研究
动态大规模网络同步
从感知到行动
乔尼·赫沃宁
1,2
, 西莫·蒙托
1,†
1
, 盛华. 王
1
, J. 马蒂亚斯·帕尔瓦
1
, 和萨图·帕尔瓦
1赫尔辛基生命科学研究所, 神经科学中心, 赫尔辛基大学, 芬兰
2生物磁学实验室, HUS医学影像中心, 赫尔辛基大学中心医院, 芬兰
†目前隶属关系: 跨学科脑研究中心, 心理学系, 于韦斯屈莱大学, 芬兰
关键词: 乙二醇, 同步, 体感, 沟通, 洞察力, 行动
开放访问
杂志
抽象的
感官引导的行动需要处理感官信息, 感性的产生
决定, 以及适当行动的产生. 这些背后的神经元活动
过程被分配到感觉, 额顶叶, 和运动脑区, 分别.
神经元处理如何在这些大脑区域之间协调以支持功能
从感知到行动仍然未知. 我们研究了相位是否同步
在大型网络中协调这些过程. 我们记录了人类皮质活动
脑磁图检查 (乙二醇) 在执行微弱体感刺激的任务时
未被察觉或被察觉. 然后我们评估了相的动态演化
通过使用源重建 MEG 数据在大规模网络中进行同步
与图论相结合的高级分析方法. 在这里我们展示了感知
弱体感刺激的报告与持续加强的体感刺激相关
Delta/Theta 中同时进行大规模同步 (3–7赫兹) 和伽玛 (40–60赫兹) 频率
乐队. 在数据驱动的网络本地化中, 我们发现这种同步是动态的
连接任务相关的, 那是, 额顶叶, 感官的, 和电机系统. 这
区域间同步的强度和时间模式也与
响应时间. 因此,这些数据表明,感知的关键大脑区域,
决策, 动作通过大规模动态阶段瞬时连接
delta/theta 和 gamma 波段同步.
作者总结
我们研究了区域间相位同步在
感知到行动的级联. 为此,我们记录了人类皮质活动
脑磁图 (乙二醇) 在执行微弱体感刺激的任务期间
被察觉或未被察觉. 我们确定了时间分辨的大规模网络
通过使用源重建 MEG 数据与高级分析工具进行同步
图论. 我们发现,对微弱体感刺激的感知和报告
与同时大规模相位同步的持续加强相关
δ/θ (3–7赫兹) 和伽玛 (40–60赫兹) 频带. 同步
表征感知刺激将感觉运动系统与额顶叶联系起来
注意力系统. 这些数据表明大规模网络同步可能
在感知过程中协调跨大脑区域的神经元处理, 决策,
当对微弱的体感刺激做出反应时.
引文: 希尔沃宁, J。, 数量, S。, 王,
S. H。, 崇拜, J. M。, & 崇拜, S. (2018).
动态大规模网络
从感知到同步
行动. 网络神经科学, 2(4),
442–463 https://doi.org/10.1162/
netn_a_00039
DOI:
https://doi.org/10.1162/netn_a_00039
支持信息:
https://doi.org/10.1162/netn_a_00039
已收到: 11 八月 2017
公认: 19 十二月 2017
利益争夺: 作者有
声明不存在竞争利益
存在.
通讯作者:
一帕尔瓦
satu.palva@helsinki.fi
处理编辑器:
奥拉夫·斯波恩斯
版权: © 2018
麻省理工学院
在知识共享下发布
归因 4.0 国际的
(抄送 4.0) 执照
麻省理工学院出版社
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
A
_
0
0
0
3
9
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
从感知到行动的大规模同步
脑磁图 (乙二醇):
测量产生的磁场
通过相干电流流入
神经元群.
源码重构:
MEG 传感器数据的逆向建模
估计底层皮质
源电流分布和时间
系列.
相位同步:
表示一致的相位
振荡之间的关系,
这里在不同的大脑区域之间.
介绍
了解控制意识知觉转化的神经机制
将感官信息转化为感知决策并生成适当的行动是
神经科学的重要挑战. 使用功能磁共振的几项研究
成像 (功能磁共振成像; 布兰肯堡等人。, 2003; 德阿纳和尚古克斯, 2011; 赫格纳等人。, 2016;
李·赫格纳等人。, 2015), 人体颅内脑电图 (脑电图; 费什等人。, 2009;
盖拉德等人。, 2009), 和非侵入性源重建电- 和脑磁-
拉菲 (脑电图和脑磁图; 希尔沃宁 & 崇拜, 2016; 金等人。, 2016; 盐等人。, 2015) 显示
前额叶的神经元活动 (全氟碳化物) 和后顶叶皮质 (个人电脑) 与那个一起在
感觉区域与意识感知呈正相关. 金等人. (2016) 最近
使用 MEG 数据的解码方法来显示大脑区域以及时间演化-
神经元处理的变化对于感知是不同的, 维护, 和可见度
以及看不见的视觉刺激.
还, 感性决策, 决定报告参议员的存在或缺席-
一些刺激或目标, 涉及人体 PPC 和 PFC 的活性 (唐纳等人。, 2009乙;
卡普兰等人。, 2017; 托索尼等人。, 2008), 猴 (西格尔等人。, 2015), 和鼠标 (戈阿尔
等人。, 2016) 局部场电位数据. 更具体地说, 有人建议参议员-
睡眠运动决策过程涉及参与准备睡眠的神经元结构-
关联动作, 例如猴子的顶叶和前额皮质的动眼神经区域
(沙尔, 2001; 哪个 & 沙德伦, 2007) 以及与反应相关的 PPC 区域和运动皮层
在人类中 (唐纳等人。, 2009乙; 古尔德等人。, 2012; 卡普兰等人。, 2017; 托索尼等人。, 2008)
(梅登多普等人。, 2011). 电机动作, 反过来, 动态调整处理
感官信息 (古特林等人。, 2011; 沃尔施拉格, 2000), 并且在行为层面,
视觉感知的节奏与动作的节奏相结合 (托马西尼等人。, 2015).
因此,这些证据表明感知和认知之间存在多层次的耦合。
行动, 但这种合作如何在神经元水平上实现仍然不完全
明白了. 实现这种协调的一种候选机制是神经元同步-
扎化, 已被证明可以调节神经元通讯 (薯条, 2015; 格雷戈里奥
等人。, 2009; 马里斯等人。, 2016; 西格尔等人。, 2012; 歌手, 2009) 并提供一个临时的
基于相关性的框架,用于将信息处理集成到神经元回路中
多尺度 (薯条, 2015; 施罗德 & 锁匠, 2009; 西格尔等人。, 2012). 在人类中, 新-
大脑皮层大部分区域的脑电波活动可以通过脑电图无创地记录
和乙二醇, 但使用这些方法来估计远程同步需要
一种基于源重建的途径和方法来解决残余物的挑战
信号泄漏 (布鲁克斯等人。, 2012; 崇拜 & 崇拜, 2012; 锄 & 总的, 2009). 预利姆-
来自视觉形态的初步证据表明,
视觉和额叶大脑区域确实与视觉感知相关 (希普等人。, 2011)
以及运动皮质区域和周围神经系统之间的执行
运动动作 (锄 & 总的, 2009). 尽管这些研究提供了部分支持
感知-行动循环可以通过相位同步来实现的想法, 这
连接感知和行动的完整网络架构仍未得到解决.
我们假设动态大规模网络相位同步是
从感知到感知决策和行动的神经元处理的协调
通过调节 PFC 之间的神经元处理, 个人电脑, 以及感觉和运动系统.
为了检验这个假设, 我们在体感刺激检测任务中记录了 MEG
其中受试者在脱粒处接受恒定电流的体感刺激-
被感知或未被感知的旧知觉 (希尔沃宁 & 崇拜, 2016).
网络神经科学
443
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
A
_
0
0
0
3
9
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
从感知到行动的大规模同步
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
A
_
0
0
0
3
9
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
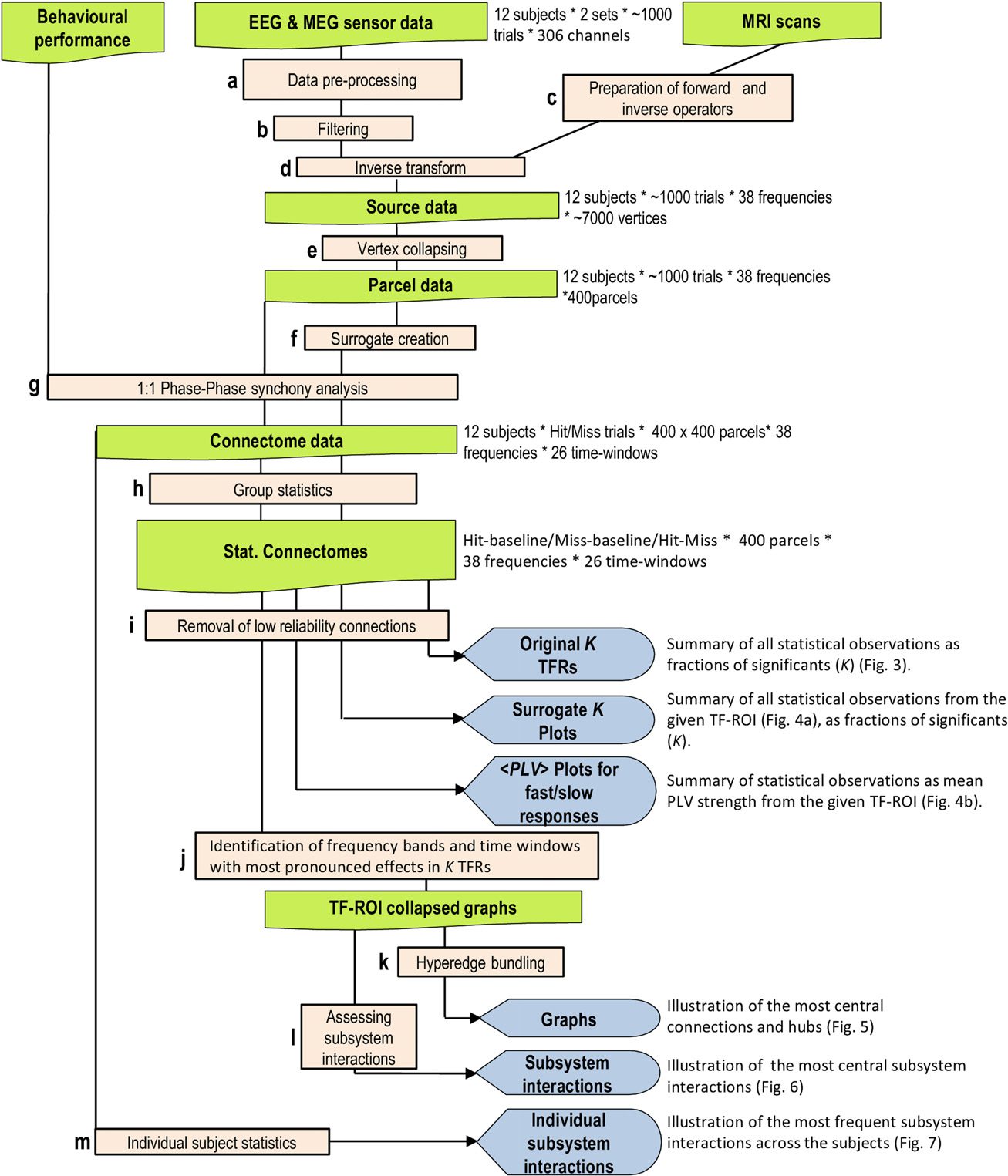
数字 1. 分析流程的示意图. 该图显示了不同的分析
步骤和结果输出 (上午至下午, 参见方法).
然后我们使用基于 MEG 源建模的全相耦合分析 (数字 1) 到
识别表征感知-动作序列相关神经元的皮质网络
加工. 我们发现并发 delta/theta- 和伽玛波段同步连接
体感, 额顶叶, 和运动皮质区域感知到的刺激
以及未检测到的刺激的这种耦合的缺失. 这些研究结果支持-
证据表明神经元相位同步在功能上发挥着重要作用
感觉的配合, 决定, 从感知到行动的运动阶段 (数字 1).
