研究
The arrow of time of brain signals in cognition:
Potential intriguing role of parts of the
default mode network
Gustavo Deco1,2,3,4, Yonatan Sanz Perl1,5, Laura de la Fuente5, Jacobo D. Sitt6,
乙. 时间. Thomas Yeo7, Enzo Tagliazucchi5,8, and Morten L. Kringelbach9,10,11
1Center for Brain and Cognition, Computational Neuroscience Group, Department of Information and
Communication Technologies, Universitat Pompeu Fabra, 巴塞罗那, 西班牙
2Institució Catalana de la Recerca i Estudis Avançats (ICREA), 巴塞罗那, 西班牙
3神经心理学系, 马克斯·普朗克人类认知和脑科学研究所, 莱比锡, 德国
4School of Psychological Sciences, Monash University, 墨尔本, Clayton VIC, 澳大利亚
5Department of Physics, University of Buenos Aires, Buenos Aires, 阿根廷
6Sorbonne Université, Institut du Cerveau – Paris Brain Institute – ICM, Inserm, 法国国家科学研究中心, APHP,
Hôpital de la Pitié Salpêtrière, 巴黎, 法国
7Centre for Sleep & 认识, Centre for Translational MR Research, 电气与计算机工程系,
N.1. Institute for Health and Institute for Digital Medicine, National University of Singapore, 新加坡
8Latin American Brain Health Institute (BrainLat), Universidad Adolfo Ibanez, 圣地亚哥, 智利
9Centre for Eudaimonia and Human Flourishing, Linacre College, 牛津大学, 牛津, 英国
10Department of Psychiatry, 牛津大学, 牛津, 英国
11Center for Music in the Brain, Department of Clinical Medicine, Aarhus University, Aarhus, 丹麦
关键词: Thermodynamics, Neuroimaging, Brain, Default mode network, Orchestration
抽象的
A promising idea in human cognitive neuroscience is that the default mode network (DMN) 是
responsible for coordinating the recruitment and scheduling of networks for computing and
solving task-specific cognitive problems. This is supported by evidence showing that the physical
and functional distance of DMN regions is maximally removed from sensorimotor regions
containing environment-driven neural activity directly linked to perception and action, 哪个
would allow the DMN to orchestrate complex cognition from the top of the hierarchy. 然而,
discovering the functional hierarchy of brain dynamics requires finding the best way to measure
interactions between brain regions. In contrast to previous methods measuring the hierarchical
flow of information using, 例如, transfer entropy, here we used a thermodynamics-
inspired, deep learning based Temporal Evolution NETwork (TENET) framework to assess the
asymmetry in the flow of events, ‘arrow of time’, in human brain signals. This provides an
alternative way of quantifying hierarchy, given that the arrow of time measures the directionality
of information flow that leads to a breaking of the balance of the underlying hierarchy. 反过来, 这
arrow of time is a measure of nonreversibility and thus nonequilibrium in brain dynamics. 什么时候
applied to large-scale Human Connectome Project (HCP) neuroimaging data from close to a
thousand participants, the TENET framework suggests that the DMN plays a significant role in
orchestrating the hierarchy, 那是, levels of nonreversibility, which changes between the resting
state and when performing seven different cognitive tasks. 此外, this quantification of the
hierarchy of the resting state is significantly different in health compared to neuropsychiatric
disorders. 全面的, the present thermodynamics-based machine-learning framework provides
vital new insights into the fundamental tenets of brain dynamics for orchestrating the interactions
between cognition and brain in complex environments.
开放访问
杂志
引文: 德科, G。, Sanz Perl, Y。, de la
Fuente, L。, Sitt, J. D ., 杨, 乙. 时间. T。,
Tagliazucchi, E., & 克林格尔巴赫, 中号. L.
(2023). The arrow of time of brain
signals in cognition: Potential
intriguing role of parts of the default
mode network. 网络神经科学,
7(3), 966–998. https://doi.org/10.1162
/netn_a_00300
DOI:
https://doi.org/10.1162/netn_a_00300
支持信息:
https://doi.org/10.1162/netn_a_00300
已收到: 22 六月 2022
公认: 14 十二月 2022
利益争夺: 作者有
声明不存在竞争利益
存在.
Corresponding Authors:
Gustavo Deco
gustavo.deco@upf.edu
Morten L. 克林格尔巴赫
morten.kringelbach@psych.ox.ac.uk
处理编辑器:
Martijn van den Heuvel
版权: © 2023
麻省理工学院
在知识共享下发布
归因 4.0 国际的
(抄送 4.0) 执照
麻省理工学院出版社
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
The arrow of time of brain signals in cognition
作者总结
Here we used a thermodynamics-inspired, deep learning based Temporal Evolution NETwork
(TENET) framework to assess the asymmetry in the flow of events, ‘arrow of time’, in human
brain signals. This was applied to large-scale HCP neuroimaging data which showed
significant changes between the hierarchy of orchestration for the resting state and seven
different cognitive tasks. 相似地, the hierarchy of the resting state is significantly different in
health compared to neuropsychiatric disorders. This framework provided new insights into the
orchestrating of brain dynamics in different brain states.
介绍
A major aim of cognitive neuroscience is to discover the physical underpinnings of cognition
and behaviour. Early studies used recordings and lesions in animal models (LeDoux, 2000;
穆雷 & Rudebeck, 2018), while human research has been mostly constrained to study
the consequences of neurological disorders leading to relatively precise cognitive and emo-
tional deficits (Adolphs, 2016). The invention of human neuroimaging started an avalanche of
studies monitoring the changes in brain activity during cognitive tasks (Poldrack, 2012), 哪个
led to a better understanding of the processing in sensorimotor regions and during tasks. 在
添加, these studies curiously also led to the discovery of a network of regions deactivated
during task (Raichle et al., 2001; Shulman et al., 1997), which came to be known as the default
mode network (DMN) and which includes the precuneus/posteromedial cortex (PMC) 和
angular gyrus, regions of the inferior frontal gyrus, the medial prefrontal cortex (MPFC) 和
the anterolateral middle temporal cortex. Paradoxically, despite the DMN’s apparent deacti-
vation during task, subsequent careful studies of the DMN have instead led to this network
becoming a leading candidate for the orchestration of cognition during task (Raichle, 2006;
Raichle et al., 2001). According to this view, the DMN is responsible for coordinating the
recruitment and scheduling of networks computing and solving the task-specific cognitive
问题 (Smallwood et al., 2021). 换句话说, rather than the DMN being deactivated
during tasks, findings from recent studies have suggested that in certain task contexts the
DMN can activate, 例如, during memory guided decision-making (Murphy et al.,
2018, 2019). Note that this view is not incompatible with a static deactivation of the DMN
during tasks over longer time periods (Raichle et al., 2001; Shulman et al., 1997) but could
reflect a more dynamic view of how the DMN is stable across tasks and therefore able to
orchestrate activity (Smallwood et al., 2021).
重要的, Margulies and colleagues have demonstrated that the physical and functional
distance of the regions of the DMN are maximally removed from sensorimotor regions contain-
ing extrinsically driven neural activity directly linked to perception and action (Margulies et al.,
2016). This would make sense in terms of an evolutionary drive for more complex behaviour,
more decoupled from the here-and-now and able to make long-term predictions assuring
survival. 像这样, regions furthest away from externally driven regions would be able to take
on roles that are both more complex and less directly influenced by the external environment,
allowing for the orchestration of more complex behaviour (巴克纳 & 克里宁, 2013).
Hierarchy:
Here defined as the asymmetrical
relationship between forward and
backward interactions between brain
地区.
Despite these important findings and hypotheses, we are still missing a better understanding
and quantification of how the functional hierarchy changes when we engage in tasks compared
to the resting state, both of which require cognition but of different kinds. Hierarchy can be
defined from the causal interactions between different brain regions, which changes in tasks
according to the required computations. This in turn changes the direction of information flow,
网络神经科学
967
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
The arrow of time of brain signals in cognition
which in effect means that the asymmetry in the underlying causal interactions change. Prom-
ising research has used a plethora of ways to directly quantify the hierarchy through determining
the underlying causal interactions between brain regions, with methods ranging from Granger
causality (Seth et al., 2013), transfer entropy (Brovelli et al., 2015; Deco et al., 2021乙), 和
dynamic causal modelling (Frässle et al., 2017; Friston et al., 2003; Prando et al., 2020).
这里, we propose to use an alternative thermodynamics-inspired approach to determine
the differences in hierarchical organisation in resting state and seven tasks. 换句话说, 这
method allows for the quantification of the hierarchy defined as the asymmetrical relationship
between forward and backward interactions between brain regions.
The key idea is to be able to assess the asymmetry in the flow of events in human brain
signals. In thermodynamics this is called ‘arrow of time’ and is a direct measure of hierarchy
since this directly provides the directionality of information flow, or ‘breaking the detailed
balance’ as this is known in physics and systems biology. 这样, a flat hierarchy is char-
acterised by a low level of breaking the detailed balance, since the information flow is mostly
symmetrical. When breaking the directionality of information flow, 那是, when breaking the
detailed balance, this results in a high level of hierarchical organisation.
重要的, such processes happen at every level in biology, where all living systems must
break detailed balance to survive. At a general level, the process of breaking the detailed
balance is achieved by consuming energy and producing entropy through a whole host of
molecular and cellular functions, including sensing, adaptation, and transportation (林恩
等人。, 2021). It is important to realise the difference between entropy as a measure of disorder,
那是, the variability of the states of a system, and the concept of production entropy, 哪个
directly measures the asymmetry in time of the evolution of the states in a nonequilibrium
系统. The latter is well suited to elucidate the differences in hierarchical organisation of
different systems given that it quantifies the level of nonreversibility.
进一步来说, our new framework estimates hierarchical organisation, not using the
production entropy but using a direct way of measuring the ‘arrow of time’, central to thermo-
dynamics in physics, which was originally popularised by Arthur Eddington (Eddington, 1928)
and since studied in great detail in a number of fields (Crooks, 1998; 冯 & Crooks, 2008;
Jarzynski, 2011; Maragakis et al., 2008; Seif et al., 2021; Shirts et al., 2003). In the context of
神经科学, there has been considerable interest in using production entropy and related
concepts to characterise the time reversibility of brain signals (Deco et al., 2022; 林恩
等人。, 2021; Palus, 1996; Sanz Perl et al., 2021; Zanin et al., 2019). 然而, there are sig-
nificant statistical problems arising when the fluctuations are high, which makes it difficult to
determine the direction of the arrow of time. 这里, we applied the excellent idea of turning the
quantification of the direction of arrow of time into a problem of statistical inference for a
physical system (Crooks, 1998; 冯 & Crooks, 2008; Jarzynski, 2011; Maragakis et al.,
2008; Seif et al., 2021; Shirts et al., 2003). Among others, Seif and colleagues demonstrated
that deep learning can be used to measure the arrow of time in forward and time-reversed time
系列, compare the two, and provide a quantitatively measure of the reversibility of signals
(Seif et al., 2021). They were able to show that deep learning is able to capture time’s arrow
in relatively simple physical systems where the ground truth is known.
Here we use a deep learning for human brain signals using a Temporal Evolution NETwork
(TENET) framework to discover the asymmetry in the flow of events, 那是, arrow of time. 这
face validity of this approach has been demonstrated in the pioneering work by Seif, Hafezi,
and Jarzynski using two model physical examples in nonequilibrium (Seif et al., 2021). Please
note that recent progress in thermodynamics has allowed for the study describing the
Thermodynamics:
A branch of physics dealing with the
relationship between heat and other
forms of energy, such as work. 这里
used to measure the transfer of
energy/information from one brain
region to another, and the ways in
which this transfer affects the brain.
Detailed balance:
In thermodynamics, a concept
describing the equilibrium state of a
system in which the forward and
reverse reactions are occurring at the
same rate.
Production entropy:
A measure of the uncertainty or
unpredictability of a process. 这
amount of production entropy
produced in any nonreversible
processes directly measures the
asymmetry in time of the evolution of
states in a nonequilibrium system.
Nonequilibrium:
A system out of equilibrium.
Nonreversibility (or irreversibility):
The asymmetry in the flow of events,
also known as the ‘arrow of time’.
深度学习:
A broad class of machine-learning
methods based on artificial neural
networks able to learn from data in a
hierarchical manner using multiple
layers of interconnected nodes to
extract increasingly complex features
of the data at each layer.
网络神经科学
968
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
The arrow of time of brain signals in cognition
dynamics of open systems driven out of equilibrium rather than merely isolated systems
(Jarzynski, 2011; Seif et al., 2021).
这里, we wanted to identify the functional hierarchy of the brain at rest and during tasks.
We achieved this by using the TENET framework to assess the level of nonreversibility (箭
时间的) in brain dynamics during the resting state and seven different tasks from the large
Human Connectome Project (HCP) neuroimaging fMRI dataset of 990 healthy human partic-
爱普茨. TENET was trained on the data from 890 参与者, and the results were generated
from a generalisation set of data from the remaining 100 randomly selected participants. 这
相同的 100 participants were used for all comparisons across conditions.
Given the importance of hierarchical organisation for the successful orchestration of a given
brain state, we wanted to discover how the functional hierarchy changes in neuropsychiatric
紊乱. We therefore used exactly the same approach to study time’s arrow in health and
disease in a UCLA dataset of 261 neuropsychiatric patients (ADHD, schizophrenia, and bipo-
lar disorder as well as controls). 这里, again TENET was trained on 90% 数据的, 和
results were generated using a generalisation set of data from the remaining 10% 数据的.
The results showed that in healthy participants, the whole-brain levels of nonreversibility
are at higher levels during task than when resting. 有趣的是, in neuropsychiatric disorders,
the brain is less hierarchical with lower levels of nonreversibility during resting state than in
healthy individuals, suggestive of less specific computation. In disease, brain processing is less
efficient, which is reflected in a lower level of asymmetry of interactions between brain
地区, resulting in a different hierarchical organisation leading to the lower levels of nonre-
versibility. In healthy participants, we also found significant differences at the system level and
regional levels between resting state and the different tasks. Most importantly, consistent with
other compelling anatomical and functional neuroimaging findings, we were able to demon-
strate that across the seven tasks, DMN contains the most endogenous regions in terms of
stability across conditions. This suggests that key DMN regions can be found at the top of
the brain hierarchy, providing some support for the hypothesis that the DMN is involved in
orchestrating cognition. 全面的, beyond identifying the potential role of the DMN, TENET
provides a general, convenient framework for assessing the functional hierarchy in any given
brain state.
结果
Brain hierarchy can be defined as the asymmetrical relationship between forward and back-
ward interactions between brain regions. TENET uses the thermodynamic concept of ‘arrow of
time’ (also known as nonreversibility, or irreversibility) to reveal the hierarchical brain organi-
sation in different brain states. In the language of thermodynamics, this estimates the ‘breaking
of the detailed balance’ in the time series across the whole brain. The second law of thermo-
dynamics states that production entropy increases over time, including in an open nonequi-
librium system, such as the brain. The Clausius inequality of classical thermodynamics
predicts that the external work performed on the system will be no less than the free energy
difference between the terminal states (Jarzynski, 2011). This inequality precisely links produc-
tion entropy and nonreversibility, establishing the arrow of time in nonequilibrium systems
(Jarzynski, 2011; Seif et al., 2021). This has been extensively used for problems related to
thermodynamics of system in nonequilibrium including biological problems such as protein
folding (Collin et al., 2005). 的确, the production entropy is a measure of nonreversibility
and arrow of time, when defined as the Kullback–Leibler distance HP =
我,j Pijlog(Pij /Pji),
where Pij is the probability of transition between states i at time t to j at time t + 1. 其他
磷
969
Clausius inequality:
The mathematical relationship
describing the relationship between
heat and work in a thermodynamic
system and stating that the change in
entropy of a system is always greater
than or equal to the amount of heat
transferred to the system, divided by
the temperature at which the heat
transfer occurs.
Free energy:
A thermodynamic concept, 还
known as Gibb’s free energy, used to
describe the amount of energy in a
system that is available to do work.
网络神经科学
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
The arrow of time of brain signals in cognition
字, production entropy is directly measuring the difference between forward and backward
evolution of states over time.
这里, rather than directly estimating the production entropy, which as mentioned earlier is
very difficult, we created a deep learning based Temporal Evolution NETwork (TENET) frame-
work to discover the asymmetry in the flow of events, 那是, ‘arrow of time’, in human brain
signals. TENET was used in different brain states in health and disease to provide a quantifi-
cation of the role of the DMN in orchestrating cognition.
