
prior to stroke
正确的
M2086
M4138
M2088
M2044
M2087
M2155
M2030
M2156
M2158
M2159
M2160
M2094
M2162
M2031
M2103
M4180
M2164
M2106
M2110
M2109
M2114
M2111
M2029
M4209
M2119
M2121
M2117
M2127
71
67
47
53
69
62
62
39
51
72
49
52
59
61
64
66
68
60
55
67
63
52
48
56
48
63
44
59
中号
中号
中号
F
中号
F
中号
F
F
中号
F
中号
中号
中号
中号
中号
中号
中号
F
F
中号
中号
中号
中号
中号
中号
中号
中号
18
14
18
13
16
16
16
12
18
16
16
20
13
18
16
12
16
16
12
16
16
16
16
18
16
12
16
12
正确的
正确的
正确的
正确的
正确的
正确的
左边
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
正确的
左边
正确的
Aphasia type
Anomic
Conduction
Anomic
Anomic
Broca’s
Broca’s
没有任何
Broca’s
没有任何
没有任何
没有任何
没有任何
Broca’s
没有任何
Wernicke’s
Conduction
Conduction
没有任何
Anomic
Anomic
Wernicke’s
全球的
Anomic
Broca’s
Conduction
没有任何
没有任何
Broca’s
Transcortical
感官的
WAB: AQ
87.6
73.5
77.8
87.5
74.8
48.9
98.9
57.2
96.7
99.6
99.1
96.6
64.6
98.4
31.2
82.9
45.2
97
90.1
91.3
47.8
15.2
93.4
43
74.6
97.5
98.1
49.1
57.8
语言的神经生物学
324
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
detect medium to strong effects in the majority of brain areas (Kimberg et al., 2007). 参加者
were at least 6 months post-stroke (4.02 years ± 4.4), and a mean age at time of testing of 58.75
years ± 9.68. The Western Aphasia Battery ( WAB; Kertesz, 2007) Aphasia Quotient mean score
曾是 75.7 ± 23.6. All participants signed informed consent, and the University of South Caro-
lina Institutional Review Board approved the research.
Materials and Procedure
Auditory sentence sensibility
One hundred canonical declarative and 50 low-meaningfulness or nonsense sentences were
presented auditorily to the participants. Sentences were sound files recorded in a noise atten-
uated room by a male speaker who was instructed to read as clearly and naturally as possible,
as if they were reading aloud to someone else. Sentences were denoised using Audacity audio
editing software (https://www.audacityteam.org/) and ranged from 2–3.5 s in length. To pre-
serve a more naturalistic sound for the stimuli, speech rate was not artificially modified
post-recording. In a sound attenuated room, sentences were played aloud to participants on
a laptop PC running E-prime software (版本 1.2, 心理学软件工具, 公司; https://
pstnet.com/products/e-prime/). Ten practice sentences were presented before the real trials
to ensure that the participant could hear the sentences and understood the task. The partici-
pant was instructed to determine as quickly and accurately as possible whether the sentence
made sense or not by pressing one of two response buttons. Participants had 10 s from the
onset of the sentence to respond.
Fifty of the sensible sentences were literal. Half of the literal sentences described physical
hand/arm actions (The repairman bent the cable for her), while the other half were more
abstract or cognitive in nature (The bank ignored the pleas from her). 另一个 50 sensible
sentences were figurative. Half of the figurative sentences were idioms (The defense picked
holes in the argument), and half were metaphors related to physical action (The discovery lifted
this nation out of poverty). The task was originally designed to examine literal/figurative sen-
tence processing and action-relatedness (Fernandino et al., 2013). In the current group of
患者, t tests revealed that response times (RT) and accuracies (ACC) did not significantly
不同 (RT p = 0.35; ACC p = 0.24) between the literal (M RT = 4,802 多发性硬化症, SD RT = 963 多发性硬化症;
M ACC = 77.7%, SD ACC = 15.3%) and figurative conditions (M RT = 4,638 多发性硬化症, 标准差= 954 多发性硬化症;
M ACC = 80.9%, SD ACC = 14.5%), so the two conditions were combined. There is evidence
那, in sentences, high action-relatedness and figurative language may involve distributed
brain areas in addition to classic language regions (例如, Johari et al., 2021), but core
language-related regions (the focus of the current study) are still expected to be involved in
the majority of language processing (活页夹 & 德赛, 2011; Binds等人。, 2009), 尤其
after collapsing all sentence types together.
Sentences followed subject-verb-object order except for four sentences that lacked an
目的. All verbs were transitive or ambitransitive and were used transitively except for the four
aforementioned sentences. For sensible sentences, 25% of the subject nouns and 10% 的
object nouns were grammatically animate. For low-sensibility sentences, 28% of the subject
名词和 4% of the object nouns were grammatically animate. Low-sensibility sentences
were well formed grammatically but constructed such that the verb was incompatible with
one or both of its arguments (The taxpayer seized the planets with his small arm; 公司
twisted the shot in the dark; The tape rejected the air in the sky). Detecting these sentences as
nonsensible involved understanding the subject/object nouns, 动词, and their semantic as well
as grammatical relationships. Sensible and nonsense sentences were matched in length
语言的神经生物学
325
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
桌子 2. 方法 (and standard deviations) for characteristics of sensible and nonsense sentences
信件
Syllables
字
Sensible
43.8 (7.3)
11.1 (2.2)
7.9 (1.2)
Nonsense
44.5 (8.9)
10.6 (2.4)
7.8 (1.7)
Accuracy in healthy adults
0.91 (0.08)
0.91 (0.08)
t test p
0.61
0.25
0.84
0.54
(number of letters, number of syllables, and number of words) and difficulty, 如
response accuracy from a pilot study using healthy adults (桌子 2; Fernandino et al., 2013).
Control tasks were as follows.
Visual lexical decision
Visual lexical decision consisted of 80 动词, 80 名词, 和 160 phonologically plausible
pseudowords. Pseudowords were chosen from the English Lexicon Project (ELP; Balota
等人。, 2007). Words and pseudowords were matched in number of letters, bigram frequency,
orthographic neighborhood size, and visual lexical decision accuracy (桌子 3). Visual lexical
decision consisted of the presentation of a fixation cross (500 多发性硬化症), a mask (‘########’,
100 多发性硬化症), a prime (50 多发性硬化症), 面具 (100 多发性硬化症), followed by the target. The prime was the same
as the target word/pseudoword in capital letters for half of the stimuli, and a consonant string
also in capital letters for the other half. For the purposes of the present investigation, 我们不
investigate priming effects and collapse primed and unprimed trials together. 参与者是
instructed to indicate as quickly and as accurately as possible whether the target was a real
word or not by pressing one of two buttons. Participants had 5 s to respond.
Half of nouns were manipulable objects (the phone, 笔), while the other half were
concrete but comparatively nonmanipulable (海洋, the stadium), as determined by
body-object interaction ratings (Pexman et al., 2019). All were inanimate. Half of the verbs
referred to voluntary hand/arm actions (to pour, to pinch) while the other half referred to sen-
sory or cognitive concepts (to observe, to notice). All verbs were transitive or ambitransitive
except for four (5% of verb stimuli) that were intransitive. This task was originally designed to
examine priming effects, as well as effects of action-relatedness and manipulability (德赛
等人。, 2015). The current study focuses on core language-related regions that are expected
to be involved in lexical-semantic processing for many word types (活页夹 & 德赛, 2011;
Binds等人。, 2009), especially when collapsing across word categories. 有 160 试验,
桌子 3.
visual lexical decision task
Psycholinguistic variable means (and standard deviations) for words and nonwords in the
长度
Orthographic neighborhood
字
5.53 (1.48)
3.75 (4.85)
Nonwords
5.36 (1.74)
3.59 (3.92)
Bigram frequency
1,607.78 (713.06)
1,580.48 (738.27)
LD ACC
0.96 (0.05)
0.97 (0.03)
笔记. LD = Lexical decision measures from the ELP database.
t test p
0.51
0.81
0.81
0.33
326
语言的神经生物学
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
桌子 4. 方法, standard deviations (in parenthesis), and ranges for response times, 准确性, 和d 0 for all tasks
RT
ACC
d 0
Auditory sentence
感性
4,690 (920)
2,814–6,737
0.79 (0.14)
0.31–0.98
1.88 (1.09)
−0.86–4.15
Visual semantic
相似
2,944 (616)
1,849–4,030
0.78 (0.16)
0.45–0.98
–
Visual lexical
决定
1,015 (327)
543–2,029
0.96 (0.07)
0.7–1
3.5 (1.1)
0.2–5
笔记. Bolded boxes indicate the measures used in the neuroanatomical analyses.
Auditory word
理解
–
0.88 (0.19)
0.13–1
–
向前
digit span
–
6.6 (1.95)
0–12
–
divided equally between words and pseudowords, in both the verb and noun versions of the
visual lexical decision task. This task was the least demanding task in the battery with respect
to executive and semantic processing, as it merely required participants to recognize a word as
real or not, and there were no distractor items (reflected by high accuracies and low response
次; 桌子 4). Semantic demands for lexical decision are relatively low compared to the
other tasks included here, as participants do not need to explicitly access word meaning in
order to successfully complete the task.
Visual semantic similarity judgment
The visual semantic similarity judgment task consisted of sets of 240 verbs and 240 名词 (看
桌子 5 for lexical characteristics). Each set was organized into 80 verb and 80 noun triplets
这样, for each triplet, the target word was more similar in meaning to one of the two
选择 (例如, to thrill, to excite, to harm; bold indicates the target word). Similar to visual lex-
ical decision, the nouns were equally divided into manipulable and concrete nonmanipulable,
and the verbs into hand/arm actions and cognitive/sensory concepts. All nouns were inani-
伴侣. All verbs were transitive or ambitransitive except for two (2.5% of verbs). This task
was originally designed to detect deficits related to manipulability and action-relatedness in
患者 (Riccardi et al., 2019, 2020). Collapsing noun and verb performance together across
all categories provides a measure of general lexical-semantic processing and is expected to
桌子 5.
judgment tasks
Lexical characteristics of words in the visual lexical decision and semantic similarity
信件
Phonemes
Syllables
Log F
LD RT
LD ACC
词汇
决定
5.32
3.4
1.26
1.18
640.75
0.965
Semantic similarity
判断
5.40
4.32
1.47
1.09
655.5
0.945
笔记. Log F = logarithmic frequency, LD = Lexical decision measures from the ELP database.
语言的神经生物学
327
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
involve core lexical-semantic language regions. This task is also relatively executively
demanding, as participants must choose the correct response in the presence of a distractor
item that is from a similar semantic category, matching one of two words to the target instead
of making a binary judgment. These increased lexical-semantic and executive demands are
reflected by low accuracy scores and high response times compared to other control tasks
(桌子 4).
