RESEARCH ARTICLE
Neural Oscillations Reflect Meaning Identification
for Novel Words in Context
开放访问
杂志
1Joint Doctoral Program in Language and Communicative Disorders, San Diego State University and UC San Diego,
圣地亚哥, CA, 美国
2School of Speech, 语言, and Hearing Sciences, San Diego State University, 圣地亚哥, CA, 美国
Jacob Pohaku Momsen1
and Alyson D. Abel2
关键词: ERSP, neural oscillations, semantic memory, sentence processing, theta power, beta
力量
抽象的
During language processing, people make rapid use of contextual information to promote
comprehension of upcoming words. When new words are learned implicitly, 信息
contained in the surrounding context can provide constraints on their possible meaning. 在里面
current study, EEG was recorded as participants listened to a series of three sentences, each
containing an identical target pseudoword, with the aim of using contextual information in the
surrounding language to identify a meaning representation for the novel word. In half of the
试验, sentences were semantically coherent so that participants could develop a single
representation for the novel word that fit all contexts. Other trials contained unrelated sentence
contexts so that meaning associations were not possible. We observed greater theta band
enhancement over the left hemisphere across central and posterior electrodes in response to
pseudowords processed across semantically related compared to unrelated contexts.
此外, relative alpha and beta band suppression was increased prior to pseudoword
onset in trials where contextual information more readily promoted pseudoword meaning
协会. Under the hypothesis that theta enhancement indexes processing demands
during lexical access, the current study provides evidence for selective online memory
retrieval for novel words learned implicitly in a spoken context.
介绍
The ability to learn new words and develop an extensive lexicon is a well-studied phenome-
non of cognition. Broadly speaking, word learning can be achieved in a number of different
ways—via explicit instruction or alternatively through implicit means. When new words are
encountered on the fly, meaning has to be inferred from the surrounding linguistic context
(Fukkink, 2005; Shtyrov, 2011). 例如, if exposed to the unfamiliar word shap in the
following sentence, The boy held the shap tightly as he fell asleep, interpretations would be
constrained to potential meanings that are plausibly related to other familiar words in the sen-
张力 (例如, pillow, blanket). While there is evidence that people can become sensitive to the
meaning of a new word after being exposed to it in context only once, long-lasting memory for
new words becomes robust after multiple exposures (Horst, 2013).
引文: Momsen, J. P。, & Abel, A. D.
(2022). Neural oscillations reflect
meaning identification for novel words
in context. Neurobiology of Language,
3(1), 132–148. https://doi.org/10.1162
/nol_a_00052
DOI:
https://doi.org/10.1162/nol_a_00052
已收到: 15 二月 2021
公认: 27 七月 2021
利益争夺: 作者
have declared that no competing
interests exist.
通讯作者:
Jacob Pohaku Momsen
jmomsen@ucsd.edu
处理编辑器:
Alec Marantz
版权: © 2021
麻省理工学院
在知识共享下发布
归因 4.0 国际的
(抄送 4.0) 执照
麻省理工学院出版社
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
哦
_
A
_
0
0
0
5
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
Meaning identification for novel words in context
Learning words from context is often studied using written language (Nagy et al., 1985,
1987), 但, 重要的是, many new words are learned by hearing them used in everyday
speech (凯里 & Bartlett, 1978; Jenkins et al., 1984; McLeod & McDade, 2011; 藏红花
等人。, 1997). Considering the level of exposure people get to new words over their life span
simply by listening to others talk, it is vital to establish an understanding of the brain mecha-
nisms that underpin contextual learning during online speech processing. Extant neuroscientific
research on implicit word learning has mostly focused on written text; the present study instead
focuses on spoken language processing.
Neural Oscillations and Speech Processing
An increasingly popular approach to examining EEG data in studies of language processing is
using event related spectral perturbations (ERSPs)—a transformation of EEG data that allows for
the inspection of both phase and non-phase locked neural activity, which is unobservable
using more traditional event-related potential (ERP) 分析 (Pfurtscheller & 席尔瓦, 1999).
ERSPs are formed from a decomposition of the continuous EEG signal to examine how neural
activity across various frequency bands changes as a function of different experimental vari-
埃布尔斯. There have been increasing efforts to explain how the neural oscillatory activity works
as a vehicle for language-related cognitive operations (see Prystauka & Lewis, 2019).
Both in and out of the context of language processing, theta band activity (~4–7 Hz) 有
been most frequently associated with memory retrieval operations (Bastiaansen & Hagoort,
2006; Bastiaansen et al., 2002, 2010; Hald et al., 2006; 施耐德 & Maguire, 2018). 在一个
2005 学习, researchers found contrasts between the oscillatory response to open-class versus
closed-class words during a passive reading task (Bastiaansen et al., 2005). The differences
were most stark in theta band activity: Open-class words elicited increases in theta power over
left temporal regions of the scalp while comparable modulation was absent for closed-class
字. This led to the conclusion that the observed changes were primarily reflective of re-
trieval processes from long-term memory elicited by semantically rich language. Other studies
show theta power enhancement in response to stimuli that are semantically incongruent with
preceding language (Bastiaansen & Hagoort, 2006; Bastiaansen et al., 2005; Hald et al., 2006).
As for sentence-level dynamics, Lam and colleagues found relative decreases in theta power
for words that appeared late in a sentence compared to those that occurred earlier (Lam et al.,
2016). Taken together under a functional interpretation, these results suggest that theta power
may index effort related to semantic memory retrieval demands, where enhancement during
word processing increases if a relevant context does not facilitate lexical access.
此外, alpha (8–12 Hz) and beta (13–30 Hz) frequencies have been discussed in light
of a number of processes likely to play a relevant role during language processing in context,
such as memory retrieval, efforts related to controlled attention, predictive processing, 和
working memory operations (Gao et al., 2017; Hanslmayr et al., 2012; Klimesch, 2012;
Klimesch & Schack, 2003; Piai et al., 2014; 韦斯 & Mueller, 2012). 例如, alpha sup-
pression appears to support a range of functions—displaying sensitivity to the complexity of
syntactic configurations (Vassileiou et al., 2018), the predictability of upcoming language
(Rommers et al., 2017; 王等人。, 2018), and speech intelligibility (Obleser & 魏兹,
2012)—and it is generally associated with conditions where cognitive resources are devoted
more heavily to the current task (詹森 & Mazaheri, 2010; Klimesch et al., 2007). 学习
observing beta oscillations posit a role in actively reflecting the maintenance of context-
sensitive meaning representations via top-down predictive signaling (刘易斯等人。, 2016).
This predictive coding account hypothesizes beta band suppression in response to syntactic
Neurobiology of Language
133
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
哦
_
A
_
0
0
0
5
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
Meaning identification for novel words in context
Neural oscillations:
In the current study, this refers
to large-scale repetitive and
synchronized neural activity
that can be measured with
scalp-recorded EEG.
or semantic perturbations during sentence processing (Bastiaansen et al., 2010; Lewis &
Bastiaansen, 2015; 王等人。, 2012). 例如, Kielar and colleagues (2014) found that
semantic violations in a sentence such as “A new computer will paint for many years” (Kielar
等人。, 2014, p. 3) elicit a decrease in beta and alpha power following the onset of the seman-
tically incongruent word. 一起, alpha and beta band suppression may arise in response to
increased cognitive demands related to processing words across semantically unrelated
contexts—that is to say, contexts that only weakly support contextual learning or inference.
Neural Oscillations and Word Learning
In addition to assessing learning performance using behavioral measures (IE。, whether a par-
ticipant can correctly report a new word’s meaning), research has used real-time processing
measures to index the brain response to words after a learning opportunity. Previous studies
using electrophysiological measures have demonstrated that adults show neural evidence of
rapid altered sensitivity to unfamiliar words after being exposed to them in context only once
(Batterink & Neville, 2011; Mestres-Missé et al., 2007; Perfetti et al., 2005). While only a few
studies have used ERSP measures to compare the processing of unknown relative to known
字, theta power appears to exhibit preferential sensitivity to known words especially over
the left hemisphere (Krause et al., 2006; Marinkovic et al., 2012), not unlike the greater en-
hancement seen for semantically rich words over and above closed-class words (Bastiaansen
等人。, 2005). These studies offer a valuable perspective on the neural oscillations that relate to
unfamiliar language.
One notable study provides evidence that theta band activity can act as a useful proxy for
some components of the word learning process. In a multisession word learning study, Bakker
and colleagues (2015) observed significantly reduced theta band activity when comparing
the initial neural response to unfamiliar pseudowords with responses to real words. After a
learning session that exposed participants to explicit word definitions, this relative power
difference was still present, albeit reduced. When participants were tested on the learned
words the following day after an opportunity for overnight memory consolidation, this low
frequency response to pseudowords was enhanced such that there was comparable ERSP
activity between these newly learned pseudowords and previously known real words. 这是
currently unknown how oscillatory mechanisms actively support other types of familiarization
and learning conditions used to eventually learn words—especially as this occurs in the
context of spoken language processing. The current study examines whether effectively using
context to build up meaning associations with novel spoken words can instigate a similar tran-
sition in the oscillatory response as unfamiliar words eventually trigger online semantic retrieval.
