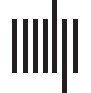研究
Age-related differences in functional brain
network segregation are consistent with
a cascade of cerebrovascular, 结构性的,
and cognitive effects
Tania S. Kong1,2, Caterina Gratton 3,4, Kathy A. 低的 1, Chin Hong Tan 1,5,6,
Antonio M. Chiarelli
1,7, 马克A. 弗莱彻
1, Benjamin Zimmerman 1,
Edward L. Maclin 1, Bradley P. Sutton 1,8, Gabriele Gratton 1,2, and Monica Fabiani
1,2
1Beckman Institute, University of Illinois at Urbana-Champaign, 伊尔, 美国
2Psychology Department, University of Illinois at Urbana-Champaign, 伊尔, 美国
3心理学系, Northwestern University, 伊尔, 美国
4神经内科, Northwestern University, 伊尔, 美国
5Division of Psychology, Nanyang Technological University, 新加坡
6Department of Pharmacology, National University of Singapore, 新加坡
7Department of Neuroscience, Imaging and Clinical Sciences, University G. D’Annunzio of Chieti-Pescara, Chieti, 意大利
8Department of Bioengineering, University of Illinois at Urbana-Champaign, 伊尔, 美国
关键词: 老化, Resting-state functional connectivity (rsFC), Cerebrovascular health, Optical brain
arterial pulse (pulse-DOT), White matter signal abnormalities (WMSAs), Cortical thickness
抽象的
Age-related declines in cognition are associated with widespread structural and functional
brain changes, including changes in resting-state functional connectivity and gray and white
matter status. Recently we have shown that the elasticity of cerebral arteries also explains
some of the variance in cognitive and brain health in aging. 这里, we investigated how
network segregation, cerebral arterial elasticity (measured with pulse-DOT—the arterial pulse
based on diffuse optical tomography) and gray and white matter status jointly account for
age-related differences in cognitive performance. We hypothesized that at least some of the
variance in brain and cognitive aging is linked to reduced cerebrovascular elasticity, leading
to increased cortical atrophy and white matter abnormalities, 哪个, 反过来, are linked to
reduced network segregation and decreases in cognitive performance. Pairwise comparisons
between these variables are consistent with an exploratory hierarchical model linking them,
especially when focusing on association network segregation (compared with segregation in
sensorimotor networks). These findings suggest that preventing or slowing age-related
changes in one or more of these factors may induce a neurophysiological cascade beneficial
for preserving cognition in aging.
作者总结
Age-related declines in cognition are associated with widespread structural and functional
brain changes as well as changes in the elasticity of cerebral arteries. In this study, using an
exploratory hierarchical model as a guide, and novel measures of cerebral arterial elasticity
(pulse-DOT—the arterial pulse based on diffuse optical tomography), we show, for the first
时间, that cerebral arterial stiffness is strongly correlated with measures of functional brain
network segregation, even after partialing out the effects of age. These findings suggest that
preventing cerebral arterial stiffening could induce a neurophysiological cascade beneficial
for preserving cognition in aging.
开放访问
杂志
引文: 孔, 时间. S。, Gratton, C。,
低的, K. A。, Tan, C. H。, Chiarelli, A. M。,
弗莱彻, 中号. A。, . . . Fabiani, 中号. (2020).
Age-related differences in functional
brain network segregation are
consistent with a cascade of
cerebrovascular, structural and
cognitive effects. 网络
神经科学, 4(1), 89–114. https://
doi.org/10.1162/netn_a_00110
DOI:
https://doi.org/10.1162/netn_a_00110
支持信息:
https://doi.org/10.1162/netn_a_00110
已收到: 23 四月 2019
公认: 21 九月 2019
利益争夺: 作者有
声明不存在竞争利益
存在.
通讯作者:
Monica Fabiani
mfabiani@illinois.edu
处理编辑器:
Lucina Uddin
版权: © 2019
麻省理工学院
在知识共享下发布
归因 4.0 国际的
(抄送 4.0) 执照
麻省理工学院出版社
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
A
_
0
0
1
1
0
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Network segregation in aging
介绍
老化, even in the apparent absence of disease, is often accompanied by cognitive de-
cline, albeit with large individual differences (Fabiani, 2012; Salthouse, 2012; Shaw, Schultz,
Sperling, & Hedden, 2015). These age-related decreases in cognitive functioning have been
linked to functional and structural brain changes,
including alterations in resting-state
功能连接 (rsFC; see Ferreira & Busatto, 2013, for a review), cortical atrophy
(Salat et al., 2004; Thambisetty et al., 2010), white matter health (DeCarli et al., 1995;
Sullivan & Pfefferbaum, 2006; 哪个, Tsai, 刘, 黄, & 林, 2016), and cerebral arterial
elasticity (Chiarelli et al., 2017; Fabiani et al., 2014; Tan et al., 2017, 2016). Resting-state
FC is an important tool for gaining insight into the intrinsic organization of the brain. 这是
now widely understood that, even in the absence of any explicit tasks or goals, brain regions
spontaneously group into intrinsic connectivity networks with temporally correlated activity
(力量, 巴恩斯, 斯奈德, 施拉加尔, & 彼得森, 2012; 力量, 施拉加尔, & 彼得森, 2014; 杨
等人。, 2011). Aging is known to be accompanied by widespread changes in rsFC (see Ferreira &
Busatto, 2013 for a review), manifesting as decreased rsFC within brain networks and increased
rsFC across networks. These changes may be quantified as decreased network modularity or
system segregation (Betzel et al., 2014; Chan, 公园, Savalia, 彼得森, & 假发, 2014; Geerligs,
Renken, Saliasi, Maurits, & Lorist, 2015). They appear to be particularly prominent in the de-
fault mode network (DMN; Damoiseaux et al., 2008) and in networks related to higher level
cognitive functions (as compared with networks involved in sensory and motor processing;
例如, Andrews-Hanna et al., 2007; Chan et al., 2014).
There is also evidence that age-related differences in rsFC are linked to age-related differ-
ences in cognition (Andrews-Hanna et al., 2007; Chan et al., 2014; Geerligs et al., 2015).
例如, using seed analysis, Andrews-Hanna et al. (2007) demonstrated that the rsFC
between DMN seeds in the medial-prefrontal cortex and in the posterior-cingulate was pos-
itively correlated with performance in a composite memory measure. 相似地, using graph
theory analyses, Chan et al. (2014) demonstrated that the segregation of association system
networks was predictive of memory function, with greater system segregation associated with
higher memory function. This was not the case for the segregation of networks related to sen-
sorimotor processing. Other studies have also reported similar relationships between rsFC and
cognition by using other network measures such as modularity and local efficiency, 虽然
these results do not always survive correction for multiple comparisons (例如, Geerligs et al.,
2015).
最后, it has been reported that network connectivity measures may also be predictive
of cognitive improvements as a result of training in older adults. 例如, 老年人
with higher baseline measures of network segregation (or modularity) showed greater cognitive
gains as a result of cognitive training (Gallen et al., 2016) and aerobic-based training (Baniqued
等人。, 2018). 因此, converging evidence suggests that rsFC not only changes systematically
with aging but also plays an important role in age-related changes in cognition.
Although this evidence points to the importance of rsFC for understanding age-related
differences in brain function, other factors are also known to contribute to individual differ-
ences in age-related cognitive decline. 例如, cortical thinning occurs during aging
(Salat et al., 2004; Thambisetty et al., 2010) and has been shown to correlate with indi-
vidual differences in cognitive performance. In a longitudinal study collecting MRI data over
8 年, participants who had the greatest amount of cortical thinning at baseline (compared
with participants having less evidence of thinning) later exhibited clinical levels of impairments
in cognition (Pacheco, Goh, Kraut, Ferrucci, & Resnick, 2015). 相似地, in an extensive series
网络神经科学
90
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
A
_
0
0
1
1
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Network segregation in aging
Diffuse optical tomography (DOT):
DOT is an imaging method based
on near-infrared light. Using a
spectroscopic approach, it images
changes in oxy- and deoxy
hemoglobin. Data can be
reconstructed in 3D (tomography).
Pulse-DOT:
The arterial pulse wave measured
with DOT. Three parameters of the
pulse wave can be measured:
振幅, pulse transit time (analog
to pulse wave velocity), and PreFx.
Pulse relaxation function (PReFx):
A measure of the shape of the pulse
wave, reflecting the relative overlap
between the forward (systolic) 和
the backward (diastolic) waves. A
small overlap, resulting in a large
PreFx value, indicates high arterial
elasticity.
of studies conducted by Dickerson and colleagues, cortical thinning has also been shown to be
predictive of Alzheimer’s disease onset and progression (Bakkour, 莫里斯, & Dickerson, 2009;
Dickerson et al., 2011; Racine, Brickhouse, Wolk, & Dickerson, 2018).
White matter integrity also generally decreases with age (Fletcher et al., 2016; Gordon
等人。, 2008; Raz, Ghisletta, Rodrigue, 肯尼迪, & Lindenberger, 2010; Sullivan & Pfefferbaum,
2006; 杨等人。, 2016) and is correlated with individual differences in cognition in normally
aging adults (G. Gratton, Wee, Rykhlevskaia, Leaver, & Fabiani, 2008; Jolly et al., 2017, 2016;
Sullivan & Pfefferbaum, 2006; 杨等人。, 2016). 有, 实际上, evidence that age-related
declines in white matter integrity obtained with MR diffusivity methods may be related to
age-related reductions in rsFC. 例如, it has been shown that white matter integrity
was positively correlated with within-DMN rsFC even after controlling for age (Andrews-Hanna
等人。, 2007). 然而, another study examining the relationship between FC in seven networks
and white matter MR diffusivity measures including mean anisotropy, fractional anisotropy, 和
tract length did not find any relationship between FC and structural connectivity (Tsang et al.,
2017).
In light of these contradictory results, it is possible to hypothesize that white matter lesions,
manifesting as white matter signal abnormalities (WMSAs; 例如, DeCarli et al., 1995; Tan et al.,
in press) may be a more sensitive measure for examining the relationship between white mat-
ter integrity and age-related rsFC in older adults. WMSAs are based on the identification of
spots of hypointense (on T1 images) or hyperintense (in T2 images) MR signal and are thought
to reflect isolated areas of demyelination. Recent studies show that the presence of WMSAs
is closely related to amyloid deposition measures and is indicative of preclinical Alzheimer’s
disease in healthy older adults (Kandel et al., 2016; Lindemer, Greve, 菲舍尔, Augustinack,
& 礼拜, 2017). WMSAs are also inversely related to measures of fluid intelligence (Leaper
等人。, 2001). These findings, coupled with the temporal match between increasing WMSAs
and conversion to mild cognitive impairment in older adults (DeCarli et al., 1995), 建议
that WMSAs may be a stronger indicator of advanced stages of cognitive decline than other
measures of white matter integrity (Maniega et al., 2015).
White matter lesions such as WMSAs are thought to be caused primarily by vascular dys-
功能 (Bots et al., 1993; Longstreth et al., 1996; Moroni et al., 2018), largely related to
arteriosclerosis (IE。, stiffening of the arteries). Cortical thinning may also be related to arterial
问题, although the mechanisms for this relationship are less well established (例如, 马歇尔,
Asllani, Pavol, 张, & Lazar, 2017). Good cardiovascular and cardiorespiratory health are
also known to promote and maintain cognitive performance in aging (例如, Colcombe et al.,
2004; Crichton, Elias, Davey, & Alkerwi, 2014; Gordon et al., 2008). 然而, studies of
the impact of vascular health on cognition thus far have most often been based on periph-
eral indices of arterial elasticity (例如, by measuring the carotid-femoral pulse delay; 看
Maillard et al., 2016; Badji et al., 2019). To more directly assess cerebrovascular health, 我们
have recently developed measures of the cerebral arterial pulse based on diffuse optical to-
mography (pulse-DOT; Chiarelli et al., 2017; Fabiani et al., 2014; Tan et al., 2017, 2016).
This approach allows for the assessment and mapping of arterial elasticity in the brain, 两个都
globally and regionally. In the current study, we use this novel pulse-DOT approach to assess
arterial elasticity within the cerebral vascular tree, therefore allowing for the investigation of
more direct associations between cerebrovascular status and brain measures such as cortical
thinning, WMSAs, and rsFC parameters. One of the indices of arterial elasticity afforded by
pulse-DOT is a measure of the shape of the pulse wave during the interval between a peak
systole and the subsequent peak diastole (which we refer to as pulse relaxation function, 或者
网络神经科学
91
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
A
_
0
0
1
1
0
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Network segregation in aging
PReFx). PReFx describes the way in which arteries return to their original size, after dilating
to accommodate the oxygenated blood bolus generated by a heart pulsation. This reflects the
relative overlap of the forward wave generated by the systole and the backward wave gener-
ated by the peripheral resistance (IE。, arterioles). This overlap (or lack thereof ) is largely due
to the speed of propagation of the pulse wave between the measurement point and the point
of peripheral resistance, 哪个, 反过来, is determined by the rigidity (or lack of elasticity, IE。,
arterial stiffness or arteriosclerosis) of the arterial wall (Oliver & Webb, 2003): The greater the
rigidity, the greater the pulse-wave velocity, and the greater the temporal overlap between the
forward and backward waves, as indicated by the convexity of the pulse-wave shape (IE。,
PReFx) during the systole-diastole interval (Chiarelli et al., 2017; Fabiani et al., 2014).
Our previous work has shown that cerebral arterial elasticity measured with pulse-DOT is
not only associated with age and cardiorespiratory fitness, but also with behavioral measures
of cognitive flexibility (Fabiani et al., 2014; Tan et al., 2017). 此外, these studies have
also shown correlations between pulse-DOT measures and white and gray matter volumes,
not only across individuals but also across different brain regions within the same individuals
(Chiarelli et al., 2017). As WMSAs have been linked to vascular factors such as hypertension
(例如, Longstreth et al., 1996; Moroni et al., 2018), it can be hypothesized that a reduction
in arterial elasticity (IE。, arterial stiffness or arteriosclerosis) is an important contributor to the
development of WMSAs (Tan et al., in press) and cortical thinning in aging.
