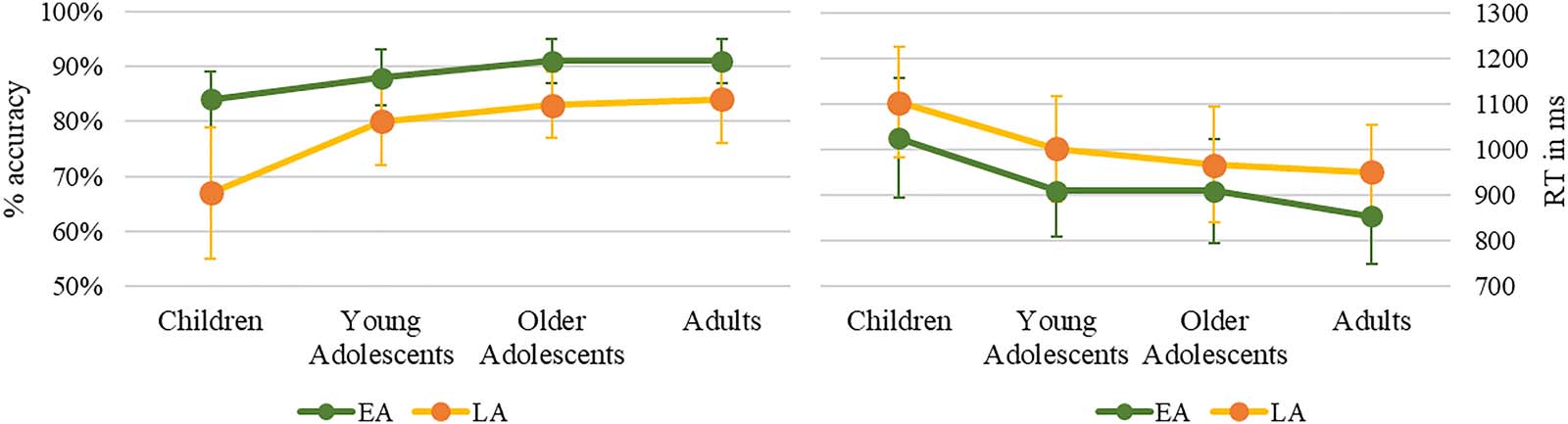RESEARCH ARTICLE
Dynamics of Word Production in the Transition
from Adolescence to Adulthood
开放访问
杂志
Tanja Atanasova1
, Raphaël Fargier2
, Pascal Zesiger1
, and Marina Laganaro1
1Faculty of Psychology and Educational Sciences, University of Geneva, 日内瓦, 瑞士
2Laboratoire Parole et Langage, Aix-Marseille University, Marseille, 法国
关键词: ERP, language production, 青春期, language development, topographic maps,
electrophysiology
抽象的
Changes in word production occur across the lifespan. Previous studies have shown
electrophysiological, 颞, and functional differences between children and adults
accompanying behavioral changes in picture-naming tasks (Laganaro, Tzieropoulos,
Fraunfelder, & Zesiger, 2015). 因此, a shift toward adult-like processes in referential word
production occurs somewhere between the ages of 13 和 20. Our aim was to investigate
when and how children develop adult-like behavior and brain activation in word production.
Toward this aim, performance and event-related potentials (ERP) in a referential word production
task were recorded and compared for two groups of adolescents (aged 14 到 16 和 17 到 18),
孩子们 (aged 10 到 13), and young adults (aged 20 到 30). Both groups of adolescents displayed
adult-like production latencies, which were longer only for children, while accuracy was lower
in the younger adolescents and in children, compared to adults. ERP waveform analysis and
topographic pattern analysis revealed significant intergroup differences in key time-windows on
stimulus-locked ERPs, both early (150–220 ms)—associated with pre-linguistic processes—and
晚的 (280–330 ms)—associated with lexical processes. The results indicate that brain activation
underlying referential word production is completely adult-like in 17-year-old adolescents,
whereas an intermediate pattern is still observed in adolescents aged 14 到 16 years old, 虽然
their production speed, but not their accuracy, is already adult-like.
介绍
The social aptitude of speaking is universal and unique to our species; converting prelinguistic
concepts into articulated words and sentences within an ephemeral time period represents one of
the major human-specific abilities. This ability is manifested from the moment children say their
first word (IE。, around the twelfth month after birth), yet the accuracy and the speed of word pro-
duction evolve during the lifespan, along with the size of the vocabulary. The lexicon grows from
大约 300 entries after one year of life (戴尔 & Fenson, 1996) 到 10,000 by the age
of six (克拉克, 1993), 以及关于 60,000 words in adulthood (磨坊主, 1996).
Compared to adults, school-aged children present reduced vocabulary, lower accuracy, 和
slower processing speed. 尽管如此, the same psycholinguistic variables, namely lexical
频率, concept familiarity, word age of acquisition (AoA), visual complexity of pictures, 和
word length, seem to predict both accuracy and production latencies in children (Cycowicz,
弗里德曼, Rothstein, & Snodgrass, 1997), as well as in adults (D’Amico, Devescovi, & Bates,
2001). This suggests that the same encoding processes underlie single-word production in children
引文: Atanasova, T。, Fargier, R。,
Zesiger, P。, & Laganaro, 中号. (2020).
Dynamics of word production in the
transition from adolescence to
成年期. Neurobiology of Language.
Advance publication. https://doi.org
/10.1162/nol_a_00024
DOI:
https://doi.org/10.1162/nol_a_00024
支持信息:
https://doi.org/10.1162/nol_a_00024
已收到: 3 九月 2020
公认: 9 九月 2020
利益争夺: 作者有
声明不存在竞争利益
存在.
通讯作者:
Tanja Atanasova
tanja.atanasova@unige.ch
处理编辑器:
Kate Watkins
版权: © 2020 马萨诸塞州
Institute of Technology. 已发表
under a Creative Commons Attribution
4.0 国际的 (抄送 4.0) 执照.
麻省理工学院出版社
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
哦
_
A
_
0
0
0
2
4
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
and adults, regardless of the behavioral differences between the two groups. Such a claim is chal-
lenged by functional neuroimaging results showing that differences between children and adults
are not limited to processing speed; 实际上, 一些, but not all, mental processes underlying word
production in picture-naming tasks seem to differ between children and adults (Laganaro,
Tzieropoulos, Fraunfelder, & Zesiger, 2015). Electrophysiological event-related potentials (ERPs)
of referential word production differ both qualitatively (IE。, different underlying neural generators)
and quantitatively (IE。, different durations of similar processes) between school-aged children (7 到
8 和 10 到 12 years old) and young adults (Laganaro et al., 2015), leaving open the issue of when
the brain mechanisms underlying word production become adult-like. 所以, the present study
intends to fill the gap between childhood and adulthood, by investigating the functional and tem-
poral dynamics of changes in word production from childhood to adulthood, focusing on
青春期.
Most of the psycholinguistics models of speech production based on adult behavior agree that
word production involves three main levels of processing, namely conceptual, 词汇的, 和
phonological-phonetic processes (戴尔, 1986; Levelt, 1989). The dynamics of these mental pro-
cesses have largely been investigated on populations of young adults, both behaviorally and with
neuroimaging techniques. Indefrey and Levelt (2004; see also Indefrey, 2011) have framed pre-
cise spatial and temporal functional features of word production through a meta-analysis of
previously published behavioral and event-related EEG/MEG studies using pictures to elicit
utterance production. The authors proposed distinctive markers of lexical ((西德:1)200 ms after picture
presentation) and phonological ((西德:1)450 ms after picture presentation) processing consistent with
the abovementioned language production models. Other authors have proposed different
dynamics, involving parallel activation of lexical and phonological information (Miozzo,
Pulvermüller, & Hauk, 2014; Strijkers, Costa, & Pulvermuller, 2017), but empirical evidence is
not yet conclusive in favor of one or the other model (Munding, Dubarry, & Alario, 2016).
In referential naming tasks, 然而, children and adults largely differ in accuracy and speed:
Children display about 300 到 400 ms longer production latencies than adults (D’Amico et al.,
2001). The similarity in the psycholinguistic predictors of referential naming may suggest that the
same processes underlie word production in children and adults, and that only timing differs—
那是, the same underlying processes are scaled in time and that the differences between adults’
and children’s speech production systems are quantitative rather than qualitative. This intuition
has been challenged by some qualitative differences observed in neuroimaging studies directly
comparing utterance production in children and adults. Laganaro et al. (2015) compared
children to adults in a picture-naming task using global field pattern (topographic) ERP analyses
to establish whether the differences in production speed were due to functional dissimilarity or to
analogous processes merely shifted in time. The authors showed that the longer production
latencies in 10- to 12-year-old children relative to adults were associated with both quantitative
(temporal changes, same topographies shifted in time) and qualitative differences (functional
变化, different topographies). 尤其, qualitative functional differences were observed
in early time-windows, around the P1 and N1 components and likely related to pre-linguistic
流程. Other ERP studies investigating language production in children reported mainly
temporal shifts in 12-year-old children relative to adults in a verb production task (Budd,
Paulmann, Barry, & Clahsen, 2013) or enhanced amplitudes in children relative to adults in a
picture-naming and word-reading task (Greenham & Stelmack, 2001). In terms of spatial
resolution, functional magnetic resonance imaging (功能磁共振成像) comparing language production in
children and adults showed fundamentally overlapping activations between the two groups,
but also higher hemodynamic response in children (Brown et al., 2005), just as the opposite
(Brown et al., 2005; Krishnan, 水蛭, Mercure, Lloyd-Fox, & 迪克, 2014). Krishnan et al. (2014)
Neurobiology of Language
2
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
哦
_
A
_
0
0
0
2
4
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
suggested that differences in the activation patterns among children and adults underlay the qual-
itative changes in processes, affecting the delay and the strength of the spatial activation.
因此, previous studies have shown clear behavioral changes between school-age children
and adults, both in terms of accuracy and speed, whereas it is still unclear whether different brain
processes in word production underpin such changes. Even more unclear is when, during devel-
选项, performance and brain processes underlying word production become adult-like.
Adolescence is likely the transition period when a shift toward adult-like processes occurs both
in behavior and in brain processes. It is certainly well known that the brain itself undergoes mas-
sive changes during the teenage years. MRI studies have shown that the cerebral cortex seems to
shrink during adolescence, when significant region-specific structural and functional changes are
observed, leading to a unique pattern of brain responses and cognition (Dumontheil, 2016).
Structural changes are detected in both grey and white matter, with grey matter volume reaching
its peak during mid- to late childhood, decreasing during adolescence (Dumontheil, 2016).
Cortical thinning is believed to be associated with underlying synaptic pruning (Alemán-
Gómez et al., 2013); counterintuitively, a higher degree of pruning seems to be observed in highly
used brain processes, which would allow associations requiring constant updating to have a
larger palette of neuronal connections, linked with higher cognitive functions such as language
(Riley et al., 2018). This phenomenon is predominantly detected in frontal and occipital corti-
ces, as a consequence of sulcal widening, linked to cortical thinning, which is associated with
gyral white matter expansion (Alemán-Gómez et al., 2013). White matter has been shown to
increase linearly during the first two to three decades of life (Dumontheil, 2016). White matter’s
growth probably reflects changes in myelination and axonal diameter (Alemán-Gómez et al.,
2013), with white matter pathways showing progressive maturation during adolescence and
consequently reflecting the changes in cognitive performance (Riley et al., 2018). These changes
can also be detected with EEG.
