RESEARCH ARTICLE
Examining Individual Differences in Language
学习: A Neurocognitive Model of
Language Aptitude
Sabrina Turker1, Annemarie Seither-Preisler2,3
, and Susanne Maria Reiterer4
1Lise Meitner Research Group Cognition and Plasticity, 马克斯·普朗克人类认知和脑科学研究所,
莱比锡, 德国
2Centre for Systematic Musicology, University of Graz, Graz, 奥地利
3BioTechMed Graz, Graz, 奥地利
4语言学系, 维也纳大学, 维也纳, 奥地利
关键词: individual differences, language learning, neuroimaging, neurobiology, arcuate fascicle
抽象的
A common practice in the cognitive neurosciences is to investigate population-typical
现象, treating individuals as equal except for a few outliers that are usually discarded from
analyses or that disappear on group-level patterns. Only a few studies to date have captured the
heterogeneity of language processing across individuals as so-called “individual differences”;
fewer have explicitly researched language aptitude, which designates an individual’s ability for
acquiring foreign languages. Existing studies show that, relative to average learners, very gifted
language learners display different task-related patterns of functional activation and connectivity
during linguistic tasks, and structural differences in white and grey matter morphology, 并在
white matter connectivity. Despite growing interest in language aptitude, there is no recent
comprehensive review, nor a theoretical model to date that includes the neural level. To fill this
gap, we review neuroscientific research on individual differences in language learning and
language aptitude and present a first, preliminary neurocognitive model of language aptitude. 我们
suggest that language aptitude could arise from an advantageous neurocognitive profile, 哪个
leads to high intrinsic motivation and proactive engagement in language learning activities. 在
the neural level, interindividual differences in the morphology of the bilateral auditory cortex
constrain individual neural plasticity, as is evident in the speed and efficiency of language
学习. We suggest that language learning success is further dependent upon highly efficient
auditory-motor connections (speech-motor networks) and the structural characteristics of dorsal
and ventral fibre tracts during language learning.
介绍
Among the numerous social, cognitive, and affective variables that influence second language
learning success (see Ellis, 2004, for an overview), research suggests that motivation, age of
onset, and language aptitude are by far the most influential predictors (Abrahamsson &
Hyltenstam, 2008; Birdsong & Molis, 2001; Dörnyei & Skehan, 2003). 换句话说, 在下面
the same learning circumstances, differences in an individual’s initial state of readiness and
capacity to acquire foreign languages, 那是, their language aptitude (卡罗尔, 1981), 将要
largely determine their ultimate attainment (Doughty, 2019). In our own understanding,
开放访问
杂志
引文: Turker, S。, Seither-Preisler, A。,
& Reiterer, S. 中号. (2021). 正在检查
individual differences in language
学习: A neurocognitive model of
language aptitude. Neurobiology of
语言, 2(3), 389–415. https://doi.org
/10.1162/nol_a_00042
DOI:
https://doi.org/10.1162/nol_a_00042
已收到: 30 七月 2020
公认: 13 可能 2021
利益争夺: 作者有
声明不存在竞争利益
存在.
通讯作者:
Sabrina Turker
turker@cbs.mpg.de
turker.sabrina@gmail.com
处理编辑器:
Patrick Wong
版权: © 2021
麻省理工学院
在知识共享下发布
归因 4.0 国际的
(抄送 4.0) 执照
麻省理工学院出版社
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
Language aptitude:
An individual’s (largely innate)
capacity for acquiring foreign
languages without much effort and
faster as compared to peers.
Phonetic coding ability:
A component of language aptitude
that includes the identification of
声音, the formation of associations
between sounds and letters, 和
ability to retain these associations.
language aptitude is at least partly genetically determined. Most researchers, 然而, empha-
size that the concrete nature of language aptitude is yet to be determined through future re-
搜索, and describe it as something between a stable, fixed trait (see evidence provided by
韦尔斯, 1986) and a plastic, malleable dynamic state (see discussions in DeKeyser, 2019, 和
文等人。, 2017). Language aptitude is further believed to comprise a set of abilities, 包括
language analytical abilities, phonetic coding ability, rote memory (文等人。, 2017), 和
工作记忆 (Wen, 2019). The most widely accepted models of language aptitude (和-
marized in Wen et al., 2017), namely Skehan’s cognitive “Processing Stages” (Skehan, 1998,
2002, 2016) and Robinson’s interactive and pragmatically oriented “Aptitude Complex
Hypothesis” (磷. 罗宾逊, 2001, 2012), address several cognitive and environmental vari-
埃布尔斯, but do not explicitly address the neurocognitive basis of their frameworks. 因此, 它是-
mains unclear which brain areas and networks are involved in language aptitude, to what
extent they modulate language learning success, and how they develop, and hence rely on
potentially innate and/or prenatal factors. 同样地, research on individual differences in sec-
ond language learning has for a long time focused on observable differences in behaviour (看
a recent summary in Kidd et al., 2018), although it is clear that these differences are influenced
by domain-general abilities (例如, 智力, 记忆; Ellis, 2004), which are rooted in brain
structure and function (坎贝尔 & Tyler, 2018).
一般来说, the brain areas most likely implicated in language aptitude are those that con-
tribute centrally to language comprehension and production (Price, 2010, 2012), 认识,
和, 记忆 (例如, see review by Biedron(西德:1), 2015). These are primarily left frontal and
perisylvian regions (inferior frontal, temporal and inferior parietal; as presented in the model
of Hickok & Poeppel, 2004; Petrides, 2014), with contributions of right homologous regions as
出色地 (Vigneau et al., 2011). In the past two decades, the number of studies exploring the neu-
robiological bases of language aptitude and individual differences in language learning has
constantly increased. 然而, uncovering the neural basis of language aptitude requires ad-
dressing its development both through nature and nurture—its genetic basis, in utero influ-
恩塞斯, peri- and postnatal factors, as well as its further development from childhood to
青春期. Even if research with infants and children is scarce, studies on individual differ-
ences in language processing and learning on a more general level can provide vital insights
into the neural basis of language aptitude.
We here provide a comprehensive review of research on the neurobiology of language ap-
titude and individual differences in language learning, and a first, preliminary neurocognitive
model of language aptitude. 第一的, we discuss previous research exploring differences in struc-
tural and functional connectivity, functional activation patterns, and brain morphology.
第二, we discuss the nature and nurture of language aptitude, which serves as a basis for
the following model. 第三, we present our model and elaborate on the neurocognitive basis of
language aptitude, as well as the neurobiological differences and changes associated with lan-
guage learning and high aptitude. 最后的, we present future avenues for research.
背景
Only a few studies to date have investigated the neural underpinnings of language aptitude.
Considerably more but still limited research has aimed to capture the heterogeneity of lan-
guage processing across individuals as so-called “individual differences” in language learning,
which are evident in all domains and components of the language system (基德等人。, 2018;
于 & Zellou, 2019), and comprise neurophysiological, neuroanatomical, cognitive, 和每-
ceptual levels (磷. C. 中号. 黄 & Ettlinger, 2011). For the following presentation of previous
Neurobiology of Language
390
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
Language analytic abilities:
A component of language aptitude
that designates the ability to analyse
语言 (mostly explicitly) 和
arrive at linguistic generalizations
(IE。, draw conclusions and infer rules
and regularities).
Sound-symbol learning:
One part of the LLAMA language
aptitude battery in which subjects
have to learn a new sound-symbol
系统 (IE。, build associations
between new linguistic units and
their corresponding auditory form).
Arcuate fascicle:
The major language-related white
matter fibre tract that connects
temporal areas with frontal areas via
the inferior parietal lobe.
研究, we divided research studies according to whether they explored structural or func-
tional underpinnings, but we emphasize the constant interaction of the two in our model pre-
sented later (see the section, A Neurocognitive Model of Language Aptitude).
The Neurobiology of Language Aptitude
Brain structure as an indicator for high language learning abilities
Structural characteristics related to high language aptitude have been found in grey and white
matter volumes of the left inferior parietal lobe (IPL), the auditory cortices, and the left inferior
frontal cortex. A study by Reiterer et al. (2011) reported higher grey matter volumes in the left IPL
and the left inferior frontal/motor compound in more gifted speech imitators during the imitation
of an unknown language. 相似地, higher grey matter volumes and a stronger gyrification (IE。, A
more frequent occurrence of multiple gyri) were found in the right auditory cortices of children
and adults with high speech imitation skills and high overall language aptitude (包括
language analytical abilities, vocabulary learning, and phonetic coding ability) (Turker et al.,
2017, 2019). In these two studies, possessing a single gyrus as opposed to multiple gyri in the
right hemisphere was associated with low language aptitude scores. 最近, higher cortical
thickness in Broca’s area (left inferior frontal gyrus/IFG) and the left posterior-medial frontal lobe
were related to higher language analytic abilities in adults. The thicker the cortex was, the higher
the language analytic ability scores were (Novén et al., 2019).
Investigating white matter connectivity between language-related brain areas, Xiang et al.
(2012) reported that each of four administered language aptitude tasks was differentially related
to the strength of different white matter tracts. Sound-symbol correspondence learning, 为了
实例, could be best predicted by the interhemispheric connections between left and right
posterior IFG. Structural connections in the left temporal pathway (connecting the left inferior
frontal cortex and temporal areas) predicted grammatical inferencing abilities, while connec-
tions in the left parietal pathway (connecting the left anterior IFG and the IPL) 预测的
vocabulary learning and sound-symbol learning. In another study, higher fractional anisotropy
of the left arcuate fascicle correlated with high speech imitation abilities, while higher volume
of the posterior right arcuate fascicle correlated with low speech imitation ability (Vaquero
等人。, 2017). 反过来, the anterior segment of the right arcuate fascicle was linked to high
grammatical analytical abilities in another study (Kepinska, Lakke, 等人。, 2017), meaning that
speech imitation and analytical abilities could be differentially reliant on segments of the
arcuate fascicle.
总共, whereas the auditory cortex seems to be important for overall language aptitude and
speech imitation, inferior frontal and motor areas might be particularly important for language
analytic abilities and speech imitation. The role of the left IPL is potentially related to phono-
logical processes rooted in the temporo-parietal junction; and different segments of the arcuate
fascicle, both in the right and left hemisphere, seem to be related to specific skills associated
with language aptitude. 然而, more research is needed to verify these preliminary results
and specifically, to determine the role of right-hemispheric white matter fibre tracts.
Brain function as an indicator for high language learning abilities
Studies that investigated functional activation have reported divergent and partially contradicting
results so far. Reiterer et al. (2011) found remarkable individual differences in how subjects
employed left-hemispheric speech areas during speech imitation tasks. Those with high
speech imitation ability showed decreased brain activation in left frontal and parietal areas.
This supports the hypothesis that high neural efficiency is associated with more focal brain
Neurobiology of Language
391
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
activation (Neubauer & Fink, 2009). 反过来, 胡等. (2013) reported more widespread
and higher functional activation in auditory perceptual and speech motor areas in learners
with better pronunciation. In this study, subjects’ English pronunciation skills, as well as their
phonetic coding ability, were assessed and related to brain activation during a speech imita-
tion task. On the behavioural level, phonetic coding ability was strongly tied to pronunciation
proficiency. On the neural level, brain activation during speech imitation differed significantly
between those with high and low pronunciation proficiency. More widespread activation was
also found in learners with high language analytic ability (Kepinska, de Rover, 等人。, 2017),
who displayed greater activation in terms of magnitude and extent in the left IPL and the right
temporal cortex. The involvement of the right hemisphere in high language learning abilities
was confirmed in a later EEG study (Kepinska, Pereda, 等人。, 2017), where higher proficiency
during artificial grammar learning was supported by stronger local synchronisation in the right
hemisphere, combined with less mental effort in the learners with high language analytic abil-
性 (for further evidence that low aptitude speakers might have fewer cognitive resources and
struggle with increasing cognitive load, see Antoniou & 磷. C. 中号. 黄, 2015).
总结一下, studies investigating language aptitude with neuroimaging have primarily
assessed speech imitation ability, pronunciation, and language analytic ability. For speech im-
itation and pronunciation, left-hemispheric language and in particular speech motor and au-
ditory areas seem to play a dominant role, whereas language analytical abilities seem to be
more tied to left inferior frontal and right-hemispheric brain activation. The observed differ-
ences between these studies (more focal vs. more widespread activation) could reflect differ-
ences in assessment of skills (IE。, which tasks were used to determine high or low language
learning status) and applied tasks outside and inside of the scanner.
The Neurobiology of Individual Differences in Language Learning
Similarly to language aptitude, individual differences in language learning success present as dif-
ferences in grey matter volume, functional activation and connectivity patterns, and even func-
tional connectivity in language-related areas during rest (see the review by Li & 授予, 2016).
Brain structure and individual differences in language learning
Research on individual differences in language learning suggests that the morphology of the
left IPL, IFG, and auditory cortices (尤其, Heschl’s gyrus (HG)) is strongly related to lan-
guage learning success. Longitudinal evidence suggests that increasing grey matter density in
the left IPL predicts foreign language learning success (Della Rosa et al., 2013). 此外,
there is evidence for the importance of the auditory cortices for various language learning-
related processes and mechanisms. 例如, higher white matter density in the left audi-
tory cortex was found to be significantly linked to novel speech sound learning (Golestani
等人。, 2002; see summary in Golestani, 2014). In the study by Golestani et al. (2007), differ-
ences in the gyrification of the left HG were also found between good and poor phonetic
learners. 相似地, higher grey and white matter volumes in left and right HG also distin-
guished expert phoneticians from a control group (Golestani et al., 2011). The same study also
reported a higher likelihood of two complete HGs in phoneticians. Apart from the auditory
cortex, the left IFG, a hub for neural activities (哪个 & 李, 2012), has been linked to success
in artificial grammar learning (Flöel et al., 2009). In that study, white matter integrity in fibres
arising from the left IFG, but not the right IFG, correlated with interindividual variability of
grammar learning performance, hinting towards a role of that area for grammar rule extraction.
而且, higher grey matter volumes in the left IFG were associated with an increase in pro-
ficiency during intensive foreign language learning, which is why the authors of that study
Neurobiology of Language
392
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
(Stein et al., 2012) suggested that an individual’s amount of learning was reflected in brain
structure changes regardless of absolute proficiency.
Other areas structurally relevant for L2 learning seem to include the anterior cingulate
(Legault, 授予, 等人。, 2019), although differences in this study depended on the learning context
还有 (Legault, Fang, 等人。, 2019). 相似地, another study suggests that multilingualism is
linked to grey matter volumes of the bilateral caudate, and an enlargement of the dorsal and
anterior portions of the left caudate nucleus prelearning (Hervais-Adelman, Egorova, &
Golestani, 2017).
Multiple fibre tracts in the human brain are involved in successful language processing and
thus most probably in language learning. 例如, Dick and Tremblay (2012) differentiate
between two dorsal (superior longitudinal fasciculus and arcuate fasciculus) and five ventral
fibre pathways (see also discussions in Catani et al., 2005, and Glasser & Rilling, 2008). 这
linguistic model proposed by Hagoort (2014) includes a selection of these tracts, and he
emphasizes the importance of the (subdivisions of the) arcuate fascicle, the inferior fronto-
occipital fasciculus, and the frontal aslant tract for all linguistics tasks (for a different proposal,
see Catani & Bambini, 2014). Concerning white matter fibre tracts and their relationship to in-
dividual differences, a single study found that fractional anisotropy of the left parieto-temporal
路径 (dorsal fibre tract and part of the arcuate fascicle) was positively correlated with sound-
to-word learning (F. C. K. Wong et al., 2011). In the same study, a ventral pathway involving the
extreme capsule and the left inferior longitudinal fasciculus, 另一方面, mediated
auditory comprehension.
总共, grey and white matter differences in the auditory cortices have been reported to
relate to speech sound learning, and studies have emphasized the involvement of the left
IFG for language-specific individual differences, especially grammatical learning. 个人
studies further suggest the involvement of left inferior parietal regions, the anterior cingulate
cortex, the bilateral caudates, and the left parieto-temporal pathway for high language learning
能力.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Brain function and individual differences in language learning
Individual differences in brain function comprise differences in neural efficiency, neural adapt-
能力, and functional synchronization (Prat et al., 2007, 2019; Prat, 2011; Prat & Just, 2011).
Prat and colleagues reported differences in resting state brain rhythms (accounting for 26–60%
of variance during intensive language learning; Prat et al., 2019); an increase of functional
connectivity due to higher demands (IE。, an interaction between synchronization and adapt-
能力); and generally greater efficiency and synchronization in several language-related areas
in successful language learners (Prat et al., 2007; Prat & Just, 2011). Greater or optimized cor-
tical processing efficiency has also been shown in polyglots (例如, see Jouravlev et al., 2021)
and second language learning experts (Reiterer, Berger, 等人。, 2005; Reiterer, Hemmelmann,
等人。, 2005). Reiterer et al. (2009) further found that the non-language students showed an
increase in phase synchrony of the lower gamma frequency range, particularly in the right
hemisphere. These significant increases involved right frontal and parietal regions: 一般来说,
the higher the proficiency level, the lower the synchronization density. 相似地, higher intrin-
sic functional connectivity within left posterior temporal areas (Chai et al., 2016) and increased
global network efficiency with distinct network patterns (Sheppard et al., 2012) were found to
be predictive of L2 word learning capacity.