Here we hypothesized that at least some of the effects of aging on the structural integrity
of the brain, its functional network segregation, and cognitive performance may be due to the
effects of arteriosclerosis. As such we tested pairwise correlations between cerebral arterial
elasticity, as measured by PReFx, rsFC segregation, cortical thinning, WMSAs, and cognitive
表现. Specifically, we hypothesized that individual differences in network segrega-
tion would be correlated with differences in PReFx, with lower arterial elasticity (indexed by
a smaller PReFx) being associated with reduced segregation. Although this relationship may
be driven in part by age, as older adults are likely to exhibit both lower arterial elasticity and
lower network segregation, we predicted that this relationship would also explain residual vari-
ance when age was controlled for. The mechanisms linking PReFx and rsFC segregation are
likely to be complex and multiply determined. Individual differences in arteriosclerosis, caused
by long-term lifestyle factors such as diet, exercise, and stress (Bowie, Clements, Gratton, &
Fabiani, in press), and exacerbated by aging, are known to attack brain structure, and espe-
cially the white matter, which is essential to efficient connectivity. 反过来, this is likely to result
in lower cognitive performance, especially in domains that are more vulnerable to aging (例如,
episodic memory and reasoning, with the latter being a key component of fluid intelligence;
Lindenberger & Baltes, 1997). Maintaining a healthy and elastic cerebral arterial system, 那里-
fore, is likely to preserve brain and cognitive function, including the maintenance of network
segregation, which is typical of younger adults and correlated with higher cognitive perfor-
曼斯. In the final part of this paper we introduce, and provide an initial test for, an exploratory
hierarchical model that links these structural, functional, and cognitive factors together.
材料和方法
参加者
Forty-nine healthy right-handed (as assessed by the Edinburgh Handedness Inventory; Oldfield,
1971) adults, evenly distributed by age and gender (大约 8 people per decade, 50%
女性; total sample: 23 males, mean age: 47.29; age range: 18–75 years) were recruited from
the Urbana-Champaign community. These were the same participants included in the Chiarelli
网络神经科学
92
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
A
_
0
0
1
1
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Network segregation in aging
等人. (2017) and Tan et al. (2017, in press) publications focused on arterial elasticity data. 这
current study includes, for the first time, functional connectivity analyses based on a resting-
state fMRI protocol, which have not been previously analyzed or published. Participants re-
ported no history of neurological or psychiatric disorders and had no signs of dementia (作为
assessed by a score >51 on the modified Mini-Mental Status Examination [mMMSE]; Mayeux,
Stern, 罗森, & Leventhal, 1981) or depression (as assessed by the Beck’s Depression Inven-
保守党; Beck, Steer, & 棕色的, 1996). Informed consent was provided by each participant. 全部
procedures were approved by the Institutional Review Board of the University of Illinois.
Data from two participants were excluded because of excess movement during the resting-
state fMRI data acquisition (IE。, participants with fewer than 50 frames remaining for at least
one of the two runs were excluded; see section Functional connectivity and Supporting Infor-
mation S1.1 for specific details). Data from one additional participant were excluded because
of technical issues during optical data acquisition. All analyses and results described below
are based on a final sample of 46 参与者 (21 males, mean age: 46.41 [标准差= 17.32]; 年龄
范围: 18–75).
Data Acquisition
Data for each participant were collected during a cognitive testing session, an optical imaging
session, and a session during which both structural and functional MRI data were acquired.
In this session the following neuropsychological tests were admin-
Cognitive testing session.
istered: Logical Memory I and II tasks from the Wechsler Memory Scale–Fourth Edition (WMS-
IV; Wechsler, 2009) to measure episodic memory; the Trail Making Tests A and B (Corrigan
& Hinkeldey, 1987), to measure processing speed and working memory; the Controlled Oral
Word Association subtest of the Multilingual Aphasia Examination (a measure of verbal fluency
using the letters CFL; Benton & Hamsher, 1989); the OSPAN task (Unsworth, Heitz, Schrock,
& Engle, 2005) to assess working memory capacity under load; Raven’s progressive matri-
ces (Raven, Raven, & 法庭, 2003) and the Kaufmann Brief Intelligence Test Second Edition
(K-BIT2; 考夫曼 & 考夫曼, 2004) to assess, 分别, cognitive flexibility and IQ; 这
vocabulary subtest of the Shipley Institute of Living Scale (Shipley, 1940).
Participants underwent a scanning session in a 3-Tesla
Structural and functional MRI session.
Siemens Trio MR scanner, using a 12-channel head coil. Resting-state images were acquired
during an echo planar imaging sequence with the following pulse parameters: repetition time
(TR) = 2,000 多发性硬化症; echo time (TE) = 25 多发性硬化症; 38 contiguous interleaved 3-mm slices; flip angle
(F A) = 90◦; voxel sizes = 2.6 × 2.6 × 3.0 毫米. Two 6-minute scans were collected during
rest while participants fixated on a cross in the center of the screen.
A 3D T1-weighted anatomical scan for each participant was also acquired using an MPRAGE
sequence with the following pulse parameters: TR = 1,900 多发性硬化症; = 2.32 多发性硬化症; 192 sagittal
slices; slice thickness = 0.90 毫米; FA = 9◦; voxel sizes = 0.9 × 0.9 × 0.9 毫米, field-of-view
(FOV) = 172.8 × 230 × 230 毫米. The FOV for both MPRAGE and resting-state scans covered
the entire head.
Arterial elasticity (pulse-DOT) data were obtained during a resting-
Optical imaging session.
state optical imaging session, in which seated participants fixated on a cross in the center
of a screen. Optical data were acquired with a multichannel frequency-domain oximeter (ISS
网络神经科学
93
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
A
_
0
0
1
1
0
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Network segregation in aging
Imagent, 原野, 伊利诺伊州) equipped with 128 laser diodes (64 emitting light at 690 nm and
64 在 830 nm) 和 24 photomultiplier tubes. Time-division multiplexing was employed so that
each detector picked up light from 16 different sources at different times within a multiplexing
cycle at a sampling rate of 39.0625 赫兹. The light was transmitted to the scalp by using single-
optic fibers (0.4-mm core) and from the scalp back to the photomultiplier tubes by using fiber
bundles (3-mm diameter). The fibers were held in place using soft, but semirigid, custom-built
helmets, fitted to participants based their head circumference.
During this session, after the helmet was set up, the locations of the optodes were marked
digitally to improve spatial accuracy during later data processing. Fiducial markers were placed
on each participant’s left and right preauricular points and on the nasion. These fiducial points,
optode locations, and other scalp locations were digitized with a Polhemus FastTrak 3D digi-
tizer (准确性: 0.8 毫米; Colchester, VT) by using a recording stylus and three head-mounted
receivers, which allowed for small movements of the head in between measurements. Optode
locations and structural MRI data were then coregistered using the fiducials and a surface-
fitting Levenberg and Marquardt algorithm (Chiarelli, Maclin, Fabiani, & Gratton, 2015).
The pulse-DOT measurements were based on a high-density, large FOV optode montage,
covering the majority of the cortical mantle. Two 6-minute resting-state blocks were recorded
for each of four different optical recording montages, which were aggregated during analysis for
maximum cortical coverage. The helmet was never removed across the entire optical session to
remain faithful to the digitized locations of the optical sensors. A total of 384 channels (192 在
830 nm and 192 在 690 nm) were acquired for each montage, with source-detector distances
varying between 15 和 80 毫米, for a total of 1,536 channels covering most of the scalp
surface. The FOV for the pulse-DOT measures is related to the montage used (哪个, 在这个
案件, covered the entire scalp) and to the penetration of diffuse optical imaging (大约
30 mm from the head’s surface, encompassing most of the outer cortex).
Concurrently with pulse-DOT data acquisition (but not during the MR scanning) we also
recorded the electrocardiogram (EKG), using a Brain-Vision recorder and a Brain-Vision pro-
fessional BrainAmp integrated amplifier system (Brain Products, 德国). This concurrent
and time-locked acquisition allowed us to synchronize the optical pulse data to the R wave of
the EKG and ensure that the same pulse was examined regardless of location. Specifically, 带领
1 of the EKG (left wrist referenced to right wrist) was recorded with a sampling rate of 200 赫兹
and a band-pass filter of 0.1–100 Hz. The exact timing of each R-wave peak was determined
by searching for peak points exceeding a voltage threshold (scaled for each participant) 和
dismissing any peak points outside the normal range of interbeat intervals. The identification
of each peak was verified by visual inspection, and false detections were eliminated.
Data Processing
Cognitive testing data. We adapted the methods described in the Supplementary Results sec-
tion of Chan et al. (2014) and categorized our cognitive tests into the same a priori constructs
(Chan et al., 2014; 见表 1). In line with Chan et al. (2014), construct scores were calculated
by averaging the z-scores across the different cognitive tests that made up a construct. 自从
age effects are of interest in our study, we performed this calculation on raw scores instead of
age-adjusted scores, even for tests that normally adjust their final scores for age (IE。, KBIT-2
and CFL). 此外, since the Trail Making test A used for the processing speed construct
gives higher scores (IE。, longer times) for slower participants, we inverted its z-scores to make
it comparable to the results of Chan et al. (2014), since they used the digit comparison and
网络神经科学
94
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
A
_
0
0
1
1
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Network segregation in aging
桌子 1. A priori cognitive constructs
推理
Processing speed
Verbal fluency
Working memory
A priori cognitive construct
Episodic memory
Associated cognitive test(s)
WMS-IV Logical memory (immediate and delayed)
WMS-IV Verbal pairing (immediate and delayed)
Trail Making task A
CFLˆ
Trail Making test A minus Trail Making test B
OSPAN
Raven’s progressive matrices
KBIT-2 non-verbal scoresˆ
Shipley vocabulary subtest
KBIT-2 verbal scoresˆ
笔记. We composed a priori constructs based on the methods and construct categories of Chan
等人. (2014). Scores for the tests associated with each construct were converted to z-scores and
averaged with other tests associated with the same construct. Tests whose scores are normally
adjusted for age are indicated withˆ. Since we are interested in age effects, we performed the
z-score conversions on the raw scores instead of the age-adjusted scores. WMS = Wechsler
Memory Scale; CFL = verbal fluency test (letters CFL); KBIT = Kaufman Brief Intelligence Test.
Verbal ability
WAIS digit symbol tests where participants with lower performance had lower scores. We did
not have tests that were similar to those used for the Mental Control construct in Chan et al.
(2014; CANTAB Stop Signal Task and ETS Card Rotation) so we did not include this construct.
Cortical reconstruction and volumetric
White matter signal abnormalities and cortical thickness.
segmentation were performed on the structural MPRAGE images by using the FreeSurfer 5.3
image analysis suite (http://surfer.nmr.mgh.harvard.edu/; 菲舍尔 & 戴尔, 2000). This same au-
tomated procedure also yielded estimates of total intracranial volume (Buckner et al., 2004).
Cortical thickness estimates provided by FreeSurfer were obtained for each of the 50 地区
of the Desikan-Killiany atlas that are superficial and can be investigated using diffuse optical
imaging methods (to make these analyses consistent with the Pulse-DOT analyses, see be-
低的). The average cortical thickness across all these regions was used as an estimate of overall
cortical thickness for each individual. White matter signal abnormalities (WMSAs), which ap-
pear as hypointense on T1-weighted images, were labeled automatically using FreeSurfer 5.3’s
probabilistic procedure (Fischl et al., 2002). The results of this automatic segmentation were
inspected for errors and corrected where needed (http://surfer.nmr.mgh.harvard.edu/fswiki/
FsTutorial/TroubleshootingData). The WMSA variable was log-transformed because of its pos-
itively skewed distribution and adjusted for intracranial volume and sex.
Arterial elasticity was quantified using PReFx
Pulse-DOT measures of arterial stiffness.
(Chiarelli et al., 2017; Fabiani et al., 2014). As mentioned in the Introduction, PReFx describes
the temporal overlap between the forward and backward waves generated during each cardiac
循环. A greater overlap between these two waves is associated with low PReFx values and in-
dexes arteriosclerosis, whereas a small overlap is associated with high PReFx values and higher
arterial elasticity. 还, as a reminder, PReFx refers to arterial elasticity in the arterial tract con-
necting the point of measurement (which in optical recordings is close to the surface of the
cortex) with the place where peripheral resistance occurs (and therefore the backward wave
is generated), 那是, downstream relative to the point where the measurements are taken.
所以, PReFx, although measured superficially, is not only sensitive to arterial stiffening
occurring in superficial regions but also to stiffening occurring in deeper regions, such as those
where WMSAs are most likely to occur. This is different from other pulse parameters (例如
网络神经科学
95
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
A
_
0
0
1
1
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Network segregation in aging
pulse amplitude or transit time) that are instead more sensitive to arterial elasticity at the point
of measurement (振幅) and at points upstream from the point of measurement (transit
时间). This is the reason we focus on the PReFx parameter in the current paper.
To derive the PReFx measures, consistent with Chiarelli et al. (2017), the optical DC intensity
数据 (IE。, the average measures of the amount of light produced by a specific source and
reaching a specific detector during a multiplexed 1.6-ms interval) 在 830 nm were normalized,
movement corrected (Chiarelli et al., 2015), and band-pass filtered between 0.5 和 5 赫兹
by using a Butterworth filter. The arterial pulse waveform for each channel was obtained by
averaging the DC light intensity time locked to the peak of the R wave of the EKG, ensuring that
the same pulse cycle was measured at all locations. Three-dimensional reconstruction of the
pulse waveform across the head was estimated using a finite element method (FEM) applied to
the diffusion equation (Ishimaru, 1989; Paulsen & Jiang, 1995) for the forward model, 和
inverse procedure introduced by Chiarelli et al. (2016) was used for the inverse model. 这
allowed light intensity measurements to be localized in voxel space.
PReFx was computed as the area under the pulse wave between the peak systole and peak
diastole, normalized for interbeat interval and peak amplitude to avoid confounds, 进而
subtracted of .5 (a value that would correspond to a hypothetical “linear” relaxation function,
indicating constant speed of relaxation of the arterial wall during the interval between peak
systole and diastole). As mentioned, PReFx is more positive for elastic arteries, and less positive
(or even negative) for stiffer arteries. PReFx was estimated for each voxel for which the sensi-
活力 (measured by the average Jacobian) was greater than 1/1,000 (60 分贝) of the maximum
价值, allowing us to disregard voxels too deep to provide useful data (approximately >3 cm
from the scalp) as well as voxels that were not covered by the optical montage. 此外,
only voxels within the cortex (as identified by FreeSurfer) were considered. PReFx was com-
puted as the average value across the 50 regions of the Desikan-Killiany atlas covered by the
optical montages. 因此, we employed a global measure of PReFx to quantify cerebral arterial
elasticity in this study.
The resting-state MRI scans underwent standard fMRI preprocessing
Functional connectivity.
and FC analyses based on recommendations by Power et al. (2014) to reduce motion artifacts.
The preprocessing steps were carried out using SPM12 (http://www.fil.ion.ucl.ac.uk/spm).