In terms of EEG power, the elimination of active synapses and changes in white matter may be
the reflection of the neural processes driving the age-related decreases in low frequencies (在
absolute power measure) and increases in high frequencies (only in relative power; Goossens,
Vercammen, Wouters, & van Wieringen, 2016). The whole process is, 然而, indicative of
continuing development of the thalamo-cortical and cortico-cortical networks that generate
EEG activity (Cragg et al., 2011). Axonal myelination and the integrity of white matter pathways
have been suggested to underlie peak alpha frequency shifts (Miskovic et al., 2015). From early to
late childhood, alpha-band electro-cortical connectivity becomes more integrated, less function-
ally segregated, and more spatially variable: Older children are less spatially homogenous, 和
the variability of cortical states being related to improved performance on task-based measures
(Miskovic et al., 2015). Changes in EEG and very large changes in early ERP components reported
in cognitive tasks (Holcomb, 科菲, & Neville, 1992) are in line with the continued changes in
brain structure and function that have been observed throughout the adolescent period
(Segalowitz, Santesso, & Jetha, 2010). Adolescence thus constitutes the key age that should
be investigated relative to the question raised above, namely when word production becomes
adult-like.
The Current Study
To fill the gap between previous studies comparing school-age children and adults, here we
investigate ERP changes underlying word production in school-age children, adolescents, 和
adults in a referential word production task. Given that the processing speed is bound to differ
across groups, it was necessary to target a specific word-encoding process in the ERP time-course;
we therefore manipulated a variable known to similarly affect word production in children and
Neurobiology of Language
3
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
哦
_
A
_
0
0
0
2
4
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
adults, namely AoA of words. Word AoA has been identified as one of the most influential
predictors of word production latencies in adults (Chalard, Bonin, Méot, 博耶, & Fayol, 2003;
Johnston & Barry, 2006; Juhasz, 2005) and in children (Morrison, Chappell, & Ellis, 1997). 它有
also been reported as having a solid effect on ERPs in time-windows associated with lexical-
phonological encoding in adults (see Perret, Bonin, & Laganaro, 2014; 薛, 刘, Marmolejo-
Ramos, & 裴, 2017).
In the present study, we therefore analyze local waveform and global topographic changes
associated with picture naming on early- and late-acquired words in four groups of speakers:
孩子们 (aged 10–12), young adolescents (aged 14–16), older adolescents (aged 17–18), 和
young adults (aged 20–30). The manipulation of AoA will allow us to follow the onset of the
processes modulated by the same psycholinguistics factors across groups. Built on the spatio-
temporal segmentation of the signal, the global topographic analyses enable the summary of
neural activity in terms of stable electrophysiological activities at scalp (Lehmann & Skrandies,
1984). Combining local waveform and global topographic analyses thus constitutes a great tool
to evaluate quantitative (IE。, same topographic patterns, varying in duration) and/or qualitative
(IE。, different topographic patterns) differences in neural events across groups of individuals.
MATERIALS AND METHOD
参加者
A total of 80 participants divided into 4 团体 (孩子们, young adolescents, older adolescents,
and adults) participated in the study. 孩子们 (n = 20; 8 女性) were aged from 10 years and zero
months to 12 years and 11 月 (average: 11;05 ± 0;83). Adolescents were recruited in two
团体, with a one-year gap in between used to clearly separate the groups: Young adolescents
(n = 20; 7 女性) were aged from 14 到 16 年 (average: 15;26 ± 0;73). Older adolescents (n =
20; 12 女性) were aged from 17 到 18 年 11 月 (average: 17;45 ± 0;51 月). 这
participants in the adult group (n = 20; 13 女性) were aged from 20 到 30 (average: 24;45 ±
2;98). All participants were right-handed and French native speakers without diagnosed language
impairment or delay or neurological disease. Adults were recruited through announcements
posted at the university, whereas children and adolescents were recruited through personal con-
tacts of the authors. This study was approved by the ethical committee of the Faculty of Psychology
and Educational Science of the University of Geneva. Written informed consent from adolescents
and adult participants, and additional parents’ approval for children and adolescents, 是
collected, in accordance with the Declaration of Helsinki.
Tasks and Material
Picture naming
The picture naming task used 120 black and white drawings and their corresponding modal names,
selected from two French databases (Alario & 费朗, 1999; Bonin, Peerman, Malardier, Méot, &
Chalard, 2003). The stimuli corresponded to words with an AoA range of 1.19–3.55 on a five-
point scale (1: learned between 0 和 3 years–4: learned between 9 和 12 年; 5: 学到了
after the age of 12 was not included) to ensure that all the words were known even by the
youngest group of participants (see Laganaro et al., 2015) and with high name agreement
(mean = 92.5%). 这 120 items were split in two groups of 60 items each, corresponding to early-
acquired words (mean = 1.73, 标准差= 0.25) and late-acquired words (mean = 2.72, 标准差= 0.59).
早期的- and late-acquired words were matched on the other relevant psycholinguistic variables
from the two mentioned databases (name agreement and visual complexity of the pictures) 和
cumulative frequency, lexical frequency, number of phonological neighbors, and on four
Neurobiology of Language
4
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
哦
_
A
_
0
0
0
2
4
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
sublexical variables (length in phonemes, sonority values of the first phoneme, syllables, 和
phonemes frequencies) from the French lexical database Lexique (新的, Pallier, Brysbaert, &
费朗, 2004).
Cognitive assessments
In addition to the experimental tasks, the participants underwent several cognitive tests in order
to verify that each participant was in the normal range of their respective age and to assess age-
related changes in our groups: the vocabulary test and the digit span from the Wechsler Adult
Intelligence Scale (Wechsler, 1997) for adolescents from 16 years old and adults, and from the
Wechsler Intelligence Scale for Children (朱 & 韦斯, 2005) for the younger participants; 二
verbal fluency tests (animals and letter P in two minutes) (Cardebat, Doyon, Puel, Goulet, &
Joanette, 1990); and a reading fluency test (a short text made up of 265 字). Participants also
underwent two reaction time (RT) 测试: a simple RT test, consisting in hitting a specific key when
a cross appeared on screen, and a choice RT test consisting in having two possible stimuli and
two possible responses, in this case hitting “D” on the keyboard when the longer line appearing
on the screen was on the left, and “L” when the longest line appeared on the right.
程序
The participants from the four groups underwent the same picture-naming experiment under
EEG/ERP recording in a dimly lit, sound-attenuated room. For the picture-naming task, 刺激
were presented on a 17-inch screen (refreshment rate: 50 赫兹) at a viewing distance of about
60 厘米. The black line drawings appeared on a white foreground, sized at 9.5 × 9.5 厘米,
against a grey background. The task sequence was controlled by a PC running the E-Prime
软件 (E-Studio). Each trial started with a fixation cross presented for 500 ms in the center of
屏幕, followed by the appearance of the picture for 3,000 多发性硬化症, during which the subjects
had to overtly produce the word corresponding to the picture as fast and accurately as pos-
兄弟姐妹. The interstimulus interval was set at 2,000 多发性硬化症. All items were presented in two different
controlled orders (regular and reversed), with a self-managed break in the middle of the task
(后 60 刺激). The controlled order was preferred to a complete randomization to avoid suc-
cession of stimuli from the same semantic category or with high phonological overlap. Four
warm-up filler trials were set at the beginning of the experiment and after the break. Overt word
productions were recorded by a dynamic microphone, digitally amplified, and the signal was
redirected to a computer. Production latencies (RT in milliseconds, i.e. the time separating the
onset of the picture and the onset of the speech wave) were systematically checked with speech
analysis software (CheckVocal 2.2.6, Protopapas, 2007).
The cognitive assessments were run in a standard room in a face-to-face setting with the
experimenter. All the tests were administered in a standard paper-and-pencil setting, 除了
the two RT tests, which were computer-based.
Behavioral Data Analyses
For the picture naming task, 异常值 (RTs shorter than 500 ms or longer than 2,000 多发性硬化症) 和
response errors were removed from the dataset. Instances when the answer did not exactly match
the expected response, as well as any articles or hesitation marks preceding the answer, 是
considered errors. RT data were fitted with a linear regression mixed model (Baayen, 2008)
and accuracy data were fitted with a generalized linear mixed-effects model for binomially
distributed outcomes (Jaeger, 2008) with the R-software (R-project, R Development Core
Team, 2005), including participants and items as random effect variables and groups as fixed
variable.
Neurobiology of Language
5
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
哦
_
A
_
0
0
0
2
4
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
For the cognitive assessments, standard scores were calculated when possible (IE。, 任务
from the Wechsler assessment), otherwise raw results were reported.
EEG Acquisition and Preprocessing
The EEG was recorded using the Active-Two Biosemi EEG system (Biosemi V.O.F. 阿姆斯特丹,
荷兰) with a 128-electrode cap. Sampling frequency was set at 512 赫兹 (filters: 直流
到 104 赫兹, 3 dB/octave slope). The custom online reference of the system is the common
mode sense active electrode; the driven right leg passive electrode drives the average po-
tentials as close as possible to the amplifier zero (for details on this setup see http://万维网
. b i o s e m i . c o m ) . The preprocessing was conducted with Cartool software 3.60 (Brunet,
穆雷, & Michel, 2011). Epochs of 300 time-frames (TFs) time-locked to 50 TFs before the
picture onset (stimulus-locked) and epochs of 250 TFs time-locked to 50 TFs before the vocal
onset (response-locked) were extracted and averaged for each subject across conditions.
Aligning ERPs to 50 TF (大约 100 多发性硬化症) before the vocal onset of each single trial is
done to remove prearticulatory motor artifacts (see Fargier, Buerki, Pinet, Alario, & Laganaro,
2018). A minimum of 55 trials was averaged per participant. Data were high-pass filtered at
0.2 Hz and low-pass filtered at 30 赫兹 (a second order acausal Butterworth filter with −12 dB/
octave roll-off ) and averaged for each participant and condition (early- or late-acquired
字). Stimulus-locked epochs were extracted without baseline correction, and with
baseline correction on the 50 prestimulus TFs. Response-locked epochs are without base-
line correction, given that there is no consensus on whether and over which period base-
line correction should be applied in response-locked ERPs. All epochs related to correct
productions were recalculated against the average reference, visually inspected, 和交流电-
cepted only in the absence of artifact, such as eyeblinks, motor artifacts, or large amplitude
variations. Only trials with artifact-free stimulus- and response-locked epochs were re-
泰内德. Contaminated electrodes (max 20% 的 128 electrodes) were interpolated with
a 3-D spline interpolation (Perrin, Pernier, Bertrand, Giard, & Echaller, 1987).
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
哦
_
A
_
0
0
0
2
4
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
EEG Data Analyses
All analyses were run on average-referenced data. The ERPs were first subjected to a sam-
pling point-wise ERP waveform analysis to determine the time-periods presenting local dif-
ferences in ERP amplitudes between age-groups and between AoA conditions.
Comparisons of each electrode and for each time-point were run separately on stimulus-
locked and response-locked ERPs using 4 × 2 analysis of variance (ANOVAs) on amplitudes
comparing the four groups in the two conditions using the sten toolbox (https://zenodo.org
/record/1167723#.X3IoDmgzaUm). An effect was considered significant if it lasted for at
至少 10 consecutive TFs and was present on five adjacent electrodes, alpha being set to
0.01.
Because waveform differences do not inform us of the qualitative versus quantitative nature of
the observed changes, the main analysis consisted in the spatio-temporal segmentation (topo-
graphic or microstate analysis). Considering that periods of stable global electric fields likely
correspond to particular periods in mental information processing (尚古克斯 & Michel, 2004;
Koukou & Lehmann, 1987; Lehmann & Skrandies, 1984; Lehmann, Strik, Henggeler, Koenig, &
Koukkou, 1998), topographic differences across groups would indicate different underlying
brain processes. The spatio-temporal segmentation was performed on the group-averaged
ERPs of each subgroup and condition, and was statistically validated in the responses of single
participants as described below.