数字 1 和图 2 provide a schematised version of the general TENET paradigm used
这里. The key concept of the arrow of time in nonequilibrium systems is demonstrated in
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
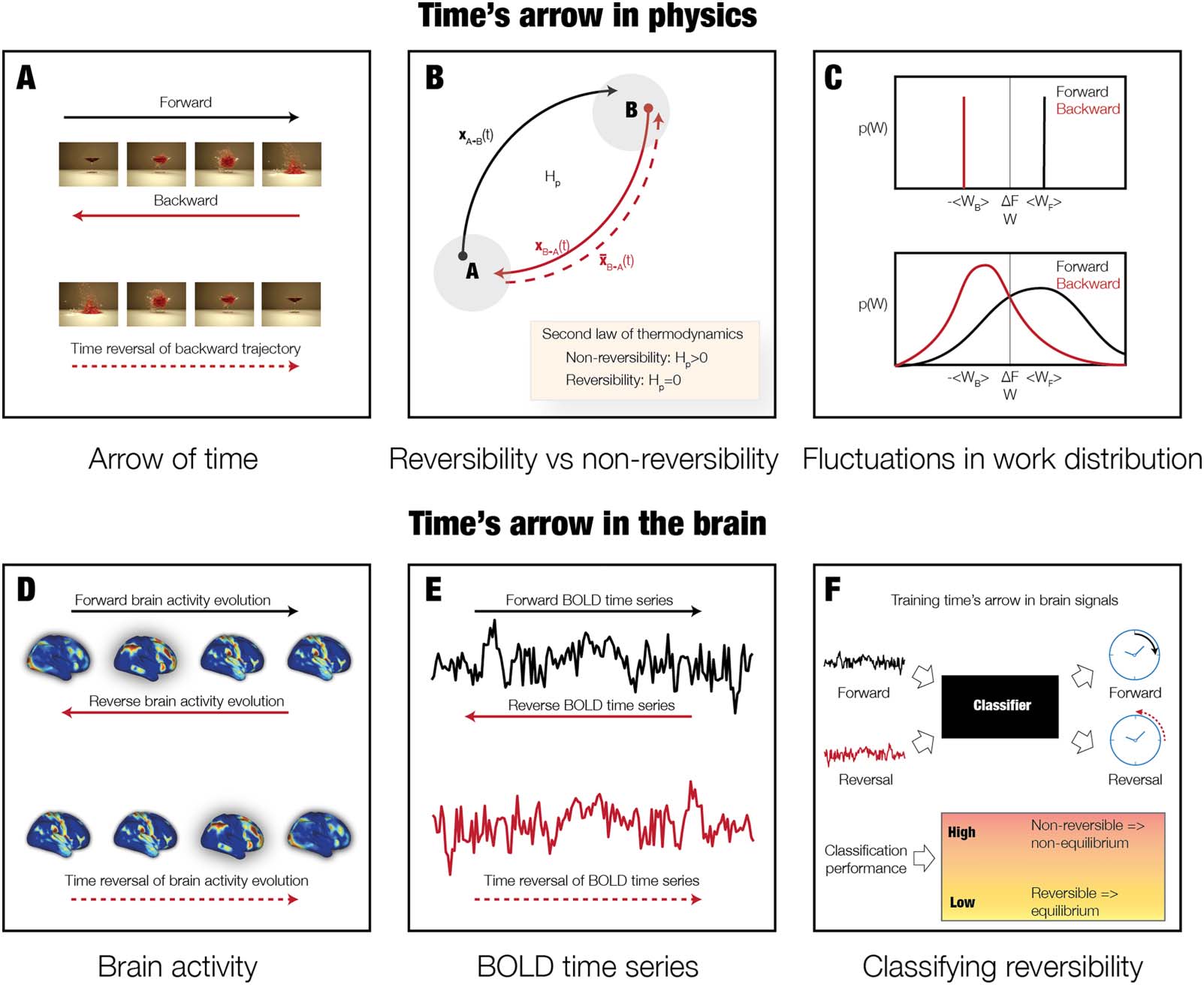
数字 1. The arrow of time in physics and brain dynamics. (A) The sequence of the four top images shows a glass being shattered by a bullet,
and we clearly perceive the causal passage of time, also called the arrow of time. 相比之下, this cause and effect is shattered by showing
these images backward—by time reversing the backward evolution. This means that this process is nonreversible. (乙) In thermodynamics,
nonreversibility can be associated with the production of entropy. The figure shows a nonequilibrium system with two states A and B and
the associated trajectories evolving during forward (A → B, black arrow) and backward (B → A, red arrow) 流程. Both the forward and
backward trajectories can be depicted as the movie shown in the top of panel A, but with a different arrow of time. 相比之下, the time reversal
of the backward trajectory (red stippled arrow) can be imagined as the movie of the backward trajectory that is played forward in time (看
bottom of panel A). If the forward and time reversal of the backward trajectories are different, this corresponds to nonreversibility of the
过程. The second law of thermodynamics uses the entropy production to describe this. If the entropy production is larger than zero, 这
corresponds to nonreversibility of a nonequilibrium system. 相比之下, if there is no entropy production, this is a reversible, equilibrium
系统. (C) 进一步来说, when small systems undergo thermodynamic processes, the fluctuations are nonnegligible and the second
law of thermodynamics expresses this in terms of averages. (D) 第一的, we used large-scale empirical whole-brain neuroimaging data from over
1,000 participants when resting and performing seven different tasks. (乙) From this data, we were able to extract the forward time series as well
as constructing the time reversal of the backward time series for a given parcellation. (F) This procedure provides a clear arrow of time for a
given time series and allows us to train a classifier to identify the forward and the time reversal for a given time series of any length. 这
classification performance provides a measure of the degree of nonreversibility and nonequilibrium.
网络神经科学
970
The arrow of time of brain signals in cognition
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
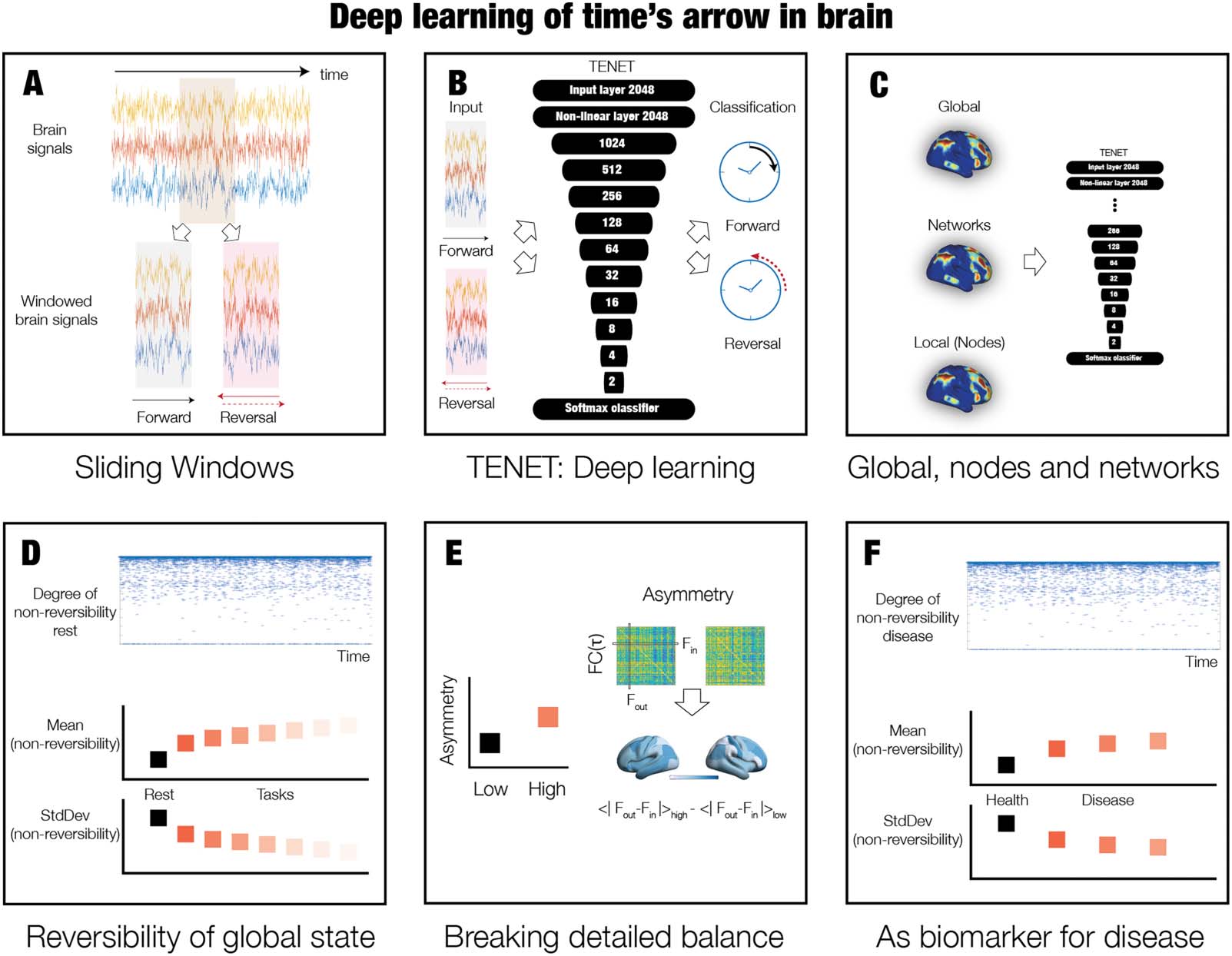
数字 2. Deep learning the arrow of time in health and disease. In order to discover the arrow of time in brain dynamics in health and
疾病, we designed a deep-learning pipeline named Temporal Evolution NETwork (TENET). (A) 具体来说, we used sliding windows of
brain signal time series from all brain regions in all participants. (乙) These sliding windows were then used in the TENET, a deep-learning
network classifier with 13 layers for classifying the arrow of time. (C) This strategy allowed us to study nonreversibility and nonequilibrium at
different levels of granularity, from global (all signals) to system level to individual node-level signals. (D) After training, TENET was able to
characterise the degree of reversibility, 那是, nonreversibility for each sliding window (top panel). We performed this procedure on data
resting and seven tasks and computed the means of the levels of certainty of the classifier output (across time) as a measure of the degree
of nonreversibility (middle panel). The standard deviation of this measure establishes stability of this nonreversibility across time. 鉴于
nonequilibrium states are already nonstationary, this provides the second order of nonstationarity (参见方法). (乙) Nonequilibrium is asso-
ciated with the breaking of detailed balance of a system. We estimated this by selecting windows of low and high reversibility, and computing
the FC(t), 那是, the time-delayed functional connectivity between all pairs of brain regions. 具体来说, the degree of asymmetry of the FC(t)
matrix is a proxy for the breaking of the detailed balance with more asymmetry corresponding to more unbalance. The level of asymmetry can
also be rendered on the brain (参见方法). (F) 最后, we used TENET on resting-state data from neuropsychiatric patients with diagnoses of
schizophrenia, ADHD, and bipolar disorder, as well as age-matched controls. Different levels of nonreversibility provide a potential biomarker
of neuropsychiatric disease.
Figure 1A, which shows four sequential images from a film of a glass being shattered by a
bullet. 以下, the same four images are shown in a sequence in an opposite direction, 那
是, in time reversal of the backward trajectory of the film. When comparing the two films, 这
arrow of time is very clear, which is the signature of a nonreversible physical process produc-
ing entropy in nonequilibrium. More general, as shown in Figure 1B, the field of thermody-
namics in physics can be used to describe such processes associated with the production of
entropy and consequently with nonequilibrium/nonreversibility. The figure shows the evolu-
tion over time of a nonequilibrium system with two states A and B and their associated trajec-
保守党. The forward and backward trajectories of the movies in Figure 1A are described as
向前 (A → B, black arrow) and backward (B → A, red arrow) 流程. The time reversal
of the backward trajectory (red stippled arrow) can be thought of as the movie of the backward
trajectory that is played forward in time (see bottom of Figure 1A). A nonreversible process
网络神经科学
971
The arrow of time of brain signals in cognition
机器学习:
A subset of artificial intelligence
involving the use of algorithms and
statistical models to enable a system
to improve its performance on a
specific task over time.
results from the ability to differentiate between the trajectories in time described by the forward
(black arrow) and time reversal (stippled red arrow). The second law of thermodynamics
(usually attributed to Rudolph Clausius and Sadi Carnot) states that if the entropy production
is larger than zero, this corresponds to nonreversibility of a nonequilibrium system. 相比之下,
if there is no entropy production, this describes a reversible, equilibrium system.
In thermodynamics, the Clausius inequality establishes that the work W associated with the
过程 (averaged over many repetitions) is larger than the change in its free energy ΔF.
Figure 1C shows distributions of the work p( 瓦) for the average of the work associated with
the forward and backward trajectories, denoted
ible macroscopic processes (like the movie shown in Figure 1A) fluctuations are negligible and
the distinction is clear between the distribution of work (top of panel) and therefore the arrow
of time is easy to establish. 相比之下, in microscopic systems (which includes brain signals)
the average work is similar, but the fluctuations are more pronounced and therefore the dif-
ferences in distribution less clear. In such cases it is much harder to establish the arrow of time,
and thus establish whether a system is nonequilibrium and nonreversible.
This uncertainty is a perfect case for which to use advanced machine-learning techniques
(Sejnowski, 2018). 这里, we used deep learning in empirical brain-imaging data to detect the
reversibility of the system. Figure 1D and Figure 1E illustrate how we used whole-brain activity
from large-scale empirical whole-brain neuroimaging data from over 1,000 participants to
construct the forward and time-reversed time series needed to establish the arrow of time
and hence nonequilibrium by detecting the level of nonreversibility. 具体来说, Figure 1F
illustrates how we extracted the forward time series as well as constructing the time reversal
of the backward time series for the DK80 parcellation (参见方法). The forward and reverse
time series were used to train a classifier to predict whether a given time series is forward or
reversed in time. If the classification performance is high, this provides evidence for nonrever-
sibility and nonequilibrium, while low performance implies the opposite. We hypothesized
that brain regions at the bottom of the functional hierarchy will exhibit greater nonreversibility
given that these regions will be driven to nonequilibrium. 所以, this approach can be
used to identify the functional hierarchy of the human brain.
数字 2 specifies the full learning pipeline using a deep-learning TENET to establish the arrow
时间的. Figure 2A shows how we used sliding windows of brain signal time series from all brain
regions in all participants. Figure 2B shows how these sliding windows were then used in TENET,
a deep-learning network classifier with 13 layers for classifying the arrow of time. Figure 2C
shows how this strategy allowed us to study nonreversibility and nonequilibrium at different
levels of granularity, from global (all signals) to system level to individual nodes. TENET allows
to quantify the information transfer within the levels of granularity but not between levels.
Figure 2D shows how TENET should be able to characterise the degree of reversibility, 那
是, nonequilibrium for each sliding window (top panel). We trained the TENET on a large data-
set of data resting and seven tasks and, on a validation dataset, computed the means of the
prediction performance of the classifier output (across time) as a measure of the degree of non-
reversibility (middle panel). 重要的, all datasets were shortened to the same task duration
to avoid the potential confound of one condition gaining undue prominence due to more data
being available. The standard deviation of prediction performance establishes stability of this
nonreversibility across time. Given that nonequilibrium states are already nonstationary, 这
provides the second order of nonstationarity (参见方法).
Figure 2E shows that nonreversibility is associated with the breaking of detailed balance of
a system. In order to test the breaking of the detailed balance of the system, we selected
网络神经科学
972
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
The arrow of time of brain signals in cognition
windows of low and high reversibility and computing the time-delayed functional connectivity
between all pairs of brain regions. 具体来说, the degree of asymmetry of this matrix is a
proxy for the breaking of the detailed balance with more asymmetry corresponding to more
unbalance. 最后, Figure 2F shows how TENET can be used on resting-state data from neu-
ropsychiatric patients with diagnoses of schizophrenia, ADHD, and bipolar disorder, 还有
as age-matched controls. 重要的, computing the different levels of nonreversibility could
provide a potential biomarker of neuropsychiatric disease and reveal the underlying funda-
mental problem with interacting with the environment.
In the following, we established the role of the DMN in cognition by applying the TENET
framework in healthy participants engaged in rest and seven tasks. We first show the results at
the whole-brain level, followed by the system level and the regional node level. 最后, 我们
apply the TENET framework to resting state in three neuropsychiatric disorders.
Significant Global Differences in Brain-Environment Interactions for Rest and Seven Tasks
For the global level of analysis of how the environment is driving the brain out of equilibrium,
we extracted BOLD time series from the DK80 parcellation covering the whole brain in rest
and the seven tasks. For each of HCP participant, we extracted forward and backward patterns
in sliding windows with a length of 20 TRs (的 0.72 秒), which were then shifted 3 TRs
向前. Each of the sliding windows consisted of two input patterns containing (1) 向前
和 (2) time-reversed backward-sliding windowed time series, which was each labelled with
an output class label of forward and backward, 分别.
For the training of TENET, in order to perfectly balance the data and avoid any potential
source of bias, we used 890 HCP participants with the longest possible duration available in
all conditions (176 TRs). For generalisation, we performed the data analysis on a separate 100
HCP participants. The data analysis consisted of computing the level of nonequilibrium/
nonreversibility, 右(t), using the output of TENET on this generalisation set after being trained
on the bulk of the data.
As specified in detail in the Methods, 右(t) is computed as the accuracy of classification of
forward and time reversal of backward trajectory of the global time series (across sliding win-
dows at time t and across participants). Perfect classification of maximal nonreversibility is thus
assigned a value of 1 以及哪里 0 corresponds to full reversibility.
