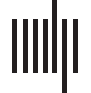研究
Fluid and flexible minds: Intelligence reflects
synchrony in the brain’s intrinsic
network architecture
迈克尔·A. Ferguson1,2∗
, Jeffrey S. Anderson2, 和R. Nathan Spreng1∗
1Laboratory of Brain and Cognition, Human Neuroscience Institute, Department of Human Development, 康奈尔大学,
伊萨卡岛, 纽约, 14853
2Departments of Bioengineering and Neuroradiology, University of Utah, Salt Lake City, UT, 84132
关键词: 智力, 功能磁共振成像, resting state functional connectivity, 机器学习, 认识
开放访问
杂志
抽象的
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
A
_
0
0
0
1
0
p
d
t
.
Human intelligence has been conceptualized as a complex system of dissociable cognitive
流程, yet studies investigating the neural basis of intelligence have typically emphasized
the contributions of discrete brain regions or, 最近, of specific networks of
functionally connected regions. Here we take a broader, systems perspective in order to
investigate whether intelligence is an emergent property of synchrony within the brain’s
intrinsic network architecture. Using a large sample of resting-state fMRI and cognitive data
(n = 830), we report that the synchrony of functional interactions within and across
distributed brain networks reliably predicts fluid and flexible intellectual functioning. 经过
adopting a whole-brain, systems-level approach, we were able to reliably predict individual
differences in human intelligence by characterizing features of the brain’s intrinsic network
建筑学. These findings hold promise for the eventual development of neural markers to
predict changes in intellectual function that are associated with neurodevelopment, 普通的
老化, and brain disease.
作者总结
In our study, we aimed to understand how individual differences in intellectual functioning
are reflected in the intrinsic network architecture of the human brain. We applied statistical
方法, known as spectral decompositions, in order to identify individual differences in the
synchronous patterns of spontaneous brain activity that reliably predict core aspects of
human intelligence. The synchrony of brain activity at rest across multiple discrete neural
networks demonstrated positive relationships with fluid intelligence. 相比之下, 全球的
synchrony within the brain’s network architecture reliably, and inversely, predicted mental
flexibility, a core facet of intellectual functioning. The multinetwork systems approach
described here represents a methodological and conceptual extension of earlier efforts that
related differences in intellectual ability to variations in specific brain regions, 网络, 或者
their interactions. Our findings suggest that the neural basis of complex, integrative cognitive
functions can be most completely understood from the perspective of network neuroscience.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Mapping the neural substrates of human intelligence could better inform our understand-
ing of how brain development influences lifespan development across multiple functional
域, both in health and in disease (Furnham, 2008; Gottfredson, 2004; Schutte, 2014).
The concept of general intelligence—Spearman’s g, or simply “g”—was first postulated by
Spearman at the turn of the previous century (Spearman, 1904). Tests for cognitive performance
引文: Ferguson, 中号. A。, 安德森,
J. S。, & Spreng, 右. 氮. (2017). Fluid
and flexible minds: 智力
reflects synchrony in the brain’s
intrinsic network architecture.
网络神经科学, 1(2), 192–207.
https://doi.org/10.1162/netn_a_00010
DOI:
https://doi.org/10.1162/netn_a_00010
支持信息:
已收到: 25 十月 2016
公认: 14 行进 2017
利益争夺: 作者有
声明不存在竞争利益
存在.
通讯作者:
迈克尔·A. Ferguson
maf378@cornell.edu
处理编辑器:
Danielle Bassett
版权: © 2017
麻省理工学院
在知识共享下发布
归因 4.0 国际的
(抄送 4.0) 执照
麻省理工学院出版社
Aspects of intelligence reflected by network synchrony in the brain
share ubiquitous positive correlations, suggesting that a common “intelligence” factor may ex-
plain individual differences across a range of cognitive domains. Over decades of inquiry,
it has been demonstrated that individual differences in Spearman’s g indeed robustly predict
performance on laboratory tests of cognitive function, as well as adaptive behavior in real-
world contexts (Kuncel, Hezlett, & Ones, 2004; Spearman, 1927). 最近, 与
advent of functional neuroimaging methods, investigating the neural basis of “g” has been an
active area of inquiry (Deary, Penke, & 约翰逊, 2010; Duncan, 2005; Duncan et al., 2000;
Schultz & Cole, 2016).
For decades, cognitive science has described intelligence as an emergent property of dis-
sociable processes (Churchland, 1986; Gregory & Zangwill, 1987; Sternberg & Detterman,
1986). Yet efforts to identify neural markers of “g” have typically focused on individual brain re-
祇翁 (Duncan, 2005; Duncan et al., 2000; Gray, Chabris, & Braver, 2003) or specific
networks of functionally connected brain regions (Cole, Yarkoni, Repovs, Anticevic, & Braver,
2012; 李等人。, 2006; Song et al., 2008). Frontoparietal networks have received the widest at-
tention as a possible single-network solution to understanding the biological underpinnings of
individual differences in intellectual ability (荣格 & Haier, 2007; Langeslag et al., 2013). 早期的
efforts to characterize the neural basis of cognitive functioning, which assumed a one-to-one
brain–behavioral mapping, have been increasingly challenged by models of whole-brain net-
work interactions, particularly with respect to complex, integrative cognitive capacities such
as intelligence (Bullmore et al., 2009; McIntosh, 1999; 斯波恩斯, Chialvo, Kaiser, & 希尔格塔格,
2004; van den Heuvel, 斯塔姆, Kahn, & Hulshoff Pol, 2009). This suggests that a systems-level,
network-based approach may prove fruitful in identifying reliable neural markers of individual
differences in human intellectual function.
Early localization studies investigated brain–behavior relationships in circumscribed brain
regions because methodological constraints, including low temporal and spatial resolution,
as well restrictions in computational capacity necessitated a univariate analytical approach
(Duncan, 2005; Duncan et al., 2000).
These analyses identified specific brain regions
where the magnitude of activity varied as a function of performance on intelligence tasks
(Duncan et al., 2000). Advances in fMRI data acquisition and analytical methods have
dramatically improved the temporal and spatial resolution of neuroimaging protocols, 使能
the measurement of regional interactions within distributed brain networks to characterize the
neural architecture of intelligence (李等人。, 2006; 史密斯等人。, 2013).
最近, studies have begun to characterize multinetwork dynamics, or the network
architecture of the brain, as a neural marker of intelligence (Cole, Ito, & Braver, 2015). Prop-
erties of large-scale, distributed networks observed in the brain during a wakeful resting state
have been identified as factors contributing to individual differences in intelligence (Finn et al.,
2015; Hearne et al., 2016; 史密斯等人。, 2015). Building from these earlier studies, here we ap-
ply spectral decompositions to resting-state functional connectivity (RSFC) from fMRI in order
to derive estimates of the brain’s spatially overlapping functional architecture that may be sensi-
tive to individual differences in “g.” Resting-state MRI is a powerful tool to detect and dissociate
functional brain networks from patterns of interregional correlations in neuronal variability, 英语-
timated by variations in BOLD signal and measured in the absence of explicit task demands
(巴克纳, 克里宁, Castellanos, Diaz, & 杨, 2011; Fox et al., 2005). Previous studies have
identified distributed contributions to intelligence in the brain by several methods, 包括
the examination of pairwise connections without imposed network definitions (Hearne et al.,
2016; 史密斯等人。, 2015), or the exploration of functional relationships both with and without
a priori network definitions (Finn et al., 2015). In these analyses, the functional correlations
193
Spectral decompositions:
Operations that determine the
orthogonal basis set of a matrix, 例如,
eigendecomposition, singular-value
分解, principal-component
分析
Resting-state functional connectivity:
An fMRI method for mapping
correlated patterns in spontaneous
blood oxygen level dependent signal
across the brain during wakeful rest
网络神经科学
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
A
_
0
0
0
1
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Aspects of intelligence reflected by network synchrony in the brain
Basis set:
Functions, represented as vectors,
whose combinations describe a
system in terms of its computational
and theoretical components
Intrinsic network architecture:
Large-scale patterns of resting-state
functional connectivity across
distributed anatomical regions
Fluid intelligence:
A component of general intelligence
that operationalizes pattern
recognition and completion in
cognitive testing
Cognitive flexibility:
The mental ability to switch between
different systems of representation,
operationalized by the Dimensional
Change Card Sort Task
Persistent synchrony:
Sustained functional covariance of
brain regions, or nodes, 在一个
网络
between pairwise regions represent the edges of distributed, spatially nonoverlapping net-
作品 (Finn et al., 2015; Hearne et al., 2016; 史密斯等人。, 2015). Here we seek to identify a
basis set of spatially overlapping functional networks in the resting brain and to determine the
contributions of within-network and across-network interactivity to intelligence. We identified
spatially overlapping network maps in an effort to characterize the one-to-many functional or-
ganization of the human brain that we believe is essential for understanding the relationships
between cognition and neural systems (Bullmore et al., 2009; McIntosh, 1999; Sporns et al.,
2004; van den Heuvel et al., 2009).