With regard to temporal and inferior parietal areas, Veroude et al. (2010) and Assaneo et al.
(2019) reported individual differences in implicit auditory/phonological learning mechanisms.
Neurobiology of Language
393
Neurobiology of language aptitude
In the latter study, subjects who implicitly aligned their own concurrent syllable production to
a perceived syllable rate (termed “high synchronizers” by the authors) outperformed those who
remained impervious to external rhythm during a word learning task. On the neural level, 这
high synchronizers showed a highly interconnected audio-motor network and better synchro-
nization in left frontal areas (see also Poeppel & Assaneo, 2020). Further studies have shown
that the degree and local efficiency of the left superior temporal gyrus (STG) is linked to sound-
to-word learning performance, predicting future spoken language learning success (Deng
等人。, 2016). 而且, pretest functional connectivity between the left insula/frontal opercu-
lum and the left superior temporal lobe predicted non-native sound contrast learning (Ventura-
Campos et al., 2013). In a study by P. C. 中号. 黄, Perrachione, and Parrish (2007), 学习
to detect and use lexical pitch led to an increase in activation in the left posterior STG in the
successful learners, who already showed higher activation in bilateral temporal areas and the
right inferior temporal gyrus pretraining. The less successful learners, 另一方面, 前任-
hibited an increase in activation in the right STG and the right IFG, as well as prefrontal and
medial frontal areas. With regard to the left IPL, Barbeau et al. (2017) reported learning-
induced plasticity effects during intensive language learning. In their study, functional activa-
tion of the left IPL before the onset of training predicted posttraining attainment. 而且, 一个
increase in activation of the left IPL (具体来说, the supramarginal gyrus) accompanied lan-
guage learning, and higher activation in that region correlated with second language reading
speed postlearning.
Studies have also provided evidence for a role of the left IFG in tonal vocabulary learning
(杨等人。, 2015), lexical pitch learning (Qi et al., 2019), and statistical word segmentation
学习 (Karuza et al., 2013). 进一步来说, Yang et al. (2015) found that successful
learners of Chinese showed distinct patterns in core language areas during tonal and lexical
information processing and an overall more coherent and better integrated multipath brain
网络. Qi et al. (2019), 另一方面, found that greater pretraining activation of the
right IFG was associated with better ultimate attainment. The key to success in these learners
was greater pre- to postreduction of right IFG activation, coupled with enhanced resting-state
connectivity between the right and left IFG and the left superior parietal lobe. 全面的, 学习
was accompanied by increasing activation in left IFG and left superior parietal areas. 最后的,
Karuza et al. (2013) reported significant activity during statistical word segmentation learning
in pars opercularis and pars triangularis of the left inferior frontal cortex, and suggested poten-
tial parallels to the frontal/subcortical network involved in implicit sequence learning.
而且, when progress is made during foreign language learning, shorter activation of left
IFG occurs, which has been interpreted as a faster L2 processing (Stein et al., 2006).
Studies show that individual differences in brain function often present as greater efficiency
and higher synchronization of language-related brain areas in both hemispheres. 具体来说,
learning success involves the functioning of left temporal, left parietal, and left inferior frontal
地区. 然而, right-hemispheric areas seem to be involved in a multitude of tasks and in
successful as well as nonsuccessful learners, which requires further research.
A potential role of subcortical structures for language learning
Individual differences in language learning have also been confirmed at subcortical levels, 对于前任-
充足, in the inferior colliculus for the encoding of non-native phonemes (Chandrasekaran et al.,
2012) or the left hippocampus for vocabulary acquisition due to its role in episodic learning
(Breitenstein et al., 2005). As discussed later on, implicit and explicit learning systems are vital
for foreign language learning (see a recent summary on the model proposed by Ullman, 2015).
The findings that implicit, subcortical learning systems (例如, in the basal ganglia; see review in
Neurobiology of Language
394
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
Ullman, 2004) may be altered in individuals with developmental language disorders and dyslexia
(Krishnan et al., 2016) suggest that these systems may play a significant role in various language-
learning tasks in children and adults. 这, 反过来, could mean that language aptitude is highly
dependent upon subcortical structures, which certainly requires future research. The same could
be true for procedural learning, which is very likely to be implicated in rule-learning.
THE NEUROCOGNITIVE BASIS OF LANGUAGE APTITUDE AND ITS HERITABILITY
The Auditory Cortex and Its Role in Language Learning
From the current status of knowledge, the individual morphology of the human brain is at least
partly genetically determined (Pol et al., 2006; Strike et al., 2015), although genetic influences
on cortical morphology are a highly complex issue (陈等人。, 2013). Prenatal experience with
speech shapes speech perception and production in newborns so that speech processing in
newborns already displays a specialization for it (Gervain, 2015). Already in the second and
third trimester of pregnancy, language-related areas show distinct patterns and asymmetries:
Most sulci, including the Sylvian fissure, become visible between gestational weeks 21 到 27
(Bernard et al., 1988) and even individual gyral characteristics in structures like HG appear be-
tween gestational weeks 18 和 41 (López Ramón y Cajal, 2019). The primary cortical shapes
and sulcal folding patterns are prenatally determined and under strong spatio-temporal genetic
控制 (陈等人。, 2013; Thompson et al., 2001; for an in-depth review, see Im & 授予, 2019).
While Bartley et al. (1997) reported that only 7–17% of gyral patterning of the entire brain
appears to be due to genetic influences, studies with mono- and dizygotic twins have shown
that the anatomy of HG is estimated to have a higher genetic determination (例如, see Peper
等人。, 2007), with estimates of up to 80% (Pol et al., 2006); this genetic stability is supported
by longitudinal behavioural and neural observations (Seither-Preisler et al., 2014). 然而,
the specific genetic variants involved in HG morphology remain yet to be further determined
(Cai et al., 2014) . It is assumed that the total variance in interindividual brain anatomy can
be explained by (A) genetic, (乙) in utero/prenatal, 和 (C) external postnatal factors (Carmelli
等人。, 2002). Dubois et al. (2010) reported large asymmetries in the superior temporal sulcus
(also confirmed in Glasel et al., 2011), the planum temporale, and the anterior region of the
Sylvian fissure (inferior frontal) in newborns from 26 到 36 weeks of gestation, providing evi-
dence that the functional lateralization of language processing begins to manifest quite early.
The auditory cortex presents with large intra- and interindividual differences as revealed in
early postmortem investigations (奥尔巴赫, 1906; Heschl, 1878) and neuroimaging studies
(Geschwind & Levitsky, 1968; Marie et al., 2016; Penhune et al., 1996; Rademacher et al.,
2001; 磷. Schneider et al., 2002, 2005). These studies showed that HG appears as a single gyrus,
a common stem duplication (partial division by a sulcus intermedius), a complete duplication
(IE。, two complete gyri), or in the form of multiple gyri (Benner et al., 2017; da Costa et al., 2011).
Duplications occur in every second or third individual (Marie et al., 2016) and the presence of
multiple gyri has been linked to developmental conditions (Seither-Preisler et al., 2014), 还有
as to high musical ability (Benner et al., 2017; 磷. Schneider et al., 2005; Wengenroth et al.,
2014).
Previous research on language-related skills (speech perception, sound learning) 已经表明
the significant involvement of the bilateral auditory cortices (also concerning the number of
gyri) in phonological processing, pitch pattern learning (磷. C. 中号. Wong et al., 2008), and sec-
ond language learning (Golestani et al., 2007, 2011; Ressel et al., 2012). These findings com-
prised both the structural and functional level. Ressel and colleagues found a significant
correlation between larger grey matter volumes in left HG and bilingualism, indicating that
Neurobiology of Language
395
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
bilingual language experience might alter grey matter volume in the auditory cortex (Ressel
等人。, 2012). In the study by Wong and colleagues (2008), less successful foreign pitch pattern
learners possessed smaller grey and white matter volumes in the left auditory cortex only (不是
in the right, 出奇). In their review, Wong and Ettlinger (2011) reported differences in
lexical pitch learning on the neuroanatomical (larger HG volumes) and the neurofunctional
等级 (higher bilateral activation). In the studies by Golestani and colleagues, higher white mat-
ter volumes in the left auditory cortices were associated with better foreign speech sound
学习 (Golestani et al., 2007). 而且, they found a link between the occurrence of mul-
tiple and split HGs in the bilateral auditory cortices of expert phoneticians (Golestani et al.,
2011). 相似地, in previous studies on language aptitude, the occurrence of multiple gyri and
higher grey matter volumes of HG in the right hemisphere predicted high overall language
aptitude and high speech imitation ability in adults and children, whereas the occurrence
of single gyri was linked to low overall language aptitude (Turker et al., 2017, 2019).
Golestani and colleagues suggested that the presence of morphological differences in the
auditory cortex could be interpreted as an intermediate phenotype for auditory-related
domain-specific aptitude (Golestani et al., 2011). The idea that auditory cortex morphology is
predetermined and not a result of learning experience is further supported by several studies
(例如, Benner et al., 2017; Seither-Preisler et al., 2014; Serrallach et al., 2016). Seither-Preisler
等人. (2014), 例如, found large interindividual differences in the grey matter volume of
right HG, which were associated with musical aptitude. 然而, the longitudinal data re-
vealed that musical training itself did not result in any neuroanatomical changes (那些是
only observed on the neurofunctional level of neural efficiency).
But why would auditory areas be so important for language aptitude? Auditory processing is
the first capacity that develops in the fetus and the correct functioning of the auditory cortex is
crucial for spoken language development (Mueller et al., 2012; Skeide & Friederici, 2016).
Already during gestational weeks 28–33, the bilateral posterior STG shows mismatch re-
sponses to sounds and syllables, reflecting early auditory/phonetic abilities in the fetus
(Mahmoudzadeh et al., 2013). Since there is a bias towards language-specific frequency spec-
tra at that early developmental age, it is highly likely that primary, intrauterine speech percep-
tion is genetically driven (Skeide & Friederici, 2016). Soon after birth, the auditory areas are
connected to the motor cortex through a myelinated fibre tract, which is crucial for developing
phoneme representations in the brain (Dubois et al., 2010; Perani et al., 2011). The right sec-
ondary auditory cortex has been shown to respond to sentences, which might indicate a reli-
ance on suprasegmental information (例如, stress, melody, or intonation) during the first stage of
language development (Homae et al., 2006). During months 6–12, infants are already capable
of detecting phonological word forms, but at the same time perceptual narrowing takes place,
meaning that an infant’s phonological system is established and their brain becomes special-
ized for the language they encounter daily (Pons et al., 2009). Few children, so-called “early
talkers,” even show full-sentence highly developed verbal communication skills at 2 years of
年龄. This likely reflects a proactive willingness to communicate and high intrinsic motivation
to engage in language activities, which is claimed to be a marker for enhanced language ap-
titude (总的, 1999; Winner, 1996).
The Heritability of Language Aptitude
In the past years, researchers have started to identify the genes involved in speech and language,
but we are still at the beginning (see the review by Fisher, 2017). Twin and heritability studies
suggest that the rate of language acquisition and the linguistic proficiency attained by an indi-
vidual are largely determined by genetic factors (Stromswold, 2001; Verhoef et al., 2020; 看
Neurobiology of Language
396
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
Learning efficiency:
An individual’s capacity to learn,
store, and consolidate new
信息, comprising primarily
associative memory in the CHC
模型.
also discussion on the latter in Chow & S. 瓦. 黄, 2021), which is supported by neuroscien-
tific studies finding that second language learning correlates with white matter fibre tracts and
genetic variation (Mamiya et al., 2016). With regard to general cognitive ability, heritability has
been found to increase linearly from child- to adulthood (Plomin et al., 2016), with genetic in-
fluence accounting for 41% of individual differences in behaviour in 9-year-old children, 55%
in 12-year-olds, 和 66% in adolescents aged 17 (Haworth et al., 2010). 最后, 最多 80% 的
variance in full-scale IQ, also comprising language-related skills and general intelligence, 能
be accounted for by genetic variance components in adults (Plomin & Deary, 2015). Focusing
on second/foreign language learning specifically, twin studies with children, teenagers, 和
young adults suggest moderate to high heritability estimates of 42–72% (42% in Dale et al.,
2012; 67% in Dale et al., 2010; 71% in Vinkhuyzen et al., 2009; 72% in Coventry et al.,
2012). Rimfeld et al. (2015) reported that all measures of second language learning showed high
heritability (36–62%) even for different first languages and that one third of the genetic influence
in second language learning was shared with intelligence. Targeting much younger children,
Rice et al. (2018) calculated heritability estimates in the range of 44–92% in 6-year-old children,
with the highest estimate being at 0.92 for grammar, and an increase in heritability from 4 到
6 年龄.
It seems very likely that adults will gain higher levels of foreign language mastery the higher
their predetermined aptitude and the more prominent the neural perquisites and changes.
These conclusions are supported by studies that found that native-like proficiency in foreign
language learning is rare and most likely a result of language aptitude, not age of onset (看
Abrahamsson & Hyltenstam, 2008, 2009). Although inherited genetic information may set
limits on what can be achieved by an individual, it is the environment that determines what
individuals actually accomplish (Dörnyei, 2014; for a discussion on gene-environment inter-
actions in language learning, see Onnis et al., 2018). 过去十年, the emerging field of
epigenetics has allowed valuable new insights into the interplay between nature and nurture,
also on the neural level (例如, refer to Dehaene-Lambertz et al., 2006). It has shown not only
that genes have an effect on learning efficiency and general cognitive abilities, but also that
learning experience has a reciprocal effect on gene expression (Bjorklund & Causey, 2018),
which is particularly interesting for education (Mc Ewen, 2015).
全面的, it seems that language-related abilities, such as language aptitude, are highly her-
itable and genetics might exert a larger influence at a later age. 然而, more research is
needed to pinpoint the genetic underpinnings of language, and further describe gene-
environment interactions with regard to language.
The Role of Cognitive Abilities, Musicality, and Memory During Language Learning
Cognitive abilities develop and come into play at different stages of language learning. 自从
hearing develops in the fetus long before birth (around gestational week 27; Hepper &
Shahidullah, 1994), it is not surprising that fetuses already show mismatch responses to unex-
pected sounds and tones (Huotilainen et al., 2005). 有趣的是, even in that early stage slight
individual differences have been observed (Draganova et al., 2005). One-year-old infants have
been shown to possess working memory in the visual (Ross-Sheehy et al., 2003) and auditory
modality (Ross-Sheehy et al., 2003), but still show clear capacity limits. Concerning other cog-
nitive capacities, Demetriou et al. (2014) postulated that fluid intelligence develops in four
reconceptualization cycles between 2 和 16 years of age and suggested that working mem-
ory and processing speed are vital for fluid intelligence development. While a strong link be-
tween the three is likely, other researchers have proposed that the developmental increases in
Neurobiology of Language
397
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
Fluid reasoning:
The ability to perform non-automatic
mental operations comprising
induction, sequential reasoning, 和
quantitative reasoning.
working memory arise primarily from improvements in other cognitive domains, such as at-
注意力, encoding, processing speed, and retrieval (Towse & Hitch, 2007). Most likely, 在职的
记忆, the portal to long-term memory, underpins cognitive processes (考恩, 2014), 和
largely interacts with learning efficiency and fluid reasoning (see discussion of the specific
cognitive components that are essential for language learning in the section, Environmental
Influences and Gene-Environment Interactions).
Musicality has also been shown to impact language learning (see recent review on interac-
tions between language aptitude and music on the neural level by Turker & Reiterer, 2021). 在
the early stages of language learning, infants are acutely sensitive to prosodic patterns, 哪个
enables later phonological development (Gervain & Mehler, 2010). This suggests that this
quasi-musical auditory structuring ability (Karma, 1994) can be advantageous for language
学习. In line with our assumption that cognitive abilities are largely genetically driven, 高的
musicality in the form of better rhythmic and melodic perception could be well linked to lan-
guage aptitude at the genetic level. Recent genomic studies, 例如, have hinted towards
a link between musical aptitude-related genes and auditory perception, cognitive perfor-
曼斯, 记忆, and language acquisition (see Järvelä, 2018, for a review). 像这样, 语言
and music would directly impact one another, but both would unfold and develop side by
边, making a classical one-before-the-other-distinction unnecessary. This could perhaps take
the form of an overarching compound of speech-music abilities, tied together by auditory and
fine motor abilities with a common root. 像这样, advanced musical processing abilities could
lead to better language perception, which in turn would result in higher musical discrimination
能力 (Bowles et al., 2016; Delogu et al., 2008; Nardo & Reiterer, 2009; Schön et al., 2004).