结果
行为表现
受试者在 MEG 记录期间的任务是检测弱信号, 恒强度电
在两次 30 分钟的训练中,以随机间隔对右手食指指尖进行刺激
之间 1.5 和 4.5 s. 刺激强度已校准至检测阈值
记录之前并在 MEG 记录期间保持恒定. 因此, 我期间
录音, 这些刺激被不同程度地检测到 (点击数) 或仍未被发现 (未命中数)
(图2A). 命中率 (人力资源) 曾是 38.32 ± 15.26% [平均值±标准差 (标清)], 和
网络神经科学
444
从感知到行动的大规模同步
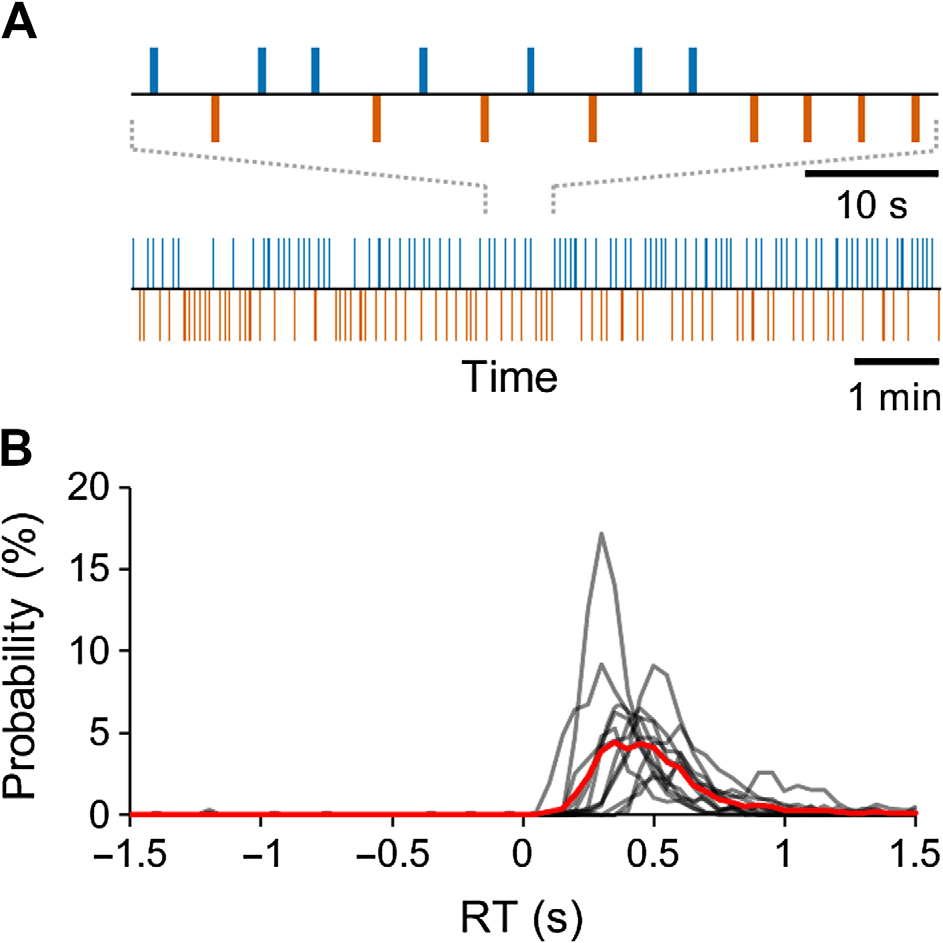
数字 2. 任务和行为表现. (A) 实验段落示意图-
digm 具有一系列恒定强度的体感刺激 1.5- 至 4.5 秒间隔
以及代表性受试者相应的行为命中-未命中时间序列. 蓝色条表示
有意识地感知刺激, 红色条表示未察觉的刺激. (乙) 个人 (黑色的
线) 和组 (红线) 响应时间 (RT) 分布.
反应时间 (RT) 曾是 399 ± 135 多发性硬化症 (平均值±标准差). 我们估计了错误的比例
警报 (F A), 那是, 没有刺激时的反应, 从之前的时间窗口开始-
电刺激 (图2B, 参见方法部分). 基于有限状态模型
方法, 我们估计 FA 率为 0.2 ± 0.3% (平均值±标准差) 所有试验中, 这给出了一个真实的
刺激检测率 38.12%.
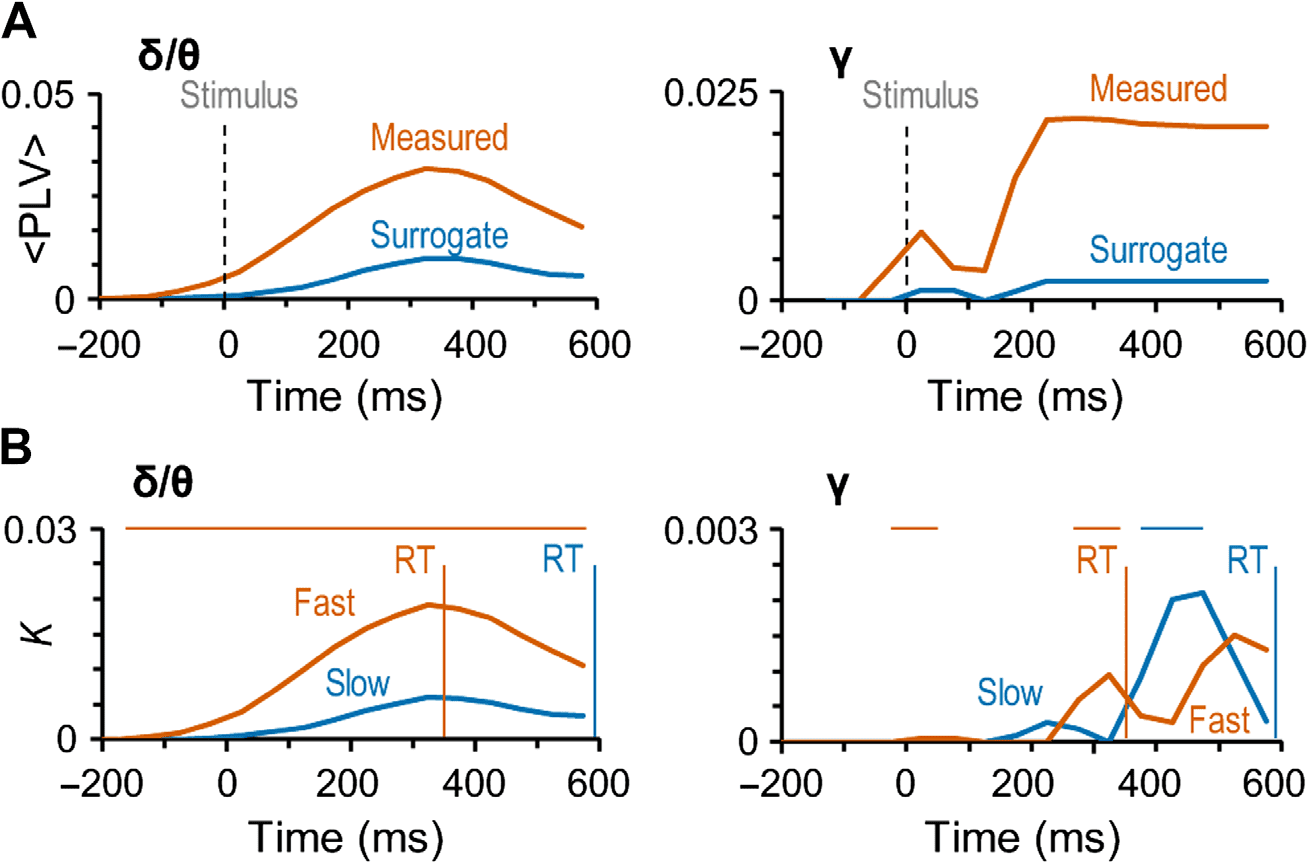
持续的大规模 Delta 和 Gamma 同步是感知-行动循环的特征
研究大规模同步是否会表征神经元过程-
从感知到行动,并绘制这些假定的解剖结构和频率模式
同步, 我们测试了同步是否会表征
那些被报告感知到的刺激. 因此,我们表征了相位同步
每个受试者的所有皮质区域, 时间窗口, 和频段分别进行试验
近感官刺激被认为是感知的 (点击数) 或不为人知
(未命中数). 我们将区域间相位同步的统计显着观察表示为
皮质区域为顶点、重要连接为边缘的图表 (布莫尔
& 斯波恩斯, 2009; 帕尔瓦等人。, 2010; 鲁比诺夫 & 斯波恩斯, 2010). 同步程度为
用连接密度估计 (K), 这是显着区域间的比例-
来自所有可能的成对相互作用的相互作用 400 我们皮质中的大脑区域
分割. K 的频谱表明,中伽马同步
频带 (中γ, 40–60赫兹) 在 150–500 ms 和 Delta/Theta 中 (d–th, 3–7赫兹) 乐队
从 0 到 500 刺激开始后的毫秒数被加强到高于基线水平
(Wilcoxon 符号秩检验, p < 0.05, corrected for multiple comparisons) when interareal
synchronization was measured with the phase-locking value (PLV; Figure 3A). An essentially
identical result was obtained when the imaginary part of PLV (iPLV) that is insensitive to source
mixing was used to measure synchronization (Figure 3B), which showed that delta and gamma-
band synchronization were not attributable to signal mixing and artificial synchronization.
445
Connection density:
A proportion of connections in a
graph out of all possible connections;
here connections were defined by
their statistical significance.
Parcellation:
Division of the cortical surface into
parcels that here correspond to
neuroanatomically named brain
areas.
Phase-locking value (PLV):
The absolute value of complex
PLV, quantifies the strength
of phase synchronization.
Imaginary part of complex
PLV (iPLV):
A phase synchronization measure
that is sensitive only to nonzero
and non-pi phase lag couplings
and thereby excluded the direct
effects of linear mixing.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
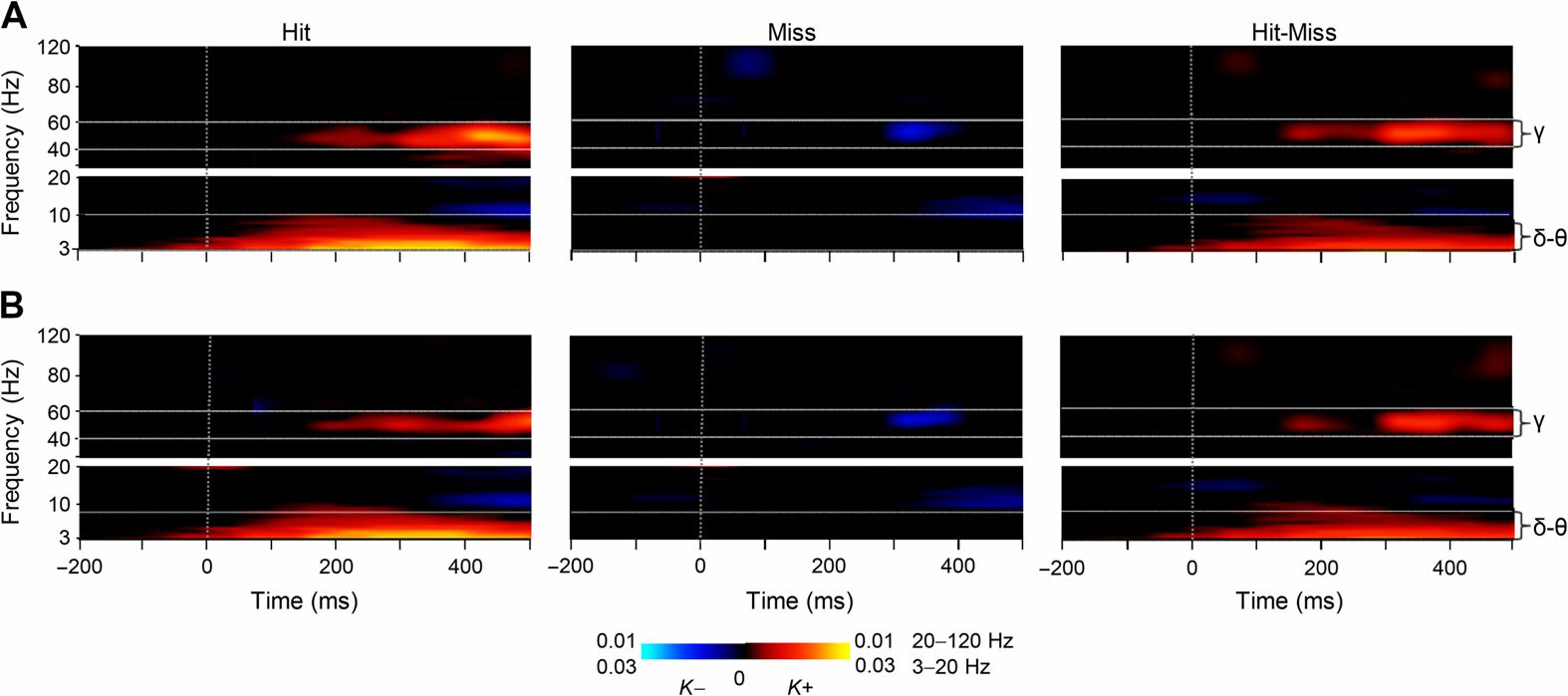
In contrast, interareal synchronization was largely insignificant (Wilcoxon signed-rank test,
p < 0.05, corrected for multiple comparisons) for Misses and in the gamma band even briefly
suppressed below the baseline level both when estimated with PLV (Figure 3A) and iPLV
(Figure 3B). Importantly, this suppression shows that the lack of synchronization for Misses was
not due to poor signal-to-noise ratio for missed stimuli, but that Hits and Misses were associ-
ated with genuinely distinct synchronization patterns. A comparison between Hits and Misses
confirmed that both large-scale delta- and gamma-band synchrony were indeed stronger for
Hits than Misses (Wilcoxon signed-rank test, p < 0.05, corrected) estimated with both PLV
(Figure 3A) and iPLV (Figure 3B).
