RESEARCH ARTICLE
Linking Lysosomal Enzyme Targeting Genes and
Energy Metabolism with Altered Gray Matter
Volume in Children with Persistent Stuttering
Ho Ming Chow1,2,3, Emily O. Garnett3
Dennis Drayna5
, Diane Chugani1, and Soo-Eun Chang3,6,7
, Hua Li2, Andrew Etchell3, Jorge Sepulcre4,
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
哦
_
A
_
0
0
0
1
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
1Department of Communication Sciences and Disorders, University of Delaware, Newark, DE
2Katzin Diagnostic & Research PET/ MRI Center, Nemours/Alfred I. duPont Hospital for Children, Wilmington, DE
3Department of Psychiatry, 密歇根大学, 安娜堡, MI
4Gordon Center for Medical Imaging, 放射科, Massachusetts General Hospital,
哈佛医学院, 波士顿, 嘛
5National Institute on Deafness and Other Communication Disorders, NIH, Bethesda, 医学博士
6Cognitive Imaging Research Center, 放射科, Michigan State University, East Lansing, MI
7Department of Communicative Sciences and Disorders, Michigan State University, East Lansing, MI
关键词: longitudinal, lysosome, metabolism, stuttering, voxel-based morphometry
抽象的
Developmental stuttering is a childhood onset neurodevelopmental disorder with an unclear
etiology. Subtle changes in brain structure and function are present in both children and adults
who stutter. It is a highly heritable disorder, and 12–20% of stuttering cases may carry a
mutation in one of four genes involved in intracellular trafficking. To better understand the
relationship between genetics and neuroanatomical changes, we used gene expression data
from the Allen Institute for Brain Science and voxel-based morphometry to investigate the
spatial correspondence between gene expression patterns and differences in gray matter
volume between children with persistent stuttering (n = 26, 和 87 scans) and their fluent
同行 (n = 44, 和 139 scans). We found that the expression patterns of two stuttering-related
基因 (GNPTG and NAGPA) from the Allen Institute data exhibited a strong positive spatial
correlation with the magnitude of between-group gray matter volume differences. 额外的
gene set enrichment analyses revealed that genes whose expression was highly correlated with
the gray matter volume differences were enriched for glycolysis and oxidative metabolism
in mitochondria. Because our current study did not examine the participants’ genomes,
these results cannot establish the direct association between genetic mutations and gray
matter volume differences in stuttering. 然而, our results support further study of the
involvement of lysosomal enzyme targeting genes, as well as energy metabolism in
stuttering. Future studies assessing variations of these genes in the participants’ genomes may
lead to increased understanding of the biological mechanisms of the observed spatial
relationship between gene expression and gray matter volume.
介绍
Fluid, effortless speech production forms the basis for communication and is considered a fun-
damental human ability. Stuttering significantly disrupts fluent speech production, often lead-
ing to negative psychosocial and economic consequences throughout life (Blumgart, 特兰, &
Craig, 2010; Craig, Blumgart, & 特兰, 2009; Yaruss, 2010). Developmental stuttering typically
开放访问
杂志
引文: Chow, H. M。, 加内特, 乙. 奥。, 李,
H。, Etchell, A。, 墓, J。, Drayna, D .,
Chugani, D ., & 张, S.-E. (2020).
Linking lysosomal enzyme targeting
genes and energy metabolism with
altered gray matter volume in children
with persistent stuttering.
Neurobiology of Language, 1(3),
365–380. https://doi.org/10.1162/
nol_a_00017
DOI:
https://doi.org/10.1162/nol_a_00017
支持信息:
https://doi.org/10.1162/nol_a_00017
已收到: 24 可能 2019
公认: 13 可能 2020
利益争夺: 作者有
声明不存在竞争利益
存在.
通讯作者:
Ho Ming Chow
hmc@udel.edu
处理编辑器:
Simon Fisher
版权: © 2020 马萨诸塞州
Institute of Technology. 已发表
under a Creative Commons Attribution
4.0 国际的 (抄送 4.0) 执照.
麻省理工学院出版社
Genetics and anomalous neuroanatomy in stuttering
has an onset in early childhood, affecting more than 5% of preschool-age children and persist-
ing in about 1% of adults (Craig, Hancock, 特兰, Craig, & Peters, 2002; Månsson, 2000; Yairi,
& Ambrose, 2013). Persistent stuttering is highly heritable, with estimations of genetic contri-
bution exceeding 80% in some studies (Dworzynski, Remington, Rijsdijk, 豪厄尔, & Plomin,
2007; Fagnani, Fibiger, Skytthe, & Hjelmborg, 2011; Ooki, 2005; Rautakoski, Hannus,
Simberg, Sandnabba, & Santtila, 2012; van Beijsterveldt, Felsenfeld, & Boomsma, 2010).
Genes causative of persistent stuttering have begun to be identified (Kang et al., 2010; Raza
等人。, 2015). 迄今为止, four such genes, designated GNPTG, GNPTAB, NAGPA, and AP4E1
have been found, and they may cumulatively account for 12–20% of unrelated individuals
with persistent stuttering (see Frigerio-Domingues & Drayna, 2017 for a comprehensive
review).
This group of genes is known to play a role in lysosomal enzyme trafficking. GNPTG,
GNPTAB, and NAGPA are involved in marking lysosomal hydrolases and several nonlysoso-
mal proteins with a mannose-6-phosphate (M6P) tag that is important for intracellular traffick-
英 (Barnes et al., 2011). Homozygous mutations in GNPTG and GNPTAB genes are known to
cause the rare inherited lysosomal storage disorders Mucolipidosis types II and III (Kornfeld,
2001), which affect many parts of the body including the brain. 然而, in most of the cases,
people who stutter only carry heterozygous mutations in these genes, and do not have the
signs or symptoms typically seen in Mucolipidosis types II and III. AP4E1 is a member of a
family of adaptor proteins that are involved in vesicle formation and sorting member proteins
for transporting lysosomal enzymes from the trans Golgi network to late endosomes and lyso-
somes (Dell’Angelica, Mullins, & Bonifacino, 1999). Mutations in AP4E1 have been associated
with hereditary spastic paraplegia and cerebral palsy (Abou Jamra et al., 2011; Kong et al.,
2013). Why mutations in these genes can specifically affect the ability to produce fluent
speech but not other cognitive or neurologic functions remains unknown. 然而, neuroim-
aging studies have shown that persistent stuttering is associated with subtle functional and an-
atomical anomalies (Chow & 张, 2017; Etchell, Civier, Ballard, & Sowman, 2018; 加内特
等人。, 2018; Neef, 安旺德, & Friederici, 2015).
The Current Study
How genetic factors relate to brain anomalies in stuttering is not yet clear. To pursue this ques-
的, we compared the differences in spatial patterns of gray matter volume (GMV) in children
with persistent stuttering with the regional expression of the four genes thus far associated with
stuttering using data provided by the Allen Institute for Brain Science (AIBS; http://www.brain-
map.org/). This approach has been used to reveal gene–brain relationships in a number of
recent studies. A seminal study was published in 2015, in which the authors used gene ex-
pression data and resting-state fMRI to identify 136 genes associated with intrinsic functional
networks in the brain (Richiardi et al., 2015). In another study, Ortiz-Terán et al. (2017) used a
similar method to demonstrate that the neural reorganization in blind children measured by
resting-state functional connectivity is associated with a set of known neuroplasticity-related
基因. 而且, this approach was also employed to study neurological disorders. 例如,
McColgan et al. (2017) identified genes associated with Huntington’s disease by comparing
regional white matter loss and gene expression in patients with the disorder.