The sentence sensibility, visual lexical decision, and visual semantic similarity judgment
tasks were presented on a laptop PC running E-prime software (版本 1.2, Psychology Soft-
ware Tools, 公司). Participants indicated their response by pressing one of two buttons. 这
position of the bottom words was counterbalanced across participants. Participants could
use whichever hand they preferred and were asked to respond as quickly and accurately as
可能的. The words remained on the screen for 5 s, after which the next triplet was presented.
有 80 trials in both the verb and noun versions.
Auditory word comprehension
Auditory word comprehension was administered by a licensed speech-language pathologist as
part of the WAB (Kertesz, 2007). It consists of 60 real objects and pictures coming from 10
类别: real objects, drawn objects, 形式, 字母, 数字, 颜色, furniture, 身体部位,
手指, and right-left body parts. There are six stimuli per category. The speech-language
pathologist speaks aloud the name of one of the pictures/objects, and the participant must
point to the correct item. Participants are given a point for each item that they correctly point
到, for a maximum of 60 点. Auditory word comprehension requires participants to com-
prehend a spoken word, and then to correctly recognize its corresponding visual form. 不像
the semantic similarity judgment task, it does not require explicit access to deeper semantic
features in order to successfully complete the task.
Forward digit span
A series of digits (1 通过 9) were read aloud to the participant. The participant was
instructed to repeat as many of the digits as they could, 为了, with series length increasing
after successful repetition.
MRI数据获取
MRI data were obtained with a Siemens 3T Trio System with a 12-channel head coil and a
Siemens 3T Prisma System with a 20-channel coil. Participants underwent two anatomical
MRI sequences: (我) T1-weighted imaging sequence with a magnetization-prepared rapid-
gradient echo (MPRAGE) turbo field echo (TFE) sequence with voxel size = 1 MM3, 场地
的观点 (FOV) = 256 × 256 毫米, 192 sagittal slices, 9°翻转角, 重复时间 (tr) =
2,250 多发性硬化症, inversion time (TI) = 925 多发性硬化症, 回声时间 (这) = 4.15 多发性硬化症, generalized autocalibrat-
ing partial parallel acquisition (GRAPPA) = 2, 和 80 reference lines; 和 (二) T2-weighted
MRI with a 3D sampling perfection with application optimized contrasts by using different
flip angle evolutions protocol with the following parameters: 体素尺寸= 1 MM3, fov =
256 × 256 毫米, 160 sagittal slices, variable flip angle, TR = 3,200 多发性硬化症, = 212 多发性硬化症, 和
no slice acceleration. The same slice center and angulation were used as in the T1 sequence.
Diffusion tensor imaging (DTI) scans consisted of two scans with a 180° flip, TR = 4,987 多发性硬化症,
= 79.2 多发性硬化症, 矩阵= 90 × 90, fov = 207 × 207 MM2, 切片厚度= 2.3 毫米, 和
50 transversal slices.
语言的神经生物学
328
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
Preprocessing of Structural Images
Lesions were defined in native space by a neurologist (L. Bonilha) in MRIcron (Rorden et al.,
2012) on individual T2-weighted images. Preprocessing started with coregistration of the
T2-weighted images to match the T-weighted images, allowing the lesions to be aligned to
native T1 space. Images were warped to standard space using enantiomorphic (Nachev
等人。, 2008) segmentation-normalization (Ashburner & 弗里斯顿, 2005) custom Matlab script
(https://github.com/rordenlab/spmScripts/blob/master/nii_enat_norm.m) to warp images to an
age-appropriate template image found in the Clinical Toolbox for SPM (https://www.nitrc.org
/scm/?group_id=881). The normalization parameters were used to reslice the lesion into stan-
dard space using linear interpolation, with subsequent lesion maps stored at 1 × 1 × 1-mm
resolution and binarized using a 50% 临界点. (Because interpolation can lead to fractional
probabilities, this step confirms that each voxel is categorically either lesioned or unlesioned
without biasing overall lesion volume.) Normalized images were visually inspected to verify
质量.
Preprocessing of DTI Data
Diffusion data were processed in the method described in Bonilha et al. (2015). MRTrix tools
were used to perform Gibbs artifacts removal (Kellner et al., 2016) and de-noising (Veraart
等人。, 2016). FMRIB Software Library’s (FSL) TOPUP (Andersson et al., 2003) and eddy
(安德森 & 索蒂罗普洛斯, 2015) were used to attenuate spatial distortion. FSL’s dtifit was
used to compute tensors, 分数各向异性 (F A), and mean diffusivity maps, and bedpost
(Hernandez et al., 2013) was used to model fibers. As discussed in the previous section, the T1 scan
used Statistical Parametric Mapping’s (SPM; https://www.fil.ion.ucl.ac.uk/spm/) unified normaliza-
tion and segmentation. This allowed warping of atlases from standard space to the patient’s space.
This was warped to native diffusion space by nonlinearly warping the T1 scan to the FA map (哪个
has similar contrast). This allowed back-projection of our regions of interest (罗伊斯) into the native
diffusion space. 最后, probtrackx (Hernandez-Fernandez et al., 2019) quantified connectivity.
This evaluated the connectivity between each and every region in the atlas.
DTI Connectome Creation
As described in Gleichgerrcht et al. (2017), a unique probabilistic DTI connectome was con-
structed for every participant using the 189 cortical regions defined by the Johns Hopkins
大学 ( JHU) atlas (Faria et al., 2012; 莫里等。, 2005; Wakana等。, 2004), 结果
在一个 189 × 189 correlation matrix for each participant, where positive values signify greater
white matter connectivity of two regions. Every participant’s probabilistic white matter map
excluding the lesion was used as a mask for the estimation of their tractography. For each pair
地区, the number of streamlines arriving in one region when another region was seeded
was calculated, and the connectivity was defined as the average between the number of
streamlines arriving in region A when region B was seeded and the number of streamlines arriv-
ing in region B when region A was seeded. The connectivity between the regions was corrected
based on the sum of the volumes of the two regions to control for larger regions inherently
having a higher number of streamlines than smaller regions within the atlas. 结果
在一个 189 × 189 correlation matrix of weighted connections, which was used for CLSM.
利益区域
Nine ROIs (数字 1), based on the JHU atlas (Faria et al., 2012; 莫里等。, 2005; 理智
等人。, 2004), were used for lesion-deficit analysis. We analyzed several ROIs each from LIFC,
语言的神经生物学
329
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
数字 1. 罗伊斯 (左边; 最小. 8, 最大限度. 33) and voxelwise lesion incidence map (最小. 8, 最大限度. 40).
LATL, and LpT-iP. From LIFC/Broca’s area, we examined two ROIs: IFGoper and IFGtri. 从
LATL, we examined three ROIs: middle and superior temporal poles (MTGpole, STGpole) 和
the anterior portion of the inferior temporal gyrus (ITG). From LpT-iP, we examined four ROIs:
posterior middle and superior temporal gyri (PMTG, pSTG), 超边缘回 (SMG), 和
角回 (Ag).
Experimental Design and Statistical Analysis
Behavioral data
Our primary aim was to inspect the relative impairment of the sentence comprehension task
(auditory sentence sensibility) factoring out the contribution of other tasks (visual lexical deci-
锡安, visual semantic similarity judgment, auditory word comprehension, forward digit span).
For the auditory sentence sensibility and visual lexical decision tasks, d 0 was calculated as the
difference between the z transforms of the proportion of hits (IE。, when a sensible sentence
was identified as sensible) and the proportion of false alarms (IE。, when a nonsensible sen-
tence was identified as sensible). A d 0 score deviating from 0 for a participant reflects a sep-
aration between the number of correct hits and false alarms. For visual semantic similarity
judgment and auditory word comprehension, the proportion out of the total possible correct
was recorded. For forward digit span, the total number of digits correctly recalled, 为了,
被使用.
地区- and voxel-based lesion-symptom mapping
RLSM was used within the nine ROIs (described in Regions of Interest) to identify damage
related to greater impairment of the sentence comprehension task as compared to the other
four tasks by regressing out performance in one condition from the other using NiiStat software
(www.nitrc.org/projects/niistat/). RLSM measures the relationship between percentage of vox-
els damaged within an ROI and a behavioral measure. Nuisance regression used the
Freedman-Lane method (弗里德曼 & Lane, 1983), allowing for permutation-based control
for family-wise error (Winkler et al., 2014). RLSM results were corrected for multiple compar-
isons using permutation analysis ( p < 0.05, 1,000 permutations). Permutation analysis is a
Neurobiology of Language
330
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
o
_
a
_
0
0
0
6
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
nonparametric significance test that compares a test statistic to a null distribution that is
created by randomly permuting the real data (Baldo & Dronkers, 2018; Baldo et al., 2012;
Kimberg et al., 2007).
Given the theoretical importance of LIFC, to increase power to detect effects VLSM was
used within a restricted IFGoper and IFGtri ROI (combined into a single region for this analysis
only) to investigate whether damage to voxels within this region was associated with worse
sentence comprehension. VLSM binarily demarcates each voxel as either lesioned or
unlesioned and tests the probability that damage to a voxel is associated with behavioral per-
formance (Bates et al., 2003). VLSM results were thresholded at p < 0.001 voxel-wise and
cluster-corrected to p < 0.05 using permutation analysis as correction for multiple comparisons
(1,000 permutations). To improve power and minimize spatial bias, only voxels where at least
10% of patients had damage were considered (Baldo & Dronkers, 2018; Karnath et al., 2018).
Region and voxelwise lesion incidence maps showed that we had sufficient coverage in all
areas of interest (Figure 1).
Connectivity-based lesion symptom mapping
CLSM was used to investigate whether white matter connectivity between all regions included
in the LIFC, LATL, and LpT-iP network of interest was specifically predictive of sentence com-
prehension impairment compared to the other tasks using nuisance regression, as described
above. Left-to-left and left-to-right connections (117 in total) between the regions were consid-
ered to test for possible contributions from undamaged inter- or intra-hemispheric regions.
White matter connectivity strengths were used in a general linear model to predict task per-
formance. Alpha was set to 0.05, and significance was determined with permutation correc-
tion for multiple comparisons (1,000 permutations).