To identify a basis set of functional networks in the resting brain, we introduced the appli-
cation of a multivariate statistical method, spectral decomposition (Alter, 棕色的, & Botstein,
2000; Yeung, Tegner, & 柯林斯, 2002), to characterize an RSFC (IE。, intrinsic) network archi-
tecture in a large population of healthy young adults. This technique identifies the spec-
trum of spatially overlapping networks that covary across the duration of the resting-state
fMRI scan. Early brain-imaging studies had applied spectral decompositions to task-based
fMRI data (Bullmore et al., 1996; 弗里斯顿, Frith, 弗莱彻, Liddle, & Frackowiak, 1996). To our
知识, this is the first application of spectral analysis to RSFC data in order to describe the
functional dynamics of intrinsic brain networks and predict individual differences in cognitive
能力.
To evaluate whether individual differences in resting-state network dynamics may be a
marker of intellectual functioning, we drew upon two measures that have previously been used
to characterize intellectual capacity: fluid intelligence (IE。, cognitive control) and cognitive
flexibility (斯科特, 1962). In our study we investigated whether individual differences in these
core facets of human intelligence are associated with differences in the intrinsic functional ar-
chitecture of the brain, operationalized as the intrinsic synchrony or connectivity of functional
网络. We predicted that greater synchrony within and among functional networks at rest
would predict better performance on measures of fluid intelligence and of cognitive flexibility.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
A
_
0
0
0
1
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
结果
Spectral Decomposition for Multivariate Pattern Identification
Spectral decomposition was used to determine the synchrony and structure of intrinsic brain
网络. Spectral decompositions are a family of closely related analyses that describe the
dominant components of a complex system. These include principal-component analysis
(PCA), singular value decomposition, and eigendecompositions. Spectral decomposition iden-
tifies the spatial patterns of functional synchrony across brain regions, as well as their archi-
tecture or hierarchical organization, as determined by the synchrony strength, or persistent
synchrony, of these networks. The resulting principal components (PCs) are ranked numeri-
cally according to their prominence within the resting-state architecture (Figure 1A). Hetero-
geneity in the subject-level PC topology was inversely related to singular values (IE。, PC rank);
PC 1 was the most homogeneous across the population, and PC 10 the most heterogeneous
(Figure 1B). The whole-brain networks described by RSFC PCs show features of well-
characterized neurocognitive systems (Fox et al., 2005; 史密斯等人。, 2009; Spreng, 墓,
车工, Stevens, & Schacter, 2013; Yeo et al., 2011). 例如, the group-mean RSFC
components show synchrony of sensory and motor cortices (PC 1), the default network (PC 2),
the salience network (ventral attention) relative to default regions (PC 3), the visual system
(PC 4), and the dorsal attention network (PC 5). PC 6 comprises motor and default regions.
PC 7 demonstrates a functionally independent mode of the left relative to right frontoparietal
网络神经科学
194
Aspects of intelligence reflected by network synchrony in the brain
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
A
_
0
0
0
1
0
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
数字 1. Principal components (PCs) of resting-state functional connectivity.
(A) Top ten PCs
(across-group FWE-corrected p < .05). Reliable positive and negative features are shown for each
(B) Correspondence of the most homologous single-
component. Color bars indicate t-values.
subject PCs to the group-average PCs, shown for 600 subjects from the Human Connectome Project
900-subject release. The least variation across individuals exists in PC1, and variation across indi-
vidual PC differences increases consecutively with PC order.
control network. PCs 8–10 represent frontoparietal functional ensembles, including posterior
attention regions as aspects of canonical intrinsic networks (Chen et al., 2013).
All subcortical structures (thalamus, caudate, putamen, pallidum, hippocampus, amygdala,
and nucleus accumbens) demonstrated bilateral positive associations with group-mean PCs 1
and 2.
In both PCs 3 and 5, amygdala and hippocampus were functionally correlated and
showed anticorrelation relative to all other subcortical structures. Subcortical functional asso-
ciations between the hemispheres were largely symmetric across principal components, with
the notable exception of PC 7.
In PC 7, a left-lateralized subcortical configuration corre-
sponded to left-lateralized patterns in cortical activity (Table 1).
Eigenvalue:
A singular-value representation for
the amount of system variance
explained by the corresponding
eigenvector within a basis set of
functions
Eigenvalues and Behavior
Correlation scores were calculated between cognitive measures and eigenvalues (i.e., mea-
sures of network synchrony) for PCs 1–10 (n = 600). PC eigenvalues demonstrated significant
correlations with numerous measures of cognitive performance when corrected for multiple
comparisons (Figure 2). As predicted, fluid intelligence demonstrated the greatest correla-
tions with functional network eigenvalues. All correlations between eigenvalues (measures
of within-network synchrony) and fluid intelligence were positive, indicating that greater
Network Neuroscience
195
Aspects of intelligence reflected by network synchrony in the brain
Table 1. Principal-component (PC) subcortical associations
Thalamus
Caudate
Amygdala
Hippocampus
Pallidum
Nucleus accumbens
Putamen
PC 1
L+ R+
L+ R+
L+ R+
L+ R+
L+ R+
L+ R+
L+ R+
PC 2
L+ R+
L+ R+
L+ R+
L+ R+
L+ R+
L+ R+
L+
PC 3
L+ R+
L+ R+
L– R–
L– R–
L+ R+
–
PC 4
L–
L– R–
L–
L+ R+
L–
–
L+ R+
L– R–
PC 5
L– R–
L– R–
L+ R+
–
L– R–
L– R–
L– R–
PC 6
PC 7
PC 8
PC 9
PC 10
L+ R+
L+
–
–
L+ R+
R+
L+ R+
–
L+ R+
L+
L+
L+
L+
L+ R+
–
–
L– R–
L– R–
–
R–
–
–
–
–
L+
–
–
–
L– R–
L– R–
L– R–
L– R–
–
–
L– R–
The table shows associations between PCs 1–10 and bilateral subcortical structures. Positive and negative functional
relationships with left (L) and right (R) subcortical structures are shown only for significant regions.
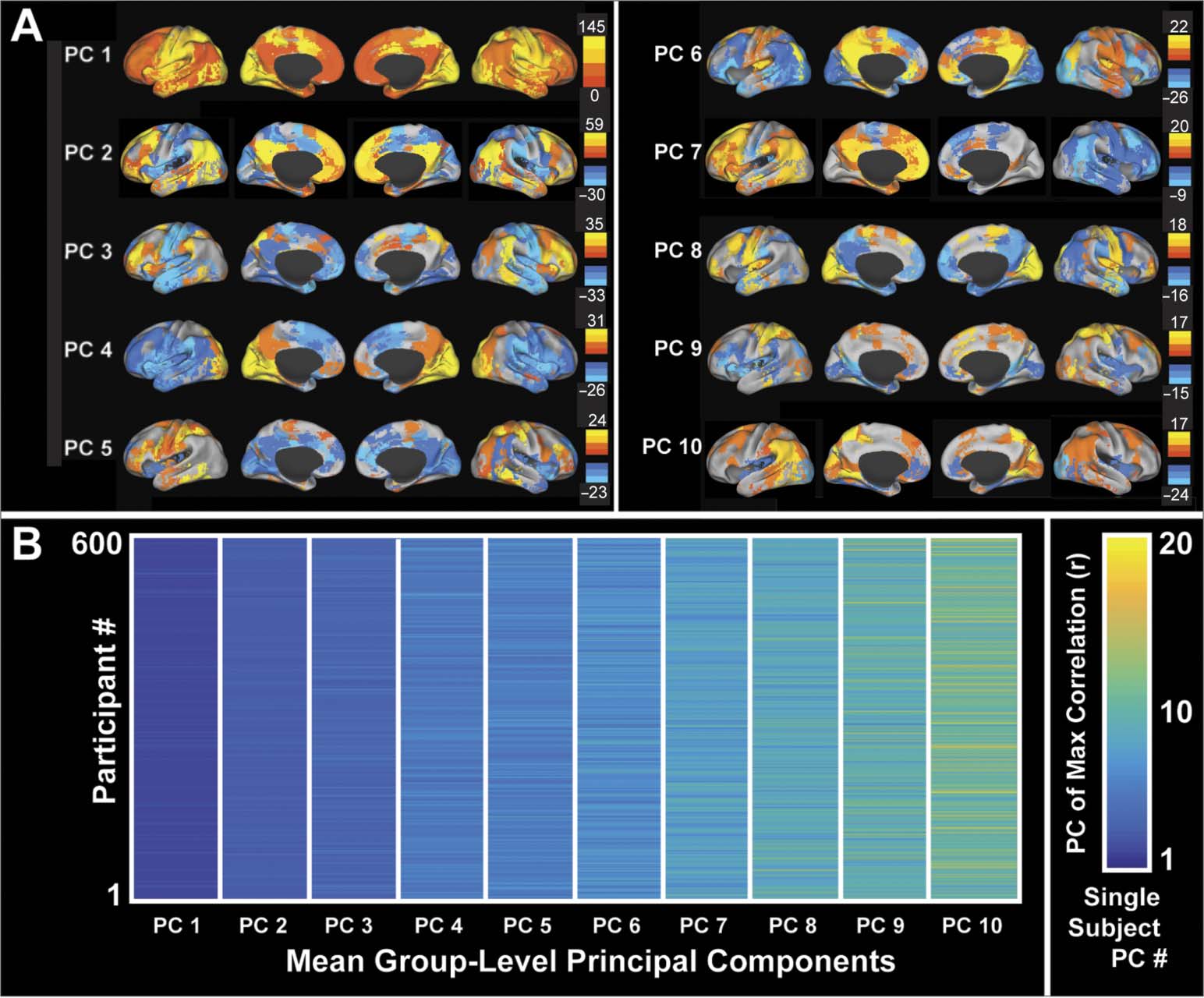
within-network synchrony across a range of networks is predictive of fluid intelligence (means
[95% CIs]: PC 3, r = 0.08 [0.015, 0.15]; PC 4, r = 0.13 [0.06, 0.19]; PC 5, r = 0.12 [0.05,
0.18]; PC 6, r = 0.09 [0.02, 0.15]; PC 7, r = 0.12 [0.06, 0.19]; PC 8, r = 0.14 [0.06, 0.21];
PC 9, r = 0.13 [0.07, 0.20]; PC 10, r = 0.13 [0.07, 0.20]; see Figure 2). Cognitive flexibility
demonstrated a negative correlation with the eigenvalue of PC 1, indicating that greater global
synchrony is associated with lower cognitive flexibility (r = –0.09, 95% CI [–0.16, –0.03];
Figure 2).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
a
_
0
0
0
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Cognitive measures and PC eigenvalues.