Observations of large-scale synchronization can also arise artificially from evoked responses
or phase locking of ongoing activity to the stimulus onsets (Palva & Palva, 2012). In the present
data, early transient phase locking of ongoing oscillations to stimulus onset as well as transient
amplitude increase was observed for both Hits and Misses and was stronger for Hits than
Misses between 3 and 30 Hz (Hirvonen & Palva, 2016) (Supporting Information Figure 2,
Hirvonen, Monto, Wang, Palva, & Palva, 2018). Sustained phase locking of oscillations to
stimulus onset and sustained oscillation amplitude increases during the time windows of large-
scale delta/theta- and gamma-band phase synchronization were observed only in the low-
alpha and delta/theta frequencies. In the delta/theta band both phase locking and oscillation
amplitudes were increased, whereas in the alpha band, phase locking to stimulus onset was
increased but the oscillations amplitudes were more suppressed for the Hits than for Misses.
Evoked/stimulus-locked activity:
A cortical response with a
consistent latency after stimuli
and thus observable in
trial-averaged response.
These findings hence indicate qualitatively that large-scale gamma-band synchronization
cannot be caused by evoked or stimulus phase-locked activity. To confirm this as well as to
assess the contribution of evoked activity in the observed delta/theta band large-scale syn-
chronization, we compared synchronization data against surrogate data where non-stimulus-
locked phase relationships were eliminated but evoked/stimulus-locked components were
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Large-scale delta- and gamma-band synchronization characterizes neuronal process-
ing of perceived but not unperceived stimuli. (A) Time-frequency representations of the extent of
significant interareal synchronization as estimated with PLV for perceived (Hit) stimuli, unperceived
(Miss), as well as for their difference (Hit-Miss) compared with the prestimulus baseline (N = 12)
(Wilcoxon signed-ranked test, p < 0.05, corrected for multiple comparisons). Sustained gamma-
band (40–60 Hz) synchronization and delta/theta- (δ/θ, 3–7 Hz) band synchronization were stronger
for Hits than Misses. The color indicates the connection densities of positive (K+) and negative (K−)
observations, that is, the fractions of connections with a statistically significant positive or negative
difference from the baseline level, respectively. (B) Time-frequency representations of the extent of
significant interareal synchronization as estimated with iPLV.
Network Neuroscience
446
Large-scale synchronization from perception to action
Figure 4. Gamma- and delta/theta-band synchronization is not artificial and predicts the speed
of sensorimotor decisions. (A) The mean coupling strength (
原著中的热门歌曲 (红线) 和替代数据 (蓝线) δ/θ- (不适用) 和伽玛- (C)
频带. (乙) 连接密度 (K) 作为点击之间差异的时间函数
和 分别用于快速试验的错过 (红色的) 且缓慢 (蓝色的) RT. 垂直线显示平均值
这两类的RT: 356 ± 122 多发性硬化症 (各受试者的平均值±标准差) 用于快速 RT 的试验
和 594 ± 229 慢速 RT 试验的毫秒数. 图表上方的水平条显示时间
快速和慢速试验 RT 的试验之间存在显着差异的窗口 (威尔科克森签名排名
测试, p < 0.05, corrected for multiple comparisons). The trials were split at the median RT of each
subject and the mean of these median RTs was 464 ± 142 ms (mean ± SD across subjects).
preserved (see Methods). We compared the strengths of phase synchrony in original data
against the corresponding mean values of surrogate data. Importantly,
in the delta/theta
(Figure 4A, left) and specifically in the gamma-band (Figure 4A, right) synchronization was
significantly stronger in the original than surrogate data for Hits. These analyses thus unequiv-
ocally indicate that neither the large-scale delta/theta- or gamma-band synchronization ob-
served for detected stimuli can be explained by artificial synchronization caused either by
phase locking of ongoing oscillations to stimulus onset or additive evoked responses, or by
confounders such as stimulus-altered local source topographies or autocorrelations.
Correlation of Large-Scale Synchronization with the Motor Responses
Our main goal was to reveal the networks underlying the coordination of processing from per-
ception to action. To thus identify whether and when synchronization is locked to actions, that
is, the motor responses rather than related to perceptual processing, we investigated whether
the strength of the observed large-scale delta/theta and gamma-band synchronization would be
correlated with RTs. We divided the responses within each subject into “fast” (356 ± 122 ms)
and “slow” (594 ± 229 ms) response categories by the individual median RT (464 ± 142 ms,
mean ± SD across subjects). We then estimated the strength of synchronization separately for
these two categories of responses. This analysis indicated that for fast and slow responses both
delta/theta band and gamma bands had distinct synchronization patterns (Figure 4B). For in
delta/theta, synchronization was stronger for the fast than for the slow responses during the
whole response time (Wilcoxon signed-ranked test, p < 0.05, corrected for multiple compar-
isons). However, gamma-band synchronization differed for the fast and slow responses only
Network Neuroscience
447
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
Subsystem:
A set of brain areas defined by
intrinsic connectivity and typically
corresponding to areas with
associated functional roles.
during the different peak latencies, which showed that for the fast responses, gamma-band
synchronization began around 200 ms from stimulus onset, whereas for the slow responses
it began 350 ms from stimulus onset (Wilcoxon signed-ranked test, p < 0.05, corrected for
multiple comparisons). These very different patterns of correlations with the RTs suggest that
although delta/theta synchronization coordinates directly the motor actions, the functional
significance of gamma-band synchronization in coordinating motor actions is dependent on
perceptual and/or decision processes that have large temporal variability.
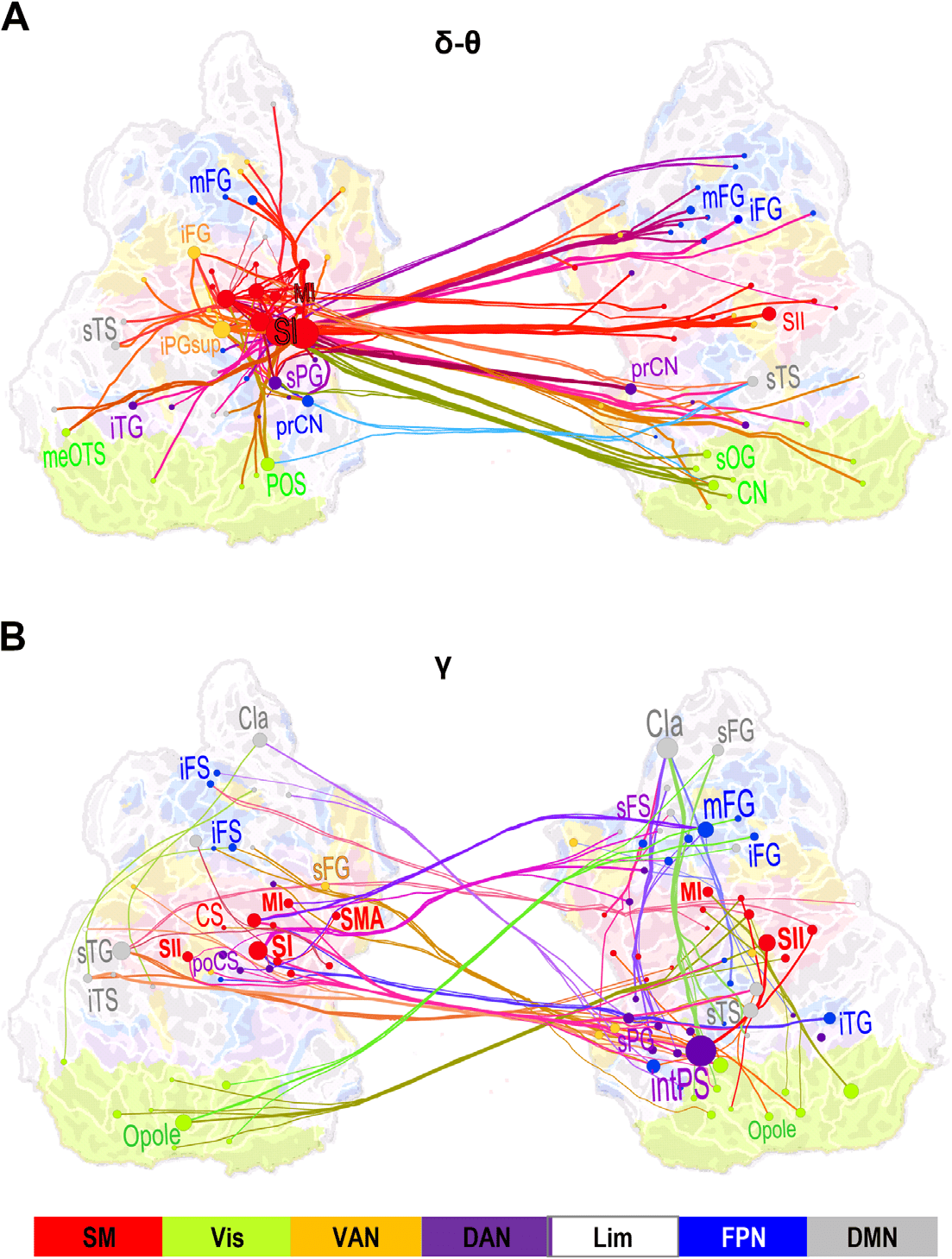
Large-Scale Synchrony Connects Sensorimotor Areas with Prefrontal and Posterior Parietal Cortices
To identify the cortical areas connected by large-scale delta/theta- and gamma-band synchro-
nization, we identified both the most central of the significant interareal connections and key
cortical areas, that is, the network hubs, by using PageRank for those time-frequency win-
dows where synchronization was significantly greater for Hits than for Misses. To obtain a
coarse localizer for the functional subsystems of the cerebral cortex, we co-localized our cor-
tical parcellations with the seven predominant systems defined by intrinsic fMRI BOLD signal
correlations (Yeo et al., 2011). In the delta/theta band, the strongest connections for the differ-
ence between Hits and Misses were observed bilaterally in the sensorimotor (SM) system in the
parcels corresponding to the contralateral primary sensory area (SI), secondary somatosensory
areas (SII), and motor area (MI). Additionally, delta/theta-band synchronization connected SM
with the bilateral middle frontal gyrus (mFG) in the dorsolateral prefrontal cortex (dlPFC) and
with nodes in the visual cortex (Figure 5A). In the mid-gamma band, the most central con-
nections were observed between contralateral SM, the most central hubs being SI, SII, and
MI, and the ipsilateral mFG and bilateral inferior frontal gyrus (iFG) of the dPLFC as well as
with the intPS of the PPC (Figure 5B). Functionally, mFG and iFG belong to the fronto-parietal
network (FPN) whereas intPS belongs to the dorsal attention network (DAN). Gamma-band
synchronization hence connected SM to FPN and DAN. In addition, mid-gamma-band syn-
chronization connected mFG and intPS ipsilaterally.