Figure 3A (left panel) contains a box plot showing that the brain dynamics during REST
have significantly lower levels of reversibility than in tasks (所有p < 0.01, Wilcoxon rank
sum, corrected for multiple comparisons with FDR). As can be seen, the highest level of non-
reversibility is found in the SOCIAL task, reflecting how the environment is forcing a stronger
arrow of time and thus nonreversibility. But, equally, the other tasks, ordered by levels of non-
reversibility (RELATIONAL, EMOTION, GAMBLING, MOTOR, WM (working memory) and
LANGUAGE) are significantly more hierarchically structured than REST, related to the
increase in the breaking of the balance directly related to the necessity of more structured
computation. It is interesting to note the significant differences between the tasks too (all
p < 0.01, Wilcoxon rank sum, corrected for multiple comparisons with FDR; all significant
comparisons between conditions except for MOTOR vs. LANGUAGE, MOTOR vs. WM,
and WM and GAMBLING).
In addition, the right panel shows the level of nonstationarity, which is the standard devi-
ation of the levels of nonreversibility across time. The differences between rest and tasks were
similar to the results of the mean of the nonreversibility in the sense that there were significant
Network Neuroscience
973
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The arrow of time of brain signals in cognition
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
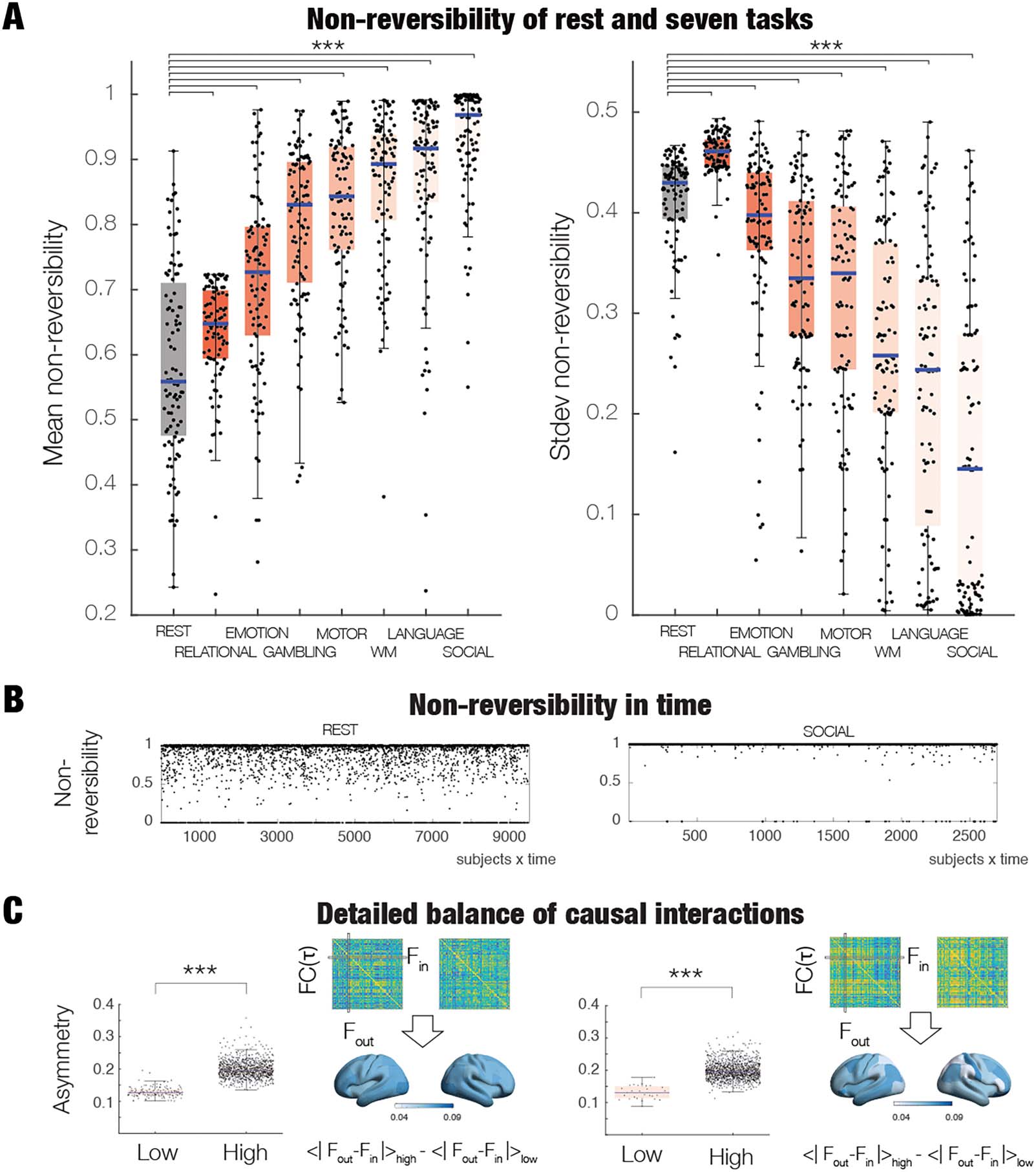
Figure 3. Global nonreversibility in HCP rest and seven tasks. (A) Left panel shows the mean nonreversibility for rest and the seven tasks
ordered by the increase in their mean level of nonreversibility. The level of nonreversibility, R(t ), is computed using the output of TENET on a
10% validation set after being trained on 90% of the data. In brief, R(t ) is computed as the accuracy of classification of forward and time
reversal of backward trajectory of global timeseries (across sliding windows at time t), where a value of 1 corresponds to perfect classification,
that is, maximal nonreversibility (see Methods). As can be seen from the box plot, brain dynamics during rest exhibits significantly lower levels
of reversibility than that found in tasks (all p < 0.01, Wilcoxon rank sum, corrected for multiple comparisons with FDR). The highest level of
nonreversibility is found in the Social task, reflecting a stronger arrow of time. In other words, the brain dynamics in tasks are showing more
nonreversibility than rest and therefore more hierarchical organisation underlying specific computations. The right panel shows the stability of
this nonreversibility across time, that is, providing a measure of second order of nonstationarity. Brain activity during rest is showing signif-
icantly more variability in the second order of nonstationarity than tasks (all p < 0.01, Wilcoxon rank sum, corrected for multiple comparisons
with FDR). (B) The panel shows the level of nonreversibility, R(t), over time for rest (left panel) and the social task (right panel). Note how the
evolution of R(t) is more variable in rest. (C) Interactions vary across time and consistently show a significantly stronger breaking of the detailed
balance in windows with high compared to low levels of nonreversibility (compare low and high box plots, p < 0.001) for both rest (left panel)
and the social task (right panel). This is measured as the asymmetry of the time-shifted functional connectivity (see Methods). The renderings of
brains reflect which brain regions are showing more symmetry breaking between low and high levels of nonreversibility. The brain shows more
heterogenous patterns of change during the social task than in rest (compare right with left panel).
Network Neuroscience
974
The arrow of time of brain signals in cognition
differences between all conditions (all p < 0.01, Wilcoxon rank sum; corrected for multiple
comparisons with FDR, except between MOTOR vs. LANGUAGE and WM and GAMBLING),
but importantly for this measure, the SOCIAL task had the lowest variability over time, which
was much lower than REST. On the other hand, REST is showing one of the largest levels of
nonstationarity, which is consistent with the idea that resting state involves less computation
and thus less breaking of the detailed balance. This can also be appreciated from Figure 3B,
where the two panels show time evolution of the levels of nonequilibrium/nonreversibility,
R(t), for REST (left) and the SOCIAL task (right).
Please note that the increase in nonreversibility during tasks, linked to the increase in pro-
duction entropy, is a measure of the increase in asymmetrical interactions. This should not be
confounded with the findings of a decrease in entropy for tasks found in the literature (He,
2013; Ponce-Alvarez et al., 2015). The entropy measured in these studies is a measure of
the variability of the state, which has been shown to decrease in task. As such, entropy and
production entropy are complementary measures of the system.
Differences in Asymmetry Breaking Between Rest and Tasks
As mentioned above, equilibrium is associated with the fluxes of transitions between different
states, that is, how the detailed balance of the transitions between the underlying states disap-
pear in completely equilibrium. In thermodynamics, a nonequilibrium system contains net
fluxes between the states as a function of broken balance, which is the source of nonreversi-
bility and thus of the arrow of time (Crooks, 1998; Feng & Crooks, 2008; Maragakis et al.,
2008; Seif et al., 2021; Shirts et al., 2003). In order to establish a quantitative link between
our measure of nonequilibrium/nonreversibility and broken detailed balance, we measured
the asymmetry of the time-shifted functional connectivity (see Methods).
In brief, in order to measure a proxy for the causal interactions, we selected patterns from sliding
windows of low and high reversibility, and computed the time-delayed functional connectivity
matrix, FC(τ), between all pairs of brain regions, over all participants and all sliding windows
for each condition of HCP REST and the SOCIAL task, which is the task with the most nonrever-
sibility. The global level of asymmetry was computed for each sliding window as the mean value
over the elements of the difference between this matrix and its transposed. In contrast, for the node
level of asymmetry, we first computed the incoming and outgoing regional flow for each sliding
window and then computed the average over all sliding windows and participants of the absolute
difference between the two regional flows (see Methods for detailed information). We render the
change between high and low levels of the node-level asymmetry.
As can be seen in Figure 3C, we found significantly stronger breaking of the detailed balance
in windows with high compared to low levels of nonequilibrium/nonreversibility (compare low
and high box plots, p < 0.001, Wilcoxon rank sum). On the right of the box plot, we show an
example of the asymmetry matrices for a single participant at a given time point. Below render-
ings are shown of the change between low and high levels of the node-level asymmetry.
Consistent with the close link between symmetry breaking and our measure, we found more
heterogenous patterns of change during the SOCIAL task than in REST. This again demonstrates
that when engaged in a task, the environment is driving the brain in very specific ways to higher
levels of nonreversibility. Please note, however, that in general, the breaking of the detailed
balance reflects the level of nonreversibility, although there could, of course, be rare cases of sys-
tems where the absence of directed information flow could still lead to nonreversibility (Ao, 2008).
Please note that the findings of TENET that tasks have higher nonreversibility than resting
state make it highly unlikely the results are biased by the onset of task blocks. Four lines of
Network Neuroscience
975
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The arrow of time of brain signals in cognition
argumentation supports this: (1) The asymmetry of the HRF associated with task blocks could be
a potential confound, especially if the windows used for classification are synchronised with
the task blocks. However, crucially, here we are using the same windows for the classification
of rest and all tasks, where the task onsets are completely different. Furthermore, similar to tasks,
resting state is also composed of spontaneous neuronal activity, which is also convolved with
HRF. Ultimately, if HRF was a confound, it would not be specific to task or rest. In fact, there is a
large literature showing how resting state can be obtained using neural discrete event convo-
lution (e.g., Tagliazucchi et al., 2016). (2) The results show that the SOCIAL task is the task with
the highest nonreversibility and RELATIONAL has the lowest. Yet, SOCIAL has less blocks than
RELATIONAL, making it unlikely that task blocks are important for the estimation of reversibility.
(3) TENET uses sliding windows of 20 TRs (14 sec) with increase of 2 or 3 TRs, which are smaller
than most task blocks and makes it unlikely that the results are biased by the task blocks. (4) As
shown in Figure 3C, the windows with high nonreversibility are the ones with maximal asymme-
try. Note that this shows that the temporal asymmetry correlates with the hierarchical organisation,
that is, asymmetry of interactions. This is consistent with the examples using spin models used by
Lynn and colleagues, showing that nonreversibility (production entropy) correlates with the break-
ing of the detailed balance, that is, the asymmetry of interactions. In other words, different tasks
require different hierarchical organisation (asymmetry interactions), which is detected by the
nonreversibility.
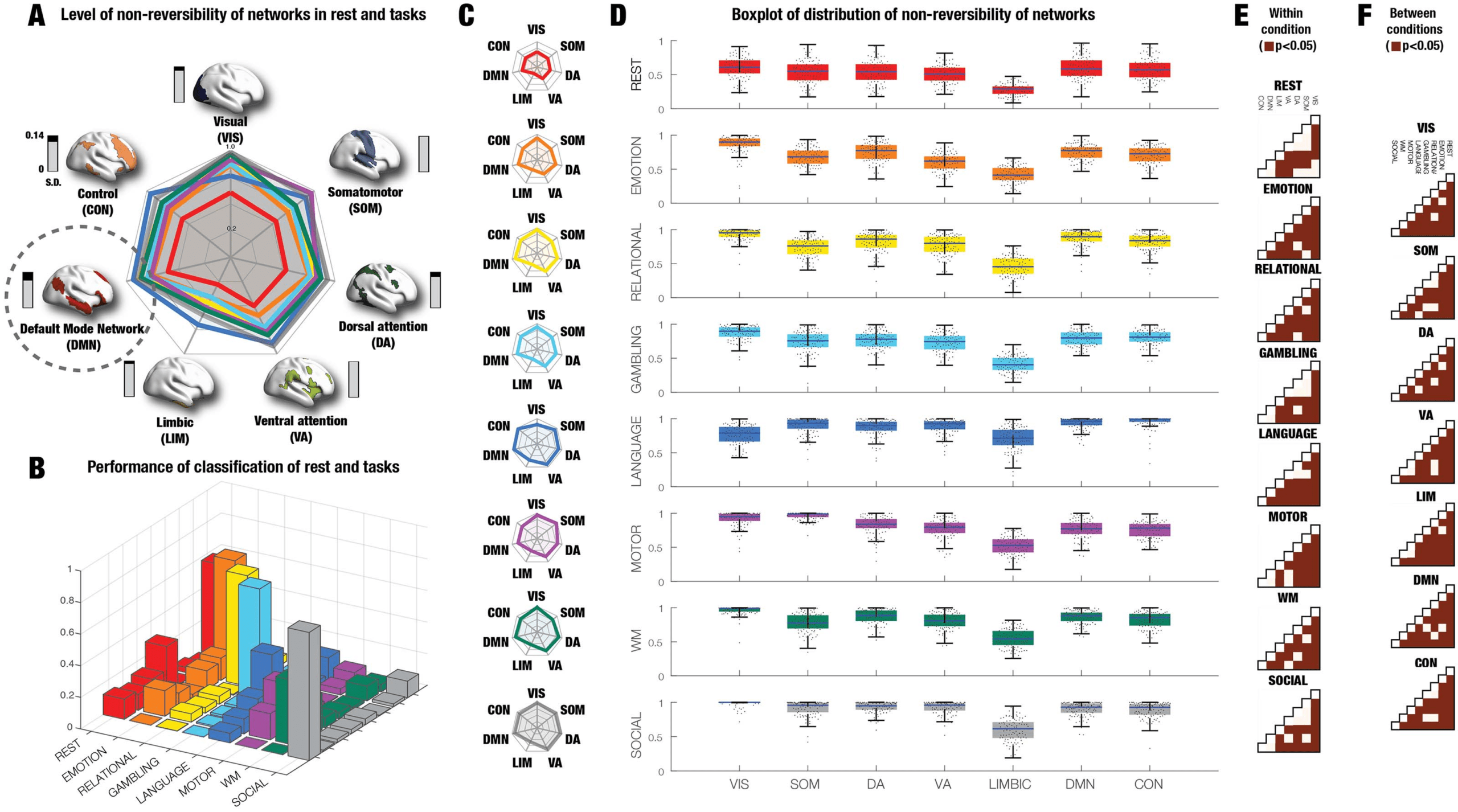
System-Level Analysis of HCP Data Shows That DMN is the Most Endogenous Network
In order to assess the system level of the brain-environment interactions between rest and task,
we used TENET framework in the same manner as in the global analysis, but now used on the
parcels belonging to each Yeo network in the Schaefer 500 parcellation. Again, in order to
balance the data, we used 890 HCP participants with the longest possible duration available
in all conditions (176 TRs). The results are from the generalisation that was performed on a
separate 100 HCP participants (see Methods). We used the same sliding window size and
shifting of this window as in the global-level analysis, but now the input is the window size
multiplied by the number of parcels for a given level analysis of HCP rest and tasks. Analysis of
the nonreversibility of the seven Yeo resting-state networks showed differential responses
between rest and tasks for the seven resting-state networks.
Figure 4A shows the combined spider plot of the different levels of nonreversibility for each
Yeo network in rest and the seven tasks, with a rendering of each Yeo network using separate
colour coding. The bar next to the renderings shows the standard deviation across rest and
tasks (ranging from 0 and 0.14). Importantly, the smallest standard deviation is found for
DMN (circled), which is therefore the most stable and endogenous network. This corroborates
the compelling anatomical and functional neuroimaging evidence from Margulies and others
showing that the physical and functional distance of the regions of the DMN are maximally
removed from sensorimotor regions (containing extrinsically driven neural activity directly
linked to perception and action). Taken together this points to DMN being a strong candidate
for orchestrating cognition.
Further, we investigated the possibility for classifying the conditions based on the system-level
TENET output. Using a support vector machine (SVM) with Gaussian kernels on the 100 HCP
participants used for generalisation. For the SVM, we subdivided the 100 participants into
90% training and 10% validation, repeated and shuffled 100 times. The SVM had seven inputs
(the Yeo resting-state networks) corresponding to the output produced by the system-level TENET.