This could explain the frequent co-occurrence of speech imitation talent and musicality in
adult language learners (Christiner & Reiterer, 2013; Nardo & Reiterer, 2009) even as early as
in 4–6 year old children (Christiner & Reiterer, 2018). 而且, research has further pointed
towards a link between musical experience and higher success during the learning of phonetic
aspects of languages (例如, perception of pitch patterns; 磷. C. 中号. 黄 & Perrachione, 2007),
potentially due to their advantage in the discrimination of musical features, like tones.
It remains to be uncovered how (什么时候) declarative and procedural memory, 二
domain-general, cognitive, long-term memory storage systems, are related to foreign language
learning success. 全面的, it seems that declarative memory could be highly relevant in early
L2 learning stages, and procedural memory in later learning stages (例如, Faretta-Stutenberg &
Morgan-Short, 2018). 此外, another study found that declarative memory was more
relevant in implicit, exposure-based learning circumstances, while procedural memory was
vital during incidental, immersive context-based learning (Antoniou et al., 2016) (see a
thorough discussion in Buffington & Morgan-Short, 2019). 最近, Hamrick et al. (2018) 亲-
vided clear evidence for a significant role of general-purpose learning systems in both first and
foreign language acquisition in several meta-analyses. 全面的, they reported that lexical abil-
ity was associated with declarative learning in all learners, while grammar was linked to de-
clarative memory in low ability second language learners, and to procedural learning in the
highly competent learners. Neurally, Morgan-Short and colleagues (Morgan-Short et al., 2014,
2015) found that some learners used the neural circuits of their first language and their proce-
dural memory circuits when implicitly learning an L2, while others engaged extralinguistic
neural circuits. It has also been suggested that procedural memory shares neural substrates
with grammar learning (例如, Broca’s area and basal ganglia; Ullman, 2004), since procedural
memory has been associated with performance for simple words during a morphophonologi-
cal grammar learning task (Antoniou et al., 2016; Ettlinger et al., 2014). Declarative and pro-
cedural learning, alongside working memory, could therefore be central ingredients to
Neurobiology of Language
398
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
language aptitude. Considering these developments together with overall language learning, 它
becomes clear that all are essential prerequisites of first language acquisition, which in turn
shows large long-term crosslinguistic transfers to foreign language learning (Sparks, 2012;
Sparks et al., 2009).
Neural Plasticity and Language Aptitude
学习, the basis of intelligent behaviour, is caused by plastic changes in neural assemblies
(Partanen et al., 2013). As Zhang and Wang (2007) summarize, learning-induced plasticity can
present as (1) higher neural sensitivity, (2) increased neural specificity (IE。, a process-specific
specialization of regions/pathways; Johnsrude et al., 2000), (3) stronger neural connections,
和 (4) enhanced neural efficiency (which may include changes in (1) 到 (3) 同时,
leading to faster and shorter activation; as found in Zhang et al., 2005). 然而, the concept of
neural efficiency is still debated, and how the brain adaptively reallocates its resources is con-
特罗西尔. 此外, learning-specific changes and enhancements are hard to separate from
changes in attentional, cognitive, and memory-related processes (see discussion in Zhang &
王, 2007).
The auditory cortex, which is a major region involved in language processing, shows per-
ceptual narrowing during specific sensitive periods early on (Ortiz-Mantilla et al., 2016).
Auditory learning leads to the formation and strengthening of long-term memory traces, 哪个
in turn positively influence discrimination skills that are the basis of speech perception and
comprehension (Partanen et al., 2013). 因此, infants learn very early how to encode auditory
features in the primary auditory cortex, group speech sounds they perceive into language-
specific phonetic categories, and process musical rhythms and harmonic relationships
(White et al., 2013). The auditory cortex is likely to have an extended period of heightened
developmental plasticity throughout childhood, where changes in cellular organization occur
(Kral & Eggermont, 2007). It is thought that during such sensitive periods (IE。, epochs during
which experiences cause enhanced, long-lasting effects on behaviour and the brain; Penhune,
2011), neural representations are first broadly tuned and subsequently become more refined
and respond preferentially to more frequently encountered features and stimuli in the environ-
蒙特 (Scott et al., 2007). With regard to language learning, 然而, it seems that the neural
systems involved therein are first established for optimal processing of the first language
(frequently encountered input), and have to be adapted for the successful and efficient pro-
cessing of another language (White et al., 2013).
尽管如此, a major question concerns the interaction between experiential and matura-
tional factors that either restrict or facilitate language learning across the lifespan. Regardless of
environmental stimulation and extrinsic motivation, the potential for successful late L2 acqui-
sition is significantly reduced on the neural level due to maturational declines in synaptic den-
城市, decreased levels of brain metabolism (Bates et al., 1992), and increased axon myelination
(Pulvermüller & Schumann, 1994). Concerning explicit learning mechanisms, it might be the
case that purely bottom-up (implicit) learning is not sufficient for later learners to change the
phonetic representations built during L1 learning (Archila-Suerte et al., 2012). On the other
手, goal-oriented explicit training (IE。, progressive adaptation to performance, feedback,
and directed attention to relevant features of the new language) may enhance post-sensitive
period L2 learning (White et al., 2013).
全面的, much more research is needed to confirm or potentially help improve current
models on language learning. A major problem in designing and interpreting studies is to fur-
ther disentangle the influence of cognitive, memory-related, and attention-related processes
Neurobiology of Language
399
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
and their impact on neural resources. 全面的, increases and enhancements in neural speci-
ficity, efficiency, 灵敏度, and connectivity are the basis of learning-induced changes in the
脑.
A NEUROCOGNITIVE MODEL OF LANGUAGE APTITUDE
Based on previous findings, we here present a preliminary neurocognitive model of language
aptitude (见图 1). In our model, language aptitude is treated as an overarching ability
based on neural and cognitive characteristics that are partly biologically determined and un-
fold and develop in interaction with the social environment. The model is an array of ideas
based on previous research (including our own) and comprises two major profiles, namely a
language aptitude profile and a language competence profile (in analogy to Seither-Preisler
Learning-induced neural plasticity:
Changes on the neural level caused
by learning processes (例如, learning a
new language).
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
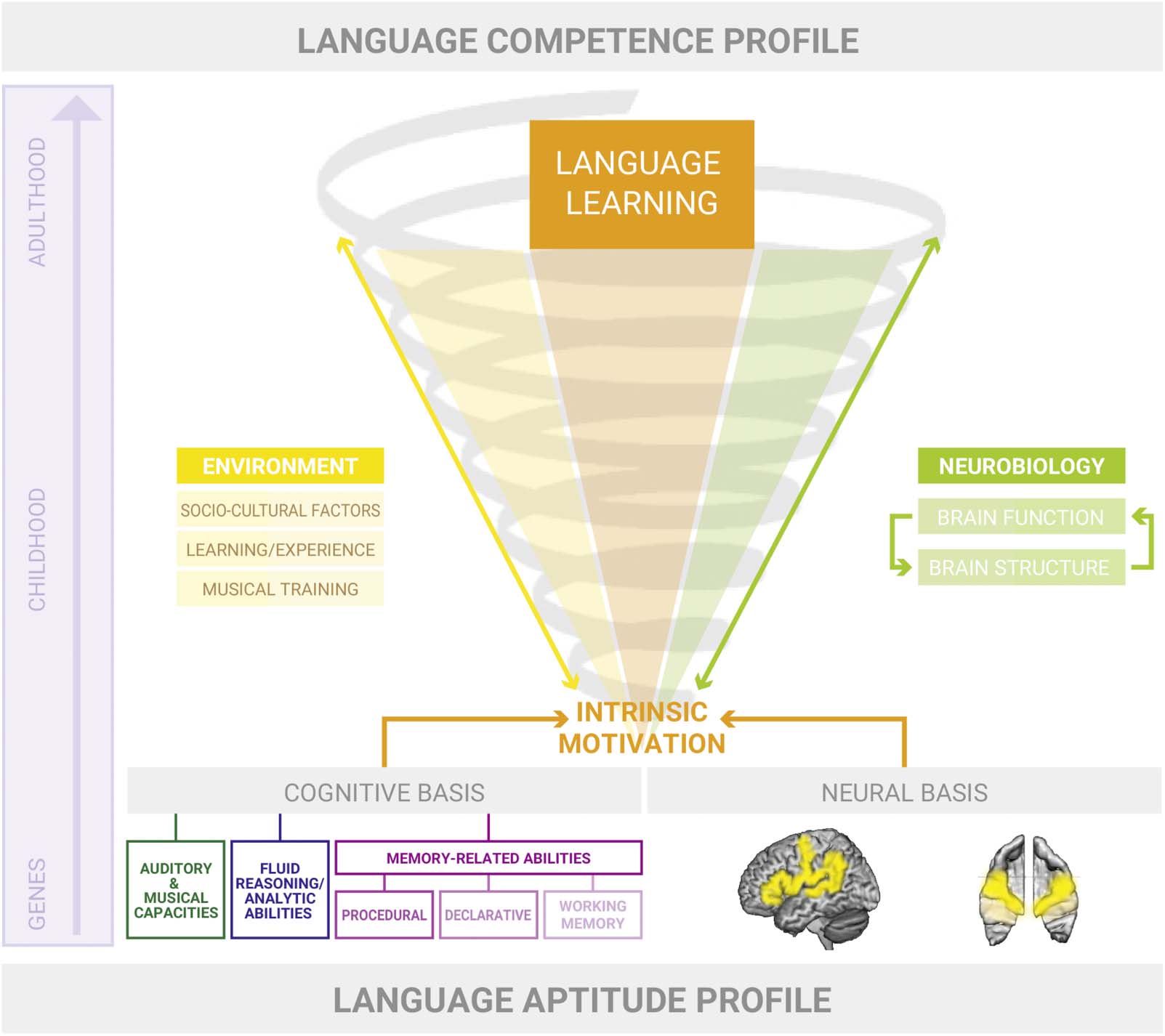
数字 1. A neurocognitive model of language aptitude. Our model proposes that a specific, largely genetically and prenatally predetermined
language aptitude profile progressively develops into a language competence profile. The language aptitude profile comprises a set of cog-
nitive predispositions for language learning and its neural basis, which is visible as anatomical characteristics in the auditory cortex and other
language-related regions (例如, left IPL or IFG). In our model, advantageous neurocognitive predispositions foster intrinsic motivation and thus a
general interest in language learning. Environmental factors (黄色的; left side of the spiral) such as sociocultural factors, learning/experience,
and musical training significantly contribute to the language learning process and interact with it on all levels. During development from early
infancy into adulthood, the brain undergoes major neuroplastic changes that are partly biologically induced (maturational aspect) and partly
learning-induced (biographical aspect). The extent of learning-induced neural plasticity is a function of the predisposed language aptitude
profile (neural characteristics and its cognitive correlates) and supportive environmental factors.
Neurobiology of Language
400
Neurobiology of language aptitude
等人。, 2014). 随着时间的推移, due to biological maturation processes and in interaction with the
环境 (例如, sociocultural factors, 教育, language/learning experience, musical
训练), the initial aptitude profile develops into a manifest competence profile (in accor-
dance with the competence level proposed in models by Gagné, 2004). In our model, 我们
assume that genetic attributes influence how individuals experience and interpret their envi-
罗门特, that is these attributes fundamentally guide the selection, modification, and creation
of experiences in an individual, thus steadily complementing their competence profiles
(Bjorklund & Causey, 2018; Scarr & McCartney, 1983). 像这样, an advantageous innate,
or congenital, aptitude profile leads to a positive attitude towards language learning, 哪个
in turn fosters intrinsic motivation and eagerness to engage in language learning activities
(as proposed in the socioeducational model of second language learning by Gardner,
2010). Previous research has shown that learning the full set of grammatical rules of one’s first
language may take up to 17 年 (Hartshorne et al., 2018), which is why we emphasize the
fluent transition of all learning stages and from first to second and further foreign language
学习. We further suggest that the individual amount of neural plasticity, which determines
the speed and success of language learning, is proportional to language-relevant predisposi-
tions in the aptitude profile, which are determined by genetic, epigenetic, and intrauterine
factors prior to birth. Although there is no doubt that the cellular mechanisms underlying
experience-dependent structural changes in the human brain are crucial as well (for a sum-
mary, see Zatorre et al., 2012), the focus of the present model shall lie on stable morpholo-
gical characteristics of cortical regions.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Cattell-Horn-Carroll (CHC) 模型:
A model of human cognitive
performance and intellectual ability.
Auditory scene analysis:
The ability to group auditory stimuli
relative to background noise in a
meaningful way on the basis of
spectral and temporal acoustic
参数.
The Language Aptitude Profile
The cognitive basis
In our view, the cognitive basis of language aptitude comprises the set of abilities/capacities
that are essential for facilitating later language learning and that an individual possesses either
before prenatal influences come into play (genetically predisposed), or that develop in early
prenatal stages (influenced by genetical predispositions and intrauterine factors). The cognitive
basis of our model is influenced by the Cattell-Horn-Carroll (CHC) model of human cognitive
performance and intellectual ability (卡罗尔, 1993; Horn & Cattell, 1966; J. 瓦. 施耐德 &
McGrew, 2018) and is intricately tied to language aptitude. In our view, it comprises (1) 非盟-
ditory and musical processing, (2) fluid reasoning/analytical abilities, (3) memory-related abil-
实体, 例如 (A) procedural memory/ learning (例如, learning efficiency), (乙) declarative
memory/learning (例如, comprehension-knowledge), 和 (C) 工作记忆.
Auditory processing, as suggested in the CHC model, comprises basic auditory hearing
功能, language-specific abilities (phonetic coding, speech sound discrimination, resis-
tance to auditory stimulus distortion, hearing/speech threshold), and music-specific abilities
(processing of pitch, 音色, musical intervals, harmonies, melodies, 节奏). We therefore
subsume them under auditory and musical processing. These higher language- and music-
relevant auditory functions represent aspects of auditory scene analysis, 那是, the ability to
group auditory stimuli relative to background noise in a meaningful way on the basis of spec-
tral and temporal acoustic parameters (for a definition, see Bregmann, 1990, 2015). Fluid rea-
soning designates the ability to perform mental operations, and comprises induction,
sequential reasoning, and quantitative reasoning (J. 瓦. 施耐德 & McGrew, 2018). 因此,
it is not only vital for intelligence and general cognitive mechanisms, but also for learning a
语言, especially for language analytic abilities. Due to the strong link between fluid rea-
soning and analytical thinking, we have subsumed the two in one category.
Neurobiology of Language
401
Neurobiology of language aptitude
Comprehension-knowledge:
A memory base built for
continuously incorporating and
communicating cultural knowledge,
including language in the CHC
模型.
The first memory-related ability is procedural memory/learning, which includes the concept of
learning efficiency (see also a discussion on procedural memory underlying learning in Ullman,
2015, 2016). According to the CHC model (J. 瓦. 施耐德 & McGrew, 2018), learning efficiency
describes an individual’s capacity to learn, store, and consolidate new information, comprising
primarily associative memory. Conceptually, learning efficiency is strongly tied to fluid reasoning
and is reliant on working memory, both visuo-spatial and auditory working memory (Wen, 2019).
The second ability, namely declarative memory/learning (see Ullman, 2015, 2016 for a distinc-
tion between declarative and procedural memory), includes comprehension-knowledge, 哪个
is defined as a memory base built for continuously incorporating and communicating cultural
知识, including language (associated with an often-neglected aspect of language aptitude,
namely pragmatic language aptitude; Sedaghatgoftar et al., 2019). Less likely to play a role at the
initial stages of language learning (an observation we made in our own studies that was already
proposed in the model by Ullman, 2004, 2015, 2016), declarative memory becomes greatly
important after the onset of first language acquisition. It includes metalevel abilities such as lexical
知识, listening ability, communication ability, and grammatical sensitivity, which have
been considered a part of language aptitude since the early models of Carroll (卡罗尔, 1981;
文等人。, 2017). There has been an ongoing discussion on how to best integrate working
memory in a model of language aptitude, and in the present model, we have decided to include
it as a memory-related ability in the largely predetermined aptitude profile.
The neural basis
In the present model, the starting point of language aptitude is the auditory cortex (HG), 哪个
in previous studies was found to be a neuroanatomical marker for individual differences in
speech sound learning and phonetic experience (Golestani et al., 2002, 2007, 2011), 语言
aptitude (Turker et al., 2017, 2019; see also discussion in Turker & Reiterer, 2021), 和
musical aptitude (Benner et al., 2017; Seither-Preisler et al., 2014). Since interindividual var-
iation of the auditory cortex can even be observed at a prenatal stage, this cortical structure
bears the potential to boost auditory processing for speech and language learning from very
early periods on. 像这样, these differences in morphometry could foster and aid an early and
efficient development of cortical connections between auditory and motor areas (Skeide &
Friederici, 2016). These auditory-motor connections are the basis of an efficient neural anal-
ysis of speech sounds, and thus of language processing (Möttönen et al., 2013). Auditory-
motor connections can lead to strong speech-motor association networks (IE。, strongly and
reciprocally interconnected neural networks; 例如, Pulvermüller & Fadiga, 2010; Watkins &
Paus, 2004) in left superior temporal, temporo-parietal, and frontal areas, also sometimes re-
ferred to as Hebbian learning circuits (Hebb, 1949). It is very likely that in the case of high
language aptitude, these highly specialized functional units develop earlier, faster, 和更多
efficiently through language learning experience.