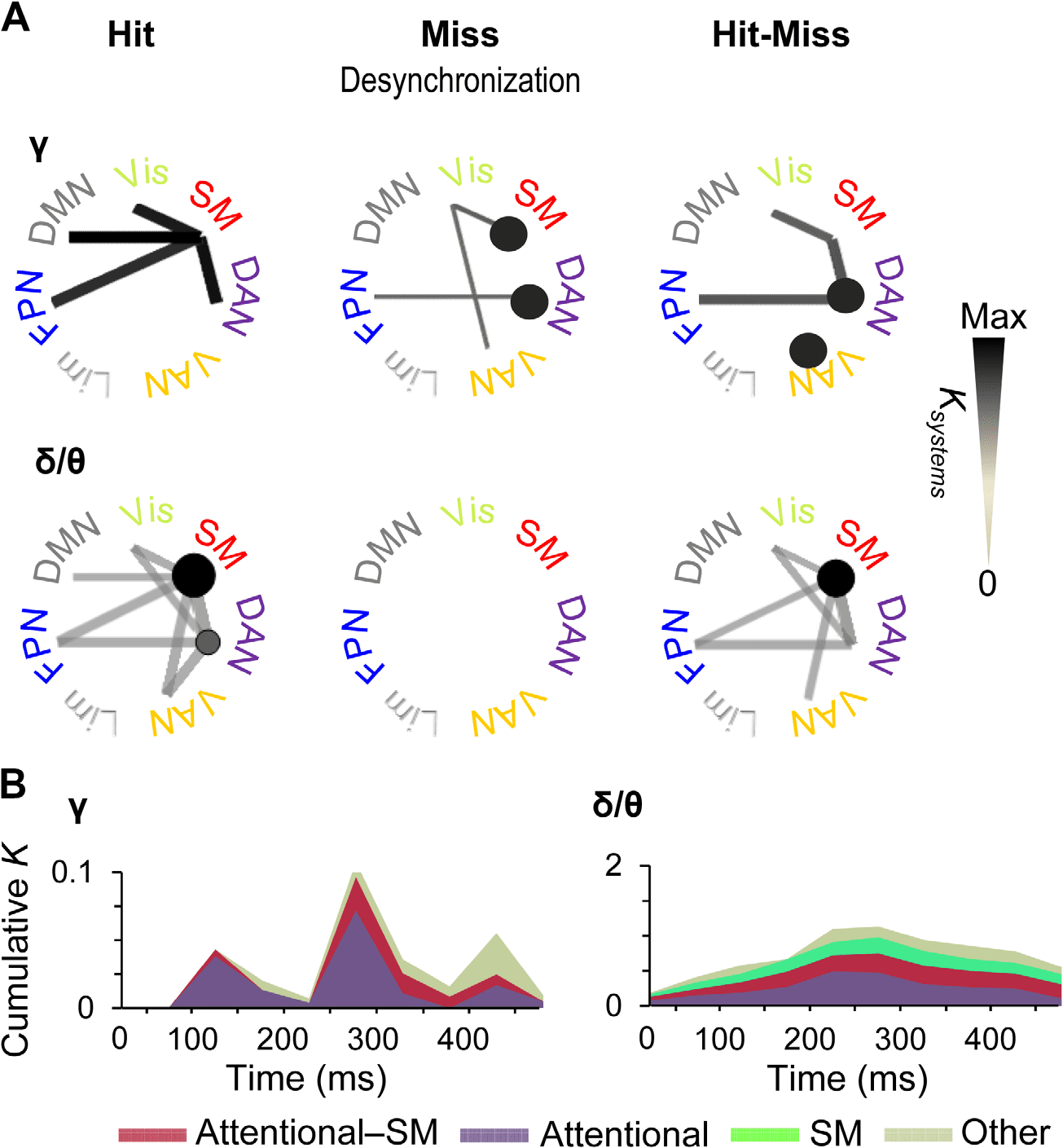
Large-Scale Synchrony Among Functional Subsystems
To complement the analyses of strongest connections, we obtained a systems-level view of all
significant observations of interareal connectivity by identifying which functional brain sys-
tems were coupled by greater fractions of significant connections (Ksystems) than expected by
chance (multiple-comparisons controlled graph permutation test, p < 0.05, see Methods).
This analysis showed that for Hits, gamma-band synchronization connected the SM system
with DAN and FPN as well as with the default mode network and visual system (Figure 6A).
Interestingly, for Misses, the suppression of synchronization below baseline levels was found
both within SM and DAN as well as among SM, ventral attention network (VAN), and the
visual system, and between DAN and FPN. Hence, when the stimuli were not detected, atten-
tional networks were uncoupled by suppression of gamma-band synchronization. In addition,
the significant differences in synchronization between Hits and Misses were more prevalent
between DAN and SM, within DAN and within VAN, and between DAN and FPN than ex-
pected for comparable random networks. Thus, these system-level results corroborate the
findings of the most central connections and showed that gamma synchronization among so-
matosensory and attentional systems predicts stimulus detection in both the delta/theta- and
gamma bands. Hit-related synchronization was found among SM and attentional systems for
both frequency bands, the key difference being that in the delta/theta band, within-system
synchronization was found in SM, whereas in the gamma band it was robust within DAN and
VAN.
Network Neuroscience
448
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Large-scale synchronization connects somatosensory and attentional brain systems. (A)
Graph of the significant differences in the strength of interareal phase synchrony as estimated with
PLV between Hits and Misses in the delta/theta-frequency band (δ/θ, 3–7 Hz) and in the time win-
dow of 125–275 ms from stimulus onset (cf. Figure 2C). Lines connect the coupled parcels and line
colors are determined by the parcel brain systems (see below). Delta/theta-band synchronization
was centered on the contralateral (left hemispheric) sensorimotor (SM, red) system, and in particu-
lar, on the primary somatosensory cortex (SI) therein that was strongly and bilaterally coupled with
frontal and parietal regions. (B) Graph of gamma-band synchronization (γ, 40–60 Hz) reveal sig-
nificantly stronger connections for Hits than Misses over 225–375 ms from stimulus onset. Stronger
gamma-band synchronization for Hits than Misses was observed within SM and between SM and
the ipsilateral frontoparietal and dorsal (FP and DA, blue and purple) attention networks. Graphs
are displayed on an inflated and flattened cortical surface with 300 (A) and 200 (B) of the most
central edges based on parcel PageRank centralities selected for visualization. SI is primary and
SII is secondary somatosensory area. MI and SMA are primary and supplementary motor areas, re-
spectively. Parcel and corresponding node colors indicate the Yeo-atlas brain systems derived from
BOLD intrinsic connectivity connectome. SM = somatomotor (SM), green = visual (Vis), yellow =
ventral attention network (VAN), purple = dorsal attention network (DAN), white = limbic (Lim),
gray = default mode network (DMN).
To illustrate the temporal evolution of the system-scale gamma synchronization and specif-
ically to compare quantitatively the contributions of task-positive attention and sensory net-
works, we estimated the total connection densities of significant system-system couplings
separately for within-attentional, attentional-sensorimotor, and all other systems (Figure 6B).
Network Neuroscience
449
Large-scale synchronization from perception to action
Here, attentional systems included FPN, DAN, and VAN, whereas the “other” systems included
the default mode, visual, and limbic systems. We found system-level gamma synchronization
to emerge in three waves. In the first wave at around 100–150 ms from stimulus onset, connec-
tivity was almost completely dominated by coupling within the attentional systems, with only a
minor sensory-attentional fraction (Figure 6B, left). In the second wave at around 250–300 ms,
coupling within-attentional systems remained predominant, but sensory-attentional coupling
became much more salient. In the third wave, at around 450 ms from stimulus onset in which
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Time-varying subsystem connectivity within and between attentional systems and SM.
(A) Connection densities of significant interareal gamma-band (40–60) Hz and delta/theta-band
(3–7 Hz) synchronization among Yeo-atlas brain systems for Hits and Misses compared with base-
line and for their difference. Only such system-system connections are shown that exhibit greater
connection densities than expected by chance in shuffled graphs (p < 0.05, permutation statistics,
see Methods). The color, line width, and radius of circles of the system-system connections indicates
the connection density of significant couplings (Ksystems) within (circles) or between (lines) the func-
tional systems in a time window of 225–375 ms. (B) Time-resolved cumulative connection densi-
ties (K) of gamma- and delta/theta-band synchronization estimated separately for within-attentional
(DAN, FNP, and VAN), between SM and attentional, and all other functional subsystems. Synchro-
nization in task-positive sensory and attentional systems predicted subsequent conscious perception
in gamma band, whereas synchronization in the delta/theta band was observed in SM along with
connections to and within-attentional systems throughout the time course of stimulus detection.
Network Neuroscience
450
Large-scale synchronization from perception to action
on average also the behavioral responses were given, connectivity in other systems was most
prevalent. In contrast, delta synchronized emerged in one wave, which peaked between 200
and 300 ms after stimulus onset. Furthermore, it predominantly comprised synchronization
within SM and attentional systems and between these two (Figure 6B, right).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
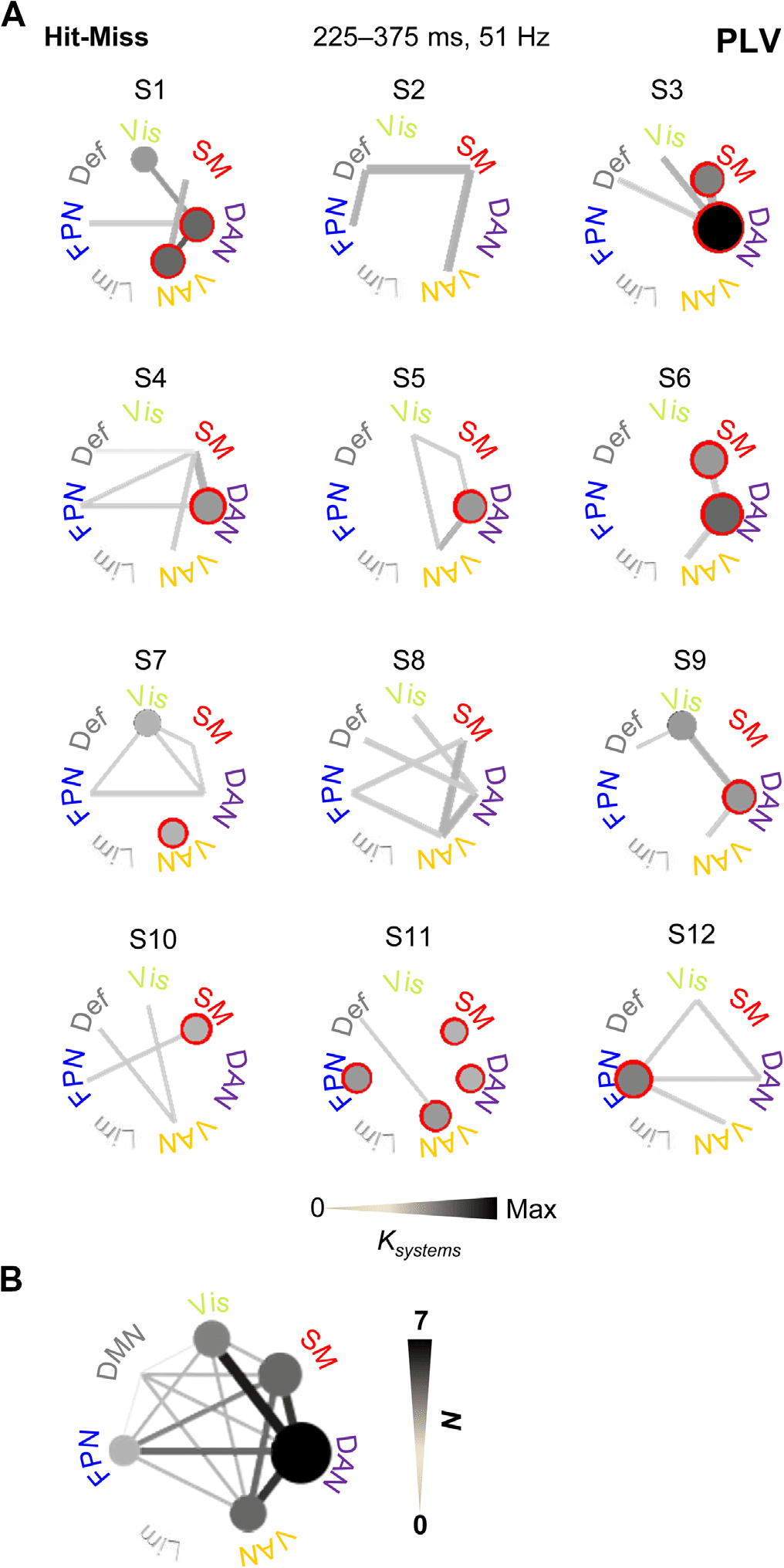
Figure 7. Synchrony within and between attentional and sensorimotor systems is frequently ob-
served in individual subject’s statistical analyses. (A) All significant connections among functional
subsystems separately for each subject. Significant connections are shown with gray tone that is
scaled relative to the connection density between (lines) or within (circles) the subsystems. Red
surroundings indicate the most common within-subsystem connections across the subjects. (B) All
significant connections across all subjects in individual subject statistical analyses. Line color indi-
cates the number of subjects (N) in which significant connections were were observed.