RESULTS
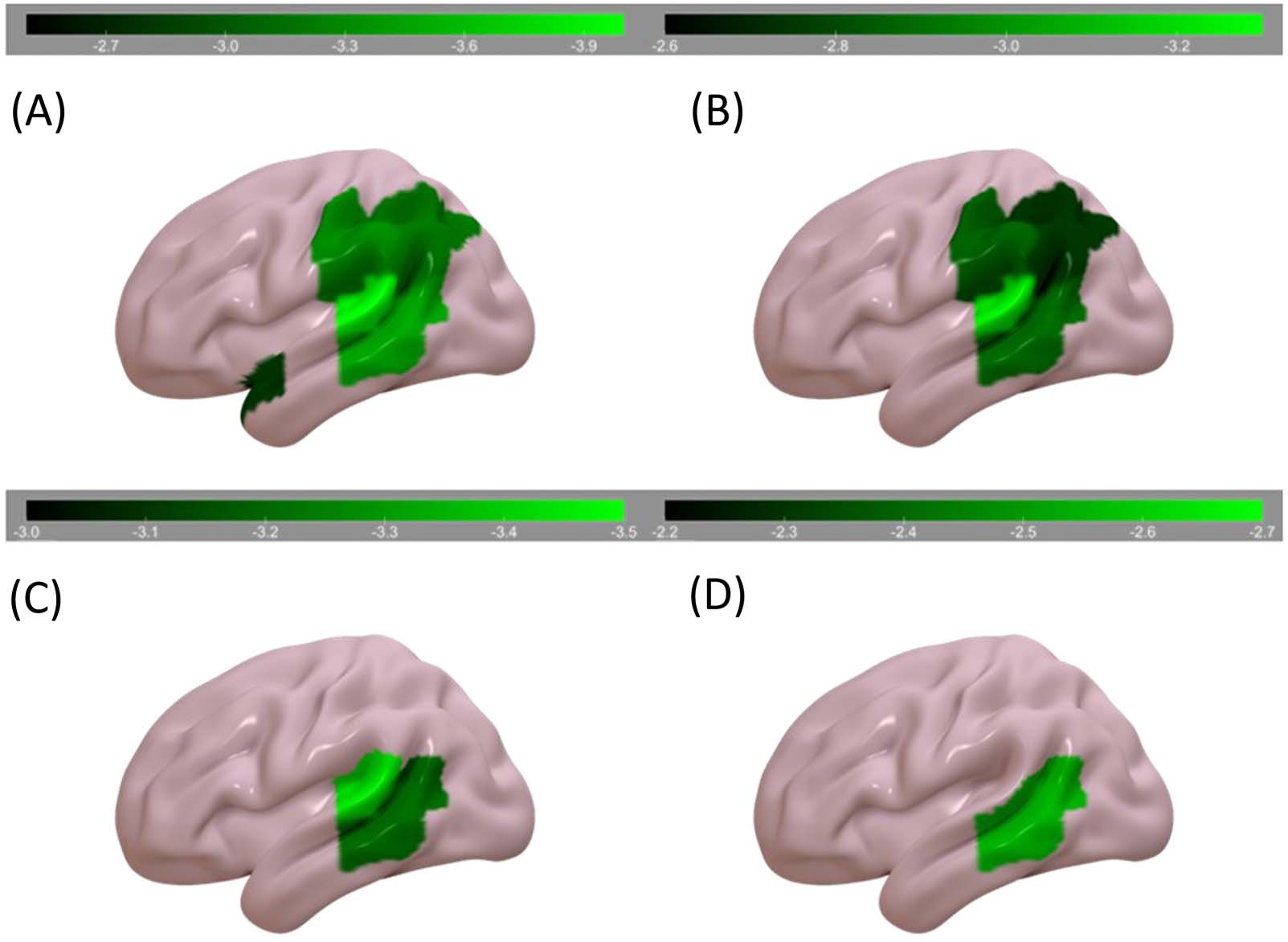
RLSM
A summary of the behavioral data can be found in Table 4. We first examined regions asso-
ciated with the auditory sentence sensibility task, without including the other behavioral tasks
as covariates. Auditory sentence sensibility performance was associated with percentage of
voxels damaged in STGpole, SMG, AG, pSTG, and pMTG (Table 6; Figure 2). It was not sig-
nificantly associated with damage in either of the LIFC ROIs.
Next, we individually included data from each task as a covariate, partially accounting for
potential contributions of auditory-verbal STM, lexical processing, executive function, seman-
tics, and input modality (Table 6; Figure 2). Worse auditory sentence sensibility performance,
controlling for forward digit span (auditory-verbal STM), was associated with percentage of
voxels damaged in the SMG, AG, pMTG, and pSTG. Controlling for visual lexical decision
(lexical processing with relatively low semantic and executive demands), it was associated
with percentage of voxels damaged in the pSTG and pMTG. Controlling for visual semantic
similarity judgment (lexical processing with relatively high semantic and executive demands),
it was associated with percentage of voxels damaged in the pMTG. No areas were significantly
associated with worse auditory sentence sensibility performance when controlling for auditory
word comprehension (lexical task in the auditory modality with moderate executive and
semantic demands).
Neurobiology of Language
331
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
o
_
a
_
0
0
0
6
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
Condition
Auditory sentence sensibility
Region
STGpole
z score
−2.7
Table 6.
Significant RLSM regions
Auditory sentence sensibility controlling for forward digit span
Auditory sentence sensibility controlling for visual lexical decision
Auditory sentence sensibility controlling for visual semantic similarity judgment
Note. Anatomical labels are according to the JHU atlas (Faria et al., 2012; Mori et al., 2005; Wakana et al., 2004).
AG
SMG
pMTG
pSTG
AG
SMG
pMTG
pSTG
pMTG
pSTG
pMTG
−3.0
−3.1
−3.4
−3.8
−2.7
−2.9
−2.9
−3.3
−3.2
−3.5
−2.5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
o
_
a
_
0
0
0
6
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
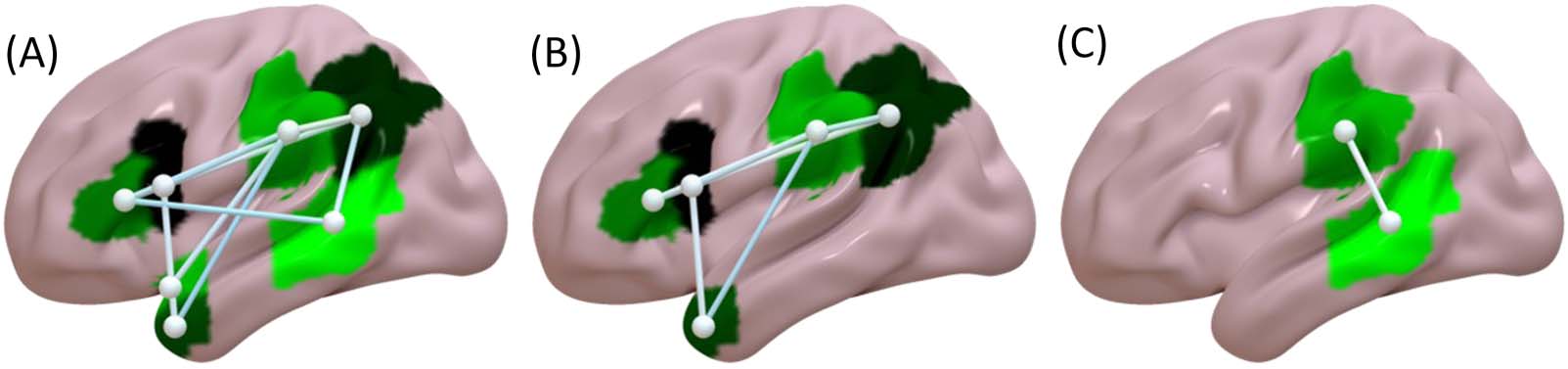
Figure 2. RLSM results (color bar represents z scores). Areas where damage was associated with: (A) worse sentence comprehension, (B)
worse sentence comprehension controlling for forward digit span, (C) worse sentence comprehension controlling for visual lexical decision,
and (D) worse sentence comprehension controlling for visual semantic similarity judgment.
Neurobiology of Language
332
Canonical sentence processing and the inferior frontal cortex
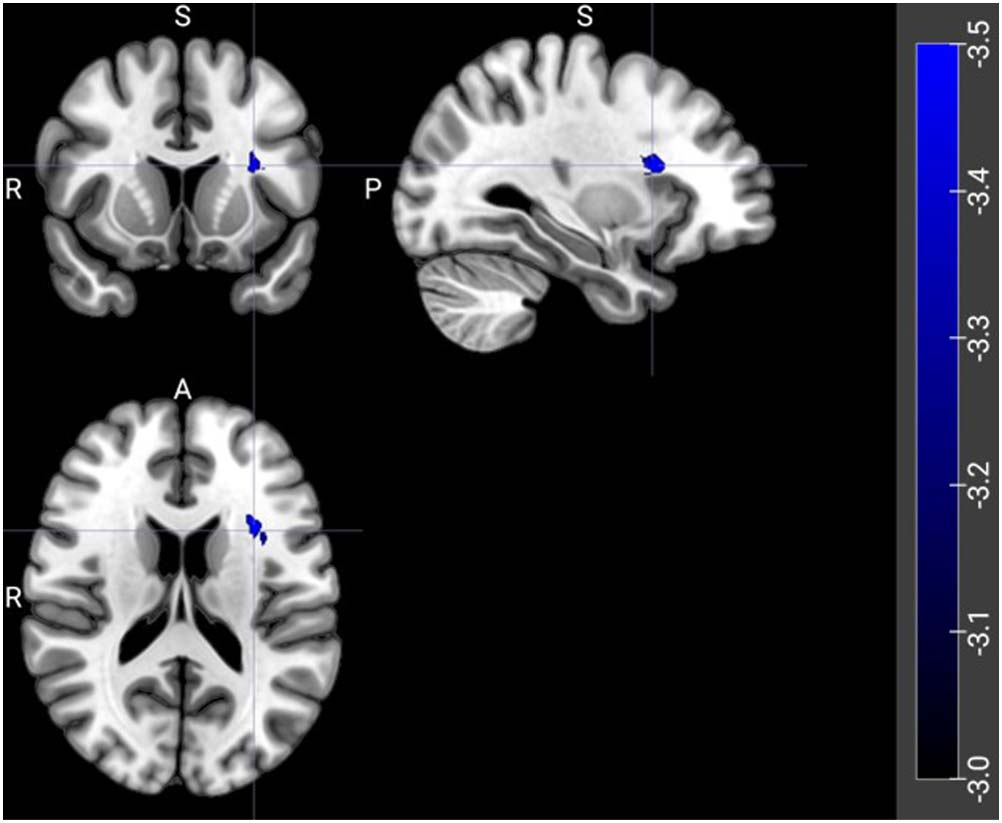
Table 7.
Significant VLSM peaks and Talairach coordinates
Location
IFGtri
Cluster size
(1 mm3 voxels)
364
Peak-z
score
−3.8
x
−27
y
18
z
16
Condition
Visual semantic similarity
judgment controlling for
auditory sentence sensibility
VLSM
VLSM, restricted to the LIFC for increased power, showed that no voxels were associated with
worse auditory sentence sensibility performance by itself, or when including any other tasks as
covariates. However, worse visual semantic similarity judgment performance, controlling for
auditory sentence sensibility, was associated with a cluster of voxels in the LIFC (Table 7;
Figure 3).
CLSM
Worse performance in the auditory sentence sensibility task, without including the other
behavioral tasks as covariates, was associated with disruption of 10 white matter connections
within the left hemisphere (Figure 4A). This included connections within and between LIFC,
LATL, and LpT-iP.
Worse auditory sentence sensibility performance, when including forward digit span,
visual lexical decision, or auditory word comprehension as nuisance covariates, was asso-
ciated with disruption of the same seven white matter connections within the left hemi-
sphere. This included connections within and between LIFC, LATL, and LpT-iP (Figure 4B).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
o
_
a
_
0
0
0
6
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. VLSM results for visual semantic similarity judgment controlling for auditory sentence
sensibility. Blue voxels represent where damage is associated with worse visual semantic similarity
judgment performance.