The Current Study
The ability to attach meaning to unfamiliar words while considering the wider linguistic con-
text is fundamental to successful implicit word learning, and thus warrants a better understand-
ing of the brain mechanisms involved. 因此, the purpose of the current study was to investigate
the electrophysiological markers associated with changes in response to novel spoken words
in context. This is to say, our aim is not to investigate long-term implicit word learning per se,
but instead focuses on one central component of this process: estimating a plausible meaning
for a new word based solely on the context it is discovered in. 进一步来说, we tested the
hypothesis that exposure to a novel word embedded across semantically coherent sentences
(意义 (+)) will promote more word-like oscillatory responses than novel words embedded
across semantically unrelated sentences (意义 (-)).
Neurobiology of Language
134
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
哦
_
A
_
0
0
0
5
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
Meaning identification for novel words in context
As a supplement to this primary motivation, we also examined unfolding changes in neural
activity prior to pseudoword onset to characterize how oscillatory markers of sentence pro-
cessing and contextual inference correspond to differences in the semantic relatedness across
multiple sentences. Participants were exposed to a series of sentence triplets, each containing
an identical target pseudoword. Participants were then instructed to identify the meaning of
the target pseudoword, requiring them to use adjacent contextual cues embedded across the
句子. In half of the trials, sentence contexts were related to each other so that a represen-
tation for the unknown word could fit with all the sentence contexts that the pseudoword had
appeared in. Sentence contexts were unrelated in the other half of the trials so that a single
meaning could not be associated with pseudowords.
ERSP effects in the theta band typically associated with real-word comprehension provided
the primary motivation for our hypothesis. We hypothesized that successfully associating mean-
ing to pseudowords would coincide with increased theta power enhancement to these words
between their first and final presentation, and that a similar increase would be diminished or not
occur to pseudowords lacking a meaning association. We also expected alpha and beta suppres-
sion to disproportionately occur as participants processed words in unrelated contexts, that is to
say when expectations about upcoming words are more frequently violated (Kielar et al., 2014).
方法
参加者
The current study included a cohort of 32 monolingual English-speaking, right-handed adult
参与者. All adults were administered a nonword repetition task (Dollaghan & 坎贝尔,
1998), which served as an index of phonological working memory ability (percentage of con-
sonants correct M = 94.0%; 标准差= 4.31%). Data from four participants were removed from the
final analysis due to excessive artifacts during recording, resulting in a final sample of 28 par-
ticipants (Mage = 20.5, SDage = 2.17; all female). Participants gave informed consent in accor-
dance with the San Diego State University Institutional Review Board. All participants gave
informed consent for their participation and were compensated with academic course credit.
Stimulus Materials
Our paradigm used a total of 300 spoken sentence stimuli in which the terminal word was
always a noun generated from a database of nouns commonly acquired by the age of 30 月
(MacArthur-Bates Communicative Developmental Inventory; Fenson et al., 2006). 句子
were all 6–9 words in length, and the terminal noun was preceded by either a determiner
(A, 这) or a possessive (我的, your, his/her).
For the word learning paradigm, trial stimuli were created by combining sentences into sets
of three (described in more detail below). The sentence-terminal noun in all three sentences
within a given trial was replaced by the same pseudoword, generated from a database (Storkel,
2013) of consonant-vowel-consonant sequences. None of the pseudowords had word-initial
sounds of /s/ or /sh/ to improve the time-locking accuracy to the pseudoword. 此外,
there were no phonetic constraints for vowels or word-final consonants.
All sentence stimuli were normed in an offline sentence completion task given to a cohort
的 248 adults to produce cloze probability ratings for each sentence-final word. The cloze
probability of a word is defined as the percentage of people who spontaneously choose to
finish a sentence with that word in an offline norming task. Cloze probability information
from the norming task was used to contrive two conditions. The Meaning (+) 状况
135
Phonological:
Refers to the system of speech sounds
that constitute words.
Cloze probability:
The probability that a particular word
will appear in a target sentence
position given its prior context.
Neurobiology of Language
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
哦
_
A
_
0
0
0
5
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
Meaning identification for novel words in context
incrementally facilitated learning for the target pseudoword within a trial by increasing the
amount of contextual support for each sentence. This was achieved by using sentences whose
final noun had low cloze probabilities in the offline norming task as the first out of three sen-
tences in a trial, and high cloze probabilities as the trial-final sentences (Mean cloze proba-
bilities for target words in first Meaning (+) sentences = 4.0%, 标准差= 6.0%; in second Meaning
(+) sentences = 40.6%, 标准差= 10.0%; in third Meaning (+) sentences = 88.1%, 标准差= 11.2%).
重要的, the Meaning (+) condition used sentences from the norming task with the same
final real word within each trial, making the sentences complimentary in terms of their support
for one target meaning. In the Meaning (-) 状况, the pseudowords also replaced the
sentence-final word in each sentence stimuli; 然而, the real word that the pseudoword re-
placed was a different real word for each sentence in the triplet (见表 1) (Mean cloze prob-
abilities for target words in first Meaning (-) sentences = 17.5%, 标准差= 17.7%; in second Meaning
(-) sentences = 20.3%, 标准差= 20.2%; in third Meaning (-) sentences = 17.1%, 标准差= 15.6%).
Each participant was administered 100 试验 (sentence triplets), 50 意义 (+) trials and
50 意义 (-) 试验. Confounds introduced from by-item effects were controlled for by using
each pseudoword stimuli in both the Meaning (+) and Meaning (-) conditions across partic-
爱普茨. All sentences were recorded in a sound-attenuated chamber by a female native English
speaker, and were produced at a natural pace to promote naturalistic listening conditions
(Mean sentence length prior to pseudoword Meaning (+) = 1,788.5 多发性硬化症, 标准差= 342.0 多发性硬化症;
意义 (-) = 1,883.4 多发性硬化症, 标准差= 347.4 多发性硬化症; Mean pseudoword length Meaning (+) = 544 多发性硬化症,
标准差= 81 多发性硬化症; 意义 (-) = 534 多发性硬化症; 标准差= 78 多发性硬化症).
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
哦
_
A
_
0
0
0
5
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
程序
The experimental paradigm was adopted from a 2007 study investigating implicit word learn-
ing in adults (Mestres-Missé, 等人。, 2007). Participants listened to sets of three naturally-paced
句子. Participants were instructed to attend to the three sentences and to attempt to iden-
tify the meaning of the unknown pseudoword by using contextual information provided across
the sentence triplet. At the beginning of a trial, a fixation cross appeared on a monitor screen
Sentence order
1
2
3
桌子 1.
Example triplets from each condition
意义 (+) 状况
Sentence presented
Her parents bought her a pav
The sick child spent the day in his pav
Mom piled the pillows on the pav
意义 (-) 状况
Real word replaced
床
床
床
Sentence order
Sentence presented
Real word replaced
1
2
3
Don’t drop and break the pav
I spilled dinner all over my pav
You have to study hard to be a pav
cup
pants
doctor
136
Neurobiology of Language
Meaning identification for novel words in context
为了 600 多发性硬化症. After this fixation cross, a spoken sentence stimulus was presented through
speakers arranged approximately 1 meter from the participant. After the offset of the first sen-
张力, participants saw a screen with multiple fixation crosses indicating that they could push
a button to initiate the presentation of the next sentence. After a button push to progress the
审判, another fixation cross appeared on the screen for 600 ms prior to the onset of the next
sentence to ensure that each sentence onset had a baseline period without stimulus presenta-
的. After the third sentence in a trial was presented, participants gave a button push to initiate
the response phase of the trial, in which they were asked by an experimenter if they believed the
target pseudoword was meaningful (IE。, if a single word was semantically congruent with the
entire sentence triplet). 重要的, these questions were not verbalized until after participants
ended the trial with a self-paced button push. If the participant believed the target pseudoword
was meaningful, they gave an oral response to the experimenter in an attempt to identify the
specific word that would fit best to replace the pseudoword in that particular trial.
Trials in the Meaning (+) condition were only counted as correct if the participant success-
fully identified the target word that the pseudoword replaced or gave an answer that was rea-
sonably similar (例如, a response “truck” for the target “car”). Trials in the Meaning (-)
condition were counted as correct if the participant reported that the target pseudoword could
not represent a noun that was semantically congruent across all three sentences within the trial
(例如, “the word is not meaningful”). Only correct trials were used in the EEG analysis. Prior to
testing, a training session was administered, such that each participant was exposed to an
example trial in the Meaning (+) and Meaning (-) 状况. Feedback was given for perfor-
mance during the training session but not for the actual experiment. 见图 1 for a visual
representation of a single trial.
EEG Acquisition and Processing
E E G r e c o r d i n g w a s p e r f o r m e d w i t h a 6 4 e l e c t r o d e N e u r o s c a n Q u i c k c a p
(compumedicsneuroscan.com/) arranged according to the 10–20 International Standard con-
figuration system. The EEG signal was referenced online at a central electrode site near the ver-
tex of the head and rereferenced offline to the average of two lateral electrode sites approximate
to the mastoids (CB1, CB2). During recording, EEG signals were low-pass filtered (200 赫兹) 和
sampled at 1 千赫. Data were later resampled offline at 512 赫兹. A 40 Hz low-pass and 1 赫兹
high-pass filter were applied to the data and then independent component analysis was per-
formed on the continuous EEG recording for each subject in order to identify sources of
数字 1. Example of one Meaning (+) 审判.