The output was eight classes corresponding to the conditions (rest and seven tasks). Figure 4B
Network Neuroscience
976
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The arrow of time of brain signals in cognition
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. TENET system-level analysis of HCP rest and tasks. Similar to the whole-brain level analysis, we found different brain-environment
interactions between rest and task at the system level. Here our focus was on revealing how the seven Yeo networks are changing between rest
and tasks. (A) The combined spider plot shows the different levels of nonreversibility for each Yeo network in rest and the seven tasks. Each Yeo
network is rendered on the brain with a separate colour coding. Most importantly, the bar next to the rendering shows the standard deviation
across rest and tasks (ranging from 0 and 0.14). Importantly, the smallest standard deviation is found for DMN (circled), which is therefore the
most endogenous network. This can be linked with the compelling anatomical and functional neuroimaging evidence from Margulies and
others showing that the physical and functional distance of the regions of the DMN are maximally removed from sensorimotor regions (con-
taining extrinsically driven neural activity directly linked to perception and action). Together this provides evidence that the DMN is a strong
candidate for orchestrating cognition. (B) The system-level TENET results also allowed for a classification of conditions (rest and seven tasks). As
can be seen from the confusion matrix, the SVM provides excellent classification results much above chance level. Average classification for the
diagonal is 59% with a chance level of 12.5%. (C) Further probing the nonreversibility of the seven Yeo resting-state networks is demonstrated by
differential responses between rest and tasks for the seven resting-state networks shown by the spider plots of the level of nonreversibility for each
Yeo network in rest and seven tasks (colour coded similar to panel A). (D) This can be seen even more clearly in the box plots (for the validation
data), where, similar to the global-level analysis, there are lower levels of nonreversibility for rest compared to the seven tasks, suggesting, as
expected, that REST is more intrinsic and thus more in equilibrium. It is of interest to note that REST is characterised by having the highest levels
of nonreversibility in the DMN and Visual ( VIS) network. Equally, across the tasks, except for LANGUAGE, the sensory networks ( VIS and SOM)
show the highest level of nonreversibility. Interestingly, again except for LANGUAGE, the limbic network (LIM) exhibit the lowest levels of
nonreversibility. Overall, of the seven tasks, the Yeo networks in EMOTION and RELATIONAL show almost as low levels of nonreversibility levels
as REST. (E) In order to show the statistical significance, we show the comparisons between the level of nonreversibility of the seven Yeo networks
within condition (rest and the seven tasks). The figure shows the significance in the lower quadrangle of the matrices with brown squares signifying
p < 0.05, Wilcoxon rank sum. As can be seen almost all comparisons within conditions are significant but less so for REST. (F) Similar, we show the
statistical significance between conditions for the level of nonreversibility of the seven Yeo networks. The figure shows the lower quadrangle of the
matrices (with brown squares signifying p < 0.05, Wilcoxon rank sum). Almost all comparisons across conditions are significant.
shows the resulting confusion matrix, which provides excellent classification results much above
chance level with an average classification accuracy of 59% (on the diagonal) compared with
the chance level of 12.5% (using permutation testing; see Methods). Interestingly, the results of
classifying rest versus all tasks, produced a very high accuracy of 93.1% on the generalisation
dataset, using exactly the same procedure as for classifying the individual tasks.
Network Neuroscience
977
The arrow of time of brain signals in cognition
Figure 4C and 4D show the differential responses between rest and tasks for the seven
resting-state networks by presenting the spider plots and box plots of the level of nonreversi-
bility for each Yeo network in rest and seven tasks (for the validation data, colour coded similar
to Figure 4A). Similar to the global-level analysis, the lower levels of nonreversibility were
found for rest compared to the seven tasks, suggesting that REST is more reversible and there-
fore less hierarchical. Overall, of the seven tasks, the Yeo networks in the tasks EMOTION and
RELATIONAL show almost as low levels of nonreversibility levels as REST. Interestingly, the
VIS and SOM networks exhibit very high levels of nonreversibility in all tasks, mostly likely
reflecting the computational requirements for the sensory regions to work out the complexity
of the environment. Similarly, the LIMBIC network has lower levels of nonreversibility in all
tasks, perhaps reflecting the intrinsic nature of this network.
Figures 4E and 4F show the statistical significance within and across conditions, respec-
tively. Both figures show the significance in the lower quadrangle of the matrices with brown
squares signifying p < 0.05, Wilcoxon rank sum. As can be seen almost all comparisons within
and across conditions are significant.
Node-Level Analysis of Healthy Individuals Reveals DMN Orchestration of Cognition
Beyond the global and system-level analyses, we were interested in studying what endogenous
brain regions are common across rest and tasks, and thus able to orchestrate cognition inde-
pendently of the environment.
To this end, we applied the TENET framework at the node level using exactly the same
amount of data across rest and seven tasks, similar as above (see Methods). For the training
of TENET, in order to perfectly balance the data and avoid any potential source of bias, we
used 890 HCP participants with the longest possible duration available in all conditions
(176 TRs). For generalisation, we performed the data analysis on a separate 100 HCP
participants.
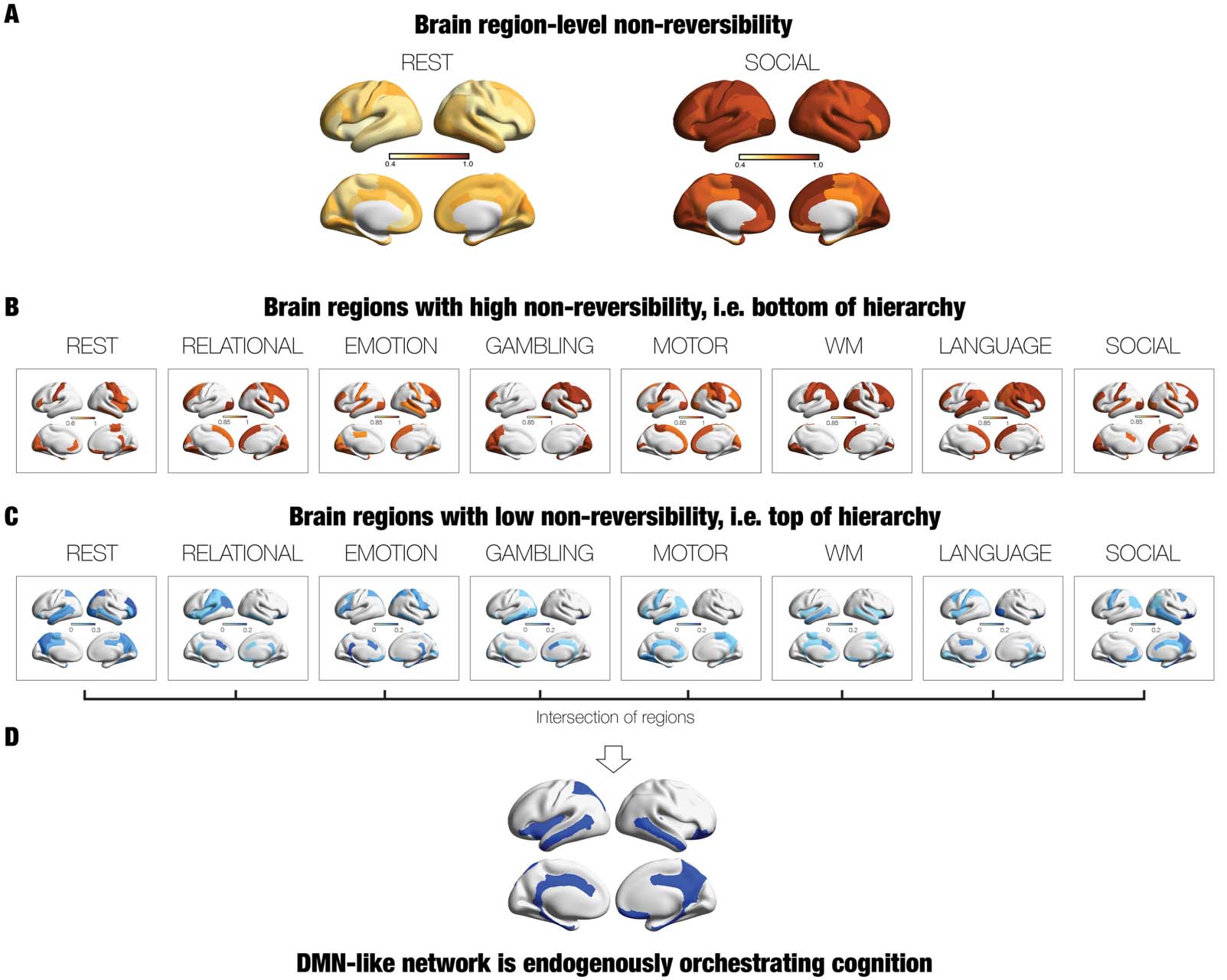
The node-level rendering for nonreversibility for REST and the SOCIAL task is shown in
Figure 5A. Similar to the global results, these are the two conditions with the lowest and highest
levels of nonreversibility (compare lighter shades of brown for REST to the darker for SOCIAL).
However, here we were also able to draw out the interregional heterogeneity. To further draw
out the differences between tasks, in Figure 5B we render the thresholded node level of
nonreversibility for all the seven tasks (thresholded to include the upper 30% quantile). This
demonstrates that sensorimotor regions are among the most stable regions and that there are
clear differences between the tasks.
Our main goal here, however, was to identify the endogenous regions at the top of the
hierarchy. Figure 5C shows the lower 30% quantile levels of nonreversibility for rest and all
the seven tasks, where darker blue colours show the less nonreversible brain regions. Note
how these regions are primarily located in higher order regions on the midline of the brain.
Confirming this finding, Figure 5D shows the brain regions common to the eight conditions
(rest and seven tasks) by selecting the lower 25% quantile levels for each condition and com-
puting the intersection between conditions for brain regions with low nonreversibility, that is,
top of hierarchy. This revealed a set of brain regions (including precuneus/posteromedial, tem-
poral, and ventromedial orbitofrontal cortices, mostly overlapping with the DMN), which are
common across all conditions and thus orchestrating cognition independently of the environ-
ment. Interestingly, this intersection also included regions not normally associated with the
DMN such as the insula and superior parietal cortex.
Network Neuroscience
978
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The arrow of time of brain signals in cognition
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Node-level analysis reveals that a DMN-like network is endogenously orchestrating cognition. Applying TENET framework at the
brain region level can distinguish the hierarchical organisation between rest and tasks. (A) The figure shows a brain rendering of the mean
node-level nonreversibility (across participants) for resting and the social task, which show the lowest and highest levels of global nonrever-
sibility, respectively. This is equally true at the node level but with significant interregional heterogeneity (compare the different shades in a
common colour map from yellow to brownish red). (B) The figure shows the upper 30% quantile levels of nonreversibility for brain regions in
rest and all the seven tasks, which shows that sensorimotor regions clearly at the bottom of the hierarchy. There are also clear differences
between the tasks. For instance, the working memory ( WM) task shows high levels of nonreversibility in prefrontal regions, while the
LANGUAGE task shows high levels of nonreversibility in known language areas, consistent with the existing extensive literature. (C) In contrast
to the previous renderings, this figure shows the brain regions at the top of the hierarchy, that is, more endogenous, rendering the lower 30%
quantile levels of nonreversibility for rest and all the seven tasks. In the rendering, darker blue colours are more endogenous and thus at the top
of the hierarchy. (D) Combining these eight conditions (rest and seven tasks) by selecting the intersection (see text) revealed that a set of regions
(including precuneus/posteromedial, temporal, and ventromedial orbitofrontal cortices), mostly overlapping with the DMN is common across
all conditions and thus orchestrating cognition.
In Figure S1 we further investigated these finding, demonstrating that our new measure
characterises the engagement across the whole brain rather than just in sensory regions. We
compared them directly to the myelinisation ratio (T1w/T2w ratio, obtained from HCP data),
which contains high values in the sensory regions of visual, somatomotor and auditory (Burt
et al., 2018). The nonsignificant correlations between the node levels of nonreversibility for
both REST and SOCIAL task (left row) with this map could indicate that the new measure is not
just linked to sensory but primarily to higher associative brain regions across the whole brain.
On the other hand, computing the full intersection as the mean across all conditions and cor-
relating this with the myelinisation ratio yielded a significant correlation (r = 0.26, p < 0.05,
nonparametric). Given that the top of the hierarchy of the intersection (Figure 5C) consists of
key regions in the DMN, the correlation with myelinisation provides further evidence for the
role of the DMN in orchestrating brain function.
Network Neuroscience
979
The arrow of time of brain signals in cognition
Further investigating links to other source of heterogeneity led us to investigate the various
forms of gene expressions in the brain as obtained from the Allen Human Brain Atlas
(Arnatkeviciute et al., 2019; Deco et al., 2021a; Fornito et al., 2019; Hawrylycz et al., 2012).
The middle row of Figure S1 shows the correlations between and the first PCA component of
all genes and the node level of nonreversibility of REST (top) and SOCIAL task (bottom). Interest-
ingly, there was a significant correlation between the PCA genes values and node levels in the
SOCIAL task (r = 0.47, p < 0.001, nonparametric), but not with the node levels in REST. Again,
there was a significant correlation (r = 0.48, p < 0.001, nonparametric) with the intersection
between conditions for brain regions with low nonreversibility, that is, top of hierarchy.
We also investigated another major source of heterogeneity, namely, the excitation-
inhibition (E-I) ratio given by the gene expression for genes coding for the excitatory AMPA
and NMDA receptors and inhibitory GABA-A receptor isoforms and subunits. In contrast to the
PCA maps, the rightmost row of Figure S1 shows a significant correlation between the node
level of nonreversibility in REST and the E-I values (r = 0.23, p < 0.04, nonparametric) but not
for the node levels in the SOCIAL task.
Using the Arrow of Time in Neuropsychiatric Disease
Given that the TENET framework by design measures how the environment is driving the
brain, and its high level of sensitivity demonstrated above, it would appear a promising avenue
for better characterising the differences between health and neuropsychiatric diseases. We
therefore applied the TENET framework on the large public UCLA dataset of neuropsychiatric
patients with schizophrenia, bipolar, and ADHD and matched control group of participants.
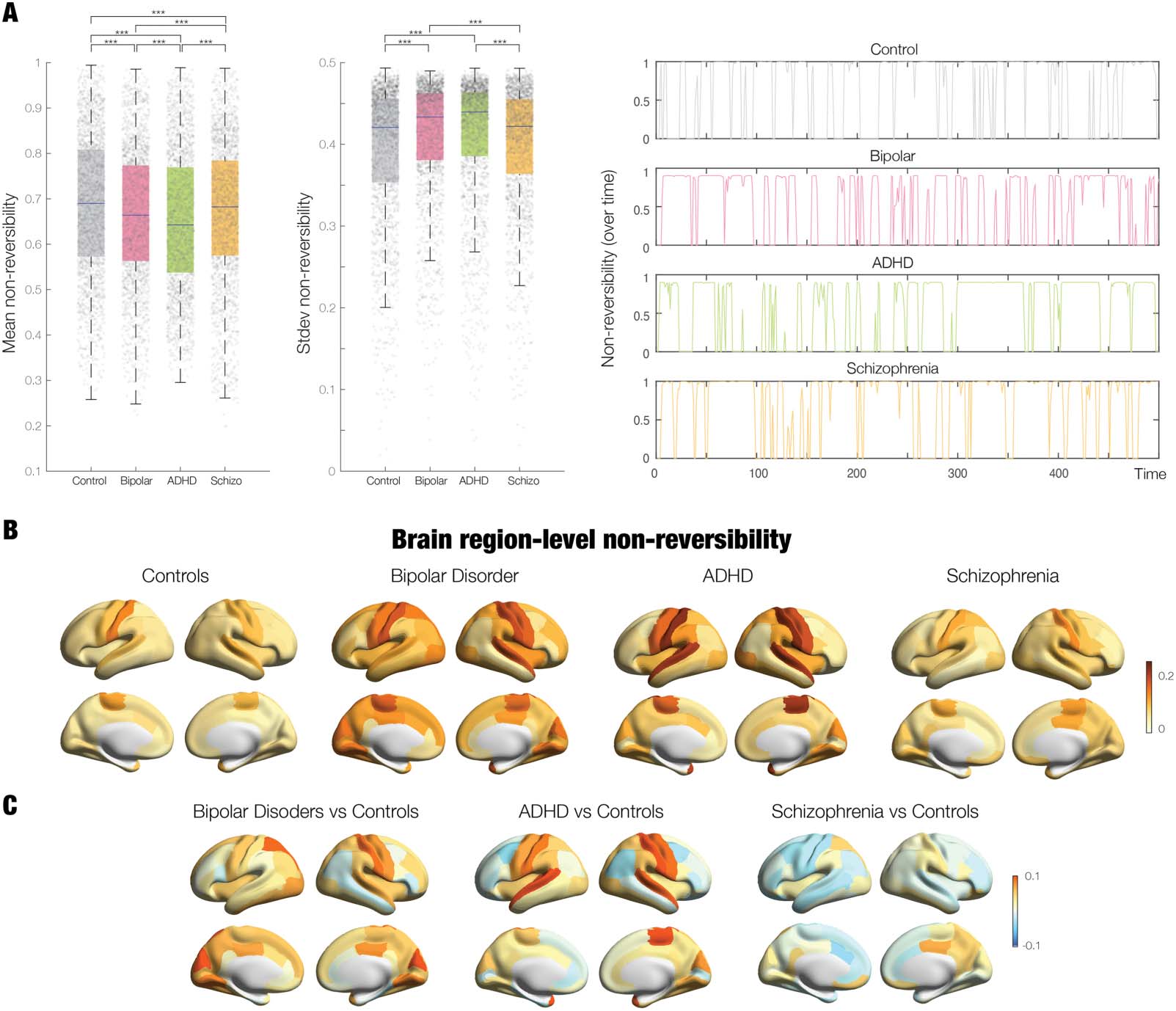
Figure 6 shows the results of using the TENET framework to establish the reversibility on the
global and local node levels for the four groups. The left panel of Figure 6A shows box plots of
the average reversibility across time, where the control group was significantly higher than
each of the neuropsychiatric groups (all p < 0.05, Wilcoxon rank sum, corrected for multiple
comparisons with FDR). This suggests that neuropsychiatric disease reduces the levels of non-
reversibility, suggesting that the brain is less hierarchical. Furthermore, each neuropsychiatric
disease group was significantly different from each other (all p < 0.05, Wilcoxon rank sum,
corrected for multiple comparisons with FDR). Given that the median values are similar but
significant, we also provide the effect size for each comparison: control versus bipolar: 0.1005;
control versus ADHD: 0.1606; control versus schizophrenia: 0.0529; bipolar versus ADHD:
0.0693; bipolar versus schizophrenia: 0.0512; and ADHD versus schizophrenia: 0.1130.