Neurobiology of Language
333
Canonical sentence processing and the inferior frontal cortex
(A) White matter connections where disruption was associated with worse auditory sentence sensibility performance. (B) White
Figure 4.
matter connections where disruption was associated with worse auditory sentence sensibility performance, controlling for forward digit span,
visual lexical decision, and auditory word comprehension. (C) White matter connections where disruption was associated with worse auditory
sentence sensibility performance, controlling for visual semantic similarity judgment. Colors are for display only.
Worse auditory sentence sensibility performance, controlling for visual semantic similarity
judgment, was associated with disruption of a single white matter connection within the left
hemisphere; SMG to pMTG (Figure 4C). CLSM results for all analyses are summarized in
Table 8.
Table 8.
Significant white matter connections. Disruption of these connections is associated with worse task performance.
Condition
Auditory sentence sensibility
Connection
IFGoper
IFGtri
SMG
AG
IFGoper
IFGtri
SMG
SMG
Auditory sentence sensibility controlling for forward digit span,
visual lexical decision, and auditory word comprehension
Auditory sentence sensibility controlling for visual semantic
similarity judgment
Neurobiology of Language
SMG
AG
MTGpole
SMG
AG
pMTG
AG
STGpole
MTGpole
pMTG
SMG
AG
MTGpole
SMG
AG
AG
MTGpole
pMTG
z score
3.7
3.9
4.0
3.8
3.8
3.6
4.1
4.0
3.6
3.8
2.8
3.1
3.0
3.0
2.9
2.9
2.7
2.9
334
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
o
_
a
_
0
0
0
6
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
DISCUSSION
By using tasks that vary in their executive, lexical, and phonological demands, we interrogated
the contributions of the LIFC, and two other language-related regions, in canonical sentence
comprehension while controlling for these related cognitive processes.
LIFC
Focusing on pars opercularis and triangularis, we examined the effects of damage to LIFC, and
also of lesions affecting the white matter connectivity of LIFC within the a priori language net-
work. RLSM and VLSM provided little evidence supporting the contribution of the LIFC to
canonical sentence comprehension. Damage to LIFC was not significantly associated with
worse sentence comprehension in any of the V/RLSM analyses. Results here are similar to sev-
eral prior studies. For example, in the classic investigation by Dronkers et al. (2004), damage
to BA 44/45 was not associated with sentence comprehension deficits for any of the several
included sentence types. Similar results were reported by Newhart et al. (2012) and Rogalsky
et al. (2018).
Here, we take these findings a step further by including a lexical task with relatively high
semantic and executive demands, visual semantic similarity judgment. A more restrictive
VLSM analysis, meant to increase power to detect effects within the LIFC, did not find a
significant association between LIFC damage and sentence comprehension, even when not
controlling for performance in the other tasks. However, damage to voxels within LIFC was
associated with worse performance in the visual semantic similarity judgment task even after
accounting for sentence comprehension performance. Visual semantic similarity judgment
was one of the most semantically and executively demanding tasks included in our battery,
suggesting a link between LIFC damage and impaired semantic access or control (Chiou et al.,
2018; Jackson, 2021; Whitney et al., 2011).
In contrast with the V/RLSM results, CLSM demonstrated that structural connectivity of the
LIFC with other regions in the a priori language network was related to sentence comprehen-
sion performance. Disruption of left-lateralized white matter connections within and between
the LIFC, LATL, and LpT-iP was associated with worse sentence comprehension, not control-
ling for other tasks. After controlling for visual lexical decision, forward digit span, and audi-
tory word comprehension, seven white matter connections, again consisting of links between
the left LIFC, LATL, and LpT-iP, remained significantly associated with sentence comprehen-
sion. These findings demonstrate that connections between multiple left-hemisphere brain
areas are important for canonical sentence comprehension, likely reflecting that successfully
comprehending a sentence requires the coordination of numerous cognitive processes. Impor-
tantly, the LIFC was part of this network after controlling for auditory word comprehension,
lexical access, and auditory-verbal STM. This suggests that the role of the LIFC within the sen-
tence comprehension network goes beyond those demands. These findings do not rule out the
contribution of the LIFC to these subprocesses, but they do suggest that the LIFC contributes to
canonical sentence comprehension in an additional way.
Using the visual semantic similarity judgment task as a covariate allowed control for seman-
tic and executive abilities, resulting in the absence of LIFC and LATL connections that were
uniquely associated with sentence comprehension. This suggests that the LIFC and LATL
connections are shared neural substrates for the sentence sensibility and semantic similarity
judgment tasks. Considering this, the LIFC and LATL may contribute to canonical sentence
comprehension via semantic knowledge/control and executive processes, likely associated
with the task demands of the sentence sensibility task. The sentence sensibility task oriented
Neurobiology of Language
335
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
o
_
a
_
0
0
0
6
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
attention to the meaningfulness of the sentences, requiring participants to identify instances
where the verb and subject/object fit together to create a coherent, sensible whole. Relating
to the hypotheses discussed in the Introduction, our findings closely align with the neuroim-
aging research suggesting that the LIFC is especially involved in sentence comprehension
when the task explicitly focuses attention on semantics or is semantically demanding (Hagoort
& Indefrey, 2014; Hasson et al., 2006; Love et al., 2006; Rogalsky & Hickok, 2009). In this
framework, the LIFC contributes to canonical sentence comprehension via executive task-
related demands, especially pertaining to detection of syntactic or semantic violations.
An alternative hypothesis partially supported by the current results is that the LIFC contrib-
utes to sentence processing via a variety of lexical-semantic operations, which are also needed
by semantic similarity judgment (Fiebach et al., 2002; Heim et al., 2009; Kotz et al., 2002,
2010; Ruff et al., 2008). However, LIFC connectivity was still associated with sentence com-
prehension after accounting for performance in two other lexical-semantic tasks that were
comparatively less demanding (visual lexical decision, auditory word comprehension), sug-
gesting that deeper task-related semantic analysis and control may be the main factors driving
LIFC connectivity involvment in the current sentence comprehension task.
The V/RLSM finding that damage to LIFC is not associated with canonical sentence com-
prehension, when contrasted with the positive CLSM finding that structural disconnection of
the LIFC from the language network is associated with worse performance, seems paradoxical
at first. One possible explanation is that the anatomy-function relationship within the LIFC has
been shown to be heterogenous between individuals (Amunts et al., 1999; Fedorenko &
Blank, 2020), which may impact power to detect group-level effects in fine-grained analyses
such as VLSM that require overlap at the voxelwise level. However, the negative results of the
RLSM anlaysis, which should not require fine-grained anatomy-function homogeneity within a
region to detect effects, makes this possibility unlikely. A more likely explanation is that, as
discussed in the Introduction, V/RLSM can only detect areas of overlapping necrosis/gliosis
but does not consider long-range white matter tract damage that can lead to cortical discon-
nection and associated behavioral impairments (Gleichgerrcht et al., 2017). CLSM comple-
ments V/RLSM by being able to detect effects of disrupted white matter connectivity resulting
from damage anywhere along white matter tracts that connect two grey matter regions, even if
those regions are spared by the lesion. In sum, the results here build upon previous studies
suggesting that disrupted LIFC connectivity has negative consequences for sentence compre-
hension (den Ouden et al., 2019; Fridriksson et al., 2018), and expand those findings by dem-
onstrating that these consequences extend even to relatively simple canonical sentences and
may be related to task-specific demands such as semantic analysis.
LpT-iP
Damage to the LpT-iP, especially the pMTG, was significantly associated with worse sentence
comprehension, even after controlling for performance on tasks that measured auditory-verbal
STM, semantic knowledge and control, and lexical access. This aligns with the previously dis-
cussed neuropsychological evidence suggesting that the pMTG is involved in sentence com-
prehension, even for canonical sentences (Magnusdottir et al., 2013; Rogalsky et al., 2018).
The pMTG has been identified as an important area for sentence processing, with possible
contributions being mapping wordforms to their meanings (“lexical interface;” Hickok &
Poeppel, 2004, 2007), processing syntax (Griffiths et al., 2013; Snijders et al., 2009), and rep-
resenting semantic knowledge (Binder & Desai, 2011; Binder et al., 2009). RLSM revealed that
damage to pMTG was associated with worse sentence comprehension when controlling for all
Neurobiology of Language
336
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
o
_
a
_
0
0
0
6
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
tasks except for auditory single-word comprehension, a task with relatively high phonological
demands. Additionally, CLSM revealed that pMTG connectivity was part of a larger network
subserving sentence comprehension, but this pMTG connectivity disappeared when control-
ling for performance in visual lexical decision, forward digit span, or auditory word compre-
hension. When controlling for visual semantic similarity judgment, disruption of a single
connection (pMTG to SMG) was associated with worse sentence comprehension. Considering
that the pMTG did not survive any analysis using auditory word comprehension as a covariate,
our results suggest that the pMTG may be a common neural substrate for auditory single-word
and sentence comprehension. This aligns with the “lexical interface” hypothesis (Hickok &
Poeppel, 2004, 2007), as well as studies suggesting that the pMTG may play a special role
in specifically auditory language comprehension (Pillay et al., 2017). However, given the
wealth of processes attributed to the pMTG, it is also possible that this area performs many
different functions or that it serves as a general hub of connectivity within the language com-
prehension network (Turken & Dronkers, 2011).
Previous studies have demonstrated that the pSTG plays a vital role in phonological pro-
cessing (Buchsbaum et al., 2001; Graves et al., 2008) and auditory-verbal STM (Leff et al.,
2009; Richardson et al., 2011), both of which are related to sentence comprehension. RLSM
in the current study found that pSTG damage was associated with worse sentence comprehen-
sion, but this association did not survive when controlling for auditory word comprehension
performance. This supports the well-established role of pSTG in acoustic-phonological
analysis (Hickok & Poeppel, 2000, 2007, 2016; Robson et al., 2012). A somewhat surprising
finding was that pSTG damage was associated with sentence comprehension even after con-
trolling for forward digit span, a measure of auditory-verbal STM. We do not interpret this as
evidence against a role of pSTG in auditory-verbal STM. Instead, this finding was possibly due
to forward digit span being a production task, meaning that some participants likely have dis-
played worse performance due to deficits in production despite relatively spared auditory-
verbal STM abilities, limiting power to detect effects in brain areas related to auditory-verbal
STM. Digit span is a widely used measure of auditory-verbal STM in neuropsychological stud-
ies of language comprehension (Caplan et al., 2016; Leff et al., 2009; Newhart et al., 2012;
Pettigrew & Hillis, 2014; Pisoni et al., 2019; Salis, 2012), but its production component is an
inherent limitation when comparing it directly to comprehension tasks. In sum, it is likely that
pSTG contributes to auditory sentence comprehension via both acoustic-phonological analy-
sis and auditory-verbal STM.