Neurobiology of Language
137
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
哦
_
A
_
0
0
0
5
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
Meaning identification for novel words in context
nonbrain-related electrical activity (Infomax ICA; 钟 & Sejnowski, 1995). Components iden-
tified as being primarily related to blink and movement artifacts were subsequently removed
after manual inspection (Mean components rejected = 1.77; 标准差= 0.91). Malfunctioning elec-
trodes were corrected by spherical interpolation based on adjacent electrode data. Any remain-
ing artifacts related to eye movements and flatlining were flagged with an automatic artifact
detection algorithm (EEGLAB; Delorme & Makeig, 2004) and removed from analysis.
此外, epochs corresponding to trials with an incorrect response from the behavioral task
were removed from the analysis. (中号 (标清) number of trials per subject entering into the analyses:
意义 (+) 1st sentence = 36.4 (4.1); 意义 (+) 2nd sentence = 37.6 (4.3); 意义 (+) 3rd
sentence = 37.8 (4.2); 意义 (-) 1st sentence = 39.1 (3.9); 意义 (-) 2nd sentence = 39.9
(3.8); 意义 (-) 3rd sentence = 40.8 (3.9).
Time–Frequency Analysis
Statistical analyses of time–frequency data used nonparametric cluster-based permutation tests
(马里斯 & Oostenveld, 2007). This approach is particularly appropriate for analyses in which
there is less certainty regarding the nature and time course of the effects of interest. For con-
ditional comparisons of interest, a t value was derived from each data point across two subject-
channel-frequency-time matrices. A three-dimensional cluster statistic was derived from
summed t values adjacent in location, 时间, and frequency (alpha cluster threshold =
0.025). Cluster-level statistics were compared with a null distribution of cluster statistics obtained
via a randomized permutation procedure (N= 1,000). Monte Carlo p values were acquired by
comparing the observed cluster statistic to this distribution. Electrode neighbors were defined via
triangulation method (6.3 average neighbors per electrode). Time–frequency representations
were computed via short-time fast Fourier transformation using Fieldtrip code implemented in
MATLAB (版本. R2019b) (mathworks.com; Oostenveld et al., 2011). A 500 ms sliding Hann
window was applied to epochs using 32 ms time steps and across frequencies from 2–30 Hz
at steps of 2 赫兹.
Data segments used in the cluster analyses were generated using epochs that were time-
locked to the onset of the critical pseudoword in each sentence. These epochs included data
spanning from 1,000 ms prior to and 750 ms after pseudoword onset. To help delineate if
effects were specific to pseudoword processing rather than their preceding context, we per-
formed similar but separate analyses of target data recorded prior to (−1,000 to 0 多发性硬化症) and after
(0 ms to 750 多发性硬化症) the pseudoword onset in each sentence. To observe differences in pseudo-
word processing relative to task-related changes in brain activity that accumulate within trials,
pre-sentence baselines were used to normalize target activity related to pseudowords.
Separate epochs time-locked to sentence onset were extracted from continuous data to gen-
erate time–frequency epochs for baseline corrections. Data from 250 ms to 50 ms prior to
sentence onset was averaged across Meaning (+) 和 (-) conditions for each sentence presen-
站, resulting in three time–frequency baseline arrays used for point-by-point decibel trans-
形成 (10*log10(activity/baseline)) of pseudoword related activity to create ERSPs for all
reported statistical analyses and visual representations of the data.
We investigated the neural correlates related to novel word processing in context by ana-
lyzing the neural response to pseudowords embedded across either semantically coherent or
incoherent sentences. To test whether being associated with a meaning representation changes
the trajectory of the neural response to pseudowords, we tested an interaction between
Meaning and sentence presentation, 那是, a comparison between the Meaning (+) 和 (-)
conditions for the difference between the power at pseudoword onset in the first and final
Neurobiology of Language
138
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
哦
_
A
_
0
0
0
5
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
9
S
e
p
e
米
乙
e
r
2
0
2
3
Meaning identification for novel words in context
句子. 此外, we examined direct comparisons of pre-pseudoword activity at each
sentence presentation (句子 1, 2, 和 3) to identify how our Meaning manipulation mod-
ulated online sentence processing dynamics as participants attempted to use contexts to inform
novel word processing.
结果
行为表现
Trial accuracy across both the Meaning (+) and Meaning (-) conditions were relatively high.
Participants correctly identified a meaning for the target pseudoword in the Meaning (+) 骗局-
dition on 83.8% (标准差= 7.9%) 的考验. Participants also correctly reported that the pseudoword
did not represent a plausible word across sentence contexts in the Meaning (-) condition on
91.0% (标准差= 5.6%) 的考验. A paired t test suggested that task performance was slightly
enhanced in the Meaning (-) 状况 (t = 3.9; p < 0.01). Overall high performance sug-
gests that the memory and learning demands posed by the current task were relatively man-
ageable for our cohort of healthy adults.
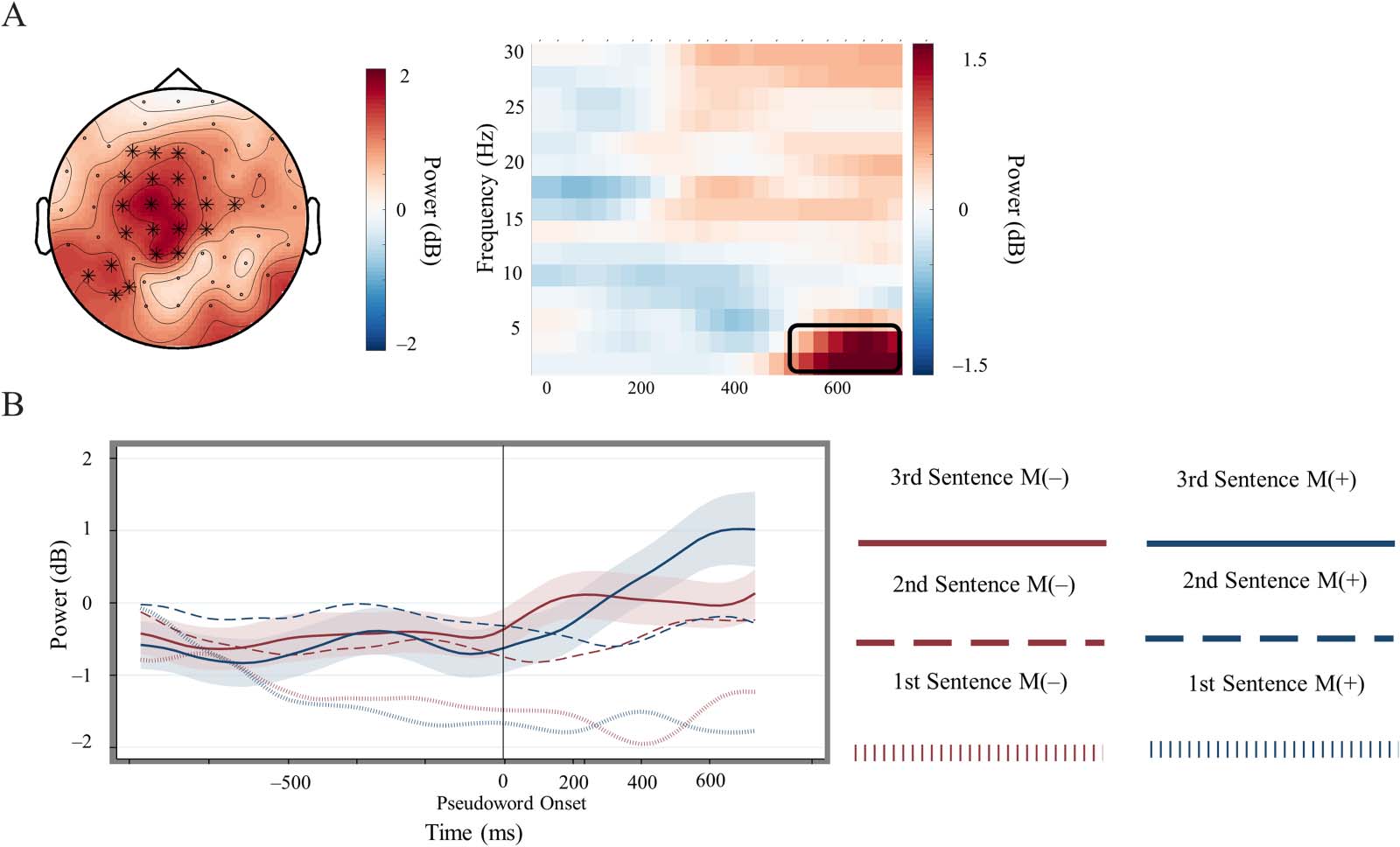
Cluster-Based Permutation Results
Cluster-based permutation tests comparing the difference in pseudoword processing from ini-
tial to final presentation across Meaning (+) and (−) trials indicated rejection of the null hypoth-
esis of exchangeability across data distributions (p = 0.049). This nonparametric analysis
generated a cluster estimate in the positive direction that included low frequency estimates
(2–4 Hz) and lasted from approximately 500 ms post-pseudoword onset to the end of the
epoch. This result suggests that the pseudowords that eventually triggered semantic retrieval
by being positioned in meaningful contexts elicited greater increases of low-frequency power
in the theta band compared to pseudowords embedded in contexts that precluded a similar
semantic representation from being retrieved. Similar cluster-based tests directly comparing
pre-pseudoword activity across Meaning (+) and (−) trials did not identify significant differences
for first or second sentences. A test comparing Meaning (+) and (−) for the third sentences indi-
cated a significant difference, revealing a cluster in the negative direction ( p = 0.002). This test
was associated with a cluster estimate predominantly characterized by frequency values in the
alpha and low beta range (8–20 Hz), which began near the initial time sample and lasted until
approximately 300 ms prior to pseudoword onset.