The neurofunctional efficiency of the auditory cortex seems to be both a function of preex-
isting neuroanatomical traits and of explicit auditory training (Seither-Preisler et al., 2014;
Serrallach et al., 2016), thus revealing the direct interaction of nature and nature on the level
of the auditory cortex. 像这样, we suggest that individual structural variation in left and right
auditory cortices accounts for individual variation in language learning, and thus language
aptitude, which in turn significantly influences all later language learning processes. 在阿迪-
的, we assume that structural variation in other left-hemispheric perisylvian areas (例如, 左边
IPL), less researched in research to date, could be related to language aptitude as well (看
bottom right of Figure 1, where the language areas in the left hemisphere are marked in yellow).
然而, the relative contributions of predisposition and experience in this case need to be
Neurobiology of Language
402
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
further researched, since only limited research has been conducted to date (例如, see Hervais-
Adelman, Moser-Mercer, 等人。, 2017; Stein et al., 2012).
As earlier mentioned, genes influence (社会的) behaviour (G. 乙. Robinson et al., 2008).
最后, genetic propensities potentially guide children to individually select, modify,
and create their experiences (see further evidence in Plomin, 1994). We argue that children
and adolescents with an advantageous language aptitude profile will feel particularly motivated
to learn foreign languages due to the ease with which they acquire them and the progress they
make without substantial effort (卡罗尔, 1981). The above-described facility for learning
languages is likely to result in primarily positive associations with language learning and height-
ened metalinguistic awareness (Jessner, 2008, 2014), which in turn facilitates future language
learning through higher comprehension-knowledge and learning efficiency (IE。, both declarative
and procedural learning would be affected) within this domain. According to our model, congen-
ital abilities and acquired skills (nature-driven and nurture-enhanced processes) constantly inter-
act with one another, 发展, reinforce, and influence one another in a spiral-like fashion or like
moving upwards/downwards in a spiral (which is displayed in Figure 1, where all components of
the spiral, because of their interconnectedness, influence the top and bottom of the spiral contin-
uously and at the same time). If a lot of experience or training falls on enhanced grounds that is
based upon better audio-lingual starter conditions, exceptional skills can be expected. 上
另一方面, untrained or barely trained positive starting conditions will not be enough to develop
high language competence, as in the opposite case of less gifted learners. In the absence or with a
very low level of auditory-language abilities (例如, language impairments), even high levels of
training will not easily compensate and bring forth high competence levels. Only with sufficient
effort and time, individuals with low aptitude can learn and improve their language skills.
Environmental Influences and Gene-Environment Interactions
In our view, the three environmental variables that most significantly influence language learn-
ing are (1) sociocultural and socioeconomic factors, which are particularly relevant during
early language learning in childhood, (2) previous (语言) learning experience (包括
the quality and quantity of language input), 和 (3) musical training. The first includes all
variables that relate to both the social and cultural background and the socioeconomic
environment of an individual (including socioeconomic status, parenting style, parent-child
相互作用, 和教育) and that shape early language learning experiences (Hayiou-
托马斯, 2008; Hoff, 2006). 同时, research has shown that the neural circuits
of language are shaped by previous language learning experience (Pierce et al., 2015;
Sharpe et al., 2021), which supports our suggestion that any form of learning experience either
directly or indirectly guides and influences future learning situations (例如, by exerting a direct
influence on intrinsic motivation. Behaviourally, first language learning difficulties, 阅读
deficits, or potentially negative experiences in foreign language learning classes, coupled with
anxiety, negative feedback, and a loss in motivational intensity, can negatively impact future
foreign language learning (see discussion in Ellis, 2004; Fonseca-Mora & Machancoses, 2016).
像这样, the quality and quantity of the language experience will exert an influence on long-
term second language learning attainment (see also proposal of Moyer, 2008). While we do
not propose a specific model for language learning experience, we believe that language
learning closely matches general learning patterns and mechanisms, which are thought to rely
on attention, the enhancement of expectancies, and autonomy, which largely influence intrin-
sic motivation (for an interesting proposal on motor learning, see Wulf & Lewthwaite, 2016).
A third essential environmental variable, namely musical training, influences language
learning on the neural level, as well as on the cognitive level (Turker & Reiterer, 2021).
Neurobiology of Language
403
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
From a neural perspective, both language and music recruit an array of brain networks involv-
ing motor, auditory, visual, and memory-related mechanisms. Auditory processing at subcor-
tical and cortical levels benefits from musical training, leading to stronger brainstem responses
(例如, to pitch; Kraus & Chandrasekaran, 2010; Moreno & Bidelman, 2014; 磷. C. 中号. 黄,
Skoe, 等人。, 2007) and faster and bilaterally more synchronous cortical auditory evoked re-
响应 (Seither-Preisler et al., 2014; Serrallach et al., 2016). Musicianship has further been
shown to enhance speech perception, linguistic skills, and high-level cognitive processing
(Moreno et al., 2011; Schellenberg & Winner, 2011), leading to positive and long-lasting ben-
efits on auditory functioning. The inextricable link between language and music (Patel, 2012;
Schellenberg & Peretz, 2008) has even led researchers to explore whether precursors of the
two domains have evolved together or from a common signal system (例如, “musilanguage” in
棕色的, 2000). 任何状况之下, it should be expected that both domains are strongly related on a
neuropsychological level, an assumption that has been corroborated in a multitude of studies
(Sammler, 2020).
Preliminary Hypotheses and Predictions
The current model suggests that a specific language aptitude profile may strongly impact the
level of intrinsic motivation. 像这样, an advantageous profile would lead to a heightened
motivational intensity, which in turn fosters the language learning process (加德纳, 2010).
While intrinsic motivation is displayed as the basis of all language learning, we emphasize that
intrinsic motivation is presumably more involved in language learning processes other than first
language learning, which is most likely largely driven by the biological need to communicate
within the social environment. It thus seems meaningful to discuss the particular neurobiological
changes and mechanisms related to the unfolding of language aptitude separate from the
concrete interaction of environmental variables with cognition and language learning.
The neural properties of the linguistically gifted brain could manifest in various ways, 从
(partly) innate morphological differences to thicker myelination around language-related fibre
tracts to higher efficiency during linguistic tasks. Such efficiency could manifest as more focal
activation in tasks requiring convergent thinking, or more widespread activation in tasks re-
quiring divergent thinking. We have formulated some hypotheses regarding brain function and
结构, and their relation to the development of a language aptitude profile into a compe-
tence profile, as displayed in Figure 1.
Neurofunctional underpinnings of language aptitude
As discussed earlier, only a few studies to date have directly related functional brain activation to
language aptitude, and so far, these have yielded mixed results. Studies have reported either more
focal or more widespread neural processing indicative of high language talent and competence
(Hu et al., 2013; Kepinska, de Rover, 等人。, 2017; Reiterer et al., 2011). Although it is not self-
evident how both observations can be true at the same time, they are not mutually exclusive.
第一的, these discrepant results could be consequences of task choice, meaning that artificial gram-
mar learning (grammatical analytical task), being a more complex and cognitive task compared to
speech imitation, could require more widespread activation and it might thus be harder to develop
more efficient processing. 同时, 然而, Kepinska, de Rover, 等人. (2017) 报道
only differences between high and average grammatical ability learners since they found no
differences between high and low ability learners. This might stem from other limitations of that
学习 (sample size, specific task choice, characteristics of the learner groups). Another explanation
for the differences in results could be the learning stage or level (beginner vs. 先进的).
Neurobiology of Language
404
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
We agree with Prat et al. (2019), who suggests that individual differences in language learn-
英, including high language learning ability, manifest in neural efficiency, neural synchroni-
扎化, and neural adaptability. Concerning brain activation, we therefore hypothesize that
brain activation should be more focal in high ability learners compared to low ability learners
due to previous learning experience and expertise in the specific language-related tasks. 这
should hold true for all domains of language learning, from pronunciation and sound learning
to grammatical analytic abilities. While we believe that studying differences in functional
activation within language-specific brain areas certainly provides interesting results as to the
engagement of relevant areas, they can only provide us with a short glimpse into how func-
tioning machinery can look. If there are distinct functional brain networks for speech articu-
关系, sensory language processing, and higher-level language processing, as suggested by
Fedorenko and Thompson-Schill (2014), individual differences related to language aptitude
need to be investigated in these networks. Previous investigations on differences in functional
and effective connectivity, absent in language aptitude research but extensively studied
regarding individual differences in language learning, portray a much clearer picture. In highly
successful language learners, a variety of language- and domain-general areas in the human
brain have been found to work in concert and show greater functional and effective connec-
活力, often in the absence of clear functional activation patterns (例如, Prat et al., 2019). 在
特别的, connectivity of the left IFG and left STG seem to be frequently observed within
language learning tasks. We propose that more efficient global language networks, be it during
rest or during/after extensive foreign language learning, could be indicators of and at the same
time the result of language aptitude. In accordance with our view that the posteromedial HG
and the posterior STG, hosting primary and secondary auditory areas, are crucial for language
aptitude, we see functional connectivity differences at rest or during linguistic processes as
potential indicators that successful language learning is the result of well-myelinated connec-
tions between these auditory structures and inferior frontal areas.
Structural underpinnings of language aptitude
Evidence so far suggests that structural characteristics of the bilateral auditory cortices
(Golestani, 2014; Golestani et al., 2002, 2007, 2011; Turker et al., 2017, 2019) and the left
inferior frontal cortex (Novén et al., 2019) are involved in high language aptitude. 而且,
portions of the left and right arcuate fascicle have been found to relate to language aptitude as
出色地 (Kepinska, Lakke, 等人。, 2017; Vaquero et al., 2017; Xiang et al., 2012). 最后,
we suggest that specific structural characteristics (例如, higher grey matter and higher gyrifica-
的) should be found in individuals with high language learning ability. 而且, we believe
that learning experiences will lead to faster changes in these areas (IE。, individuals with higher
language learning abilities will show faster response to training and possess a heightened neu-
ral plasticity).
Regarding structural connectivity, we would like to elaborate on ideas related to the mat-
uration of the arcuate fascicle and how it could foster language learning. Language accuracy
and processing speed have been shown to depend upon the maturational status of the arcuate
fascicle (Skeide et al., 2016). Fractional anisotropy, and other maturational indicators, 对于前任-
充足, myelination (Mukherjee et al., 2001), axon growth (Paus, 2010), and increasing fibre
density (Scholz et al., 2009), are essential for the refinement of the dorsal syntax network. 这
dorsal pathway from left temporal areas to posterior Broca’s area (left IFG) is important for
higher-order language functions since it is weak in nonhuman primates and weaker in children
than in adults (Friederici, 2009). Learning a new or an artificial grammar is closely tied to
sentence-level syntax, which is primarily driven by the left IFG modulating the posterior
Neurobiology of Language
405
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
STG (Makuuchi & Friederici, 2013), and syntactic information is exchanged dorsally along the
arcuate fascicle (Friederici et al., 2011; Saur et al., 2008). 然而, the two dorsal fibre tracts
that interconnect left temporal and frontal areas in adults do not develop simultaneously in
婴儿. Only the fibre tract to the premotor cortex, thought to be responsible for the integration
of sensory and motor representations during babbling and the development of phonemic rep-
resentations, is already myelinated in infants of 2–5 months of age (Dubois et al., 2010; Perani
等人。, 2011). We thus hypothesize that an earlier maturation of the left arcuate fascicle, 还有
as a stronger left lateralization of specific segments of this fibre tract (具体来说, the left pos-
terior segment) could be a marker for high language aptitude. 更重要的是, higher mye-
lination of that fibre tract could be a direct indicator for faster processing between posterior
STG and IFG, which should result in more efficient communication between these areas and
thus likely reflects high language learning abilities.
Apart from the arcuate fascicle, we hypothesize that the frontal aslant tract (pronunciation
and imitation), the uncinate fasciculus (potentially involved in memory retrieval and learning
efficiency), and the inferior fronto-occipital longitudinal fasciculus (phonetic coding, phono-
logical processes, as well as semantic-related learning) could be key tracts to further investi-
gate with respect to language aptitude.
经验- and learning-dependent plasticity effects
Although grey and white matter volumes in most areas of the brain increase in early child-
兜帽, declines in frontal, 顶叶, and temporal areas both in terms of volume and thickness
have been reported across adolescence (Foulkes & Blakemore, 2018). What should not be
forgotten, 然而, is that these primarily group-level findings may be subject to considerable
interindividual differences, resulting from both the genes and the environment. With regard to
language learning, a plethora of studies have reported differences in neural plasticity (例如,
cortical thickness) related to various forms of language learning, 例如, due to language
learning onset (Vaquero et al., 2020), novel word learning (Shtyrov et al., 2010), or intense
language learning studies (Mårtensson et al., 2012). 迄今为止, very little overlap regarding the
brain regions affected by cortical thinning/thickening has been reported. Leading to more con-
fusion, changes due to learning have been shown in numerous studies (例如, 张等人。,
2009, reporting brain activation changes due to phonetic learning in adulthood) but little is
known about whether these changes pertain and lead to life-long modifications or are just
temporarily built during intensive learning phases.
It is widely accepted that learning, regardless of whether it is of linguistic nature, leads to a
modification of biochemical processes (at the subcortical and cortical level through a modu-
lation of synaptic plasticity; Sharpe et al., 2021). Such changes due to learning occur in all
个人, regardless of their specific capacity for learning and often even irrespective of
the learning period and intensity. Although it seems logical to assume that neuroplasticity is
more a sign of learning than of language aptitude per se, it is likely that neuroplastic changes
happen faster and are more effective in those with high language aptitude. 而且, 研究
suggests that the amount of learning-induced neuroplasticity is directly proportional to biolog-
ically predisposed aptitude (Seither-Preisler et al., 2014; Serrallach et al., 2016), 哪个, 如何-
曾经, requires more research.
CONCLUSIONS AND FUTURE DIRECTIONS
Despite the substantial improvements in research on individual differences and cognitive ca-
pacities, very few studies to date have explicitly tackled the investigation of language aptitude
Neurobiology of Language
406
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
and its relation to neurobiological patterns and mechanisms. Studies with only adults provide a
short and limited glimpse into already established, functioning machinery, and so far, 语言
aptitude and individual differences have only been investigated in spoken languages, 然而
sign languages have been completely left out. 当然, uncovering the neural bases of language
aptitude, be it in the spoken, auditory, or signed domain, requires addressing the development of
language aptitude both through nature and nurture in utero, and in peri- and postnatal stages and
up to childhood and adolescence. The ideas we present in this model are based on spoken lan-
guage mainly and cannot account for signed languages yet.
As mentioned in the abstract, the model presented here is preliminary and shall be refined as
more research on the topic emerges. To confirm the above formulated hypotheses, further be-
havioural and neurobiological longitudinal and molecular genetic studies are desperately
需要的, especially with young children and adolescents. Other avenues for future research
include investigating relative contributions of experience-dependent plasticity, especially in
contrast to potentially preexisting indicators for learning and plasticity (例如, domain-specific
aptitude; Golestani, 2014; Golestani et al., 2011). 此外, we suggest that future func-
tional neuroimaging studies should investigate the involvement of functionally specialized and
domain-general language-related networks to explore language aptitude on a network level
(Fedorenko & Thompson-Schill, 2014). 此外, even if the arcuate fascicle is one of the
most prominent fibre tracts for higher-level language processing and cognition, we propose to
further investigate other fibre tracts in relation to specific language subskills, 例如, 这
frontal aslant tract for speech imitation and pronunciation proficiency, the uncinate fasciculus
for semantic-related memory retrieval (例如, necessary in vocabulary learning), or the inferior
fronto-occipital fasciculus for specific phonetic and phonology-related skills. In the future it
might also be worthwhile to incorporate intergenerational genetic transmission patterns and
their variation in men and women, 例如, as has been shown for corticolimbic circuity
(Yamagata et al., 2016), which would add an additional perspective to the present model.
资金信息
Sabrina Turker, Alexander von Humboldt-Stiftung (https://dx.doi.org/10.13039/100005156),
奖项ID: Postdoc Fellowship.
作者贡献
Sabrina Turker: Writing – original draft: 平等的; 写作——复习 & 编辑: 平等的. Annemarie
Seither-Preisler: Writing – original draft: 平等的; 写作——复习 & 编辑: 平等的. Susanne
Maria Reiterer: Writing – original draft: 平等的; 写作——复习 & 编辑: 平等的.
参考
Abrahamsson, N。, & Hyltenstam, K. (2008). The robustness of apti-
tude effects in near-native second language acquisition. 学习
in Second Language Acquisition, 30(4), 481–509. https://doi.org
/10.1017/S027226310808073X
Abrahamsson, N。, & Hyltenstam, K. (2009). Erratum: Age of onset
and nativelikeness in a second language: Listener perception ver-
sus linguistic scrutiny (Language Learning: 59:2 (249–306)).