Neurobiology of Language
6
Word production in the transition from adolescence to adulthood
Before running the topographic analysis, we performed topographic consistency tests
(Koenig & Melie-García, 2010) on stimulus-locked and response-locked ERPs to ensure that
different topographies were not driven by random noise in the within-group data. This analysis
allowed us to determine that topographic patterns were indeed consistent across subjects
(see Supplementary Material in the online supporting information, located at https://万维网
.mitpressjournals.org/doi/suppl/10.1162/nol_a_00024).
The first topographic analysis consisted in identifying periods of significant topographic
modulation between groups and conditions on each sampling point on stimulus- 并重新-
sponse-locked ERPs. This procedure is called topographic analysis of variance (tANOVA); 它
consists in a nonparametric randomization test based on global dissimilarities between electric
fields, 哪个, in contrast to channel comparisons, computes global dissimilarity of all electric
field topographies and tests for the significance of the said topographic differences at each time-
观点 (see an example of its computation in Murray, Brunet, & Michel, 2008). The permutation
of the data is accomplished by randomly reassigning the topographic maps of single subjects to
the different conditions. The global dissimilarity of these random group-averaged ERPs is com-
pared time-point by time-point with the values of topographic dissimilarity of the actual con-
版本. The criterion we applied was 12 TFs (关于 24 多发性硬化症) of consecutive significant
差异. Neurophysiologically, differences in topography directly indicate changes in the
configuration of the active neuronal sources in the brain. In order to test for differences in to-
pography, tANOVA is used. The comparison of the tANOVA result with the microstate seg-
mentation is therefore important to disentangle topographic differences that are yielded by
temporal shifts of the same patterns of global distribution of the signal at scalp from topo-
graphic differences that are the consequences of different underlying sources.
For the common topographic analysis of stimulus- and response-locked ERPs, the “overlap”
between stimulus-locked ERPs and response-locked ERPs was removed based on average
RTs of each group, following the procedure described in Laganaro and Perret (2011) 和
Laganaro (2014). The same procedure was applied to each individual ERP and used in the fitting
procedure (见下文). 因此, the combination of stimulus- and response-locked ERPs
covered the exact time-interval from picture onset to 50 TF before articulation onset for each
group and for each participant in each condition.
Scalp electric field topography remains in a stable configuration for a limited period of time:
Differences in topography indicate changes in the configuration of the active neuronal sources
in the brain (Murray et al., 2008). This analysis involves a spatio-temporal segmentation of the
ERPs over periods of electrophysiological stability, by compressing the variability of ERPs in a series
of topographical maps that summarize data and indicate which template explains ERPs in each
团体. A temporal atomized and agglomerate hierarchical clustering algorithm (Murray et al.,
2008) to identify the template maps was applied to the group-averaged data. With the aim of
selecting the optimal number of maps most likely explaining the ERP signal, the minimum
remaining time of a given topography was 12 TFs (大约 24 多发性硬化症) 和 95% correlation.
Microstate segmentation was applied to the eight group- and condition-averaged ERP data sets.
To determine whether the differences observed on the grand averages reflect a single participant’s
neural responses, a comparison between grand averages and individual data was conducted.
This “fitting” procedure, provides the information required to understand how much a topo-
graphic map observed in a given group actually explains an individual participant’s ERP data.
Measures of map (microstate) presence and global explained variance (GEV) in each individual
ERP were obtained and used to test statistical differences among groups and conditions. When a
topographic map was consistently present, its duration in the participants’ ERP signal was also
used to compare conditions.
Neurobiology of Language
7
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
哦
_
A
_
0
0
0
2
4
p
d
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
结果
Behavioral Results
The performances of all the participants were within the norms on the vocabulary test and the digit
span memory test. On the other cognitive tasks, where no standard score was available, no subject
obtained outlying results. On the three tests where a comparison was possible (due to age adap-
站, some of the tests were not identical, see Materials and Method), an increase of performance
can be observed across the age groups. 桌子 1 displays the results per group on these cognitive
测试. In both verbal fluencies (animals and letter), a group effect was observed—respectively,
Kruskal-Wallis test H(3, N= 80) = 14.80443, p = 0.0020, and Kruskal-Wallis test H(3, N=
80) = 21.73078, p = 0.0001—with children providing fewer items per category than older ad-
olescents and adults (动物: z = 3.54, p = 0.002, and z = 3.06, p = 0.01; letter: z = 3.65, p =
0.001 and z = 4.32, p = 0.00009), but without significant difference with young adolescents
(动物: z = 2.27, p = 0.15; letter: z = 2.64, p > 0.05). A group effect was also observed on
reading fluency—Kruskal-Wallis test H(3, 79) = 18.55257, p = 0.0003—with slower reading
times in children relative to older adolescents and adults (z = 3.893, p = 0.0006, and z =
3.335, p = 0.005). Young adolescents did not differ from the other groups. Similar results ap-
peared on simple and choice RT—group effect simple: Kruskal-Wallis test: H(3, N= 80) =
14.76358, p = 0.002; group effect choice: Kruskal-Wallis test: H(3, N= 80) = 21.09, p =
0.0001. Older adolescents and adults differed from children, who displayed the slowest process-
ing speed (simple older adolescents: z = 3.368, p = 0.005; simple adults: z = 3.035, p = 0.01;
choice older adolescents: z = 4.062, p = 0.0003; choice adults: z = 3.538, p = 0.002), 然而
young adolescents did not differ from children, nor from older adolescents, nor from adults.
In the picture-naming task (见图 1), production accuracy was lower in children relative to
adults (z = −4.508, p < 0.0001, (cid:1) = −0.83, SE = 0.184) and in young adolescents relative to adults
(z = −2.133, p < 0.05, (cid:1) = −0.405, SE = 0.1897), but without significant difference between the
two groups of adolescents (z = −0.364, p = 0.716, (cid:1) = −0.0714, SE = 0.19611), both differing from
children (young adolescents: z = −4.588, p < 0.0001, (cid:1) = −1.104, SE = 0.24; older adolescents: z =