Theta Power Enhancement Across Pseudoword Presentation
Compared to the difference between pseudowords upon their initial presentation, final pseu-
dowords embedded within semantically cohesive sentence sets and consequently associated
with meaning elicited greater theta enhancement compared to pseudowords that appeared
across a series of mutually unsupportive sentences (Figure 2). Our nonparametric analysis sug-
gested that this effect manifested relatively late, from approximately 500 ms after pseudoword
onset until the end of the epoch (see Sassenhagen & Draschkow, 2019, for discussion about
the interpretive limitations of cluster-based tests). The twenty-one electrodes comprising this
cluster estimate were predominately over central electrode locations, but also included elec-
trodes over left posterior scalp regions (Figure 3). Paired-sample t tests using subject-averaged
data from third sentences indicated that theta power in Meaning (+) trials was greater than
zero, indicating a relative power enhancement compared to the pre-sentence baseline (t =
2.34, SD = 2.14, p < 0.05). Alternatively, the theta response to third pseudowords in
Meaning (−) trials was not significantly enhanced compared to the baseline (t = 0.05; p = 0.9).
Neurobiology of Language
139
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
o
_
a
_
0
0
0
5
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Meaning identification for novel words in context
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
o
_
a
_
0
0
0
5
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
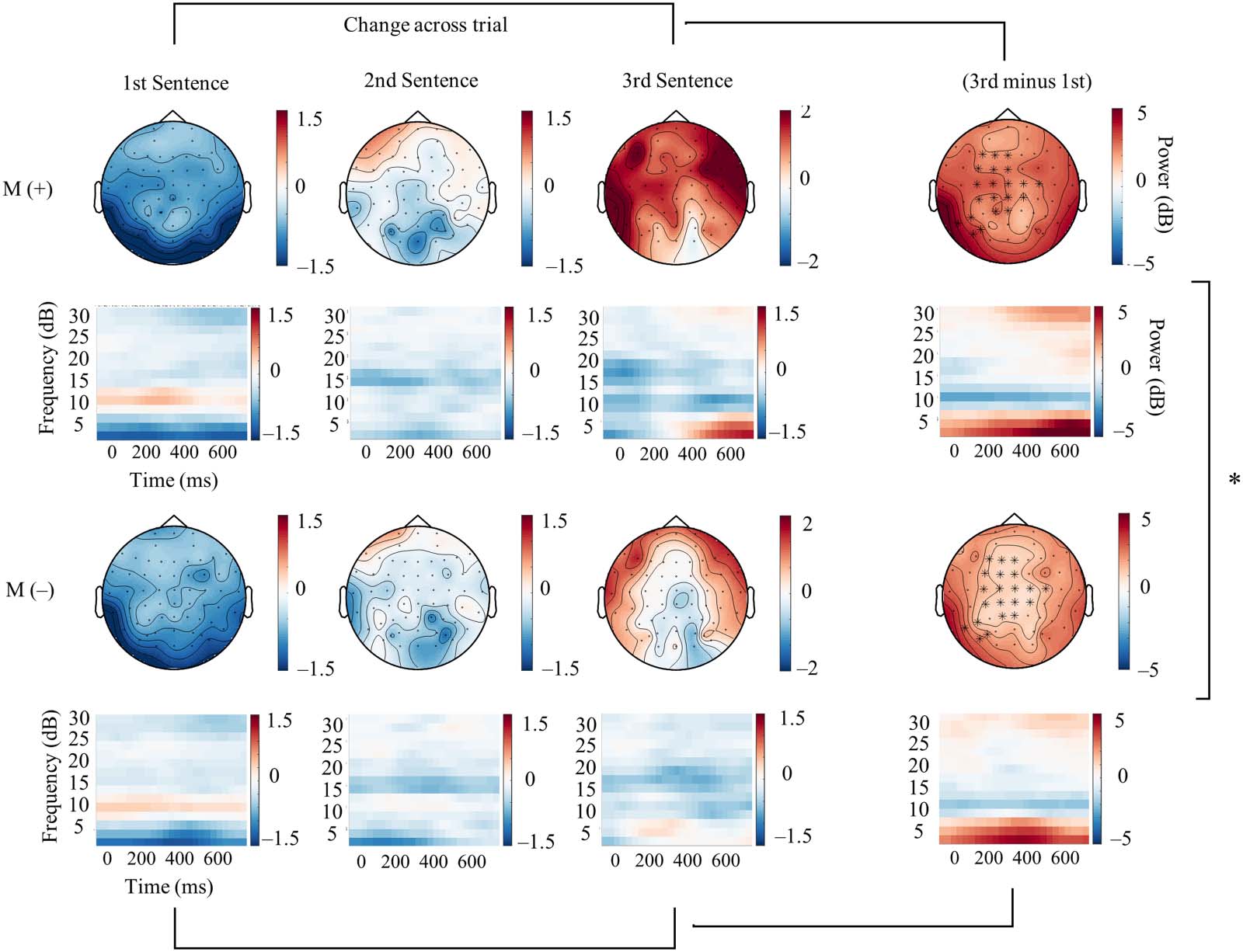
Figure 2. A series of topographical distributions and time–frequency plots representing the change in theta band power occurring after pseudo-
word onset relative to the pre-sentence baseline. An interaction between Sentence presentation (transition between 1st and 3rd sentences)
and the Meaning manipulation (Meaning (+) vs. (–) trials) revealed greater changes in theta enhancement for pseudowords associated with
meaning (far right). The shown topographical distributions of theta enhancement correspond to the spatiotemporal features of the cluster
estimate revealed by this interaction (2–4 Hz; approximately 500–750 ms post-pseudoword onset). The time–frequency plots display ERSP
activity averaged across all channels identified in the interaction, which are indicated by the asterisks on the far-right scalp plots.
Relative Alpha and Beta Power Suppression Prior to Final Pseudowords
Our analyses indicated relative alpha and beta band (8–20 Hz) suppression preceding final
pseudoword onset in Meaning (+) trials relative to Meaning (−). The separate calculation of
group means for alpha (8–12 Hz) and beta (14–20 Hz) band activity suggested that differences
prior to pseudoword onset were driven predominately by beta band suppression in the
Meaning (+) condition (t = −2.79, SD = 0.56, p < 0.01; Figure 4B). Electrodes in the significant
cluster indicated a widely distributed effect unlike the relative left-lateralization seen for the
theta band enhancement.