The middle panel of Figure 6A shows box plots of the standard deviation of the reversibility
across time for each of the four group, reflecting the levels of nonstationarity. This is signifi-
cantly reduced for the control participants compared to neuropsychiatric groups and between
them (all p < 0.05, Wilcoxon rank sum, corrected for multiple comparisons with FDR except
controls vs. schizophrenia and bipolar vs. ADHD), that is, the brains of patients with ADHD
and bipolar disorder are more nonstationary than controls. To further appreciate the differ-
ences between groups, right panel of Figure 6A plots examples of the temporal evolution of
the global reversibility computed by TENET for all the four groups.
These promising results prompted us to use the TENET framework to compute the mean
node-level reversibility for each group. Figure 6B shows the corresponding renderings on
the human brain. We also computed the differences between the mean in the control group
with the three neuropsychiatric groups, shown rendered in Figure 6C. As can be seen, and
interpreted in details in the discussion, there are clear differences between groups, which sug-
gest that the node level of nonreversibility might be useful as a biomarker for disease.
Network Neuroscience
980
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The arrow of time of brain signals in cognition
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. The arrow of time in neuropsychiatric disease. We used the nonreversibility on the global and local node levels on the large public
UCLA dataset of neuropsychiatric patients with schizophrenia, bipolar, and ADHD and matched control group of participants. (A) First, we
used the TENET framework to compute the level of nonreversibility at the global level for each group. The left panel of box plots shows that the
average reversibility across time for the control group is significantly higher than each of the neuropsychiatric groups (all p < 0.05, Wilcoxon
rank sum, corrected for multiple comparisons with FDR). In addition, each neuropsychiatric disease group are significantly different from each
other (all p < 0.05, Wilcoxon rank sum, corrected for multiple comparisons with FDR). The middle panel of box plots shows that the standard
deviation of the reversibility across time is significantly reduced for the control participants compared to neuropsychiatric groups and that
between them, there are also significant differences (all p < 0.05, Wilcoxon rank sum, corrected for multiple comparisons with FDR, except
for comparisons between controls vs. schizophrenia and bipolar vs. ADHD). The right panel shows examples of the temporal evolution of the
reversibility computed by TENET for a participant from each of the four groups. (B) Complementing these findings at the global level, we used
the TENET framework to compute the node-level reversibility for each group and show the corresponding thresholded renderings. (C) In order
to stress the differences between the control group and the three neuropsychiatric disorders, we show renderings of these differences.
DISCUSSION
Here we developed a thermodynamics-inspired, deep-learning TENET framework designed to
identify the hierarchical organisation of any brain state. This allowed us to address a central,
challenging problem in human cognitive neuroscience, namely, what brain networks are coor-
dinating the recruitment and scheduling of networks for computing and solving task-specific cog-
nitive problems. The leading hypothesis in human cognitive neuroscience is that the DMN is
responsible for this orchestration (Smallwood et al., 2021). Using the TENET framework, we were
able to demonstrate the breaking of detailed balance in rest and cognition. We found that an
endogenous network largely overlapping with the DMN is responsible for the orchestration of
cognition across rest and tasks. The TENET framework quantified the asymmetry in the flow of
events, ‘arrow of time’, in human brain signals. The results reveal the independent, endogenous
Network Neuroscience
981
The arrow of time of brain signals in cognition
regions able to orchestrate activity, in contrast to the sensorimotor regions at the bottom of the
hierarchy. These findings are in agreement with the findings of Margulies and colleagues, who
demonstrated that the physical and functional distance of the regions of the DMN are maximally
removed from sensorimotor regions (Margulies et al., 2016; Smallwood et al., 2021).
Whole-Brain Differences in Brain-Environment Interactions in Health and Disease
We found higher global levels of nonreversibility in different tasks than during resting state in
the large-scale HCP neuroimaging dataset (Figure 3). Similarly, we showed lower and less var-
iable global levels of nonreversibility across time in three different neuropsychiatric patient
groups (ADHD, schizophrenia, and bipolar disorder) in the large-scale UCLA neuroimaging
dataset compared to healthy participants (Figure 6). This suggests a flattening of the functional
hierarchy in disease, perhaps reflecting the lower asymmetry in functional interactions
between brain regions. As speculation, this could directly influence the computation and lead
to the more rigid and less flexible behavioural repertoire found in neuropsychiatric disorder.
At the System Level, DMN is the Most Endogenous Network
Complementary to investigating the hierarchical organisation of brain states by quantifying the
nonreversibility at the global level, we aimed to discover if there is more information to be
extracted from the system level, specifically regarding differences between rest and task. We
used the TENET framework on the seven Yeo resting-state networks in the Schaefer 500 par-
cellation. The results again showed lower levels of nonreversibility for rest compared to the
seven tasks, suggesting that REST is more in equilibrium, while the tasks have higher levels of
nonreversibility in the sensory networks ( VIS and SOM).
Interestingly, the REST condition shows the highest levels of nonreversibility in the DMN and
visual (VIS) network. Similarly, the sensory networks ( VIS and SOM) are showing the highest level
of nonreversibility across the seven tasks (except for LANGUAGE), and thus at the bottom of the
hierarchy. The lowest levels of nonreversibility are found in the limbic network (LIM), signalling
that the LIM network is less endogenous. Of the seven tasks the lowest levels of nonreversibility
are found in the EMOTION and RELATIONAL tasks, which are almost as low as REST.
We wanted to measure the stability of the different networks across the conditions, which is
captured by the standard deviation in the level of nonreversibility across time. In other words,
having the smallest standard deviation across time implies stability. Importantly, as shown in
Figure 4A in the combined spider plot in Yeo networks across all conditions, the DMN showed
the smallest standard deviation. This confirmed that the DMN is indeed the most endogenous
and at the top of the hierarchy, fitting well with the topographical evidence showing that the
DMN is located in regions furthest away from those contributing to sensory and motor systems
(Margulies et al., 2016; Smallwood et al., 2021). As such this provides further evidence that the
DMN is strong candidate for leading the orchestration of cognition.
In addition, the system-level analysis can also be used to accurately classify condition.
Using a SVM with Gaussian kernels to classify the conditions (rest and seven tasks), the results
showed a very high accuracy of 59% compared to the 12.5% chance level. Equally, just clas-
sifying rest compared to all tasks the level of accuracy to 93.1%, showing that system-level
TENET is an excellent method for distinguishing different cognitive brain states.
Node-Level Identification of the Most Endogenous Brain Regions
We also used the TENET framework at the node level, which revealed that a network of endog-
enous brain regions largely overlapping with the DMN is orchestrating behaviour across rest
Network Neuroscience
982
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The arrow of time of brain signals in cognition
and tasks. This provides further confirmation of the DMN as a strong candidate for leading the
orchestration of cognition.
In contrast to these results investigating the more endogenous brain regions, we were also
able to find brain regions with higher nonreversibility. Similar to the findings at the global scale,
the findings at the local scale reveal clear interregional heterogeneities between rest and tasks
(see Figure 5). The findings show that in many cases higher associative brain regions are more
nonreversible during task performance in the HCP dataset. Specifically, Figure 5A shows the
node-level rendering for nonreversibility for REST and the SOCIAL task. Like the results for the
global level, these are the two conditions with the lowest and highest levels of nonreversibility.
The power of the TENET framework is perhaps illustrated in the results for the MOTOR task,
where the thresholded results show selective engagement of the somatomotor regions as
expected (Barch et al., 2013), but equally the results show engagement in visual cortices
and midline medial prefrontal regions. Another good example is the LANGUAGE task, where
the results show broad engagement of ventral lateral prefrontal cortex, superior and inferior
temporal cortex—including the anterior temporal poles bilaterally (Barch et al., 2013; Binder
et al., 2011). Also, as expected from this primarily auditory task, the visual regions are not in
the top 15% of the regions with the highest nonreversibility.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Similarly, the WM task shows high levels of driving in regions including the MPFC, poste-
rior cingulate, and the occipital-parietal junction, fully consistent with the literature (Barch
et al., 2013; Drobyshevsky et al., 2006). In particular recent studies have demonstrated the
importance of activity in MPFC for memory guided decision-making (Murphy et al., 2018,
2019). In these tasks, the decision is not possible based on the received input and thus dem-
onstrates how the DMN orchestrates behaviour. In addition, the activity in the MPFC was also
linked to self-reports indicating a greater focus on task-relevant detail (Murphy et al., 2019;
Turnbull et al., 2019).
The findings support the view that the spatiotemporal dynamics of the brain cannot be ade-
quately captured by solely separable intrinsic and task-evoked dichotomy, but rather a
dynamic interplay of task-appropriate functional reconfigurations (Bolt et al., 2018).
Heterogeneity
We further investigated the heterogeneity found at the node level by comparing the TENET
results to various known forms of heterogeneity such as the gene expressions maps obtained
from the Allen Human Brain Atlas (Arnatkeviciute et al., 2019; Deco et al., 2021a; Fornito et al.,
2019; Hawrylycz et al., 2012). We found a significant correlation between the PCA genes
values and node levels in the SOCIAL task, but not with the node levels found in REST (see
Figure S1). The reverse was true for excitation-inhibition ratio given by the gene expression,
which was correlated with the node levels in REST but not in SOCIAL. These significant differ-
ences between rest and task show how the environment is directing changes in the functional
hierarchy. It is of considerable interest that the E-I ratio is significantly correlated with node
levels in REST and not the node levels in task, since this suggest that REST is more intrinsically
shaped. On the other hand, the significant correlation between the node levels in the SOCIAL
task (but not with REST), suggests that the level of driving by the environment is not fully free but
still constrained to a certain degree by genetics (here captured by the first PCA of the genes).
Further investigating the heterogeneity, we compared REST and SOCIAL directly to the
myelinisation ratio (T1w/T2w ratio), which contains high values in the sensory regions of
visual, somatomotor, and auditory (Burt et al., 2018). We did not find a correlation to the node
Network Neuroscience
983
The arrow of time of brain signals in cognition
levels of nonreversibility for both REST and SOCIAL task. This result clearly demonstrates that
the environment is not driving the brain to nonreversibility by solely affecting the sensory
regions but equally through driving higher associative brain regions across the whole brain.
This fits well with the evidence that the three sources of heterogeneity investigated here are
correlated among themselves (Burt et al., 2018; Deco et al., 2021a), yet influencing the func-
tional hierarchy at the node level of nonreversibility differently under different conditions.
Overall, these results provide evidence for the severe constraints in terms of degrees of free-
dom that the brain has to operate within.
Node-Level Investigation of Neuropsychiatric Disorders
As shown above, at the global level, the three neuropsychiatric patient groups (ADHD, schizo-
phrenia, and bipolar disorder) had lower levels of nonreversibility compared to healthy par-
ticipants. Further investigating this finding, we also found significant local heterogenous
node-level changes differentiating between the different disorders. Interestingly, in schizophre-
nia compared to controls, we found local decreases across the brain but primarily located in
the temporal, parietal, and prefrontal cortices. These regions clearly have less nonreversibility
compared to controls, which is compatible with the literature showing that the disorder is asso-
ciated with more isolation, as a function of the loss of balance between intrinsic and extrinsic
activity (van den Heuvel & Fornito, 2014).
In contrast, while in bipolar disorder the overall level of nonreversibility is lower than
controls, the somatomotor regions show increases in nonreversibility, perhaps linked to the
findings in the literature of large, sudden swings in mood, given that the brain is in more non-
equilibrium (Furman et al., 2011; Menon & Uddin, 2010). Comparing ADHD with controls
shows larger local levels of nonreversibility in somatosensory, temporal, parietal, and insular
cortices. In particular, the somatomotor regions are more nonreversible than in controls, sug-
gesting a potential route for hyperactivity, while the lower nonreversibility in parietal regions
could be linked to the known attentional deficits in the disorder (Konrad & Eickhoff, 2010).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Discussion of TENET Framework
The results produced by the TENET framework relies on two essential elements, namely, the
concept of reversibility (as captured by the arrow of time) and how machine learning (and for
example deep learning) is able to quantify the reversibility of brain signals. In terms of the arrow
of time, this popularisation of this idea is usually credited to the physicist Arthur Eddington
(Eddington, 1928). Here we showed that this key idea from physics and thermodynamics can
equally well be applied in neuroscience. The second law of thermodynamics, as immortalised
by Rudolph Clausius (Clausius, 1865) and Sadi Carnot (Carnot, 1824) states a nonequilibrium is
characterised by the arrow of time which indicates the nonreversibility of a system. In fact, the
second law of thermodynamics can be expressed by the Clausius inequality, which establishes that
the work associated with the process (averaged over many repetitions) is larger than the change in
its free energy, which is the same as stating that the system is nonreversible and in nonequilibrium.
As shown by Seif and colleagues, rather than computing the production entropy, the arrow
of time can be cast as a game in which a player is shown either a forward or a backward
trajectory and has to guess the direction of the arrow of time. The only information available
is the trajectory, and from this a player must guess was generated by a forward or reverse pro-
cess. The accuracy is then given by the ratio of correct predictions to the total number of sam-
ples. This statistical problem can be very hard when the fluctuations are high, and therefore the
direction of the arrow of time is very difficult to ascertain (Crooks, 1998; Feng & Crooks, 2008;
Network Neuroscience
984
The arrow of time of brain signals in cognition
Jarzynski, 2011; Maragakis et al., 2008; Seif et al., 2021; Shirts et al., 2003). One way of solv-
ing this is to use a machine-learning algorithm trained such as deep learning to infer the direc-
tion of the arrow time. In fact, Seif and colleagues have demonstrated that classification
between forward and reverse time series can identify the arrow of time in systems where
the direction is known, and therefore establishes a one-to-one relationship between this clas-
sification, production entropy, and nonreversibility (Seif et al., 2021). Even more, Parrondo
and colleagues demonstrated this relationship analytically (Parrondo et al., 2009).
However, given that the learning of the classifier could fail, this classification method is a
necessary but not sufficient condition for assessing the arrow of time. Yet, as we show here this
can be mitigated by using the same deep-learning network with the same training and gener-
alisation data samples that balance the method and provide robust results when used to com-
pare nonreversibility across different brain states. Note that other machine-learning methods
(like SVM) could equally well be used. Indeed, as shown by de la Fuente and colleagues,
different machine-learning architectures can successfully be used to determine the temporal
irreversibility of neural dynamics of different brain states (de la Fuente et al., 2023). Yet, not all
machine-learning architectures are equally good. Hence, why we took great care to build a
suitable deep-learning network utilising the same parameters for comparisons. We note that
the main results obtained here are in full agreement with the published literature. Lynn and
colleagues used a very different approach to measure the production entropy and arrived at
the same result for a subset of the HCP data used here, namely, that task is more nonreversible
than rest. A similar confirmation of this result (using the full HCP dataset) was also obtained
using the INSIDEOUT framework of classifying forward and reverse time series of the same
dataset but not using machine learning (Deco et al., 2022).
In terms of the deep-learning framework, this method has received a lot of attention over
the last couple of years. This powerful machine-learning technique has proven highly useful
for providing solutions to a number of difficult computational problems ranging from vision to
playing Go (Sejnowski, 2018, 2020; Silver et al., 2018; Yang & Wang, 2020). However, some
criticisms have been raised over the largely black box nature of these advances, which have
had considerable practical utility for solving complex problems, but have produced little in
way of new insight into how this is achieved mechanistically (Marcus, 2018). Recent research
has, however, started to harness the power of deep learning for discovering useful underlying
mechanisms (Seif et al., 2021).