Network Neuroscience
451
Large-scale synchronization from perception to action
Single-Subject Statistics Corroborate The Robustness of Gamma-Band Synchronization in the
Sensorimotor Decision Process.
To corroborate the group-level analyses and to assess the robustness of gamma-band synchro-
nization in single subjects, we investigated individual interareal synchrony with single-subject
permutation statistics for the mid-gamma band (51 Hz) in the time window of 225–375 ms
where synchronization was most extensive in the group data. We estimated subsystem-level
connectivity as above separately for significant connections across subjects. This approach re-
vealed that synchronization was most prominent within SM, FPN, DAN, VAN, and the visual
system, and most repeatedly connected SM with DAN as well as DAN with VAN and the visual
system (Figure 7A). Even with considerable interindividual variability, in 9 out of 12 participants
the SM system was connected with an attentional system, and in all subjects synchronization
was observed within and/or between these functional systems (Figure 7B). These findings thus
confirmed that the correlation of large-scale gamma-band synchronization among SM, FPN,
DAN, and VAN with perception is a robust phenomenon even in individual subjects.
DISCUSSION
We aimed in this study to identify the large-scale networks of phase-locked cortical regions
supporting the neuronal processing from perception to perceptual decisions and to actions.
Building on accumulating evidence for the putative essential roles of large-scale neuronal inter-
action networks in cognitive integration (Baker et al., 2014; de Pasquale et al., 2015; Deco
et al., 2015; Petersen & Sporns, 2015; Siegel et al., 2012) and of neuronal phase coupling
in regulating the information transfer (Fries, 2015), we assessed the role of dynamic large-
scale phase synchronization in the emergence of coordinated neuronal processing that ac-
complishes the transition from sensory perception to action generation.
This is the first study to show, using human source-reconstructed noninvasive MEG or EEG
data, that dynamic interareal synchronization characterizes the neuronal activity cascade from
perception of sensory stimuli, to perceptual decisions, and to actions. We revealed the spa-
tiotemporal structure of this synchronization by using time-resolved synchrony analyses and
localized the key cortical areas of synchronization networks. Importantly, we found that the
strength of this synchronization was correlated with both detection performance (HR) and RTs,
and hence is functionally significant.
We observed that two concurrent networks of transient synchronization, one in the delta/
theta- and another in the gamma-frequency band, connected the sensory, fronto-parietal, and
motor systems from perception to action. Delta/theta-band synchronization was pronounced
within the SM system and between SM and attentional systems. In contrast, gamma-band syn-
chronization was first observed within the fronto-parietal attention networks, and subsequently
also between attention networks and the SM system, but it was not found to connect nodes
within SM. These couplings connecting the key task-relevant cortical structures suggest that
they could be functionally significant in the perception-action cycle. In line with their func-
tional significance, the strength of delta-band and the timing of gamma-band synchronization
were associated with the intertrial variability of the subsequent response times. These data thus
demonstrate that the large-scale network interaction mechanisms achieving perception, per-
ceptual decisions, and generation of actions may be achieved by phase coupling of neuronal
oscillations in delta/theta and gamma bands.
These data now show that in addition to evoked responses and local modulations of
oscillation amplitudes (Blankenburg et al., 2003; Hirvonen & Palva, 2016; Jones et al., 2007;
Network Neuroscience
452
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
King et al., 2016; Nierhaus et al., 2015; Taskin et al., 2008), somatosensory perception is also
characterized by concurrent induced neuronal synchronization connecting the task-relevant
cortical areas. In summary, these data provide evidence for the overarching hypothesis that
neuronal synchronization may coordinate communication during cognitive operations (Fries,
2015) and that long-range synchronization may play a key role in the emergence of conscious
sensory perception (Dehaene & Changeux, 2011; Engel & Singer, 2001; Tallon-Baudry, 2012)
and sensorimotor decisions (Donner et al., 2009a; Engel & Fries, 2010).
Long-Range Synchrony May Coordinate Processing from Perception to Action
The interest toward large-scale phase coupling as a putative mechanism for the emergent
coordination of distributed processing into coherent cognitive operations stems from two
established lines of evidence. First, phase coupling—synchronization—in a given neuronal
assembly endows greater postsynaptic impact than internally uncoupled, incoherent assem-
blies (Konig et al., 1995; Singer, 1999). Second, interareally organized phase relationships or
phase coupling may play a powerful role in opening or closing “channels” for communication
among brain areas by facilitating communication during the high-excitability phases of neu-
ronal oscillations (Fries, 2015). Cellular-level and animal model studies thus provided a strong
mechanistic framework for the likely functional roles of interareal phase coupling. None-
theless, because of significant methodological challenges, many of the resulting hypotheses
have remained unaddressed at the level of large-scale cortical interaction networks and their
relationship with human cognitive operations.
Prior fMRI-EEG studies have shown that activity in sensory and fronto-parietal brain areas is
related to both conscious perception and generation of actions in response to somatosensory
stimuli (Blankenburg et al., 2003; Nierhaus et al., 2015; Taskin et al., 2008), whereas EEG and
MEG studies have shown that early evoked responses are stronger for detected somatosensory
stimuli (Hirvonen & Palva, 2016; Jones et al., 2007; Nierhaus et al., 2015). Yet, the role of large-
scale network synchronization in coordinating the perception and actions has remained poorly
understood. A seminal study using intracranial EEG (iEEG) recordings from epileptic patients
found that long-distance beta synchronization between frontal, parietal, and visual areas char-
acterized visually presented words when these were consciously perceived but not if they re-
mained undetected (Gaillard et al., 2009). Whether similar long-range synchronization would
characterize also the perception of sensory stimuli in other modalities, in different experimen-
tal paradigms, and in MEG/EEG data obtained over the whole cortex has remained elusive.
Our central aim was to investigate whether large-scale networks of neuronal phase syn-
chronization would be essential in the transformation of sensory processing to sensorimotor
decision and actions. To this end, we used MEG recordings combined with several advanced
data analysis approaches and graph theory (Bullmore & Sporns, 2009) to identify the spa-
tiotemporal patterns of network synchronization from the perception of weak somatosensory
stimuli presented at a threshold of detection to the action. We found that as hypothesized,
dynamic large-scale synchronization characterized this process. Large-scale synchronization
was observed concurrently in delta/theta and gamma bands showing that even the simplest
perceptual process involves dynamic synchronization in multiple frequencies. Using a novel
parcellation scheme (Korhonen et al., 2014), combined with the novel edge-bundling
approach (Wang et al., 2018), we also identified the network structures of these networks.
Delta/theta-band synchronization connected the contralateral SI with ipsilateral SI and SII bi-
laterally and thereby the brain regions underlying the processing of somatosensory information
in monkeys (Haegens et al., 2014; Tauste Campo et al., 2015) and humans (Hegner et al., 2016;
Network Neuroscience
453
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
Hirvonen & Palva, 2016; Jones et al., 2007; Jones et al., 2009; Li Hegner et al., 2015; Papadelis
et al., 2016). This connectivity pattern positions delta/theta-band synchronization to directly
support the processing of somatosensory information. These nodes were also connected to the
motor cortex (MI) as well as to the contralateral PFC supporting a close link and dynamic inter-
actions between perceptual (Gutteling et al., 2011; Wohlschlager, 2000) and decision-making
processes (Donner et al., 2009b; Gould et al., 2012; Kaplan et al., 2017; Tosoni et al., 2008)
with the coordination of motor actions. Delta/theta-band synchronization could thus underlie
the coordination of neuronal processing achieved collectively in motor and sensory cortices.
In contrast, gamma-band synchronization did not connect cortical areas within SM but
coupled the contralateral SI and SII with ipsilateral sFS, mFG, and intPS. These areas are the
key nodes in the fronto-parietal and DAN, FPN, and DAN, as indicated both by the whole-
brain as well as by the subsystem analyses (Murakami & Okada, 2006; Power et al., 2011;
Spadone et al., 2015), respectively. These FPN/DAN nodes were also mutually connected
in both hemispheres, suggesting that gamma-band synchronization could here coordinate
neuronal communication specifically within the attentional system and between the atten-
tional and sensory systems. In summary, these data revealed many subnetworks among the key
cortical areas that were correlated with conscious somatosensory perception, decisions, and
actions in great anatomical and temporal precision. Unexpectedly, the data-driven all-to-all
data analysis approach also revealed the visual system to be consistently connected with the
sensorimotor system both in delta/theta and gamma bands. This may reflect the responsive-
ness of the visual cortex also to the somatosensory stimulation (Nordmark et al., 2012) and/or
complementary representation of the perceived somatosensory stimuli in the visual cortex
(Orlov et al., 2010), which is also plausible as the subjects had eyes closed during the task.
Suppression of Gamma-Band Synchronization for Misses
Intriguingly, we found gamma-band synchronization to be suppressed below baseline levels
for stimuli that remained undetected. These stimuli were hence subliminal in the terms that
they modulated brain activity but in an inadequate manner to reach conscious perception
(Dehaene et al., 2006; Dehaene & Changeux, 2011). Prior studies have reported that sublimi-
nal stimuli evoke neuronal activity in primary sensory cortices but not later in sensory process-
ing hierarchy for visual stimuli (Dehaene et al., 2001; Del Cul et al., 2007; Grill-Spector et al.,
2000). Our data showed that not only is the synchronization stronger for the consciously per-
ceived and reported than for unperceived and unreported stimuli, but also that synchronization
patterns between Hits and Misses are qualitatively and genuinely distinct. The suppression of
gamma activity for Misses is, however, well in line with prior fMRI studies reporting negative
BOLD fMRI signal responses in contralateral SI, SII, and SMA for subthreshold somatosen-
sory stimuli (Blankenburg et al., 2003; Nierhaus et al., 2015; Taskin et al., 2008). Suppres-
sion of synchronization may reflect active uncoupling of task-relevant cortical areas to block
conscious access from subliminal stimuli or it may reflect non-conscious stop-signals to inter-
rupt motor responses (van Gaal et al., 2008).
Temporal Pattern of Large-Scale Synchronization and Correlation with RT Supports Distinct Functions
for Delta/Theta and Gamma-Band Synchronization in Perceptual Decisions and Actions
Finally, to reveal whether network synchronization would be stable or show dynamic changes
as a function of time, we used time-resolved analysis of network synchronization. Interestingly,
despite the stable profile of gamma-band synchronization in the time-frequency representa-
tions, this analysis showed that the gamma-band connectivity patterns changed as a function
Network Neuroscience
454
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
of time while those of delta-band synchronization pattern were more stable. During the early
time windows, gamma-band synchronization connected nodes only within the attention net-
works, after which the gamma-band network comprised also the SM system, including motor
cortex (M1). This temporal pattern suggests that in the present task, attentional systems drive
sensory perception in line with prior studies, which suggest that perception is related to differ-
ences in the prestimulus network properties (Weisz et al., 2014; Frey et al., 2016) and predicted
by infraslow fluctuations of electrical activity (Monto et al., 2008). Furthermore, fMRI BOLD
signal fluctuations in attentional but not in sensory system are also correlated with percep-
tual performance in a threshold-stimulus detection task like ours (Sadaghiani et al., 2009). As
PFC and PPC coordinate both perceptual decisions (Donner et al., 2009b; Goard et al., 2016;
Hegner et al., 2016; Kaplan et al., 2017; Siegel et al., 2015; Tosoni et al., 2008) and conscious
perception (Dehaene & Changeux, 2011; Gaillard et al., 2009; Hirvonen & Palva, 2016;
Li Hegner et al., 2015; Salti et al., 2015), we speculate that gamma-band synchronization
during the later time windows, however, could underlie conscious perceptual decisions of the
weak somatosensory stimuli. The role of gamma-band synchronization in coordinating percep-
tual decisions was also supported by the distinct temporal profiles of synchronization for fast
and slow actions. Interestingly, fast and slow responses were associated with distinct gamma-
band patterns, which illustrates the presence of large intertrial variability in the functionally
significant gamma-band synchronization patterns. More specifically, the neuroanatomical and
temporal patterns of gamma-band synchronization could plausibly reflect a sequence of net-
work reconfigurations from conscious sensory decisions to the coordination of motor actions.