−6.092, p < 0.0001, (cid:1) = −1.488, SE = 0.24). Furthermore, accuracy was better for early-acquired
words than for late-acquired words, in all groups (children: z = 4.905, p < 0.0001, (cid:1) = 1.4598, SE =
0.2976; young adolescents: z = 2.533, p < 0.05, (cid:1) = 0.7774, SE = 0.30687; older adolescents: z =
2.306, p < 0.02, (cid:1) = 0.725, SE = 0.314; adults: z = −1.969, p < 0.05, (cid:1) = −0.625, SE = 0.3172).
Production latencies also differed across groups, F(3, 79.8) = 10.035, p < 0.0001, and word
AoA F(1, 118.8) = 23.2385, p < 0.0001, without interaction between groups and AoA,
F(3, 7829.3) < 1. Between-group differences were significant for children as compared to
both adolescent groups, young adolescents: t(86.4477) = 3.298, p = 0.001; older adolescents:
t(86.2356) = 4.337, p < 0.0001, and to adults, t(79.985) = 5.075, p < 0.0001. Both adolescent
Table 1. Mean results on the cognitive tests for each age group
Children
Young adolescents
Older adolescents
Adults
Verbal fluency (score)
Letter
Category
19
28
Reading fluency
(seconds)
123
Reaction time (ms)
Choice
Simple
469
325
35
40
37
25
28
28
87
78
75
315
290
278
418
369
358
Note. Each score represents the number of correct answers provided by the group.
Neurobiology of Language
8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
Figure 1. Naming accuracy (in %) and reaction times (RT; in ms) for groups and conditions. EA: early-acquired words, LA: late-acquired words.
groups did not differ from adults, young adolescents: t(79.674) = 1.666, p = 0.0996; older ado-
lescents: t(79.498) = 0.972, p = 0.334, nor among them, t(85.02) = −1.04, p = 0.3.
ERP Results
Waveforms
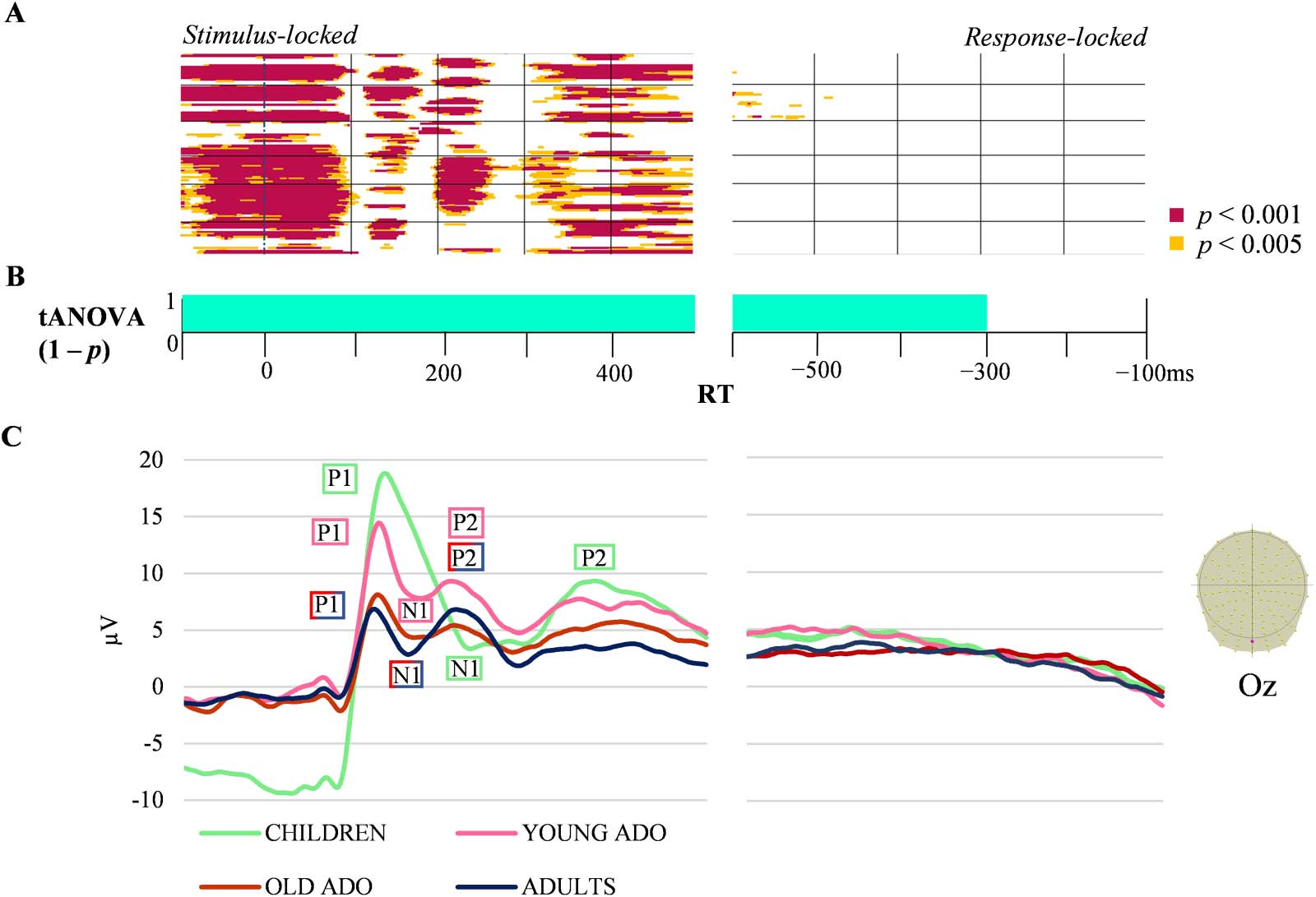
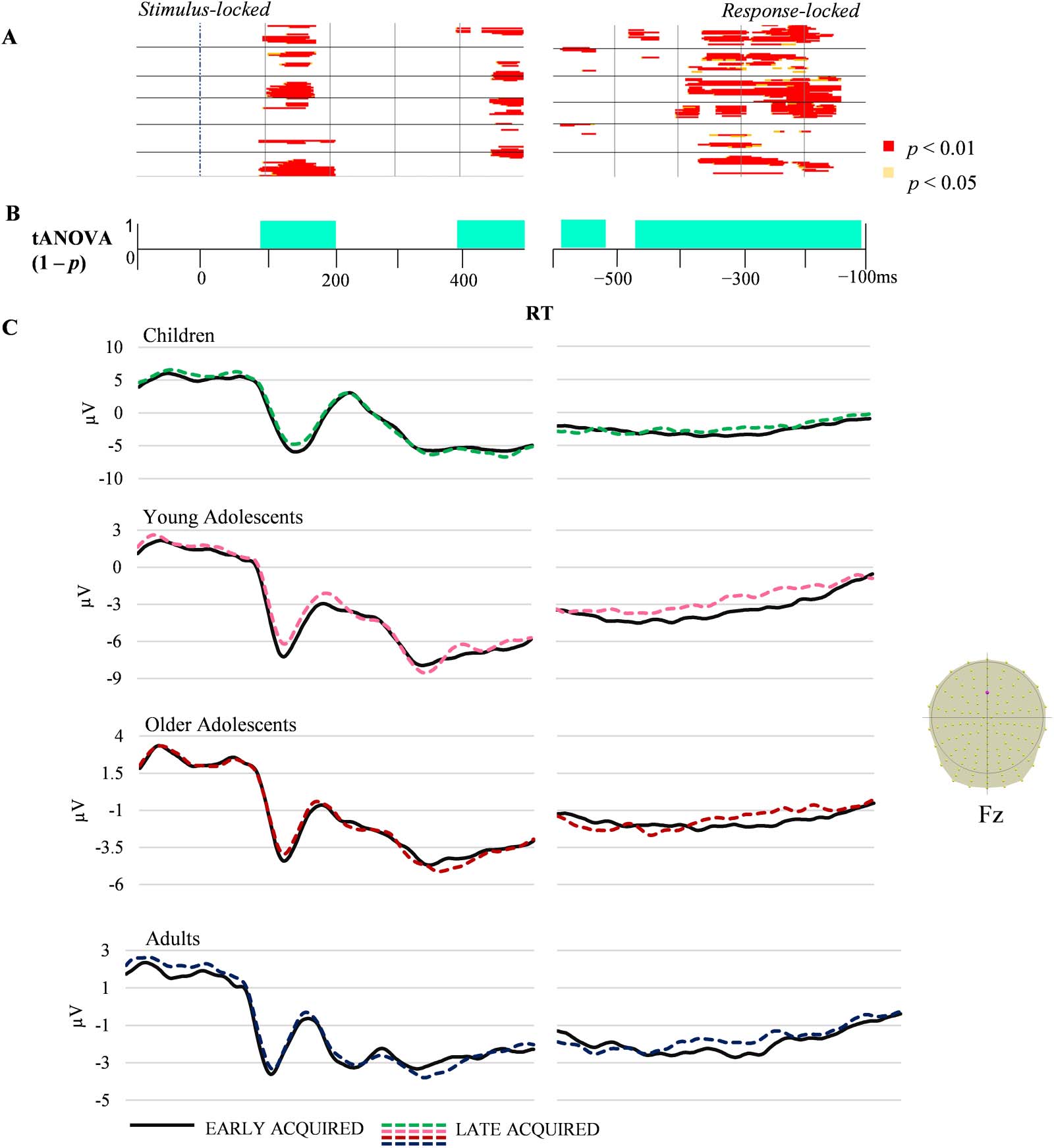
The 4 × 2 ANOVA showed significant time-periods of group effect (Figure 2; see Supplementary
Materials in the online supporting information for stimulus-locked periods of differences in
amplitudes and exemplar of the group-averaged waveform [Oz] for the four groups) and also of
AoA effect (Figure 3) on ERP waveforms, without interaction between groups and AoA condition.
The group effect yielded significant differences in amplitudes on most electrodes throughout the
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Children, young & older adolescents and adults group-averaged stimulus-aligned and response-aligned event-related potentials (ERPs; 128
electrodes). (A) Periods of significant differences in amplitudes across groups on each electrode and time-frame for the stimulus-locked and response-
locked ERPs. (B) Periods of significant differences in the tANOVA analysis ( p < 0.01) in turquoise. (C) Exemplars of group-averaged ERP waveforms (Oz)
for the four groups are plotted in microvolts in function of time (bottom panel). The shifting of the components in the stimulus-locked ERPs is labeled (with
older adolescents (ADO) and adults sharing the label due to similar amplitudes and components). RT: reaction time.
Neurobiology of Language
9
Word production in the transition from adolescence to adulthood
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Condition-averaged stimulus- and response-locked event-related potentials (ERPs; 128 electrodes). (A) Periods of significant differences
in amplitudes between early- and late-acquired words on each electrode and time-frame for the stimulus-locked and response-locked ERPs. (B)
Periods of significant differences in the tANOVA analysis ( p < 0.01) are displayed in turquoise. (C) Exemplar condition-averaged ERP waveform
(Fz) for the four groups plotted in microvolts in function of time. RT: reaction time.
entire stimulus-locked time-period, whereas virtually no significant differences were found in the
response-locked ERPs (Figure 2). The extended statistical differences in time and space in the
stimulus-locked ERPs reflect the larger amplitudes and latency shifts between the groups
observed on the exemplar of waveforms in Figure 3. The younger groups were characterized
by higher amplitudes and delayed latencies of ERP components. The results of waveform analyses
computed on two consecutive groups at a time (see Appendix A in the online supporting infor-
mation) indicated that amplitudes differ between children and young adolescents in the very first
time-window (−100 to 100 ms) and from about 190 to 250 ms. Between young and older
adolescents, amplitudes were significantly different only on a short time-window around 110 ms,
but waveform amplitude differed between young adolescents and adults in the 300 to 350 ms
time-window on a large proportion of electrodes.
Neurobiology of Language
10
Word production in the transition from adolescence to adulthood
To validate the visually-observed latency shift, the latency of the P1 component (around 100 ms
post picture onset) was also extracted for each age group based on the latency of maximal global
field power value in the 80 to 160 ms time-window. A group effect was observed, Kruskal-Wallis
test H(3, 160) = 20.74, p = 0.0001, with a later P1 peak in children relative to older adolescents
and to adults (z = 3.96, p = 0.0004 and z = 3.84, p = 0.0007), but without significant difference
with young adolescents (z = 2.13, p = 0.2). Given these results, we computed the correlation
between the P1 peak and age. On the entire group of participants, the negative correlation
was low although significant (r = −0.21, p < 0.05), whereas on the two youngest groups the
correlation was not significant (r = −0.14, p > 0.05).
The AoA effect yielded different amplitudes between early- and late-acquired words around
100 ms after stimulus appearance and towards the end of the stimulus-locked signal, 后 400 多发性硬化症,
on most electrodes (数字 3). On the response-locked ERPs, a long period covering from 480 到
100 ms prior to participant response showed significant differences on most electrodes, and a
shorter significance period of 80 ms starting around 600 ms before articulation was observed
on central bilateral electrodes (数字 3).
tANOVA
The tANOVA analysis also revealed differences across groups throughout all the stimulus-locked
ERPs, 但是也 600 到 300 ms prior to the vocal onset for the response-locked ERPs (见图 2).
For AoA, the tANOVA was significant in the same time-windows described for amplitudes.
Microstate analysis
The microstate segmentation applied on the eight grand averages (4 groups × 2 状况)
revealed a best model explaining 95% of variance with 12 different topographic maps from
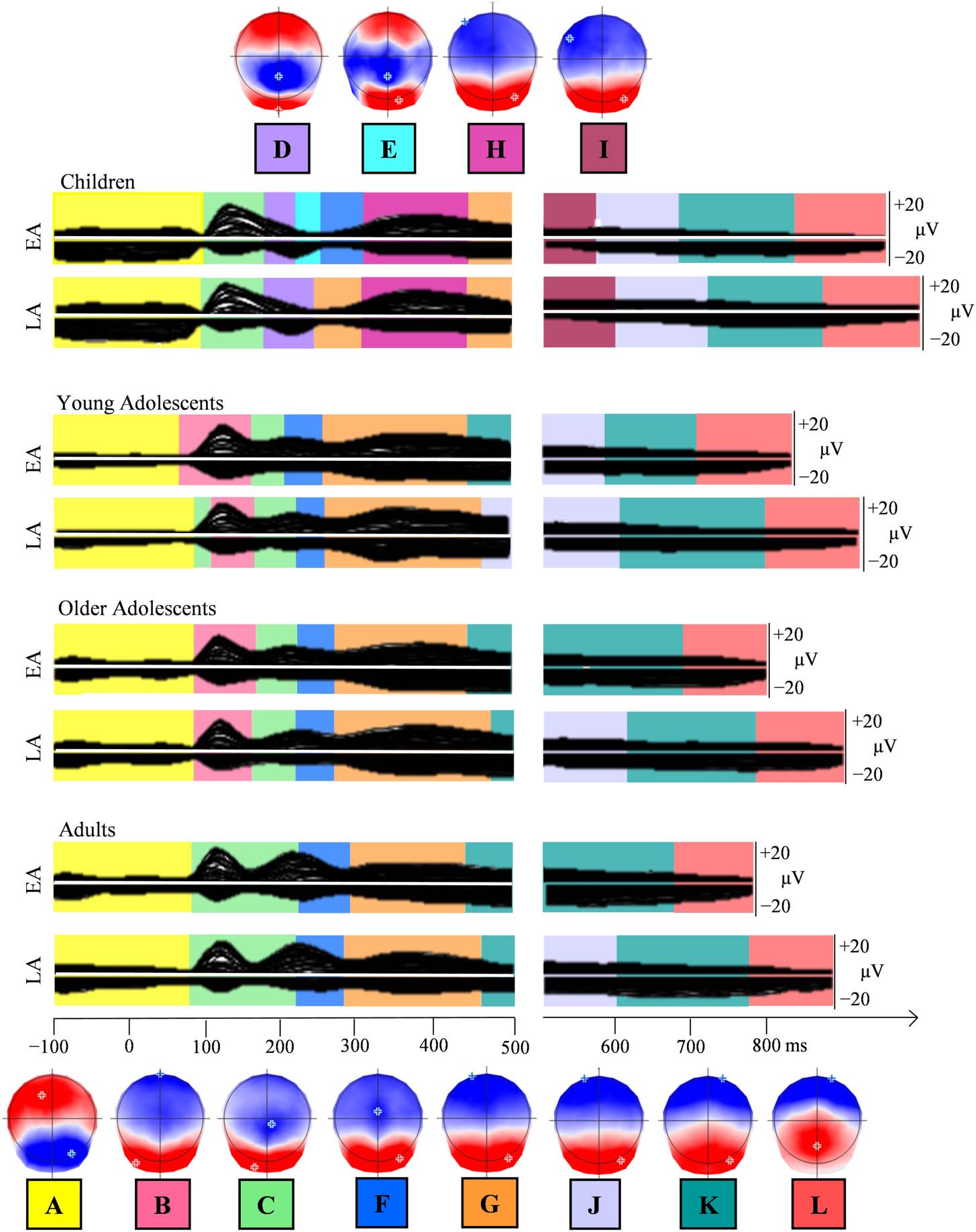
100 ms before picture onset to 100 ms before vocal onset (数字 4). On the grand averages,
different periods of topographic stability appeared for children, as compared to older groups, 在
early time-windows (maps D, 乙, H in Figure 4). Both groups of adolescents displayed similar maps
to adults, after the P1 component. On response-locked epochs, the sequence of topographic maps
was similar across age-groups.