Spearman correlations were calculated
between cognitive performance measures and unscaled eigenvalues for PCs 1–10 (n = 600).
Correlations with 95% confidence intervals that do not cross 0 and that survive multiple-
comparison correction are highlighted. The cognitive measures demonstrating the broadest cor-
relations with PC eigenvalues are those for fluid intelligence. Because larger eigenvalues indicate
stronger within-network synchrony, the results demonstrate that increased within-network syn-
chrony for a broad range of PCs is positively correlated with fluid intelligence.
Network Neuroscience
196
Aspects of intelligence reflected by network synchrony in the brain
Scaled Eigenvalues and Behavior
To investigate the relationship between intellectual functioning and individual differences in
network hierarchies, we also correlated the cognitive performance measures with the scaled
eigenvalues for PCs 2–10. (Because the scaled eigenvalue for PC 1 is by definition equal to 1,
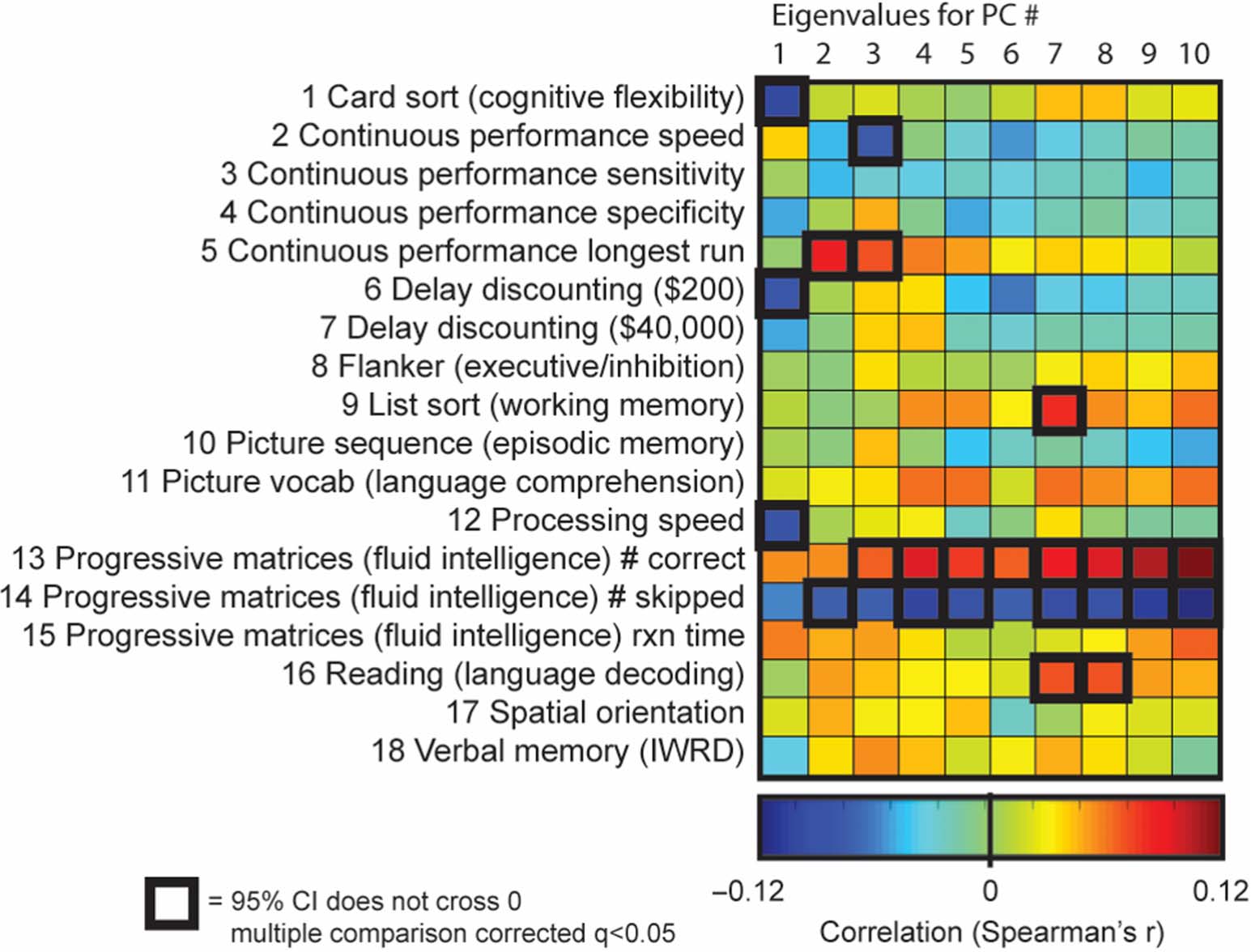
PC 1 is not included in the scaled-eigenvalue comparisons.) Cognitive flexibility demonstrated
positive correlations with each of the scaled eigenvalues for PCs 2 to 10 (means [95% CIs]:
PC 2, r = 0.09 [0.03, 0.16]; PC 3, r = 0.09 [0.03, 0.16]; PC 4, r = 0.10 [0.03, 0.16]; PC 5,
r = 0.09 [0.03, 0.17]; PC 6, r = 0.10 [0.03, 0.16]; PC 7, r = 0.10 [0.04, 0.17]; PC 8, r =
0.10 [0.04, 0.17]; PC 9, r = 0.10 [0.03, 0.17]; PC 10, r = 0.10 [0.03, 0.17]; Figure 3). This
indicates that cognitive flexibility is supported by persistent synchrony across a broad array of
functional networks. In contrast, as we identified above, global synchrony, or the baseline level
of sustained synchrony across the whole brain, is associated with lower cognitive flexibility.
A similar pattern was observed for processing speed, suggesting that both aspects of cognitive
functioning are enhanced by greater synchrony in specific networks but negatively impacted
by higher levels of global synchrony.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
a
_
0
0
0
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Cognitive measures and scaled eigenvalues for the PCs. Spearman correlations were
calculated between cognitive performance measures and the scaled eigenvalues for PCs 2–10 (n =
600). Scaled eigenvalues were calculated by dividing each PC’s eigenvalue by the eigenvalue for
PC 1 in each individual subject. Correlations with 95% confidence intervals that do not cross 0 and
that survive multiple-comparison correction are highlighted. The cognitive measures demonstrating
the broadest correlations with PCs are those for cognitive flexibility and processing speed. Larger
scaled eigenvalues indicate stronger within-network synchrony relative to the global synchrony of
an individual’s brain. Stronger within-network synchrony relative to the global synchrony (PC 1)
is positively correlated with increased cognitive flexibility and more rapid processing speed across
PCs 2–10.