Language Learning, 59(4), 928. https://doi.org/10.1111/j.1467
-9922.2009.00530.X
Antoniou, M。, Ettlinger, M。, & 黄, 磷. C. 中号. (2016). Complexity,
training paradigm design, and the contribution of memory
subsystems to grammar learning. PLOS ONE, 11(7), 1–20. https://
doi.org/10.1371/journal.pone.0158812, 考研: 27391085
Antoniou, M。, & 黄, 磷. C. 中号. (2015). Poor phonetic perceivers
are affected by cognitive load when resolving talker variability. 这
Journal of the Acoustical Society of America, 138(2), 571–574.
https://doi.org/10.1121/1.4923362, 考研: 26328675
Archila-Suerte, P。, Zevin, J。, Bunta, F。, & Hernandez, A. 乙. (2012). 年龄
of acquisition and proficiency in a second language independently
influence the perception of non-native speech. Bilingualism, 15(1),
190–201. https://doi.org/10.1017/S1366728911000125, 考研:
30197550
Neurobiology of Language
407
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
Assaneo, 中号. F。, Ripollés, P。, Orpella, J。, 林, 瓦. M。, de Diego-
Balaguer, R。, & Poeppel, D. (2019). Spontaneous synchroniza-
tion to speech reveals neural mechanisms facilitating language
学习. 自然神经科学, 22(4), 627–632. https://doi.org
/10.1038/s41593-019-0353-z, 考研: 30833700
奥尔巴赫, S. (1906). Beitrag zur Lokalisation des musikalischen
Talentes im Gehirn und am Schädel [Contribution to the locali-
zation of musical talent in the brain and on the skull]. Archives of
Anatomy and Physiology, 197–230.
Barbeau, 乙. B., Chai, X. J。, 陈, J. K., Soles, J。, Berken, J。, Baum, S。,
Watkins, K. E., & 克莱因, D. (2017). The role of the left inferior
parietal lobule in second language learning: An intensive lan-
guage training fMRI study. Neuropsychologia, 98, 169–176.
https://doi.org/10.1016/j.neuropsychologia.2016.10.003,
考研: 27725166
Bartley, A. J。, 琼斯, D. W., & 温伯格, D. 右. (1997). Genetic
variability of human brain size and cortical gyral patterns.
Brain, 120(2), 257–269. https://doi.org/10.1093/ brain/120.2
.257, 考研: 9117373
Bates, E., Thal, D ., 芬莱, B., Clancy, B., Origins, 磷. D ., & Int,
氮. S. F. (1992). Early language development and its neural cor-
relates. In I. Rapin & S. Segalowitz (编辑。), Handbook of
Neuropsychology (2nd 版。, PP. 69–110). 爱思唯尔.
Benner, J。, Wengenroth, M。, Reinhardt, J。, Stippich, C。, 施耐德,
P。, & Blatow, 中号. (2017). Prevalence and function of Heschl’s
gyrus morphotypes in musicians. Brain Structure and Function,
222(8), 3587–3603. https://doi.org/10.1007/s00429-017-1419-x,
考研: 28397108
Bernard, C。, Droulle, P。, Didier, F。, Gérard, H。, Larroche, J. C。,
Plenat, F。, Bomsel, F。, Roland, J。, & Hoeffel, J. C. (1988).
Echographic aspects of cerebral sulci in the ante- and perinatal
时期. Journal de Radiologie, 69(8–9), 521–532. 考研:
3057186
Biedron(西德:1), A. (2015). Neurology of foreign language aptitude. 学习
in Second Language Learning and Teaching, 5(1), 13–40. https://
doi.org/10.14746/ssllt.2015.5.1.2
Birdsong, D ., & Molis, 中号. (2001). On the evidence for maturational
constraints in second-language acquisition. Journal of Memory
and Language, 44(2), 235–249. https://doi.org/10.1006/jmla
.2000.2750
Bjorklund, D. F。, & Causey, K. 乙. (2018). Children’s thinking:
Cognitive development and individual differences (6第三版。).
SAGE Publications.
鲍尔斯, A. R。, 张, C. B., & Karuzis, V. 磷. (2016). Pitch ability as
an aptitude for tone learning. Language Learning, 66(4), 774–808.
https://doi.org/10.1111/lang.12159
Bregmann, A. S. (1990). Auditory scene analysis: The perceptual
organization of sound. 与新闻界. https://doi.org/10.7551
/mitpress/1486.001.0001
Bregmann, A. S. (2015). Progress in understanding auditory scene
分析. Music Perception, 33(1), 12–19. https://doi.org/10.1525
/mp.2015.33.1.12
Breitenstein, C。, Jansen, A。, Deppe, M。, Foerster, A. F。, 索默, J。,
Wolbers, T。, & Knecht, S. (2005). Hippocampus activity differen-
tiates good from poor learners of a novel lexicon. 神经影像,
25(3), 958–968. https://doi.org/10.1016/j.neuroimage.2004.12
.019, 考研: 15808996
棕色的, S. (2000). The “musilanguage” model of language evolu-
的. 在S. 棕色的, 乙. Merker, & 氮. L. Wallin (编辑。), The origins
of music (PP. 271–300). 与新闻界. https://doi.org/10.7551
/mitpress/5190.003.0022
Buffington, J。, & Morgan-Short, K. (2019). Declarative and proce-
dural memory as individual differences in second language
aptitude. In Z. Wen, 磷. Skehan, A. Biedron(西德:1), S. 李, & 右. L.
Sparks (编辑。), Language aptitude: Advancing theory, testing,
research and practice (PP. 215–237). 劳特利奇. https://doi.org
/10.4324/9781315122021-11
Cai, D. C。, Fonteijn, H。, Guadalupe, T。, Zwiers, M。, Wittfeld, K.,
Teumer, A。, Hoogman, M。, Arias-Vásquez, A。, 哪个, Y。,
Buitelaar, J。, Fernández, G。, Brunner, H. G。, van Bokhoven, H。,
弗兰克, B., Hegenscheid, K., Homuth, G。, Fisher, S. E., Grabe,
H. J。, Francks, C。, & Hagoort, 磷. (2014). A genome-wide search
for quantitative trait loci affecting the cortical surface area and
thickness of Heschl’s gyrus. Genes, Brain and Behavior, 13(7),
675–685. https://doi.org/10.1111/gbb.12157, 考研:
25130324
坎贝尔, K. L。, & Tyler, L. K. (2018). Language-related domain-
specific and domain-general systems in the human brain.
Current Opinion in Behavioral Sciences, 21, 132–137. https://
doi.org/10.1016/j.cobeha.2018.04.008, 考研: 30057936
Carmelli, D ., Swan, G. E., DeCarli, C。, & 芦苇, 时间. (2002).
Quantitative genetic modeling of regional brain volumes and
cognitive performance in older male twins. Biological
心理学, 61(1–2), 139–155. https://doi.org/10.1016/S0301
-0511(02)00056-X, 考研: 12385673
卡罗尔, J. 乙. (1981). Twenty-five years of research on foreign lan-
guage aptitude. In K. C. Diller (埃德。), Individual differences and
universals in language learning aptitude (PP. 83–118). Newbury
房子.
卡罗尔, J. 乙. (1993). Human cognitive abilities: A survey of factor-
analytic studies. 剑桥大学出版社. https://doi.org/10
.1017/CBO9780511571312
Catani, M。, & Bambini, V. (2014). A model for social communica-
tion and language evolution and development (SCALED).
神经生物学的当前观点, 28, 165–171. https://doi.org
/10.1016/j.conb.2014.07.018, 考研: 25156623
Catani, M。, 琼斯, D. K., & Ffytche, D. H. (2005). Perisylvian lan-
guage networks of the human brain. Annals of Neurology, 57(1),
8–16. https://doi.org/10.1002/ana.20319, 考研: 15597383
Chai, X. J。, Berken, J. A。, Barbeau, 乙. B., Soles, J。, Callahan, M。,
陈, J. K., & 克莱因, D. (2016). Intrinsic functional connectivity
in the adult brain and success in second-language learning.
神经科学杂志, 36(3), 755–761. https://doi.org/10
.1523/JNEUROSCI.2234-15.2016, 考研: 26791206
Chandrasekaran, B., Kraus, N。, & 黄, 磷. C. 中号. (2012). 人类
inferior colliculus activity relates to individual differences in spo-
ken language learning. 神经生理学杂志, 107(5),
1325–1336. https://doi.org/10.1152/jn.00923.2011, 考研:
22131377
陈, C. H。, Fiecas, M。, Gutiérrez, 乙. D ., Panizzon, 中号. S。, Eyler,
L. T。, Vuoksimaa, E., 汤普森, 瓦. K., Fennema-Notestine, C。,
Hagler, D. J。, Jernigan, 时间. L。, Neale, 中号. C。, Franz, C. E., Lyons,
中号. J。, 菲舍尔, B., Tsuang, 中号. T。, 戴尔, A. M。, & Kremen, 瓦. S.
(2013). Genetic topography of brain morphology. 会议记录
of the National Academy of Sciences of the United States of
美国, 110(42), 17089–17094. https://doi.org/10.1073/pnas
.1308091110, 考研: 24082094
Chow, 乙. 瓦. Y。, & 黄, S. 瓦. (2021). What does genetic re-
search tell us about the origins of language and literacy develop-
蒙特? A reflection on Verhoef et al. (2020). Journal of Child
Psychology and Psychiatry, 62(6), 739–741. https://doi.org/10
.1111/jcpp.13399, 考研: 33631837
Christiner, M。, & Reiterer, S. 中号. (2013). Song and speech:
Examining the link between singing talent and speech imitation
能力. 心理学前沿, 4, 文章 874. https://doi.org/10
.3389/fpsyg.2013.00874, 考研: 24319438
Neurobiology of Language
408
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
Christiner, M。, & Reiterer, S. 中号. (2018). Early influence of musical
abilities and working memory on speech imitation abilities:
Study with pre-school children. Brain Sciences, 8(9), 169.
https://doi.org/10.3390/brainsci8090169, 考研: 30200479
Coventry, W., Antón-Méndez, 我。, Ellis, 乙. M。, Levisen, C。, Byrne, B.,
van Daal, V. H. P。, & Ellis, 氮. C. (2012). The etiology of individual
differences in second language acquisition in Australian school
学生: A behavior-genetic study. Language Learning, 62(3),
880–901. https://doi.org/10.1111/j.1467-9922.2012.00718.x
考恩, 氮. (2014). Working memory underpins cognitive develop-
蒙特, 学习, 和教育. Educational Psychology Review,
26(2), 197–223. https://doi.org/10.1007/s10648-013-9246-y,
考研: 25346585
da Costa, S。, van der Zwaag, W., Marques, J. P。, Frackowiak, 右. S. J。,
克拉克, S。, & Saenz, 中号. (2011). Human primary auditory cortex
follows the shape of Heschl’s Gyrus. 神经科学杂志,
31(40), 14067–14075. https://doi.org/10.1523/ JNEUROSCI
.2000-11.2011, 考研: 21976491
戴尔, 磷. S。, Harlaar, N。, 霍沃斯, C. 中号. A。, & Plomin, 右. (2010).
Two by two: A twin study of second-language acquisition.
心理科学, 21(5), 635–640. https://doi.org/10.1177
/0956797610368060, 考研: 20483839
戴尔, 磷. S。, Harlaar, N。, & Plomin, 右. (2012). Nature and nurture in
school-based second language achievement. Language Learning,
62(s2), 28–48. https://doi.org/10.1111/j.1467-9922.2012.00705.x
Dehaene-Lambertz, G。, Hertz-Pannier, L。, & Dubois, J. (2006).
Nature and nurture in language acquisition: Anatomical and func-
tional brain-imaging studies in infants. Trends in Neurosciences,
29(7), 367–373. https://doi.org/10.1016/j.tins.2006.05.011,
考研: 16815562
DeKeyser, 右. (2019). The future of language aptitude. In Z. Wen, 磷.
Skehan, A. Biedron(西德:1), S. 李, & 右. L. Sparks (编辑。), Language aptitude:
Advancing theory, testing, research and practice (PP. 317–329).
劳特利奇. https://doi.org/10.4324/9781315122021-16
Della Rosa, 磷. A。, Videsott, G。, Borsa, 中号. 五、, Canini, M。, Weekes,
乙. S。, Franceschini, R。, & Abutalebi, J. (2013). A neural interactive
location for multilingual talent. Cortex, 49(2), 605–608. https://土井
.org/10.1016/j.cortex.2012.12.001, 考研: 23294573
Delogu, F。, Lampis, G。, & Olivetti Belardinelli, 中号. (2008). From melody
to lexical tone: Musical ability enhances specific aspects of foreign
language perception. European Journal of Cognitive Psychology,
22(1), 46–61. https://doi.org/10.1080/09541440802708136
Demetriou, A。, Spanoudis, G。, Shayer, M。, van der Ven, S。,
Brydges, C. R。, Kroesbergen, E., Podjarny, G。, & 斯旺森,
H. L. (2014). Relations between speed, 工作记忆, 和
intelligence from preschool to adulthood: Structural equation
modeling of 14 学习. 智力, 46(1), 107–121. https://土井
.org/10.1016/j.intell.2014.05.013
Deng, Z。, Chandrasekaran, B., 王, S。, & 黄, 磷. C. 中号. (2016).
Resting-state low-frequency fluctuations reflect individual differ-
ences in spoken language learning. Cortex, 76, 63–78. https://土井
.org/10.1016/j.cortex.2015.11.020, 考研: 26866283
迪克, A. S。, & Tremblay, 磷. (2012). Beyond the arcuate fasciculus:
Consensus and controversy in the connectional anatomy of lan-
规格. Brain, 135(12), 3529–3550. https://doi.org/10.1093/brain
/aws222, 考研: 23107648
Dörnyei, Z. (2014). The psychology of the language learner:
Individual differences in second language acquisition. 劳特利奇.
https://doi.org/10.4324/9781410613349
Dörnyei, Z。, & Skehan, 磷. (2003). Individual differences in second
language learning. 在C中. J. Doughty & 中号. H. 长的 (编辑。), 这
handbook of second language acquisition (PP. 589–630).
布莱克威尔. https://doi.org/10.1002/9780470756492.ch18
Doughty, C. J. (2019). Cognitive language aptitude. 语言
学习, 69(s1), 101–126. https://doi.org/10.1111/lang.12322
Draganova, R。, Eswaran, H。, 墨菲, P。, Huotilaiinen, M。, 洛厄里,
C。, & Preissl, H. (2005). Sound frequency change detection in
fetuses and newborns, a magnetoencephalographic study.
神经影像, 28(2), 354–361. https://doi.org/10.1016/j
.neuroimage.2005.06.011, 考研: 16023867
Dubois, J。, Benders, M。, Lazeyras, F。, Borradori-Tolsa, C。, Leuchter,
右. H. 五、, 曼金, J. F。, & Hüppi, 磷. S. (2010). Structural asymme-
tries of perisylvian regions in the preterm newborn. 神经影像,
52(1), 32–42. https://doi.org/10.1016/j.neuroimage.2010.03
.054, 考研: 20362679
Ellis, 右. (2004). Individual differences in second language learning.
在一个. 戴维斯 & C. Elder (编辑。), The handbook of applied linguis-
抽动症 (卷. 17, p. 866). 布莱克威尔. https://doi.org/10.1002
/9780470757000.ch21
Ettlinger, M。, Bradlow, A. R。, & 黄, 磷. C. 中号. (2014). Variability
in the learning of complex morphophonology. Applied
Psycholinguistics, 35(4), 807–831. https://doi.org/10.1017
/S0142716412000586
Faretta-Stutenberg, M。, & Morgan-Short, K. (2018). The interplay of
individual differences and context of learning in behavioral and
neurocognitive second language development. 第二
Language Research, 34(1), 67–101. https://doi.org/10.1177
/0267658316684903
Fedorenko, E., & Thompson-Schill, S. L. (2014). Reworking the lan-
guage network. 认知科学的趋势, 18(3), 120–126.
https://doi.org/10.1016/j.tics.2013.12.006
Fisher, S. 乙. (2017). Evolution of language: Lessons from the ge-
nome. Psychonomic Bulletin and Review, 24(1), 34–40. https://
doi.org/10.3758/s13423-016-1112-8, 考研: 27432000
Flöel, A。, de Vries, 中号. H。, Scholz, J。, Breitenstein, C。, & Johansen-
伯格, H. (2009). White matter integrity in the vicinity of Broca’s
area predicts grammar learning success. 神经影像, 47(4),
1974–1981. https://doi.org/10.1016/j.neuroimage.2009.05.046,
考研: 19477281
Fonseca-Mora, 中号. C。, & Machancoses, F. H. (2016). Music and
language learning: Emotions and engaging memory pathways. 在
磷. D. MacIntyre, 时间. Gregersen, & S. 美世 (编辑。), Positive psy-
chology in SLA (PP. 359–373). Multilingual Matters. https://土井
.org/10.21832/9781783095360-017
Foulkes, L。, & Blakemore, S. J. (2018). Studying individual differ-
ences in human adolescent brain development. 自然
神经科学, 21(3), 315–323. https://doi.org/10.1038/s41593
-018-0078-4, 考研: 29403031
Friederici, A. D. (2009). Pathways to language: Fiber tracts in the
人脑. 认知科学的趋势, 13(4), 175–181.
https://doi.org/10.1016/j.tics.2009.01.001, 考研: 19223226
Friederici, A. D ., Brauer, J。, & Lohmann, G. (2011). Maturation of
the language network: From inter- to intrahemispheric connectiv-
实体. PLOS ONE, 6(6), 1–7. https://doi.org/10.1371/journal.pone
.0020726, 考研: 21695183
Gagné, F. (2004). Transforming gifts into talents: The DMGT as a
developmental theory. High Ability Studies, 15(2), 119–147.
https://doi.org/10.1080/1359813042000314682
加德纳, 右. C. (2010). Motivation and second language acquisi-
的: The socio-educational model. Peter Lang.