Specifically, the T2*-weighted images for each run were slice-time corrected, realigned, 和
coregistered to each subjects’ structural MRI. The scans were then normalized to the MNI152
template brain by using 3-mm resampling. FC processing steps were then carried out in
MATLAB. A censoring mask was first created by marking high motion frames using a variant
of frame-wise displacement (FD), referred to here as FDfilt. FDfilt was calculated by low-pass
filtering the six motion parameters at 0.1 赫兹, finding the frame-wise displacement for each
of them and then summing the absolute values across them for each frame. We chose to use
FDfilt as it is more sensitive to differentiating motion from respiration as compared with FD
(Fair et al., 2018). Any frames with FDfilt > .1 were marked as high motion. The first and last
five frames each run were also marked for removal, as were any contiguous segments less
than five frames long. 下一个, each run was demeaned, followed by nuisance regression ignor-
ing the frames in the censoring mask. Nuisance variables included the global signal, average
cerebral spinal fluid, and average white matter time courses, and the Volterra expansion of
the six motion parameters (弗里斯顿, 威廉姆斯, 霍华德, Frackowiak, & 车工, 1996; IE。, 这
6 motion parameters of current and preceding volumes, and each of these values squared).
After nuisance regression, data were linearly interpolated across censored frames to preserve
网络神经科学
96
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
A
_
0
0
1
1
0
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Network segregation in aging
Segregation:
A measure used in graph theory
for summarizing the relative
independence of brain networks.
data integrity for the next step of band-pass filtering, where a Butterworth filter (band-pass:
0.009–0.08 Hz) of order 1 was applied. The functional images were then spatially smoothed
using a 6-mm FWHM Gaussian kernel. 最后, the frames initially marked for censoring that
were temporarily interpolated with data were removed. To ensure that we had sufficient data
for each participant, only participants with at least 50 frames remaining for each of the two
runs were used (>100 frames total). 因此, two participants were excluded. The average
number of frames remaining for our final sample of participants (N= 46 after the various
post–data collection exclusions; see section Participants) across both runs was 297.91 frames
(range = 169–320; see section S1.1 in the Supporting Information for how much censoring
was done using solely the FDfilt exclusion criterion; C. Gratton et al., 2018, 2019).
We based our regions of interest (ROIs) on previous work, which sorted 264 brain areas
into functionally distinct networks (Power et al., 2011). We focused on 11 out of the original
14 网络. Specifically, we used the 219 ROIs associated with the auditory, cingulo-opercular,
dorsal attention, default, 额顶叶, memory-related, sensorimotor hand, sensorimotor
嘴, salience, ventral attention, and visual networks, leaving out the 45 ROIs that were
undefined or belonging to the cerebellar and subcortical networks. The reason why subcor-
tical networks were excluded from the analysis is because these networks cannot be easily
explored with pulse-DOT measures (which tend to be limited to cortical areas).
The average time series within a 10-mm-diameter sphere was extracted for each of these
219 ROIs for each subject. A Fisher Z-transformed matrix was created for each subject based
on the correlations between the time series for each pair of ROIs. To avoid using negative corre-
lations, since their biological significance is currently debated (例如, 墨菲, Birn, Handwerker,
琼斯, & Bandettini, 2009), and to replicate the methods used by Chan et al. (2014) 在他们的
study of age-related differences in network segregation, all negative values in each participants’
Z-matrix were set to zero. All further FC analyses were derived from these Fisher Z-transformed
matrices.
Zw
System segregation was calculated using the formula of Chan et al. (2014) (IE。, system seg-
regation = Zw−Zb
, where Zw is the mean of the within-system Fisher Z-transformed r’s, and Zb
is the mean of the between-system Fisher Z-transformed r’s). Segregation was measured sepa-
rately for association systems (IE。, cingulo-opercular, dorsal attention, default, 额顶叶,
memory-related, salience, and ventral attention networks) and sensorimotor systems (IE。,
auditory, sensorimotor hand, sensorimotor mouth, and visual networks).
最后, it should be noted that the above seeds from Power et al. (2011) were derived using
younger adult samples, as is true of most other available parcellations. C. Gratton et al. (2018)
have shown that this network assignment of nodes holds well even in older adults and people
with Parkinson’s disease. Han and colleagues (2018) recently established age cohort–specific
parcellations to further improve the validity of rsFC across cohorts. 因此, we also calculated
association and sensorimotor system segregation by using the parcellations of Han et al. (2018).
Method and results obtained using the parcellations of Han et al. (2018) are presented as
支持信息 (see sections S2.1 and S2.2 and related tables and figures). In brief,
similar patterns of results were obtained across the two parcellation approaches.
统计分析
Based on our hypotheses, we first performed pairwise correlations between age and all other
factors in the model to ensure that we could replicate the previously demonstrated effects
of aging. Specifically, we computed pairwise correlations between age and PReFx, WMSAs,
网络神经科学
97
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
A
_
0
0
1
1
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Network segregation in aging
桌子 2. Complete correlation matrix
PReFx
WMSAs
Cortical
thickness
协会
系统
segregation
Sensorimotor
系统
segregation
Episodic
记忆
加工
速度
Verbal
fluency
Working
memory Reasoning
年龄
−0.381**
0.211
0.346*
0.563**
0.504**
−0.326*
−0.629**
−0.425**
−0.454**
−0.707**
−0.383**
0.525** −0.378**
0.427** −0.314*
0.154
PReFx
WMSAs
Cortical
thickness
协会
系统
segregation
Sensorimotor
系统
segregation
Episodic
记忆
加工
速度
Verbal
fluency
Working
记忆
推理
Verbal
能力
笔记. PReFx = pulse relaxation function; WMSAs = white matter signal abnormalities.
*p < .05; **p < .01, both one-tailed.
0.092
0.415** −0.182
−0.313*
0.283
−0.601**
−0.295*
−0.087
−0.280
−0.070
−0.157
−0.263
−0.223
−0.168
−0.048
−0.091
0.529**
0.442**
0.404**
0.437**
0.450**
0.392**
0.322*
0.269
0.196
0.047
0.209
0.181
0.251
0.184
0.045
0.131
0.097
0.140
0.234
0.008
0.345*
0.431**
0.070
0.334*
0.249
0.565**
0.248
0.294*
−0.140
−0.085
0.162
0.498**
0.187
0.061
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 3. Complete correlation matrix after partialing out age
Cortical
thickness
Association
system
segregation
Sensori-motor
system
segregation
Episodic
memory
Perceptual
speed
Verbal
fluency
Working
memory
Reasoning
WMSAs
PReFx
−0.180
−0.120 −0.081
0.340* −0.176
0.328*
0.066 −0.269
0.165
0.370*
0.074
0.032
0.153
0.150
−0.090 −0.285
0.160
0.216
0.207
0.046
0.016
0.064
0.064
0.337
0.246
−0.044 −0.191
0.170
0.116
0.411*
−0.185 −0.078
−0.028
0.129
0.095
0.147
0.232
0.001
0.229
0.099
0.330
0.076
0.281
0.222
0.318
0.264
0.561*
0.287
0.301
−0.113
−0.007
0.119
0.467*
0.315
0.428*
WMSAs
Cortical
thickness
Association
system
segregation
Sensorimotor
system
segregation
Episodic
memory
Perceptual
speed
Verbal
fluency
Working
memory
Reasoning
Verbal
ability
Note. PReFx = PReFx = pulse relaxation function; WMSAs = white matter signal abnormalities.
*p < .05.
Network Neuroscience
98
Network segregation in aging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Relationship between age and PReFx (A), cortical thickness (B), and WMSAs (C). Shad-
ing indicates the 95% bootstrap confidence interval for the linear regression function (solid line);
**p < .01. PReFx = pulse relaxation function; WMSAs = White matter signal abnormalities.
cortical thickness, association system segregation, sensorimotor system segregation, and each
of the a priori–defined cognitive construct scores.
Next, we measured the relationship between segregation and the other brain factors (i.e.,
cerebral arterial elasticity, WMSAs, cortical thickness). Specifically, we performed separate
pairwise comparisons between the two segregation indices (association system and sensori-
motor system segregation) and measures of WMSAs, cortical thickness, and PReFx.
Finally, we investigated the relationship between rsFC and cognition by correlating the six
cognitive constructs defined a priori separately for association system segregation and senso-
rimotor system segregation.
For all of the above analyses, we report one-tailed p values since we had specific hypotheses
about the direction of the age effects. Furthermore, we controlled for motion for both measures
of system segregation (i.e., association or sensorimotor) by partialing out mean FDfilt as a
control variable.
Network Neuroscience
99
Network segregation in aging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
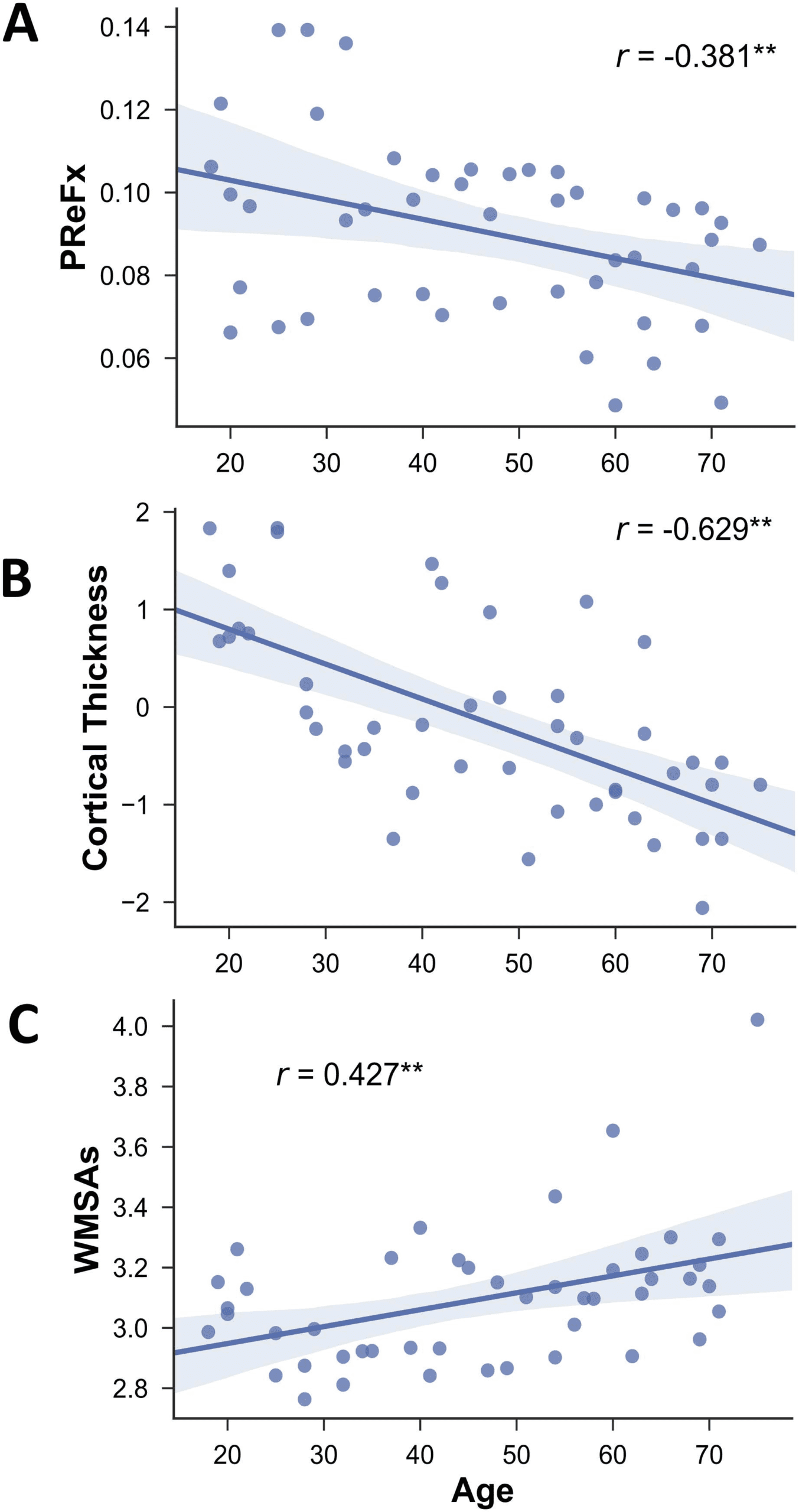
Figure 2. Mean functional connectivity matrices for younger (age range = 18–37 years, N = 16),
middle-aged (age range = 39–57 years, N = 15), and older adults (age range = 58–75 years,
N = 15). Color scale indicates the mean Fisher Z-transformed connectivity values. Aud = auditory
network; CON = cingulo-opercular network; DAN = dorsal attention network; DMN = default
mode network; FPN = fronto-parietal network; SN = salience network; VAN = ventral attention
network; Vis = visual network; Memory = memory network; SMHand = sensorimotor hand net-
work; SMMouth = sensorimotor mouth network.
Network Neuroscience
100
Network segregation in aging
We also computed correlations of all the relevant variables in the study with sex. All these
correlations were very small (|r′s| < .25), and none of them was significant at p < .05 level
(with the exception of intracranial volume, which was used in the computation of the WMSA
score; the correlation of the adjusted WMSA score with sex was, however, not significant;
r = −.187, p = .212). For these reasons we decided not to use sex as a covariate in the current
study, in order not to reduce the degrees of freedom of the analyses unnecessarily.
RESULTS
The correlation matrix including all variables measured in the study is presented in Table 2.
Table 3 reports the same matrix of correlations with age partialed out.