DISCUSSION
The current study set out to explore oscillatory neural dynamics that support implicit word
learning from speech. We tested the hypothesis that adults can develop an online sensitivity
to the meaning of unfamiliar words embedded across meaningful spoken sentence contexts by
analyzing oscillatory signatures of lexical processing. We hypothesized that greater theta power
enhancement would occur in association with pseudowords after being embedded across
semantically coherent as opposed to unrelated sentences. Our data revealed that pseudowords
Neurobiology of Language
140
Meaning identification for novel words in context
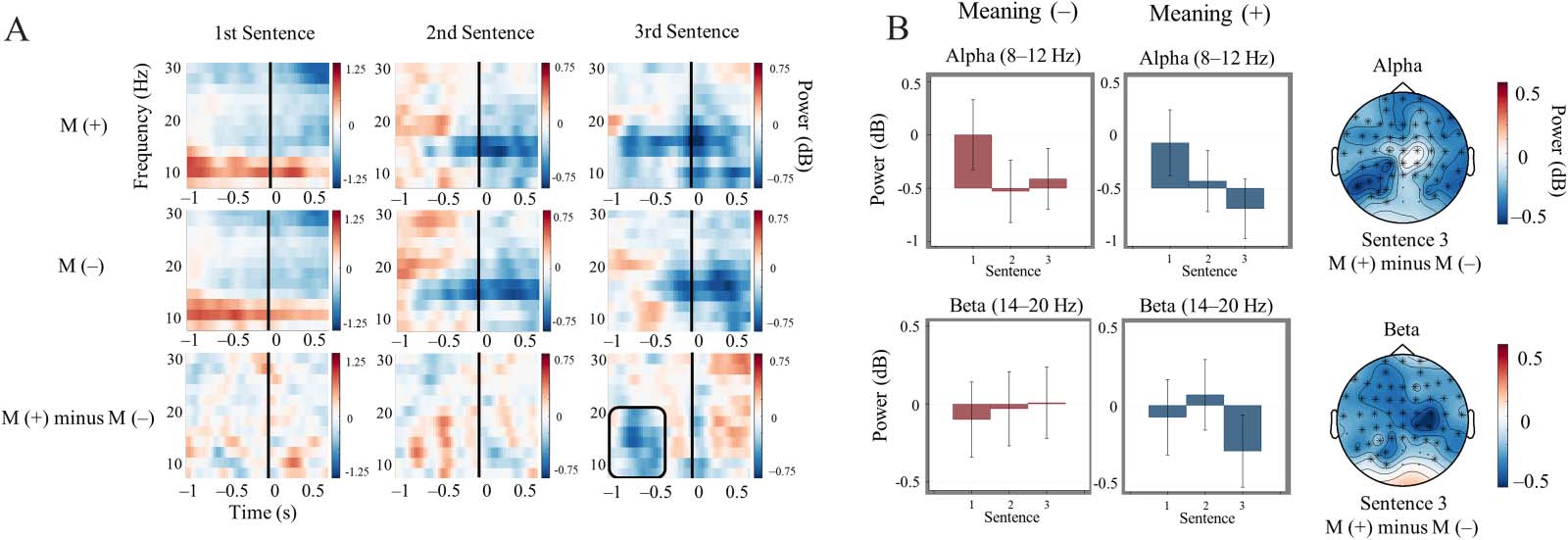
(A) Topographical distribution and time–frequency plot depicting the difference in theta enhancement observed across Meaning (+)
Figure 3.
relative to pre-sentence baseline relative to Meaning (−) trials. The scalp map shows averaged theta power over from approximately 500 ms
post-pseudoword onset to the end of the epoch. The spectrogram includes ERSP data averaged across all channels indicated by asterisks in the
scalp plot. (B) Time course of theta power averaged within spatiotemporal cluster boundaries for each sentence across Meaning conditions.
Shading for third sentence data indicates ±1 standard error of the mean and illustrates that theta power related to pseudowords completing
Meaning (+) trials was uniquely enhanced relative to pre-sentence baseline. 0 ms corresponds to pseudoword onset.
eliciting a meaning representation were associated with greater theta power enhancement
across trials relative to pseudowords presented in contexts where meaning was not similarly
activated. Additionally, we observed more alpha and beta power suppression prior to final
pseudowords in Meaning (+) trials relative to Meaning (−).
(A) Spectrograms including data averaged across significant cluster between third Meaning (+) and (−) sentences. Time and
Figure 4.
frequency range identified by the significant cluster are highlighted by the box in the third sentence difference plot (right side, bottom row).
(B) Topographical distribution of the difference in alpha and beta activity across Meaning conditions prior to pseudoword onset reflected in
the cluster analysis (−950 to −330 ms in the figure). Bar plots on left display alpha and beta activity corresponding to the observed cluster
estimate across sentences. Lines in these plots provide a time stamp of pseudoword onset.
Neurobiology of Language
141
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
o
_
a
_
0
0
0
5
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Meaning identification for novel words in context
Pseudowords Associated with Meaning Elicit Theta Band Enhancement
When novel or unfamiliar words are processed for the first time, low frequency activity in the
ERSP response lacks features common to real-word comprehension (Bakker et al., 2015;
Marinkovic et al., 2012). The present study found that processing novel spoken words in con-
text can engage oscillatory mechanisms related to real-word processing soon after their initial
presentation. While similar effects had been demonstrated in response to written text (e.g.,
Mestres-Missé et al., 2007), the current study provides a novel contribution by examining this
processing in the context of spoken language. This finding is significant, especially considering
the importance of word learning from speech. Importantly, we found that these effects depended
on the association between novel words and meaning representations—a direct product of the
context that pseudowords were presented in. A “meaningful” pseudoword embedded across
semantically coherent sentences eventually elicited greater theta band enhancement on average
than pseudowords embedded across incoherent contexts where meaning identification was not
achieved. This activity is unlikely the result of pseudoword repetition across sentences; indeed,
the effect displayed spatiotemporal features similar to those found in previous studies investigat-
ing ERSP correlates of language processing that suggest this low frequency enhancement is
linked to lexical retrieval processes (Bastiaansen et al., 2002, 2005; Hald et al., 2006; Kielar
et al., 2014).
Other neuroimaging work on incidental word learning in the written modality has provided
evidence of canonical language network activation (e.g., left middle temporal and inferior
frontal gyri) in conditions where novel words were meaningful in their provided context but
also in cases where context did not support meaning identification (Mestres-Missé et al.,
2008). A 2012 study used high density MEG and EEG to estimate source generators for theta
effects during a lexical decision task using real and pseudowords. Researchers observed more
theta band activation in left temporal and left inferior frontal regions for real words than pseu-
dowords between approximately 400–500 ms post-word-onset (Marinkovic et al., 2012).
More recent work established a similar generator for pseudoword related activity after a word
learning opportunity (Bakker-Marshall et al., 2018). In line with this work, the current effect
exhibited a lateralized bias and extended over left posterior temporal areas, reminiscent of the
left-biased topography common to EEG studies of lexical-semantic retrieval (Bastiaansen et al.,
2005; Marinkovic et al., 2012; Salisbury & Taylor, 2012).
Prior work also demonstrated activity across several other sites in the left hemisphere that
was unique to processing new words across meaningful relative to non-meaningful contexts,
including anterior parahippocampal gyrus, thalamus, and precuneus (Mestres-Missé et al.,
2008). There is growing support for a role of the medial temporal lobe during online language
processing—not only for learning but also when ambiguities about word meaning need to be
resolved (Duff & Brown-Schmidt, 2012; Pu et al., 2020). Links between theta rhythms and
functional coordination involving the hippocampus during memory retrieval are also consis-
tent with the observed effects in the Meaning (+) trials, considering that meaning identification
in the current paradigm requires a sensitivity to information delivered in prior contexts
(Herweg et al., 2016).
Previous evidence tracking the emergence of theta enhancement to unknown words after
meaning acquisition was demonstrated using explicit learning opportunities, and this effect of
learning was heightened after a period of overnight consolidation (Bakker et al., 2015; Bakker-
Marshall et al., 2018). The present study extends previous work by demonstrating that a similar
effect can be achieved via implicit learning mechanisms: As adults used sentence contexts to
successfully associate meaning with novel word forms, theta band activation was enhanced by
Neurobiology of Language
142
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
o
_
a
_
0
0
0
5
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Meaning identification for novel words in context
Sentence constraint:
Related to the highest cloze
probability that a sentence affords.
the third time that pseudowords were heard. Importantly, because we did not test participants
on their memory for pseudoword-meaning associations after the experiment, we cannot make
claims about the extent to which new words were learned—indeed, the task was a meaning
identification task not a word learning task per se. The importance of our findings lies in the
discovery that people can quickly develop an online sensitivity to the meaning of unfamiliar
words after hearing someone use them appropriately—words elicited an online neural signa-
ture suggestive of meaning acquisition from context only when words appeared in the
Meaning (+) condition.
An important caveat to our findings involves the role of contextual expectations during sen-
tence or discourse level processing. Our effect may be similar to that observed by Rommers
and colleagues (2017), who crossed effects of sentence constraint and word predictability and
found that target-word theta band enhancement was most strongly elicited by unexpected
words when participants were better able to generate expectations for them (i.e., when com-
pleting high constraint sentences). Because our goal was to experimentally contrive success
rates for implicit pseudoword-meaning associations, target real words completing Meaning
(+) trials did have higher cloze-probabilities on average than target words in Meaning (−) trials.
Thus, a current limitation of this study is that we cannot determine how much of this effect is a
product of these differences in context-driven expectations and their subsequent violation.
Indeed, this leaves the door open for an alternative interpretation: The effect could be taken
to reflect increased information processing demands as a result of competition between different
lexical representations partially activated by the end of Meaning (+) trials. If Meaning (+) trials
drove participants to activate one or multiple related candidates for the meaning identification
task, competition among different meaning representations or even between retrieved lexical
entries and the novel word itself could potentially explain increases in theta band activity. For
example, using a single-word auditory lexical decision task, Strauß et al. (2014) observed more
theta enhancement in response to phonologically ambiguous pseudowords (e.g., “banene”)
compared to real words (banana) or other pseudowords with novel phonological constructions.
Meaning associations unique to Meaning (+) trials may have had a similar effect by resulting in
co-activation between lexical representations in long-term memory and unfamiliar wordforms.
Thus, theta enhancement may index a conflict resolution process elicited by activated com-
peting lexical representations.
Lastly, theta activity after final pseudowords in Meaning (−) trials corresponding to the spa-
tiotemporal estimates of the cluster interaction was not significantly different from zero, sug-
gesting that a similar effect was not generated when pseudowords remained “meaningless.”