A core presumption of this current study is that the expression patterns of the genes known
to be associated with stuttering, to a certain extent, reflect the pattern of anatomical anomalies
in the disorder. While the genetic causes of stuttering are likely to be heterogeneous, 他们的
effects at the neuroanatomical level may be similar because the disorder affects speech
Neurobiology of Language
366
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
哦
_
A
_
0
0
0
1
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
Genetics and anomalous neuroanatomy in stuttering
production specifically. 而且, in support of the previous argument, neuroimaging studies
of people who stutter with unknown genetic status have shown some consistent results. 在
特别的, different groups of researchers have independently demonstrated that the aniso-
tropic diffusivity in the corpus callosum and the superior longitudinal fasciculus is decreased in
people who stutter compared with matched controls (Neef et al., 2015). A similar presumption
was made in a previous study showing that the expression of risk genes for schizophrenia were
positively correlated with the anatomical disconnectivity defined by diffusion tensor imaging
tractography in patients with schizophrenia, but not in patients with bipolar disorder (Romme,
de Reus, Ophoff, Kahn, & van den Heuvel, 2017).
In our recent study, using a similar approach, we have shown that the pattern of functional
connectivity differences between people who stutter and controls and the expression pattern of
GNPTG are spatially correlated (Benito-Aragón et al., 2019). In the current study, we further
examined whether the expression pattern of the genes associated with stuttering are correlated
with the structural differences as reflected in the pattern of regional GMV. We hypothesized
that the expression of the four stuttering-related genes from the AIBS data would be spatially
correlated with the GMV differences in children with persistent stuttering. 而且, we used
gene set enrichment analysis to identify genes whose expression is highly associated with the
GMV differences in order to explore the potential biological processes and pathways involved in
stuttering. It is important to note that in these previous studies and our current study, 因为
genomes of the participants were not studied, the spatial relationship between brain measures
and gene expression, 如果有的话, cannot provide evidence of a direct relationship between gene
expressions and brain anomalies.
材料和方法
Participants were monolingual English speakers primarily recruited from the East Lansing,
Michigan, area and within a surrounding 50-mile radius, as part of an ongoing longitudinal study
conducted at Michigan State University. Subjects were between three and ten years old when
they entered the study, and participated in 1 到 4 longitudinal visits, with an intervisit interval of
大约 12 月. All research procedures were approved by the Michigan State
University Institutional Review Board, which follows the ethical standards described in the
Belmont Report and complies with the requirements of the Federalwide Assurance for the
Protection of Human Subjects from the United States Department of Health and Human
Services. Written consent was obtained from all parents of the participating children, and assents
were obtained from all children in verbal or written format depending on reading level. 孩子们
were paid a nominal remuneration, and were given small prizes (例如, stickers) for their
参与.
Participants were recruited via printed advertisements in newspapers and magazines, parent
email listservs, study flyers, social media or online ads (例如, Facebook, Craigslist), speech-
language pathologist (SLP) referral, and word of mouth. A total of 226 children were contacted
and screened for eligibility by an SLP on the study team. Of those screened, 128 were eligible
and willing to participate in the study. Exclusion criteria for all children included (A) hearing
loss, (乙) a history of diagnosis of a developmental, psychological, neurological, or speech
紊乱, except for stuttering (例如, autism spectrum disorder), (C) currently taking medication
affecting central nervous functioning, (d) language impairment based on standardized tests (<−2
SD), (e) cognitive impairment based on standardized tests (<−2 SD), (f ) bilingualism from a
young age, (g) pregnancy, and (h) any type of metal implant or braces. Inclusion criteria for fluent
controls included (a) no history of speech disorder at any time, and (b) stuttering-like
Neurobiology of Language
367
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
o
_
a
_
0
0
0
1
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Genetics and anomalous neuroanatomy in stuttering
disfluencies (SLDs) less than 3%. For children who stutter, they must have been stuttering for
at least six months. The details of the standardized tests and behavioral evaluations are listed
below.
Standardized Tests and Speech Samples
All children were tested by trained, licensed speech language pathologists for cognitive, speech-
language, and articulation abilities through a battery of standardized assessments, including the
Wechsler Primary and Preschool Scale of Intelligence (3rd Ed.), for children 2:6–7:3 (Wechsler,
2002), the Wechsler Abbreviated Scale of Intelligence for children 7 and up (Wechsler, 1999),
the Peabody Picture Vocabulary Test (PPVT-4) for receptive vocabulary ability (Dunn & Dunn,
2007), the Expressive Vocabulary Test (EVT-2; Williams, 2007), and the Goldman-Fristoe Test of
Articulation 2 (GFTA-2; Goldman & Fristoe, 2000). The results of these tests are listed in Table 1.
Further, all children performed within normal range on the Oral Speech Mechanism Screening
Examination Revised (St Louis & Ruscello, 1987), which tests oral facial structure and function,
as well as diadochokinetic rates. Participants’ phonological processing skills were not formally
tested in this cohort, although clinicians made note of any phonological delay or processes
based on their interaction with the child during conversation and test administration. All chil-
dren were required to pass a hearing screening that was administered using an audiometer
(Beltone model 119) at pure tone frequencies 500, 1,000, 2,000, and 4,000 Hz at the 20 dB
threshold. The analyses of the above-mentioned tests were conducted according to instrument
guidelines by trained, licensed SLPs.
For study inclusion, participants had to score above −2 SD of the nominative mean on all
standardized tests. The reason for using this threshold for inclusion was to ensure that we
recruited a representative sample of children who stutter (CWS), who have been reported to
exhibit greater comorbidity of articulation and language deficits and dissociated development
among language areas (Choo, Burnham, Hicks, & Chang, 2016; Coulter, Anderson, &
Conture, 2009). Nevertheless, in our sample, none of the subjects fell at or below −2 SD on
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
o
_
a
_
0
0
0
1
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. Demographics, intelligent quotient (IQ), and language test scores averaged across longitudinal visits for each participant
Age at the first scan (years)
IQa
PPVT-4b
EVT-2c
GFTA-2d
SSI-4e
Controls n = 44 (21 boys)
Mean (SD)
Range
3.3–10.8
6.5 (2.0)
Persistent n = 26 (18 boys)
Mean (SD)
Range
3.6–10.3
6.5 (1.9)
Recovered n = 17 (9 boys)
Mean (SD)
Range
3.1–9.4
5.4 (1.9)
114 (14.1)
119 (12.7)
115 (11.8)
104 (6.6)
-
84–144
95–141
93–142
84–115
-
106 (15.5)
110 (13.5)
106 (12.2)
102 (4.2)
21 (8.3)
81–138
86–146
86–138
92–110
12–48
106 (13.1)
114 (10.3)
109 (9.2)
106 (7.3)
13 (2.9)
88–128
93–131
89–129
91–115
7–19
a Wechsler Primary and Preschool Scale of Intelligence (3rd Ed.) or Wechsler Abbreviated Scale of Intelligence). No significant difference between any two
groups (t tests, p > 0.05).
b The Peabody Picture Vocabulary Test (4th Ed.). Scores significantly higher in control than persistent groups (two-sample t tests, p < 0.05).
c The Expressive Vocabulary Test (2nd Ed.). Scores significantly higher in control than both persistent and recovered groups (two-sample t tests, p < 0.05).
d The Goldman-Fristoe Test of Articulation 2. Scores significantly higher in control than persistent groups (two-sample t tests, p < 0.05).
e Stuttering Severity Instrument. Scores significantly higher in persistent and recovered groups (two-sample t tests, p < 0.05).
Neurobiology of Language
368
Genetics and anomalous neuroanatomy in stuttering
any of the assessments, and only two from each group tested at or below −1 SD on the tests (1
CWS for GFTA and 1 CWS for PPVT; 2 controls on GFTA).
Determination of Stuttering Status and Eventual Persistence versus Recovery
The Stuttering Severity Instrument (SSI-4; Riley, 2009), as well as offline analysis of recorded
speech samples comprising both conversation (parent, clinician samples separately) and nar-
rative samples elicited through storytelling using the wordless book, Frog, where are you?