Three fitting periods were applied to the individual combined stimulus- and response-locked
ERPs to statistically assess the differences observed on the grand averages. The first fitting period
was meant to target the microstate underlying the P1 component, thus spanned from 35 TF to
103 TF (68 到 202 多发性硬化症) after picture onset, testing for Maps B and C. The second period spanned
从 100 TF (~200 ms) to the end of the stimulus-locked signal and included maps D, 乙, F, G, 和H.
In the third period covering the remaining individual ERPs locked to response onset, maps I, J, K,
and L were fitted. The results of the fitting in the individual ERPs, in terms of presence, 全球的
explained variance, and duration, are displayed in Appendix B in the online supporting information.
Group comparisons. Among the two microstates observed around the P1 component, Map B was
present in most participants from the three older groups and in only half of the children, 然而
Map C was observed in 19 在......之外 20 children but also in a high proportion of participants from the
other groups (see Presence in Appendix B in the online supporting information). 不同的
distribution of the two P1 maps across groups was statistically confirmed by the interaction
between groups and maps on the GEV, F(3, 76) = 6.07, p = 0.009. Pearson’s correlation
computed between age and GEV of Map B on all participants showed no significant correlation
(r = −0.14, p > 0.05), whereas a significant negative correlation was observed on Map C (r =
−0.49, p < 0.05), further showing that the presence of Map C decreases with age.
Neurobiology of Language
11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Group-averaged event-related potentials (ERPs; 128 electrodes) for each group (children, young and older adolescents, adults) and
condition (EA & LA) from 100 ms before the stimulus onset to 100 ms before vocal onset (after the concatenation process, which removed the
overlap between stimulus-locked and response-locked ERPs according to each group average response latencies). The temporal distribution of the
topographic maps in each data set revealed by the spatio-temporal segmentation analysis is superposed with code colors. The 12 corresponding
template maps are displayed with positive values in red and negative values in blue. EA: early-acquired words, LA: late-acquired words.
In the second fitting period, Map D and—to a lesser extent—Map H are predominantly present
in children as opposed to all other groups (see Presence in Appendix B in the online supporting
information). This was confirmed by the statistical analyses, with children differing from all the
other groups ( p < 0.05) on GEV, D: F(3, 76) = 7.846, p = 0.0001, MSE = 0.0004; H: F(3, 76) =
7.318, p = 0.001, MSE = 0.015, except for the comparison with young adolescents on Map H,
where there is no significant difference. There was actually a significant correlation between GEV
and age on these two groups (children and young adolescents: r = −0.44, p < 0.05), showing that
Neurobiology of Language
12
Word production in the transition from adolescence to adulthood
the presence of Map H decreases between the ages of 10 and 16. Presence in the individual ERP
signal was very low for Map E and will not be analyzed further. Map F was present in about half of
the participants from each group (see Presence in Appendix B in the online supporting informa-
tion) without statistical difference across groups.
Map G was consistently observed in all the groups, but with between-group differences in the
explained variance, F(3, 76) = 5.958, p = 0.001, MSE = 0.048, and on duration, F(3, 76) = 6.4735,
p = 0.0005, MSE = 19,174, with children differing from all the other groups on GEV ( p < 0.05) and
young adolescents differing from older adolescents and adults ( p < 0.05). Map G actually better
characterized the ERP signal in the young adolescent group relative to the other groups (see GEV
and Duration in Appendix B in the online supporting information).
Maps K and L were present in the ERP signal of the participants from all groups. This was less
the case for Maps I and J, which were more present in children and adolescents than in adults. In
fact, the GEV for Map J yielded a group effect, F(3, 76) = 3.21, p = 0.02, MSE = 0.006, with young
adolescents differing from adults. Map K did not present any significant effect on either GEV or
duration. Map L presented a group effect on GEV, F(3, 76) = 2.87, p < 0.05, MSE = 0.003, with
children differing only from adults.
In terms of AoA, there was a significant effect on the P1 Maps,
Age of Acquisition comparisons.
F(1, 76) = 25.20, p < 0.0001, with higher explained variance for early-acquired words, and no
interaction between AoA and Maps or between AoA and groups. Map D, which was specific to
children, was modulated by AoA on GEV, F(1, 76) = 9.8586, p = 0.003, MSE = 0.00001, with
higher GEV for late-acquired words. AoA modulated both GEV and duration on Map G, GEV:
F(1, 76) = 5.958, p = 0.04, MSE = 0.048; duration: F(1, 76) = 6.8975, p = 0.01, MSE = 2,690,
with slightly longer lasting and higher GEV for early-acquired words (8 ms). No interaction was
observed between AoA and groups. On Map I an AoA effect was observed on GEV, F(1, 76) =
5.134, p = 0.02, MSE = 0.0006, with late-acquired words presenting a larger GEV, but no group
effect or interaction. Map L yielded an AoA effect on duration, F(1, 76) = 16.78, p = 0.0001,
MSE = 13,708, with longer duration for late-acquired words.
To summarize, behavioral results indicated that only children differed from all other groups in
picture-naming speed. On accuracy and on the other cognitive tasks, children statistically
differed only from the two older groups, with young adolescents being somehow intermediate.
In the ERP waveform analyses, young adolescents displayed intermediate amplitudes between
children and older adolescents and adults on the stimulus-locked ERPs. Different patterns under-
lying the P1 component were observed in children (Map C) relative to other groups (Map B).
Different time distributions of a specific pattern underlying the P2 component (see Figure 3C)
were observed in children (Map H), and Map G differed in adults relative to the older group of
adolescents. Age of acquisition effects were observed on topographies approximately in the
same time-windows as amplitude differences in all groups, namely around the P1 component
and after 400 ms.
DISCUSSION
The aim of the present study was to investigate ERP differences in single-word production in two
groups of adolescents compared to children and adults. In particular, we intended to determine
when and how the behavioral and neurophysiological processes underlying word production in
a picture-naming task become adult-like. Reaction times were significantly slower only in chil-
dren as compared to adolescents and adults. Both adolescent groups did not differ significantly
from adults, suggesting an already adult-like behavioral outcome from 14-year-old adolescents.
Neurobiology of Language
13
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
In terms of accuracy of responses, children as well as the younger group of adolescents (14 to
16 years old) were less precise than adults, whereas the older group of adolescents (17 to 18 years
old) did not differ from adults. The same results were also observed on the other cognitive tasks
(verbal fluency, RTs, reading fluency), where the group of young adolescents did not differ
significantly either from children or from older adolescents and adults.
At the neurophysiological level, waveform amplitudes clearly decreased across the first three
age-groups, with the young adolescent group being intermediate between children and the two
other groups. Also, children displayed different global field potentials in early time-windows
(P1-N1), all other groups being alike in these time-windows. However, young adolescents also
yielded some differences relative to the two older groups on some topographies in the stimulus-
locked data, in particular around 300–400 ms after picture onset. No group differences appeared
in the response-locked data on amplitudes, and an extremely similar sequence of stable global
electric fields at scalp was observed in the four groups, with only different time distribution.
A first interpretation of these results is that brain activity underlying referential word production
is completely adult-like in the adolescents over 17 years old, whereas a somehow intermediate
pattern is still observed in the group aged 14 to 16 years old. In the following we will discuss
changes across groups, first on amplitudes, then on microstates, and integrate the functional
interpretation in terms of word-planning processes by integrating the results of the manipulated
psycholinguistic factor (word AoA).
Amplitude and Latency Decrease from Childhood to Adolescence
The present results indicate larger amplitudes in children and young adolescents compared to
adults and older adolescents, whose signal is equivalent. Amplitudes and latency of the P1 com-
ponent are intermediate between children and adults in the 14- to 16-year-old adolescents and
are adult-like after the age of 17. Such differences in amplitudes are observed only in the stimulus-
locked ERPs.
Developmental changes, especially in the P1 time-range of the visual evoked potentials
(VEP; a positive ERP component on posterior electrodes, occurring approximately 100 ms after
stimulus presentation) have been well documented in the literature. Amplitudes decrease with
age, and have been linked to the progressive thickening of children’s skulls (Chauveau et al.,
2004; Picton & Taylor, 2007), as well as to the growing age-related automaticity in the processes
required by the task (Durston & Casey, 2006), automatization being a result of the combination of
myelination (Picton & Taylor, 2007) and synaptic pruning (Itier & Taylor, 2004; Taylor, Edmonds,
McCarthy, & Allison, 2001). The latency shift of the P1 component also decreases with matura-
tion, most likely reflecting the acceleration of stimulus processing (Itier & Taylor, 2004; Taylor,
et al., 2001). Moreover, some studies (Hopfinger & Mangun, 1998; Luck, Woodman, & Vogel,
2000) agree on the fact that attention has an effect on modulating visual-spatial cortical sensory
processing, which results in shifted latencies of the P1 among the youngest participants.
Developmental changes have also been reported in the N1-N170 time-window, with amplitudes
becoming more negative with age and the latency decreasing in visual discrimination tasks
(Batty & Taylor, 2006; Itier & Taylor, 2004). N1 is another major VEP component, characterized
by a central negative peak around 170 milliseconds after stimulus presentation (Creel, 2019).
Similarly to our findings, previous VEP studies showed that children have different ERPs relative
to adolescents and adults. In particular, an additional component is observed in children’s ERPs in
the N170-like time-window (Campbell & Sharma, 2016; Yadav, Poudel, Limbu, Thakur, &
Yadav, 2015). Here, the period of electrophysiological stability observed in this time-window
(see topography of map D, Figure 4) has previously been reported for N1 (N170-like) adult
Neurobiology of Language
14
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
ERPs (Fargier & Laganaro, 2016; Laganaro, 2017; Rossion & Caharel, 2011); we therefore inter-
preted the stability as a single N170-like component.
The stabilization of the VEP component latency is specified to occur only around the twen-
tieth year of age (Allison, Hume, Wood, & Goff, 1984; Emmerson-Hanover, Shearer, Creel, &
Dustman, 1994; Shaw & Cant, 1981). In our data, this seems to occur earlier, because the P1
peak appears already adult-like in the older adolescent group. Nonetheless, other studies af-
firm not showing reliable change across young age (Mahajan & McArthur, 2012; Snyder,
Dustman, & Shearer, 1981). Here, amplitude decreases also concern the P2 component,
which will be further discussed in relationship with the corresponding microstate.
In sum, the ongoing amplitude shrinking is coherent with the literature although stabilization
seems to occur earlier, which might be linked to stimulus processing, thus not observed in the
response-locked amplitudes.
Microstates
As for amplitudes, topographic differences across groups are observed only in early time-
windows. The first topographic differences observed on the averaged ERPs appeared in
the P1 time-window. In this time-window, the topographic map that was typically specific
to the children’s signal (Map C in Figure 4) showed decreased presence with age, and young
adolescents differed from the rest of the groups because they displayed the most consistent
presence of the microstate corresponding to Map B in Figure 4. The presence of a different
P1 microstate in children as compared to adults has been reported previously (Holcomb,
et al., 1992; Laganaro, et al., 2015; Shaywitz et al., 2007) and has been attributed to more
bilateral occipital activation in children.