Similar to the pSTG, previous studies implicate the SMG in phonological processing,
auditory-verbal STM, and the processing of lexical and sub-lexical cues in both visual and
auditory modalities (Deschamps et al., 2014; Hartwigsen et al., 2010; Oberhuber et al.,
2016; Sliwinska et al., 2012). Damage to SMG was associated with worse sentence compre-
hension, and disrupted connectivity between SMG and AG, IFG, and MTGpole predicted
worse sentence comprehension after controlling for auditory-verbal STM, auditory word
comprehension, and visual lexical decision performance. We interpret the SMG connectivity
as reflecting the coordination of complex acoustic-phonological analysis in SMG with distrib-
uted lexical-semantic and executive processes required for successful completion of the sen-
tence sensibility task. Further support for this interpretation was found when these connections
to LATL, AG, and IFG did not survive after correcting for visual semantic similarity judgment, a
task with relatively high lexical-semantic and executive demands. Instead, a single connection
(SMG to pMTG) was associated with sentence comprehension. This likely reflects that the sen-
tence comprehension task was auditory, while the semantic similarity judgment was visual,
considering the previously mentioned studies indicating that SMG and pMTG play important
Neurobiology of Language
337
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
o
_
a
_
0
0
0
6
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
roles in acoustic-phonological processing and auditory language comprehension,
respectively.
Finally, the AG is a well-established part of the lexical-semantic system, being associated
with the representation of semantic knowledge across many different tasks and concept types
(Binder & Desai, 2011; Binder et al., 2009). RLSM revealed that damage to the AG was asso-
ciated with worse sentence comprehension, but this did not survive after controlling for any of
the lexical-semantic tasks (visual lexical decision, visual semantic similarity judgment, audi-
tory word comprehension). CLSM showed that disconnection of the AG from IFG and SMG
was associated with worse sentence comprehension after controlling for all tasks except for
visual semantic similarity judgment. The RLSM findings demonstrate that the AG is a shared
neural substrate for a variety of lexical-semantic tasks, while CLSM suggests that connectivity
from AG to the larger language comprehension network may be shared neural correlates spe-
cifically for explicitly semantic tasks (semantic similarity and sentence sensibility judgment).
LATL
RLSM revealed that damage to the STGpole was associated with worse sentence comprehen-
sion when not controlling for the other behavioral measures. This finding lends support to
theories that implicate the LATL in sentence processing (Brennan & Pylkkanen, 2017; Brennan
et al., 2012; Humphries et al., 2001, 2005). However, the fact that LATL damage was not asso-
ciated with worse sentence comprehension when adding the other behavioral measures as
covariates could reflect that the LATL contributes to multiple functions in addition to sentence
processing, such as lexical or semantic retrieval (Lambon Ralph et al., 2017; Mesulam et al.,
2013). It is also possible that regions specialized for sentence-related syntactic processing are
located more inferiorly in the LATL (Humphries et al., 2005), where relatively few patients had
damage, limiting power to detect effects. CLSM revealed that disconnection of the MTGpole
from the IFGoper and SMG was associated with worse sentence comprehension after control-
ling for all tasks except for visual semantic similarity judgment. These RLSM and CLSM results,
considering the already well-established role of the LATL in lexical-semantic access and
retrieval (Lambon Ralph et al., 2017; Mesulam et al., 2013), suggest that the LATL contributes
to canonical sentence comprehension through the representation or access of lexical-semantic
knowledge.
Limitations
This study leveraged preexisting data to interrogate the contribution of various language-
related areas to canonical sentence comprehension and related cognitive processes. The tasks
were not explicitly designed for the current investigation, and the retrospective nature of the
study inherently limits our control over the presence of certain psycholinguistic variables. For
example, while the auditory word comprehension task is a widely used clinical measure of
word recognition impairments, it contains words from semantic categories that were not
included in the other tasks (e.g., colors) and lacks a verb component. While our analysis
focused on core language-related areas that are expected to be involved in the processing
of many different types of words, it is possible that some effects observed here are associated
with those lexical-semantic differences between tasks.
The current study consisted of canonical declarative sentences. Inclusion of other types of
sentences, including noncanonical and more syntactically complex sentences would provide
additional information about neural substrates of sentence comprehension when syntactic
demands are manipulated. Additionally, it is possible that participants employed a number
Neurobiology of Language
338
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
o
_
a
_
0
0
0
6
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
of strategies to aid in their performance of the sentence sensibility task, such as using lexical-
semantic associations between words within the sentences, as this was not explicitly con-
trolled for, or by using animacy clues (e.g., does the subject noun typically perform the action
described by the verb). However, half of the sensible sentences were figurative (The bank
pulled the plug on the deal ). In these sentences, subject/object nouns and verbs are combined
in ways that are not sensible literally, and the subject and object nouns usually have low
lexical-semantic associations (e.g., bank and plug). This means that participants who heavily
relied on the aforementioned strategies would still likely perform poorly overall on the task.
Further, using the other lexical-semantic tasks included in the current study as covariates
would also help control for this possibility.
A single meaningfulness judgment task was used. Adding other tasks, such as sentence-
picture matching, could allow for direct comparisons of task type and demands. Additionally,
“executive control” involves many processes, and having multiple tasks that stress control in
conjunction with visual semantic similarity judgment to probe those specific processes would
be helpful in understanding the specific role of the LIFC. Finally, the spatial resolution of lesion
studies is inherently limited, and a fine-grained (e.g., millimeter scale) organization of function
is better studied with methods such as functional magnetic resonance imaging (fMRI).
Conclusion
VLSM and RLSM analyses suggested that the left pars opercularis and triangularis regions are
not vital for canonical sentence comprehension in the context of a sensibility judgment task.
LIFC damage was associated instead with impairments in a semantic similarity judgment task
that had high semantic and executive demands. However, the complementary CLSM method
revealed that disruption of left-lateralized white matter connections from LIFC to LATL and
LpT-iP was associated with worse sentence comprehension after controlling for performance
in tasks related to lexical access, auditory word comprehension, and auditory-verbal STM.
Semantic similarity judgment on single words explained similar variance to sentence sensibil-
ity judgment in LIFC connectivity, consistent with the hypothesis that LIFC’s contribution to
sentence comprehension is related to task-related processes when semantic demands are high.
Damage to the LpT-iP, especially pMTG, predicted worse sentence comprehension after con-
trolling for lexical access, semantic knowledge, and auditory-verbal STM, but not auditory
word comprehension. This supports previous studies demonstrating that the pMTG is a vital
region for sentence comprehension and suggests that the pMTG may contribute to canonical
sentence comprehension by acting as an auditory “lexical interface" or general hub for audi-
tory language comprehension.
ACKNOWLEDGMENTS
This work was supported by NIH/ NIDCD grants R01DC010783 (Rutvik H. Desai),
R56DC010783 (Rutvik H. Desai), R01DC017162 (Rutvik H. Desai), and P50DC014664 ( Julius
Fridriksson). We thank three reviewers for their insightful comments.
FUNDING INFORMATION
Rutvik H. Desai, National Institute on Deafness and Other Communication Disorders (https://
dx.doi.org/10.13039/100000055), Award ID: R01DC010783. Rutvik H. Desai, National Insti-
tute on Deafness and Other Communication Disorders (https://dx.doi.org/10.13039
/100000055), Award ID: R56DC010783. Rutvik H. Desai, National Institute on Deafness
and Other Communication Disorders (https://dx.doi.org/10.13039/100000055), Award ID:
Neurobiology of Language
339
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
o
_
a
_
0
0
0
6
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
R01DC017162. Julius Fridriksson, National Institute on Deafness and Other Communication
Disorders (https://dx.doi.org/10.13039/100000055), Award ID: P50DC014664.
AUTHOR CONTRIBUTIONS
Nicholas Riccardi: Conceptualization: Supporting; Formal analysis: Lead; Investigation: Lead;
Visualization: Lead; Writing – original draft: Lead; Writing – review & editing: Equal. Chris
Rorden: Data curation: Lead; Funding acquisition: Supporting; Methodology: Supporting;
Resources: Equal; Software: Lead; Writing – review & editing: Equal. Julius Fridriksson:
Conceptualization: Equal; Data curation: Supporting; Funding acquisition: Equal; Project
administration: Lead; Resources: Lead; Supervision: Supporting; Writing – review & editing:
Equal. Rutvik H. Desai: Conceptualization: Lead; Formal analysis: Supporting; Funding
acquisition: Equal; Investigation: Equal; Project administration: Supporting; Supervision: Lead;
Writing – original draft: Supporting; Writing – review & editing: Equal.
REFERENCES
Amunts, K., Schleicher, A., Burgel, U., Mohlberg, H., Uylings,
H. B., & Zilles, K. (1999). Broca’s region revisited: Cytoarchitec-
ture and intersubject variability. Journal of Comparative Neurol-
ogy, 412(2), 319–341. https://doi.org/10.1002/(SICI)1096-9861
(19990920)412:2<319::AID-CNE10>3.0.一氧化碳;2-7, 考研:
10441759
安德森, J. L. R。, Skare, S。, & Ashburner, J. (2003). How to correct
susceptibility distortions in spin-echo echo-planar images: Appli-
cation to diffusion tensor imaging. 神经影像, 20(2), 870–888.
https://doi.org/10.1016/S1053-8119(03)00336-7, 考研:
14568458
安德森, J. L. R。, & 索蒂罗普洛斯, S. 氮. (2015). Non-parametric
representation and prediction of single- and multi-shell
diffusion-weighted MRI data using Gaussian processes. 神经-
图像, 122, 166–176. https://doi.org/10.1016/j.neuroimage
.2015.07.067, 考研: 26236030
Ashburner, J。, & 弗里斯顿, K. J. (2005). Unified segmentation. 神经-
图像, 26(3), 839–851. https://doi.org/10.1016/j.neuroimage
.2005.02.018, 考研: 15955494
大胆的, J. 五、, & drungers, 氮. F. (2018). Lesion studies. 在一个. 中号. 乙.
de Groot & 磷. 哈科特 (编辑。), Research methods in psycholin-
guistics and the neurobiology of language: A practical guide
(PP. 310–329). 威利.
大胆的, J. 五、, 威尔逊, S. M。, & drungers, 氮. (2012). 揭开
neural substrates of language: A voxel-based lesion-symptom
mapping approach. 在米. Faust (埃德。), The handbook of the neu-
ropsychology of language (PP. 582–594). Wiley-Blackwell.
https://doi.org/10.1002/9781118432501.ch28
Balota, D. A。, 是的, 中号. J。, Cortese, 中号. J。, 和记黄埔, K. A。, Kessler,
B., Loftis, B., Neely, J. H。, 纳尔逊, D. L。, 辛普森, G. B., &
Treiman, 右. (2007). The English lexicon project. Behavior Research
方法, 39(3), 445–459. https://doi.org/10.3758/BF03193014,
考研: 17958156
贝茨, E., 威尔逊, S. M。, 塞金, A. P。, 迪克, F。, 塞雷诺, 中号. 我。,
骑士, 右. T。, & drungers, 氮. F.