However, upon inspecting the transition from initial to final pseudowords in this condition, we
found that these stimuli do exhibit relatively increased theta power compared to pre-sentence
baseline across presentations; and a post hoc cluster-permutation test directly comparing first
and third pseudowords in these trials confirms this observation ( p < 0.001). We find it feasible
that low frequency activity tied to working memory operations may account for differences as
the trials progress. Maintaining relevant information from earlier sentences within the same
trial is presumably important for task performance, and memory demands during Meaning
(−) trials may even be heightened due to the semantic incongruence between sentences.
Theta activity reflecting more general working-memory encoding processes would align it with
studies that observed theta power enhancement during successful memory performance, espe-
cially when working memory content involved sequential information (Axmacher et al., 2006;
Kleberg et al., 2014; Klimesch et al., 2001; Nyhus & Curran, 2010; Roux & Uhlhaas, 2014;
Scholz et al., 2017). Because we used naturally paced speech and because our task required
that all trials involve a correct assessment of contextual fit, we believe it is possible that
Neurobiology of Language
143
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
o
_
a
_
0
0
0
5
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Meaning identification for novel words in context
Monotonic:
Describes a function that only
entirely increases or decreases
in value.
working-memory-related theta activity could contribute to ERSP activity in either Meaning
condition.
Alpha and Beta Suppression Within Meaningful Contexts
In addition to the observed effects in the theta band, beta and alpha suppression was relatively
heightened prior to final pseudoword onset in Meaning (+) trials compared to Meaning (−).
Across different sensorimotor domains, a number of proposals have posited beta power as
an index reflecting the preparation for processing anticipated events—inversely related to
the expected probability of voluntary action execution or of stimulus presentation
(Jenkinson & Brown, 2011; Van Ede et al., 2011). Applied to language processing, interplay
between top-down predictions, instantiated by beta activity, and bottom-up error signaling
might play a role in maintaining and updating a representation of discourse context important
for flexible online comprehension (Lewis et al., 2016; Meyer, 2018). Within this framework,
beta band suppression may have occurred selectively as people updated relevant information
about trial content or pseudoword meaning, whereas in Meaning (−) trials the semantic inco-
herence across sentences prevented stable contextual representations from being formed to
begin with—which is likely the reason we did not find evidence for similar suppression effects
in response to the pseudowords themselves
Additionally, recent work using MEG directly tested the relationship between power sup-
pression effects during sentence encoding and word predictability, and found that neither alpha
nor beta band activity held a monotonic relationship with the level of contextual constraint
provided by the sentence context (Terporten et al., 2019). This indicates these mechanisms
are not merely an index of upcoming word predictability and may instead relate more closely
to information updating during sentence processing. We believe a compatible description can
be framed around general long-term memory encoding and retrieval, in which both alpha and
beta activity act as an important mechanism for long-term memory access (Hanslmayr et al.,
2012; Klimesch, 2012; Klimesch et al., 2005). For example, alpha and beta suppression can
increase as a function of the number of items retrieved from memory (Khader & Rösler,
2011), and more relevantly, low beta suppression has been observed in response to real words
compared to unknown novel words (Bakker et al., 2015). Together, the fact that Meaning (+)
trials saw disproportionately enhanced beta and alpha suppression may reflect more robust
semantic activation or updating compared to Meaning (−).
Other potential interpretations could be mounted from evidence showing beta power as
closely related to reward processing (Lansink et al., 2016; Schwerdt et al., 2020). This may
be relevant to our current findings, considering that previous work has shown that subcortical
nuclei important for reward processing can be engaged during implicit language learning even
in the absence of explicit feedback about learning performance (Ripollés et al., 2014, 2016).
Lastly, it is also possible these effects were in part driven by differences in general attentional
engagement during these sentences (Jensen & Mazaheri, 2010; Klimesch et al., 2007; Shahin
et al., 2009).
Conclusions
We presented data from one of the first studies to focus on oscillatory brain dynamics related
to processes important for implicit word learning from speech. In summary, a cohort of 28
healthy adults were able to integrate contextual information across a series of three sentences
in order to estimate a viable association between a novel spoken word and a meaningful con-
cept. Sensitivity to context was reflected by a greater progressive enhancement of theta band
Neurobiology of Language
144
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
o
_
a
_
0
0
0
5
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Meaning identification for novel words in context
activity in trials with meaningful pseudowords compared to trials where pseudowords re-
mained meaningless. This work helps make progress toward understanding how people learn
from hearing words used appropriately by other speakers by showing that the brain adapts
quickly to speech to support the application of meaning associations to previously unknown
word forms.
ACKNOWLEDGMENTS
The authors would like to acknowledge Julie Schneider for her feedback on this manuscript,
members of the Language Learning Lab at San Diego State University for their assistance with
data collection and processing, and the research participants. Jacob Pohaku Momsen was sup-
ported by the Tribal Membership Initiative Fellowship through UC San Diego. Alyson D. Abel
was supported by the National Institute of Deafness and other Communication Disorders of
the National Institutes of Health under award number R21 DC018865, the San Diego State
University Grants Program, and the National Science Foundation under award number
BCS-1551770.
FUNDING INFORMATION
Alyson Abel, National Science Foundation (https://dx.doi.org/10.13039/100000001), Award
ID: Grant BCS-1551770. Alyson Abel, National Institute of Deafness and other
Communication Disorders of the National Institutes of Health (https://dx.doi.org/10.13039
/100000055), Award ID: R21 DC018865.
AUTHOR CONTRIBUTIONS
Jacob Pohaku Momsen: Methodology; Data curation; Formal analysis; Writing—review &
editing. Alyson D. Abel: Conceptualization; Data curation; Methodology; Writing—review
& editing; Funding acquisition; Supervision.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
o
_
a
_
0
0
0
5
2
p
d
.
/
l
REFERENCES
Axmacher, N., Mormann, F., Fernández, G., Elger, C. E., & Fell J.
(2006). Memory formation by neuronal synchronization. Brain
Research Reviews, 52(1), 170–182. https://doi.org/10.1016/j
.brainresrev.2006.01.007, PubMed: 16545463
Bakker, I., Takashima, A., van Hell, J. G., Janzen, G., & McQueen,
J. M. (2015). Changes in theta and beta oscillations as signatures
of novel word consolidation. Journal of Cognitive Neuroscience,
27(7), 1286–1297. https://doi.org/10.1162/jocn_a_00801,
PubMed: 25761007
Bakker-Marshall, I., Takashima, A., Schoffelen, J. M., van Hell, J. G.,
Janzen, G., & McQueen, J. M. (2018). Theta-band oscillations in
the middle temporal gyrus reflect novel word consolidation.
Journal of Cognitive Neuroscience, 30(5), 621–633. https://doi
.org/10.1162/jocn_a_01240, PubMed: 29393716
Bastiaansen, M.[C.], & Hagoort, P. (2006). Oscillatory neuronal dy-
namics during language comprehension. In C. Neuper & W.
Klimesch (Eds.), Event-related dynamics of brain oscillations
(pp. 179–196). Elsevier. https://doi.org/10.1016/S0079-6123(06)
59012-0
Bastiaansen, M. C., Linden, M. V., Keurs, M. T., Dijkstra, T., &
Hagoort, P. (2005). Theta responses are involved in lexical-
semantic retrieval during language processing. Journal of
Cognitive Neuroscience, 17(3), 530–541. https://doi.org/10
.1162/0898929053279469, PubMed: 15814011
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Bastiaansen, M.[C.], Magyari, L., & Hagoort, P. (2010). Syntactic
unification operations are reflected in oscillatory dynamics dur-
ing on-line sentence comprehension. Journal of Cognitive
Neuroscience, 22(7), 1333–1347. https://doi.org/10.1162/jocn
.2009.21283, PubMed: 19580386
Bastiaansen, M. C., van Berkum, J. J., & Hagoort, P. (2002).
Syntactic processing modulates the θ rhythm of the human
EEG. NeuroImage, 17(3), 1479–1492. https://doi.org/10.1006
/nimg.2002.1275, PubMed: 12414287
Batterink, L., & Neville, H. (2011). Implicit and explicit mechanisms
of word learning in a narrative context: An event-related potential
study. Journal of Cognitive Neuroscience, 23(11), 3181–3196.
https://doi.org/10.1162/jocn_a_00013, PubMed: 21452941
Bell, A. J., & Sejnowski, T. J. (1995). An information-maximization
approach to blind separation and blind deconvolution. Neural
Computation, 7(6), 1129–1159. https://doi.org/10.1162/neco
.1995.7.6.1129, PubMed: 7584893
Carey, S., & Bartlett, E. (1978). Acquiring a single new word. Papers
and Reports on Child Language Development, 15, 17–29.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including inde-
pendent component analysis. Journal of Neuroscience Methods,
134(1), 9–21. https://doi.org/10.1016/j.jneumeth.2003.10.009,
PubMed: 15102499
Neurobiology of Language
145
Meaning identification for novel words in context
Dollaghan, C., & Campbell, T. F. (1998). Nonword repetition and
child language impairment. Journal of Speech, Language, and
Hearing Research, 41(5), 1136–1146. https://doi.org/10.1044
/jslhr.4105.1136, PubMed: 9771635
Duff, M. C., & Brown-Schmidt, S. (2012). The hippocampus and
the flexible use and processing of language. Frontiers in
Human Neuroscience, 6, Article 69. https://doi.org/10.3389
/fnhum.2012.00069, PubMed: 22493573
Fenson, L., Marchman, V. A., Thal, D. J., Dale, P. S., Reznick, J. S.,
& Bates, E. (2006). The MacArthur-Bates communicative devel-
opment inventories user’s guide and technical manual (2nd ed.).