Microstates also differed across groups in the N1-like time-window. Here again, children are
the only group statistically different from the other 3 groups (Map D and E). Children are actually
the only group in which an N1 map is consistently present (Maps D and E), although it is tempo-
rally shifted by the delay of the P1 latency described in the previous section. Interindividual
inconsistency in the N1 time-window in picture-naming tasks has been previously reported in
(young) adults (see for instance Laganaro, 2017, on a large group of participants). Here, the results
show that such inconsistency is already present in the young adolescent group (i.e., from the age
of 14). The N1/N170 component has been associated with conceptual processes for pictorial
stimuli (Schendan & Kutas, 2003). It has been shown to be modulated by the category of the
picture (Thorpe, 2009), and this modulation could be due, among other factors, to the expertise
level of subjects for a given category (Taylor, Batty, & Itier, 2004). The present results suggest
that picture-to-concept processes are different in children only and are already adult-like in
younger adolescents. Such modulation across ages may reflect either changes in visual-conceptual
processes, which are not specific to picture naming, or visuo-conceptual activation that specif-
ically guides lexical selection.
The P2 peak, the second component most likely underlying Levelt’s (2001) concept-to-lexical
processes (selection of the lemma), confirmed by ERP studies on picture naming (Aristei, Melinger,
& Abdel Rahman, 2011; Laganaro et al., 2015; Laganaro, Valente, & Perret, 2012; Maess,
Friederici, Damian, Meyer, & Levelt, 2002; Strijkers et al., 2010), also showed differences be-
tween children and the older groups, in terms of latencies and underlying topographies (Map H
in children and G in the other groups). Whereas different latencies in children can be the
consequence of previous shifts (P1 and N1), there is a significant change in the underlying
microstate already between children and young adolescents, whose ERP signal is better explained
by the same map that is in the other older groups. The present results in children differ from
Neurobiology of Language
15
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
previous results in Laganaro, et al. (2015), where the same P2 Map was observed in children and
adults. The two P2 topographies (Maps G and H) are however very close and highly correlated
(95%). The younger adolescents display the adult P2—Map G. Actually, as already observed on
the P1 map, the P2 “adult” map is also more consistent in young adolescents than in older
adolescents and in adults. In general, along the whole signal, young adolescents seem indeed
to present considerably different topographies from older adolescents and adults, although their
amplitudes remain intermediate.
No significant between-group differences are observed beyond the P2 map in terms of presence.
There are hence only differences in duration on the rest of the microstates, which will be discussed
in further detail in relationship with the AoA effect.
Dynamics of Word Planning
The manipulation of the AoA was aimed at pinpointing lexical-phonological processes (Perret
et al., 2014), and thus, at interpreting the time-course of different encoding processes from child-
hood to adolescence and to adulthood. Amplitudes and spatio-temporal analyses converged in
identifying that AoA consistently modulated ERPs after 430 ms and mainly in the response-locked
ERPs, except for an auxiliary time-window around the P1 component. Crucially, there was no
interaction with groups, showing that AoA affects the same time-windows at all ages. While
the P1 modulation is very likely due to different picture properties across early- and late-acquired
words (see Perret & Laganaro, 2012, for similar results), the later AoA effect likely reflects the time-
window of word-form encoding. The large time-window modulated by AoA also corresponds to
the period in which no significant differences appeared on ERPs across groups on either ampli-
tudes or topographies.
As highlighted above, qualitative differences between groups appear mainly for children in the
P1-N1-P2 time-window, whereas the corresponding microstates are already adult-like in the
signal of the younger adolescents. On the other hand, amplitudes are still intermediate between
children and adults in the young adolescents in these time-windows. According to the results of
the AoA effect, as well as to previous estimates on the time-course of word production in picture
naming (Indefrey, 2011), these early time-windows correspond to prelexical (visual and concep-
tual; P1 component), and to word (lemma) retrieval (P2 component). These results suggest that
brain processes are the same for all groups for lexical and postlexical word-encoding processes
and that visual-to-conceptual, and possibly conceptual-to-lexical, encoding differs only in chil-
dren before the age of 14, at least in the sequential approaches to word planning processes. A
different interpretation would stem in the framework of theories claiming a parallel activation of
lexical and phonological information (Miozzo et al., 2014; Strijkers et al., 2017). In such
accounts, lexical-semantic and phonological-articulatory processes emerge together rapidly
(i.e., the firing hypothesis), drawing in parallel on temporal and frontal cortex around 150–200 ms
after the presentation of the picture. In this interpretation, the differences between groups may
reflect different patterns of information flow between children and adults, either serial versus
parallel processes or different degrees of overlap between serial processes.
The intermediate amplitudes in these same time-windows in the 14 to 16 year olds reflect
either the ongoing brain maturation as discussed above or an increased recruitment of the same
network as in adults, to deliver the same performance on the same task. Visually mediated pro-
cesses around 100 ms after stimulus onset, and supposedly lexical-semantic selection processes
around 300 ms, are then undergoing changes. These brain processes do not seem to develop at
the same pace (visual processes being already wired among young adolescents before other
later-developing processes, possibly related to lexical-semantic encoding), and they seem to
Neurobiology of Language
16
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
recruit different brain regions. The middle temporal gyrus, and partly, the inferior frontal gyrus,
are allegedly involved in the lexical-semantic processes (Indefrey & Levelt, 2004). The develop-
mental curve of these two regions is not analogous, with the cortical gray matter of the temporal
cortex peaking at 16 years old (Giedd et al., 1999; Lenroot & Giedd, 2006), which may explain
the ongoing maturation. In fact, when looking back at the question of when children become
adult-like, there is no strong assumption that all processes should mature at once, but rather they
should be gradual because different processes involve different brain regions with some pro-
cesses becoming adult-like before others.
Conclusions
The present study analyzed the waveform amplitudes and the periods of stable electric fields
(topographies) in the ERP signal from picture onset to the individual onset of articulation in
children, adolescents, and adults. This allowed us to track simultaneously the behavioral, func-
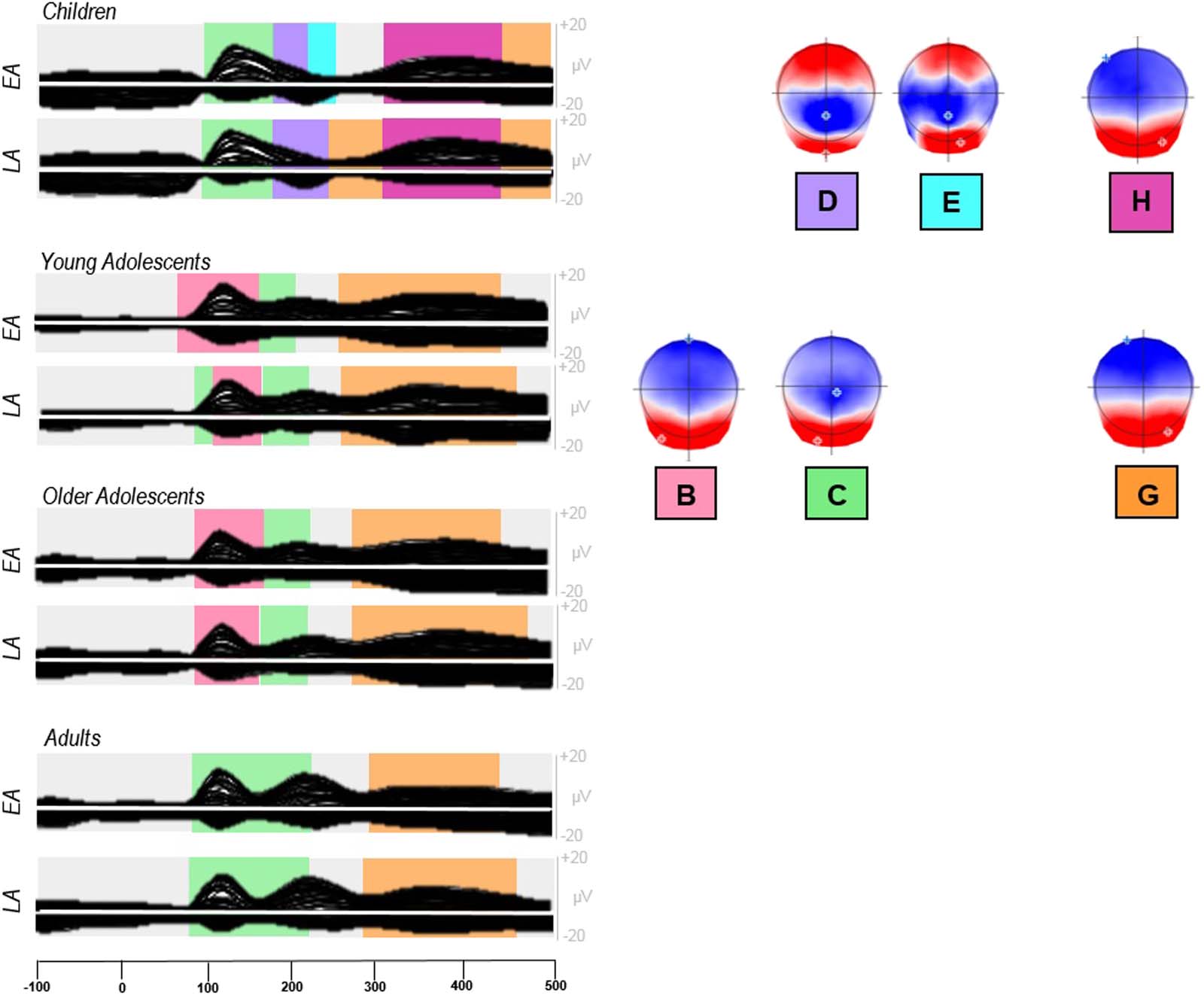
tional, and temporal changes in the development of the word-encoding processes. Our results
show that children are outliers compared to the older participants in all measures (in terms of
naming latency and accuracy, and in ERP amplitudes, topography, and morphology; see
Figure 2 and Figure 5). Moreover, adolescents from the age of 17 years old seem to have reached,
not only behaviorally, but also temporally and functionally, adult-like brain activation. Young
adolescents (aged 14 to 16), although being closer to adults than to children in terms of processing
speed and microstates, still display intermediate results on accuracy and ERP amplitudes. An
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Group-averaged event-related potentials for each group (children, young adolescents, older adolescents, adults) and condition (EA
& LA) isolating and only showing the topographical maps of interest. EA: early-acquired words, LA: late-acquired words.
Neurobiology of Language
17
Word production in the transition from adolescence to adulthood
important shift in brain signals and in referential word production processing speed seems thus to
occur between the ages of 12 and 14. This period should therefore be further investigated with
improved granularity and on additional word production tasks.
ACKNOWLEDGMENTS
This work was supported by Swiss National Science Foundation (SNSF) grant no. 100014_165647.
FUNDING INFORMATION
Marina Laganaro, Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung
(http://dx.doi.org/10.13039/501100001711), Award ID: 100014_165647.
AUTHOR CONTRIBUTIONS
Tanja Atanasova: Conceptualization, Methodology, Validation, Data curation, Formal analysis,
Investigation, Writing – Original draft, Visualization. Raphaël Fargier: Conceptualization,
Methodology, Writing – Review & editing. Pascal Zesiger: Conceptualization, Methodology,
Writing – Review & editing, Funding acquisition. Marina Laganaro: Conceptualization,
Methodology, Validation, Resources, Writing – Review & editing, Supervision, Project adminis-
tration, Funding acquisition.