(2003). Voxel-based
lesion-symptom mapping. 自然神经科学, 6(5), 448–450.
https://doi.org/10.1038/nn1050, 考研: 12704393
活页夹, J. R。, & 德赛, 右. H. (2011). The neurobiology of semantic
记忆. 认知科学的趋势, 15(11), 527–536. https://
doi.org/10.1016/j.tics.2011.10.001, 考研: 22001867
活页夹, J. R。, 德赛, 右. H。, 格雷夫斯, 瓦. W., & Conant, L. L. (2009).
语义系统在哪里? 批判性审查和
荟萃分析 120 功能性神经影像学研究. 大脑
皮质, 19(12), 2767–2796. https://doi.org/10.1093/cercor
/BHP055, 考研: 19329570
Bonilha, L。, & Fridriksson, J. (2009). Subcortical damage and white
matter disconnection associated with non-fluent speech. 脑,
132(6), Article e108. https://doi.org/10.1093/ brain/awn200,
考研: 18723562
Bonilha, L。, Gleichgerrcht, E., Fridriksson, J。, Rorden, C。, Breedlove,
J. L。, Nesland, T。, Paulus, W., 头盔, G。, & Focke, 氮. K. (2015).
Reproducibility of the structural brain connectome derived from
diffusion tensor imaging. PLOS一个, 10(8), Article e0135247.
https://doi.org/10.1371/journal.pone.0135247, 考研:
26332788
Bonilha, L。, Nesland, T。, Rorden, C。, Fillmore, P。, Ratnayake, 右. P。,
& Fridriksson, J. (2014). Mapping remote subcortical ramifica-
tions of injury after ischemic strokes. Behavioral Neurology,
2014, 文章 215380. https://doi.org/10.1155/2014/215380,
考研: 24868120
Bonilha, L。, Rorden, C。, & Fridriksson, J. (2014). Assessing the clin-
ical effect of residual cortical disconnection after ischemic
strokes. Stroke, 45, 988–993. https://doi.org/10.1161
/STROKEAHA.113.004137, 考研: 24619391
Brennan, J. [R。], 尼尔, Y。, Hasson, U。, 马拉奇, R。, 歌曲, D. J。, &
Pylkkanen, L. (2012). Syntactic structure building in the anterior
temporal lobe during natural story listening. 大脑和语言,
120(2), 163–173. https://doi.org/10.1016/j.bandl.2010.04.002,
考研: 20472279
Brennan, J. R。, & Pylkkanen, L. (2017). MEG evidence for incre-
mental sentence composition in the anterior temporal lobe. 齿轮-
nitive Science, 41(S6), 1515–1531. https://doi.org/10.1111/cogs
.12445, 考研: 27813182
黄杨木, 乙. R。, 希科克, G。, & 汉弗莱斯, C. (2001). Role of left
posterior superior temporal gyrus in phonological processing for
speech perception and production. 认知科学, 25(5),
663–678. https://doi.org/10.1207/s15516709cog2505_2
Caplan, D ., Michaud, J。, Hufford, R。, & 马克里斯, 氮. (2016). Deficit-
lesion correlations in syntactic comprehension in aphasia. 脑
and Language, 152, 14–27. https://doi.org/10.1016/j.bandl.2015
.10.005, 考研: 26688433
Caramazza, A。, Capasso, R。, Capitani, E., & Miceli, G. (2005). Pat-
terns of comprehension performance in agrammatic Broca’s
aphasia: A test of the trace deletion hypothesis. Brain and
语言的神经生物学
340
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
语言, 94(1), 43–53. https://doi.org/10.1016/j.bandl.2004.11
.006, 考研: 15896382
Catani, M。, Dell’Acqua, F。, 比兹, A。, Forkel, S. J。, 威廉姆斯, S. C。,
西蒙斯, A。, 墨菲, D. G。, & Thiebaut de Schotten, 中号. (2012).
Beyond cortical localization in clinico-anatomical correlation.
皮质, 48(10), 1262–1287. https://doi.org/10.1016/j.cortex
.2012.07.001, 考研: 22995574
Catani, M。, & ffytche, D. H. (2005). The rises and falls of discon-
nection syndromes. 脑, 128(10), 2224–2239. https://doi.org
/10.1093/brain/awh622, 考研: 16141282
Chiou, R。, 汉弗莱斯, G. F。, 荣格, J。, & Lambon Ralph, 中号. A.
(2018). Controlled semantic cognition relies upon dynamic and
flexible interactions between the executive ‘semantic control’
and hub-and-spoke ‘semantic representation’ systems. 皮质,
103, 100–116. https://doi.org/10.1016/j.cortex.2018.02.018,
考研: 29604611
den Ouden, D. B., Malyutina, S。, Basilakos, A。, Bonilha, L。,
Gleichgerrcht, E., Yourganov, G。, Hillis, A. E., 希科克, G。,
Rorden, C。, & Fridriksson, J. (2019). Cortical and structural-
connectivity damage correlated with impaired syntactic process-
ing in aphasia. 人脑图谱, 40(7), 2153–2173.
https://doi.org/10.1002/hbm.24514, 考研: 30666767
德赛, 右. H。, Herter, T。, Riccardi, N。, Rorden, C。, & Fridriksson, J.
(2015). Concepts within reach: Action performance predicts
action language processing in stroke. 神经心理学, 71,
217–224. https://doi.org/10.1016/j.neuropsychologia.2015.04
.006, 考研: 25858602
德赛, 右. H。, & Riccardi, 氮. (2021). Cognitive neuroscience of
语言. 在w. 徐 & J. 右. 泰勒 (编辑。), The Routledge handbook
of cognitive linguistics (PP. 615–642). 劳特利奇. https://doi.org
/10.4324/9781351034708-42
Deschamps, 我。, 鲍姆, S. R。, & Gracco, V. L. (2014). On the role of
the supramarginal gyrus in phonological processing and verbal
工作记忆: Evidence from rTMS studies. Neuropsycholo-
gia, 53, 39–46. https://doi.org/10.1016/j.neuropsychologia.2013
.10.015, 考研: 24184438
drungers, 氮. F。, Plaisant, 奥。, Iba-Zizen, 中号. T。, & Cabanis, 乙. A.
(2007). Paul Broca’s historic cases: High resolution MR imaging
of the brains of Leborgne and Lelong. 脑, 130(5), 1432–1441.
https://doi.org/10.1093/brain/awm042, 考研: 17405763
drungers, 氮. F。, Wilkins, D. P。, Van Valin, 右. D ., 小。, Redfern, 乙. B.,
& Jaeger, J. J. (2004). Lesion analysis of the brain areas involved
in language comprehension. 认识, 92(1–2), 145–177.
https://doi.org/10.1016/j.cognition.2003.11.002, 考研:
15037129
Fadiga, L。, Craighero, L。, & D’Ausilio, A. (2009). Broca’s area in
语言, 行动, 和音乐. Annals of the New York Academy
科学系, 1169(1), 448–458. https://doi.org/10.1111/j.1749
-6632.2009.04582.X, 考研: 19673823
Faria, A. 五、, 乔尔, S. E., 张, Y。, Oishi, K., van Zjil, 磷. C。, 磨坊主,
中号. 我。, Pekar, J. J。, & 森, S. (2012). Atlas-based analysis of
resting-state functional connectivity: Evaluation for reproducibil-
ity and multi-modal anatomy-function correlation studies. 新-
Kingage, 61(3), 613–621. https://doi.org/10.1016/j.neuroimage
.2012.03.078, 考研: 22498656
Fedorenko, E., & 空白的, 我. A. (2020). Broca’s area is not a natural
种类. 认知科学的趋势, 24(4), 270–284. https://doi.org
/10.1016/j.tics.2020.01.001, 考研: 32160565
费尔南迪诺, L。, Conant, L. L。, 活页夹, J. R。, Blindauer, K., Hiner, B.,
Spangler, K., & 德赛, 右. H. (2013). Where is the action? 行动
sentence processing in Parkinson’s disease. 神经心理学,
51(8), 1510–1517. https://doi.org/10.1016/j.neuropsychologia
.2013.04.008, 考研: 23624313
Fiebach, C. J。, 弗里德里克, A. D ., 穆勒, K., & 来自Cramon, D. 是.
(2002). fMRI evidence for dual routes to the mental lexicon in
visual word recognition. 认知神经科学杂志,
14(1), 11–23. https://doi.org/10.1162/089892902317205285,
考研: 11798383
弗里德曼, D. J。, & Lane, D. (1983). A nonstochastic interpretation
of reported significance levels. Journal of Business & 经济的
统计数据, 1(4), 292–298. https://doi.org/10.2307/1391660
Fridriksson, J。, Bonilha, L。, & Rorden, C. (2007). Severe Broca’s
aphasia without Broca’s area damage. Behavioural Neurology,
18(4), 文章 785280. https://doi.org/10.1155/2007/785280,
考研: 18430982
Fridriksson, J。, den Ouden, D. B., Hillis, A. E., 希科克, G。, Rorden,
C。, Basilakos, A。, Yourganov, G。, & Bonilha, L. (2018). Anatomy
of aphasia revisited. 脑, 141(3), 848–862. https://doi.org/10
.1093/brain/awx363, 考研: 29360947
Fridriksson, J。, Fillmore, P。, 郭, D ., & Rorden, C. (2015). Chronic
Broca’s aphasia is caused by damage to Broca’s and Wernicke’s
地区. 大脑皮层, 25(12), 4689–4696. https://doi.org/10
.1093/cercor/bhu152, 考研: 25016386
弗里德里克, A. D. (2012). The cortical language circuit: From audi-
tory perception to sentence comprehension. 认知趋势
科学, 16(5), 262–268. https://doi.org/10.1016/j.tics.2012.04
.001, 考研: 22516238
Gleichgerrcht, E., Fridriksson, J。, Rorden, C。, & Bonilha, L. (2017).
Connectome-based lesion-symptom mapping (CLSM): A novel
approach to map neurological function. 神经影像: Clinical,
16, 461–467. https://doi.org/10.1016/j.nicl.2017.08.018,
考研: 28884073
Graessner, A。, Zaccarella, E., & Hartwigsen, G. (2021). Differential
contributions of left-hemispheric language regions to basic seman-
tic composition. Brain Structure and Function, 226(2), 501–518.
https://doi.org/10.1007/s00429-020-02196-2, 考研: 33515279
格雷夫斯, 瓦. W., Grabowski, 时间. J。, 梅塔, S。, & 古普塔, 磷. (2008). 这
left posterior superior temporal gyrus participates specifically in
accessing lexical phonology. 认知神经科学杂志,
20(9), 1698–1710. https://doi.org/10.1162/jocn.2008.20113,
考研: 18345989
格里菲斯, J. D ., Marslen-Wilson, 瓦. D ., Stamatakis, 乙. A。, & 泰勒,
L. K. (2013). Functional organization of the neural language sys-
TEM: Dorsal and ventral pathways are critical for syntax. 大脑
皮质, 23(1), 139–147. https://doi.org/10.1093/cercor/bhr386,
考研: 22275482
哈科特, P。, & Indrey, 磷. (2014). The neurobiology of language
beyond single words. 神经科学的年度审查, 37,
347–362. https://doi.org/10.1146/annurev-neuro-071013
-013847, 考研: 24905595
Hartwigsen, G。, Baumgaertner, A。, 价格, C. J。, Koehnke, M。, Ulmer,
S。, & Siebner, H. 右. (2010). Phonological decisions require both
the left and right supramarginal gyri. 国家会议录
科学院, 107(38), 16494–16499. https://doi.org/10
.1073/pnas.1008121107, 考研: 20807747
Hasson, U。, Nusbaum, H. C。, & 小的, S. L. (2006). Repetition sup-
pression for spoken sentences and the effect of task demands.