Network Neuroscience
197
Aspects of intelligence reflected by network synchrony in the brain
Least absolute squares shrinkage
operator (LASSO):
A statistical learning method for
selecting predictive features from
large sets of independent variables
through iterative regression
Connectome:
Graph theory model for connections
across the brain; the connectome can
be represented as layers of spatially
overlapping functional networks
Network Neuroscience
Behavior/RSFC Network Interactions
Using a LASSO regression determined from a training set of 600 participants, 33 of the 256
possible unique combinations of network interactions were used as predictors for fluid intel-
ligence in an independent testing set (n = 230). The motivation for identifying combinations
of networks related to intelligence is the hypothesis that individual differences in intellectual
abilities may be represented better by the interactions of functional networks. A correlation
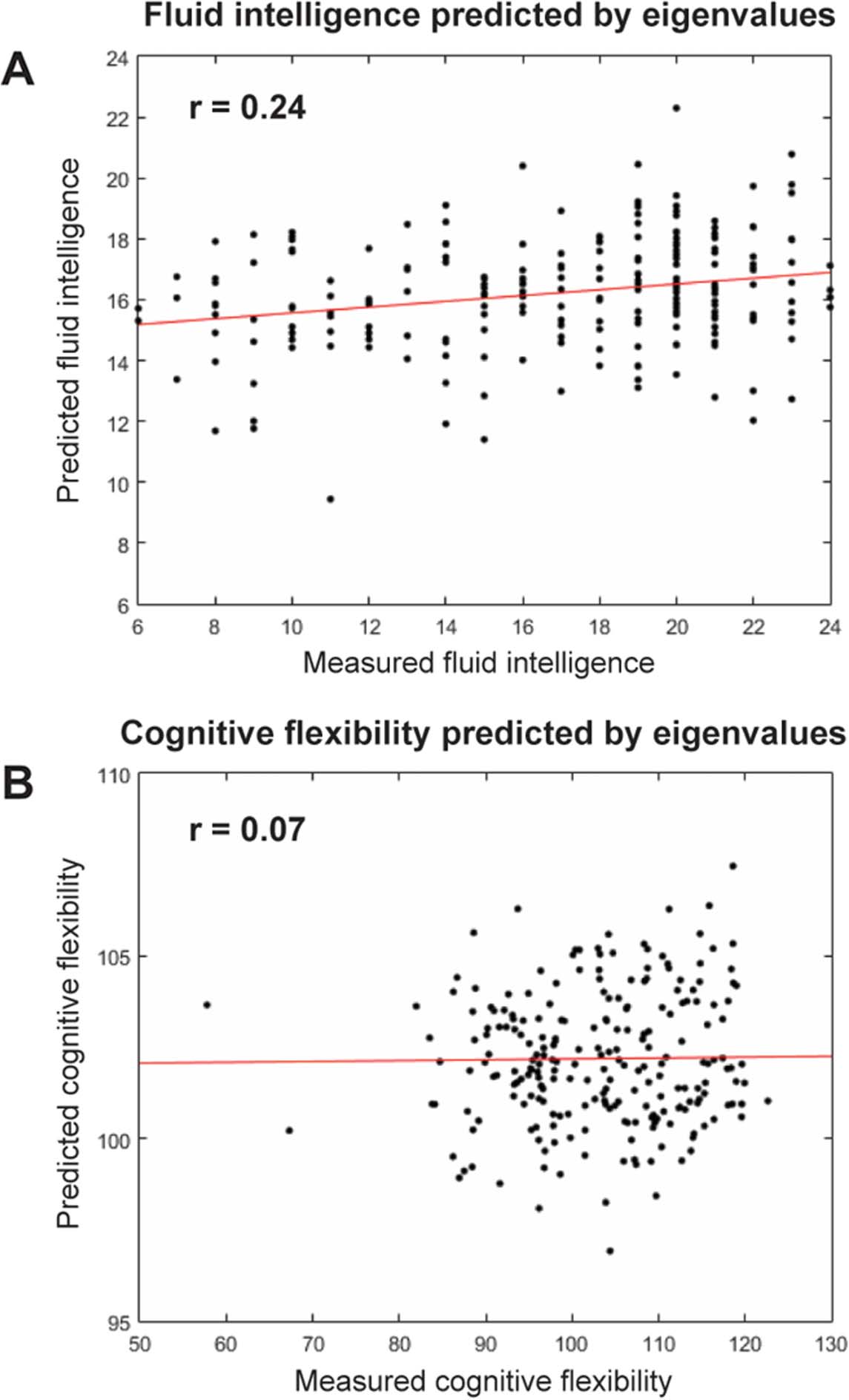
score of r = 0.24 (p < .001) between the measured and predicted values of fluid intelligence
was observed in the testing set (Figure 4A). For cognitive flexibility, 48 of the 1,024 possible
unique combinations of network interactions were selected by LASSO regression as the best-fit
model of cognitive flexibility from the PC eigenvalues. A correlation score of r = 0.07 (p = .28)
was observed between the predicted and measured scores of cognitive flexibility (Figure 4B).
However, predictions for cognitive flexibility based on eigenvalue products failed to reach sig-
nificance, indicating that the eigenvalue magnitude for PC 1—that is, the strength of global
synchrony—is a better predictor of cognitive flexibility than are models based on network in-
teractions. Together, these results suggest that fluid intelligence and cognitive flexibility are
associated with greater network stability or synchrony for the networks related to PCs 3–10.
In contrast, cognitive flexibility, but not fluid intelligence, is negatively associated with overall
synchrony, or stability, within the brain’s intrinsic functional architecture.
DISCUSSION
We investigated how individual differences in the intrinsic network architecture of the brain
are associated with human intelligence. Building from previous studies (Finn et al., 2015;
Hearne et al., 2016; Smith et al., 2013), here we applied spectral decomposition methods to
large-scale functional brain networks to determine whether spatially overlapping patterns of
synchrony within these networks corresponded with two core aspects of intellectual func-
tioning: fluid intelligence and cognitive flexibility. Both fluid intelligence and cognitive flex-
ibility were reliably associated with the functional architecture of intrinsic brain networks.
Synchrony within multiple functional networks reliably predicted fluid intelligence. Greater
relative synchrony of networks within the network architecture (i.e., scaled eigenvalues) was
associated with greater cognitive flexibility, whereas greater whole-brain baseline synchrony
levels were associated with reduced flexibility. These results are consistent with our prediction
that systems-level descriptors of the intrinsic functional architecture of the brain can provide
reliable markers of human intellectual functioning.
Properties of the brain’s intrinsic neural architecture have been used to predict a range of
cognitive capacities and may be a powerful predictor of more stable, or metacognitive, capaci-
ties such as intelligence (Stevens & Spreng, 2014). Previous reports examining the associations
between resting-state functional connectivity and intelligence have identified pairs of nodes
(i.e., network edges) that are predictive of cognitive ability using multiple regressions for fea-
ture selection (Finn et al., 2015; Hearne et al., 2016; Smith et al., 2013; Smith et al., 2015).
Conceptually, our approach is consistent with these studies, in the sense that we characterize
features of the brain’s functional architecture in order to identify large-scale distributed net-
works that support intellectual abilities. Our approach also builds upon these previous studies
in several key ways. First, we identified whole-brain distributed networks using spectral de-
composition applied to the resting-state connectome (Figure 1), rather than using pairwise
correlation values as the predictive features for our model. We then used the singular val-
ues (i.e., eigenvalues) of the first ten principal components (eigenvectors), representing the
persistence of within-network synchrony, to measure relationships between whole-network
198
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
a
_
0
0
0
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Aspects of intelligence reflected by network synchrony in the brain
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
a
_
0
0
0
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Predicting aspects of intelligence from resting-state functional connectivity A least-
absolute-squares shrinkage operator (LASSO) regression trained in a subsample of n = 600 pre-
dicted fluid intelligence in an independent testing set (n = 230) with a correlation of r = 0.24 (p <
0.001; panel A). Using the same training set (n = 600) and testing set (n = 230) subsamples, LASSO
regression predicted cognitive flexibility with a correlation of r = 0.07 (p = 0.28; panel B). The
differential in predictive power between fluid intelligence and cognitive flexibility indicates that the
spectral features associated with fluid intelligence represent more unique cognitive variance than
do the spectral features associated with cognitive flexibility.
synchrony and intelligence (Figures 2 and 3). This method supports the findings from previous
network approaches to identify the predictors of intelligence (Finn et al., 2015; Hearne et al.,
2016) and positive (or negative) real-life functioning (Smith et al., 2015). The present study
corroborates these earlier reports that whole-brain network connectivity and interactivity are
biomarkers of fluid intelligence (Finn et al., 2015; Hearne et al., 2016; Smith et al., 2015) and
Network Neuroscience
199
Aspects of intelligence reflected by network synchrony in the brain
supports the conclusion that its large-scale network architecture reflects individual differences
in human intellectual functioning.
As we noted above, singular values for PCs are interpreted as reflecting the temporal stabil-
ity of brain networks.
In simulations iterating brain activity from a set of initial conditions
based on functional connectivity between brain regions (Ferguson & Anderson, 2012), the
PCs of functional connectivity matrices by definition correspond to patterns of activity that
are invariant to change in a Markov model based on functional connectivity between regions.
We predicted that more temporally stable patterns of brain activity would be associated with
higher intelligence across a broad range of brain networks.