In contrast, the temporal profile of delta/theta-band synchronization was more stable. This
may be caused by more stable network configuration of synchronization or the poorer temporal
resolution of wavelets compared with the gamma band. Synchronization was confined to SM
system across time. This together with an increase in the strength of synchronization but not
a change in temporal profile in response to faster RTs suggest that delta-band synchronization
is related to evidence accumulation of sensory information and coordination of motor actions
but not the achieving conscious sensory perception per se. Overall, in line with prior stud-
ies of local neuronal processing in humans (Donner et al., 2009b; Gould et al., 2012; Kaplan
et al., 2017; Medendorp et al., 2011; Tosoni et al., 2008) our data suggest that the sensorimotor
decision process and the associated actions involve overlapping neuronal circuits also at the
large-scale network level.
Conclusions
In summary, these findings constitute evidence for the hypothesis that dynamic large-scale
network synchronization plays an essential role in the coordination of neuronal processing to
achieve conscious somatosensory perception, perceptual decisions, and initiation of a motor
response.
METHODS
An overview of the workflow is given in Figure 1. All analyses, if not stated otherwise, were
analyzed with LabVIEW software (National Instruments), available on request.
Subjects and Recordings
MEG was recorded from 12 healthy right-handed subjects (27.5 ± 4.5 years of age, mean ±
standard deviation; 7 women) with a 306 channel MEG (Elekta Neuromag, Helsinki, Finland)
Network Neuroscience
455
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
at 600-Hz sampling rate as described earlier in (Hirvonen & Palva, 2016). Electromyogram
(EMG) was recorded to detect the thumb movement responses and T1-weighted anatomical
MRI scans were obtained for source localization. The study was approved by the Coordinat-
ing Ethical Committee of the Helsinki University Hospital, and the subjects gave a written,
informed consent prior to their participation in the experiment.
Task
We used a continuous stimulus detection task in which 0.2-ms electrical stimuli were given
with an intensity (mean 4.3 ± 0.6 mA) at the threshold of detection (Hirvonen and Palva 2016;
Monto et al., 2008; Palva et al., 2005). The intensity was individually calibrated before the
recordings to a level that yielded a ∼50% detection rate and then maintained constant during
the recordings. The stimuli were delivered at uniformly random 1.5- to 4.5-s intervals to the
right index finger in two separate approximately 30-min blocks. The subject’s task was to report
with a right thumb twitch whenever he or she perceived the stimulus. On average, 1,073 ±
207 (mean ± SD, n = 12) trials were acquired per subject. No trials were rejected.
Behavioral Performance
HR was defined to be the proportion of detected stimuli of all stimuli. The stimuli were clas-
sified as consciously perceived, or “Hits,” if the subject performed the right thumb twitch
response within 0.1–1.5 s after the stimulus onset and “Misses” if stimulus was not associated
with response (Figure 2A). The responses were identified from filtered (low- and high-pass fi-
nite impulse-response filter with pass band 30–190 Hz) and absolute valued EMG recordings
using 10 baseline standard deviations as the threshold for a response and the first crossing
of this threshold as the RT (Hirvonen & Palva, 2016; Monto et al., 2008; Palva et al., 2005).
Stimuli to which no response was observed were categorized as unperceived, or “Misses.” To
estimate the false alarm (FA) rate, we estimated the number of responses between −1.5 s to 0 s
before stimulus onset, during which there should not have been any responses to somatosen-
sory stimuli. For this estimation, only the trials with a distance at least 3 s to previous and next
trial were selected. FA estimate was then computed as the proportion of these responses from
all the stimulus events (Figure 2B). Delayed responses and FA were not analyzed because of
the exceedingly small number of trials in these categories.
MEG Data Preprocessing, Filtering, and Source Analysis
The signal space separation method (tSSS) was used to remove extracranial noise from the
raw MEG recordings, and independent component analysis was used to identify and exclude
components associated with eye movements and cardiac artifacts (Figure 1, a). In short, pre-
processed MEG sensor time series were filtered using Morlet wavelets into 38 frequency bands
covering 3–120 Hz with equal distances between neighboring frequencies on the log scale
by using time-frequency compromise parameter m = 5 (Figure 1, b). We used FreeSurfer
software (http://surfer.nmr.mgh.harvard.edu/ ) and minimum-norm estimate (MNE) toolkit for
volumetric segmentation of MRI images, and reconstruction of anatomical surfaces and cor-
tical parcellation (Destrieux et al., 2010; Fisch et al., 2002) (Figure 1, c). The MNE toolkit
(http://www.nmr.mgh.harvard.edu/martinos/userInfo/data/sofMNE.php) was used to create
three-layer boundary element models, cortically constrained source models, MEG-MRI co-
localization and for preparation of the forward model and MNE inverse operators (Hamalainen
& Ilmoniemi, 1994) (Figure 1, d). The source models had dipole orientations fixed to the pial
Network Neuroscience
456
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
surface normals and a 7-mm source-to-source separation throughout the cortex, which yielded
models containing 6,000–8,000 source vertices.
We used the MNE inverse operators in the form of dynamic statistical parametric map
(dSPM) operators (Dale et al., 2000) so that the noise-covariance matrices (NCM) were ob-
tained from the baseline data 0.75–0.2 s prior to stimulus onsets and by using 0.05 as the
regularization constant. The NCMs and hence the inverse operators were prepared separately
for each Morlet-wavelet filter frequency by using lambda as 0.05. We used an atlas-based
analysis strategy (Hillebrand et al., 2012; Palva et al., 2010), in which the preprocessed and
Morlet-filtered MEG sensor time series were inverse modeled sample-by-sample into source
time series (Figure 1, d) that were then collapsed into time series of 400 cortical parcels by
using individually optimized collapse operators (Figure 1, e) (Korhonen et al., 2014). In these
collapse operators, the source vertices were weighted with parcel-signal reconstruction accu-
racy, and only the set of vertices that yielded best reconstruction accuracy were used
(Korhonen et al., 2014). For creating these weighted collapse operators, and more precisely, to
assess parcel-signal reconstruction accuracy, another set of NCM and inverse operators were
computed. To obtain NCM only for this “fidelity optimization” inverse-forward modeling,
we applied 0.1–45 Hz pass-band filtering to MEG time series by using low- and high-pass
finite impulse-response filters. The 400-parcel parcellation was obtained from a precursor
“Destrieux” atlas of 148 parcels (Destrieux et al., 2010) by iteratively splitting the largest parcels
along their most elongated axis and using the same parcel-wise splits for all subjects. Using
neuroanatomical labeling as the anatomical “coordinate system” eliminates the need for inter-
subject morphing in group-level analyses, which would have compromised individual anatom-
ical accuracy.
Forward and Inverse-Modeled Trial-Shuffled Surrogates for Evoked Component Estimation
To account for the artificial synchronization attributable to evoked responses and/or phase
locking of ongoing activity to the stimuli, we created trial-shuffled surrogate data (Figure 1, f).
To reconstruct the effects of signal mixing at MEG acquisition and inverse modeling, we ap-
plied a forward-inverse modeling–based surrogate construction approach that reconstructs the
evoked components and their spatial spread caused by the signal mixing inherent to MEG/EEG
and preserves local source topography changes, amplitude dynamics, and changes in autocor-
relation structures that may constitute significant confounders (Palva & Palva, 2012). This ap-
proach hence provides a good surrogate for selectively identifying the true induced interareal
interactions for which the conventional trial shuffling (Lachaux et al., 1999) is insufficient in
the presence of signal mixing.
We used the source-modeled single-trial data in the 400-parcel parcellation as a new parcel
time series so that first in forward modeling the activity time series of each source vertex of
a given parcel for each trial was simulated with the parcel time series of a random trial.
In
each new “trial” of the surrogate data, the cortical parcels were simulated with random-trial
time series of the original data. We then source reconstructed these sensor-level surrogate
data with procedures identical to those used for real data. This procedure thus abolishes true
non-stimulus-locked phase correlations between parcels but reconstructs both the evoked and
stimulus-phase-locked components as well as the spread of the signals caused by MEG data
acquisition and inverse modeling. Phase correlation analyses (Figure 1, g) were then performed
with these surrogate source data identically to those of real data for 10 independent realizations
of the surrogate data. The means of surrogate data were compared against the corresponding
real data in Fig. 3 (Figure 1, f).
Network Neuroscience
457
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Large-scale synchronization from perception to action
Analysis of Phase Synchrony
All-to-all phase coupling–based functional connectivity, that is, phase synchrony, between all
parcel pairs, was estimated in reconstructed source space for all 38 narrow-band time series.
We first estimated the complex-valued phase-locking value (cPLV) between all parcel pairs.
The cPLV is defined as
cPLV =
1
N ∑N
n=1
(cid:2)
ei(θX(n)−θY(n))
(cid:3)
(1)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
a
_
0
0
0
3
9
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
where N denotes sample number, θX and θY are phases of narrow-band time series of X and
Y, and I denotes the imaginary unit (Lachaux et al., 1999; Palva et al., 2005). We assess here
phase synchrony both by using the PLV, PLV = |cPLV|, and the imaginary part of cPLV (iPLV),
iPLV = |im(cPLV)|, where im indicates the imaginary value operator. PLV, a commonly used
measure for phase correlations, is essentially equally sensitive to phase coupling at all phase
lags and thus is sensitive to artificial zero-phase-lag correlations caused by MEG signal mixing
and spatial leakage in inverse modeling. iPLV, on the other hand, is insensitive to near-zero
and near-pi phase lags and thus does not yield false positives attributable exclusively to signal
mixing (Brookes et al., 2014) (Figure 1, g).
To compensate for the fact that both PLV and iPLV are biased by the number of samples, the
numbers of Hit and Miss trials were balanced within subjects before the PLV/iPLV analyses by
keeping only those events of the larger conditions, which are closest to the onset latencies of
the smallest condition. For each frequency, the cPLV values were obtained across samples in
100-ms time windows and across trials. Using 50% overlap, we obtained 26 time windows
from −725 ms to 525 ms around the stimulus.
Group Statistical Analyses
Group statistical contrasts between conditions or between prestimulus baseline and post-
stimulus periods were performed separately for each frequency, time window, and parcel pair.
Before group statistics, mean baseline values from 125 to 225 ms prior to stimulus onset were
subtracted. The significance of the difference between stimulus processing and baseline or
between Hits and Misses was estimated with the Wilcoxon signed-rank test (p < 0.05). To re-
duce false discoveries arising from multiple comparisons, we pooled significant observations
for each contrast across all cortical parcels and time windows but separately for each frequency
band. Then we discarded as many least significant observations across parcels and across time
windows as were predicted by the alpha level (α) and the total number of comparisons (Nc),
α × Nc (Figure 1, h). This false discovery reduction was done separately for each frequency
band.
Removal of Parcels and Connections with Low Reliability
The poor source reconstruction accuracy of some cortical parcels limits the overall quality of
all-to-all interaction analyses. In this study, we utilized a simulation-based method to assess
the reliability of local source time-series reconstruction and interareal interaction estimates
(Korhonen et al., 2014). We used the subject cohort’s source models, that is, forward and
inverse operators that were derived from real MEG recordings, simulated independent parcel
time series as the ground-truth data, and then simulated a virtual MEG recording and source
reconstruction by forward and inverse modeling the ground-truth time series. Thus, assessing
the correlations between simulated and reconstructed data yields quantitative estimates of the
Network Neuroscience
458
Large-scale synchronization from perception to action
reconstruction accuracy and signal mixing between parcels. We used parcel f idelity ( fu) to
quantify the source reconstruction accuracy of a parcel u; it is defined as follows:
fu = |re(cPLV(xu, ˆxu))|
(2)
where xu is the simulated original time series of parcel u, and ˆxu is the source-reconstructed
time series of xu.