Here, we were not aiming to use deep learning as a technique for revealing underlying
brain mechanisms but rather simply as a tool for providing the level of the reversibility of
the arrow of time in brain signals. In other words, in our framework the important question
is to determine the level of distinction between forward and time-reversed back time series,
but not how this is achieved. As such, here the black box nature of deep learning was not
relevant for solving the nontrivial problem of determining nonreversibility.
As mentioned above, the TENET framework can be studied at all spatial scales. Here, how-
ever, we focused on three levels: global, system level, and node level, where the global level
considers all the whole-brain signals, while the node level considers the signals in each brain
region separately and the system level considers the typical large-scale resting-state networks
(Biswal et al., 1995, 2010; Smith et al., 2013). This allowed us to focus on different aspects of
nonequilibrium, that is, the interactions between the environment and different brain scales,
ranging from whole-brain to large-scale networks and to regions. Indeed, the node level turned
out to be a highly sensitive measure of quantifying and interpreting different cognitive brain
states and differences between health and multiple diseases.
Network Neuroscience
985
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The arrow of time of brain signals in cognition
Use of Thermodynamics for Assessing the Environment Through Brain Signals
The TENET framework is especially powerful, given that thermodynamics can quantify the
influence of the environment by the level of nonreversibility. More broadly, this is related to
a fundamental question in general biology, namely, how survival is a key characteristic of life
and requires the ability to find order in a complex, largely disordered environment. As proposed
by the Austrian physicist and Nobel Laurate Ernst Schrödinger, survival is predicated on avoid-
ing equilibrium: “How does the living organism avoid decay? … By eating, drinking, breathing
and … assimilating. The technical term is metabolism” (Schrödinger, 1944). The avoidance of
decay thus requires nonequilibrium interactions with the complex environment—and the
brain is at the heart of these interactions.
There is a long history of understanding how the brain is able to interact with the environ-
ment. The initial extrinsic perspective proposed that the brain is primarily, reflexively driven by
momentary stimulation from the environment in a task-driven manner (Raichle, 2010;
Sherrington, 1906; Yuste et al., 2005). A more recent, complementary perspective was proposed
by Marcus Raichle, which holds that the brain is mainly intrinsic, resting but switching between
states whilst “interpreting, responding to, and even predicting environmental demands”
(Raichle, 2006; Raichle et al., 2001). The evidence is clear that the brain’s metabolic energy
budget for maintaining the intrinsic resting activity is large (Zhang & Raichle, 2010). In fact, by
some estimates, 20% of the total energy consumption is taken up by the brain, which only rep-
resents 2% of body weight (Attwell & Laughlin, 2001; Clarke & Sokoloff, 1999; Magistretti et al.,
1999), which has led to Raichle’s poetic proposal of “dark energy” (Raichle, 2006).
The brain’s energy budget governs the flow of energy between the brain and environment,
which is the ultimate cause of the nonequilibrium essential to the proposals of Schrödinger and
Raichle. Any living system requires the breaking of the detailed balance of the transitions
between the underlying states (Gnesotto et al., 2018; Schrödinger, 1944). In a system with
detailed balance, the fluxes of transitions between different states disappear (Lynn et al.,
2021; Sanz Perl et al., 2021). This is conveniently described in the language of thermodynam-
ics, where a system ceases to produce entropy and becomes reversible in time (Jarzynski,
2011). In contrast, a nonequilibrium system—where the balance is broken—shows net fluxes
between the underlying states, and thus becomes irreversible, establishing an arrow of time
(Crooks, 1998; Feng & Crooks, 2008; Maragakis et al., 2008; Seif et al., 2021; Shirts et al.,
2003). This is closely linked to turbulence, a classical example of nonequilibrium, which
has been shown to be highly useful for optimally transferring energy/information over
space-time due to its mixing properties (Frisch, 1995). Turbulence has recently been demon-
strated in the human brain, where the resulting information cascade is crucial for extracting
order from disorder (Deco & Kringelbach, 2020; Sheremet et al., 2019).
The ideas for the present framework comes from physics and thermodynamics, where non-
equilibrium is intrinsically linked to nonreversibility (Seif et al., 2021) and the production of
entropy, leading to the arrow of time, as originally popularised by Arthur Eddington
(Eddington, 1928) and since studied in great detail (Crooks, 1998; Feng & Crooks, 2008;
Jarzynski, 2011; Maragakis et al., 2008; Seif et al., 2021; Shirts et al., 2003). In fact, a simple
yet powerful way of assessing nonequilibrium in the brain is to quantitatively estimate the
arrow of time of the brain signals rather than the more difficult way of estimating the produc-
tion of entropy (Lynn et al., 2021; Sanz Perl et al., 2021).
The nonreversibility of a physical process and the arrow of time is clearly illustrated when
watching a film of a glass being shattered, which is very different from watching the same film
in reverse. In thermodynamics, this can be elegantly described in terms of the entropy
Network Neuroscience
986
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The arrow of time of brain signals in cognition
production, which increases when a system becomes disordered. If the total entropy produc-
tion is larger than zero, it means that the system is nonreversible and in nonequilibrium. In
cases such when a glass is being shattered, the nonreversibility is very clear for all to see.
In contrast, a film of colliding billiards balls can be watched equally forward and backward,
making it very difficult to distinguish the correct arrow of time in the film. This process could
potentially be fully reversible and not producing entropy.
In most processes, however, like the evolution of brain signals, the level of reversibility is
much less clear. Here, we therefore used the power of deep machine learning to detect the
level of reversibility of empirical brain-imaging data. This allowed us to assess the level of
nonequilibrium in brain dynamics in different states. Specifically, for whole-brain data we
extracted the normal forward time series as well as constructing the time reversal of the
backward time series for a given parcellation. This procedure provides a clear arrow of time
for a given time series for which a deep-learning classifier (here named Temporal Evolution
NETwork, TENET) could be trained to identify the forward and the time reversal for a given
time series of any length. The performance of TENET provides a reliable measure of the degree
of nonreversibility and nonequilibrium at different levels of global and local brain organisation.
Perspectives
The present thermodynamics-based deep-learning TENET framework opens up for an exami-
nation of differences of nonreversibility for different levels of consciousness, from sleep and
anaesthesia to altered states of consciousness induced by psychedelics and meditation. The
framework can easily be extended to other neuroimaging modalities such as MEG/EEG. It can
even be used with LFP and other types of cell recordings in animals.
One particularly challenging question relates to the role of brain processing of reversible
external stimuli such as, for example, watching the forward and backward versions of the
movie of a glass being shattered. How will the nonreversibility in brain dynamics change with
identical stimuli, but where the order has been changed such that the arrow of time has been
violated? Will this elicit different nonreversibility in brain dynamics when showing the forward
and backward versions of a movie of billiard balls moving, which is not in any clear way vio-
lating the arrow of time? These stimuli are experienced radically different, which must be
linked to the interactions between the extrinsic stimulation with the dynamics of intrinsic pre-
dictability formed by prior experiences. However, intrinsic predictability is known to be
affected in neuropsychiatric disease, and it would be of considerable interest to study how
the reversibility of external stimulation influences the nonreversibility in brain dynamics. In
fact, this could potentially reveal new information about the interactions between intrinsic
and extrinsic dynamics, which are known to be compromised. The reversal of external stimuli
could also take place at higher cognitive levels, such as when inverting the nodes of Bach’s
fugue or that of a complex narrative.
From its conception, the arrow of time is coupled to the deep notion of causality. Thermo-
dynamics offers important tools for establishing the causal directionality of information flow
through the concept reversibility and entropy. There is of course a large literature on causality,
best summarised in the seminal work by Judea Pearl (Pearl, 2009), where he shows that any
framework of causal inference is based on inferring causal structures that are equivalent in
terms of the probability distributions they generate; that is, they are indistinguishable from
observational data, and could only be distinguished by manipulating the whole system.
In neuroscience, there have been numerous attempts to capture causality in brain dynam-
ics. One influential concept is ignition, the idea that a stimulus can ignite a causal chain of
Network Neuroscience
987
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The arrow of time of brain signals in cognition
events propagating across the brain (Dehaene et al., 1998). This ignition can happen as a result
of extrinsic stimulation (Joglekar et al., 2018; Mashour et al., 2020; van Vugt et al., 2018) or as
part of intrinsic events (Deco et al., 2017, 2021a). More sophisticated approaches use proba-
bilistic principles of mutual information (Brovelli et al., 2015; Pereda et al., 2005; Quiroga
et al., 2000, 2002) to determine the directional causality underlying the functional hierarchical
organisation of brain function (Deco et al., 2021b).
The concept of the arrow of time has also been investigated from the perspective of chaos
theory, originating with the work of Henri Poincaré who published the first description of cha-
otic motion in 1890 (Poincaré, 1890). Later work has confirmed that one key characteristic of
chaos is the infinite sensitivity to initial conditions (Strogatz, 2018). Given this extreme sensitivity,
even if a classic mechanic deterministic chaotic system is in principle reversible, in practice this is
in fact nonreversible. In other words, chaos makes it very difficult to establish the computational
reversibility and thus causality. Turbulence is a classic example of a spatiotemporal chaotic sys-
tem that is associated with nonequilibrium and thus nonreversibility. Interestingly, turbulence is a
highly useful dynamical regime for optimally transferring energy/information over space-time,
and it has recently been shown that brain dynamics are indeed turbulent (Deco & Kringelbach,
2020). The turbulent regime supports the information cascade that is crucial for extracting order
from disorder.
Overall, the novel thermodynamics-based deep-learning TENET framework can provide
detailed information of the varying levels of nonstationary and nonequilibrium nature of brain
dynamics in health and disease. The TENET framework offers a quantitative account of
differences in nonreversibility. Future work could integrate this with causal mechanistic
whole-brain modelling in a turbulent regime to deepen our understanding of how brain
dynamics organise human behaviour in the face of the second law of thermodynamics in
health and disease.
METHODS
Neuroimaging Ethics
For the HCP dataset, the Washington University–University of Minnesota ( WU-Minn HCP)
Consortium obtained full informed consent from all participants, and research procedures
and ethical guidelines were followed in accordance with Washington University institutional
review board approval.
For the UCLA dataset, as detailed in Poldrack et al. (2016), the Consortium for Neuropsy-
chiatric Phenomics recruited neuropsychiatric participants and healthy controls who gave
written informed consent following procedures approved by the Institutional Review Boards
at UCLA and the Los Angeles County Department of Mental Health.
Neuroimaging Participants HCP Rest and Task
The dataset used for this investigation was selected from the March 2017 public data release
from the HCP where we chose a sample of 990 participants from the total of 1,003 partici-
pants, since not all participants performed all tasks.
The HCP Task Battery of Seven Tasks
The HCP task battery consists of seven tasks: working memory, motor, gambling, language,
social, emotional, and relational, which are described in details on the HCP website (Barch
et al., 2013). HCP states that the tasks were designed to cover a broad range of human
Network Neuroscience
988
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The arrow of time of brain signals in cognition
cognitive abilities in seven major domains that sample the diversity of neural systems: (1)
visual, motion, somatosensory, and motor systems; (2) working memory, decision-making,
and cognitive control systems; (3) category-specific representations; (4) language processing;
(5) relational processing; (6) social cognition; and (7) emotion processing. In addition to
resting-state scans, all 1,003 HCP participants performed all tasks in two separate sessions (first
session: working memory, gambling and motor; second session: language, social cognition,
relational processing, and emotion processing). As a test-retest control condition, a small sub-
sample of 45 HCP participants performed the paradigm twice.
Neuroimaging Participants UCLA Rest
Consortium for Neuropsychiatric Phenomics published a dataset with neuroimaging as well as
phenotypic information for 272 participants. We used the preprocessed data with a total of
261 participants, since seven of the participants were missing T1-weighted scans (Gorgolewski
et al., 2017), and three healthy controls and one ADHD patient were missing resting-state
scans. The total population analysed consists of 122 healthy controls, as well as participants
with diagnoses of adult ADHD (40 patients), bipolar disorder (49 patients), and schizophrenia
(50 patients).
Neuroimaging Structural Connectivity and Extraction of Functional Time Series
HCP preprocessing and extraction of functional time series in fMRI resting-state and task data. The
preprocessing of the HCP resting-state and task datasets is described in detail on the HCP web-
site. Briefly, the data is preprocessed using the HCP pipeline, which is using standardized
methods using FSL (FMRIB Software Library), FreeSurfer, and the Connectome Workbench
software (Glasser et al., 2013; Smith et al., 2013). This standard preprocessing included cor-
rection for spatial and gradient distortions and head motion, intensity normalization and bias
field removal, registration to the T1 weighted structural image, transformation to the 2-mm
Montreal Neurological Institute (MNI) space and using the FIX artefact removal procedure
(Navarro Schröder et al., 2015; Smith et al., 2013). The head motion parameters were
regressed out, and structured artefacts were removed by ICA + FIX processing (independent
component analysis followed by FMRIB’s ICA-based X-noiseifier (Griffanti et al., 2014; Salimi-
Khorshidi et al., 2014). Preprocessed time series of all grayordinates are in HCP CIFTI gray-
ordinates standard space and available in the surface-based CIFTI file for each participants for
resting-state and each of the seven tasks.
We used a custom-made MATLAB script using the ft_read_cifti function (Fieldtrip toolbox;
Oostenveld et al., 2011) to extract the average time series of all the grayordinates in each
region of the DK80 parcellation, which are defined in the HCP CIFTI grayordinates standard
space. The BOLD time series were filtered using a second-order Butterworth filter in the range
of 0.008–0.08 Hz.
UCLA preprocessing and extraction of functional time series in fMRI resting-state data. The prepro-
cessing of UCLA resting-state datasets is described in detail on the website and in the paper by
Gorgolewski and colleagues (Gorgolewski et al., 2017). Briefly, the preprocessing was per-
formed using FMRIPREP version 0.4.4 (https://fmriprep.readthedocs.io). This robust prepro-
cessing pipeline is based on the Nipype workflow engine7 and aims to combine different
implementations of various MR signal processing algorithms (from established software pack-
ages) to deliver a robust spatial normalization and nuisance estimation workflow.
Network Neuroscience
989
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The arrow of time of brain signals in cognition
Parcellations
For the analysis at the global and node level, all neuroimaging data was processed using the
DK80 standard parcellations (Deco et al., 2021b). Briefly, this was constructed using the
Mindboggle-modified Desikan–Killiany parcellation (Desikan et al., 2006) with a total of 62
cortical regions (31 regions per hemisphere) (Klein & Tourville, 2012). We added the 18 sub-
cortical regions, that is, nine regions per hemisphere: hippocampus, amygdala, subthalamic
nucleus (STN), globus pallidus internal segment (GPi), globus pallidus external segment (GPe),
putamen, caudate, nucleus accumbens, and thalamus. This provided a total of 80 regions in
the DK80 parcellation, also precisely defined in the common HCP CIFTI grayordinates stan-
dard space. For the analysis at the system level we also used the Schaefer500, where each
parcel is marked with the seven resting-state network in the Yeo7 parcellation (Schaefer
et al., 2018; Yeo et al., 2011).
TENET Deep-Learning Framework and Associated Methods
The TENET deep-learning framework is a general method that can use many types of data.
However, it is important that the data for training and generalisation are balanced between
different conditions. We used TENET to carry out two independent analyses using two different
datasets from HCP and UCLA, and below we show how we avoided any potential problems
with bias.
HCP Dataset
For the large-scale HCP dataset, we performed the reversibility and nonequilibrium analyses at
three different spatial scales: Global, system level, and local node level. For each scale, we
trained and analysed three different TENETs, one for each spatial scale. We used roughly
90% of the data for the training set (890 participants) and the remaining 10% for the test
and validation sets (100 participants). Note that the sliding windows used for training belong
to different participants than the ones used for test and validation, hence there are no overlap-
ping windows.
For the global level, the input patterns consisted of the windowed time series across the
whole brain in the DK80 parcellation, that is, each pattern consisted of 80 windowed time
series.
For the system level, the input patterns consisted of the windowed time series in the
Schaefer500 parcellation, that is, each parcel belonged to one of the seven Yeo resting-state
networks. We performed the analysis for the Schaefer500 parcels belonging to each of the
seven Yeo networks and thus obtained a measure of nonequilibrium for each Yeo network.
Finally, for the finest spatial scale at the node level, the input patterns consisted of the
windowed time series for one parcel in the DK80 parcellation, that is, each of the patterns
consisted of one windowed time series for that parcel. We performed this for each of the
80 parcels in the DK80 separately such that we obtained a measure of nonreversibility for each
brain region. For the analysis of the HCP data, we carried out these three spatial-scale analyses
for HCP rest and seven tasks, that is, a total of 24 different analyses.
The input to TENET consisted of sliding windows from the BOLD neuroimaging data from
all the participants, shortened to the shortest duration of a task (EMOTION task with 176 TRs),
always starting from the beginning. We generated these sliding windows with slightly different
parameters for the three types of analysis: for global and system-level analysis, we used a win-
dow size of 20 TRs (each lasting 0.72 sec), and for node-level analysis, we used a window size
Network Neuroscience
990
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The arrow of time of brain signals in cognition
of 150 TRs. These were then shifted forward until the end of the datasets (with a length of 3 TRs
for global and system-level analysis and 2 TRs for node-level analysis). In all cases, the param-
eters (window size and overlap) were found by varying them and finding the most robust
results for generalisation.