Effects of Aging
As expected, older age was associated with decreased PReFx and cortical thickness (r(44) =
−.381, p = .005; Figure 1A; and r(44) = −0.629, p < .0001; Figure 1B), and with increased
WMSAs (r(44) = .427, p = .002; Figure 1C). Furthermore, aging was related to reduced
segregation (see Figure 2 and Figure 3), especially for association networks, as previously
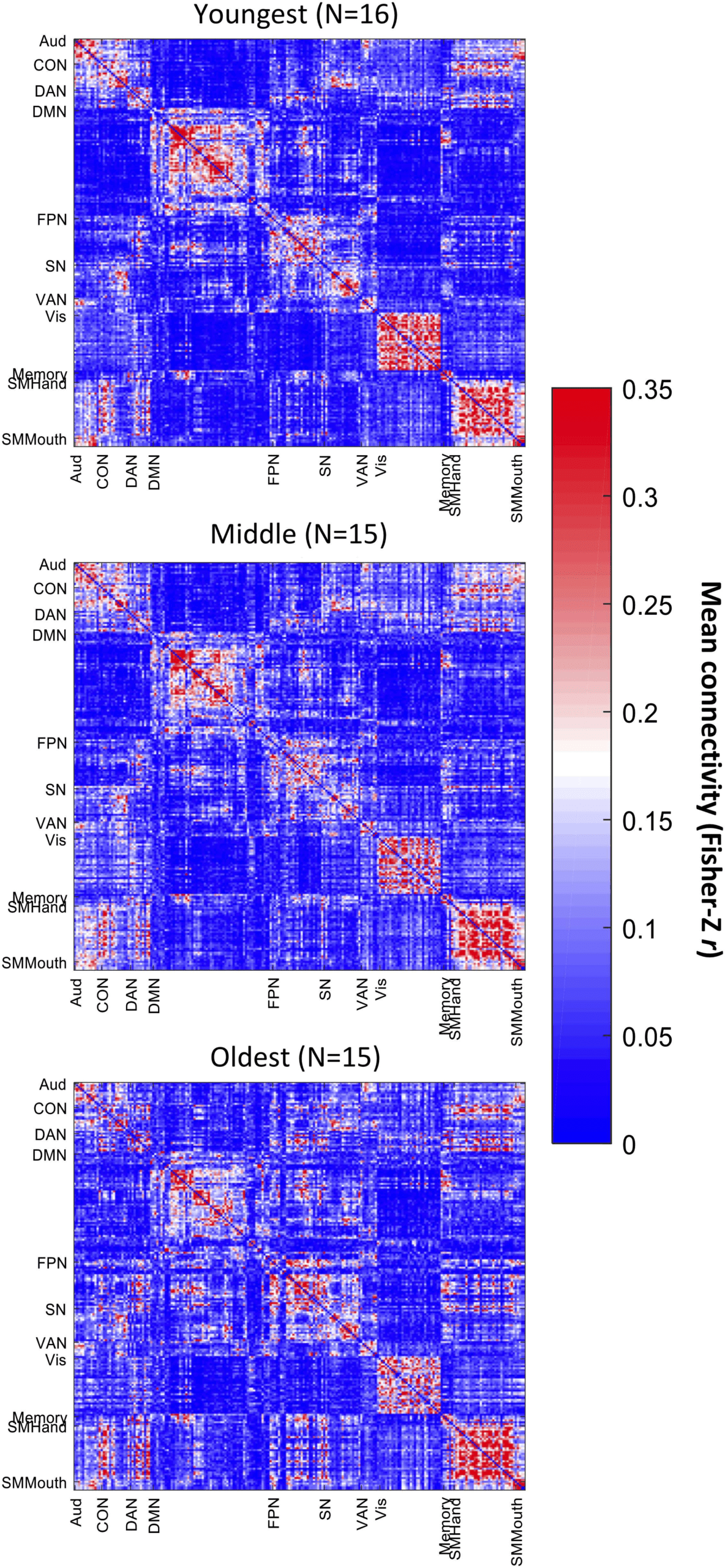
reported by Chan et al. (2014). Specifically, there was an age-related decrease for associa-
tion system segregation (r(43) = −.707, p < .0001) and sensorimotor system segregation
(r(43) = −.425, p = .002). However, the decrease was larger for association networks than
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Relationship between age and association system segregation (A) and sensorimotor
system segregation (B). Shading indicates the 95% bootstrap confidence interval for the linear re-
gression function (solid line); **p < .01.
Network Neuroscience
101
Network segregation in aging
for sensorimotor networks, as confirmed by a paired two-sample t-test (t(44) = −10.510,
p < .001).
The relationship between cognition and aging varied depending on the cognitive con-
struct. Whereas performance for episodic memory and reasoning decreased with age (r(44) =
−0.454, p = .001 and r(44) = −0.601, p < .0001, respectively), verbal ability increased with
age (r(44) = 0.415, p = 0.003), as is typically found (Baltes, Lindenberger, Schubert, Stober, &
Weilandt, 1997). There were no significant age effects for the other cognitive constructs after
correcting for multiple comparisons.
Relationship Between PReFx, WMSAs, Cortical Thickness, and System Segregation
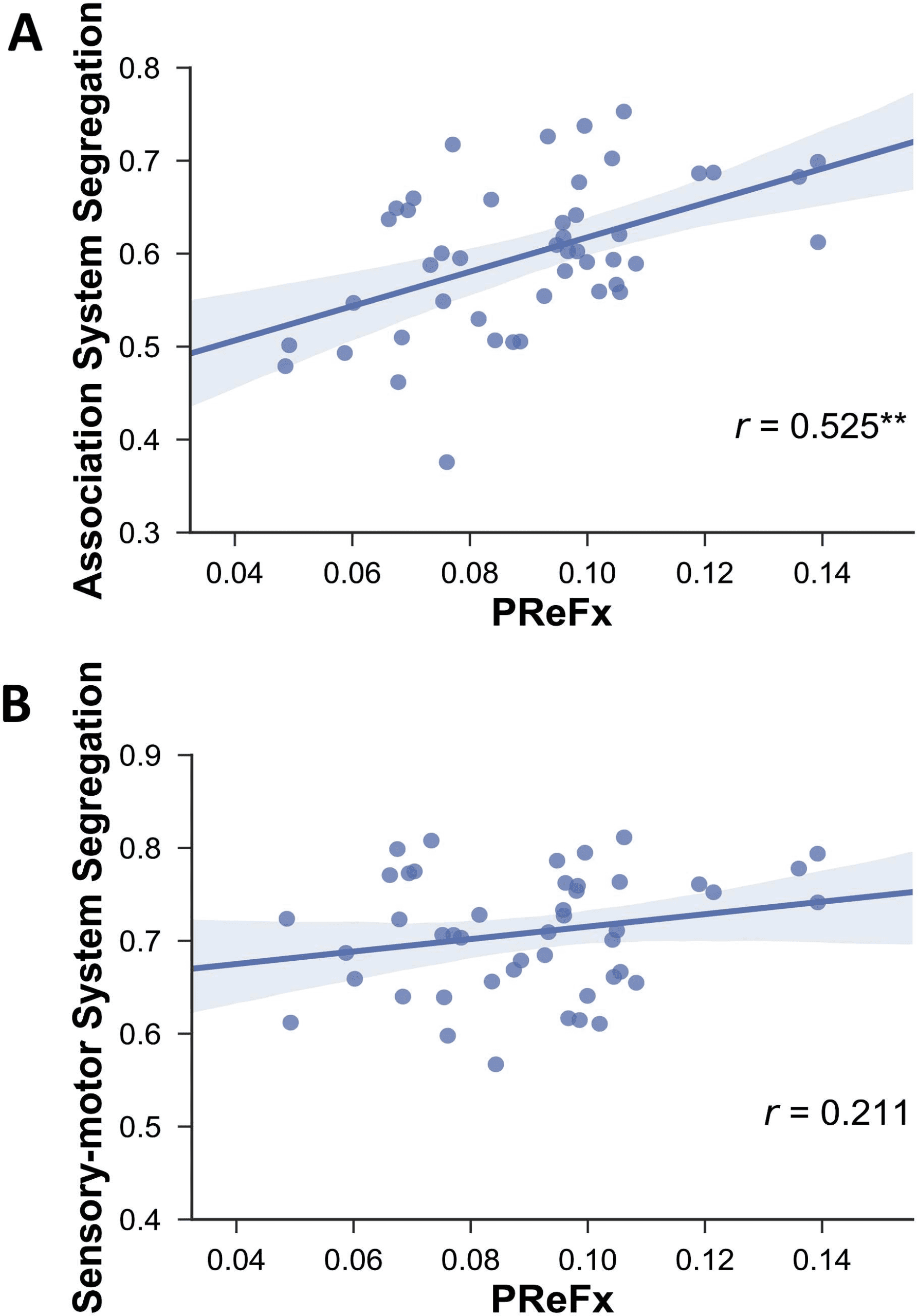
Our results revealed that, as predicted, greater network segregation was related to greater ar-
terial elasticity (Figure 4). Specifically, PReFx was significantly correlated with association sys-
tem segregation (r(43) = .525, p < .001). The correlation between PReFx and sensorimotor
system segregation was in the expected direction but did not reach statistical significance
(r(43) = .211, p = .087).
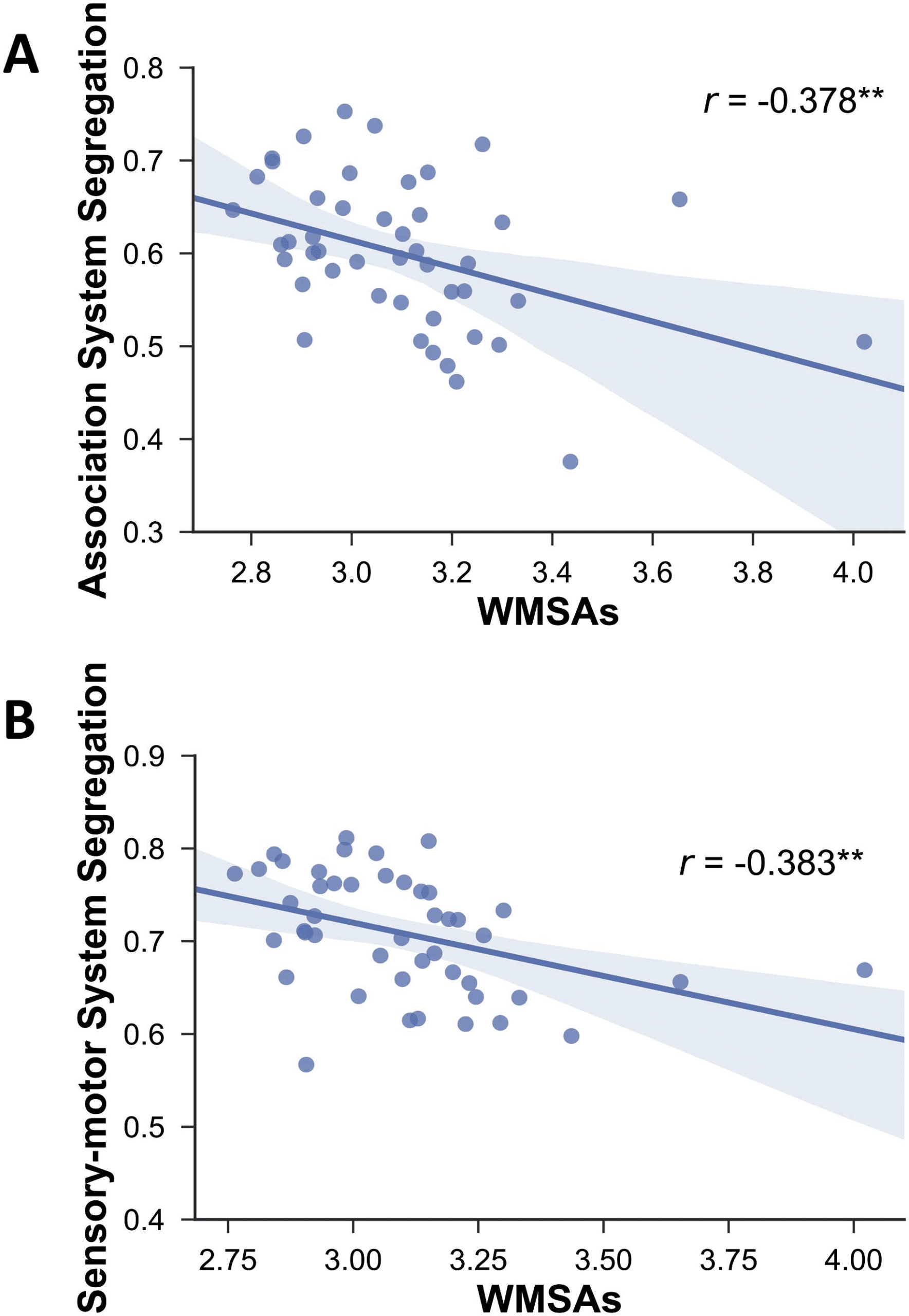
Furthermore, and also as predicted, our results showed that network segregation was sig-
nificantly associated with WMSAs (Figure 5). Specifically, WMSAs were negatively correlated
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Relationship between PReFx and association system segregation (A) and sensorimo-
tor system segregation (B). Shading indicates the 95% bootstrap confidence interval for the linear
regression function (solid line); **p < .01. PReFx, pulse relaxation function.
Network Neuroscience
102
Network segregation in aging
with association system segregation (r(43) = −.378, p = .006) and sensorimotor system seg-
regation (r(43) = −.383, p = .006).