We used parcel-to-parcel infidelity (iuv) to quantify the amount of mixing from parcels u to
another parcel v, and it is defined as follows:
iuv = |re(cPLV(xu, ˆxv))|
(3)
where xu is the original time series of parcel u, and ˆxv is the forward- and inverse-modeled time
series of another parcel v. Thereby iuv quantifies the amount of signal from u that is observed in
the reconstructed signal of another parcel v, thus describing degree of signal leak from parcel u
to parcel v. To decrease the probability of reporting artificial and spurious synchronization due
to poor source reconstruction accuracy, we first removed parcels with fidelity lower than 0.11.
These parcels were located mostly in deep and/or inferior parts of the cortex and are known to
generate weak signals in M/EEG (Supporting Information Figure 1, Hirvonen, Monto, Wang,
Palva, & Palva, 2018). Next, we excluded cortical parcels close to the eyes because they are
known to include oculomotor artifacts in MEG. These parcels are mostly located afar (e.g.,
>5 厘米) 来自眶额等传感器, 前部, 和下颞叶和我-
表盘结构. 总共, 20% 的此类地块被排除在分析之外. 此外, 我们
还排除了以低保真度连接任何地块的地块与地块之间的相互作用, 那是,
MEG 不太可能观察到的相互作用. 在分析数据中, 这些
因此,耦合更有可能反映来自其他信号的混合信号相互作用。
来源,从而产生误报. 合在一起, 38% 的相互作用被消除
可能会受到较差的重建精度和线性混合的污染.
(数字 1, 我).
可视化
总结我们的结果, 我们使用连接密度, K, 作为频带的函数
指示给定实验条件对比中统计显着边缘的分数
(数字 3). 这些数据用于识别感兴趣的频段 (数字 1, j). 图表
(数字 5) 然后将与这些数据相对应的可视化,以便大多数中心连接
由连接节点的平均 PageRank 中心度来确定 (鲁比诺夫 & 斯波恩斯,
2010). 用于图形可视化, 我们首先评估每条边的中心性 ei(你, v) 在里面
通过对源节点和目标节点的 PageRank 中心性求和来绘制图表:
这些 = 总和(普鲁 + PRv)
(4)
其中 PR 表示 PageRank 中心性. PageRank 是衡量以下内容重要性的指标
一个节点, 因此,Cei 衡量连接重要节点的边的权重更大
在图表中. 随后, 我们对组边缘应用了一种新颖的超边缘捆绑方法
通过信号混合中的相邻性将其分成束 (王等人。, 2018), 这是源自
前面描述的地块到地块不保真度函数 iuv (数字 1, k). 简单地说, 这种方法
将具有大信号混合的边缘捆绑在一起,因此是杂散的和人工的-
由真正的神经元连接正式诱导. 因此, 超边代表下边-
真实的相互作用及其由信号空间泄漏引起的虚假反射. 我们排除了
网络神经科学
459
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
A
_
0
0
0
3
9
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
从感知到行动的大规模同步
少于四个边的束, 因为真正的区域间同步很可能反映在
每个真实连接周围的虚假连接导致大量信号传播. 到
提高神经束的神经解剖分辨率, 我们丢弃了中心较低的边缘-
性 (< 50%) within each bundle. The starting number of edges was selected so that the final
edge number became 300 and 200 for graphs in Figure 5 so that the largest number of edges
was selected for graphs with largest K and smallest for smallest K.
To estimate interactions between the seven functional subsystems (Yeo et al., 2011), we
morphed the original 400 × 400 adjacency matrices into 7 × 7 subsystem interaction matri-
ces (Figures 6A and 7), and evaluated K of connections among each subsystem (Figure 1, l). To
test whether these K were greater than expected by chance for a random graph, we computed
5,000 randomizations of the same 400 × 400 matrices, keeping the numbers of significant
edges constant. K values of the original subsystems interaction matrix were reported as signif-
icant if they exceeded the 95th percentile of the K values in the randomized graphs.
Individual Subject Statistics
To estimate statistical significance of synchronization of the center frequency that was identi-
fied in the group-level analysis in single subjects, we used nonparametric permutation statistics
(Figure 1, m). We computed the difference in mean PLV between Hits and Misses, δI, within
each time window and for each pair of parcels. To obtain the permutation-based statistics, we
randomly redistributed the trials into Hitperm and Missperm and computed δIperm, the mean
PLV between the permuted conditions. The random permutation and PLV recomputation was
repeated for Nperm = 5,000 times. To obtain the subject-wise permutation test statistic, pperm,
we counted the portion of permutations that resulted in higher absolute difference in PLV than
the unpermuted conditions:
pperm = #(|δIperm| > |δI|)/恩佩尔姆,
(5)
在哪里 #() 表示计数操作. 然后对 ppm 统计数据进行校正以预测
错误发现率如上所述,使用 α = 0.05 罗斯福= 0.05.
致谢
我们感谢博士. Alexander Zigalov 和 MSc Santeri Rouhinen 贡献了分析和
可视化工具.
作者贡献
乔尼·赫沃宁: 数据分析; 可视化; 资金获取; 稿件写作. 盛
王: 软件设计. 西莫·蒙托: 数据采集; 软件设计; 数据分析; 维-
实际化; 稿件写作. 一帕尔瓦: 实验设计; 数据收集; 资金
收购; 稿件写作. 马蒂亚斯·帕尔瓦: 实验设计: 软件设计; 资金
收购; 稿件写作.
资金信息
这项工作得到了赫尔辛基大学研究补助金的支持, 芬兰科学院
(在 267030, 在 266402 和SA 273807 至 SP 和 SA 253130 和 256472 国会议员), 研究
赫尔辛基大学向 SP 和 JH 成立, 和 Sigrid Juselius 基金会到 SP 和
支持这项研究的议员.
网络神经科学
460
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
A
_
0
0
0
3
9
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
从感知到行动的大规模同步
参考
贝克, A. P。, 布鲁克斯, 中号. J。, 雷泽克, 我. A。, 史密斯, S. M。, 贝伦斯,
T。, 普罗伯特·史密斯, & 磷. J。, 伍尔里奇, 中号.
(2014). 快速瞬态
人脑自发活动中的网络. 亿生活, 3, e01867.
布兰肯堡, F。, 塔斯金, B., 鲁本, J。, 莫斯曼, M。, 里特尔, P。,
古玩, G。, & 狂野的召唤者, A. (2003). 难以察觉的刺激和
感觉处理障碍. 科学 (纽约, 纽约), 299,
1864.
布鲁克斯, 中号. J。, 伍尔里奇, 中号. W., & 巴恩斯, G. 右. (2012). 测量
MEG 中的功能连接: 多变量方法 insen-
对线性源泄漏敏感. 神经影像, 63, 910–920.
布鲁克斯, 中号. J。, 奥尼尔, G. C。, 大厅, 乙. L。, 伍尔里奇, 中号. W., 贝克,
A。, 宫角, S。, 罗布森, S. E., 莫里斯, PG, & 巴恩斯,
G. 右. (2014). 测量时间, 光谱和空间变化
电生理脑网络连接. 神经影像,
91, 282–299.
布莫尔, E., & 斯波恩斯, 氧. (2009). 复杂的大脑网络: 图形
结构和功能系统的理论分析. 自然
评论 神经科学, 10, 186–198.
戴尔, A. M。, 刘, A. K., 菲舍尔, 乙. R。, 巴克纳, 右. L。, 贝利沃, J. W.,
勒万, J. D ., & 哈尔格伦, 乙.
(2000). 动态统计参数-
度量映射: 结合 fMRI 和 MEG 以获得高分辨率
皮质活动成像. 神经元, 26, 55–67.
帕斯卡莱, F。, 笔下的, S。, 斯波恩斯, 奥。, 罗马尼人, G. L。,
& 科尔贝塔, 中号. (2015). 动态的核心网络和全球
人脑休息时的效率. 大脑皮层, 26,
4015–4033.
德科, G。, 托诺尼, G。, 博利, M。, & 克林格尔巴赫, 中号. L. (2015).
的贡献
重新思考隔离与融合:
全脑建模. 自然评论. 神经科学, 16,
430–439.
德阿内, S。, & 尚古克斯, J. 磷. (2011). 实验与理论-
意识加工的自然方法. 神经元, 70, 200–227.
德阿内, S。, 尚古克斯, J. P。, 纳克卡什, L。, 萨克尔, J。, & 军士, C.
(2006). 有意识的, 前意识, 和潜意识处理: A
可测试的分类法. 认知科学的趋势, 10, 204–211.
德阿内, S。, 纳克卡什, L。, 科恩, L。, 碧涵, D. L。, 曼金,
J. F。, 波琳娜, J. B., & 里维埃, D. (2001). 大脑机制
词语掩蔽和无意识重复启动. 自然
神经科学, 4, 752–758.
德尔库尔, A。, 巴耶莱特, S。, & 德阿内, S. (2007). 大脑动力学
进入意识的非线性阈值的基础.
公共科学图书馆生物学, 5, e260.
德斯特里厄, C。, 菲舍尔, B., 戴尔, A。, & 哈尔格伦, 乙. (2010). 汽车-
使用标准对人类皮质回和脑沟进行自动分割
解剖学命名法. 神经影像, 53, 1–15.
唐纳, 时间. H。, 西格尔, M。, 薯条, P。, & 恩格尔, A. K. (2009A). 积聚
人类运动皮层的选择预测活动
感性决策. 现代生物学, 19, 1581–1585.
唐纳, 时间. H。, 西格尔, M。, 薯条, P。, & 恩格尔, A. K. (2009乙). 积聚
人类运动皮层的选择预测活动
感性决策. 现代生物学, 19, 1581–1585.
恩格尔, A. K., & 薯条, 磷. (2010). Beta 波段振荡——信号传导
现状? 神经生物学的当前观点, 20, 156–165.
恩格尔, A. K., & 歌手, 瓦. (2001). 时间结合和神经
感官意识的相关性. 认知科学的趋势, 5,
16–25.
费施, L。, 私下里, E., 拉莫特, M。, 哈雷尔, M。, 尼尔, Y。, 基珀瓦瑟, S。,
安德尔曼, F。, 诺伊菲尔德, 中号. Y。, 克莱默, U。, 油炸, 我。, & 马拉奇, 右.
(2009). 神经“点火”: 与 percep 相关的增强激活-
人类腹侧流视觉皮层的双重意识. 神经元,
64, 562–574.
菲舍尔, B., 礼拜, D. H。, 所以, E., 阿尔伯特, M。, 迪特里希, M。,
哈塞尔格罗夫, C。, 范德库韦, A。, 基利亚尼, R。, 肯尼迪, D .,
克拉夫内斯, S。, 蒙蒂略, A。, 马克里斯, N。, 罗森, B., & 戴尔, A. 中号.
(2002). 全脑分割: 神经元自动标记-
人脑的解剖结构. 神经元, 33, 341–355.
弗雷, J. N。, 鲁瑙, P。, 闪闪发光, S。, 西格尔, M。, 布劳恩, C。, & 魏兹,
氮. (2016). 意识的触觉窗口的特点是
通过初级的频率特定集成和隔离
体感皮层. 科学报告, 6, 20805.
薯条, 磷.
(2015). 认知节奏: 沟通通过
连贯性. 神经元, 88, 220–235.
盖拉德, R。, 德阿内, S。, 亚当, C。, 克列孟梭, S。, 哈斯本,
D ., 鲍拉克, M。, 科恩, L。, & 纳克卡什, L. (2009). 汇聚
意识通路的颅内标记. 公共科学图书馆生物学, 7, e61.
戈阿尔, 中号. J。, 河粉, G. N。, 伍德森, J。, & 在, 中号. (2016). 清楚的
视觉的角色, 顶叶, 和记忆中的额叶运动皮层-
引导感觉运动决策. 亿生活, 5, 10.7554/埃莱夫.13764.
古尔德, 我. C。, 高贵, A. C。, 怀亚特, 五、, & 拉什沃斯, 中号. F. (2012).
决策变量和顶内刺激的影响
人脑中的感觉运动振荡活动. 杂志
神经科学, 32, 13805–13818.
格雷戈里奥, G. G。, 戈茨, S. J。, 周, H。, & 德西莫内, 右.
(2009).
通过与 atten 同步进行远程神经耦合-
的. 脑研究进展, 176, 35–45.
烧烤-斯佩克特, K., 库什尼尔, T。, 活动, T。, & 马拉奇, 右. (2000). 这
对象选择性激活的动态与识别相关-
人类的表现. 自然神经科学, 3, 837–843.
小男孩, 时间. P。, 凯尼曼斯, J. L。, & 内格斯, S. F. (2011). 抓握
准备增强方向变化检测. 公共科学图书馆一号, 6,
e17675.