Brookes. https://doi.org/10.1037/t11538-000
Fukkink, R. G. (2005). Deriving word meaning from written con-
text: A process analysis. Learning and Instruction, 15(1), 23–43.
https://doi.org/10.1016/j.learninstruc.2004.12.002
Gao, Y., Wang, Q., Ding, Y., Wang, C., Li, H., Wu, X., Qu, T., & Li,
L. (2017). Selective attention enhances beta-band cortical oscil-
lation to speech under “cocktail-party” listening conditions.
Frontiers in Human Neuroscience, 11, Article 34. https://doi
.org/10.3389/fnhum.2017.00034, PubMed: 28239344
Hald, L. A., Bastiaansen, M. C., & Hagoort, P. (2006). EEG theta
and gamma responses to semantic violations in online sentence
processing. Brain and Language, 96(1), 90–105. https://doi.org
/10.1016/j.bandl.2005.06.007, PubMed: 16083953
Hanslmayr, S., Staudigl, T., & Fellner, M. C. (2012). Oscillatory
power decreases and long-term memory: The information via de-
synchronization hypothesis. Frontiers in Human Neuroscience,
6, Article 74. https://doi.org/10.3389/fnhum.2012.00074,
PubMed: 22514527
Herweg, N. A., Apitz, T., Leicht, G., Mulert, C., Fuentemilla, L., &
Bunzeck, N. (2016). Theta-alpha oscillations bind the hippocam-
pus, prefrontal cortex, and striatum during recollection: Evidence
from simultaneous EEG-fMRI. Journal of Neuroscience, 36(12),
3579–3587. https://doi.org/10.1523/JNEUROSCI.3629-15.2016,
PubMed: 27013686
Horst, J. S. (2013). Context and repetition in word learning.
Frontiers in Psychology, 4, Article 149. https://doi.org/10.3389
/fpsyg.2013.00149, PubMed: 23580347
Jenkins, J. R., Stein, M. L., & Wysocki, K. (1984). Learning vocab-
ulary through reading. American Educational Research Journal,
21(4), 767–787. https://doi.org/10.3102/00028312021004767
Jenkinson, N., & Brown, P. (2011). New insights into the relation-
ship between dopamine, beta oscillations and motor function.
Trends in Neurosciences, 34(12), 611–618. https://doi.org/10
.1016/j.tins.2011.09.003, PubMed: 22018805
Jensen, O., & Mazaheri, A. (2010). Shaping functional architecture
by oscillatory alpha activity: Gating by inhibition. Frontiers in
Human Neuroscience, 4, Article 186. https://doi.org/10.3389
/fnhum.2010.00186, PubMed: 21119777
Khader, P. H., & Rösler, F. (2011). EEG power changes reflect dis-
tinct mechanisms during long-term memory retrieval.
Psychophysiology, 48(3), 362–369. https://doi.org/10.1111/j
.1469-8986.2010.01063.x, PubMed: 20624249
Kielar, A., Meltzer, J. A., Moreno, S., Alain, C., & Bialystok, E.
(2014). Oscillatory responses to semantic and syntactic viola-
tions. Journal of Cognitive Neuroscience, 26(12), 2840–2862.
https://doi.org/10.1162/jocn_a_00670, PubMed: 24893735
Kleberg, F. I., Kitajo, K., Kawasaki, M., & Yamaguchi, Y. (2014).
Ongoing theta oscillations predict encoding of subjective mem-
ory type. Neuroscience Research, 83, 69–80. https://doi.org/10
.1016/j.neures.2014.02.010, PubMed: 24602686
Klimesch, W. (2012). (cid:1)-band oscillations, attention, and controlled
access to stored information. Trends in Cognitive Sciences, 16(12),
606–617. https://doi.org/10.1016/j.tics.2012.10.007, PubMed:
23141428
Klimesch, W., Doppelmayr, M., Yonelinas, A., Kroll, N. E., Lazzara,
M., Roehm, D., & Gruber, W. (2001). Theta synchronization dur-
ing episodic retrieval: Neural correlates of conscious awareness.
Cognitive Brain Research, 12(1), 33–38. https://doi.org/10.1016
/S0926-6410(01)00024-6, PubMed: 11489606
Klimesch, W., Sauseng, P., & Hanslmayr, S. (2007). EEG alpha os-
cillations: The inhibition–timing hypothesis. Brain Research
Reviews, 53(1), 63–88. https://doi.org/10.1016/j.brainresrev
.2006.06.003, PubMed: 16887192
Klimesch, W., & Schack, B. (2003). Activation of long-term mem-
ory by alpha oscillations in a working-memory task? Behavioral
and Brain Sciences, 26(6), 743. https://doi.org/10.1017
/S0140525X03370165
Klimesch, W., Schack, B., & Sauseng, P. (2005). The functional sig-
nificance of theta and upper alpha oscillations. Experimental
Psychology, 52(2), 99–108. https://doi.org/10.1027/1618-3169
.52.2.99, PubMed: 15850157
Krause, C. M., Grönholm, P., Leinonen, A., Laine, M., Säkkinen, A. L.,
& Söderholm, C. (2006). Modality matters: The effects of stimu-
lus modality on the 4- to 30-Hz brain electric oscillations during
a lexical decision task. Brain Research, 1110(1), 182–192.
https://doi.org/10.1016/j.brainres.2006.06.100, PubMed:
16901470
Lam, N. H., Schoffelen, J., Uddén, J., Hultén, A., & Hagoort, P.
(2016). Neural activity during sentence processing as reflected
in theta, alpha, beta, and gamma oscillations. NeuroImage,
142, 43–54. https://doi.org/10.1016/j.neuroimage.2016.03.007,
PubMed: 26970187
Lansink, C. S., Meijer, G. T., Lankelma, J. V., Vinck, M. A., Jackson,
J. C., & Pennartz, C. M. (2016). Reward expectancy strengthens
CA1 theta and beta band synchronization and hippocampal-
ventral striatal coupling. Journal of Neuroscience, 36(41),
10598–10610. https://doi.org/10.1523/ JNEUROSCI.0682-16
.2016, PubMed: 27733611
Lewis, A. G., & Bastiaansen, M. (2015). A predictive coding frame-
work for rapid neural dynamics during sentence-level language
comprehension. Cortex, 68, 155–168. https://doi.org/10.1016/j
.cortex.2015.02.014, PubMed: 25840879
Lewis, A. G., Schoffelen, J. M., Schriefers, H., & Bastiaansen, M.
(2016). A predictive coding perspective on beta oscillations dur-
ing sentence-level language comprehension. Frontiers in Human
Neuroscience, 10, Article 85. https://doi.org/10.3389/fnhum
.2016.00085, PubMed: 26973500
Marinkovic, K., Rosen, B. Q., Cox, B., & Kovacevic, S. (2012).
Event-related theta power during lexical-semantic retrieval and
decision conflict is modulated by alcohol intoxication: Anatomically
constrained MEG. Frontiers in Psychology, 3, Article 121. https://
doi.org/10.3389/fpsyg.2012.00121, PubMed: 22536192
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical testing
of EEG- and MEG-data. Journal of Neuroscience Methods, 164(1),
177–190. https://doi.org/10.1016/j.jneumeth.2007.03.024,
PubMed: 17517438
McLeod, A. N., & McDade, H. L. (2011). Preschoolers’ incidental
learning of novel words during storybook reading. Communication
Disorders Quarterly, 32(4), 256–266. https://doi.org/10.1177
/1525740109354777
Mestres-Missé, A., Camara, E., Rodriguez-Fornells, A., Rotte, M., &
Münte, T. F. (2008). Functional neuroanatomy of meaning acqui-
sition from context. Journal of Cognitive Neuroscience, 20(12),
2153–2166. https://doi.org/10.1162/jocn.2008.20150, PubMed:
18457509
Neurobiology of Language
146
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
o
_
a
_
0
0
0
5
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Meaning identification for novel words in context
Mestres-Missé, A., Rodriguez-Fornells, A., & Münte, T. F. (2007).