认知神经科学杂志, 18(12), 2013–2029. https://
doi.org/10.1162/jocn.2006.18.12.2013, 考研: 17129188
Heim, S。, Eickhoff, S. B., 弗里德里克, A. D ., & Amunts, K. (2009). 左边
cytoarchitectonic area 44 supports selection in the mental lexi-
con during language production. Brain Structure and Function,
213(4–5), 441–456. https://doi.org/10.1007/s00429-009-0213-9,
考研: 19621242
埃尔南德斯, M。, Guerrero, G. D ., Cecilia, J. M。, 加西亚, J. M。,
Inuggi, A。, jbabdi, S。, 贝伦斯, 时间. E., & 索蒂罗普洛斯, S. 氮.
语言的神经生物学
341
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
(2013). Accelerating fibre orientation estimation from diffusion
weighted magnetic resonance imaging using GPUs. PLOS
ONE, 8(4), Article e61892. https://doi.org/10.1371/journal.pone
.0061892, 考研: 23658616
Hernandez-Fernandez, M。, Reguly, 我。, jbabdi, S。, 贾尔斯, M。, 史密斯, S。,
& 索蒂罗普洛斯, S. 氮. (2019). Using GPUs to accelerate computa-
tional diffusion MRI: From microstructure estimation to tractogra-
phy and connectomes. 神经影像, 188, 598–615. https://土井
.org/10.1016/j.neuroimage.2018.12.015, 考研: 30537563
希科克, G。, & 船尾, D. (2000). Towards a functional neuroanat-
omy of speech perception. 认知科学的趋势, 4(4),
131–138. https://doi.org/10.1016/s1364-6613(00)01463-7,
考研: 10740277
希科克, G。, & 船尾, D. (2004). Dorsal and ventral streams: A
framework for understanding aspects of the functional anatomy
语言的. 认识, 92(1–2), 67–99. https://doi.org/10.1016/j
.cognition.2003.10.011, 考研: 15037127
希科克, G。, & 船尾, D. (2007). The cortical organization of
speech processing. 自然评论神经科学, 8(5), 393–402.
https://doi.org/10.1038/nrn2113, 考研: 17431404
希科克, G。, & 船尾, D. (2016). The neural basis of speech
洞察力. 在g. 希科克 & S. 小的 (编辑。), 神经生物学
语言 (PP. 299–310). 爱思唯尔. https://doi.org/10.1016/B978
-0-12-407794-2.00025-0
汉弗莱斯, C。, 活页夹, J. R。, Medler, D. A。, & Liebenthal, 乙. (2006).
Syntactic and semantic modulation of neural activity during
auditory sentence comprehension. Journal of Cognitive Neuro-
科学, 18(4), 665–679. https://doi.org/10.1162/jocn.2006.18.4
.665, 考研: 16768368
汉弗莱斯, C。, 爱, T。, Swinney, D ., & 希科克, G. (2005).
Response of anterior temporal cortex to syntactic and prosodic
manipulations during sentence processing. 人脑图谱-
平, 26(2), 128–138. https://doi.org/10.1002/ hbm.20148,
考研: 15895428
汉弗莱斯, C。, Willard, K., 黄杨木, B., & 希科克, G. (2001).
Role of anterior temporal cortex in auditory sentence compre-
hension: An fMRI study. NeuroReport, 12(8), 1749–1752.
https://doi.org/10.1097/00001756-200106130-00046, 考研:
11409752
杰克逊, 右. L. (2021). The neural correlates of semantic control
revisited. 神经影像, 224, 文章 117444. https://doi.org/10
.1016/j.neuroimage.2020.117444, 考研: 33059049
Johari, K., Riccardi, N。, Malyutina, S。, Modi, M。, & 德赛, 右. H.
(2021). HD-tDCS over motor cortex facilitates figurative and lit-
eral action sentence processing. 神经心理学, 159, 文章
107955. https://doi.org/10.1016/j.neuropsychologia.2021
.107955, 考研: 34252418
Karnath, H. 奥。, Sperber, C。, & Rorden, C. (2018). Mapping human
brain lesions and their functional consequences. 神经影像,
165, 180–189. https://doi.org/10.1016/j.neuroimage.2017.10
.028, 考研: 29042216
凯尔纳, E., 迪塔尔, B., 基谢列夫, V. G。, & 旅行过, 中号. (2016). Gibbs-
ringing artifact removal based on local subvoxel-shifts. Magnetic
Resonance in Medicine, 76(5), 1574–1581. https://doi.org/10
.1002/mrm.26054, 考研: 26745823
Kemmerer, D. (2021). Cognitive neuroscience of language (2nd 版。).
Psychology Press.
Kertesz, A. (2007). Western Aphasia Battery—Revised. 皮尔逊.
https://doi.org/10.1037/t15168-000
Kimberg, D. Y。, Coslett, H. B., & 施瓦茨, 中号. F. (2007). Power in
voxel-based lesion-symptom mapping. Journal of Cognitive Neu-
roscience, 19(7), 1067–1080. https://doi.org/10.1162/jocn.2007
.19.7.1067, 考研: 17583984
Kinno, R。, Muragaki, Y。, Hori, T。, Maruyama, T。, 川村, M。, &
Sakai, K. L. (2009). Agrammatic comprehension caused by a gli-
oma in the left frontal cortex. 大脑和语言, 110(2), 71–80.
https://doi.org/10.1016/j.bandl.2009.05.001, 考研:
19573900
(2014). Differential
Kinno, R。, ohta, S。, Muragaki, Y。, Maruyama, T。, & Sakai,
K. L.
three syntax-
related networks induced by a left frontal glioma. 脑, 137(4),
1193–1212. https://doi.org/10.1093/ brain/awu013, 考研:
24519977
reorganization of
婴儿床, S. A。, Cappa, S. F。, 来自Cramon, D. Y。, & 弗里德里克, A. D.
(2002). Modulation of the lexical-semantic network by auditory
semantic priming: An event-related functional MRI study. 神经-
图像, 17(4), 1761–1772. https://doi.org/10.1006/nimg.2002
.1316, 考研: 12498750
婴儿床, S. A。, D’Ausilio, A。, Raettig, T。, Begliomini, C。, Craighero, L。,
Fabbri-Destro, M。, Zingales, C。, Haggard, P。, & Fadiga, L. (2010).
Lexicality drives audio-motor transformations in Broca’s area.
大脑和语言, 112(1), 3–11. https://doi.org/10.1016/j
.bandl.2009.07.008, 考研: 19698980
Kristinsson, S。, Thors, H。, Yourganov, G。, Magnusdottir, S。, Hjaltason,
H。, 斯塔克, 乙. C。, Basilakos, A。, den Ouden, D. B., Bonilha, L。,
Rorden, C。, 希科克, G。, Hillis, A。, & Fridriksson, J. (2020). 脑
damage associated with impaired sentence processing in acute
aphasia. 认知神经科学杂志, 32(2), 256–271.
https://doi.org/10.1162/jocn_a_01478, 考研: 31596169
Lambon Ralph, 中号. A。, Jefferies, E., 帕特森, K., & 罗杰斯, 时间. 时间.
(2017). The neural and computational bases of semantic cogni-
的. 自然评论神经科学, 18(1), 42–55. https://doi.org
/10.1038/nrn.2016.150, 考研: 27881854
Leff, A. P。, Schofield, 时间. M。, Crinion, J. T。, Seghier, 中号. L。, Grogan, A。,
绿色的, D. W., & 价格, C. J. (2009). The left superior temporal
gyrus is a shared substrate for auditory short-term memory and
speech comprehension: 证据来自 210 patients with stroke.
脑, 132(12), 3401–3410. https://doi.org/10.1093/ brain
/awp273, 考研: 19892765
爱, T。, Haist, F。, Nicol, J。, & Swinney, D. (2006). A functional
neuroimaging investigation of the roles of structural complexity
and task-demand during auditory sentence processing. 皮质,
42(4), 577–590. https://doi.org/10.1016/s0010-9452(08)70396-4,
考研: 16881268
Lukic, S。, 汤普森, C. K., Barbieri, E., Chiappetta, B., Bonakdarpour,
B., Kiran, S。, Rapp, B., 帕里什, 时间. B., & Caplan, D. (2021).
Common and distinct neural substrates of sentence production
and comprehension. 神经影像, 224, 文章 117374. https://
doi.org/10.1016/j.neuroimage.2020.117374, 考研:
32949711
Magnusdottir, S。, Fillmore, P。, den Ouden, D. B., Hjaltason, H。,
Rorden, C。, Kjartansson, 奥。, Bonilha, L。, & Fridriksson, J.
(2013). Damage to left anterior temporal cortex predicts impair-
ment of complex syntactic processing: A lesion-symptom map-
ping study. 人脑图谱, 34(10), 2715–2723. https://
doi.org/10.1002/hbm.22096, 考研: 22522937
匹配, W., Hammerly, C。, & 刘, 乙. (2017). The role of the IFG
and pSTS in syntactic prediction: Evidence from a parametric
study of hierarchical structure in fMRI. 皮质, 88, 106–123.
https://doi.org/10.1016/j.cortex.2016.12.010, 考研:
28088041
Mazoyer, 乙. M。, Tzourio, N。, Frak, 五、, Syrota, A。, Murayama, N。,
Levrier, 奥。, Salamon, G。, 德阿内, S。, 科恩, L。, & Mehler, J.
(1993). The cortical representation of speech. Journal of Cogni-
tive Neuroscience, 5(4), 467–479. https://doi.org/10.1162/jocn
.1993.5.4.467, 考研: 23964919
语言的神经生物学
342
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
Mesulalam, 中号. M。, Wieneke, C。, Hurley, R。, Rademaker, A。, 汤普森,
C. K., Weintraub, S。, & Rogalski, 乙. J. (2013). Words and objects at
the tip of the left temporal lobe in primary progressive aphasia.
脑, 136(2), 601–618. https://doi.org/10.1093/brain/aws336,
考研: 23361063
森, S。, 理智, S。, 范·齐尔, 磷. C. M。, & Nagae-Poetscher, L. 中号.
(2005). MRI atlas of human white matter (1st ed.). 爱思唯尔.