As an example of how we chose the two parameters for each level of analysis, Supporting
Information Table S1 shows the global-level analysis for all resting-state data in all participants,
the Accuracy and Information Loss for the Training set, and the Accuracy and Information Loss
for the Cross-validation Set using varying window sizes [10, 20, 30] and overlaps [1, 3, 6]
(both in TRs). In general, as can be seen, the best results were found for window sizes [10,
20], TRs, and overlaps [3, 6]. Our criteria were to take the smallest overlap and the largest
possible window size, hence why we chose a window size of 20 TRs with an overlap of 3
TRs. Nevertheless, the results are stable and robust within this range.
For each of the sliding windows, we generated two input patterns containing (1) forward
and (2) time-reversed backward-sliding windowed time series, and for the supervised
learning, we associated each pattern with the output class label of forward and backward,
respectively.
UCLA Dataset
For the analysis of the smaller scale UCLA dataset with coarser TR (2 sec) and shorter duration
(152 TRs), we carried out different spatial-scale analyses at the level of global and node level
for the resting-state data of control and three neuropsychiatric disorders (schizophrenia, bipo-
lar disorder, and ADHD). For this much smaller dataset of 261 participants, we used roughly
90% of the data for the training set (40 participants in each of the four conditions) and the
remaining data for the test and validation sets (i.e., 36 participants for the global and 35 par-
ticipants for node-level analysis). For the global analysis, we shuffled the data 500 times in
order to perform nonparametric significance testing (for the box plot in Figure 6A). For the
node analysis we shuffled the data 10 times and computed the average node level of nonre-
versibility. We do not provide statistics for the node analysis, and note that it is computation-
ally very expensive to add more shuffles. Nevertheless, we show the rendered node-level
results on a qualitative basis.
The input to TENET consisted of sliding windows from the BOLD neuroimaging data from
all the participants. We generated these sliding windows with slightly different parameters for
the two types of analysis: for global-level analysis, we used a windows size of 10 TRs of 2 sec
and for node-level analysis, we used a window size of 20 TRs. These were then shifted forward
(in steps of 1 TR) until the end of the dataset.
Training and Generalisation in Deep-Learning Network TENET
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
a
_
0
0
3
0
0
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
This data was then used with TENET for training and generalisation in the following way. After
training, we measure the reversibility of each pattern in the validation set. This is computed by
comparing the trained output for the forward and backward versions of each pattern. More
specifically, for all three spatial scales, we compute the level of reversibility, R(t), for a given
sliding window at time t by using the following equation:
(cid:2)
(cid:3)
oforward 2ð Þ − oforward 1ð Þ
R tð Þ ¼
Þ
þ obackward 1ð Þ − obackward 2ð Þ
ð
2
Here oforward (i) is the output i of the final output classification layer, when the forward pattern
is presented in the input layer. Similarly, obackward (i) is the output for node i of the final output
Network Neuroscience
991
The arrow of time of brain signals in cognition
classification layer, but now when a backward pattern is presented in the input layer. Given
that the forward and backward categories are associated with the outputs oforward = [0, 1] and
obackward = [1, 0], respectively, R(t) thus represents the degree of reversibility for a particular
sliding window at time t. If either the degrees of right classification of forward (oforward (2) −
oforward(1)) or backward (obackward (1) − obackward (2)) are smaller than 0, this means that the
classification is incorrect and in this case we set R(t ) = 0. The degree of reversibility R(t ) is
measuring the degree of nonreversibility at time t. We also report the mean and the standard
deviation over time, which corresponds to what is called the mean nonreversibility and the
standard deviation of nonreversibility (across time) in the results and figures.
For the architecture of TENET, we use the standard MATLAB architecture with an input layer
of size N * w, where N corresponds to the spatial scale (i.e., global N = 1, system level N = 7,
node level N = 80), and w is the size of the sliding window. This is followed by 10 fully con-
nected layers including (1) batch normalisation, which normalises a minibatch of data across
all observations for each channel independently, and (2) a nonlinearity operation implemented
using reluLayer, which performs a threshold operation to each element of the input, where any
value less than zero is set to zero. The dimensions of the 10 layers are [2048; 1024; 512; 256;
128; 64; 32; 16; 8; 4]. The final layer of TENET is a softmax classification layer of dimension
[2], corresponding the two possible output class labels (forward and backward).
For training TENET, we used the deep-learning algorithm ADAM, which is an algorithm for
first-order gradient-based optimization of stochastic objective functions, based on adaptive esti-
mates of lower order moments to attenuate the effects of noise as it is implemented in MATLAB
with the recommended default parameters (Kingma & Ba, 2014). These standard MATLAB train-
ing options are as follows: GradientDecayFactor: 0.9000, SquaredGradientDecayFactor:
0.9990, Epsilon: 1.0000e-08, InitialLearnRate: 1.0000e-03, LearnRateSchedule: ’none’, Learn-
RateDropFactor: 0.1000, LearnRateDropPeriod: 10, L2Regularization: 1.0000e-04, Gradient-
ThresholdMethod: ’l2norm’, GradientThreshold: Inf, MaxEpochs: 10, MiniBatchSize: 128,
Verbose: 1, VerboseFrequency: 50, ValidationData: [ ],ValidationFrequency: 30, Validation-
Patience: Inf. Shuffle: ’once’, CheckpointPath: “, ExecutionEnvironment: ’auto’, WorkerLoad: [],
OutputFcn: [],Plots: ’none’, SequenceLength: ’longest’, SequencePaddingValue: 0, Sequence-
PaddingDirection: ’right’, DispatchInBackground: 0, ResetInputNormalization: 1.
Measuring the Breaking of Detailed Balance of the System
We wanted to test if nonreversibility is associated with the breaking of detailed balance of
a system. In order to show the increase in nonbalance in nonequilibrium, we decided to
characterise the level of asymmetry in the interaction as expressed by the shifted correlation
matrix. Specifically, we selected patterns from sliding windows of low and high reversibility,
and computed the time-delayed functional connectivity matrix between all pairs of brain
regions, FC(τ)
FCij τð Þ ¼ < xi tð Þ; xj t þ τ
ð
Þ >
在哪里 <> indicates correlation over time of the fMRI time series x and we used τ = 3 (in TRs),
which was selected using the autocorrelation function, such that the chosen delay produced
sufficient decay. We computed the FCij (t) over all participants and all sliding windows for each
状况 (for HCP REST and the SOCIAL task, the most nonreversibility task). For the global
level of asymmetry, we computed for each sliding window the degree of asymmetry as the
mean value of the elements of the matrix: FCij (t) − FCij (t)时间, where the superscript T indicates
the transposition. For the node level of asymmetry, we first computed for each sliding window,
iFCij (t). The node level
jFCij (t), and the outcoming flow Fout (j ) =
the incoming flow Fin(我) =
磷
磷
网络神经科学
992
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
The arrow of time of brain signals in cognition
of asymmetry for node i is then given by <|Fout(i) − Fin(i)|>t, 在哪里 <>t indicates the average of
the node-level asymmetry over all sliding windows and participants. We render the change
between high and low levels of the node-level asymmetry.
Support Vector Machine for System-Level Classification
We used a SVM with Gaussian kernels as implemented in the MATLAB function fitcecoc. 这
function returns a full, trained, multiclass, error-correcting output codes (ECOC) 模型. 这是
achieved using the predictors in the input with class labels. The function uses K(K − 1)/2 binary
SVM models using the one-versus-one coding design, where we used K = 8 as the number of
unique class labels. 换句话说, the SVM had seven inputs (the Yeo resting-state networks)
corresponding to the output produced by the system-level TENET. The output was eight classes
corresponding to the conditions (rest and seven tasks). We used the output from the 100 HCP
participants used for generalisation, subdivided into 90% training and 10% 验证,
repeated and shuffled 100 次.
致谢
G. D. is supported Spanish national research project (ref. PID2019-105772GB-I00 MCIU AEI)
funded by the Spanish Ministry of Science, Innovation and Universities (MCIU), State Research
机构 (AEI); HBP SGA3 Human Brain Project Specific Grant Agreement 3 (grant agreement
不. 945539), funded by the EU H2020 FET Flagship programme; SGR Research Support Group
支持 (ref. 2017 SGR 1545), funded by the Catalan Agency for Management of University
and Research Grants (AGAUR); Neurotwin Digital twins for model-driven noninvasive electri-
cal brain stimulation (grant agreement ID: 101017716) funded by the EU H2020 FET Proactive
programme; euSNN European School of Network Neuroscience (grant agreement ID: 860563)
funded by the EU H2020 MSCA-ITN Innovative Training Networks; CECH The Emerging
Human Brain Cluster (Id. 001-磷-001682) within the framework of the European Research
Development Fund Operational Program of Catalonia 2014–2020; Brain-Connects: Brain
Connectivity during Stroke Recovery and Rehabilitation (id. 201725.33) funded by the Funda-
cio La Marato TV3; Corticity, FLAG–ERA JTC 2017 (ref. PCI2018-092891) funded by the Span-
ish Ministry of Science, Innovation and Universities (MCIU), State Research Agency (AEI).
MLK is supported by the Center for Music in the Brain, funded by the Danish National
Research Foundation (DNRF117), and Centre for Eudaimonia and Human Flourishing at
Linacre College funded by the Pettit and Carlsberg Foundations.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00300.
The multimodal neuroimaging data are freely available from HCP. The UCLA data are freely
available from the Consortium for Neuropsychiatric Phenomics. The code used to run the
analysis is available on GitHub (https://github.com/decolab/tenet).
作者贡献
Gustavo Deco: 概念化; 数据管理; 形式分析; 资金获取; Inves-
tigation; 方法; 项目管理; 资源; 软件; 监督; 验证;
可视化; Writing – original draft; 写作——复习 & 编辑. Yonathan Sanz Perl:
方法; 软件; Writing – original draft; 写作——复习 & 编辑. Laura de la
Fuente: 方法. Jacobo D. Sitt: 方法. Thomas Yeo: 方法; Writing –
网络神经科学
993
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
The arrow of time of brain signals in cognition
review & 编辑. Enzo Tagliazucchi: 概念化; 方法; 写作——复习 &
编辑. Morten Kringelbach: 概念化; 数据管理; 形式分析; 资金
acquisition; 调查; 方法; 项目管理; 资源; 软件; Super-
想象; 验证; 可视化; Writing – original draft; 写作——复习 & 编辑.
资金信息
Morten Kringelbach, Danmarks Grundforskningsfond (https://dx.doi.org/10.13039
/501100001732), 奖项ID: DNRF117.
参考
Adolphs, 右. (2016). Human lesion studies in the 21st century.
神经元, 90(6), 1151–1153. https://doi.org/10.1016/j.neuron
.2016.05.014, 考研: 27311080
Ao, 磷. (2008). Emerging of stochastic dynamical equalities and
steady state thermodynamics from Darwinian dynamics. 康姆-
munications in Theoretical Physics, 49(5), 1073–1090. https://
doi.org/10.1088/0253-6102/49/5/01, 考研: 21949462
Arnatkeviciute, A。, Fulcher, 乙. D ., & 假如, A. (2019). A practical
guide to linking brain-wide gene expression and neuroimaging
数据. 神经影像, 189, 353–367. https://doi.org/10.1016/j
.neuroimage.2019.01.011, 考研: 30648605
Attwell, D ., & Laughlin, S. 乙. (2001). An energy budget for signaling
in the grey matter of the brain. Journal of Cerebral Blood Flow &
Metabolism, 21(10), 1133–1145. https://doi.org/10.1097
/00004647-200110000-00001, 考研: 11598490
Barch, D. M。, 伯吉斯, G. C。, Harms, 中号. P。, 彼得森, S. E.,
施拉加尔, 乙. L。, 科尔贝塔, M。, Glasser, 中号. F。, Curtiss, S。, Dixit,
S。, Feldt, C。, Nolan, D ., Bryant, E., Hartley, T。, Footer, 奥。, Bjork,
J. M。, Poldrack, R。, 史密斯, S。, Johansen-Berg, H。, 斯奈德, A. Z。, &
Van Essen, D. C. (2013). Function in the human connectome:
Task-fMRI and individual differences in behavior. 神经影像,
80, 169–189. https://doi.org/10.1016/j.neuroimage.2013.05
.033, 考研: 23684877
Binder, J. R。, 总的, 瓦. L。, Allendorfer, J. B., Bonilha, L。, Chapin, J。,
爱德华兹, J. C。, Grabowski, 时间. J。, Langfitt, J. T。, Loring, D. W.,
Lowe, 中号. J。, Koenig, K., 摩根, 磷. S。, Ojemann, J. G。, Rorden,
C。, Szaflarski, J. P。, Tivarus, 中号. E., & Weaver, K. 乙. (2011). 测绘
anterior temporal lobe language areas with fMRI: A multicenter
normative study. 神经影像, 54(2), 1465–1475. https://doi.org
/10.1016/j.neuroimage.2010.09.048, 考研: 20884358
Biswal, B., Yetkin, F。, Haughton, 五、, & Hyde, J. (1995). Functional
connectivity in the motor cortex of resting human brain using
echo-planar MRI. Magnetic Resonance in Medicine, 34(4), 537–541.
https://doi.org/10.1002/mrm.1910340409, 考研: 8524021
Biswal, 乙. B., Mennes, M。, Zuo, X.-N., Gohel, S。, 凯莉, C。, 史密斯,
S. M。, 贝克曼, C. F。, Adelstein, J. S。, 巴克纳, 右. L。,
Colcombe, S。, Dogonowski, A.-M., Ernst, M。, Fair, D ., Hampson,
M。, Hoptman, 中号. J。, Hyde, J. S。, Kiviniemi, V. J。, Kötter, R。, 李,
S.-J., 林, C.-P., Lowe, 中号. J。, Mackay, C。, Madden, D. J。, Madsen,
K. H。, 马古利斯, D. S。, Mayberg, H. S。, 麦克马洪, K., Monk,
C. S。, Mostofsky, S. H。, Nagel, 乙. J。, Pekar, J. J。, Peltier, S. J。,
彼得森, S. E., Riedl, 五、, Rombouts, S. A. 右. B., Rypma, B.,
施拉加尔, 乙. L。, 施密特, S。, Seidler, 右. D ., Siegle, G. J。, Sorg,
C。, Teng, G.-J., Veijola, J。, 狂野的召唤者, A。, Walter, M。, 王, L。,
翁, X.-C., Whitfield-Gabrieli, S。, 威廉森, P。, Windischberger,
C。, Zang, Y.-F., 张, H.-Y., Castellanos, F. X。, & Milham, 中号. 磷.
(2010). Toward discovery science of human brain function. Pro-
ceedings of the National Academy of Sciences of the United
States of America, 107(10), 4734–4739. https://doi.org/10.1073
/普纳斯.0911855107, 考研: 20176931
Bolt, T。, 安德森, 中号. L。, & Uddin, L. 问. (2018). Beyond the
evoked/intrinsic neural process dichotomy. Network Neurosci-
恩斯, 2(1), 1–22. https://doi.org/10.1162/ NETN_a_00028,
考研: 29911670
Brovelli, A。, Chicharro, D ., Badier, J. M。, 王, H。, & Jirsa, V.
(2015). Characterization of cortical networks and corticocortical
functional connectivity mediating arbitrary visuomotor map-
平. 神经科学杂志, 35(37), 12643–12658. https://
doi.org/10.1523/ JNEUROSCI.4892-14.2015, 考研:
26377456
巴克纳, 右. L。, & 克里宁, F. 中号. (2013). The evolution of distributed
association networks in the human brain. Trends in Cognitive
科学, 17(12), 648–665. https://doi.org/10.1016/j.tics.2013
.09.017, 考研: 24210963
Burt, J. B., Demirtas, M。, Eckner, 瓦. J。, Navejar, 氮. M。, 吉, J. L。,
马丁, 瓦. J。, Bernacchia, A。, Anticevic, A。, & 穆雷, J. D.
(2018). Hierarchy of transcriptomic specialization across human
cortex captured by structural neuroimaging topography. 自然
神经科学, 21(9), 1251–1259. https://doi.org/10.1038
/s41593-018-0195-0, 考研: 30082915
Carnot, S. (1824). Reflections on the motive power of fire, and on
machines fitted to develop that power. 巴黎: Bachelier.
克拉克, D. D ., & Sokoloff, L. (1999). Circulation and energy metab-
olism of the brain. 在乙. 瓦. Agranoff & G. J. 西格尔 (编辑。), Basic
neurochemistry. Molecular, cellular and medical aspects (6第三版。,
PP. 637–670). Lippincott-Raven.
Clausius, 右. (1865). Ueber verschiedene für die Anwendung
bequeme Formen der Hauptgleichungen der mechanischen
Wärmetheorie ( Vorgetragen in der naturforsch. Gesellschaft zu
Zürich den 24. 四月 1865). Annalen der Physik und Chemie,
201(7), 353–400. https://doi.org/10.1002/andp.18652010702
Collin, D ., Ritort, F。, Jarzynski, C。, 史密斯, S. B., Tinoco, 我。, &
Bustamante, C. (2005). Verification of the Crooks fluctuation
theorem and recovery of RNA folding free energies. 自然,
437(7056), 231–234. https://doi.org/10.1038/nature04061,
考研: 16148928
Crooks, G. 乙. (1998). Nonequilibrium measurements of free energy
differences for microscopically reversible Markovian systems.