(Mayer, 1969), were used to conduct disfluency analysis, quantify the frequency of SLDs, note
any physical concomitants of stuttering (e.g., tense lips, facial muscles during stuttering), and
duration of stuttering blocks and prolongations. SSI-4 was used to examine frequency and du-
ration of disfluencies occurring in the speech sample, as well as any physical concomitants
associated with stuttering. These were incorporated into a composite stuttering severity rating
(Riley, 2009). Children were determined to be in the stuttering group if they exhibited at least
3% SLDs, for example, part-word repetitions, prolongations, and blocks (Yairi, Ambrose,
Paden, & Watkins, 2005), and scored at least in the “very mild” category based on the com-
posite SSI-4 score. To confirm reliability of the SSI-4 scores, a random subset of the speech
samples (25%) was rated by a second independent SLP. The intraclass correlation coefficient
calculated based on the two SLPs’ ratings was 0.96, indicating high reliability. In borderline
cases, expressed parental concern and clinician report were considered to confirm the diag-
nosis of stuttering. Children in the control group exhibited less than 3% SLDs, reported no
family history of stuttering, and neither clinician nor parent reported any concern regarding
stuttering.
CWS were further categorized as recovered (rCWS) or persistent (pCWS) based on their
SSI-4 scores from two or more visits. A child was considered recovered if the SSI-4 score
was 10 or below at the second visit or thereafter. A child was categorized as persistent if
the composite SSI-4 score was higher than 10 at the second visit or thereafter. Both clinician
and parent reports were required to be consistent with stuttering severity assessments in deter-
mining whether a child had recovered or was persistent. Three CWS were excluded because
they were assessed only once and, therefore their final diagnoses could not be determined.
Exclusion due to MRI Tolerance, Excess Head Movement and Incidental Findings
Thirty-three of 128 subjects did not successfully complete the MRI session for variable reasons
(e.g., uneasiness in the MRI setting and anxiety detected during mock scanner training), leav-
ing 50 CWS (20 girls and 30 boys) and 45 controls (23 girls and 22 boys) who completed
scans. The mean ages of CWS and controls at the first visit were 5.55 (SD = 2.02) and 5.99
(SD = 2.00) years, ranging from 3 to 10 years. Another CWS was excluded due to incidental
findings in the structural scan. Three CWS and one control were excluded due to excessive
head movement during scanning.
Final Data Set
The subject selection procedure described above is summarized in a flowchart (see Figure S1
in the appendix in the online supporting information located at https://www.mitpressjournals.
org/doi/suppl/10.1162/nol_a_00017), and the demographics and behavioral results of the par-
ticipants included in the final data set are listed in Table 1. The final analysis included 87 scans
from 26 pCWS (8 girls and 18 boys; mean age at the first visit = 6.5 years; SD = 1.9), 61 scans
from 17 rCWS (8 girls and 9 boys; mean age at the first visit = 5.4 years; SD = 1.9), and 139
Neurobiology of Language
369
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
o
_
a
_
0
0
0
1
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Genetics and anomalous neuroanatomy in stuttering
scans from 44 controls (23 girls and 21 boys; mean age at the first visit = 6.5 years; SD = 2.0).
These children were recruited from printed advertisements in newspapers or magazines
(5 controls, 10 persistent, 7 recovered), parent email listservs (14 controls, 2 persistent, 1 recov-
ered), study flyers (8 controls, 3 persistent, 1 recovered), SLP referral (1 control, 6 persistent,
1 recovered), word of mouth (5 controls, 2 persistent, 3 recovered), or other methods (e.g.,
Craigslist, Facebook, postcards; 2 controls, 3 persistent, 2 recovered). The specific ascertainment
information was not known for 9 controls, 3 persistent, and 4 recovered, but all were recruited
from the same general recruitment methods. Both persistent and recovered groups did not differ
from controls in chronological age, sex, handedness, or socioeconomic status. Because
GNPTAG, NAGPA, GNPTG and AP4E1 are associated with persistent stuttering, only scans col-
lected from pCWS and controls were included in the primary analysis of gene expression.
Separate secondary analyses of pCWS (males only) compared to the control group, and rCWS
compared to controls, were performed.
Voxel-based Morphometry
Anatomical images were acquired on a GE 3T Signa scanner with an 8-channel head coil at
Michigan State University. In each scan session, a whole brain three-dimensional inversion
recovery fast spoiled gradient-recalled T1-weighted images with cerebrospinal fluid (CSF) sup-
pressed was obtained using the following parameters: time of echo = 3.8 ms, time of repetition
of acquisition = 8.6 ms, time of inversion = 831 ms, repetition time of inversion = 2,332 ms, flip
angle = 8°, and receiver bandwidth = 620.8 kHz. For voxel-based morphometry (VBM) analysis,
we used the optimized procedure proposed by Good et al. (2001). In summary, anatomical images
were first segmented into different tissue partitions (Ashburner & Friston, 2005). Gray and white
matter images were nonlinearly registered to a Montreal Neurological Institute (MNI) template
using a diffeomorphic image registration algorithm (DARTEL) (Ashburner, 2007).
To accommodate for brain size differences, registrations were performed iteratively in a
coarse-to-fine manner. Volumetric changes of each voxel were obtained by multiplying (or
modulating) voxel values in the gray matter image by the deformation field derived from the
registration procedure. Individual, modulated images were resampled to 1.5 mm isotropic
voxels and spatially smoothed with a 6 mm full width half maximum (FWHM) kernel. To
account for the dependence of participants’ multiple scans in this study, GMV images were
analyzed using the sandwich estimator method, which was designed for analyzing longitu-
dinal and repeated measures data (Guillaume, Hua, Thompson, Waldorp, & Nichols, 2014).
The model included group (pCWS and controls) and group by age interaction as well as
quadratic age, sex, IQ, brain size, socioeconomic status, and stuttering severity as covariates
to control potential sources of variation. Although there was a significant difference between
CWS and controls in IQ and both language measures, PPVT-4 and EVT-2 (Table 1), only IQ
was included in the model because both measures were highly correlated with IQ (PPVT-IQ
r = 0.70, EVT-IQ r = 0.69). The overall means of each covariate, except stuttering severity,
were removed to capture the variation and potential differences between groups associated
with the covariates. Since stuttering severity is only relevant to CWS, we considered stutter-
ing severity to be zero for controls, and the mean for CWS was removed from the measure
so that it would remove the variation associated with stuttering severity without affecting the
group estimates.
Voxel-wise t statistics of the group difference were calculated. For comparing our VBM re-
sults with the findings in the literature, we also applied a threshold to visualize the significant
GMV differences. However, identifying significant GMV differences between groups is not the
Neurobiology of Language
370
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
o
_
a
_
0
0
0
1
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Genetics and anomalous neuroanatomy in stuttering
primary goal of this study. A voxel-wise height threshold p < 0.005 and a cluster-size threshold
k > 316 voxels were used to control for false positives. This set of thresholds corresponds to a
corrected p < 0.05. The cluster-size threshold was determined by AFNI 3dClustSim (version
17.2.13; https://afni.nimh.nih.gov/pub/dist/doc/program_help/3dClustSim.html). Specifically,
we first generated a non-Gaussian noise model according to the spatial smoothness of the re-
sidual images using the AFNI 3dFWHMx autocorrelation function (-acf option; https://afni.
nimh.nih.gov/pub/dist/doc/program_help/3dFWHMx.html for 3dFWHMx). Then, we used
Monte Carlo simulations implemented in AFNI 3dClustSim to estimate the false positive rate
from the noise model (Cox, Chen, Glen, Reynolds, & Taylor, 2017).
Gray Matter Volume and Gene Expression Correlation
Microarray-based gene expression data were obtained from the AIBS, which provides normal-
ized expression of 29,131 genes using a total of 58,692 probes in each of 3,702 brain samples
obtained from six adult donors (5 males, 1 female; age 24–57 years; see http://www.brain-
map.org/ for details; Hawrylycz et al., 2012). We excluded genes whose symbols could
not be identified in the HUGO Gene Nomenclature Committee database (https://www.
genenames.org/), resulting in a total of 19,174 unique genes. The T1-weighted MRIs of the
donors were segmented into different tissue partitions and normalized to the MNI template
using the same procedure used for analyzing the structural images acquired from our pediatric
subjects. Using the deformation field generated by DARTEL, the locations of brain samples in native
space were transformed into the MNI space. The samples’ locations were mapped to 90 cor-
tical and subcortical regions and the cerebellum, based on a standard atlas with automated
anatomical labeling (Tzourio-Mazoyer et al., 2002). Because samples in the right hemisphere
were taken from only two of the six donors, only supratentorial regions in the left hemisphere
were included in our GMV-gene expression analysis. Since the right cerebellum has strong
anatomical connections with the left cerebral hemisphere, and cerebellar anomalies have
been associated with stuttering, the right cerebellum was included in our analyses as a single
region. In total, 46 regions were included in the primary GMV-gene expression analysis. For
each donor, expression of the same gene at each sample location from different probes was
first averaged. Gene expression for each region was represented by the median of all the
samples in the region. This step generated a parcellated expression map for each of the
19,174 genes.