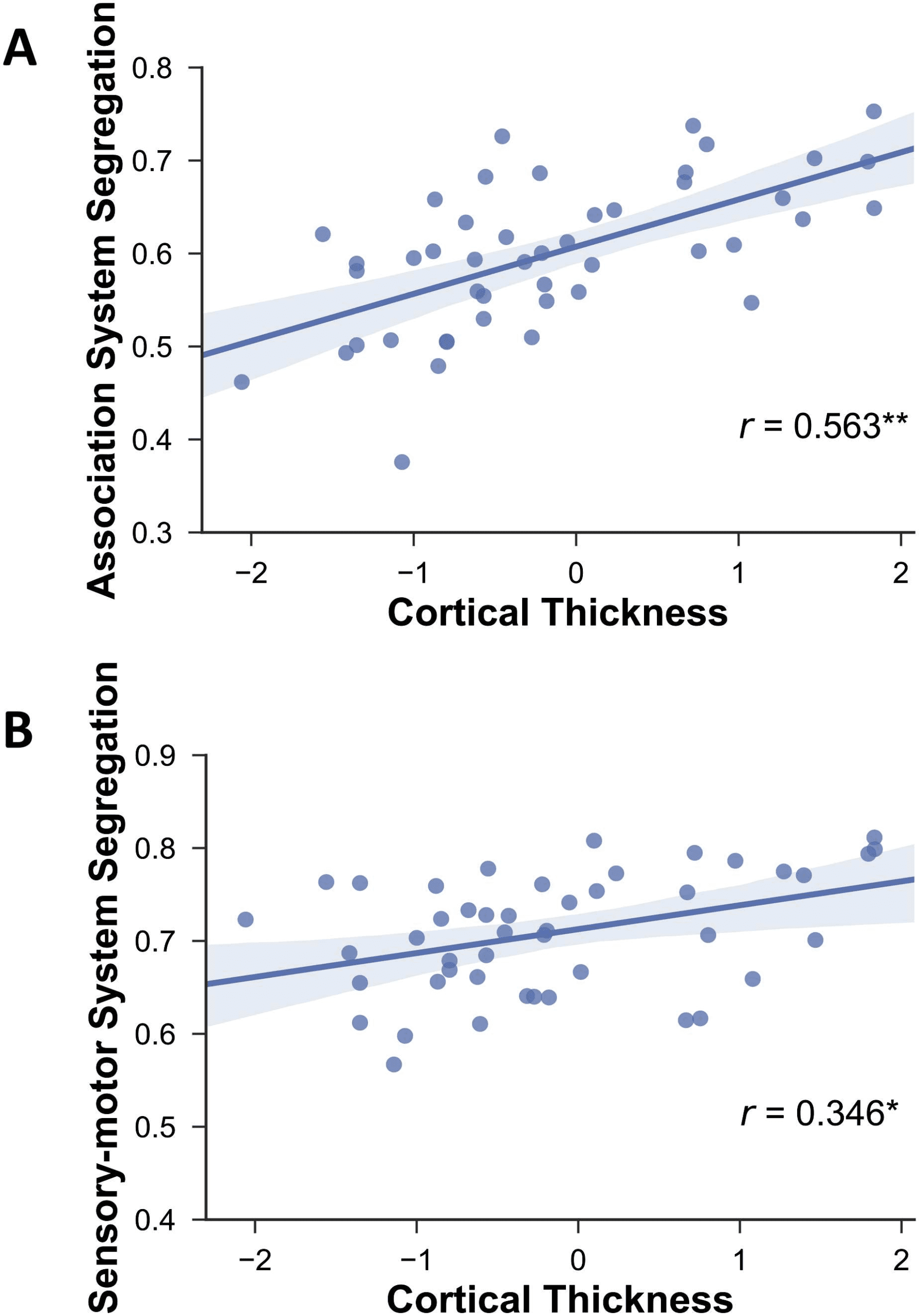
Finally, network segregation was also positively associated with cortical thickness (Figure 6;
cortical thickness vs. association system segregation: r(43) = .563, p < .0001; vs. sensorimo-
tor segregation: r(43) = .346, p = .012). However, although greater cortical thickness was
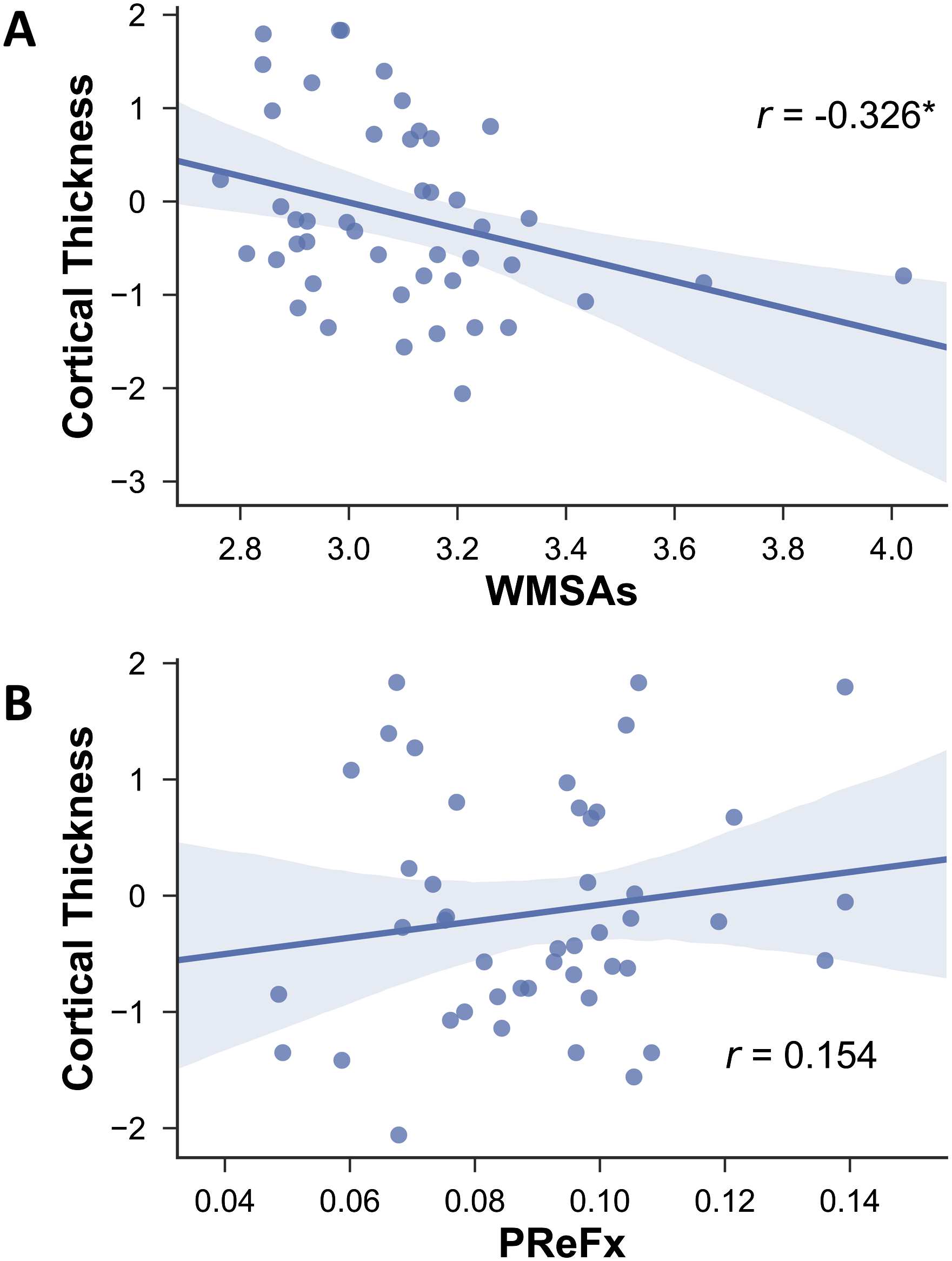
associated with fewer WMSAs as expected (r(43) = −0.326, p = 0.016; Figure 7), it was not
significantly correlated with PReFx (r(43) = .154, p = .162; Figure 7B).
Table 3 reports the correlation matrix across all the variables in the study after partialing
out age. Notably, the correlations of association system segregation with PReFx and cortical
thickness remain significant after removing the effect of age. Furthermore, the relationships
between sensorimotor system segregation and working memory also remain significant. Fi-
nally, several of the correlations between cognitive constructs remain significant even after
partialing out age.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Relationship between white matter signal abnormalities (WMSAs) and association sys-
tem segregation (A) and sensorimotor system segregation (B). Shading indicates the 95% bootstrap
confidence interval for the linear regression function (solid line); **p < .01. Note that these rela-
tionships remain significant (r = −0.477 and r = 0.414, respectively, p < .01 for both) when the
two extreme values are excluded.
Network Neuroscience
103
Network segregation in aging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
Figure 6. Relationship between cortical thickness and association system segregation (A) and
sensorimotor system segregation (B). Shading indicates the 95% bootstrap confidence interval for
the linear regression function (solid line); *p < .05, **p < .01.
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
t
.
Relationship Between Network Segregation and Cognition
Similar to the correlations with age, episodic memory and reasoning were the only cogni-
tive constructs significantly correlated with rsFC. Specifically, higher episodic memory perfor-
mance was correlated with greater association system segregation (r(43) = .450, p = .001),
whereas higher reasoning performance was correlated with both greater association system
segregation (r(43) = .529, p < .001) and greater sensorimotor segregation (r(43) = .404,
p = .004). Higher sensorimotor system segregation was also correlated with better working
memory (r(43) = .442, p = .002). However, there was no relationship between verbal abil-
ity and segregation measures. Network segregation was related to other cognitive constructs,
but these correlations were no longer significant after correcting for multiple comparisons
by adjusting for the number of cognitive constructs (see Table 2). Specifically, by using a
Bonferroni approach, the alpha-rejection criterion (p = .05) was divided by the number of
cognitive constructs (6), so the adjusted alpha-rejection levels for significant cognitive-related
results is p = 0.0083.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Summary and Exploratory Integration of Results
The results presented thus far are consistent with the presence of a cascade of phenomena,
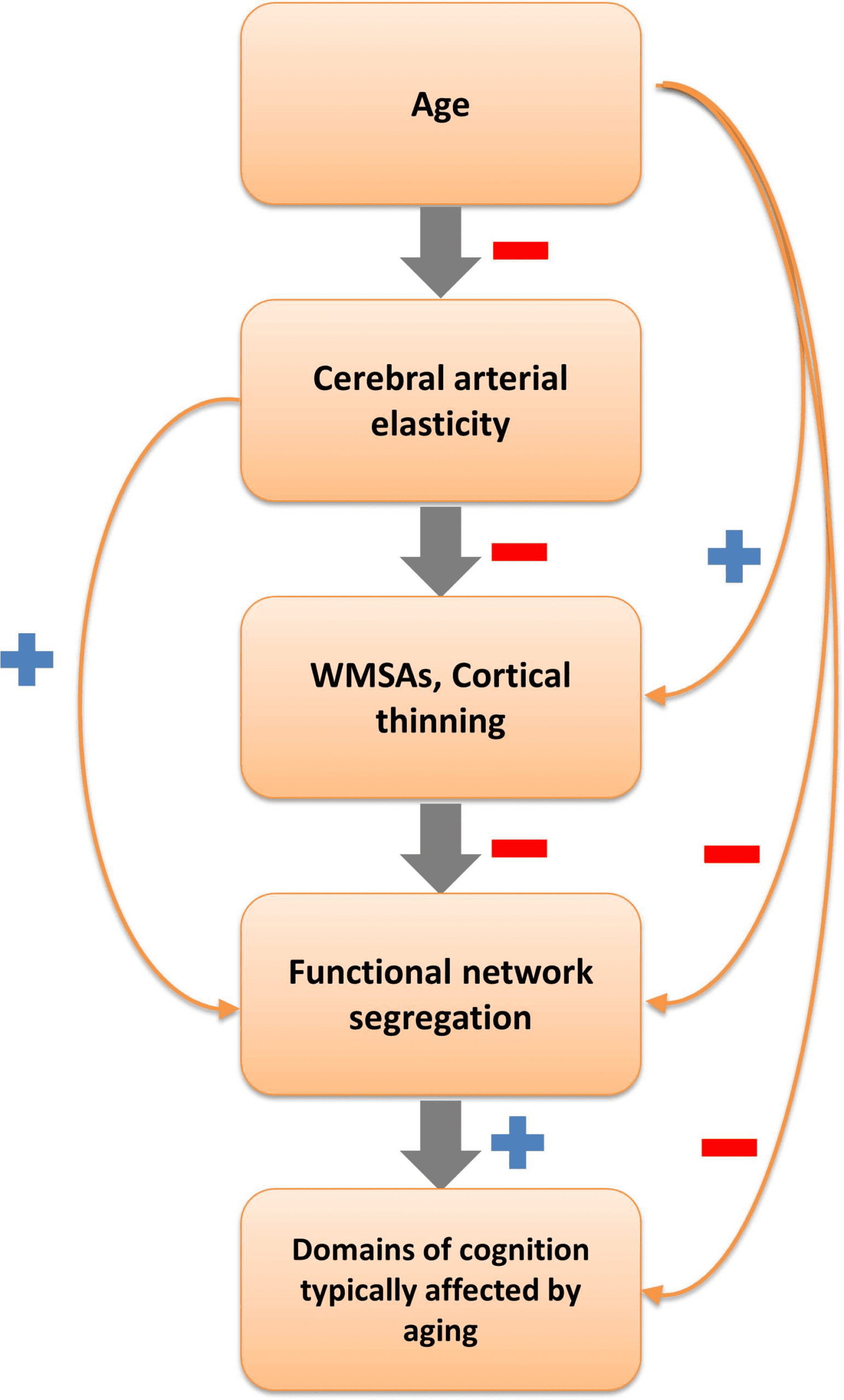
hierarchically linking the variables under study (Figure 8). This view (in line with others present
in the literature, e.g., Barnes, 2015; de la Torre, 2012) proposes that aging is linked to declines
Network Neuroscience
104
Network segregation in aging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
Figure 7. Relationship between cortical thickness and WMSAs (A) and PReFx (B). Shading indi-
cates the 95% bootstrap confidence interval for the linear regression function (solid line); *p < .05.
PReFx = pulse relaxation function; WMSAs = white matter signal abnormalities.
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
in cerebral arterial elasticity (as measured here by decreases in PReFx), which, in turn, are
tied to deterioration of white (as indexed by increased WMSAs) and gray matter structures
(measured as reduced cortical thickness), due to reduced perfusion, decreases in neurotrophic
factors (such as brain derived neurotrophic factor, BDNF, and vascular endothelial growth
factor, VEGF; Voss et al., 2013a; Voss, Vivar, Kramer, & Van Praag, 2013b), and neuronal and
myelin loss. These factors consequently weaken brain function and network organization, as
measured by the reduced rsFC network segregation, and finally manifest in lower cognitive
performance, especially in those cognitive domains that are most vulnerable to aging (e.g.,
episodic memory and reasoning/fluid intelligence).
Note that this hypothesized cascade does not exclude that aging might influence structural
brain integrity through other pathogenetic mechanisms, such as the development of plaques
and neurofibrillary tangles, neuroinflammation, and so on. It is also important to note that each
of the levels in the cascade (cerebrovascular, anatomical, functional, and cognitive) is likely
multidimensional, and that there are multiple ways in which each level can be quantified. For
example, functional network organization may be quantified using segregation (Chan et al.,
2014), participation coefficient, and modularity (Geerligs et al., 2015; see Sporns & Betzel,
2016, for an overview). However, our aim here is to highlight some specific links between
pairs of consecutive elements along the chain depicted in Figure 8, rather than to provide an
exhaustive coverage of all possible routes to cognitive aging.
Network Neuroscience
105
Network segregation in aging
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 8. Schematic representation of an exploratory hierarchical cascade of effects. The sign
next to each arrow indicates the direction of the relationship between pairwise variables (red minus
sign = negative relationship; blue plus sign = positive relationship). The main hypothesized cascade
of effects is indicated by the gray arrows. Pairwise relationships between the levels are indicated by
the orange arrows. WMSAs = white matter signal abnormalities.
In this paper, we have provided substantial evidence for correlational links between any two
consecutive levels within the chain depicted in Figure 8. However, the cross-sectional nature
of the study and limited sample size preclude the use of a systematic multilevel mediation
analysis. Instead, here we use an indirect approach to provide a preliminary exploration of the
hypothesized cascade. In fact, this hypothetical chain also suggests that the relationships be-
tween adjacent levels in the chain should be overall stronger than those between nonadjacent
levels, with the smallest correlations for the levels that are furthest apart. Thus, an examination
of the patterns of correlations across adjacent versus nonadjacent levels might provide some
very preliminary evidence in support of the proposed hierarchy.