Watching the brain during meaning acquisition. Cerebral
Cortex, 17(8), 1858–1866. https://doi.org/10.1093/cercor
/bhl094, PubMed: 17056648
Meyer, L. (2018). The neural oscillations of speech processing and
language comprehension: State of the art and emerging mecha-
nisms. European Journal of Neuroscience, 48(7), 2609–2621.
https://doi.org/10.1111/ejn.13748, PubMed: 29055058
Nagy, W. E., Anderson, R. C., & Herman, P. A. (1987). Learning
word meanings from context during normal reading. American
Educational Research Journal, 24(2), 237–270. https://doi.org
/10.3102/00028312024002237
Nagy, W. E., Herman, P. A., & Anderson, R. C. (1985). Learning
words from context. Reading Research Quarterly, 20(2), 233–253.
https://doi.org/10.2307/747758
Nyhus, E., & Curran, T. (2010). Functional role of gamma and theta
oscillations in episodic memory. Neuroscience & Biobehavioral
Reviews, 34(7), 1023–1035. https://doi.org/10.1016/j.neubiorev
.2009.12.014, PubMed: 20060015
Obleser, J., & Weisz, N. (2012). Suppressed alpha oscillations pre-
dict intelligibility of speech and its acoustic details. Cerebral
Cortex, 22(11), 2466–2477. https://doi.org/10.1093/cercor
/bhr325, PubMed: 22100354
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J. M. (2011).
FieldTrip: Open source software for advanced analysis of
M E G , E E G , a n d i n v a s i v e e l e c t r o p h y s i o l o g i c a l d a t a .
Computational Intelligence and Neuroscience, 2011, Article
156869. https://doi.org/10.1155/2011/156869, PubMed:
21253357
Perfetti, C. A., Wlotko, E. W., & Hart, L. A. (2005). Word learning
and individual differences in word learning reflected in event-
related potentials. Journal of Experimental Psychology:
Learning, Memory, and Cognition, 31(6), 1281–1292. https://
doi.org/10.1037/0278-7393.31.6.1281, PubMed: 16393047
Pfurtscheller, G., & Silva, F. L. (1999). Event-related EEG/MEG syn-
chronization and desynchronization: Basic principles. Clinical
Neurophysiology, 110(11), 1842–1857. https://doi.org/10.1016
/S1388-2457(99)00141-8, PubMed: 10576479
Piai, V., Roelofs, A., & Maris, E. (2014). Oscillatory brain re-
sponses in spoken word production reflect lexical frequency
and sentential constraint. Neuropsychologia, 53, 146–156.
https://doi.org/10.1016/j.neuropsychologia.2013.11.014,
PubMed: 24291513
Prystauka, Y., & Lewis, A. G. (2019). The power of neural oscil-
lations to inform sentence comprehension: A linguistic per-
spective. Language and Linguistics Compass, 13(9), Article
e12347. https://doi.org/10.1111/ lnc3.12347, PubMed:
33042211
Pu, Y., Cheyne, D., Sun, Y., & Johnson, B. W. (2020). Theta oscil-
lations support the interface between language and memory.
NeuroImage, 215, Article 116782. https://doi.org/10.1016/j
.neuroimage.2020.116782, PubMed: 32276054
Ripollés, P., Marco-Pallarés, J., Alicart, H., Tempelmann, C.,
Rodríguez-Fornells, A., & Noesselt, T. (2016). Intrinsic monitoring
of learning success facilitates memory encoding via the activation
of the SN/ VTA-Hippocampal loop. eLife, 5, Article e17441.
https://doi.org/10.7554/eLife.17441, PubMed: 27644419
Ripollés, P., Marco-Pallarés, J., Hielscher, U., Mestres-Missé, A.,
Tempelmann, C., Heinze, H. J., Rodríguez-Fornells, A., &
Noesselt, T. (2014). The role of reward in word learning and its
implications for language acquisition. Current Biology, 24(21),
2606–2611. https://doi.org/10.1016/j.cub.2014.09.044,
PubMed: 25447993
Rommers, J., Dickson, D. S., Norton, J. J., Wlotko, E. W., &
Federmeier, K. D. (2017). Alpha and theta band dynamics related
to sentential constraint and word expectancy. Language,
Cognition and Neuroscience, 32(5), 576–589. https://doi.org/10
.1080/23273798.2016.1183799, PubMed: 28761896
Roux, F., & Uhlhaas, P. J. (2014). Working memory and neural
oscillations: Alpha–gamma versus theta–gamma codes for dis-
tinct WM information? Trends in Cognitive Sciences, 18(1),
16–25. https://doi.org/10.1016/j.tics.2013.10.010, PubMed:
24268290
Saffran, J. R., Newport, E. L., Aslin, R. N., Tunick, R. A., & Barrueco,
S. (1997). Incidental language learning: Listening (and learning)
out of the corner of your ear. Psychological Science, 8(2), 101–105.
https://doi.org/10.1111/j.1467-9280.1997.tb00690.x
Salisbury, D. F., & Taylor, G. (2012). Semantic priming increases
left hemisphere theta power and intertrial phase synchrony.
Psychophysiology, 49(3), 305–311. https://doi.org/10.1111/j
.1469-8986.2011.01318.x, PubMed: 22176140
Sassenhagen, J., & Draschkow, D. (2019). Cluster-based permuta-
tion tests of MEG/EEG data do not establish significance of effect
latency or location. Psychophysiology, 56(6), Article e13335.
https://doi.org/10.1111/psyp.13335, PubMed: 30657176
Schneider, J. M., & Maguire, M. J. (2018). Identifying the relation-
ship between oscillatory dynamics and event-related responses.
International Journal of Psychophysiology, 133, 182–192. https://
doi.org/10.1016/j.ijpsycho.2018.07.002, PubMed: 29981766
Scholz, S., Schneider, S. L., & Rose, M. (2017). Differential effects
of ongoing EEG beta and theta power on memory formation.
PLOS ONE, 12(2), Article e0171913. https://doi.org/10.1371
/journal.pone.0171913, PubMed: 28192459
Schwerdt, H. N., Amemori, K., Gibson, D. J., Stanwicks, L. L.,
Yoshida, T., Bichot, N. P., Amemori, S., Desimone, R., Langer,
R., Cima, M. J., & Graybiel, A. M. (2020). Dopamine and beta-
band oscillations differentially link to striatal value and motor
control. Science Advances, 6(39), Article eabb9226. https://doi
.org/10.1126/sciadv.abb9226, PubMed: 32978148
Shahin, A. J., Picton, T. W., & Miller, L. M. (2009). Brain oscilla-
tions during semantic evaluation of speech. Brain and
Cognition, 70(3), 259–266. https://doi.org/10.1016/j.bandc
.2009.02.008, PubMed: 19324486
Shtyrov, Y. (2011). Neural bases of rapid word learning.
Neuroscientist, 18(4), 312–319. https://doi.org/10.1177
/1073858411420299, PubMed: 22020546
Storkel, H. L. (2013). A corpus of consonant–vowel–consonant real
words and nonwords: Comparison of phonotactic probability,
neighborhood density, and consonant age of acquisition.
Behavior Research Methods, 45, 1159–1167. https://doi.org/10
.3758/s13428-012-0309-7, PubMed: 23307574
Strauß, A., Kotz, S. A., Scharinger, M., & Obleser, J. (2014). Alpha
and theta brain oscillations index dissociable processes in spo-
ken word recognition. NeuroImage, 97, 387–395. https://doi
.org/10.1016/j.neuroimage.2014.04.005, PubMed: 24747736
Terporten, R., Schoffelen, J. M., Dai, B., Hagoort, P., & Kösem, A.
(2019). The relation between alpha/beta oscillations and the en-
coding of sentence induced contextual information. Scientific
Reports, 9(1), Article 20255. https://doi.org/10.1038/s41598
-019-56600-x, PubMed: 31882830
Van Ede, F., De Lange, F., Jensen, O., & Maris, E. (2011). Orienting
attention to an upcoming tactile event involves a spatially and
temporally specific modulation of sensorimotor alpha-and beta-
band oscillations. Journal of Neuroscience, 31(6), 2016–2024.
https://doi.org/10.1523/ JNEUROSCI.5630-10.2011, PubMed:
21307240
Neurobiology of Language
147
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
o
_
a
_
0
0
0
5
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Meaning identification for novel words in context
Vassileiou, B., Meyer, L., Beese, C., & Friederici, A. D. (2018).
Alignment of alpha-band desynchronization with syntactic struc-
ture predicts successful sentence comprehension. NeuroImage,
175, 286–296. https://doi.org/10.1016/j.neuroimage.2018.04
.008, PubMed: 29627592
Wang, L., Hagoort, P., & Jensen, O. (2018). Language prediction is
reflected by coupling between frontal gamma and posterior alpha
oscillations. Journal of Cognitive Neuroscience, 30(3), 432–447.
https://doi.org/10.1162/jocn_a_01190, PubMed: 28949823
Wang, L., Jensen, O., van den Brink, D., Weder, N., Schoffelen,
J. M., Magyari, L., Hagoort, P., & Bastiaansen, M. (2012). Beta
oscillations relate to the N400m during language comprehen-
sion. Human Brain Mapping, 33(12), 2898–2912. https://doi
.org/10.1002/hbm.21410, PubMed: 22488914
Weiss, S., & Mueller, H. M. (2012). “Too many betas do not spoil
the broth:” The role of beta brain oscillations in language pro-
cessing. Frontiers in Psychology, 3, Article 201. https://doi.org
/10.3389/fpsyg.2012.00201, PubMed: 22737138
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
3
1
1
3
2
1
9
8
6
8
3
1
n
o
_
a
_
0
0
0
5
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
148