Consistent with this possibility, our results demonstrated that across-network interactions at
rest are predictive of fluid intelligence more strongly than are correlations with the synchrony
strength of any individual network (Figure 4). Cross-network interactions between the default
network and frontoparietal networks have previously been reported to correspond positively
with individual differences in fluid intelligence (Finn et al., 2015; Hearne et al., 2016). Our
results are compatible with these findings of across-network interactions supporting fluid in-
telligence, and they extend this cross-network interaction paradigm for fluid intelligence to
include a diverse set of resting-state functional networks (see Figures 1 and 4). Although our
findings do not exclude a disproportionate effect of some connections, such as frontoparietal
region pairs, which may have influenced many of the PCs in our analysis, the data suggest that
additional information contributes to fluid intelligence in across-network synchrony.
As we demonstrated here, spectral decomposition of resting-state functional connectivity is
consistent with a “one-to-many” functional architecture, with brain regions being implicated in
multiple PCs, or networks. It is widely accepted that single brain regions can be recruited dur-
ing multiple cognitive processes (McIntosh, 1999). One constraint of common decomposition
methods such as spatial independence (Beckmann et al., 2005) or a greedy, winner-take-all
algorithm (Yeo et al., 2011) is that they may obscure this one-to-many architecture. The PCA
approach used here to identify patterns of functional synchrony (i.e., intrinsic brain networks)
enables the identification of “promiscuous” brain regions that flexibly couple with multiple
networks, providing the neural substrate for distributed parallel processing, a necessary con-
dition for higher cognitive processes associated with intellectual functioning.
Structural research on neural contributions to intelligence has indicated that greater
efficiency of physical connections within networks is positively correlated with individ-
ual differences in cognitive performance (Li et al., 2009; Pineda-Pardo et al., 2015). These
findings may provide insight into the ways that individual differences in functional syn-
chrony might arise from variations in anatomical connections.
Integration of structural and
functional data indicates that both efficiency and synchrony characterize intelligent brains
(Pineda-Pardo et al., 2015). Greater efficiency in structural connectivity may directly con-
tribute to more persistent synchrony within functional networks, in turn producing greater
cognitive capacity. Future investigations, integrating multilayer maps that can incorporate
functional synchrony, structural efficiency, and other physiological or genetic factors, will be
necessary to identify truly systems-level biomarkers of human intelligence.
A major challenge in identifying neural makers of intellectual functioning has been the vari-
ability in research methodologies and in the operationalization of intelligence across studies
(Sternberg, 2005). A prominent approach has involved the investigation of domain-specific
processes associated with general intelligence, such as working memory capacity (Ackerman,
et al., 2005; Conway et al., 2003; DeYoung et al., 2009; Edgin et al., 2010). However,
Network Neuroscience
200
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
a
_
0
0
0
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Aspects of intelligence reflected by network synchrony in the brain
mounting neural evidence is challenging such domain-specific conceptualizations of intelli-
gence and supporting the existence of a more unitary or “multiple-demand” account (Duncan,
2010; Duncan et al., 2000) of fluid intellectual functioning, implicating the frontal parietal
control network (Vincent et al., 2008). Meta-analyses of fMRI studies provide partial sup-
port for this idea. As was suggested by Duncan (2010), multiple brain regions are com-
monly recruited during cognitive control tasks associated with fluid intelligence (Basten, 2015).
However, this multidemand network flexibly couples with other brain regions depending on
the specific task context, including perceptual, mnemonic, or motor output demands (e.g.,
Niendam et al., 2012). This hybrid model, combining cognitive control and fluid intellectual
functioning more broadly, is consistent with our findings, which also suggest that multiple brain
networks (PCs 3–10) couple with core frontoparietal brain regions, yet remain differentiated
by network-specific recruitment of brain regions outside the frontoparietal control network.
Patterns of persistent frontoparietal network synchrony are found in PCs 6, 7, and 10 and
correspond to previous evidence that fluid intelligence, or “g,” may be preferentially dependent
on the integrity of frontoparietal control networks and the underlying white-matter pathways
(Penke et al., 2012, and see above). Cognitive control processes linked to fluid intelligence
are necessary to form, reconfigure, or consolidate interpretive schema and are robustly linked
with frontoparietal functional anatomy (Cole et al., 2015; Cole et al., 2013; Cole et al., 2012;
Spreng et al., 2013). The positive correlations observed with the eigenvalues for these com-
ponents suggest that fluid intelligence is supported by the cooperation of multiple spatially
overlapping functional networks defined in our model, consistent with previous meta-analyses
(e.g., Niendam et al., 2012). Furthermore, individuals with higher levels of intellectual func-
tioning have greater brain “resilience,” such that those demonstrating more spatially distributed
patterns of neural recruitment during cognitive tasks may be less susceptible to age-related
brain changes or brain insult (Santarnecchi, Rossi, & Rossi, 2015). Our results here are also
consistent with this idea, demonstrating that an intrinsic functional architecture comprising
multiple, widely distributed brain networks, organized along a continuum of differentiated
network synchrony, may provide the necessary neural foundation for the expression, and pre-
sumably the preservation, of fluid intelligence.
Interestingly, the magnitude of intrinsic global synchrony (PC 1) was negatively associated
with cognitive flexibility (Figure 2). This may suggest, perhaps somewhat intuitively, that greater
whole-brain synchrony—conceptually related to stability—is associated with lower mental
flexibility. PC 1 is unique among the components in that it did not reflect some brain regions
as positive and other brain regions as negative, so differences specific to PC 1 in our behav-
ioral correlations may reflect global synchrony versus antagonistic or competing interactions
between brain regions reflected in successive PCs. Cognitive flexibility, as such, appears func-
tionally related to lower global synchrony and more network flexibility, a pattern of neural
activity positively associated with learning and health (Bassett et al., 2011; Braun et al., 2015).
We suggest that a flexible network architecture with interacting brain networks is necessary
for flexible thought and behavior, and may be a critical element of adaptive real-world func-
tioning. Investigating this association between network and behavioral flexibility and how it
changes as a result of aging and brain disease will represent an important area of future re-
search. However, this observation also highlights the importance of studying functional brain
networks, not simply in isolation, but also in relation to other networks, as well as studying
within-subject variability in global network characteristics, as we did here. By scaling individ-
ual network synchrony values by each individual’s global network synchrony, we were able to
Network Neuroscience
201
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
a
_
0
0
0
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Aspects of intelligence reflected by network synchrony in the brain
identify robust correlations between these scaled eigenvalues and cognitive flexibility for all
identified networks (PCs 2–10 in Figure 3).
Our findings suggesting that greater network synchrony is associated with higher fluid in-
telligence are consistent with reports that the brain network configuration at rest is closely
aligned with task-driven network configurations in individuals of higher intelligence Schultz,
2016. These resting-state (or intrinsic) connections are hypothesized to reflect the persistence
of task-based synchrony patterns formed through recurrent coactivation of distributed brain
regions occurring over the course of development (Stevens & Spreng, 2014). These patterns
of coactivation may serve as the brain’s “ready state,” potentiating task-driven activity among
regions that commonly work in concert (Fair et al., 2007). The functional network organization
associated with higher cognitive functioning may position individuals in an optimized “ready
stance” that is prepared to engage cognitive tasks. However, future research will be necessary
to more directly investigate this relationship.
These measures describe spectral components of RSFC as novel features of the brain’s func-
tional architecture. Here we have shown that individual differences in intrinsic connectivity
predict core aspects of human intelligence. Mapping intelligence in the human brain with
RSFC offers a novel approach to investigating psychological functioning in health and disease.
Alterations in the absolute and relative prominences of networks within the brain’s intrinsic
functional architecture may help predict individual differences in normal cognitive function-
ing, as was demonstrated here, or predict patterns of altered cognitive abilities in neurodevel-
opment, normal aging, and brain disease.
METHODS
Resting-State Functional Connectivity
The Human Connectome Project (HCP) is an initiative by the National Institutes of Health
to generate large, open-access behavioral and fMRI datasets. RSFC was preprocessed and
analyzed for 830 subjects (mean age = 28.8 years, SD = 3.9, range = 22–37; 465 women)
from the Human Connectome Project (www.humanconnectome.org; HCP900 release). The
subjects were selected on the basis of having four complete resting-state scans and com-
plete behavioral metrics for the cognitive features of interest. The BOLD fMRI data were
acquired in four 15-min blocks. For the analysis, we used data cleaned using the FIX soft-
ware (Glasser et al., 2013; Griffanti et al., 2014; Moeller et al., 2010; Setsompop et al., 2012;
Van Essen et al., 2013).
The cerebral cortex was parcellated into 333 functionally defined regions (Gordon et al.,
2014). Fourteen subject-specific subcortical regions were added using Freesurfer-derived seg-
mentation (Fischl et al., 2002) of bilateral thalamus, caudate, putamen, amygdala, hippocam-
pus, pallidum, and nucleus accumbens. Fourteen cerebellar regions were also added (Buckner
et al., 2011). This combined parcellation scheme covering the full cortex, subcortical struc-
tures, and the cerebellum comprised a total of 361 regions. BOLD time series for each region of
interest (ROI) were extracted, and Fisher’s r-to-z transformed Pearson correlation coefficients
were obtained for each pair of ROIs in each 15-min block for each subject. The resulting
361 × 361 matrices were averaged across the four blocks for each subject and subsequently
averaged across all subjects to obtain a group-level functional connectivity matrix.