The GMV difference of each region was calculated by taking the mean of voxel-wise ab-
solute t statistics of between-group GMV differences (|t stat|) within the region. Absolute GMV
difference was used because the effect of genetic variation on GMV in stuttering is not known.
It is biologically plausible that the relationship between GMV and gene expression is direc-
tional. However, since the biological mechanism of this potential relationship is not known,
we could not rule out the possibility that the relationship is nondirectional (i.e., the level of
gene expression is only related to the magnitude of GMV difference). Thus, we chose to use
the absolute GMV difference because it is sensitive to both possibilities (i.e., the directional
and nondirectional relationships). However, this method ignores biologically plausible as-
sumptions of directional correlations, that is to say, the higher the expression, the greater/lesser
the GMV in CWS relative to controls. To further explore the potential directional relationship
between gene expression and GMV differences, we carried out a post hoc analysis using the
averaged t statistics across voxels in each region to represent the pattern of GMV difference.
As has been done previously in order to minimize the potential adverse effect of outliers,
we used Spearman’s rank correlation to assess the relationship between gene expression and
Neurobiology of Language
371
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
o
_
a
_
0
0
0
1
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Genetics and anomalous neuroanatomy in stuttering
between-group GMV difference, instead of Pearson’s correlation (Ortiz-Terán et al., 2017;
Richiardi et al., 2015). We calculated Spearman’s rank correlation coefficient (ρ) between
GMV differences and each of the 19,174 genes expressed across the 46 regions. This proce-
dure established a distribution of correlation coefficients. Statistical threshold was set at q <
0.05 (adjusted p < 0.05), corrected for multiple testing by controlling the false discovery rate
(FDR; Benjamini & Hochberg, 1995).
Gene Set Enrichment Analysis
Since genes other than GNPTG and NAGPA that are expressed in concordance with between-
group GMV differences might also be associated with persistent stuttering, we carried out a
gene set enrichment analysis to identify biological processes, molecular functions, cellular
components, or KEGG pathways (Kyoto Encyclopedia of Genes and Genomes; https://www.
genome.jp/kegg/) for which the top 2.5% of the genes that were most positively correlated
with GMV differences are enriched. The 19,174 genes were used as the input of the back-
ground set for the enrichment analyses. We used PANTHER (http://geneontology.org/) to
identify enrichment for biological processes, molecular functions, or cellular components,
and DAVID (https://david.ncifcrf.gov/) for identifying enrichment for KEGG pathways
(M. Ashburner et al., 2000; Huang, Sherman, & Lempicki, 2009; The Gene Ontology
Consortium, 2017). The redundancy of the resulting gene ontology terms were removed
by using REViGO (Supek, Bošnjak, Škunca, & Šmuc, 2011). Fisher’s exact test was used to
determine statistical significance of enrichment factors. Statistical threshold was set at q <
0.05, corrected for multiple testing by controlling the FDR (Benjamini & Hochberg, 1995).
Although the regional expression of our targeted four genes was only positively correlated
with the GMV differences, for exploratory purposes, we carried out the same enrichment
analysis using the 2.5% of the genes that were most negatively correlated with GMV
differences.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
o
_
a
_
0
0
0
1
7
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
RESULTS
We used 87 longitudinally acquired structural scans from 26 pCWS and 139 scans from 44
controls using VBM, a well-established neuroimaging technique (Ashburner, 2007; Good
et al., 2001), to estimate voxel-wise GMV differences across the whole brain. Controlling for
sex, age, quadratic age, cranial brain volume, IQ, socioeconomic status, and stuttering severity
(SSI-4 score), the longitudinal analysis model showed that in the pCWS group, GMV in the left
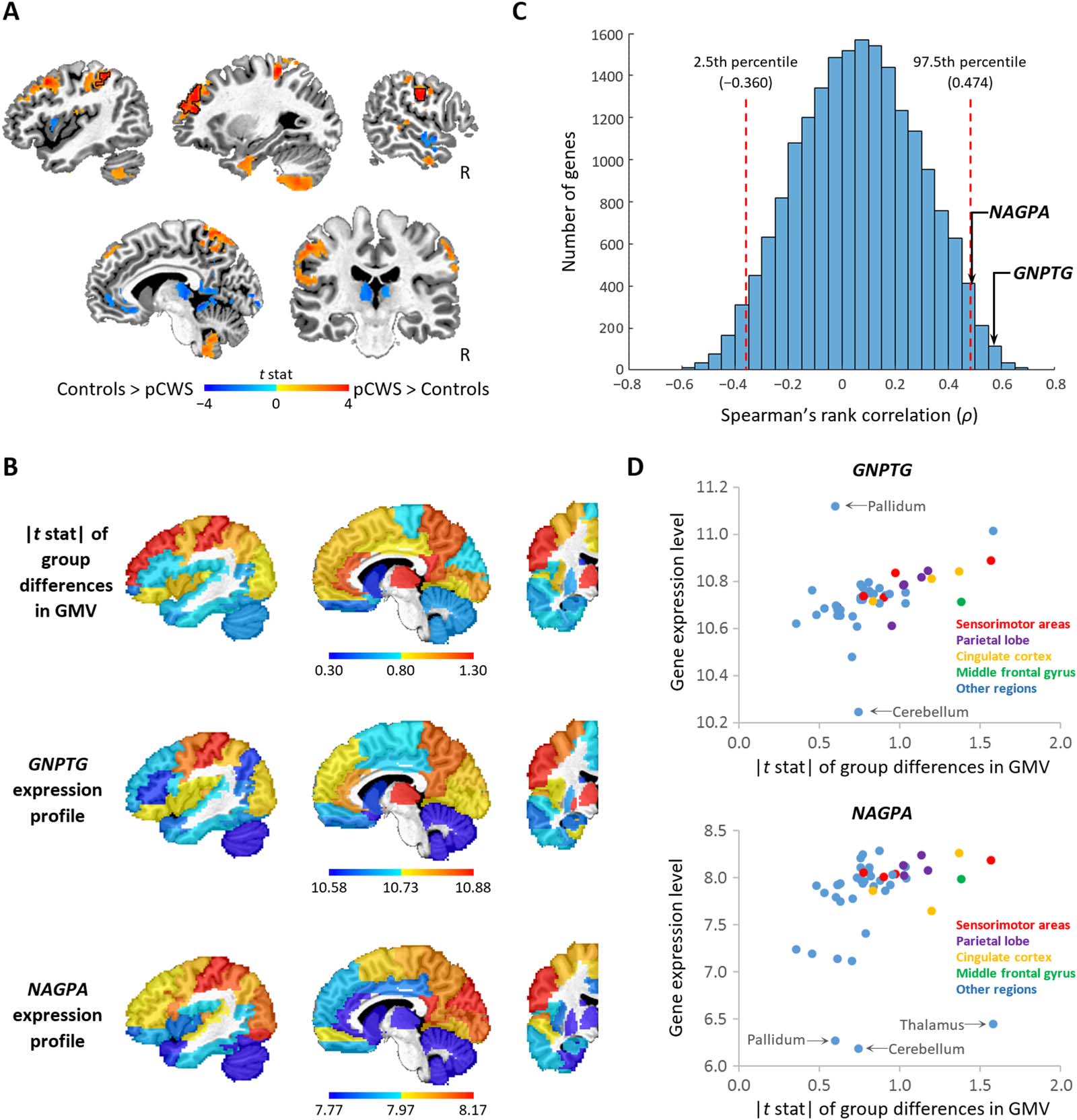
somatosensory, the left anterior prefrontal, and the right motor areas was significantly larger
than in the control group (Figure 1A). Some regions such as the thalamus and the left inferior
frontal gyrus showed a decrease in GMV, but none of them survived after correction for
multiple comparisons (Figure 1A). We examined the spatial correspondence between the
expression of the GNPTG, NAGPA, GNPTAB and AP4E1 genes and between-group differ-
ences in GMV across the 46 regions using the Spearman rank correlation (Figure 1B). The
correlation coefficients (ρ) associated with GNPTG, NAGPA, GNPTAB and AP4E1 genes
were 0.57 (q < 0.01), 0.49 (q < 0.05), −0.08 (q = 0.78), and 0.08 (q = 0.78), respectively.