An exception to this expected correlation pattern might occur for the age variable. This is
due to two reasons: (a) age can influence all other levels in multiple ways, and not necessarily
only through the chain proposed in Figure 8 (e.g., age could influence cognition via life-long
learning and other cohort effects); and (b) age can be measured with almost errorless precision,
which is not the case for any of the variables representing other levels, engendering de facto
higher correlations because of its lack of error variance. In other words, were we to include
age in this final exploratory test, age would dominate (and therefore likely obscure) all other
relationships. As such, we omitted age when examining the patterns of correlations across
Network Neuroscience
106
Network segregation in aging
levels (see Table 3 for effect of partialing out age on relationships between the other variables
in the study).
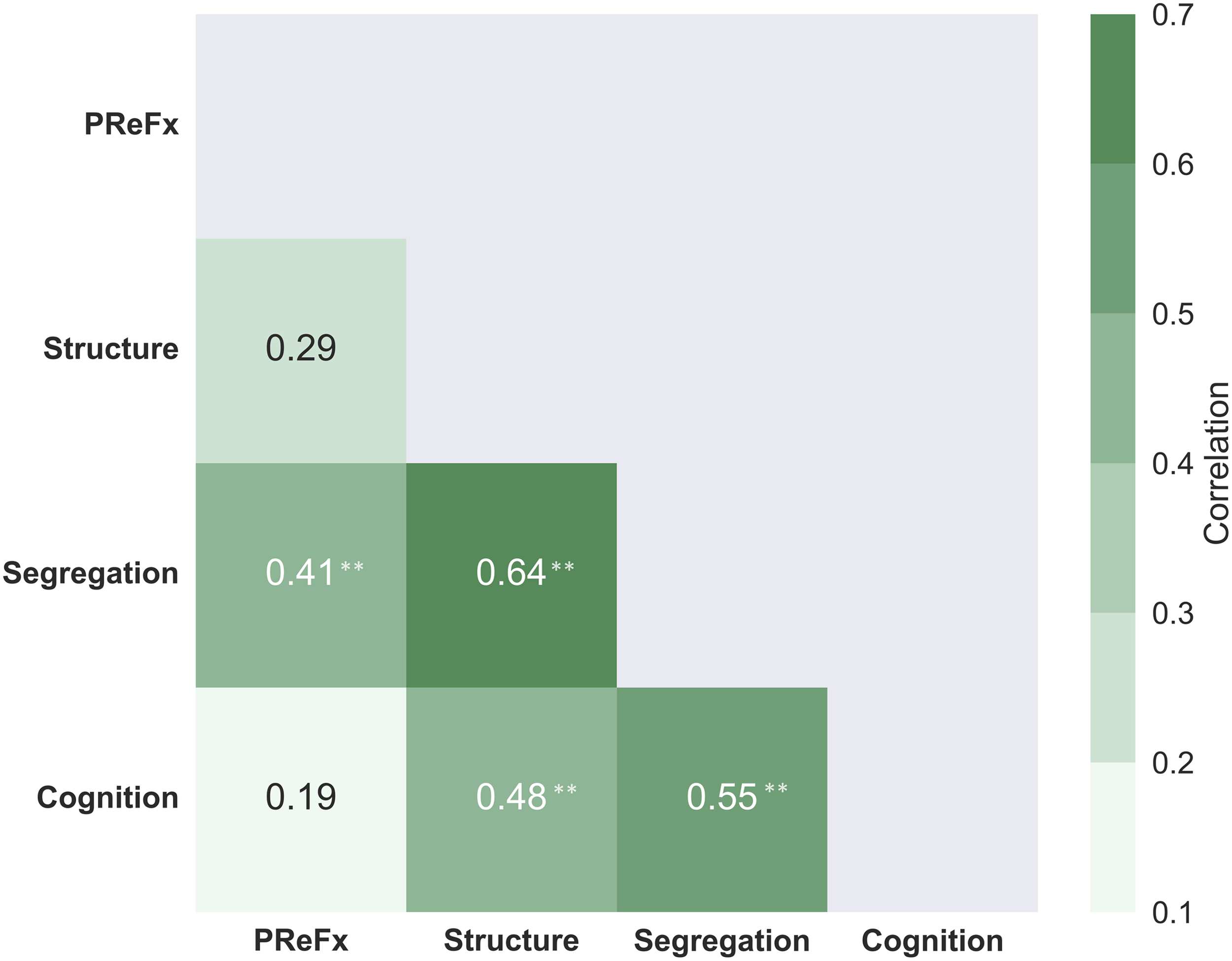
Note that multiple measures are available for several levels. For example, for the structure
level, we combined the correlations of each of the other levels with cortical thinning and
WMSAs (whose sign was changed to maintain coherence with other variables). Similarly, for
the functional segregation level, we combined correlations involving associative and senso-
rimotor networks. Finally, for the cognitive level we combined the correlations for episodic
memory and reasoning. To combine the variables for each level, we did the following: (a)
The values for each variable within each level were standardized; (b) if needed, their sign was
changed to maintain coherence; (c) the standardized values were averaged together; and (d)
correlations were computed across levels.
The resulting table of correlations is presented in Figure 9. As predicted, correlations tended
to be higher (average r = .51) between adjacent levels (1-off from the main diagonal in
Figure 9), intermediate between levels separated by one level (average r = .45), and small-
est between levels separated by two levels (r = .19). This apparent pattern is consistent with
the predictions of the hierarchical model. To provide quantitative support for this qualitative
impression, we performed a bootstrap analysis in which we generated 10,000 samples of
N = 46, taken from the subject pool with replacement. For each bootstrap sample, we cal-
culated the same 6-value correlation matrix presented in Figure 9, and then calculated the
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 9. Absolute values for the integrated pairwise correlations between levels in the exploratory
hierarchical model presented in Figure 8. The green shading indicates the strength of the correla-
tions, based on the scale presented on the right; **p < .01. PReFx = pulse relaxation function.
Network Neuroscience
107
Network segregation in aging
mean (Fisher-transformed) correlations of the elements 1-, 2-, and 3-off the diagonal, defined
as above. Finally, we compared the differences between the actual values (from Figure 9) with
the distribution of the corresponding differences obtained with the bootstrap approach. The
results indicated that both the 1-off and the 2-off correlation values were significantly greater
than the 3-off (p = .0021 and p = .0170, respectively). However, the difference between the
1-off and the 2-off values was not significant (p = .2109).
DISCUSSION
This study examined the interrelationships between cerebrovascular, structural, and functional
factors associated with age-related differences in cognition. Specifically, we found that cere-
brovascular elasticity (as measured by pulse-DOT PReFx) and structural integrity (as measured
by WMSAs and cortical thickness) were related to functional network segregation (taken as an
indicator of network integrity) and to age-related differences in cognitive domains that are typ-
ically affected by aging (episodic memory and reasoning/fluid intelligence). By using pairwise
comparisons, we showed that aging is related to reduced cerebral arterial elasticity, increases
in WMSAs, reduced cortical thickness, and reduced network segregation (especially in asso-
ciation systems, compared with sensorimotor systems). In turn, these differences are related
to reduced cognitive performance in the aforementioned domains. Finally, to integrate these
correlational results and put them in context, we used an exploratory hierarchical model as a
reference framework, and showed that adjacent levels within the model were more strongly
correlated than distant ones, when the relationship with age was not considered. Further re-
search, using larger and/or longitudinal samples, will be needed to allow for a more formal
testing of this model.
A novel and central aspect of the current study is that we showed, for the first time, a ro-
bust relationship between optical measures of the elasticity of cerebral arteries and associative
network segregation, extending the findings of Chan et al. (2014). This relationship remained
significant even after controlling for the effects of age (Table 3), supporting the idea that in-
dividual differences in cerebrovascular health may explain important variations in functional
brain organization, irrespective of age.
As mentioned, the relationships between aging, network segregation, and cognitive perfor-
mance reported in this paper closely replicated results previously reported by Chan et al. (2014)
in a larger sample, demonstrating the robustness and replicability of these findings across dif-
ferent scanner sites and different samples of participants. Importantly, we also replicated their
finding of a significant relationship between episodic memory and association system seg-
regation, despite using a subset of different tests for these constructs, demonstrating that the
relationships between system segregation and cognition may be generalizable even across
nonidentical cognitive tasks. In addition, we found that system segregation (both association
and sensorimotor) was also correlated with reasoning.
The cognitive tests used in this study focused on working memory and reasoning, consistent
with the extensive literature, indicating that these domains of cognition are more affected by
aging than others. Specifically, fluid intelligence (of which reasoning is a key component) is
more vulnerable to aging compared with crystallized intelligence (which includes vocabulary
and general knowledge). The latter has been shown to be relatively unaffected by aging un-
til very late in life, and we did not expect any age-related differences, especially given the
maximum 75 years of age of our sample. Thus, within this context, our results are typical: they
Network Neuroscience
108
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation in aging
indicate that domains of cognition that are especially affected by aging (i.e., episodic memory
and reasoning) are also affected by cerebrovascular health.
Because of the relatively small sample used in this study, we primarily relied on pairwise
correlations between variables. Although our results are consistent with the hierarchical model
presented in Figure 8, we could not explicitly test it, because of insufficient power for imple-
menting mediation analyses. In addition, the cross-sectional nature of the current study fur-
ther limits inferences regarding potential causal links. Future research should investigate this
model by using a larger longitudinal sample, paired with mediation analyses, to fully explore
the proposed relationships.
It is also important to note that the exploratory model presented in Figure 8 is based on re-
lationships between selected measures within each level/factor instead of testing for a number
of possible measures that may be available for each level (e.g., functional network organiza-
tion can be measured in several other ways). Although beyond the scope of this paper, it is
possible that a more thorough investigation of other measures within each level would allow
us to better refine the neurophysiological pathways that might be implicated. For example,
age-related declines in cerebral perfusion may play a role in how a reduced arterial elasticity
is tied to increases in WMSAs (Bernbaum et al., 2015; Brickman et al., 2011; Marstrand et al.,
2002; van Dalen et al., 2016) and decreases in brain volume and memory performance (Alosco
et al., 2013). Similarly, there is also evidence that the plasma level of VEGF is related to the
incidence of WMSAs (Pikula et al., 2013), is predictive of baseline and longitudinal hippocam-
pal volume and cognitive performance (Hohman & Jefferson, 2015), and may be modified by
exercise training (Maass et al., 2016; Voss et al., 2013a). The current results are in line with
this literature but do not fully explore these mechanisms.
Despite the above limitations, the reported findings are consistent with a hierarchical rela-
tionship between different cerebrovascular, structural, and functional variables. Demonstration
of a hierarchical relationship between these variables may suggest that preservation of cogni-
tion in healthy aging could be tied to improvements on one or more of these levels. In other
words, it is possible that the effects of interventions designed to delay the effects of age on
cognition may influence this cascade, so that variations at any level may be reflected in dif-
ferences not only at that level but also on subsequent levels in the hierarchy, including the
ultimate outcome of improved (or at least delayed decline of) cognition. Indeed, it has been
shown that physical exercise (which may modify arterial function) is linked to cortical thick-
ness (Lee et al., 2016; Williams et al., 2017) as well as improved rsFC and cognition in older
adults (Voss et al., 2010). In this sense, measures of intermediate levels within the proposed
cascade may be used to demonstrate not only the efficacy of an intervention strategy but also
point at some of the mechanisms involved in the process.
Along the same lines of reasoning, there is some evidence that rsFC measures may be
useful for predicting intervention outcomes in older adults (Baniqued et al., 2018). It has
been found that older adults exhibiting higher network modularity at baseline showed greater
improvements in synthesizing complex information after cognitive training, and this effect
was more pronounced for association systems compared with sensorimotor systems (Gallen
et al., 2016). Furthermore, there is evidence that some interventions may promote changes in
WMSAs, rsFC, and cognition as demonstrated by a study examining older adults after 6 months
of resistance training, cognitive training, or both (Suo et al., 2016). Results demonstrated that
resistance training improved scores on a clinical dementia scale, whereas cognitive training
was linked to the stabilizing of memory performance. Interestingly, resistance training was
Network Neuroscience
109
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation in aging
linked to improvements (i.e., decreases) in WMSAs, although this was not linked to any im-
provements in behavior. Cognitive training, however, was linked not only to improved hip-
pocampal rsFC but also to improvements in memory performance. Although untested, this
finding may suggest that even though all these phenomena are affected by intervention, im-
provements in rsFC measures may manifest more clearly in cognitive performance than im-
provements in WMSA measures. Although this particular study employed a sample with mild
cognitive impairment, it is still highly relevant to our understanding of these factors in normal
aging. Together with converging evidence linking exercise to improvements in cerebrovascular
function in older adults (see Voss, Nagamatsu, Liu-Ambrose, & Kramer, 2011, for a review),
these studies suggest that multiple neurobiological factors may be improved by intervention,
leading to preservation of cognitive function in older adults.
In conclusion, the current study investigates pairwise relationships between multiple neu-
rophysiological factors that may explain some of the effects of aging on cognition. Specifically,
we propose that aging compromises the cerebral vasculature, leading to declines in brain
structure, which in turn reduce brain functional integrity and finally result in reduced cogni-
tive performance (see also Barnes, 2015, and de la Torre, 2012, for similar models applied to
Alzheimer’s disease). Although a more in-depth exploration of these relationships is needed,
including explicit testing of causal links, the current findings suggest pathways by which cog-
nition can be preserved through improvement of one or more of the factors.
ACKNOWLEDGMENTS
We acknowledge the support of the Bioimaging Center of the Beckman Institute for Advanced
Science and Technology at the University of Illinois at Urbana-Champaign (UIUC-BI-BIC). We
are grateful to Dr. Gagan Wig and colleagues for providing access to their age-cohort-specific
parcellations (Han et al., 2018).
SUPPORTING INFORMATION
Supporting Information for this article is available at https://www.doi.org/10.1162/netn_a_00110.
AUTHOR CONTRIBUTIONS
Tania S. Kong: Formal analysis; Investigation; Methodology; Software; Visualization; Writing -
Original Draft. Caterina Gratton: Data curation; Methodology; Writing - Review & Editing.
Kathy A Low: Data curation; Project administration; Supervision; Writing - Review & Editing.
Chin Hong Tan: Data curation; Formal analysis; Writing - Review & Editing. Antonio M Chiarelli:
Formal analysis; Methodology; Software; Writing - Review & Editing. Mark A Fletcher: For-
mal analysis; Writing - Review & Editing. Benjamin Zimmerman: Data curation; Writing -
Review & Editing. Edward L Maclin: Data curation; Writing - Review & Editing. Bradley P.
Sutton: Methodology. Gabriele Gratton: Conceptualization; Formal analysis; Funding acquisi-
tion; Methodology; Supervision; Writing - Review & Editing. Monica Fabiani: Conceptualiza-
tion; Funding acquisition; Project administration; Supervision; Writing - Review & Editing.