Spectral decomposition of RSFC data produces functionally orthogonal principal compo-
nents (PCs). These components are synonymous with eigenvectors of RSFC matrices and iden-
tify covariance patterns in the functional brain data. As such, PCs from group-mean and
Network Neuroscience
202
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
a
_
0
0
0
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Aspects of intelligence reflected by network synchrony in the brain
single-subject RSFC matrices represent a set of hierarchically organized intrinsic brain net-
works, ranked by the amount of signal variance described by each component. A functional
network thus defined is a set of multivariate patterns that explain persistent synchrony be-
tween distributed brain regions during rest. In addition to identifying RSFC networks, spectral
decomposition also calculates a singular value, alternatively called an eigenvalue, for each
principal component vector. These are single numerical scores for the principal-component
vector length and are proportional to the amount of total variance in the RSFC matrix accounted
for by that component.
Principal components were identified using singular value decomposition of the 361 × 361
functional connectivity matrices (corresponding to the cortical-and-subcortical parcellation
scheme; see above). The first ten PCs were calculated from the group mean connectivity matrix
and back-projected onto anatomical space (Figure 1A). The first 20 PCs were identified in the
same manner for each subject. Spatial correlations between the individual and group PCs
were calculated using Pearson correlations. The single-subject PCs with the highest spatial
correlation to group PCs 1–10 were determined, in order to assess the homogeneity of the
single-subject PCs across the population (Figure 1B). The homogeneity of PCs is an indicator
of network stability and replicability across the study population. We selected the most stable,
or homogeneous, PCs in our sample for the subsequent brain and behavioral analyses—that
is, PCs 1 to 10. The reliability of the PC network architecture across the group was assessed
by one-sample t tests (Figure 1A), family-wise error-corrected for multiple comparisons (p <
0.05).
Network synchrony, the core measure of network architecture in the present study, is repre-
sented by the eigenvalue of each PC. We also calculate a scaled eigenvalue for each PC, rep-
resenting the relative dominance of that network within the network hierarchy. All scaled
eigenvalues represent the connectivity strength of that network relative to the dominant com-
ponent (PC 1). PCs with higher eigenvalues (and scaled eigenvalues) are more prominent
within an individual’s resting-state network architecture. Greater prominence suggests that the
regions that compose a functional network demonstrate more persistently synchronous activity
across the resting-state time series and explain more of the variance of temporal fluctuations
than do lower-ranked networks. PC 1 constitutes the highest-ranked component and is re-
ferred to as the global synchrony component. As we noted above, all scaled eigenvalues are
scaled relative to this global synchrony component and reflect individual differences in the
organization, or relative strengths, of networks within the overall network hierarchy.
Behavioral Data
We used the HCP behavioral measures related to cognition for our analysis. The battery
includes measures of cognitive flexibility (Gershon et al., 2013), continuous performance
(Gur et al., 2001), delay discounting (Estle, Green, Myerson, & Holt, 2006), executive inhibi-
tion (Gershon et al., 2013), working memory (Gershon et al., 2013), episodic memory
(Gershon et al., 2013), language comprehension (Gershon et al., 2013), processing speed
(Gershon et al., 2013), fluid intelligence (Bilker et al., 2012), language decoding (Gershon
et al., 2013), spatial orientation (Gur et al., 2001), and verbal memory (Gur et al., 2001). These
HCP behavioral measures largely consist of tests developed and validated for the NIH Toolbox
(Gershon et al., 2013). A full description of these measures, including assessment protocols,
normative data samples, and psychometric properties, may be found at www.healthmeasures.
net/explore-measurement-systems/nih-toolbox). Additional measures were included in the
HCP behavioral battery to further explore the dimensions of human social, emotional,
Network Neuroscience
203
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
a
_
0
0
0
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Aspects of intelligence reflected by network synchrony in the brain
and cognitive function.
For full data on these additional measures, please see www.
humanconnectome.org; HCP900 release. Behavioral data were collected from HCP partner
sites, including Washington University and the University of Minnesota.
Behavior/RSFC Eigenvalue and Scaled-Eigenvalue Correlations
To investigate whether individual differences in resting-state functional synchrony reliably pre-
dicted intellectual functioning, we divided the available 830 participants’ data into two sets:
a training set (n = 600) and a testing set (n = 230). Spearman correlation coefficients were
calculated between the individual subjects’ scores for behavior and the eigenvalues for PCs
1–10 in the training set (n = 600, Figure 2) to allow for nonnormal distributions in the behavior
scores. As described above, we calculated the scaled eigenvalue, based on each individual’s
global synchrony component, for each network component by dividing all the eigenvalues in
an individual’s resting-state network profile by the eigenvalue of PC 1. The scaled eigenvalues
for PCs 2–10 were correlated with the cognitive measures using Spearman correlation (n =
600, Figure 3). We report all correlations with 95% confidence intervals that did not include
zero and with p-values that survived multiple-comparison correction (Figure 3).
Behavior/RSFC Network Interactions
Given our overall prediction that human intelligence may be best reflected in the dynamic net-
work architecture of the brain, we hypothesized that fluid intelligence and cognitive flexibility
may be more strongly associated with dynamic network interactions than with synchrony sim-
ply within specific networks. As such, we undertook a secondary brain and behavioral analysis,
using the products of individual eigenvalues, rather than a linear model, to predict cognitive
performance.
We used a least-absolute-squares shrinkage operator (LASSO) with the training set (n = 600)
to optimize the selection of network interactions—that is, the eigenvalue product terms—to be
used in the predictor model. The designation of 600 participants in the training set and 230 par-
ticipants in the testing set was based on a 70%–30% partition scheme for machine-learning
predictions (Weinberger, Blitzer, & Saul, 2005). We evaluated all possible combinations of
networks for the RSFC principal components that correlated significantly with the cognitive
measures (see above). Eight significant PC features met these criteria for fluid intelligence
(Figure 2), and ten significant PC features met these criteria for cognitive flexibility (Figures 2
and 3). As such, there were 256 possible unique combinations of network interactions for
networks covarying with fluid-intelligence features (28), and 1,024 possible combinations of
network interactions for networks covarying with cognitive-flexibility features (210). LASSO
regression (MATLAB, version 2012b, statistical toolbox) optimized the data-fitting between
eigenvalue products and fluid-intelligence scores in the training set (n = 600) to 33 unique
combinations of network interactions that could be used as predictor terms. Coefficients and
intercepts from the training set for these 33 network interactions were used as predictors in
the independent testing set (n = 230). Of the 1,024 possible network interactions between the
cognitive-flexibility RSFC features, LASSO regression optimized the data-fitting in the training
set to 48 unique combinations of network interactions that could be used as predictor terms.
The coefficients and intercepts for these interactions were used to calculate predicted values for
fluid intelligence and cognitive flexibility in the independent testing set (n = 230). The volume
of an abstract, n-dimensional eigensubspace (a.k.a., a “hypervolume”) may be determined by
the products of orthogonal eigenvector magnitude—that is, by multiplying eigenvalues. Ulti-
mately, each of the various eigensubspace terms identified by LASSO may prove to represent
Network Neuroscience
204
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
a
_
0
0
0
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Aspects of intelligence reflected by network synchrony in the brain
a discrete, dissociable neurobiological contribution to the emergent phenomenon of general
intelligence.
ACKNOWLEDGMENTS
the NIH Blueprint
Data were provided by the Human Connectome Project, WU–Minn Consortium (PIs: David
Van Essen and Kamil Ugurbil; Grant No. 1U54MH091657), funded by the 16 NIH institutes
for Neuroscience Research; and by the
and centers that support
McDonnell Center for Systems Neuroscience at Washington University. Support for the anal-
ysis was provided by the National Institute of Mental Health (Grant No. K08 MH092697)
and the Alzheimer’s Association (Grant No. NIRG-14-320049) to R.N.S. We are grateful to
Elizabeth DuPre, Karen Spreng, and Gary R. Turner for helpful advice and discussion in the
writing of the manuscript.
AUTHOR CONTRIBUTIONS
Michael A. Ferguson: Conceptualization; Data curation; Formal analysis; Investigation;
Methodology; Software; Visualization; Writing – original draft
Jeffrey S. Anderson: Data curation; Formal analysis; Methodology; Writing – review & editing
R. Nathan Spreng: Conceptualization; Methodology; Supervision; Writing – review & editing
REFERENCES
Ackerman, P. L., Beier, M. E., & Boyle, M. O.
(2005). Work-
ing memory and intelligence: The same or different constructs?
Psychological Bulletin, 131, 30–60. doi:10.1037/0033-2909.
131.1.30
Alter, O., Brown, P. O., & Botstein, D. (2000). Singular value de-
composition for genome-wide expression data processing and
modeling. Proceedings of the National Academy of Sciences,
97, 10101–10106.