As illustrated in the frequency distribution for ρ for all 19,174 genes in our analysis (Figure 1C),
the ρ values associated with GNPTG and NAGPA genes were significantly higher than the
97.5 percentile.
Since there was a tendency for a between-group difference in sex ratio, χ2(1, N = 70) = 3.09,
p = 0.079, we conducted a follow-up analysis including only male pCWS and controls to rule
Neurobiology of Language
372
Genetics and anomalous neuroanatomy in stuttering
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
o
_
a
_
0
0
0
1
7
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Spatial relationship between gene expression and between-groups differences in gray matter volume (GMV). (A) Voxel-wise dif-
ferences between children with persistent stuttering (pCWS) and controls in GMV. Color-coded t values of group differences are overlaid on an
anatomical image. Areas exhibiting a significant between-group difference in GMV (corrected p < 0.05) are outlined by black lines. The other
colored areas are subthreshold (uncorrected p < 0.1). (B) Parcellated gene expression of GNPTG and NAGPA and absolute GMV differences in
t statistics (|t stat|) in 45 left hemispheric regions and the right cerebellum (which anatomically connects to the left hemisphere) were overlaid
on a single-subject anatomical image. The parcellation of the brain was based on a standard atlas with automated anatomical labeling (AAL).
Gene expression and |t stat| of the right cerebellum were displayed in the left cerebellum to save space. (C) Frequency plot of Spearman’s
correlation coefficients between GMV group differences and each of the 19,174 genes expressed across the regions. The red dashed lines
indicate the levels of correlation at the 2.5th and 97.5th percentiles. The probability of obtaining a correlation >0.474 或者 <−0.360 is less than
5% if a gene is randomly selected. (D) Scatter plots between gene expression and group differences in GMV in the sensorimotor areas (red
dots), the parietal lobe (purple dots), the cingulate cortex (orange dots), the middle frontal gyrus (green dots) and the rest of the regions (blue
dots). Regions in which gene expression is 2.5 standard deviations above or below the mean are labeled.
out the possibility that the observed gene-brain relationship was driven by potential sex differ-
ences. The results of the male-only analysis were very similar to the original results (i.e.,
GNPTG, ρ = 0.47, q < 0.05; NAGPA, ρ = 0.57, q < 0.01; GNPTAB, ρ = 0.03, q = 0.93, and
AP4E1, ρ = 0.01, q = 0.98), indicating that the results were not driven by a higher proportion of
female subjects in the control group.
Neurobiology of Language
373
Genetics and anomalous neuroanatomy in stuttering
The scatter plots in Figure 1D show the relationship between gene expression and between-
group differences in GMV across regions in the original analysis. We observed that the expres-
sion level of the GNPTG and NAGPA genes in the cerebellum and some subcortical regions,
such as the pallidum, deviated from the relationship seen in the other regions. Repeating the
analysis excluding the cerebellum, basal ganglia regions, and thalamus, we obtained similar
results (i.e., GNPTG, ρ = 0.57, q < 0.05; NAGPA, ρ = 0.42, q < 0.10; GNPTAB, ρ = 0.02, q =
0.58, and AP4E1, ρ = 0.08, q = 0.50), although the correlation for NAGPA was no longer
significant (see Figure S2 in the appendix in the online supporting information). Figure 1D fur-
ther illustrates the monotonic relationship between the expression of GNPTG/NAGPA and
between-group GMV differences in our original analysis. To further explore whether the rela-
tionship between gene expression and between-group differences in GMV was specific to per-
sistent stuttering, we performed the same analysis on 17 rCWS. For rCWS, the relationship was
not significant for any of the four genes (i.e., GNPTG, ρ = −0.08, q = 0.56; NAGPA, ρ = 0.20,
q = 0.38: GNPTAB, ρ = −0.24, q = 0.34; AP4E1, ρ = 0.15, q = 0.45). Since we used the
magnitude of between-group GMV differences to examine the spatial relationship with gene
expression, the results reported above could not delineate whether the observed spatial
correction was directional, for example, the higher the expression, the greater/lesser the
GMV in CWS relative to controls. To explore whether the observed spatial correlation was
directional or driven by the magnitude of between-group GMV differences, we carried out
a post hoc directional analysis, which repeated the original analysis of spatial relationship
using the directional GMV differences. We found no significant spatial correlation between
the directional GMV differences and gene expression of the four genes associated with stuttering
( p > 0.1).
Gene set enrichment analysis of the top 2.5% of the genes whose expression was most
positively correlated with the between-group differences in GMV (see Table S1 in the appen-
dix in the online supporting information) showed that this set of genes was highly enriched
with genes involved in energy metabolism and mitochondrial functions (see Tables S2 and
S3 in the online appendix). Genes in several KEGG pathways were also overrepresented in
the gene set. Forty-nine out of 479 (10.2%) of the genes analyzed were involved in metabolic
pathways, including overrepresentation of a subset of genes involved in citrate cycle
(hsa00020) and oxidative phosphorylation (hsa00190; Table S4 in the online appendix).
Genes involved in oxidative phosphorylation were also linked to a number of neurological
disorders, including Parkinson’s disease, 阿尔茨海默氏病, Huntington’s disease, 和
amyotrophic lateral sclerosis (Table S5 in the online appendix). Using the same method, 这
顶部 2.5% of the genes negatively correlated with GMV differences were significantly enriched
for the DNA packaging complex and the nucleosome (cellular components) as well as three
KEGG pathways: alcoholism (hsa05034), systemic lupus erythematosus (hsa05322) and viral
carcinogenesis (hsa05203; Tables S6 and S7 in the online appendix).
讨论
迄今为止, two VBM studies in children who stutter have been published in peer-reviewed jour-
nals (Beal, Gracco, Brettschneider, Kroll, & De Nil, 2013; 张, 埃里克森, Ambrose,
Hasegawa-Johnson, & Ludlow, 2008). Despite small sample sizes (<12 pCWS) and the use
of relatively lenient statistical thresholds, both studies showed that smaller GMV in bilateral
inferior frontal gyrus (IFG) is associated with stuttering persistence children. In current
study, decreased IFG was observed only at an uncorrected threshold
(Figure 1A). On other hand, our data with
significantly increased left prefrontal bilateral sensorimotor areas, which have
Neurobiology of Language
374
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
> 0.1). The results of this individual donor analysis
point in the same direction as our results using the gene expression aggregated from the six
donors. For readers who are interested in further examining the individual donor variability,
the regional expression patterns of the four targeted genes in each of the six donors are pre-
sented in Figures S3 and S4 (see the online appendix). 迄今为止, the expression data from AIBS
are the only source of human gene expression patterns with high spatial resolution. 作为男人-
tioned in the Introduction, AIBS gene expression data and the approach of the current study
Neurobiology of Language
376
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
哦
_
A
_
0
0
0
1
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
Genetics and anomalous neuroanatomy in stuttering
have been used to reveal relationships between genes and intrinsic brain networks,
neuroplasticity-related genes, and altered functional connectivity in blind children, 和
expression patterns of known risk genes and the brain anomalies of their associated neurolog-
ical disorders including Alzheimer’s disease, Huntington’s disease, and schizophrenia (Grothe
等人。, 2018; McColgan et al., 2017; Ortiz-Terán et al., 2017; Richiardi et al., 2015; Romme
等人。, 2017).