Gervain, J. (2015). Plasticity in early language acquisition: 这
effects of prenatal and early childhood experience. 当前的
Opinion in Neurobiology, 35, 13–20. https://doi.org/10.1016/j
.conb.2015.05.004, 考研: 26093365
Gervain, J。, & Mehler, J. (2010). Speech perception and language
acquisition in the first year of life. 心理学年度评论,
Neurobiology of Language
409
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
61, 191–218. https://doi.org/10.1146/annurev.psych.093008
.100408, 考研: 19575623
Geschwind, N。, & Levitsky, 瓦. (1968). Human brain: Left-right
asymmetries in temporal speech region. 科学, 161(3837),
186–187. https://doi.org/10.1126/science.161.3837.186,
考研: 5657070
Glasel, H。, Leroy, F。, Dubois, J。, Hertz-Pannier, L。, 曼金, J. F。, &
Dehaene-Lambertz, G. (2011). A robust cerebral asymmetry in
the infant brain: The rightward superior temporal sulcus.
神经影像, 58(3), 716–723. https://doi.org/10.1016/j
.neuroimage.2011.06.016, 考研: 21723397
Glasser, 中号. F。, & Rilling, J. K. (2008). DTI tractography of the
human brain’s language pathways. 大脑皮层, 18(11),
2471–2482. https://doi.org/10.1093/cercor/ bhn011, 考研:
18281301
Golestani, 氮. (2014). Brain structural correlates of individual differ-
ences at low- to high-levels of the language processing hierarchy:
A review of new approaches to imaging research. 国际的
Journal of Bilingualism, 18(1), 6–34. https://doi.org/10.1177
/1367006912456585
Golestani, N。, Molko, N。, 德阿内, S。, LeBihan, D ., & Pallier, C.
(2007). Brain structure predicts the learning of foreign speech
声音. 大脑皮层, 17(3), 575–582. https://doi.org/10
.1093/cercor/bhk001, 考研: 16603709
Golestani, N。, Paus, T。, & Zatorre, 右. J. (2002). Anatomical corre-
lates of learning novel speech sounds. 神经元, 35(5), 997–1010.
https://doi.org/10.1016/S0896-6273(02)00862-0, 考研:
12372292
Golestani, N。, Price, C. J。, & 斯科特, S. K. (2011). Born with an ear for
dialects? Structural plasticity in the expert phonetician brain.
神经科学杂志, 31(11), 4213–4220. https://doi.org/10
.1523/JNEUROSCI.3891-10.2011, 考研: 21411662
总的, 中号. U. 中号. (1999). Small poppies: Highly gifted children in the
early years. Roeper Review, 21, 207–214. https://doi.org/10
.1080/02783199909553963
Hagoort, 磷. (2014). Nodes and networks in the neural architecture
对于语言: Broca’s region and beyond. Current Opinion in
Neurobiology, 28, 136–141. https://doi.org/10.1016/j.conb
.2014.07.013, 考研: 25062474
Hamrick, P。, Lum, J. A。, & Ullman, 中号. 时间. (2018). Child first lan-
guage and adult second language are both tied to general-
purpose learning systems. 国家会议录
Academy of Sciences, 115(7), 1487–1492. https://doi.org/10
.1073/pnas.1713975115, 考研: 29378936
Hartshorne, J. K., Tenenbaum, J. B., & Pinker, S. (2018). 一个批评的
period for second language acquisition: 证据来自 2/3 米尔-
lion English speakers. 认识, 177, 263–277. https://doi.org
/10.1016/j.cognition.2018.04.007, 考研: 29729947
霍沃斯, C. 中号. A。, 赖特, 中号. J。, Luciano, M。, 马丁, 氮. G。, 的
Geus, 乙. J. C。, van Beijsterveldt, C. 乙. M。, Bartels, M。, Posthuma,
D ., Boomsma, D. 我。, 戴维斯, 氧. S. P。, Kovas, Y。, Corley, 右. P。,
Defries, J. C。, Hewitt, J. K., 奥尔森, 右. K., Rhea, S. A。, 沃兹沃斯,
S. J。, Iacono, 瓦. G。, McGue, M。, … Plomin, 右. (2010). 这
heritability of general cognitive ability increases linearly from
childhood to young adulthood. Molecular Psychiatry, 15(11),
1112–1120. https://doi.org/10.1038/mp.2009.55, 考研:
19488046
Hayiou-Thomas, 中号. 乙. (2008). Genetic and environmental influ-
ences on early speech, language and literacy development.
Journal of Communication Disorders, 41(5), 397–408. https://
doi.org/10.1016/j.jcomdis.2008.03.002, 考研: 18538338
Hebb, D. 氧. (1949). The organization of behavior: A neuropsycho-
logical theory. John Wiley and Sons Inc.
Hepper, 磷. G。, & Shahidullah, 乙. S. (1994). Development of fetal
hearing. Archives of Disease in Childhood, 71(2), 81–87.
https://doi.org/10.1136/fn.71.2.f81, 考研: 7979483
Hervais-Adelman, A。, Egorova, N。, & Golestani, 氮. (2017). 超过
bilingualism: Multilingual experience correlates with caudate
体积. BioRxiv, 1–13. https://doi.org/10.1101/209619
Hervais-Adelman, A。, Moser-Mercer, B., 穆雷, 中号. M。, &
Golestani, 氮. (2017). Cortical thickness increases after simulta-
neous interpretation training. Neuropsychologia, 98, 212–219.
https://doi.org/10.1016/j.neuropsychologia.2017.01.008,
考研: 28077311
Heschl, 右. L. (1878). Über die vordere quere Schläfenwindung des
menschlichen Grosshirns [About the anterior transverse temporal
convolutions of the human cerebrum]. Braumüller.
Hickok, G。, & Poeppel, D. (2004). Dorsal and ventral streams: A
framework for understanding aspects of the functional anatomy
语言的. 认识, 92(1–2), 67–99. https://doi.org/10.1016/j
.cognition.2003.10.011, 考研: 15037127
Hoff, 乙. (2006). How social contexts support and shape language
发展. Developmental Review, 26(1), 55–88. https://土井
.org/10.1016/j.dr.2005.11.002
Homae, F。, Watanabe, H。, Nakano, T。, Asakawa, K., & Taga, G.
(2006). The right hemisphere of sleeping infant perceives senten-
tial prosody. Neuroscience Research, 54(4), 276–280. https://土井
.org/10.1016/j.neures.2005.12.006, 考研: 16427714
Horn, J. L。, & Cattell, 右. 乙. (1966). Refinement and test of the theory
of fluid and crystallized general intelligences. 杂志
Educational Psychology, 57(5), 253–270. https://doi.org/10
.1037/h0023816, 考研: 5918295
胡, X。, Ackermann, H。, 马丁, J. A。, Erb, M。, Winkler, S。, &
Reiterer, S. 中号. (2013). Language aptitude for pronunciation in
advanced second language (L2) learners: Behavioural predictors
and neural substrates. Brain and Language, 127(3), 366–376.
https://doi.org/10.1016/j.bandl.2012.11.006, 考研:
23273501
Huotilainen, M。, Kujala, A。, Hotakainen, M。, 帕克科宁, L。, Taulu,
S。, Simola, J。, Nenonen, J。, Karjalainen, M。, & Näätänen, 右.
(2005). Short-term memory functions of the human fetus recorded
脑磁图检查. NeuroReport, 16(1), 81–84.
https://doi.org/10.1097/00001756-200501190-00019, 考研:
15618896
我是, K., & 授予, 磷. 乙. (2019). Sulcal pits and patterns in developing
human brains. 神经影像, 185, 881–890. https://doi.org/10
.1016/j.neuroimage.2018.03.057, 考研: 29601953
Järvelä, 我. (2018). Genomics studies on musical aptitude, music percep-
的, and practice. Annals of the New York Academy of Sciences,
1423(1), 82–91. https://doi.org/10.1111/nyas.13620, 考研:
29570792
Jessner, U. (2008). A DST model of multilingualism and the role
of metalinguistic awareness. Modern Language Journal, 92(2),
270–283. https://doi.org/10.1111/j.1540-4781.2008.00718.x
Jessner, U. (2014). On multilingual awareness or why the multilin-
gual learner is a specific language learner. 在米. Pawlak & L.
Aronin (编辑。), Essential topics in applied linguistics and multilin-
gualism: Studies in honor of David Singleton (PP. 175–184).
Springer International Publishing. https://doi.org/10.1007/978-3
-319-01414-2_10
Johnsrude, 我. S。, Penhune, V. B., & Zatorre, 右. J. (2000). Functional
specificity in the right human auditory cortex for perceiving pitch
方向. Brain, 123(1), 155–163. https://doi.org/10.1093/brain
/123.1.155, 考研: 10611129
Jouravlev, 奥。, Mineroff, Z。, 空白的, 我. A。, & Fedorenko, 乙. (2021).
The small and efficient language network of polyglots and
Neurobiology of Language
410
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
hyper-polyglots. 大脑皮层, 31(1), 62–76. https://doi.org/10
.1093/cercor/bhaa205, 考研: 32820332
Karma, K. (1994). Auditory and visual temporal structuring: 如何
important is sound to musical thinking. Psychology of Music,
22, 20–30. https://doi.org/10.1177/0305735694221002
Karuza, 乙. A。, Newport, 乙. L。, Aslin, 右. N。, Starling, S. J。, Tivarus,
中号. E., & Bavelier, D. (2013). The neural correlates of statistical
learning in a word segmentation task: An fMRI study. Brain and
语言, 127(1), 46–54. https://doi.org/10.1016/j.bandl.2012
.11.007, 考研: 23312790
Kepinska, 奥。, de Rover, M。, Caspers, J。, & Schiller, 氮. 氧. (2017).
On neural correlates of individual differences in novel grammar
学习: An fMRI study. Neuropsychologia, 98, 156–168. https://
doi.org/10.1016/j.neuropsychologia.2016.06.014, 考研:
27305834
Kepinska, 奥。, Lakke, 乙. A. J. F。, Dutton, 乙. M。, Caspers, J。, &
Schiller, 氮. 氧. (2017). The perisylvian language network and
language analytical abilities. Neurobiology of Learning and
记忆, 144, 96–101. https://doi.org/10.1016/j.nlm.2017.07
.003, 考研: 28710000
Kepinska, 奥。, Pereda, E., Caspers, J。, & Schiller, 氮. 氧. (2017).
Neural oscillatory mechanisms during novel grammar learning
underlying language analytical abilities. Brain and Language,
175, 99–110. https://doi.org/10.1016/j.bandl.2017.10.003,
考研: 29059544
Kidd, E., Donnelly, S。, & Christiansen, 中号. H. (2018). Individual dif-
ferences in language acquisition and processing. 趋势
认知科学, 22(2), 154–169. https://doi.org/10.1016/j
.tics.2017.11.006, 考研: 29277256
Kral, A。, & Eggermont, J. J. (2007). What’s to lose and what’s to
学习: Development under auditory deprivation, cochlear im-
plants and limits of cortical plasticity. Brain Research Reviews,
56(1), 259–269. https://doi.org/10.1016/j.brainresrev.2007.07
.021, 考研: 17950463
Kraus, N。, & Chandrasekaran, 乙. (2010). Music training for the
development of auditory skills. 自然评论神经科学,
11(8), 599–605. https://doi.org/10.1038/nrn2882, 考研:
20648064
Krishnan, S。, Watkins, K. E., & Bishop, D. V. 中号. (2016).
Neurobiological basis of language learning difficulties. Trends
in Cognitive Sciences, 20(9), 701–714. https://doi.org/10.1016/j
.tics.2016.06.012, 考研: 27422443
Legault, J。, Fang, S.-Y。, 兰, Y.-J., & 李, 磷. (2019). Structural brain
changes as a function of second language vocabulary training:
Effects of learning context. Brain and Cognition, 134, 90–102.
https://doi.org/10.1016/j.bandc.2018.09.004, 考研:
30429056
Legault, J。, 授予, A。, Fang, S.-Y。, & 李, 磷. (2019). A longitudinal
investigation of structural brain changes during second language
学习. Brain and Language, 197, 文章 104661. https://土井
.org/10.1016/j.bandl.2019.104661, 考研: 31376630
李, P。, & 授予, A. (2016). Second language learning success re-
vealed by brain networks. Bilingualism: 语言 & 认识,
19(4), 657–664. https://doi.org/10.1017/S1366728915000280
López Ramón y Cajal, C. (2019). Antenatal study of the Heschl’s
gyrus: The first step to understanding prenatal learning.
Medical Hypotheses, 130, 文章 109290. https://doi.org/10
.1016/j.mehy.2019.109290, 考研: 31383347
Mahmoudzadeh, M。, Dehaene-Lambertz, G。, Fournier, M。,
Kongolo, G。, Goudjil, S。, Dubois, J。, Grebe, R。, & Wallois, F.
(2013). Syllabic discrimination in premature human infants prior
to complete formation of cortical layers. 诉讼程序
美国国家科学院,
110(12), 4846–4851. https://doi.org/10.1073/pnas.1212220110,
考研: 23440196
Makuuchi, M。, & Friederici, A. D. (2013). Hierarchical functional
connectivity between the core language system and the working
memory system. Cortex, 49(9), 2416–2423. https://doi.org/10
.1016/j.cortex.2013.01.007, 考研: 23480847
Mamiya, 磷. C。, Richards, 时间. L。, 科伊, 乙. P。, Eichler, 乙. E., & Kuhl,
磷. K. (2016). Brain white matter structure and COMT gene are
linked to second-language learning in adults. 会议记录
the National Academy of Sciences of the United States of
美国, 113(26), 7249–7254. https://doi.org/10.1073/pnas
.1606602113, 考研: 27298360
Marie, D ., Maingault, S。, Crivello, F。, Mazoyer, B., & Tzourio-
Mazoyer, 氮. (2016). Surface-based morphometry of cortical
thickness and surface area associated with Heschl’s gyri duplica-
t io ns i n 43 0 hea lt hy vo lu nt e e r s . Frontiers in Human
神经科学, 10(69). https://doi.org/10.3389/fnhum.2016
.00069, 考研: 27014013
Mårtensson, J。, Eriksson, J。, Bodammer, 氮. C。, Lindgren, M。,
约翰逊, M。, Nyberg, L。, & Lövdén, 中号. (2012). Growth of
language-related brain areas after foreign language learning.
神经影像, 63(1), 240–244. https://doi.org/10.1016/j
.neuroimage.2012.06.043, 考研: 22750568
Mc Ewen, 乙. (2015). Epigenetics and learning. 趋势
Neuroscience and Education, 4(4), 108–111. https://doi.org/10
.1016/j.tine.2015.11.002
Moreno, S。, Bialystok, E., Barac, R。, Schellenberg, 乙. G。, Cepeda,
氮. J。, & Chau, 时间. (2011). Short-term music training enhances
verbal intelligence and executive function. Psychological
科学, 22 (11 ), 14 25–1 433 . h ttps://do i.org/10 .1 177
/0956797611416999, 考研: 21969312
Moreno, S。, & Bidelman, G. 中号. (2014). Examining neural plasticity
and cognitive benefit through the unique lens of musical training.
Hearing Research, 308, 84–97. https://doi.org/10.1016/j.heares
.2013.09.012, 考研: 24079993
Morgan-Short, K., Deng, Z。, Brill-Schuetz, K. A。, Faretta-Stutenberg,
M。, 黄, 磷. C. M。, & 黄, F. C. K. (2015). A view of the neu-
ral representation of second language syntax through artificial
language learning under implicit contexts of exposure. 学习
in Second Language Acquisition, 37(2), 383–419. https://doi.org
/10.1017/S0272263115000030
Morgan-Short, K., Faretta-Stutenberg, M。, Brill-Schuetz, K. A。,
Carpenter, H。, & 黄, 磷. C. 中号. (2014). Declarative and proce-
dural memory as individual differences in second language
acquisition. Bilingualism, 17(1), 56–72. https://doi.org/10.1017
/S1366728912000715
Möttönen, R。, Dutton, R。, & Watkins, K. 乙. (2013). Auditory-motor
processing of speech sounds. 大脑皮层, 23(5), 1190–1197.
https://doi.org/10.1093/cercor/bhs110, 考研: 22581846
Moyer, A. (2008). Input as a critical means to an end: Quantity and
quality of experience in L2 phonological attainment. 在T. Piske
& 中号. Young-Scholten (编辑。), Input matters in SLA (PP. 159–174).