黑根斯, S。, 巴斯克斯, Y。, 扎伊诺斯, A。, 阿尔瓦雷斯, M。, 詹森, 奥。, &
罗莫, 右. (2014). 振动触觉期间的丘脑皮质节律
检测任务. 美国国家科学院院刊
美利坚合众国, 111, E1797–E1805.
哈马兰, 中号. S。, & 科学, 右. J. (1994). 口译杂志-
大脑网络场: 最低标准估计. 医疗的 &
生物工程 & 计算, 32, 35–42.
栅栏, 是. L。, 林德纳, A。, & 布劳恩, C. (2016). 体感-
参与知觉决策的皮层区域的运动级联-
触觉模式辨别过程中的视觉形成. 人脑
测绘, 38, 1172–1181.
希勒布兰德, A。, 巴恩斯, G. R。, 森林树, J. L。, 贝伦德塞, H. W., &
斯塔姆, C. J. (2012). 频率相关的功能连接
在静息态网络内: 基于图集的 MEG 波束形成器
解决方案. 神经影像, 59, 3909–3921.
网络神经科学
461
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
A
_
0
0
0
3
9
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
从感知到行动的大规模同步
希普, J. F。, 恩格尔, A. K., & 西格尔, 中号.
(2011). 振荡同步-
大规模皮质网络的时间化预测感知.
神经元, 69, 387–396.
希尔沃宁, J。, 数量, S。, 王, S. H。, 崇拜, J. M。, & 崇拜, S. (2018).
“动态大规模网络同步”支持信息-
从感知到行动的同步化。”网络神经科学,
2, 442–463. https://doi.org/10.1162/netn_a_00039
希尔沃宁, J。, & 崇拜, S. (2016). 相位和皮层定位
振幅动态预测体感意识的获取-
内斯. 人脑图谱, 37, 311–326.
琼斯, S. R。, 普里切特, D. L。, 施弗尔比姆, S. M。, 哈马兰,
M。, & 摩尔, C. 我. (2007). 触觉检测的神经相关性:
脑磁图和生物物理学相结合
计算模型研究. 神经科学杂志, 27,
10751–10764.
琼斯, S. R。, 普里切特, D. L。, 山雀, 中号. A。, 施弗尔比姆, S. M。,
(2009). 定量分析
哈马兰, M。, & 摩尔, C. 我.
以及 MEG μ 节律的生物物理现实神经模型:
感觉诱发反应的节律发生和调节.
神经生理学杂志, 102, 3554–3572.
卡普兰, R。, 国王, J。, 科斯特, R。, 一分钱, 瓦. D ., 伯吉斯, N。, &
弗里斯顿, K. J. (2017). 预期的神经表征
空间规划和决策过程中的选择. 公共科学图书馆生物学, 15,
e1002588.
国王, J. R。, 小鱼, N。, & 德阿内, S. (2016). 大脑机制
可见和不可见的感官的短暂维持的基础
信息. 神经元, 92, 1122–1134.
柯尼格, P。, 恩格尔, A. K., 罗尔夫塞玛, 磷. R。, & 歌手, 瓦. (1995).
神经元同步有多精确. 神经计算,
7, 469–485.
科尔霍宁, 奥。, 崇拜, S。, & 崇拜, J. 中号. (2014). 稀疏权重
用于折叠皮质分区的逆解优化
M/EEG 源重建精度. 神经科学杂志
方法, 226C, 147–160.
拉绍, J. P。, 罗德里格斯, E., 马蒂内里, J。, & 瓦雷拉, F. J. (1999).
测量大脑信号的相位同步. 人脑图谱-
平, 8, 194–208.
李海格纳, Y。, 林德纳, A。, & 布劳恩, C. (2015). 皮质关联
触觉空间模式识别过程中感知决策的影响-
定罪. 人脑图谱, 36, 3339–3350.
马里斯, E., 薯条, P。, & 来自 埃德, F.
(2016). 不同的相位关系
神经元节律及其潜在功能. 趋势
神经科学, 39, 86–99.
梅登多普, 瓦. P。, 布赫霍尔茨, V. N。, 范德沃夫, J。, & 莱昂内,
(2011). 目标导向行为中的顶额环路.
F. 时间.
欧洲神经科学杂志, 33, 2017–2027.
数量, S。, 崇拜, S。, 沃皮奥, J。, & 崇拜, J. 中号. (2008). 非常慢
脑电图波动预测刺激检测的动态
人体的振荡幅度. 神经科学杂志, 28,
8268–8272.
村上, S。, & 冈田, 是. (2006). 校长的贡献
新皮质神经元脑磁图和电图-
脑电图信号. 生理学杂志, 575, 925–936.
尼尔豪斯, T。, 福尔沙克, N。, 派珀, S. K., 霍尔茨, S。, 克劳斯, T。,
塔斯金, B., 长的, X。, 施特尔策, J。, 马古利斯, D. S。, 斯坦布林克,
J。, & 狂野的召唤者, A. (2015). 难以察觉的体感刺激-
关系改变感觉运动背景节律和连通性.
神经科学杂志, 35, 5917–5925.
诺德马克, 磷. F。, 普鲁申斯基, J. A。, & 约翰逊, 右. S. (2012). 大胆的
视觉和听觉皮层对触觉刺激的反应取决于
关于刺激的频率内容. 认知杂志
神经科学, 24, 2120–2134.
奥尔洛夫, T。, 马金, 时间. R。, & 佐哈拉斯, 乙. (2010). 地形代表-
人体枕颞皮质的变化. 神经元,
68, 586–600.
崇拜, J. M。, 崇拜, S。, & 卡维拉, K. (2005). neu之间的相位同步-
人类皮层的罗纳尔振荡. 神经科学杂志,
25, 3962–3972.
崇拜, J. M。, 数量, S。, 库拉谢卡尔, S。, & 崇拜, S. (2010). 新-
罗纳尔同步揭示工作记忆网络并预测
个人记忆容量. 会议记录
全国
美国科学院, 107,
7580–7585.
崇拜, S。, & 崇拜, J. 中号. (2012). 发现振荡相互作用
M/EEG 网络: 挑战与突破. 趋势
认知科学, 16, 219–230.
崇拜, S。, 林肯卡尔-汉森, K., 纳塔宁, R。, & 崇拜, J. 中号. (2010).
神经元同步揭示了工作记忆网络和
预测个人记忆容量. 国家会议录
美国科学院, 107,
7580–7585.
帕帕德利斯, C。, 阿费勒, C。, 埃拉, S。, 我不想, G。, 卡塔内奥, L。, & 布劳恩,
C. (2016). 额下回连接视觉和运动皮质
在视觉运动精确握力任务期间. 脑研究,
1650, 252–266.
彼得森, S。, & 斯波恩斯, 氧.
(2015). 大脑网络和认知
架构. 神经元, 88, 207–219.
力量, J. D ., 科恩, A. L。, 纳尔逊, S. M。, 假发, G. S。, 巴恩斯,
K. A。, 教会, J. A。, 沃格尔, A. C。, 劳曼, 时间. 奥。, 米津, F. M。,
施拉加尔, 乙. L。, 彼得森, S. 乙.
(2011). 功能网络
人脑的组织. 神经元, 72, 665–678.
鲁比诺夫, M。, & 斯波恩斯, 氧. (2010). 复杂的网络措施
大脑连接: 用途和解释. 神经影像, 52,
1059–1069.
萨达吉亚尼, S。, 赫塞尔曼, G。, & 克莱因施密特, A. (2009). 迪斯-
正在进行的活动变化的贡献和对抗贡献-
对听觉刺激检测的影响. 神经科学杂志,
29, 13410–13417.
盐, M。, 数量, S。, 查尔斯, L。, 国王, J. R。, 帕克科宁, L。, & 德阿内,
(2015). 独特的皮质代码和时间动态
S.
用于有意识和无意识的感知. 亿生活, 4, 10.7554/
电子生活.05652.
沙尔, J. D. (2001). 决策的神经基础, 选择和行动.
自然评论神经科学, 2, 33–42.
锄, J. M。, & 总的, J. (2009). 源连通性分析
脑磁图和脑电图. 人脑图谱, 30, 1857–1865.
施罗德, C. E., & 锁匠, 磷.
(2009). 低频神经元
振荡作为感官选择的工具. 神经的趋势-
科学, 32, 9–18.
西格尔, M。, 布施曼, 时间. J。, & 磨坊主, 乙. K. (2015). 皮质信息-
灵活感觉运动决策期间的化流. 科学, 348,
1352–1355.
网络神经科学
462
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
A
_
0
0
0
3
9
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
从感知到行动的大规模同步
西格尔, M。, 唐纳, 时间. H。, & 恩格尔, A. K. (2012). 光谱手指-
互动. 自然评论
大尺度神经元
的印刷品
神经科学, 13, 121–134.
歌手, 瓦. (2009). 分布式处理和时间代码
神经元网络. 认知神经动力学, 3, 189–196.
歌手, 瓦.
(1999). 神经元同步性: 通用代码
关系的定义? 神经元, 24, 49–65, 111–25.
巨剑, S。, 笔下的, S。, 塞斯捷里, C。, 贝蒂, 五、, 托索尼, A。,
佩鲁奇, 中号. G。, 罗马尼人, G. L。, & 科尔贝塔, 中号. (2015). 动态的
visuo期间人类静息态网络的重组-
空间注意力. 美国国家科学院院刊-
美利坚合众国, 112, 8112–8117.
塔隆-鲍德里, C.
(2012). 论辅助的神经机制
意识和注意力. 心理学前沿, 2, 397.
塔斯金, B., 霍尔茨, S。, 克劳斯, T。, & 狂野的召唤者, A. (2008). 抑制药-
手指潜意识电刺激对 SI 的历史影响
相邻手指的表征和感知灵敏度.
神经影像, 39, 1307–1313.
陶斯特·坎波, A。, 马丁内斯-加西亚, M。, 纳彻, 五、, 露娜, R。,
罗莫, R。, & 德科, G. (2015). 任务驱动的内部- 和区域间
灵长类动物大脑皮层的通讯. 诉讼程序
美国国家科学院,
112, 4761–4766.
托马西尼, A。, 斯皮内利, D ., 雅科诺, M。, 桑迪尼, G。, & 莫罗内,
中号. C. (2015). 视觉对比敏感度的节律性振荡
与动作同步. 神经科学杂志, 35, 7019–7029.
托索尼, A。, 加拉塔, G。, 罗马尼人, G. L。, & 科尔贝塔, 中号. (2008).
人类顶叶皮层的感觉运动机制
任意的视觉决定. 自然神经科学, 11, 1446–1453.
范加尔, S。, 里德林克霍夫, K. R。, 继续行驶, J. J。, 斯科尔特, H. S。,
& 瘫痪, V. A. (2008). 额叶皮层无意识地调节
神经科学杂志, 28,
触发抑制控制.
8053–8062.
王, S。, 洛比尔, L。, 七只母鸡, F。, 崇拜, S。, & 崇拜, J. 中号. (2018).
超边缘捆绑: 杂散交互的实用解决方案-
MEG/EEG 源连通性分析中的系统蒸发散. 173, 610–622.
魏兹, N。, 你很美丽, A。, 莫尼托拉, G。, 德马奇, G。, 弗雷, J。, 波波夫,
T。, & 布劳恩, C. (2014). 刺激前振荡功率和缺点-
活动模式倾向于有意识的体感感知-
的. 美国国家科学院院刊
美国, 111, 4: E417–E425.
沃尔施拉格, A. (2000). 隐形交流引发视觉运动-
系统蒸发散. 视觉研究, 40, 925–930.
哪个, T。, & 沙德伦, 中号. 氮. (2007). neu 的概率推理-
罗恩. 自然, 447, 1075–1080.
杨, 乙. T。, 克里宁, F. M。, 墓, J。, 肥皂, 中号. R。, 军队, D .,
霍林斯黑德, M。, 罗夫曼, J. L。, 斯莫勒, J. W., 佐莱伊, L。, 波利梅尼,
(2011). 这
J. R。, 菲舍尔, B., 刘, H。, & 巴克纳, 右. L.
通过内因估计人类大脑皮层的组织-
sic功能连接. 神经生理学杂志, 106,
1125–1165.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
2
4
4
4
2
1
0
9
2
2
4
9
n
e
n
_
A
_
0
0
0
3
9
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
网络神经科学
463