第四, we cannot completely rule out the possibility that expression levels and patterns of
some genes in children and adults are different. If this is the case for the genes associated with
stuttering, we would expect that the spatial correlation between gene expression in the six
adult donors and the GMV patterns in children would be weak, whereas a strong relationship
was observed in our study. 而且, previous studies have suggested that the changes in
gene expression occur predominately during prenatal and infant development, and become
relatively stable by around 6 年龄 (Kang et al., 2011). Kang et al. estimated that only
9.1% of genes exhibit temporally differential expression in the first 20 years of life, 和
portion of differentially expressed genes should be even less in our participants’ age group.
Future studies using gene expression profiles in children should be pursued to refine our
understanding of the relationship between brain anomalies and gene expression, when such
data sets become available.
结论
综上所述, we showed that relative to controls, pCWS exhibited greater GMV in the left
somatosensory, the left anterior prefrontal, and the right motor areas. The nondirectional
magnitude of GMV differences (both increases and decreases) in stuttering children relative
to controls across regions in the left hemisphere and the right cerebellum was positively
correlated with the expression of lysosomal targeting genes GNPTG and NAGPA as well as
genes involved in energy metabolism. More research is warranted to further investigate
possible roles of M6P mediated intracellular and extracellular trafficking, as well as whether
metabolic functions play a role in the development of brain structural anomalies associated
with stuttering.
致谢
The authors wish to thank all the children and parents who participated in this study. 我们也
thank Kristin Hicks for her assistance in participant recruitment, behavioral testing, and help
with MRI data collection, Scarlett Doyle for her assistance in MRI data acquisition, and Ashley
Diener for her assistance in speech data analyses.
资金信息
Ho Ming Chow, National Institute on Deafness and Other Communication Disorders (http://
dx.doi.org/10.13039/100000055), 奖项ID: R21DC015853. Dennis Drayna, 国家的
Institute on Deafness and Other Communication Disorders (http://dx.doi.org/10.13039/
100000055), 奖项ID: Z1A-000046. Soo-Eun Chang, National Institute on Deafness and
Other Communication Disorders (http://dx.doi.org/10.13039/100000055), 奖项ID:
R01DC011277. Soo-Eun Chang, National Institute on Deafness and Other Communication
Neurobiology of Language
377
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
哦
_
A
_
0
0
0
1
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
Genetics and anomalous neuroanatomy in stuttering
Disorders (http://dx.doi.org/10.13039/100000055), 奖项ID: R21DC015312. Soo-Eun
张, Matthew K. Smith Stuttering Research Fund.
作者贡献
Ho Ming Chow: 概念化, 数据管理, 方法, 形式分析,
可视化, Writing—original draft, Writing—editing. Emily O. 加内特: 数据管理,
Writing—editing. Hua Li: 形式分析, Writing—editing. Andrew Etchell: Writing—editing.
Jorge Sepulcre: 方法, Writing—editing. Dennis Drayna: Writing—editing. Diane
Chugani: 概念化, 方法, Writing—editing. Soo-Eun Chang: 数据管理,
方法, Writing—editing.
参考
Abou Jamra, R。, Philippe, 奥。, Raas-Rothschild, A。, 埃克, S. H。, Graf,
E., Buchert, R。, … Colleaux, L. (2011). Adaptor protein complex
4 deficiency causes severe autosomal-recessive intellectual dis-
能力, progressive spastic paraplegia, shy character, and short
stature. American Journal of Human Genetics, 88(6), 788–795.
https://doi.org/10.1016/j.ajhg.2011.04.019
Ashburner, J. (2007). A fast diffeomorphic image registration algo-
rithm. 神经影像, 38(1), 95–113. https://doi.org/10.1016/j.
neuroimage.2007.07.007
Ashburner, J。, & 弗里斯顿, K. J. (2005). Unified segmentation.
神经影像, 26(3), 839–851. https://doi.org/10.1016/j.
neuroimage.2005.02.018
Ashburner, M。, Ball, C. A。, Blake, J. A。, Botstein, D ., 管家, H。,
Cherry, J. M。, … Sherlock, G. (2000). Gene Ontology: Tool for
the unification of biology. 自然遗传学, 25(1), 25–29.
https://doi.org/10.1038/75556
巴恩斯, J。, Lim, J. M。, Godard, A。, Blanchard, F。, 韦尔斯, L。, & Steet,
右. (2011). Extensive mannose phosphorylation on leukemia
inhibitory factor (LIF) controls its extracellular levels by multi-
Journal of Biological Chemistry, 286(28),
ple mechanisms.
24855–24864. https://doi.org/10.1074/jbc.M111.221432
Beal, D. S。, Gracco, V. L。, Brettschneider, J。, Kroll, 右. M。, & De Nil,
L. F. (2013). A voxel-based morphometry ( VBM) analysis of re-
gional grey and white matter volume abnormalities within the
speech production network of children who stutter. Cortex, 49(8),
2151–2161. https://doi.org/10.1016/j.cortex.2012.08.013
Benito-Aragón, C。, Gonzalez-Sarmiento, R。, Liddell, T。, Diez, 我。,
d’Oleire Uquillas, F。, Ortiz-Terán, L。, … Sepulcre, J. (2019).
Neurofilament-lysosomal genetic intersections in the cortical net-
work of stuttering. Progress in Neurobiology. https://doi.org/
10.1016/j.pneurobio.2019.101718
Benjamini, Y。, & Hochberg, 是. (1995). Controlling the false discov-
ery rate: A practical and powerful approach to multiple testing.
Royal Statistical Society B, 57(1), 289–300. https://www.jstor.org/
stable/2346101
Blumgart, E., 特兰, Y。, & Craig, A. (2010). An investigation into the
personal financial costs associated with stuttering. 杂志
Fluency Disorders, 35(3), 203–215.
布劳恩, A. R。, Varga, M。, Stager, S。, Schulz, G。, Selbie, S。, Maisog,
J. M。, … Ludlow, C. L. (1997). Altered patterns of cerebral activity
during speech and language production in developmental stut-
tering. An H2(15)O positron emission tomography study. Brain,
120(5), 761–784. https://doi.org/10.1093/brain/120.5.761
张, S. E., 埃里克森, K. 我。, Ambrose, 氮. G。, Hasegawa-Johnson,
中号. A。, & Ludlow, C. L. (2008). Brain anatomy differences in
childhood stuttering. 神经影像, 39(3), 1333–1344. https://
doi.org/10.1016/j.neuroimage.2007.09.067
秋, A. L。, Burnham, E., 希克斯, K., & 张, S. 乙. (2016).
Dissociations among linguistic, cognitive, and auditory-motor
neuroanatomical domains in children who stutter. 杂志
C o m mu n i c a t i o n D i s o r d e r s. ht t p s : / / d o i . o r g / 1 0 . 1 0 1 6/ j .
jcomdis.2016.03.003
Chow, H. M。, & 张, S. 乙. (2017). White matter developmental
trajectories associated with persistence and recovery of child-
hood stuttering. 人脑图谱. https://doi.org/
10.1002/hbm.23590
Coulter, C. E., 安德森, J. D ., & Conture, 乙. G. (2009). 童年
stuttering and dissociations across linguistic domains: A replica-
tion and extension. Journal of Fluency Disorders. https://doi.org/
10.1016/j.jfludis.2009.10.005
考克斯, 右. W., 陈, G。, Glen, D. R。, Reynolds, 右. C。, & 泰勒, 磷. A.
(2017). fMRI clustering and false-positive rates. 会议记录
the National Academy of Sciences, 201614961.
Craig, A。, Blumgart, E., & 特兰, 是. (2009). The impact of stuttering
on the quality of life in adults who stutter. Journal of Fluency
Disorders, 34(2), 61–71.
Craig, A。, Hancock, K., 特兰, Y。, Craig, M。, & Peters, K. (2002).