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
(2011). Dynamic reconfiguration of hu-
J. M., & Grafton, S. T.
man brain networks during learning. Proceedings of the National
Academy of Sciences, 108, 7641–7646.
Basten, U., Hilger, K., & Fiebach, C. J. (2015). Where smart brains
are different: A quantitative meta-analysis of functional and struc-
Intelligence, 51,
tural brain imaging studies on intelligence.
10–27.
Beckmann, C. F., DeLuca, M., Devlin, J. T., & Smith, S. M. (2005).
Investigations into resting-state connectivity using independent
component analysis. Philosophical Transactions of the Royal
Society B, 360, 1001–1013. doi:10.1098/rstb.2005.1634
Bilker, W. B., Hansen, J. A., Brensinger, C. M., Richard, J., Gur,
R. E., & Gur, R. C. (2012). Development of abbreviated nine-
item forms of the Raven’s standard progressive matrices test.
Assessment, 19, 354–369. doi:10.1177/1073191112446655
Braun, U., Muldoon, S. F., & Bassett, D. S.
(2015). On human
brain networks in health and disease. eLS, 1–9. doi:10.1002/
9780470015902.a0025783
Buckner, R. L., Krienen, F. M., Castellanos, A., Diaz, J. C., & Yeo,
(2011). The organization of the human cerebellum esti-
Journal of Neuro-
B. T.
mated by intrinsic functional connectivity.
physiology, 106, 2322–2345. doi:10.1152/jn.00339.2011
Bullmore, E., Barnes, A., Bassett, D. S., Fornito, A., Kitzbichler, M.,
Meunier, D., & Suckling, J. (2009). Generic aspects of complexity
in brain imaging data and other biological systems. NeuroImage,
47, 1125–1134. doi:10.1016/j.neuroimage.2009.05.032
Bullmore, E., Rabe-Hesketh, S., Morris, R. G., Williams, S. C.,
Gregory, L., Gray, J. A., & Brammer, M. J. (1996). Functional mag-
netic resonance image analysis of a large-scale neurocognitive
network. NeuroImage, 4, 16–33. doi:10.1006/nimg.1996.0026
Chen, A. C., Oathes, D. J., Chang, C., Bradley, T., Zhou, Z. W.,
Williams, L. M., . . . Etkin, A. (2013). Causal interactions between
fronto-parietal central executive and default-mode networks in
humans. Proceedings of the National Academy of Sciences, 110,
19944–19949. doi:10.1073/pnas.1311772110
Churchland, P. S. (1986). Neurophilosophy: Toward a unified sci-
ence of the mind-brain. Cambridge, MA: MIT Press.
Cole, M. W., Ito, T., & Braver, T. S. (2015). Lateral prefrontal cortex
contributes to fluid intelligence through multinetwork connec-
tivity. Brain Connections, 5, 497–504. doi:10.1089/brain.2015.
0357
Cole, M. W., Reynolds, J. R., Power, J. D., Repovs, G., Anticevic,
A., & Braver, T. S.
(2013). Multi-task connectivity reveals flex-
ible hubs for adaptive task control. Nature Neuroscience, 16,
1348–1355. doi:10.1038/nn.3470
Cole, M. W., Yarkoni, T., Repovs, G., Anticevic, A., & Braver, T. S.
(2012). Global connectivity of prefrontal cortex predicts cog-
Journal of Neuroscience, 32,
nitive control and intelligence.
8988–8999. doi:10.1523/JNEUROSCI.0536-12.2012
Conway, A. R. A., Kane, M. J., & Engle, R. W.
(2003). Working
memory capacity and its relation to general intelligence. Trends
in Cognitive Sciences, 7, 547–552. doi:10.1016/j.tics.2003.10.
005
Network Neuroscience
205
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
a
_
0
0
0
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Aspects of intelligence reflected by network synchrony in the brain
Deary, I. J., Penke, L., & Johnson, W. (2010). The neuroscience of
human intelligence differences. Nature Reviews Neuroscience,
11, 201–211. doi:10.1038/nrn2793
DeYoung, C. G., Shamosh, N. A., Green, A. E., Braver, T. S., & Gray,
J. R. (2009). Intellect as distinct from openness: Differences re-
Journal of Personality and
vealed by fmri of working memory.
Social Psychology, 97, 883–892. doi:10.1037/a0016615
Duncan, J. (2005). Frontal lobe function and general intelligence:
Why it matters. Cortex, 41, 215–217.
Duncan,
J.
(2010). The multiple-demand (MD) system of
the
primate brain: Mental programs for intelligent behaviour. Trends
in Cognitive Sciences, 14, 172–179. doi:10.1016/j.tics.2010.01.
004
Duncan, J., Seitz, R. J., Kolodny, J., Bor, D., Herzog, H., Ahmed,
A., . . . Emslie, H. (2000). A neural basis for general intelligence.
Science, 289, 457–460. doi:10.1126/science.289.5478.457
Edgin, J. O., Pennington, B. F., & Mervis, C. B.
(2010). Neuro-
psychological components of intellectual disability: The con-
tributions of
immediate, working, and associative memory.
Journal of
Intellectual Disability Research, 54, 406–417.
doi:10.1111/j.1365-2788.2010.01278.x
Estle, S. J., Green, L., Myerson, J., & Holt, D. D. (2006). Differen-
tial effects of amount on temporal and probability discounting of
gains and losses. Memory & Cognition, 34, 914–928. doi:10.
3758/BF03193437
Fair, D. A., Schlaggar, B. L., Cohen, A. L., Miezin, F. M.,
Dosenbach, N. U., Wenger, K. K., . . . Petersen, S. E.
(2007).
A method for using blocked and event-related fMRI data to
study “resting state” functional connectivity. NeuroImage, 35,
396–405. doi:10.1016/j.neuroimage.2006.11.051
Ferguson, M. A., & Anderson, J. S. (2012). Dynamical stability of
intrinsic connectivity networks. NeuroImage, 59, 4022–4031.
doi:10.1016/j.neuroimage.2011.10.062
Finn, E. S., Shen, X., Scheinost, D., Rosenberg, M. D., Huang, J.,
Chun, M. M., . . . Constable, R. T. (2015). Functional connectome
fingerprinting: Identifying individuals using patterns of brain con-
nectivity. Nature Neuroscience, 18, 1664–1671. doi:10.1038/
nn.4135
Fischl, B., Salat, D. H., Busa, E., Albert, M., Dieterich, M.,
(2002). Whole brain segmen-
Haselgrove, C., . . . Dale, A. M.
tation: Automated labeling of neuroanatomical structures in the
human brain. Neuron, 33, 341–355.
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen,
D. C., & Raichle, M. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks. Pro-
ceedings of the National Academy of Sciences, 102, 9673–9678.
doi:10.1073/pnas.0504136102
Friston, K. J., Frith, C. D., Fletcher, P., Liddle, P. F., & Frackowiak,
(1996). Functional topography: Multidimensional scal-
R. S.
ing and functional connectivity in the brain. Cerebral Cortex, 6,
156–164.
Furnham, A. (2008). Personality and intelligence at work: Explor-
ing and explaining individual differences at work. London, UK:
Routledge.
Gershon, R. C., Wagster, M. V., Hendrie, H. C., Fox, N. A., Cook,
K. F., & Nowinski, C. J. (2013). NIH toolbox for assessment of
neurological and behavioral function. Neurology, 80, S2–6. doi:
10.1212/WNL.0b013e3182872e5f
Glasser, M. F., Sotiropoulos, S. N., Wilson, J. A., Coalson, T. S.,
. . . Consortium, W. U.-M. H.
Fischl, B., Andersson,
(2013). The minimal preprocessing pipelines for the Human
Connectome Project. NeuroImage, 80, 105–124. doi:10.1016/
j.neuroimage.2013.04.127
J. L.,
Gordon, E. M., Laumann, T. O., Adeyemo, B., Huckins, J. F., Kelley,
W. M., & Petersen, S. E. (2014). Generation and evaluation of a
cortical area parcellation from resting-state correlations. Cerebral
Cortex, bhu239.
Gottfredson, L. S., & Deary, I. (2004). Intelligence predicts health
and longevity, but why? Current Directions in Psychological
Science, 13, 1–4.
Gray, J. R., Chabris, C. F., & Braver, T. S. (2003). Neural mechanisms
of general fluid intelligence. Nature Neuroscience, 6, 316–322.
doi:10.1038/nn1014
Gregory, R. L., & Zangwill, O. L. (1987). The Oxford companion
to the mind. Oxford, UK: Oxford University Press.
Griffanti, L., Salimi-Khorshidi, G., Beckmann, C. F., Auerbach, E. J.,
Douaud, G., Sexton, C. E., . . . Smith, S. M. (2014). ICA-based
artefact removal and accelerated fMRI acquisition for improved
resting state network imaging. NeuroImage, 95, 232–247. doi:
10.1016/j.neuroimage.2014.03.034
Gur, R. C., Moberg, P. D., Turner, T. H., Bilker, W. B., Kohler, C.,
(2001). Computerized neurocogni-
Siegel, S. J., & Gur, R. E.
tive scanning: I. Methodology and validation in healthy people.