REFERENCES
Alario, F. X., & Ferrand, L. (1999). A set of 400 pictures standard-
ized for French: Norms for name agreement, image agreement,
familiarity, visual complexity, image variability, and age of acquisi-
tion. Behavior Research Methods, Instruments, & Computers, 31,
531–552. DOI: https://doi.org/10.3758/BF03200732, PMID:
10502875
Alemán-Gómez, Y., Janssen, J., Schnack, H., Balaban, E., Pina-
Camacho, L., Alfaro-Almagro, F., … Desco, M. (2013). The
human cerebral cortex flattens during adolescence. The Journal
of Neuroscience, 33(38), 15004–15010. DOI: https://doi.org
/10.1523/JNEUROSCI.1459-13.2013, PMID: 24048830, PMCID:
PMC6618418
Allison, T., Hume, A. L., Wood, C. C., & Goff, W. R. (1984).
Developmental and aging changes in somatosensory, auditory
and visual evoked potentials. Electroencephalography and Clinical
Neurophysiology, 58, 14–24. DOI: https://doi.org/10.1016/0013
-4694(84)90196-2
A r i s t e i , S . , Me l i n g e r , A ., & A b d e l R a h m a n , R .
( 2 0 1 1 ) .
Electrophysiological chronometry of semantic context effects in
language production. Journal of Cognitive Neuroscience, 23(7),
1567–1586. DOI: https://doi.org/10.1162/jocn.2010.21474,
PMID: 20515409
Baayen, R. H. (2008). Analyzing linguistic data: A practical introduc-
tion to statistics using R. Cambridge, UK: Cambridge University
Press. DOI: https://doi.org/10.1017/CBO9780511801686
Batty, M., & Taylor, M. J. (2006). The development of emotional
face processing during childhood. Developmental Science, 9(2),
207–220. DOI: https://doi.org/10.1111/j.1467-7687.2006
.00480.x, PMID: 16472321
Bonin, P., Peerman, R., Malardier, N., Méot, A., & Chalard, M.
(2003). A new set of 299 pictures for psycholinguistic studies:
French norms for name agreement, image agreement, conceptual
familiarity, visual complexity, image variability, age of acquisition,
and naming latencies. Behavior Research Methods, Instruments, &
Computers, 35, 158–167. DOI: https://doi.org/10.3758/BF03195507,
PMID: 12723790
Brown, T. T., Lugar, H. M., Coalson, R. S., Miezin, F. M., Petersen,
S. E., & Schlaggar, B. L. (2005). Developmental changes in human
cerebral functional organization for word generation. Cerebral
Cortex, 15, 275–290. DOI: https://doi.org/10.1093/cercor
/bhh129, PMID: 15297366
Brunet, D., Murray, M. M., & Michel, C. M. (2011). Spatiotemporal
analysis of multichannel EEG: CARTOOL. Computational
Intelligence and Neuroscience, 2011, 813870. DOI: https://doi
.org/10.1155/2011/813870, PMID: 21253358, PMCID:
PMC3022183
Budd, M.-J., Paulmann, S., Barry, C., & Clahsen, H. (2013). Brain
potentials during language production in children and adults: An
ERP study of the English past tense. Brain and Language, 127(3),
345–355. DOI: https://doi.org/10.1016/j.bandl.2012.12.010,
PMID: 23398779
Campbell, J., & Sharma, A. (2016). Distinct visual evoked potential
morphological patterns for apparent motion processing in
school-aged children. Frontiers in Human Neuroscience, 10.
DOI: https://doi.org/10.3389/fnhum.2016.00277, PMID:
27445738, PMCID: PMC4923113
Cardebat, D., Doyon, B., Puel, M., Goulet, P., & Joanette, Y. (1990).
[Formal and semantic lexical evocation in normal subjects.
Performance and dynamics of production as a function of sex,
age and educational level.] Acta Neurologica Belgica, 90(4),
207–217.
Chalard, M., Bonin, P., Méot, A., Boyer, B., & Fayol, M. (2003).
Objective age-of-acquisition (AoA) norms for a set of 230 object
names in French: Relationships with psycholinguistic variables,
the English data from Morrison et al. (1997), and naming laten-
cies. European Journal of Cognitive Psychology, 15(2), 209–245.
DOI: https://doi.org/10.1080/09541440244000076
Changeux, J. P., & Michel, C. M. (2004). Mechanisms of neural
integration at the brain scale level: The neuronal workspace
and microstate models. In S. Grillner, & A. M. Grabyel (Eds.),
Microcircuits: The interface between neurons and global brain
function. Cambridge, MA: MIT Press.
Neurobiology of Language
18
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
Chauveau, N., Franceries, X., Doyon, B., Rigaud, B., Morucci, J. P., &
Celsis, P. (2004). Effects of skull thickness, anisotropy, and inhomo-
geneity on forward EEG/ERP computations using a spherical three-
dimensional resistor mesh model. Human Brain Mapping, 212,
86–97. DOI: https://doi.org/10.1002/hbm.10152, PMID:
14755596, PMCID: PMC6872130
Clark, E. V. (1993). The lexicon in acquisition. Cambridge, UK:
Cambridge University Press. DOI: https://doi.org/10.1017
/CBO9780511554377
Cragg, L., Kovacevic, N., McIntosh, A. R., Poulsen, C., Martinu, K.,
Leonard, G., & Paus, T. (2011). Maturation of EEG power spectra
in early adolescence: A longitudinal study. Developmental Science,
14, 935–943. DOI: https://doi.org/10.1111/j.1467-7687
.2010.01031.x, PMID: 21884309
Creel, D. J. (2019). Visually evoked potentials. In Handbook of
Clinical Neurology, 160, 501–522. DOI: https://doi.org/10.1016
/B978-0-444-64032-1.00034-5, PMID: 31277872
Cycowicz, Y. M., Friedman, D., Rothstein, M., & Snodgrass, J. G.
(1997). Picture naming by young children: Norms for name agree-
ment, familiarity, and visual complexity. Journal of Experimental
Child Psychology, 65, 171–237. DOI: https://doi.org/10.1006
/jecp.1996.2356, PMID: 9169209
Dale, P. S., & Fenson, L. (1996). Lexical development norms for
young children. Behavior Research Methods, Instruments, &
Computers, 28, 125–127. DOI: https://doi.org/10.3758/BF03203646
D’Amico, S., Devescovi, A., & Bates, E. (2001). Picture naming and
lexical access in Italian children and adults. Journal of Cognition &
Development, 2(1), 71–105. DOI: https://doi.org/10.1207
/S15327647JCD0201_4
Dell, G. S. (1986). A spreading-activation theory of retrieval in sen-
tence production. Psychological Review, 93, 283–321. DOI:
https://doi.org/10.1037/0033-295X.93.3.283, PMID: 3749399
Dumontheil, I. (2016). Adolescent brain development. Current
Opinion in Behavioral Sciences, 10, 39–44. DOI: https://doi
.org/10.1016/j.cobeha.2016.04.012
Durston, S., & Casey, B. J. (2006). What have we learned about
cognitive development from neuroimaging? Neuropsychologia,
44, 2149–2157. DOI: https://doi.org/10.1016/j.neuropsychologia
.2005.10.010, PMID: 16303150
Emmerson-Hanover, R., Shearer, D. E., Creel, D. J., & Dustman, R. E.
(1994). Pattern reversal evoked potentials: Gender differences
a n d a g e r e l a t e d c h a n g e s i n a m p l i t u d e a n d l a t e n c y .
Electroencephalography & Clinical Neurophysiology, 92, 93–101.
DOI: https://doi.org/10.1016/0168-5597(94)90049-3
Fargier, R., Buerki, A., Pinet, S., Alario, F.-X., & Laganaro, M.
(2018). Word onset phonetic properties and motor artifacts in
speech production EEG recordings. Psychophysiology, 55(2).
DOI: https://doi.org/10.1111/psyp.12982, PMID: 28850684
Fargier, R., & Laganaro, M. (2016). Neurophysiological modula-
tions of non-verbal and verbal dual-tasks interference during
word planning. PLOS ONE, 11(12), e0168358. DOI: https://doi
.org/10.1371/journal.pone.0168358, PMID: 27992586, PMCID:
PMC5167377
Giedd, J. N., Blumenthal, J., Jeffries, N. O., Castellanos, F. X., Lju,
H., Zijdenbos, A., … Rapoport, J. L. (1999). Brain development
during childhood and adolescence: A longitudinal MRI study.
Nature Neuroscience, 2, 861–863. DOI: https://doi.org
/10.1038/13158, PMID: 10491603
Goossens, T., Vercammen, C., Wouters, J., & van Wieringen, A.
(2016). Aging affects neural synchronization to speech-related
acoustic modulations. Frontiers in Aging Neuroscience, 8, 133.
DOI: https://doi.org/10.3389/fnagi.2016.00133, PMID:
27378906, PMCID: PMC4908923
Greenham, S. L., & Stelmack, R. M. (2001). Event-related potentials
and picture-word naming: Effects of attention and semantic relation
for children and adults. Developmental Neuropsychology, 20,
619–638. DOI: https://doi.org/10.1207/S15326942DN2003_5,
PMID: 12002097
Holcomb, P. J., Coffey, S. A., & Neville, H. J. (1992). Visual and
auditory sentence processing: A developmental analysis using
event-related brain potentials. Developmental Neuropsychology,
8 ( 2–3 ) , 2 0 3–2 4 1 . D O I : h t t p s : / / d o i . o r g / 1 0 . 1 0 8 0
/87565649209540525
Hopfinger, J. B., & Mangun, G. R. (1998). Reflexive attention modu-
lates processing of visual stimuli in human extrastriate cortex.
Psychological Science, 9, 441–447. DOI: https://doi.org/10.1111
/1467-9280.00083, PMID: 26321798, PMCID: PMC4552358
Indefrey, P. (2011). The spatial and temporal signatures of word
production components: A critical update. Frontiers in Psychology,
2, 255. DOI: https://doi.org/10.3389/fpsyg.2011.00255, PMID:
22016740, PMCID: PMC3191502
Indefrey, P., & Levelt, W. J. M. (2004). The spatial and temporal
signatures of word production components. Cognition, 92,
101–144. DOI: https://doi.org/10.1016/j.cognition.2002.06.001,
PMID: 15037128
Itier, R. J., & Taylor, M. J. (2004). Source analysis of the N170 to
faces and objects. NeuroReport, 15(8), 1261–1265. DOI:
https://doi.org/10.1097/01.wnr.0000127827.73576.d8, PMID:
15167545
Jaeger, T. F. (2008). Categorical data analysis: Away from ANOVAs
(transformation or not) and towards logit mixed models. Journal
of Memory and Language, 59(4), 434–446. DOI: https://doi.org/
10.1016/j.jml.2007.11.007, PMID: 19884961, PMCID:
PMC2613284
Johnston, R. A., & Barry, C. (2006). Repetition priming of access to
biographical information from faces. Quarterly Journal of
Experimental Psychology, 59(2). DOI: https://doi.org/10.1080
/02724980443000791, PMID: 16618637
Juhasz, B. J. (2005). Age-of-acquisition effects in word and picture
identification. Psychological Bulletin, 131(5), 684–712. DOI:
https://doi.org/10.1037/0033-2909.131.5.684, PMID: 16187854
Koenig, T., & Melie-García, L. (2010). A method to determine the
presence of averaged event-related fields using randomization
tests. Brain Topography, 23, 233–242. DOI: https://doi.org
/10.1007/s10548-010-0142-1, PMID: 20376546
Koukou, M., & Lehmann, D. (1987). An Information processing
perspective of psychophysiological measurements. Journal of
Psychophysiology, 1, 109–112.
Krishnan, S., Leech, R., Mercure, E., Lloyd-Fox, S., & Dick, F.
(2014). Convergent and divergent fMRI responses in children and
adults to increasing language production demands. Cerebral
Cortex, 25(10). DOI: https://doi.org/10.1093/cercor/bhu120,
PMID: 24907249, PMCID: PMC4585486
Laganaro, M. (2014). ERP topographic analyses from concept to artic-
ulation in word production studies. Frontiers in Psychology, 5, 493.