Epidemiology of stuttering in the community across the entire life
span. Journal of Speech, 语言, and Hearing Research, 45(6),
1097–1105. https://doi.org/10.1044/1092-4388(2002/088)
De Leon, D. D ., 特里, C。, Asmerom, Y。, & Nissley, 磷. (1996).
Insulin-like growth factor II modulates the routing of cathepsin
D in MCF-7 breast cancer cells. Endocrinology, 137(5), 1851–1859.
https://doi.org/10.1210/endo.137.5.8612524
Dell’Angelica, 乙. C。, Mullins, C。, & Bonifacino, J. S. (1999). AP-4, A
novel protein complex related to clathrin adaptors. 杂志
Biological Chemistry, 274, 7278–7285. https://doi.org/10.1074/
jbc.274.11.7278
Dunn, L. M。, & Dunn, D. 中号. (2007). Peabody picture vocabulary
测试 (4第三版。). Circle Pines, 明尼苏达州: AGS.
Dworzynski, K., Remington, A。, Rijsdijk, F。, 豪厄尔, P。, & Plomin,
右. (2007). Genetic etiology in cases of recovered and persistent
stuttering in an unselected, longitudinal sample of young twins.
American Journal of Speech-Language Pathology, 16(2), 161–178.
https://doi.org/10.1044/1058-0360(2007/021)
Etchell, A. C。, Civier, 奥。, Ballard, K. J。, & Sowman, 磷. F. (2018). A
systematic literature review of neuroimaging research on devel-
opmental stuttering between 1995 和 2016. Journal of Fluency
Disorders, 55, 6–45. https://doi.org/10.1016/j.jfludis.
2017.03.007
Neurobiology of Language
378
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
哦
_
A
_
0
0
0
1
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
Genetics and anomalous neuroanatomy in stuttering
Fagnani, C。, Fibiger, S。, Skytthe, A。, & Hjelmborg, J. V. 乙. (2011).
Heritability and environmental effects for self-reported periods
with stuttering: A twin study from Denmark. Logopedics
Phoniatrics Vocology, 36, 114–120. https://doi.org/10.3109/
14015439.2010.534503
Fernandez, A. M。, & Torres-Alemán, 我. (2012). The many faces of
insulin-like peptide signalling in the brain. 自然评论
神经科学, 13, 225–239. https://doi.org/10.1038/nrn3209
狐狸, 磷. T。, Ingham, 右. J。, Ingham, J. C。, Hirsch, 时间. B., Downs, J. H。,
马丁, C。, … Lancaster, J. L. (1996). A PET study of the neural
systems of stuttering. 自然, 382(6587), 158–162. https://土井.
org/10.1038/382158a0
Frigerio-Domingues, C。, & Drayna, D. (2017). Genetic contribu-
tions to stuttering: The current evidence. Molecular Genetics
and Genomic Medicine, 5, 95–102. https://doi.org/10.1002/
mgg3.276
Fusar-Poli, P。, Radua, J。, Frascarelli, M。, Mechelli, A。, Borgwardt, S。,
Di Fabio, F。, … David, S. 磷. (2014). Evidence of reporting biases
in voxel-based morphometry ( VBM) studies of psychiatric and
neurological disorders. 人脑图谱, 25(7). https://
doi.org/10.1002/hbm.22384
加内特, 乙. 奥。, Chow, H. M。, Nieto-Castañón, A。, Tourville, J. A。,
Guenther, F. H。, & 张, S.-E. (2018). Anomalous morphology
in left hemisphere motor and premotor cortex of children who
stutter. Brain, 141(9), 2670–2684. https://doi.org/10.1093/brain/
awy199
Gary-Bobo, M。, Nirdé, P。, Jeanjean, A。, Morère, A。, & 加西亚, 中号.
(2007). Mannose 6-phosphate receptor targeting and its applica-
tions in human diseases. Current Medicinal Chemistry, 14(28),
2945–2953. https://doi.org/10.2174/092986707782794005
Gennatas, 乙. D ., Avants, 乙. B., Wolf, D. H。, Satterthwaite, 时间. D .,
Ruparel, K., Ciric, R。, … Gur, 右. C. (2017). Age-related effects
and sex differences in gray matter density, 体积, 大量的, 和
cortical thickness from childhood to young adulthood. 这
神经科学杂志, 37(20), 5065–5073. https://doi.org/
10.1523/JNEUROSCI.3550-16.2017
高盛, R。, & Fristoe, 中号. (2000). Goldman-Fristoe test of articu-
关系 (2nd Ed.). Circle Pines, 明尼苏达州: American Guidance Service.
好的, C. D ., Johnsrude, 我. S。, Ashburner, J。, Henson, 右. N。, 弗里斯顿,
K. J。, & Frackowiak, 右. S. (2001). A voxel-based morphometric
s t u d y o f a g e i n g i n 4 6 5 n o r m a l a d u l t h u m a n b r a i n s .
神经影像, 14(1), 21–36. https://doi.org/10.1006/nimg.
2001.0786
Goyal, 中号. S。, Hawrylycz, M。, 磨坊主, J. A。, 斯奈德, A. Z。, & Raichle,
中号. 乙. (2014). Aerobic glycolysis in the human brain is associated
with development and neotenous gene expression. 细胞
Metabolism, 19(1), 49–57. https://doi.org/10.1016/j.cmet.
2013.11.020
Grothe, 中号. J。, 墓, J。, Gonzalez-Escamilla, G。, Jelistratova, 我。,
Schöll, M。, Hansson, 奥。, & Teipel, S. J. (2018). Molecular prop-
erties underlying regional vulnerability to Alzheimer’s disease
pathology. Brain, 141(9), 2755–2771. https://doi.org/10.1093/
brain/awy189
Guillaume, B., Hua, X。, 汤普森, 磷. M。, Waldorp, L。, & Nichols,
时间. 乙. (2014). Fast and accurate modelling of longitudinal and re-
peated measures neuroimaging data. 神经影像, 94, 287–302.
https://doi.org/10.1016/j.neuroimage.2014.03.029
Han, V. K. M。, D’Ercole, A. J。, & Lund, 磷. K. (1987). Cellular loca-
lizaton of somatomedin (insulin-like growth factor) messenger
RNA in the human fetus. 科学, 236(4798), 193–197. https://
doi.org/10.1126/science.3563497
Hawrylycz, 中号. J。, Lein, 乙. S。, Guillozet-Bongaarts, A. L。, 沉, 乙. H。,
的, L。, 磨坊主, J. A。, … Jones, A. 右. (2012). An anatomically
comprehensive atlas of the adult human brain transcriptome.
自然, 489(7416), 391–399. https://doi.org/10.1038/
nature11405
Hoeft, F。, Carter, J. C。, Lightbody, A. A。, Hazlett, H. C。, Piven, J。, &
Reiss, A. L. (2010). Region-specific alterations in brain develop-
ment in one- to three-year-old boys with fragile X syndrome.
Proceedings of the National Academy of Sciences of the United
States of America. https://doi.org/10.1073/pnas.1002762107
黄, D. W., Sherman, 乙. T。, & Lempicki, 右. A. (2009).
Systematic and integrative analysis of large gene lists using
DAVID bioinformatics resources. Nature Protocols, 4, 44–57.
https://doi.org/10.1038/nprot.2008.211
Kang, C。, Riazuddin, S。, Mundorff, J。, Krasnewich, D ., 弗里德曼, P。,
Mullikin, J. C。, & Drayna, D. (2010). Mutations in the lysosomal
enzyme–targeting pathway and persistent stuttering. 新的
England Journal of Medicine, 362(8), 677–685. https://doi.org/
10.1056/NEJMoa0902630
Kang, H. J。, Kawasawa, 是. 我。, Cheng, F。, 朱, Y。, 徐, X。, 李, M。, ……
Šestan, 氮. (2011). Spatio-temporal transcriptome of the human
脑. 自然, 478(7370), 483–489. https://doi.org/10.1038/
nature10523
Kiess, W., 托马斯, C. L。, Greenstein, L. A。, 李, L。, Sklar, 中号. M。,
Rechler, 中号. M。, … Nissley, S. 磷. (1989). Insulin-like growth
factor-II (IGF-II) inhibits both the cellular uptake of β-galactosidase
and the binding of β-galactosidase to purified IGF-II/mannose
6-phosphate receptor. Journal of Biological Chemistry, 264(8),
4710–4714.