FUNDING INFORMATION
Monica Fabiani, National Institute on Aging, Award ID: R01AG059878. Gabriele Gratton, NIH,
Award ID: R56MH097973. Gabriele Gratton, NCRR, Award ID: S10-RR029294.
Network Neuroscience
110
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation in aging
REFERENCES
Alosco, M. L., Gunstad, J., Jerskey, B. A., Xu, X., Clark, U. S.,
Hassenstab, J., . . . Sweet, L. H. (2013). The adverse effects of
reduced cerebral perfusion on cognition and brain structure in
older adults with cardiovascular disease. Brain and Behavior,
3(6), 626–636. https://doi.org/10.1002/brb3.171
Andrews-Hanna, J. R., Snyder, A. Z., Vincent, J. L., Lustig, C., Head,
D., Raichle, M. E., & Buckner, R. L. (2007). Disruption of large-
scale brain systems in advanced aging. Neuron, 56(5), 924–935.
https://doi.org/10.1016/j.neuron.2007.10.038
Badji, A., Sabra, D., Bherer, L., Cohen-Adad, J., Girouard, H.,
& Gauthier, C. J. (2019). Arterial Stiffness and Brain Integrity: A
review of MRI findings. Ageing research reviews. https://doi.org/
10.1016/j.arr.2019.05.001.
Bakkour, A., Morris, J. C., & Dickerson, B. C. (2009). The cortical
signature of prodromal AD Regional thinning predicts mild AD
dementia. Neurology, 72(12), 1048–1055.
Baltes, P. B., Lindenberger, U., Schubert, G., Stober, A., & Weilandt, M.
(1997). Emergence of a powerful connection between sensory
and cognitive functions across the adult life span: A new window
to the study of cognitive aging? Psychology and Aging, 12(1), 12–21.
Baniqued, P. L., Gallen, C. L., Voss, M. W., Burzynska, A. Z.,
Wong, C. N., Cooke, G. E., . . . D’Esposito, M. (2018). Brain
network modularity predicts exercise-related executive function
gains in older adults. Frontiers in Aging Neuroscience, 9(JAN),
1–17. https://doi.org/10.3389/fnagi.2017.00426
Barnes, J. N. (2015). Exercise, cognitive function, and aging. Ad-
vances in Physiology Education, 39(2), 55–62. https://doi.org/10.
1152/advan.00101.2014
Beck, A. T., Steer, R. A., & Brown, G. K. (1996). Manual for the Beck
Depression Inventory (2nd ed.). San Antonio, TX: The Psycholog-
ical Corporation.
Benton, A. L., & Hamsher, K. (1989). Multilingual Aphasia Examina-
tion Manual. Iowa City, IA: University of Iowa.
Bernbaum, M., Menon, B. K., Fick, G., Smith, E. E., Goyal, M., &
Frayne, R. (2015). Reduced blood flow in normal white matter
predicts development of leukoaraiosis. Journal of Cerebral Blood
Flow & Metabolism, 35(10), 1610–1615. https://doi.org/10.1038/
jcbfm.2015.92
Betzel, R. F., Byrge, L., He, Y., Goñi, J., Zuo, X.-N., & Sporns,
O. (2014). Changes in structural and functional connectivity
among resting-state networks across the human lifespan. Neuro-
Image, 102, 345–357. https://doi.org/10.1016/j.neuroimage.2014.
07.067
Bots, M. L., Breteler, M. M. B., Hofman, A., Grobbee, D. E.,
van Swieten, J. C., van Gijn, J., . . . de Jong, P. T. V. M. (1993).
Cerebral white matter lesions and atherosclerosis in the Rotter-
dam Study. The Lancet, 341(8855), 1232–1237. https://doi.org/
10.1016/0140-6736(93)91144-B
Bowie, D. C., Clements, G. M., Gratton, G., & Fabiani, M. (in press).
The effects of cardiorespiratory fitness on brain and cognitive
aging: A review. In C. Martin, V. R. Preedy & R. Rajendram (Eds.)
The Neuroscience of Aging.
Brickman, A. M., Siedlecki, K. L., Muraskin,
J.,
Luchsinger, J. A., Yeung, L. K., . . . Stern, Y. (2011). White
J., Manly,
J.
matter hyperintensities and cognition: Testing the reserve hypoth-
esis. Neurobiology of Aging, 32(9), 1588–1598. https://doi.org/
10.1016/j.neurobiolaging.2009.10.013
Buckner, R. L., Head, D., Parker, J., Fotenos, A. F., Marcus, D.,
Morris, J. C., & Snyder, A. Z. (2004). A unified approach for
morphometric and functional data analysis in young, old, and
demented adults using automated atlas-based head size nor-
malization: Reliability and validation against manual measure-
ment of total intracranial volume. NeuroImage, 23(2), 724–738.
https://doi.org/10.1016/j.neuroimage.2004.06.018
Chan, M. Y., Park, D. C., Savalia, N. K., Petersen, S. E., & Wig,
G. S. (2014). Decreased segregation of brain systems across the
healthy adult lifespan. Proceedings of the National Academy of
Sciences, 111(46), E4997–E5006. https://doi.org/10.1073/pnas.
1415122111
Chiarelli, A. M., Fletcher, M. A., Tan, C. H., Low, K. A., Maclin,
E. L., Zimmerman, B., . . . Fabiani, M. (2017). Individual dif-
ferences in regional cortical volumes across the life span are
associated with regional optical measures of arterial elastic-
ity. NeuroImage, 162(August), 199–213. https://doi.org/10.1016/
j.neuroimage.2017.08.064
Chiarelli, A. M., Maclin, E. L., Fabiani, M., & Gratton, G. (2015). A
kurtosis-based wavelet algorithm for motion artifact correction of
fNIRS data. NeuroImage, 112, 128–137. https://doi.org/10.1016/
j.neuroimage.2015.02.057
Chiarelli, A. M., Maclin, E. L., Low, K. A., Mathewson, K. E., Fabiani,
M., & Gratton, G. (2016). Combining energy and Laplacian
regularization to accurately retrieve the depth of brain acti-
vity of diffuse optical tomographic data. Journal of Biomedi-
cal Optics, 21(3), 036008. https://doi.org/10.1117/1.JBO.21.3.
036008
Colcombe, S. J., Kramer, A. F., Erickson, K. I., Scalf, P., Mcauley, E.,
Cohen, N. J., . . . Elavsky, S. (2004). Cardiovascular fitness, corti-
cal plasticity, and aging. Proceedings of the National Academy of
Sciences, 101(9), 3316–3321. https://doi.org/10.1073/pnas.97.
15.8728
Corrigan, J. D., & Hinkeldey, N. S. (1987). Relationships between
parts A and B of the Trail Making Test. Journal of Clinical Psychol-
ogy, 43(4), 402–409.
Crichton, G. E., Elias, M. F., Davey, A., & Alkerwi, A. (2014). Cardio-
vascular health and cognitive function: The Maine-Syracuse
Longitudinal Study. PLoS One, 9(3). https://doi.org/10.1371/
journal.pone.0089317
Damoiseaux, J. S., Beckmann, C. F., Arigita, E. J. S., Barkhof, F.,
Scheltens, P., Stam, C. J., . . . Rombouts, S. A. R. B. (2008). Re-
duced resting-state brain activity in the “default network” in nor-
mal aging. Cerebral Cortex, 18(8), 1856–1864. https://doi.org/10.
1093/cercor/bhm207
de la Torre, J. C. (2012). Cardiovascular risk factors promote brain
hypoperfusion leading to cognitive decline and dementia. Car-
diovascular Psychiatry and Neurology, 2012, 1–15. https://doi.
org/10.1155/2012/367516
DeCarli, C., Murphy, D. G. M., Tranh, M., Grady, C. L., Haxby,
J. V., Gillette, J. A., . . . Schapiro, M. B. (1995). The effect of
white matter hyperintensity volume on brain structure, cognitive
Network Neuroscience
111
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation in aging
performance, and cerebral metabolism of glucose in 51 healthy
adults. Neurology, 45(11), 2077–2084.
tional network effects in Parkinson disease. Cerebral Cortex,
1–15. https://doi.org/10.1093/cercor/bhy121
Dickerson, B. C., Shah, R. C., Killiany, R. J., Albert, M. S., Hyman,
B. T., & Blacker, D. (2011). Alzheimer-signature MRI biomarker
predicts AD dementia in cognitively normal adults. Neurology,
76(16), 1395–1402.
Fabiani, M. (2012). It was the best of times, it was the worst of times:
A psychophysiologist’s view of cognitive aging. Psychophysiol-
ogy, 49(3), 283–304. https://doi.org/10.1111/j.1469-8986.2011.
01331.x
Fabiani, M., Low, K. A., Tan, C.-H., Zimmerman, B., Fletcher, M. A.,
Schneider-Garces, N., . . . Gratton, G. (2014). Taking the pulse
of aging: Mapping pulse pressure and elasticity in cerebral arter-
ies with optical methods. Psychophysiology, 51(11), 1072–1088.
https://doi.org/10.1111/psyp.12288
Fair, D. A., Miranda-Dominguez, O., Snyder, A. Z., Perrone, A., Earl,
A., Van, A. N., . . . Gratton, C. (2018). Correction of respiratory
artifacts in MRI head motion estimates. Preprint. Retrieved from
https://www.biorxiv.org/content/early/2018/06/07/337360
Ferreira, L. K., & Busatto, G. F. (2013). Resting-state functional con-
nectivity in normal brain aging. Neuroscience, and Biobehavioral
Reviews, 37(3), 384–400. https://doi.org/10.1016/j.neubiorev.
2013.01.017
Fischl, B., & Dale, A. M.
(2000). Measuring the thickness of
the human cerebral cortex from magnetic resonance images.
Proceedings of the National Academy of Sciences of the United
States of America, 97, 11050–11055. https://doi.org/10.1073/
pnas.200033797
Fischl, B., Salat, D. H., Busa, E., Albert, M., Dieterich, M.,
Haselgrove, C., et al. (2002). Whole brain segmentation: au-
tomated labeling of neuroanatomical structures in the human
brain. Neuron, 33, 341–355. https://doi.org/10.1016/S0896-
6273(02)00569-X
Fletcher, M. A., Low, K. A., Boyd, R., Zimmerman, B., Gordon, B. A.,
Tan, C. H., . . . Fabiani, M. (2016). Comparing aging and fit-
ness effects on brain anatomy. Frontiers in Human Neuroscience,
10(June), 1–15. https://doi.org/10.3389/fnhum.2016.00286
Friston, K. J., Williams, S., Howard, R., Frackowiak, R. S. J., & Turner,
R. (1996). Movement-related effects in fMRI time-series. Mag-
netic Resonance in Medicine, 35(3), 346–355. https://doi.org/
10.1002/mrm.1910350312
Gallen, C. L., Baniqued, P. L., Chapman, S. B., Aslan, S., Keebler,
M., Didehbani, N., & D’Esposito, M. (2016). Modular brain net-
work organization predicts response to cognitive training in older
adults. PLoS One, 11(12), 1–17. https://doi.org/10.1371/journal.
pone.0169015
Geerligs, L., Renken, R. J., Saliasi, E., Maurits, N. M., & Lorist, M. M.
(2015). A brain-wide study of age-related changes in functional
connectivity. Cerebral Cortex, 25(7), 1987–1999. https://doi.org/
10.1093/cercor/bhu012
Gordon, B. A., Rykhlevskaia, E. I., Brumback, C. R., Lee, Y., Elavsky,
S., Konopack, J. F., . . . Fabiani, M. (2008). Neuroanatomical
correlates of aging, cardiopulmonary fitness level, and educa-
tion. Psychophysiology, 45(5), 825–838. https://doi.org/10.1111/
j.1469-8986.2008.00676.x
Gratton, C., Koller, J. M., Shannon, W., Greene, D. J., Maiti, B.,
Snyder, A. Z., . . . Campbell, M. C. (2018). Emergent func-
Gratton, C., Coalson, R., Dworetsky, A., Adeyemo, B., Barch,
D., Tranel, D., Miranda-Dominguez, O., Fair, D., Dosenbach,
N., Snyder, A., Perlmutter, J., Petersen, S., & Campbell, M.
(2019). High frequency contamination of motion estimates in
single-band fMRI. Paper presented at the 2019 Meeting of the
Human Brain Mapping Organization, Rome, Italy, June 2019.
Gratton, C., Laumann, T. O., Nielsen, A. N., Greene, D. J., Gordon,
E. M., Gilmore, A. W., . . . Dosenbach, N. U. (2018). Functional
brain networks are dominated by stable group and individual fac-
tors, not cognitive or daily variation. Neuron, 98(2), 439–452.
https://doi.org/10.1016/j.neuron.2018.03.035
Gratton, G., Wee, E., Rykhlevskaia, E. I., Leaver, E. E., & Fabiani, M.
(2008). Does white matter matter? Spatio-temporal dynamics of
task switching in aging. Journal of Cognitive Neuroscience, 21(7),
1380–1395.
Han, L., Savalia, N. K., Chan, M. Y., Agres, P. F., Nair, A. S., & Wig,
G. S. (2018). Functional parcellation of the cerebral cortex across
the human adult lifespan. Cerebral Cortex, 28(12), 4403–4423.
https://doi.org/10.1093/cercor/bhy218
Hohman, P. B. S., T. J., & Jefferson, A. L. (2015). The role of vas-
cular endothelial growth factor in neurodegeneration and cogni-
tive decline exploring interactions with biomarkers of Alzheimer
JAMA Neurology, 72(5), 520–529. https://doi.org/
disease.
10.1001/jamaneurol.2014.4761
Ishimaru, A. (1989). Diffusion of light in turbid material. Applied
Optics, 28(12), 2210. https://doi.org/10.1364/AO.28.002210
Jolly, T. A. D., Cooper, P. S., Rennie, J. L., Levi, C. R., Lenroot, R.,
Parsons, M. W., . . . Karayanidis, F. (2017). Age-related decline in
task switching is linked to both global and tract-specific changes
in white matter microstructure. Human Brain Mapping, 38(3),
1588–1603. https://doi.org/10.1002/hbm.23473
Jolly, T. A. D., Cooper, P. S., Wan Ahmadul Badwi, S. A., Phillips,
N. A., Rennie, J. L., Levi, C. R., . . . Karayanidis, F. (2016). Micro-
structural white matter changes mediate age-related cognitive
decline on the montreal cognitive assessment (moCA). Psycho-
physiology, 53(2), 258–267. https://doi.org/10.1111/psyp.12565
Kandel, B. M., Avants, B. B., Gee, J. C., McMillan, C. T., Erus,
G., Doshi, J., . . . Wolk, D. A. (2016). White matter hyperin-
tensities are more highly associated with preclinical alzheimer’s
disease than imaging and cognitive markers of neurodegener-
ation. Alzheimer’s and Dementia: Diagnosis, Assessment and
Disease Monitoring, 4, 18–27. https://doi.org/10.1016/j.dadm.