Multilingual Matters. https://doi.org/10.21832/9781847691118-011
Mueller, J. L。, Friederici, A. D ., & Mannel, C. (2012). Auditory per-
ception at the root of language learning. 诉讼程序
美国国家科学院,
1 09 ( 3 9) , 15 9 53 – 15 9 58 . h t t p s : / / d oi . o r g / 10 . 1 0 73 / p na s
.1204319109, 考研: 23019379
Mukherjee, P。, 磨坊主, J. H。, Shimony, J. S。, Conturo, 时间. E., 李, 乙. C。,
Almli, C. R。, & McKinstry, 右. C. (2001). Normal brain maturation
during childhood: Developmental trends characterized with
diffusion-tensor MR imaging. Radiology, 221, 349–358. https://
doi.org/10.1148/radiol.2212001702, 考研: 11687675
Neurobiology of Language
411
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
Nardo, D ., & Reiterer, S. 中号. (2009). Musicality and phonetic
language aptitude. In G. Dogil & S. 中号. Reiterer (编辑。),
Language talent and brain activity (PP. 213–256). Mouton de
Gruyter.
Neubauer, A. C。, & Fink, A. (2009). Intelligence and neural effi-
ciency. Neuroscience and Biobehavioral Reviews, 33(7), 1004–1023.
https://doi.org/10.1016/j.neubiorev.2009.04.001, 考研:
19580915
Novén, M。, Schremm, A。, Nilsson, M。, Horne, M。, & Roll, 中号.
(2019). Cortical thickness of Broca’s area and right homologue
is related to grammar learning aptitude and pitch discrimination
proficiency. Brain and Language, 188, 42–47. https://doi.org/10
.1016/j.bandl.2018.12.002, 考研: 30572263
Onnis, L。, Truzzi, A。, & Ma, X. (2018). Language development and
disorders: Possible genes and environment interactions. 研究
in Developmental Disabilities, 82, 132–146. https://doi.org/10
.1016/j.ridd.2018.06.015, 考研: 30077386
Ortiz-Mantilla, S。, Hämäläinen, J. A。, Realpe-Bonilla, T。, & Benasich,
A. A. (2016). Oscillatory dynamics underlying perceptual narrowing
of native phoneme mapping from 6 到 12 months of age. 杂志
神经科学, 36(48), 12095–12105. https://doi.org/10.1523
/JNEUROSCI.1162-16.2016, 考研: 27903720
Partanen, E., Kujala, T。, Näätänen, R。, Liitola, A。, Sambeth, A。, &
Huotilainen, 中号. (2013). Learning-induced neural plasticity of
speech processing before birth. 国家会议录
美国科学院, 110(37),
15145–15150. https://doi.org/10.1073/pnas.1302159110,
考研: 23980148
Patel, A. D. (2012). 语言, music and the brain: A resource-
sharing framework. 在P. Rebuschat, 中号. Rohrmeier, J. Hawkins,
& 我. 叉 (编辑。), Language and music as cognitive systems
(PP. 204–223). 牛津大学出版社. https://doi.org/10.1093
/acprof:oso/9780199553426.003.0022
Paus, 时间. (2010). Growth of white matter in the adolescent brain:
Myelin or axon? Brain and Cognition, 72(1), 26–35. https://土井
.org/10.1016/j.bandc.2009.06.002, 考研: 19595493
Penhune, V. 乙. (2011). Sensitive periods in human development:
Evidence from musical training. Cortex, 47(9), 1126–1137.
https://doi.org/10.1016/j.cortex.2011.05.010, 考研:
21665201
Penhune, V. B., Zatorre, 右. J。, MacDonald, J. D ., & 埃文斯, A. C.
(1996). Interhemispheric anatomical differences in human pri-
mary auditory cortex: Probabilistic mapping and volume mea-
surement from magnetic resonance scans. 大脑皮层, 6(5),
661–672. https://doi.org/10.1093/cercor/6.5.661, 考研:
8921202
Peper, J. S。, Brouwer, 右. M。, Boomsma, D. 我。, Kahn, 右. S。, &
Hulshoff Pol, H. 乙. (2007). Genetic influences on human brain
结构: A review of brain imaging studies in twins. 人类
Brain Mapping, 28(6), 464–473. https://doi.org/10.1002/ hbm
.20398, 考研: 17415783
Perani, D ., Saccuman, 中号. C。, Scifo, P。, Awander, A。, Spada, D .,
Baldoli, C。, Poloniato, A。, Lohmann, G。, & Friederici, A. D.
(2011). Neural language networks at birth. 诉讼程序
美国国家科学院,
1 0 8( 3 8 ) , 1 6 05 6 –1 60 6 1. ht t p s : / / d o i . or g / 1 0 . 10 7 3/ p n a s
.1102991108, 考研: 21896765
Petrides, 中号. (2014). Neuroanatomy of language regions of the human
脑. 学术出版社.
Pierce, L. J。, 陈, J. K., Delcenserie, A。, Genesee, F。, & 克莱因, D.
(2015). Past experience shapes ongoing neural patterns for lan-
规格. Nature Communications, 6, 文章 10073. https://doi.org
/10.1038/ncomms10073, 考研: 26624517
Plomin, 右. (1994). The Emanuel Miller Memorial Lecture 1993:
Genetic research and identification of environmental influences.
Journal of Child Psychology & Psychiatry & Allied Disciplines, 35(5),
817–834. https://doi.org/10.1111/j.1469-7610.1994.tb02297.x,
考研: 7962243
Plomin, R。, & Deary, 我. J. (2015). Genetics and intelligence differences:
Five special findings. Molecular Psychiatry, 20(1), 98–108. https://
doi.org/10.1038/mp.2014.105, 考研: 25224258
Plomin, R。, DeFries, J. C。, Knopik, V. S。, & Neiderhiser, J. 中号. (2016).
Top 10 replicated findings from behavioral genetics. Perspectives
on Psychological Science, 11(1), 3–23. https://doi.org/10.1177
/1745691615617439, 考研: 26817721
Poeppel, D ., & Assaneo, 中号. F.
(2020). Speech rhythms and
their neural foundations. 自然评论神经科学, 2(21),
322–334. https://doi.org/10.1038/s41583-020-0304-4, 考研:
32376899
Pol, H. 乙. H。, Schnack, H. G。, Posthuma, D ., Mandl, 右. C. W.,
Baaré, 瓦. F。, van Oel, C。, van Haren, 氮. E., 柯林斯, D. L。,
埃文斯, A. C。, Amunts, K., Bürgel, U。, Zilles, K., de Geus, E.,
Boomsma, D. 我。, & Kahn, 右. S. (2006). Genetic contributions to
human brain morphology and intelligence. 杂志
神经科学, 26(40), 10235–10242. https://doi.org/10.1523
/JNEUROSCI.1312-06.2006, 考研: 17021179
Pons, F。, Lewkowicz, D. J。, Soto-Faraco, S。, & Sebastián-Gallés, 氮.
(2009). Narrowing of intersensory speech perception in infancy.
Proceedings of the National Academy of Sciences of the United
States of America, 106(26), 10598–10602. https://doi.org/10
.1073/普纳斯.0904134106, 考研: 19541648
Prat, C. S. (2011). The brain basis of individual differences in language
comprehension abilities. Linguistics and Language Compass, 5(9),
635–649. https://doi.org/10.1111/j.1749-818X.2011.00303.x
Prat, C. S。, & Just, 中号. A. (2011). Exploring the neural dynamics under-
pinning individual differences in sentence comprehension.
大脑皮层, 21(8), 1747–1760. https://doi.org/10.1093/cercor
/bhq241, 考研: 21148612
Prat, C. S。, 凯勒, 时间. A。, & Just, 中号. A. (2007). Individual differences
in sentence comprehension: A functional magnetic resonance
imaging investigation of syntactic and lexical processing
需要. 认知神经科学杂志, 19(12), 1950–1963.
https://doi.org/10.1162/jocn.2007.19.12.1950, 考研:
17892384
Prat, C. S。, Yamasaki, 乙. L。, & 彼得森, 乙. 右. (2019). Individual dif-
ferences in resting-state brain rhythms uniquely predict second
language learning rate and willingness to communicate in adults.
认知神经科学杂志, 31(1), 78–94. https://doi.org
/10.1162/jocn_a_01337, 考研: 30240310
Price, C. J. (2010). The anatomy of language: A review of 100 功能磁共振成像
studies published in 2009. Annals of the New York Academy of
科学, 1191, 62–88. https://doi.org/10.1111/j.1749-6632
.2010.05444.X, 考研: 20392276
Price, C. J. (2012). A review and synthesis of the first 20 years of PET
and fMRI studies of heard speech, spoken language and reading.
神经影像, 62(2), 816–847. https://doi.org/10.1016/j
.neuroimage.2012.04.062, 考研: 22584224
Pulvermüller, F。, & Fadiga, L. (2010). Active perception:
Sensorimotor circuits as a cortical basis for language. 自然
评论 神经科学, 11(5), 351–360. https://doi.org/10.1038
/nrn2811, 考研: 20383203
Pulvermüller, F。, & Schumann, J. H. (1994). Neurobiological mecha-
nisms of language acquisition. Language Learning, 44(4), 681–734.
https://doi.org/10.1111/j.1467-1770.1994.tb00635.x
齐, Z。, Han, M。, 王, Y。, de los Angeles, C。, 刘, Q., Garel, K.,
陈, 乙. S。, Whitfield-Gabrieli, S。, Gabrieli, J. D. E., &
Neurobiology of Language
412
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
Perrachione, 时间. K. (2019). Speech processing and plasticity in the
right hemisphere predict variation in adult foreign language
学习. 神经影像, 192, 76–87. https://doi.org/10.1016/j
.neuroimage.2019.03.008, 考研: 30853566
Rademacher, J。, Morosan, P。, Schormann, T。, Schleicher, A。,
Werner, C。, Freund, H. J。, & Zilles, K. (2001). Probabilistic mapping
and volume measurement of human primary auditory cortex.
神经影像, 13(4), 669–683. https://doi.org/10.1006/nimg.2000
.0714, 考研: 11305896
Reiterer, S. M。, Berger, 中号. L。, Hemmelmann, C。, & Rappelsberger,
磷. (2005). Decreased EEG coherence between prefrontal elec-
特罗德斯: A correlate of high language proficiency? Experimental
脑研究, 163(1), 109–113. https://doi.org/10.1007
/s00221-005-2215-z, 考研: 15821933
Reiterer, S. M。, Hemmelmann, C。, Rappelsberger, P。, & Berger,
中号. L. (2005). Characteristic functional networks in high- 相对
low-proficiency second language speakers detected also during
native language processing: An explorative EEG coherence study
在 6 频带. Cognitive Brain Research, 25(2), 566–578.
https://doi.org/10.1016/j.cogbrainres.2005.08.010, 考研:
16185851
Reiterer, S. M。, 胡, X。, Erb, M。, Rota, G。, Nardo, D ., Grodd, W.,
Winkler, S。, & Ackermann, H. (2011). Individual differences in
audio-vocal speech imitation aptitude in late bilinguals: Functional
neuro-imaging and brain morphology. 心理学前沿, 2,
文章 271. https://doi.org/10.3389/fpsyg.2011.00271, 考研:
22059077
Reiterer, S. M。, Pereda, E., & Bhattacharya, J. (2009). 测量
second language proficiency with EEG synchronization: 如何
functional cortical networks and hemispheric involvement differ
as a function of proficiency level in second language speakers.
Second Language Research, 25(1), 77–106. https://doi.org/10
.1177/0267658308098997
Ressel, 五、, Pallier, C。, Ventura-Campos, N。, Díaz, B., Roessler, A。, Ávila,
C。, & Sebastián-Gallés, 氮. (2012). An effect of bilingualism on the
auditory cortex. 神经科学杂志, 32(47), 16597–16601.
https://doi.org/10.1523/ JNEUROSCI.1996-12.2012, 考研:
23175815
米, 中号. L。, Zubrick, S. R。, 泰勒, C. L。, Hoffman, L。, & Gayán, J.
(2018). Longitudinal study of language and speech of twins at 4
和 6 年: Twinning effects decrease, zygosity effects disap-
pear, and heritability increases. Journal of Speech, 语言,
and Hearing Research, 61(1), 79–93. https://doi.org/10.1044
/2017_JSLHR-L-16-0366, 考研: 29255901
Rimfeld, K., 戴尔, 磷. S。, & Plomin, 右. (2015). How specific is sec-
ond language-learning ability? A twin study exploring the contri-
butions of first language achievement and intelligence to second
language achievement. Translational Psychiatry, 5(9). https://土井
.org/10.1038/tp.2015.128, 考研: 26393484
罗宾逊, G. E., Fernald, 右. D ., & 克莱顿, D. F. (2008). 基因和
social behavior. 科学, 322(5903), 896–900. https://doi.org/10
.1126/science.1159277, 考研: 18988841
罗宾逊, 磷. (2001). Individual differences, cognitive abilities, apti-
tude complexes and learning conditions in second language
acquisition. Second Language Research, 17(4), 368–392. https://
doi.org/10.1177/026765830101700405
罗宾逊, 磷. (2012). Individual differences, aptitude complexes,
SLA processes, and aptitude test development. 在米. Pawlak
(埃德。), New perspectives on individual differences in language
learning and teaching (PP. 57–75). 施普林格. https://doi.org/10
.1007/978-3-642-20850-8_4
Ross-Sheehy, S。, Oakes, L. M。, & Luck, S. J. (2003). 的发展
visual short-term memory capacity in infants. Child Development,
74(6), 1807–1822. https://doi.org/10.1046/j.1467-8624.2003
.00639.X, 考研: 14669897
Sammler, D. (2020). Splitting speech and music. 科学, 367(6481),
974–976. https://doi.org/10.1126/science.aba7913, 考研:
32108099
Saur, D ., Kreher, 乙. W., Schnell, S。, Kümmerera, D ., Kellmeyera, P。,
Vrya, 中号. S。, Umarova, R。, Musso, M。, Glauche, 五、, Abel, S。,
Huber, W., Rijntjes, M。, Hennig, J。, & Weiller, C. (2008).
Ventral and dorsal pathways for language. 诉讼程序
美国国家科学院,
1 05 ( 4 6) , 18 0 35 – 18 0 40 . h t t p s : / / d oi . o r g / 10 . 1 0 73 / p na s
.0805234105, 考研: 19004769
Scarr, S。, & McCartney, K. (1983). How people make their own
环境: A theory of genotype → environment effects.
Child Development, 54(2), 424–435. https://doi.org/10.2307
/1129703, 考研: 6683622
Schellenberg, 乙. G。, & Peretz, 我. (2008). 音乐, language and cog-
尼尼申: Unresolved issues. 认知科学的趋势, 12(2),
45–46. https://doi.org/10.1016/j.tics.2007.11.005, 考研:
18178126
Schellenberg, 乙. G。, & Winner, 乙. (2011). Music training and nonmu-
sical abilities: 介绍. Music Perception, 29(2), 129–132.
https://doi.org/10.1525/mp.2011.29.2.129
施耐德, J. W., & McGrew, K. S. (2018). The Cattell-Horn-Carroll
theory of cognitive abilities. 在D中. 磷. 弗拉纳根 & 乙. 中号. McDonough
(编辑。), Contemporary intellectual assessment: Theories, tests and
问题 (PP. 73–163). The Guilford Press.
施耐德, P。, Scherg, M。, Dosch, H. G。, Specht, H. J。, Gutschalk,
A。, & Rupp, A. (2002). Morphology of Heschl’s gyrus reflects
enhanced activation in the auditory cortex of musicians. 自然
神经科学, 5(7), 688–694. https://doi.org/10.1038/nn871,
考研: 12068300
施耐德, P。, Sluming, 五、, 罗伯茨, N。, Scherg, M。, Goebel, R。,
Specht, H. J。, Dosch, H. G。, Bleeck, S。, Stippich, C。, & Rupp,
A. (2005). Structural and functional asymmetry of lateral
Heschl’s gyrus reflects pitch perception preference. 自然
神经科学, 8(9), 1241–1247. https://doi.org/10.1038
/nn1530, 考研: 16116442
Scholz, J。, 克莱因, 中号. C。, 贝伦斯, 时间. 乙. J。, & Johansen-Berg, H.
(2009). Training induces changes in white matter architecture.
自然神经科学, 12, 1370–1371. https://doi.org/10.1038
/nn.2412, 考研: 19820707
Schön, D ., Magne, C。, & Besson, 中号. (2004). The music of speech:
Music training facilitates pitch processing in both music and lan-
规格. Psychophysiology, 41(3), 341–349. https://doi.org/10
.1111/1469-8986.00172.X, 考研: 15102118
斯科特, L. S。, Pascalis, 奥。, & 纳尔逊, C. A. (2007). A domain-general
theory of the development of perceptual discrimination. 当前的
Directions in Psychological Science, 16(4), 197–201. https://土井
.org/10.1111/j.1467-8721.2007.00503.x, 考研: 21132090
Sedaghatgoftar, N。, Karimi, 中号. N。, Babaii, E., & Reiterer, S. 中号.