DOI: https://doi.org/10.3389/fpsyg.2014.00493, PMID: 24904505,
PMCID: PMC4034040
Laganaro, M. (2017). Inter-study and inter-individual consistency
and variability of EEG/ERP microstate sequences in referential
word production. Brain Topography, 30(6), 785–796. DOI:
https://doi.org/10.1007/s10548-017-0580-0, PMID: 28770364
Laganaro, M., & Perret, C. (2011). Comparing electrophysiological
correlates of word production in immediate and delayed naming
through the analysis of word age of acquisition effects. Brain
Topography, 24, 19–29. DOI: https://doi.org/10.1007/s10548
-010-0162-x, PMID: 20938730
Neurobiology of Language
19
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
Laganaro, M., Tzieropoulos, H., Fraunfelder, U. H., & Zesiger P. (2015).
Functional and time-course changes in single word production
from childhood to adulthood. NeuroImage, 111, 204–214. DOI:
https://doi.org/10.1016/j.neuroimage.2015.02.027, PMID:
25703828
Laganaro, M., Valente, A., & Perret, C. (2012). Time course of word
production in fast and slow speakers: A high density ERP topo-
graphic study. NeuroImage, 59, 3881–3388. DOI: https://doi
.org/10.1016/j.neuroimage.2011.10.082, PMID: 22079505
Lehmann, D., & Skrandies, W. (1984). Spatial analysis of evoked
potentials in man: A review. Progress in Neurobiology, 23(3),
227–250. DOI: https://doi.org/10.1016/0301-0082(84)90003-0,
PMID: 6395186
Lehmann, D., Strik, W. K., Henggeler, B., Koenig, T., & Koukkou, M.
(1998). Brain electric microstates and momentary conscious mind
states as building blocks of spontaneous thinking: I. Visual imagery
and abstract thoughts. International Journey Psychophysiology, 29,
1–11. DOI: https://doi.org/10.1016/S0167-8760(97)00098-6
Lenroot, R. K., & Giedd, J. N. (2006). Brain development in children
and adolescents: Insights from anatomical magnetic resonance
imaging. Neuroscience & Biobehavioral Reviews, 30(6), 718–729.
DOI: https://doi.org/10.1016/j.neubiorev.2006.06.001, PMID:
16887188
Levelt, W. J. M. (1989). Speaking: From intention to articulation.
Cambridge, MA: MIT Press.
Levelt, W. J. M. (2001). Spoken word production: A theory of
lexical access. Proceedings of the National Academy of Sciences
of the United States of America, 98(23), 13464–13471. DOI:
https://doi.org/10.1073/pnas.231459498, PMID: 11698690,
PMCID: PMC60894
Luck, S. J., Woodman, G. F., & Vogel, E. K. (2000). Event-related
potential studies of attention. Trends in Cognitive Sciences, 4,
432–440. DOI: https://doi.org/10.1016/S1364-6613(00)01545-X,
PMID: 11058821
Mahajan, Y., & McArthur, G. (2012). Maturation of visual evoked
potentials across adolescence. Brain & Development, 34, 655–666.
DOI: https://doi.org/10.1016/j.braindev.2011.10.009, PMID:
22105091
Maess, B., Friederici, A. D., Damian, M. F., Meyer, A. S., & Levelt,
W. J. M. (2002). Semantic category interference in overt picture
naming. Journal of Cognitive Neuroscience, 14(3), 455–462.
DOI: https://doi.org/10.1162/089892902317361967, PMID:
11970804
Miller, G. A. (1996). The science of words. New York: Scientific
American Library.
Miozzo, M., Pulvermüller, F., & Hauk, O. (2014). Early parallel acti-
vation of semantics and phonology in picture naming: Evidence
from a multiple linear regression MEG study. Cerebral Cortex,
25. DOI: https://doi.org/10.1093/cercor/bhu137, PMID:
25005037, PMCID: PMC4585490
Miskovic, V., Ma, X., Chou, C.-A., Fan, M., Owens, M., Sayama, H.,
& Gibb, B. E. (2015). Developmental changes in spontaneous
electrocortical activity and network organization from early to late
childhood. NeuroImage, 118, 237–247. DOI: https://doi.org
/10.1016/j.neuroimage.2015.06.013, PMID: 26057595, PMCID:
PMC4554821
Morrison, C. M., Chappell, T. D., & Ellis, W. A. (1997). Age of
acquisition norms for a large set of object names and their relation
to adult estimates and other variables. Quarterly Journal of
Experimental Psychology, 50A, 528–559. DOI: https://doi.org
/10.1080/027249897392017
Munding, D., Dubarry, A.-S., & Alario, F.-X. (2016). On the cortical
dynamics of word production: A review of the MEG evidence.
Language, Cognition and Neuroscience, 31(4), 441–462. DOI:
https://doi.org/10.1080/23273798.2015.1071857
Murray, M. M., Brunet, D., & Michel, C. (2008). Topographic ERP
analyses: A step-by-step tutorial review. Brain Topography, 20,
249–269. DOI: https://doi.org/10.1007/s10548-008-0054-5,
PMID: 18347966
New, B., Pallier, C., Brysbaert, M., & Ferrand, L. (2004). Lexique 2:
A new French lexical database. Behavior Research Methods,
Instruments, & Computers, 36(3), 516–524. DOI: https://doi.org
/10.3758/BF03195598, PMID: 15641440
Perret, C., Bonin, P., & Laganaro, M. (2014). Exploring the multiple-
loci hypothesis of AoA effects in spoken and written object nam-
ing using a topographic ERP analysis. Brain and Language, 135,
20–31. DOI: https://doi.org/10.1016/j.bandl.2014.04.006,
PMID: 24887390
Perret, C., & Laganaro, M. (2012). Comparison of electrophysiologi-
cal correlates of writing and speaking: A topographic ERP analysis.
Brain Topography, 25, 64–72. DOI: https://doi.org/10.1007
/s10548-011-0200-3, PMID: 21863371
Perrin, F., Pernier, J., Bertrand, O., Giard, M. H., & Echallier, J. F.
(1987). Mapping of scalp potentials by surface spline interpolation.
Electroencephalogry and Clinical Neurophysiology, 66, 75–81.
DOI: https://doi.org/10.1016/0013-4694(87)90141-6
Picton, T. W., & Taylor, M. J. (2007). Electrophysiological evaluation of
human brain development. Developmental Neuropsychology, 31,
249–278. DOI: https://doi.org/10.1080/87565640701228732,
PMID: 17559326
Protopapas, A. (2007). CheckVocal: A program to facilitate checking
the accuracy and response time of vocal responses from DMDX.
Behavior Research Methods, 39, 859–862. DOI: https://doi.org
/10.3758/BF03192979, PMID: 18183901
R Development Core Team. (2005). A language and environment for
statistical computing. Vienna, Austria: R Foundation for Statistical
Computing.
Riley, J. D., Chen, E. E., Winsell, J., Poggi Davis, E., Glynn, L. M.,
Baram, T. Z., … Solodkin, A. (2018). Network specialization
during adolescence: Hippocampal effective connectivity in boys
and girls. NeuroImage, 175, 402–412. DOI: https://doi.org
/10.1016/j.neuroimage.2018.04.013, PMID: 29649560, PMCID:
PMC5978413
Rossion, B., & Caharel, S. (2011). ERP evidence for the speed of
face categorization in the human brain: Disentangling the contri-
bution of low-level visual cues from face perception. Vision
Research, 51(12), 1297–1311. DOI: https://doi.org/10.1016
/j.visres.2011.04.003, PMID: 21549144
Schendan, H. E., & Kutas, M. (2003). Time course of processes and
representations supporting visual object identification and memory.
Journal of Cognitive Neuroscience, 15, 111–135. DOI: https://doi
.org/10.1162/089892903321107864, PMID: 12590847
J., Santesso, D. L.,
(2010).
Electrophysiological changes during adolescence: A review.
Brain and Cognition, 72, 86–100. DOI: https://doi.org/10.1016
/j.bandc.2009.10.003, PMID: 19914761
Segalowitz, S.
Jetha, M. K.
Shaw, N. A., & Cant, B. R. (1981). Age-dependent changes in the
t h e p a t t e r n v i s u a l e v o k e d p o t e n t i a l .
a m p l i t u d e o f
Electroencephalography and Clinical Neurophysiology, 51,
671–673. DOI: https://doi.org/10.1016/0013-4694(81)90212-1,
PMID: 6165569
Shaywitz, B. A., Skudlarski, P., Holahan, J. M., Marchione, K. E.,
Constable, R. T., … Shaywitz, S. E. (2007). Age-related changes
in reading systems of dyslexic children. Annals of Neurology, 61,
363–370. DOI: https://doi.org/10.1002/ana.21093, PMID:
17444510
Neurobiology of Language
20
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Word production in the transition from adolescence to adulthood
Snyder, E. W., Dustman, R. E., & Shearer, D. E. (1981). Pattern
reversal evoked potential amplitudes: Life span changes.
Electroencephalography and Clinical Neurophysiology, 52,
429–434. DOI: https://doi.org/10.1016/0013-4694(81)90026-2,
PMID: 6171408
Strijkers, K., Costa, A., & Pulvermuller, F. (2017). The cortical
dynamics of speaking: Lexical and phonological knowledge
simultaneously recruit the frontal and temporal cortex within
200 ms. NeuroImage, 163, 206–219. DOI: https://doi.org
/10.1016/j.neuroimage.2017.09.041, PMID: 28943413
Strijkers, K., Costa, A., & Thierry, G. (2010). Tracking lexical access
in speech production: Electrophysiological correlates of word
frequency and cognate effects. Cerebral Cortex, 20, 912–928.
DOI: https://doi.org/10.1093/cercor/bhp153, PMID: 19679542
Taylor, M. J., Batty, M., & Itier, R. J. (2004). The faces of development:
A review of early face processing over childhood. Journal of
Cognitive Neuroscience, 16(8), 1426–1442. DOI: https://doi.org
/10.1162/0898929042304732, PMID: 15509388
Taylor, M. J., Edmonds, G. E., McCarthy, G., & Allison, T. (2001).
Eyes first! Eye processing develops before face processing in
children. NeuroReport, 12(8), 1671–1676. DOI: https://doi.org
/10.1097/00001756-200106130-00031, PMID: 11409737
Thorpe, S. J. (2009). The speed of categorization in the human visual
system. Neuron, 62, 168–170. DOI: https://doi.org/10.1016
/j.neuron.2009.04.012, PMID: 19409262
Wechsler, D. (1997). Wechsler adult intelligence scale (3rd ed.).
San Antonio, TX: Psychological Corporation. DOI: https://doi
.org/10.1037/t49755-000
Xue, J., Liu, T., Marmolejo-Ramos, F., & Pei, X. (2017). Age of
acquisition effects on word processing for Chinese native learners’
English: ERP evidence for the arbitrary mapping hypothesis.
Frontiers in Psychology, 8. DOI: https://doi.org/10.3389/fpsyg
.2017.00818, PMID: 28572785, PMCID: PMC5435808
Yadav, R., Poudel, B., Limbu, N., Thakur, D., & Yadav, S. (2015).
Normative data of visual evoked potential in children and corre-
lation with age. Asian Journal of Medical Sciences, 7(2), 39–43.
Zhu, J., & Weiss, L. (2005). The Wechsler Scales. In D. P. Flanagan
& P. L. Harrison (Eds.), Contemporary intellectual assessment:
Theories, tests, and issues (pp. 297–324). New York: The Guilford
Press.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
/
2
1
1
1
8
9
9
6
6
0
n
o
_
a
_
0
0
0
2
4
p
d
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
21