2016.03.001
Kaufman, A. S., & Kaufman, N. L. (2004). Kaufman Brief Intelligence
Test (2nd ed.). San Antonio, TX: The Psychological Corporation.
Leaper, S. A., Murray, A. D., Lemmon, H. A., Staff, R. T., Deary,
I. J., Crawford, J. R., & Whalley, L. J. (2001). Neuropsychologic
correlates of brain white matter lesions depicted on MR im-
ages: 1921 Aberdeen Birth Cohort. Radiology, 221(1), 51–55.
https://doi.org/10.1148/radiol.2211010086
Lee, J. S., Shin, H. Y., Kim, H. J., Jang, Y. K., Jung, N., Lee, J., . . .
Kang, M. (2016). Combined effects of physical exercise and ed-
ucation on age-related cortical thinning in cognitively normal
individuals. Nature Scientific Reports, 6, 24284. https://doi.org/
10.1038/srep24284
Network Neuroscience
112
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation in aging
Lindemer, E. R., Greve, D. N., Fischl, B. R., Augustinack, J. C., &
Salat, D. H. (2017). Regional staging of white matter signal abnor-
malities in aging and Alzheimer’s disease. NeuroImage: Clinical,
14, 156–165. https://doi.org/10.1016/j.nicl.2017.01.022
Lindenberger, U., & Baltes, P. B. (1997). Intellectual Functioning
in old and very old age: Cross-sectional results from the Berlin
Aging Study. Psychology and Aging, 12(3), 410–432.
Longstreth, W. T., Manolio, T. A., Arnold, A., Burke, G. L., Bryan,
N., Jungreis, C. A., . . . Fried, L. (1996). Clinical correlates of
white matter findings on cranial magnetic resonance imaging of
3301 elderly people. Stroke, 27(8), 1274–1282. https://doi.org/
10.1161/01.STR.27.8.1274
Maass, A., Düzel, S., Brigadski, T., Goerke, M., Becke, A., &
Sobieray, U. (2016). Relationships of peripheral IGF-1, VEGF and
BDNF levels to exercise-related changes in memory, hippo-
campal perfusion and volumes in older adults. NeuroImage, 131,
142–154. https://doi.org/10.1016/j.neuroimage.2015.10.084
Maillard, P., Mitchell, G. F., Himali, J. J., Beiser, A., Tsao, C. W.,
Pase, M. P., . . . De Carli, C. (2016). Effects of arterial stiffness on
brain integrity in young adults from the framingham heart study.
Stroke, 47(4), 1030–1036. https://doi.org/10.1161/STROKEAHA.
116.012949
Maniega, S. M., Valdés Hernández, M. C., Clayden, J. D., Royle,
N. A., Murray, C., Morris, Z., . . . Wardlaw, J. M. (2015). White
matter hyperintensities and normal-appearing white matter in-
tegrity in the aging brain. Neurobiology of Aging, 36(2), 909–918.
https://doi.org/10.1016/j.neurobiolaging.2014.07.048
Marshall, R. S., Asllani, I., Pavol, M. A., Cheung, Y., & Lazar,
M. (2017). Altered cerebral hemodyamics and cortical thin-
ning in asymptomatic carotid artery stenosis. PLoS One, 12(12),
e0189727.
Marstrand, J. R., Garde, E., Rostrup, E., Ring, P., Rosenbaum, S.,
Mortensen, E. L., & Larsson, H. B. W. (2002). Cerebral perfusion
and cerebrovascular reactivity are reduced in white matter hy-
perintensities. Stroke, 33, 972–976.
Mayeux, R., Stern, Y., Rosen, J., & Leventhal, J. (1981). Depres-
sion, intellectual impairment, and Parkinson disease. Neurology,
31(6), 645–650.
Moroni, F., Ammirati, E., Rocca, M. A., Filippi, M., Magnoni, M.,
& Camici, P. G. (2018). Cardiovascular disease and brain health:
Focus on white matter hyperintensities. IJC Heart and Vascula-
ture, 19, 63–69. https://doi.org/10.1016/j.ijcha.2018.04.006
Murphy, K., Birn, R. M., Handwerker, D. A.,
Jones, T. B., &
Bandettini, P. A. (2009). The impact of global signal regression
on resting state correlations: are anti-correlated networks intro-
duced? NeuroImage, 44(3), 893–905. https://doi.org/10.1016/j.
neuroimage.2008.09.036
Oldfield, R. C. (1971). The assessment and analysis of handedness:
The Edinburgh inventory. Neuropsychologia, 9(1), 97–113.
Oliver, J. J., & Webb, D. J. (2003). Noninvasive assessment of ar-
terial stiffness and risk of atherosclerotic events. Arteriosclerosis,
Thrombosis, and Vascular Biology, 23(4), 554–566. https://doi.
org/10.1161/01.ATV.0000060460.52916.D6
Pacheco, J., Goh, J. O., Kraut, M. A., Ferrucci, L., & Resnick,
S. M. (2015). Greater cortical thinning in normal older adults
predicts later cognitive impairment. Neurobiology of Aging,
36(2), 903–908. https://doi.org/10.1016/j.neurobiolaging.2014.
08.031
Paulsen, K. D., & Jiang, H. (1995). Spatially varying optical property
reconstruction using a finite element diffusion equation approx-
imation. Medical Physics, 22, 691–701.
.
.
. Seshadri, S.
Pikula, A., Beiser, A. S., Chen, T. C., Preis, S. R., Vorgias, D.,
Decarli, C.,
(2013). Serum brain–derived
neurotrophic factor and vascular endothelial growth factor lev-
els are associated with risk of stroke and vascular brain injury:
Framingham study. Stroke, 44, 2768–2775. https://doi.org/10.
1161/STROKEAHA.113.001447
Power, J. D., Barnes, K. A., Snyder, A. Z., Schlaggar, B. L., &
Petersen, S. E. (2012). Spurious but systematic correlations in
functional connectivity MRI networks arise from subject mo-
tion. NeuroImage, 59(3), 2142–2154. https://doi.org/10.1016/
j.neuroimage.2011.10.018
Power, J. D., Cohen, A. L., Nelson, S. M., Wig, G. S., Barnes, K. A.,
Church, J. A., . . . Petersen, S. E. (2011). Functional network
the human brain. Neuron, 72(4), 665–678.
organization of
https://doi.org/10.1016/j.neuron.2011.09.006
Power, J. D., Mitra, A., Laumann, T. O., Snyder, A. Z., Schlaggar,
B. L., & Petersen, S. E. (2014). Methods to detect, characterize,
and remove motion artifact in resting state fMRI. NeuroImage, 84,
320–341. https://doi.org/10.1016/j.neuroimage.2013.08.048
Power, J. D., Schlaggar, B. L., & Petersen, S. E. (2014). Studying
brain organization via spontaneous fMRI signal. Neuron, 84(4),
681–696. https://doi.org/10.1016/j.neuron.2014.09.007
Racine, A. M., Brickhouse, M., Wolk, D. A., & Dickerson, B. C.
(2018). The personalized Alzheimer’s disease cortical thickness
index predicts likely pathology and clinical progression in mild
cognitive impairment. Alzheimer’s & Dementia, 10, 301–310.
https://doi.org/10.1016/j.dadm.2018.02.007
Raven, J., Raven, J. C., & Court, J. H. (2003). Manual for Raven’s
Progressive Matrices and Vocabulary Scales. San Antonio, TX:
Harcourt Assessment.
Raz, N., Ghisletta, P., Rodrigue, K. M., Kennedy, K. M., &
Lindenberger, U. (2010). Trajectories of brain aging in middle-
aged and older adults? Regional and individual differences.
NeuroImage,51(2),501–511.https://doi.org/10.1016/j.neuroimage.
2010.03.020
Salat, D. H., Buckner, R. L., Snyder, A. Z., Greve, D. N., Desikan,
R. S. R., Busa, E., . . . Fischl, B. (2004). Thinning of the cerebral
cortex in aging. Cerebral Cortex, 14(July), 721–730. https://doi.
org/10.1093/cercor/bhh032
Salthouse, T. (2012). Consequences of age-related cognitive de-
clines. The Annual Review of Psychology, 63, 201–226. https://
doi.org/10.1146/annurev-psych-120710-100328
Shaw, E. E., Schultz, A. P., Sperling, R. A., & Hedden, T. (2015).
Functional connectivity in multiple cortical networks is associ-
ated with performance across cognitive domains in older adults.
Brain Connectivity, 5(8), 505–516. https://doi.org/10.1089/brain.
2014.0327
Shipley, W. C. (1940). A self-administering scale for measuring
intellectual impairment and deterioration. Journal of Psychology,
9, 371–377.
Network Neuroscience
113
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation in aging
Sporns, O., & Betzel, R. F. (2016). Modular brain networks. Annual
Review of Psychology, 67(1), 613–640. https://doi.org/10.1146/
annurev-psych-122414-033634
Sullivan, E. V., & Pfefferbaum, A. (2006). Diffusion tensor imag-
ing and aging. Neuroscience and Biobehavioral Reviews, 30(6),
749–761. https://doi.org/10.1016/j.neubiorev.2006.06.002
Suo, C., Singh, M. F., Gates, N., Wen, W., Sachdev, P., Brodaty, H.,
. . . Valenzuela, M. J. (2016). Therapeutically relevant structural
and functional mechanisms triggered by physical and cognitive
exercise. Molecular Psychiatry, 21(11), 1633–1642. https://doi.
org/10.1038/mp.2016.19
Tan, C. H., Low, K. A., Kong, T., Fletcher, M. A., Zimmerman, B.,
Maclin, E. L., . . . Fabiani, M. (2017). Mapping cerebral pulse
pressure and arterial compliance over the adult lifespan with
optical imaging. Plos One, 12(2), e0171305. https://doi.org/10.
1371/journal.pone.0171305
Tan, C. H., Low, K. A., Schneider-Garces, N., Zimmerman, B.,
Fletcher, M. A., Maclin, E. L., . . . Fabiani, M. (2016). Opti-
cal measures of changes in cerebral vascular tone during vol-
untary breath holding and a sternberg memory task. Biological
Psychology, 118, 184–194. https://doi.org/10.1016/j.biopsycho.
2016.05.008
Tan, C. H., Low, K. A., Chiarelli, A. M., Fletcher, M., A., B. Z., Kong,
T. S., Zimmerman, B., Maclin, E. L., Gratton, G., & Fabiani, M.
(in press). Optical measures of cerebral arterial stiffness are as-
sociated with white matter signal abnorMalities and cognitive
performance in normal aging. Neurobiology of Aging. https://doi.
org/10.1016/j.neurobiolaging.2019.08.004
Thambisetty, M., Wan, J., Carass, A., An, Y., Prince, J. L., & Resnick,
S. M. (2010). Longitudinal changes in cortical thickness associ-
ated with normal aging. NeuroImage, 52(4), 1215–1223. https://
doi.org/10.1016/j.neuroimage.2010.04.258
Tsang, A., Lebel, C. A., Bray, S. L., Goodyear, B. G., Hafeez, M.,
Sotero, R. C., . . . Frayne, R. (2017). White matter structural
connectivity is not correlated to cortical resting-state functional
connectivity over the healthy adult lifespan. Frontiers in Ag-
ing Neuroscience, 9(MAY), 1–13. https://doi.org/10.3389/fnagi.
2017.00144
Unsworth, N., Heitz, R. P., Schrock, J. C., & Engle, R. W. (2005). An
automated version of the operation span task. Behavior Research
Methods, 37(3), 498–505.
van Dalen, J. W., Mutsaerts, H. J. M. M., Nederveen, A. J., Vrenken,
H., Steenwijk, M. D., Caan, M. W. A., . . . Richard, E. (2016).
White matter hyperintensity volume and cerebral perfusion in
older individuals with hypertension using arterial spin-labeling.
American Journal of Neuroradiology, 37(10), 1824–1830.
Voss, M. W., Erickson, K. I., Shaurya, R., Chaddock, L., Kim, J. S.,
Alves, H., . . . Kramer, A. F. (2013a). Brain, behavior, and immu-
nity neurobiological markers of exercise-related brain plasticity
in older adults. Brain Behavior and Immunity, 28, 90–99. https://
doi.org/10.1016/j.bbi.2012.10.021
Voss, M. W., Vivar, C., Kramer, A. F., & Van Praag, H. (2013b).
Bridging animal and human models of exercise-induced brain
plasticity. Trends in Cognitive Sciences, 17(10), 525–544. https://
doi.org/10.1016/j.tics.2013.08.001
Voss, M. W., Nagamatsu, L. S., Liu-Ambrose, T., & Kramer, A. F.
(2011). Exercise, brain, and cognition across the life span. Jour-
nal of Applied Physiology, 111, 1505–1513. https://doi.org/10.
1152/japplphysiol.00210.2011
Voss, M. W., Prakash, R. S., Erickson, K. I., Basak, C., Chaddock,
L., & Jennifer, S. (2010). Plasticity of brain networks in a random-
ized intervention trial of exercise training in older adults. Fron-
tiers in Aging Neuroscience, 2(August), 1–17. https://doi.org/10.
3389/fnagi.2010.00032
Wechsler, D.
(2009). Wechsler Memory Scale. Fourth Edition.
San Antonio, TX: Pearson.
Williams, V. J., Hayes, J. P., Forman, D. E., Salat, D. H., Sperling,
R. A., Verfaellie, M., & Hayes, S. M. (2017). Cardiorespiratory fit-
ness is diff erentially associated with cortical thickness in young
and older adults. NeuroImage, 146, 1084–1092. https://doi.org/
10.1016/j.neuroimage.2016.10.033
Yang, A. C., Tsai, S. J., Liu, M. E., Huang, C. C., & Lin, C. P.
(2016). The association of aging with white matter integrity and
functional connectivity hubs. Frontiers in Aging Neuroscience,
8(June), 1–13. https://doi.org/10.3389/fnagi.2016.00143
Yeo, B. T. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari,
D., Hollinshead, M., . . . Buckner, R. L. (2011). The organization
of the human cerebral cortex estimated by intrinsic functional
Journal of Neurophysiology, 106, 1125–1165.
connectivity.
https://doi.org/10.1152/jn.00338.2011
Network Neuroscience
114
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
8
9
1
8
6
6
7
4
2
n
e
n
_
a
_
0
0
1
1
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3