Nachev, P。, Coulthard, E., Jager, H. R。, Kennard, C。, & Husain, 中号.
(2008). Enantiomorphic normalization of focally lesioned brains.
神经影像, 39(3), 1215–1226. https://doi.org/10.1016/j
.neuroimage.2007.10.002, 考研: 18023365
Newhart, M。, Trupe, L. A。, 戈麦斯, Y。, Cloutman, L。, Molitoris, J. J。,
戴维斯, C。, Leigh, R。, Gottesman, 右. F。, 种族, D ., & Hillis, A. 乙.
(2012). Asyntactic comprehension, 工作记忆, and acute
ischemia in Broca’s area versus angular gyrus. 皮质, 48(10),
1288–1297. https://doi.org/10.1016/j.cortex.2011.09.009,
考研: 22079684
Oberhuber, M。, 希望, 时间. 中号. H。, Seghier, 中号. L。, Parker Jones, 奥。,
Prejawa, S。, 绿色的, D. W., & 价格, C. J. (2016). Four functionally
distinct regions in the left supramarginal gyrus support word pro-
cessing. 大脑皮层, 26(11), 4212–4226. https://doi.org/10
.1093/cercor/bhw251, 考研: 27600852
Pettigrew, C。, & Hillis, A. 乙. (2014). Role for memory capacity in
句子理解: Evidence from acute stroke. Aphasiol-
奥吉, 28(10), 1258–1280. https://doi.org/10.1080/02687038.2014
.919436, 考研: 25221377
Pexman, 磷. M。, Muraki, E., Sidhu, D. M。, Siakaluk, 磷. D ., & 是的,
中号. J. (2019). Quantifying sensorimotor experience: Body-object
interaction ratings for more than 9,000 English words. 行为
研究方法, 51(2), 453–466. https://doi.org/10.3758
/s13428-018-1171-z, 考研: 30484218
Pillay, S. B., 活页夹, J. R。, 汉弗莱斯, C。, 总的, 瓦. L。, & 书,
D. S. (2017). Lesion localization of speech comprehension defi-
cits in chronic aphasia. 神经病学, 88(10), 970–975. https://
doi.org/10.1212/ wnl.0000000000003683, 考研:
28179469
Pisoni, A。, Mattavelli, G。, Casarotti, A。, Comi, A。, 里瓦, M。, Bello,
L . , & P a p a g n o , C .
( 2 0 1 9 ) . T h e n e u r a l c o r r e l a t e s o f
a u d i t o r y – v e r b a l s h o r t – t e r m m e m o r y : A v o x e l – b a s e d
lesion-symptom mapping study on 103 patients after glioma
removal. Brain Structure and Function, 224(6), 2199–2211.
https://doi.org/10.1007/s00429-019-01902-z, 考研:
31177297
Riccardi, N。, Yourganov, G。, Rorden, C。, Fridriksson, J。, & 德赛,
右. H. (2019). Dissociating action and abstract verb comprehen-
sion post-stroke. 皮质, 120, 131–146. https://doi.org/10.1016/j
.cortex.2019.05.013, 考研: 31302507
Riccardi, N。, Yourganov, G。, Rorden, C。, Fridriksson, J。, & 德赛, 右.
(2020). Degradation of praxis brain networks and impaired com-
prehension of manipulable nouns in stroke. 认知杂志
神经科学, 32(3), 467–483. https://doi.org/10.1162/jocn_a
_01495, 考研: 31682566
理查森, F. M。, Ramsden, S。, 埃利斯, C。, Burnett, S。, Megnin, 奥。,
Catmur, C。, Schofield, 时间. M。, Leff, A. P。, & 价格, C. J. (2011).
Auditory short-term memory capacity correlates with gray mat-
ter density in the left posterior STS in cognitively normal and
dyslexic adults. 认知神经科学杂志, 23(12),
3746–3756. https://doi.org/10.1162/jocn_a_00060, 考研:
21568634
罗布森, H。, 智者, K., & Lambon Ralph, 中号. A. (2012). Wernicke’s
aphasia reflects a combination of acoustic-phonological and
semantic control deficits: A case-series comparison of Wer-
nicke’s aphasia, semantic dementia and semantic aphasia.
神经心理学, 50(2), 266–275. https://doi.org/10.1016/j
.neuropsychologia.2011.11.021, 考研: 22178742
Rogalsky, C。, & 希科克, G. (2009). Selective attention to semantic
and syntactic features modulates sentence processing networks
in anterior temporal cortex. 大脑皮层, 19(4), 786–796.
https://doi.org/10.1093/cercor/bhn126, 考研: 18669589
Rogalsky, C。, & 希科克, G. (2011). The role of Broca’s area in sen-
tence comprehension. 认知神经科学杂志, 23(7),
1664–1680. https://doi.org/10.1162/jocn.2010.21530, 考研:
20617890
Rogalsky, C。, LaCroix, A. N。, 陈, K. H。, 安德森, S. W., Damasio,
H。, 爱, T。, & 希科克, G. (2018). The neurobiology of agram-
matic sentence comprehension: A lesion study. Journal of Cog-
nitive Neuroscience, 30(2), 234–255. https://doi.org/10.1162
/jocn_a_01200, 考研: 29064339
Rogalsky, C。, 匹配, W., & 希科克, G. (2008). Broca’s area, sen-
tence comprehension, and working memory: An fMRI Study.
Frontiers in Human Neuroscience, 2, 文章 14. https://doi.org
/10.3389/神经的.09.014.2008, 考研: 18958214
Rorden, C。, Bonilha, L。, Fridriksson, J。, Bender, B., & Karnath, H. 氧.
(2012). Age-specific CT and MRI templates for spatial normaliza-
的. 神经影像, 61(4), 957–965. https://doi.org/10.1016/j
.neuroimage.2012.03.020, 考研: 22440645
拉夫, 我。, Blumstein, S. E., 迈尔斯, 乙. B., & 和记黄埔, 乙. (2008).
R e c r u i t m e n t o f a n t e r i o r a n d p o s t e r i o r s t r u c t u r e s i n
lexical-semantic processing: An fMRI study comparing implicit
and explicit tasks. 大脑和语言, 105(1), 41–49. https://
doi.org/10.1016/j.bandl.2008.01.003, 考研: 18279947
Salis, C. (2012). Short-term memory treatment: Patterns of learning
and generalisation to sentence comprehension in a person with
aphasia. Neuropsychological Rehabilitation, 22(3), 428–448.
https://doi.org/10.1080/09602011.2012.656460, 考研:
22300398
Schell, M。, Zaccarella, E., & 弗里德里克, A. D. (2017). Differential
cortical contribution of syntax and semantics: An fMRI study
on two-word phrasal processing. 皮质, 96, 105–120. https://
doi.org/10.1016/j.cortex.2017.09.002, 考研: 29024818
Sliwinska, 中号. W., Khadilkar, M。, Campbell-Ratcliffe, J。, Quevenco,
F。, & 德夫林, J. 时间. (2012). Early and sustained supramarginal gyrus
contributions to phonological processing. Frontiers in Psychol-
奥吉, 3, 文章 161. https://doi.org/10.3389/fpsyg.2012.00161,
考研: 22654779
Snijders, 时间. M。, 狐狸, T。, 活动, G。, Van Berkum, J. J。, Petersson,
K. M。, & 哈科特, 磷. (2009). Retrieval and unification of syntactic
structure in sentence comprehension: An fMRI study using
word-category ambiguity. 大脑皮层, 19(7), 1493–1503.
https://doi.org/10.1093/cercor/bhn187, 考研: 19001084
Thothathiri, M。, Kimberg, D. Y。, & 施瓦茨, 中号. F. (2012). 新的-
ral basis of reversible sentence comprehension: 证据来自
voxel-based lesion symptom mapping in aphasia. Journal of Cog-
nitive Neuroscience, 24(1), 212–222. https://doi.org/10.1162
/jocn_a_00118, 考研: 21861679
土耳其人, A. U。, & drungers, 氮. F. (2011). The neural architecture of
the language comprehension network: Converging evidence
from lesion and connectivity analyses. Frontiers in Systems Neu-
roscience, 5, 文章 1. https://doi.org/10.3389/fnsys.2011.00001,
考研: 21347218
Veraart, J。, Fieremans, E., & Novikov, D. S. (2016). 弥散磁共振成像
noise mapping using random matrix theory. Magnetic Resonance
在医学上, 76(5), 1582–1593. https://doi.org/10.1002/mrm
.26059, 考研: 26599599
理智, S。, 江, H。, Nagae-Poetscher, L. M。, 范·齐尔, 磷. C. M。, &
森, S. (2004). Fiber tract-based atlas of human white matter
语言的神经生物学
343
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Canonical sentence processing and the inferior frontal cortex
anatomy. Radiology, 230(1), 77–87. https://doi.org/10.1148
/radiol.2301021640, 考研: 14645885
Walenski, M。, 欧罗巴, E., Caplan, D ., & 汤普森, C. K. (2019).
Neural networks for sentence comprehension and production:
An ALE-based meta-analysis of neuroimaging studies. 人类
大脑映射, 40(8), 2275–2304. https://doi.org/10.1002/hbm
.24523, 考研: 30689268
惠特尼, C。, 柯克, M。, O’Sullivan, J。, Lambon Ralph, 中号. A。, & Jefferies,
乙. (2011). The neural organization of semantic control: TMS evi-
dence for a distributed network in left inferior frontal and posterior
中间回. 大脑皮层, 21(5), 1066–1075. https://
doi.org/10.1093/cercor/bhq180, 考研: 20851853
威尔逊, S. M。, Demarco, A. T。, 亨利, 中号. L。, Gesierich, B., Babiak,
M。, 磨坊主, 乙. L。, & Gorno-Tempini, 中号. L. (2016). Variable disrup-
tion of a syntactic processing network in primary progressive
aphasia. 脑, 139(11), 2994–3006. https://doi.org/10.1093
/brain/aww218, 考研: 27554388
温克勒, A. M。, Ridgway, G. R。, 韦伯斯特, 中号. A。, 史密斯, S. M。, &
尼科尔斯, 时间. 乙. (2014). Permutation inference for the general linear
模型. 神经影像, 92, 381–397. https://doi.org/10.1016/j
.neuroimage.2014.01.060, 考研: 24530839
Zaccarella, E., & 弗里德里克, A. D. (2015). Merge in the human
脑: A sub-region based functional investigation in the left pars
opercularis. 心理学前沿, 6, 文章 1818. https://土井
.org/10.3389/fpsyg.2015.01818, 考研: 26640453
Zaccarella, E., Schell, M。, & 弗里德里克, A. D. (2017). Reviewing the
functional basis of the syntactic merge mechanism for language: A
coordinate-based activation likelihood estimation meta-analysis.
神经科学 & Biobehavioral Reviews, 80, 646–656. https://土井
.org/10.1016/j.neubiorev.2017.06.011, 考研: 28743620
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
2
3
1
8
2
0
0
8
4
8
2
n
哦
_
A
_
0
0
0
6
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
语言的神经生物学
344