孔, X. F。, Bousfiha, A。, Rouissi, A。, Itan, Y。, Abhyankar, A。,
Bryant, 五、, … Boisson-Dupuis, S. (2013). A novel homozygous
p.R1105X mutation of the AP4E1 gene in twins with hereditary
spastic paraplegia and mycobacterial disease. PLOS ONE, 8(3).
https://doi.org/10.1371/journal.pone.0058286
Kornfeld, S. (2001). I-cell disease and pseudo-Hurler polydystro-
物理层: Disorders of lysosomal enzyme phosphorylation and local-
化. The Metabolic and Molecular Bases of Inherited Disease,
3469–3482.
Månsson, H. (2000). Childhood stuttering: Incidence and develop-
蒙特. Journal of Fluency Disorders, 25(1), 47–57.
Matrone, C。, Dzamko, N。, Madsen, P。, Nyegaard, M。, Pohlmann,
R。, Søndergaard, 右. 五、, … Nielsen, 中号. S. (2016). Mannose
6-phosphate receptor is reduced in -synuclein overexpressing
models of Parkinsons disease. PLOS ONE, 11(8). https://doi.org/
10.1371/journal.pone.0160501
Mayer, 中号. (1969). Frog, where are you? 纽约: Dial Press.
McColgan, P。, Gregory, S。, Seunarine, K. K., Razi, A。, Papoutsi, M。,
约翰逊, E., … Tabrizi, S. J. (2017). Brain regions showing white
matter loss in Huntington’s disease are enriched for synaptic and
metabolic genes. Biological Psychiatry, 83(5). https://doi.org/
10.1016/j.biopsych.2017.10.019
McKenna, 中号. C。, Schuck, 磷. F。, & 费雷拉, G. C. (2018).
Fundamentals of CNS energy metabolism and alterations in lyso-
somal storage diseases. Journal of Neurochemistry, 148(5).
https://doi.org/10.1111/jnc.14577
Neef, 氮. E., 安旺德, A。, & Friederici, A. D. (2015). The neuro-
biological grounding of persistent stuttering: From structure to
function. Current Neurology and Neuroscience Reports, 15.
https://doi.org/10.1007/s11910-015-0579-4
Oka, S。, Kawasaki, T。, & Yamashina, 我. (1985). Isolation and char-
acterization of mannan-binding proteins from chicken liver.
Archives of Biochemistry and Biophysics, 241(1), 95–105.
https://doi.org/10.1016/0003-9861(85)90366-2
Ooki, S. (2005). Genetic and environmental influences on stutter-
ing and tics in Japanese twin children. Twin Research and
Neurobiology of Language
379
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
哦
_
A
_
0
0
0
1
7
p
d
/
.
我
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
Genetics and anomalous neuroanatomy in stuttering
人类遗传学, 8(1), 69–75. https://doi.org/10.1375/
1832427053435409
Ortiz-Terán, L。, Diez, 我。, Ortiz, T。, Perez, D. L。, Aragón, J. 我。,
Costumero, 五、, … Sepulcre, J. (2017). Brain circuit–gene expres-
sion relationships and neuroplasticity of multisensory cortices in
blind children. 美国国家科学院院刊
科学, 201619121. https://doi.org/10.1073/pnas.1619121114
Pardo, M。, Cheng, Y。, Sitbon, 是. H。, Lowell, J. A。, Grieco, S. F。,
Worthen, 右. J。, … Barreda-Diaz, A. (2018). Insulin growth factor
2 (IGF2) as an emergent target in psychiatric and neurological
disorders. 审查. Neuroscience Research, 149. https://doi.org/
10.1016/j.neures.2018.10.012
Rautakoski, P。, Hannus, T。, Simberg, S。, Sandnabba, N。, & Santtila,
磷. (2012). Genetic and environmental effects on stuttering: A
twin study from Finland. Journal of Fluency Disorders, 37(3),
202–210. https://doi.org/10.1016/j.jfludis.2011.12.003
Raza, 中号. H。, Mattera, R。, Morell, R。, Sainz, E., Rahn, R。, Gutierrez,
J。, … Drayna, D. (2015). Association between rare variants in
AP4E1, a component of intracellular trafficking, 和坚持不懈的
stuttering. American Journal of Human Genetics, 97(5), 715–725.
https://doi.org/10.1016/j.ajhg.2015.10.007
Richiardi, J。, 阿尔特曼, A。, Milazzo, A. C。, 张, C。, Chakravarty, 中号.
M。, Banaschewski, T。, … Greicius, 中号. D. (2015). Correlated gene
expression supports synchronous activity in brain networks.
科学, 348(6240), 1241–1244. https://doi.org/10.1126/
science.1255905
Riley, G. (2009). SSI-4: Stuttering severity instrument. Austin, TX:
Pro-Ed.
Romme, 我. A. C。, de Reus, 中号. A。, Ophoff, 右. A。, Kahn, 右. S。, & van
den Heuvel, 中号. 磷. (2017). Connectome disconnectivity and cor-
tical gene expression in patients with schizophrenia. Biological
Psychiatry, 81(6), 495–502. https://doi.org/10.1016/j.
biopsych.2016.07.012
St Louis, K. 奥。, & Ruscello, D. 中号. (1987). Oral speech mechanism
screening exam–revised (OSMSE-R). Austin, TX: Pro-Ed.
Supek, F。, Bošnjak, M。, Škunca, N。, & Šmuc, 时间. (2011). REVIGO
summarizes and visualizes long lists of gene ontology terms.
PLOS ONE. https://doi.org/10.1371/journal.pone.0021800
The Gene Ontology Consortium. (2017). Expansion of the Gene
Ontology knowledgebase and resources. Nucleic Acids Research,
45(D1), D331–D338. https://doi.org/10.1093/nar/gkw1108
Tzourio-Mazoyer, N。, Landeau, B., Papathanassiou, D ., Crivello, F。,
Etard, 奥。, Delcroix, N。, … Joliot, 中号. (2002). Automated anatomi-
cal labeling of activations in SPM using a macroscopic anatomical
parcellation of the MNI MRI single-subject brain. 神经影像,
15(1), 273–289. https://doi.org/10.1006/nimg.2001.0978
van Beijsterveldt, C. 乙. M。, Felsenfeld, S。, & Boomsma, D. 我. (2010).
Bivariate genetic analyses of stuttering and nonfluency in a large
sample of 5-year-old twins. Journal of Speech Language and
Hearing Research, 53(3), 609. https://doi.org/10.1044/1092-
4388(2009/08-0202)
Wechsler, D. (1999). Wechsler abbreviated intelligence scale. 桑
Antonio, TX: Psychological Corporation.
Wechsler, D. (2002). Wechsler preschool and primary scale of in-
telligence (3rd Ed.). San Antonio, TX: Psychological Corporation.
(2007). EVT-2: Expressive vocabulary test.
威廉姆斯, K.
明尼阿波利斯, 明尼苏达州: Pearson Assessments.
Yairi, E., & Ambrose, 氮. (2013). Epidemiology of stuttering: 21st
century advances. Journal of Fluency Disorders. https://doi.org/
10.1016/j.jfludis.2012.11.002
Yairi, E., Ambrose, 氮. G。, Paden, 乙. P。, & Watkins, 右. V. (2005).
Early childhood stuttering: For clinicians by clinicians. Austin,
TX: Pro-ed.
Yaruss, J. S. (2010). Assessing quality of life in stuttering treatment
outcomes research. Journal of Fluency Disorders, 35(3), 190–202.
https://doi.org/10.1016/j.jfludis.2010.05.010
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
n
哦
/
我
/
我
A
r
t
我
C
e
–
p
d
F
/
/
/
/
1
3
3
6
5
1
8
6
7
7
3
5
n
哦
_
A
_
0
0
0
1
7
p
d
.
/
我
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
Neurobiology of Language
380