(2019). Developing and validating a second language pragmatics
aptitude test. Cogent Education, 6(1), 文章 1654650. https://土井
.org/10.1080/2331186X.2019.1654650
Seither-Preisler, A。, 帕恩卡特, R。, & 施耐德, 磷. (2014). Size and
synchronization of auditory cortex promotes musical, literacy, 和
attentional skills in children. 神经科学杂志, 34(33),
10937–10949. https://doi.org/10.1523/ JNEUROSCI.5315-13
.2014, 考研: 25122894
Serrallach, B., Groß, C。, Bernhofs, 五、, Engelmann, D ., Benner, J。,
Gündert, N。, Blatow, M。, Wengenroth, M。, Seitz, A。, Brunner, M。,
Seither, S。, 帕恩卡特, R。, 施耐德, P。, & Seither-Preisler, A. (2016).
Neural biomarkers for dyslexia, ADHD, and ADD in the auditory
Neurobiology of Language
413
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
cortex of children. Frontiers in Neuroscience, 10(324), 1–23. https://
doi.org/10.3389/fnins.2016.00324, 考研: 27471442
夏普, 中号. J。, Batchelor, H. M。, Mueller, L. E., 加德纳, 中号. 磷. H。, &
Schoenbaum, G. (2021). Past experience shapes the neural circuits
recruited for future learning. 自然神经科学, 24(3), 391–400.
https://doi.org/10.1038/s41593-020-00791-4, 考研: 33589832
Sheppard, J. P。, 王, J.-P., & 黄, 磷. C. 中号. (2012). Large-scale
cortical network properties predict future sound-to-word learning
成功. 认知神经科学杂志, 24(5), 1087–1103.
https://doi.org/10.1162/jocn_a_00210, 考研: 22360625
Shtyrov, Y。, Nikulin, V. 五、, & Pulvermüller, F. (2010). Rapid cortical
plasticity underlying novel word learning. 神经科学杂志,
30(50), 16864–16867. https://doi.org/10.1523/JNEUROSCI.1376
-10.2010, 考研: 21159957
Skehan, 磷. (1998). A cognitive approach to language learning.
牛津大学出版社.
Skehan, 磷. (2002). Theorising and updating aptitude. 在P.
罗宾逊 (埃德。), Individual differences and instructed language
学习 (PP. 69–95). John Benjamins. https://doi.org/10.1075
/lllt.2.06ske
Skehan, 磷. (2016). Foreign language aptitude, acquisitional sequences,
and psycholinguistic processes. In G. Granena, D. 氧. Jackson, & 是.
Yilmaz (编辑。), Cognitive individual differences in second language
processing and acquisition (PP. 17–41). John Benjamins. https://
doi.org/10.1075/bpa.3.02ske
Skeide, 中号. A。, Brauer, J。, & Friederici, A. D. (2016). Brain functional
and structural predictors of language performance. Cerebral
Cortex, 26(5), 2127–2139. https://doi.org/10.1093/cercor
/bhv042, 考研: 25770126
Skeide, 中号. A。, & Friederici, A. D. (2016). The ontogeny of the cor-
tical language network. 自然评论神经科学, 17(5),
323–332. https://doi.org/10.1038/nrn.2016.23, 考研:
27040907
Sparks, 右. L. (2012). Individual differences in L2 learning and long-
term L1–L2 relationships. Language Learning, 62(s2), 5–27.
https://doi.org/10.1111/j.1467-9922.2012.00704.x
Sparks, 右. L。, Patton, J。, Ganschow, L。, & Humbach, 氮. (2009).
Long-term crosslinguistic transfer of skills from L1 to L2.
Language Learning, 59(1), 203–243. https://doi.org/10.1111/j
.1467-9922.2009.00504.X
斯坦因, M。, Dierks, T。, Brandeis, D ., Wirth, M。, Strik, W., & Koenig,
时间. (2006). Plasticity in the adult language system: A longitudinal
electrophysiological study on second language learning.
神经影像, 33(2), 774–783. https://doi.org/10.1016/j
.neuroimage.2006.07.008, 考研: 16959500
斯坦因, M。, Federspiel, A。, Koenig, T。, Wirth, M。, Strik, W., Wiest, R。,
Brandeis, D ., & Dierks, 时间. (2012). Structural plasticity in the lan-
guage system related to increased second language proficiency.
Cortex, 48(4), 458–465. https://doi.org/10.1016/j.cortex.2010.10
.007, 考研: 21106192
Strike, L. T。, Couvy-Duchesne, B., Hansell, 氮. K., Cuellar-Partida,
G。, Medland, S. E., & 赖特, 中号. J. (2015). Genetics and brain
morphology. Neuropsychology Review, 25(1), 63–96. https://
doi.org/10.1007/s11065-015-9281-1, 考研: 25773500
Stromswold, K. (2001). The heritability of language: A review and
metaanalysis of twin, 采用, and linkage studies. 语言,
77(4), 647–723. https://doi.org/10.1353/lan.2001.0247
汤普森, 磷. M。, Cannon, 时间. D ., Narr, K. L。, van Erp, T。, Poutanen,
V. P。, Huttunen, M。, Lönnqvist, J。, Standertskjöld-Nordenstam,
C. G。, Kaprio, J。, Khaledy, M。, Dail, R。, Zoumalan, C. 我。, & Toga,
A. 瓦. (2001). Genetic influences on brain structure. 自然
神经科学, 4(12), 1253–1258. https://doi.org/10.1038
/nn758, 考研: 11694885
Towse, J. N。, & Hitch, G. J. (2007). Variation in working memory
due to normal development. 在一个. 右. A. 康威, C. Jarrold, 中号. J.
Kane, A. Miyake, & J. 氮. Towse (编辑。), Variation in working mem-
奥里 (PP. 109–133). 牛津大学出版社. https://doi.org/10
.1093/acprof:oso/9780195168648.003.0005
Turker, S。, & Reiterer, S. 中号. (2021). Brain, musicality and language ap-
titude: A complex interplay. Annual Review of Applied Linguistics,
41, 95–107. https://doi.org/10.1017/S0267190520000148
Turker, S。, Reiterer, S. M。, 施耐德, P。, & Seither-Preisler, A. (2019).
Auditory cortex morphology predicts language learning potential
in children and teenagers. Frontiers in Neuroscience, 13, 1–16.
https://doi.org/10.3389/fnins.2019.00824, 考研: 31447639
Turker, S。, Reiterer, S. M。, Seither-Preisler, A。, & 施耐德, 磷.
(2017). “When music speaks”: Auditory cortex morphology as
a neuroanatomical marker of language aptitude and musicality.
心理学前沿, 8, 文章 2096. https://doi.org/10.3389
/fpsyg.2017.02096, 考研: 29250017
Ullman, 中号. 时间. (2004). Contributions of memory circuits to lan-
规格: The declarative/procedural model. 认识, 92(1–2),
231–270. https://doi.org/10.1016/j.cognition.2003.10.008,
考研: 15037131
Ullman, 中号. 时间. (2015). The declarative/procedural model. 在乙.
VanPatten & J. 威廉姆斯 (编辑。), Theories in second language
acquisition: An introduction (2nd 版。, PP. 135–158). 劳特利奇.
Ullman, 中号. 时间. (2016). The declarative/procedural model: A neuro-
biological model of
language learning, 知识, and use.
In G. Hickok & S. L. 小的 (编辑。), Neurobiology of language
(PP. 953–968). 爱思唯尔. https://doi.org/10.1016/ B978-0-12
-407794-2.00076-6
Vaquero, L。, Rodríguez-Fornells, A。, & Reiterer, S. 中号. (2017). 这
左边, the better: White-matter brain integrity predicts foreign
language imitation ability. 大脑皮层, 27(8), 3906–3917.
https://doi.org/10.1093/cercor/bhw199, 考研: 27461123
Vaquero, L。, Rousseau, 磷. N。, Vozian, D ., 克莱因, D ., & Penhune,
V. 乙. (2020). What you learn & when you learn it: Impact of early
bilingual & music experience on the structural characteristics of
auditory-motor pathways. 神经影像, 213, 文章 116689.
https://doi.org/10.1016/j.neuroimage.2020.116689, 考研:
32119984
Ventura-Campos, N。, 桑胡安, A。, González, J。, Palomar-García,
中号. Á., Rodríguez-Pujadas, A。, Sebastián-Gallés, N。, 德科, G。,
& Ávila, C. (2013). Spontaneous brain activity predicts learning
ability of foreign sounds. 神经科学杂志, 33(22),
9295–9305. https://doi.org/10.1523/JNEUROSCI.4655-12.2013,
考研: 23719798
Verhoef, E., Shapland, C. Y。, Fisher, S. E., 戴尔, 磷. S。, & St Pourcain,
乙. (2020). The developmental origins of genetic factors influenc-
ing language and literacy: Associations with early-childhood
词汇. Journal of Child Psychology and Psychiatry, 62(6),
728–738. https://doi.org/10.1111/jcpp.13327, 考研:
32924135
Veroude, K., Norris, D. G。, Shumskaya, E., Gullberg, M。, &
Indefrey, 磷. (2010). Functional connectivity between brain re-
gions involved in learning words of a new language. Brain and
语言, 113(1), 21–27. https://doi.org/10.1016/j.bandl.2009
.12.005, 考研: 20116090
Vigneau, M。, Beaucousin, 五、, Hervé, P.-Y., Jobert, G。, 佩蒂特, L。,
Crivello, F。, Mellet, E., Zago, L。, Mazoyer, B., & Tzourio-
Mazoyer, 氮. (2011). What is right-hemisphere contribution to
phonological, lexico-semantic, and sentence processing?
Insights from a meta-analysis. 神经影像, 54(1), 577–593.
https://doi.org/10.1016/j.neuroimage.2010.07.036, 考研:
20656040
Neurobiology of Language
414
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of language aptitude
Vinkhuyzen, A. A. E., van der Sluis, S。, Posthuma, D ., & Boomsma,
D. 我. (2009). The heritability of aptitude and exceptional talent
across different domains in adolescents and young adults.
Behavior Genetics, 39(4), 380–392. https://doi.org/10.1007
/s10519-009-9260-5, 考研: 19288254
Watkins, K., & Paus, 时间. (2004). Modulation of motor excitability
during speech perception: The role of Broca’s area. 杂志
Cognitive Neuroscience, 16(6), 978–987. https://doi.org/10
.1162/0898929041502616, 考研: 15298785
韦尔斯, G. (1986). The meaning makers: Children learning language
and using language to learn. Hodder and Stoughton.
Wen, Z. (2019). Working memory as language aptitude: 这
phonological/executive model. In Z. Wen, 磷. Skehan, A. Biedron(西德:1),
S. 李, & 右. L. Sparks (编辑。), Language aptitude: Advancing theory,
testing, research and practice (PP. 187–214). 劳特利奇. https://
doi.org/10.4324/9781315122021-10
Wen, Z。, Biedron(西德:1), A。, & Skehan, 磷. (2017). Foreign language apti-
tude theory: Yesterday, today and tomorrow. Language Teaching,
50(1), 1–31. https://doi.org/10.1017/S0261444816000276
Wengenroth, M。, Blatow, M。, Heinecke, A。, Reinhardt, J。, Stippich,
C。, Hofmann, E., & 施耐德, 磷. (2014). Increased volume and
function of right auditory cortex as a marker for absolute pitch.
大脑皮层, 24(5), 1127–1137. https://doi.org/10.1093
/cercor/bhs391, 考研: 23302811
白色的, 乙. J。, Hutka, S. A。, 威廉姆斯, L. J。, & Moreno, S. (2013).
学习, neural plasticity and sensitive periods: Implications
for language acquisition, music training and transfer across the
lifespan. Frontiers in Systems Neuroscience, 7(90), 1–18.
https://doi.org/10.3389/fnsys.2013.00090, 考研: 24312022
Winner, 乙. (1996). Gifted children: Myths and realities. Basic
图书.
黄, F. C. K., Chandrasekaran, B., Garibaldi, K., & 黄,
磷. C. 中号. (2011). White matter anisotropy in the ventral language
pathway predicts sound-to-word learning success. 杂志
神经科学, 31(24), 8780–8785. https://doi.org/10.1523
/JNEUROSCI.0999-11.2011, 考研: 21677162
黄, 磷. C. M。, & Ettlinger, 中号. (2011). Predictors of spoken lan-
guage learning. Journal of Communication Disorders, 44(5),
564–567. https://doi.org/10.1016/j.jcomdis.2011.04.003,
考研: 21601868
黄, 磷. C. M。, & Perrachione, 时间. K. (2007). Learning pitch pat-
terns in lexical identification by native English-speaking adults.
Applied Psycholinguistics, 28(4), 565–585. https://doi.org/10
.1017/S0142716407070312
黄, 磷. C. M。, Perrachione, 时间. K., & Parrish, 时间. 乙. (2007). Neural
characteristics of successful and less successful speech and word
learning in adults. 人脑图谱, 28(10), 995–1006.
https://doi.org/10.1002/hbm.20330, 考研: 17133399
黄, 磷. C. M。, Skoe, E., Russo, 氮. M。, Dees, T。, & Kraus, 氮. (2007).
Musical experience shapes human brainstem encoding of linguis-
tic pitch patterns. 自然神经科学, 10(4), 420–422. https://
doi.org/10.1038/nn1872, 考研: 17351633
黄, 磷. C. M。, Warrier, C. M。, Penhune, V. B., Roy, A. K., Sadehh, A。,
Parrish, 时间. B., & Zatorre, 右. J. (2008). Volume of left Heschl’s gyrus
and linguistic pitch learning. 大脑皮层, 18(4), 828–836.
https://doi.org/10.1093/cercor/bhm115, 考研: 17652466
Wulf, G。, & Lewthwaite, 右. (2016). Optimizing performance
through intrinsic motivation and attention for learning: 这
OPTIMAL theory of motor learning. Psychonomic Bulletin and
审查, 23(5), 1382–1414. https://doi.org/10.3758/s13423-015
-0999-9, 考研: 26833314
Xiang, H。, Dediu, D ., 罗伯茨, L。, van Oort, E., Norris, D. G。, &
Hagoort, 磷. (2012). The structural connectivity underpinning
language aptitude, 工作记忆, and IQ in the perisylvian
language network. Language Learning, 62(s2), 110–130. https://
doi.org/10.1111/j.1467-9922.2012.00708.x
Yamagata, B., Murayama, K., 黑色的, J. M。, Hancock, R。, Mimura,
M。, 哪个, 时间. T。, Reiss, A. L。, & Hoeft, F. (2016). Female-specific
intergenerational transmission patterns of the human corticolim-
bic circuitry. 神经科学杂志, 36(4), 1254–1260. https://
doi.org/10.1523/ JNEUROSCI.4974-14.2016, 考研:
26818513
哪个, J。, Gates, K. M。, Molenaar, P。, & 李, 磷. (2015). Neural changes
underlying successful second language word learning: An fMRI
学习. Journal of Neurolinguistics, 33, 29–49. https://doi.org/10
.1016/j.jneuroling.2014.09.004
哪个, J。, & 李, 磷. (2012). Brain networks of explicit and implicit
学习. PLOS ONE, 7(8), e42993. https://doi.org/10.1371
/journal.pone.0042993, 考研: 22952624
于, A. C. L。, & Zellou, G. (2019). Individual differences in language
加工: Phonology. Annual Review of Linguistics, 5, 131–150.
https://doi.org/10.1146/annurev-linguistics-011516-033815
Zatorre, 右. J。, Fields, 右. D ., & Johansen-Berg, H. (2012). Plasticity in
gray and white: Neuroimaging changes in brain structure during
学习. 自然神经科学, 15(4), 528–536. https://doi.org
/10.1038/nn.3045, 考研: 22426254
张, Y。, Kuhl, 磷. K., Imada, T。, Iverson, P。, Pruitt, J。, Stevens,
乙. B., Kawakastu, M。, Tohkura, Y。, & Nemoto, 我. (2009).
Neural signatures of phonetic learning in adulthood: A magne-
toencephalography study. 神经影像, 46(1), 226–240. https://
doi.org/10.1016/j.n euroimage.2009.01.028, 考研:
19457395
张, Y。, Kuhl, 磷. K., Imada, T。, Kotani, M。, & Tohkura, 是. (2005).
Effects of language experience: Neural commitment to language-
specific auditory patterns. 神经影像, 26(3), 703–720. https://
doi.org/10.1016/j.neuroimage.2005.02.040 , 考研:
15955480
张, Y。, & 王, 是. (2007). Neural plasticity in speech acquisi-
tion and learning. Bilingualism, 10(2), 147–160. https://doi.org
/10.1017/S1366728907002908
Neurobiology of Language
415
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
2
3
3
8
9
1
9
5
8
6
7
8
n
哦
_
A
_
0
0
0
4
2
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3