Neuropsychoparmacology, 25, 766–776.
Hearne, L. J., Mattingley, J. B., & Cocchi, L. (2016). Functional brain
networks related to individual differences in human intelligence
at rest. Scientific Reports, 6, 32328. doi:10.1038/srep32328
Jung, R. E., & Haier, R. J.
(2007). The Parieto-Frontal Integra-
tion Theory (P-FIT) of intelligence: Converging neuroimaging evi-
dence. Behavioral and Brain Sciences, 30, 135–154; discussion
154–187. doi:10.1017/S0140525X07001185
Kuncel, N. R., Hezlett, S. A., & Ones, D. S. (2004). Academic per-
formance, career potential, creativity, and job performance: can
one construct predict them all? Journal of Personality and Social
Psychology, 86, 148–161. doi:10.1037/0022-3514.86.1.148
Langeslag, S. J., Schmidt, M., Ghassabian, A.,
Jaddoe, V. W.,
Hofman, A., van der Lugt, A., . . . White, T. J. (2013). Functional
connectivity between parietal and frontal brain regions and intel-
ligence in young children: The generation R study. Human Brain
Mapping, 34, 3299–3307. doi:10.1002/hbm.22143
Lee, K. H., Choi, Y. Y., Gray, J. R., Cho, S. H., Chae, J. H., Lee,
S., & Kim, K. (2006). Neural correlates of superior intelligence:
Stronger recruitment of posterior parietal cortex. NeuroImage,
29, 578–586. doi:10.1016/j.neuroimage.2005.07.036
Li, Y., Liu, Y., Li, J., Qin, W., Li, K., Yu, C., & Jiang, T.
(2009).
Brain anatomical network and intelligence. PLoS Computational
Biology, 5, e1000395.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
a
_
0
0
0
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
McIntosh, A. R.
(1999). Mapping cognition to the brain through
interactions. Memory, 7, 523–548. doi:10.1080/
neural
096582199387733
Moeller, S., Yacoub, E., Olman, C. A., Auerbach, E., Strupp, J.,
(2010). Multiband multislice GE-EPI
Harel, N., & Ugurbil, K.
Network Neuroscience
206
Aspects of intelligence reflected by network synchrony in the brain
at 7 tesla, with 16-fold acceleration using partial parallel imag-
ing with application to high spatial and temporal whole-brain
fMRI. Magnetic Resonance Medicine, 63, 1144–1153. doi:10.
1002/mrm.22361
Niendam, T. A., Laird, A. R., Ray, K. L., Dean, Y. M., Glahn, D. C., &
Carter, C. S.
(2012). Meta-analytic evidence for a super-
ordinate cognitive control network subserving diverse executive
functions. Cognitive, Affective, & Behavioral Neuroscience, 12,
241–268.
Penke, L., Maniega, S. M., Bastin, M. E., Hernandez, M. V., Murray,
C., Royle, N. A., . . . Deary, I. J. (2012). Brain white matter tract
integrity as a neural foundation for general intelligence. Molec-
ular Psychiatry, 17, 1026–1030.
Pineda-Pardo, J. A., Martínez, K., Román, F. J., & Colom, R. (2016).
Structural efficiency within a parieto-frontal network and cogni-
tive differences. Intelligence, 54, 105–116.
Santarnecchi, E., Rossi, S., & Rossi, A.
(2015). The smarter, the
stronger:
Intelligence level correlates with brain resilience to
systematic insults. Cortex, 64, 293–309. doi:10.1016/j.cortex.
2014.11.005
Schultz, D. H., & Cole, M. W. (2016). Higher intelligence is asso-
ciated with less task-related brain network reconfiguration. Jour-
nal of Neuroscience, 36, 8551–8561. doi:10.1523/JNEUROSCI.
0358-16.2016
Schutte, N. S., & Loi, N. (2014). Connections between emotional
intelligence and workplace flourishing. Personality and Individ-
ual Differences, 66, 134–139.
Scott, W. A. (1962). Cognitive complexity and cognitive flexibility.
American Sociological Association, 25, 405–414.
Setsompop, K., Gagoski, B. A., Polimeni, J. R., Witzel, T., Wedeen,
(2012). Blipped-controlled aliasing in par-
V. J., & Wald, L. L.
allel imaging for simultaneous multislice echo planar imaging
with reduced g-factor penalty. Magnetic Resonance Medicine,
67, 1210–1224. doi:10.1002/mrm.23097
Smith, S. M., Beckmann, C. F., Andersson, J., Auerbach, E. J.,
Bijsterbosch, J., Douaud, G., . . . Glasser, M. F. (2013). Resting-
state fMRI in the Human Connectome Project. NeuroImage, 80,
144–168. doi:10.1016/j.neuroimage.2013.05.039
Smith, S. M., Fox, P. T., Miller, K. L., Glahn, D. C., Fox, P. M.,
(2009). Correspon-
. Beckmann, C. F.
Mackay, C. E.,
dence of the brain’s functional architecture during activation and
rest. Proceedings of the National Academy of Sciences, 106,
13040–13045. doi:10.1073/pnas.0905267106
.
.
Smith, S. M., Nichols, T. E., Vidaurre, D., Winkler, A. M., Behrens,
T. E., Glasser, M. F., . . . Miller, K. L. (2015). A positive-negative
mode of population covariation links brain connectivity, demo-
graphics and behavior. Nature Neuroscience, 18, 1565–1567.
Song, M., Zhou, Y., Li, J., Liu, Y., Tian, L., Yu, C., & Jiang, T. (2008).
Brain spontaneous functional connectivity and intelligence.
NeuroImage, 41, 1168–1176. doi:10.1016/j.neuroimage.2008.
02.036
Spearman, C.
(1904). “General intelligence,” objectively deter-
mined and measured. American Journal of Psychology, 15,
201–292.
Spearman, C. (1927). The abilities of man. London, UK: Macmillan.
(2004).
Sporns, O., Chialvo, D. R., Kaiser, M., & Hilgetag, C. C.
Organization, development and function of complex brain net-
works. Trends in Cognitive Sciences, 8, 418–425. doi:10.1016/
j.tics.2004.07.008
Spreng, R. N., Sepulcre,
J., Turner, G. R., Stevens, W. D., &
Schacter, D. L. (2013). Intrinsic architecture underlying the re-
lations among the default, dorsal attention, and frontoparietal
control networks of the human brain. Journal of Cognitive Neu-
roscience, 25, 74–86. doi:10.1162/jocn_a_00281
Sternberg, R. J., & Detterman, D. K. (1986). What is intelligence?
Contemporary viewpoints on its nature and definition. Norwood,
NJ: Ablex.
Stevens, W. D., & Spreng, R. N.
(2014). Resting-state functional
connectivity MRI reveals active processes central to cognition.
Wiley Interdisciplinary Reviews: Cognitive Science, 5, 233–245.
doi:10.1002/wcs.1275
van den Heuvel, M. P., Stam, C. J., Kahn, R. S., & Hulshoff Pol, H. E.
(2009). Efficiency of functional brain networks and intellectual
performance. Journal of Neuroscience, 29, 7619–7624. doi:10.
1523/JNEUROSCI.1443-09.2009
Van Essen, D. C., Smith, S. M., Barch, D. M., Behrens, T. E.,
Yacoub, E., Ugurbil, K., & the WU–Minn H.C.P. Consortium.
(2013). The WU–Minn Human Connectome Project: An over-
view. NeuroImage, 80, 62–79. doi:10.1016/j.neuroimage.2013.
05.041
Vincent, J. L., Kahn, I., Snyder, A. Z., Raichle, M. E., & Buckner,
R. L. (2008). Evidence for a frontoparietal control system revealed
by intrinsic functional connectivity. Journal of Neurophysiology,
100, 3328–3342. doi:10.1152/jn.90355.2008
Weinberger, K. Q., Blitzer, J., & Saul, L. K. (2006). Distance met-
ric learning for large margin nearest neighbor classification. In
Y. Weiss, B. Schoelkopf, & J. Platt (Eds.), Advances in neural in-
formation processing systems (pp. 1473–1480). Cambridge, MA:
MIT Press.
Yeo, B. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D.,
Hollinshead, M., . . . Buckner, R. L. (2011). The organization of
the human cerebral cortex estimated by intrinsic functional con-
nectivity. Journal of Neurophysiology, 106, 1125–1165. doi:10.
1152/jn.00338.2011
Yeung, M. K., Tegner, J., & Collins, J. J. (2002). Reverse engineering
gene networks using singular value decomposition and robust re-
gression. Proceedings of the National Academy of Sciences, 99,
6163–6168. doi:10.1073/pnas.092576199
Network Neuroscience
207
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
1
2
1
9
2
1
0
9
1
9
2
5
n
e
n
_
a
_
0
0
0
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3