网络神经科学
994
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
The arrow of time of brain signals in cognition
Journal of Statistical Physics, 90, 1481–1487. https://doi.org/10
.1023/A:1023208217925
de la Fuente, L. A。, Zamberlan, F。, Bocaccio, H。, 克林格尔巴赫, M。,
德科, G。, Sanz Perl, Y。, Pallavicini, C。, & Tagliazucchi, 乙. (2023).
Temporal irreversibility of neural dynamics as a signature of con-
sciousness. 大脑皮层, 33(5), 1856–1865. https://doi.org/10
.1093/cercor/bhac177, 考研: 35512291
德科, G。, 克林格尔巴赫, M。, Arnatkevičiūtė, A。, Oldham, S。,
Sabaroedin, K., Rogasch, 氮. C。, Aquino, K., & 假如, A.
(2021A). Dynamical consequences of regional heterogeneity in
the brain’s transcriptional landscape. Science Advances, 7(29),
eabf4752. https://doi.org/10.1126/sciadv.abf4752, 考研:
34261652
德科, G。, & 克林格尔巴赫, 中号. L. (2020). Turbulent-like dynamics in
the human brain. Cell Reports, 33(10), 108471. https://doi.org/10
.1016/j.celrep.2020.108471, 考研: 33296654
德科, G。, Sanz Perl, Y。, Tagliazucchi, E., & 克林格尔巴赫, 中号. L.
(2022). The INSIDEOUT framework provides precise signatures
of the balance of intrinsic and extrinsic dynamics in brain states.
Communications Biology, 5(1), 572. https://doi.org/10.1038
/s42003-022-03505-7, 考研: 35688893
德科, G。, Tagliazucchi, E., Laufs, H。, 桑胡安, A。, & 克林格尔巴赫,
中号. L. (2017). Novel intrinsic ignition method measuring
local-global integration characterises wakefulness and deep
睡觉. eNeuro, 4(5), ENEURO.0106-0117.2017. https://doi.org
/10.1523/ENEURO.0106-17.2017, 考研: 28966977
德科, G。, Vidaurre, D ., & 克林格尔巴赫, 中号. L. (2021乙). Revisiting
the global workspace orchestrating the hierarchical organisation
of the human brain. Nature Human Behaviour, 5(4), 497–511.
https://doi.org/10.1038/s41562-020-01003-6, 考研:
33398141
德阿内, S。, Kerszberg, M。, & 尚古克斯, J. 磷. (1998). A neuronal
model of a global workspace in effortful cognitive tasks. Proceed-
ings of the National Academy of Sciences of the United States of
美国, 95(24), 14529–14534. https://doi.org/10.1073/pnas.95
.24.14529, 考研: 9826734
Desikan, 右. S。, 硒, F。, 菲舍尔, B., 奎因, 乙. T。, Dickerson, 乙. C。,
Blacker, D ., 巴克纳, 右. L。, 戴尔, A. M。, Maguire, 右. P。, 海曼,
乙. T。, 阿尔伯特, 中号. S。, & 基利亚尼, 右. J. (2006). An automated labeling
system for subdividing the human cerebral cortex on MRI scans
into gyral based regions of interest. 神经影像, 31(3), 968–980.
https://doi.org/10.1016/j.neuroimage.2006.01.021, 考研:
16530430
Drobyshevsky, A。, Baumann, S. B., & 施耐德, 瓦. (2006). A rapid
fMRI task battery for mapping of visual, motor, cognitive, 和
emotional function. 神经影像, 31(2), 732–744. https://土井
.org/10.1016/j.neuroimage.2005.12.016, 考研: 16488627
Eddington, A. S. (1928). The nature of the physical world. 伦敦:
Macmillan. https://doi.org/10.5962/bhl.title.5859
冯, 乙. H。, & Crooks, G. 乙. (2008). Length of time’s arrow. Physical
Review Letters, 101(9), 090602. https://doi.org/10.1103
/PhysRevLett.101.090602, 考研: 18851595
假如, A。, Arnatkeviciute, A。, & Fulcher, 乙. D. (2019). Bridging the
gap between connectome and transcriptome. Trends in Cognitive
科学, 23(1), 34–50. https://doi.org/10.1016/j.tics.2018.10
.005, 考研: 30455082
Frässle, S。, Lomakina, 乙. 我。, Razi, A。, 弗里斯顿, K. J。, Buhmann, J. M。, &
Stephan, K. 乙. (2017). Regression DCM for fMRI. 神经影像,
155, 406–421. https://doi.org/10.1016/j.neuroimage.2017.02
.090, 考研: 28259780
Frisch, U. (1995). Turbulence: The legacy of A. 氮. Kolmogorov.
剑桥, 英国: 剑桥大学出版社. https://doi.org/10
.1017/CBO9781139170666
弗里斯顿, K. J。, Harrison, L。, & 一分钱, 瓦. (2003). Dynamic causal
modelling. 神经影像, 19(4), 1273–1302. https://doi.org/10
.1016/S1053-8119(03)00202-7, 考研: 12948688
Furman, D. J。, 汉密尔顿, J. P。, & Gotlib, 我. H. (2011). Frontostriatal
functional connectivity in major depressive disorder. Biology of
Mood & Anxiety Disorders, 1(1), 11. https://doi.org/10.1186
/2045-5380-1-11, 考研: 22737995
Glasser, 中号. F。, 索蒂罗普洛斯, S. N。, Wilson, J. A。, Coalson, 时间. S。,
菲舍尔, B., 安德森, J. L。, 徐, J。, Jbabdi, S。, 韦伯斯特, M。,
波利梅尼, J. R。, Van Essen, D. C。, & 詹金森, M。, 为了
WU-Minn HCP Consortium. (2013). The minimal preprocessing
pipelines for the Human Connectome Project. 神经影像 80,
105–124. https://doi.org/10.1016/j.neuroimage.2013.04.127,
考研: 23668970
Gnesotto, F. S。, Mura, F。, Gladrow, J。, & Broedersz, C. 磷. (2018).
Broken detailed balance and non-equilibrium dynamics in living
系统: A review. Reports on Progress in Physics, 81(6), 066601.
https://doi.org/10.1088/1361-6633/aab3ed, 考研: 29504517
Gorgolewski, K. J。, Durnez, J。, & Poldrack, 右. A. (2017). Prepro-
cessed Consortium for Neuropsychiatric Phenomics dataset.
F1000Research, 6, 1262. https://doi.org/10.12688/f1000research
.11964.2, 考研: 29152222
Griffanti, L。, Salimi-Khorshidi, G。, 贝克曼, C. F。, 奥尔巴赫, 乙. J。,
Douaud, G。, Sexton, C. E., Zsoldos, E., Ebmeier, K. P。, Filippini,
N。, Mackay, C. E., Moeller, S。, 徐, J。, Yacoub, E., Baselli, G。,
Ugurbil, K., 磨坊主, K. L。, & 史密斯, S. 中号. (2014). ICA-based arte-
fact removal and accelerated fMRI acquisition for improved rest-
ing state network imaging. 神经影像, 95, 232–247. https://土井
.org/10.1016/j.neuroimage.2014.03.034, 考研: 24657355
Hawrylycz, 中号. J。, Lein, 乙. S。, Guillozet-Bongaarts, A. L。, 沉, 乙. H。,
的, L。, 磨坊主, J. A。, van de Lagemaat, L. N。, 史密斯, K. A。, Ebbert,
A。, Riley, Z. L。, Abajian, C。, 贝克曼, C. F。, Bernard, A。,
Bertagnolli, D ., Boe, A. F。, Cartagena, 磷. M。, Chakravarty,
中号. M。, Chapin, M。, Chong, J。, Dalley, 右. A。, David Daly, B.,
当, C。, Datta, S。, Dee, N。, Dolbeare, 时间. A。, Faber, 五、, 冯,
D ., Fowler, D. R。, Goldy, J。, Gregor, 乙. W., Haradon, Z。, Haynor,
D. R。, Hohmann, J. G。, Horvath, S。, 霍华德, 右. E., Jeromin, A。,
Jochim, J. M。, Kinnunen, M。, Lau, C。, Lazarz, 乙. T。, 李, C。,
柠檬, 时间. A。, 李, L。, 李, Y。, 莫里斯, J. A。, Overly, C. C。, 派克,
磷. D ., 招架, S. E., Reding, M。, Royall, J. J。, Schulkin, J。, Sequeira,
磷. A。, Slaughterbeck, C. R。, 史密斯, S. C。, Sodt, A. J。, Sunkin, S. M。,
斯旺森, 乙. E., Vawter, 中号. P。, 威廉姆斯, D ., Wohnoutka, P。,
Zielke, H. R。, Geschwind, D. H。, Hof, 磷. R。, 史密斯, S. M。, 科赫,
C。, 授予, S. G. N。, & 琼斯, A. 右. (2012). An anatomically com-
prehensive atlas of the adult human brain transcriptome. 自然,
489(7416), 391–399. https://doi.org/10.1038/nature11405,
考研: 22996553
他, 乙. J. (2013). Spontaneous and task-evoked brain activity nega-
tively interact. 神经科学杂志, 33(11), 4672–4682.
https://doi.org/10.1523/ JNEUROSCI.2922-12.2013, 考研:
23486941
Jarzynski, C. (2011). Equalities and inequalities: Irreversibility and
the second law of thermodynamics at the nanoscale. Annual
网络神经科学
995
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
The arrow of time of brain signals in cognition
Review of Condensed Matter Physics, 2, 329–351. https://doi.org
/10.1146/annurev-conmatphys-062910-140506
Joglekar, 中号. R。, Mejias, J. F。, 哪个, G. R。, & 王, X.-J. (2018).
Inter-areal balanced amplification enhances signal propagation
in a large-scale circuit model of the primate cortex. 神经元,
98(1), 222–234. https://doi.org/10.1016/j.neuron.2018.02.031,
考研: 29576389
Kingma, D. P。, & Ba, J. (2014). 亚当: A method for stochastic
优化. arXiv:1412.6980. https://doi.org/10.48550/arXiv
.1412.6980
克莱因, A。, & Tourville, J. (2012). 101 labeled brain images and a
consistent human cortical labeling protocol. Frontiers in Neuro-
科学, 6, 171. https://doi.org/10.3389/fnins.2012.00171,
考研: 23227001
Konrad, K., & Eickhoff, S. 乙. (2010). Is the ADHD brain wired dif-
ferently? A review on structural and functional connectivity in
attention deficit hyperactivity disorder. 人脑图谱,
31(6), 904–916. https://doi.org/10.1002/hbm.21058, 考研:
20496381
LeDoux, J. 乙. (2000). Emotion circuits in the brain. Annual Review
of Neuroscience, 23, 155–184. https://doi.org/10.1146/annurev
.neuro.23.1.155, 考研: 10845062
林恩, C. W., Cornblath, 乙. J。, Papadopoulos, L。, Bertolero, 中号. A。, &
Bassett, D. S. (2021). Broken detailed balance and entropy
production in the human brain. 国家会议录
美国科学院, 118(47),
e2109889118. https://doi.org/10.1073/pnas.2109889118,
考研: 34789565
Magistretti, 磷. J。, Pellerin, L。, Rothman, D. L。, & 舒尔曼, 右. G. (1999).
Energy on demand. 科学, 283(5401), 496–497. https://doi.org
/10.1126/science.283.5401.496, 考研: 9988650
Maragakis, P。, Ritort, F。, Bustamante, C。, Karplus, M。, & Crooks,
G. 乙. (2008). Bayesian estimates of free energies from nonequi-
librium work data in the presence of instrument noise. The Jour-
nal of Chemical Physics, 129(2), 024102. https://doi.org/10.1063
/1.2937892, 考研: 18624511
马库斯, G. (2018). 深度学习: A critical appraisal. arXiv:1801
.00631. https://doi.org/10.48550/arXiv.1801.00631
马古利斯, D. S。, 戈什, S. S。, Goulas, A。, Falkiewicz, M。,
Huntenburg, J. M。, Langs, G。, Bezgin, G。, Eickhoff, S. B.,
Castellanos, F. X。, Petrides, M。, Jefferies, E., & Smallwood, J.
(2016). Situating the default-mode network along a principal gra-
dient of macroscale cortical organization. 诉讼程序
美国国家科学院,
113(44), 12574–12579. https://doi.org/10.1073/pnas.1608282113,
考研: 27791099
Mashour, G. A。, 罗尔夫塞玛, P。, 尚古克斯, J.-P., & 德阿内, S.
(2020). Conscious processing and the global neuronal work-
space hypothesis. 神经元, 105(5), 776–798. https://doi.org/10
.1016/j.neuron.2020.01.026, 考研: 32135090
Menon, 五、, & Uddin, L. 问. (2010). Saliency, switching, 注意力
and control: A network model of insula function. Brain Structure
& Function, 214(5–6), 655–667. https://doi.org/10.1007/s00429
-010-0262-0, 考研: 20512370
墨菲, C。, Jefferies, E., Rueschemeyer, S. A。, Sormaz, M。, 王,
H. T。, 马古利斯, D. S。, & Smallwood, J. (2018). Distant from
输入: Evidence of regions within the default mode network
supporting perceptually-decoupled and conceptually-guided
认识. 神经影像, 171, 393–401. https://doi.org/10.1016/j
.neuroimage.2018.01.017, 考研: 29339310
墨菲, C。, 王, H. T。, Konu, D ., Lowndes, R。, 马古利斯, D. S。,
Jefferies, E., & Smallwood, J. (2019). Modes of operation: A
topographic neural gradient supporting stimulus dependent
and independent cognition. 神经影像, 186, 487–496.
https://doi.org/10.1016/j.neuroimage.2018.11.009, 考研:
30447291
穆雷, 乙. A。, & Rudebeck, 磷. H. (2018). Specializations for
reward-guided decision-making in the primate ventral prefrontal
cortex. 自然评论神经科学, 19(7), 404–417. https://
doi.org/10.1038/s41583-018-0013-4, 考研: 29795133
Navarro Schröder, T。, Haak, K. 五、, Zaragoza Jimenez, 氮. 我。,
贝克曼, C. F。, & Doeller, C. F. (2015). Functional topography
of the human entorhinal cortex. 电子生活, 4, e06738. https://doi.org
/10.7554/电子生活.06738, 考研: 26052748
Oostenveld, R。, 薯条, P。, 马里斯, E., & 锄, J. 中号. (2011). Field-
Trip: Open source software for advanced analysis of MEG, EEG,
and invasive electrophysiological data. Computational Intelli-
gence and Neuroscience, 2011, 156869. https://doi.org/10
.1155/2011/156869, 考研: 21253357
Palus, 中号. (1996). Nonlinearity in normal human EEG: Cycles, TEM-
poral asymmetry, nonstationarity and randomness, not chaos.
Biological Cybernetics, 75(5), 389–396. https://doi.org/10.1007
/s004220050304, 考研: 8983161
Parrondo, J. 中号. R。, Van den Broeck, C。, & Kawai, 右. (2009). Entropy
production and the arrow of time. New Journal of Physics, 11,
073008. https://doi.org/10.1088/1367-2630/11/7/073008
Pearl, J. (2009). Causality: 楷模, 推理, and inference. 凸轮-
桥, 英国: 剑桥大学出版社. https://doi.org/10.1017
/CBO9780511803161
Pereda, E., Quiroga, 右. Q., & Bhattacharya, J. (2005). Nonlinear
multivariate analysis of neurophysiological signals. Progress in
Neurobiology, 77(1–2), 1–37. https://doi.org/10.1016/j.pneurobio
.2005.10.003, 考研: 16289760
Poincaré, H. (1890). Sur le problème des trois corps et les équations
de la dynamique. Acta Mathematica, 13, A3–A270.
Poldrack, 右. A. (2012). The future of fMRI in cognitive neurosci-
恩斯. 神经影像, 62, 1216–1220. https://doi.org/10.1016/j
.neuroimage.2011.08.007, 考研: 21856431
Poldrack, 右. A。, Congdon, E., Triplett, W., Gorgolewski, K. J。,
Karlsgodt, K. H。, Mumford, J. A。, Sabb, F. W., Freimer, 氮. B.,
伦敦, 乙. D ., Cannon, 时间. D ., & Bilder, 右. 中号. (2016). A
phenome-wide examination of neural and cognitive function.
Scientific Data, 3, 160110. https://doi.org/10.1038/sdata.2016
.110, 考研: 27922632
Ponce-Alvarez, A。, 他, 乙. J。, 哈格曼, P。, & 德科, G. (2015).
Task-driven activity reduces the cortical activity space of the
脑: Experiment and whole-brain modeling. PLoS Computa-
tional Biology, 11(8), e1004445. https://doi.org/10.1371/journal
.pcbi.1004445, 考研: 26317432
Prando, G。, Zorzi, M。, Bertoldo, A。, 科尔贝塔, M。, Zorzi, M。, &
Chiuso, A. (2020). Sparse DCM for whole-brain effective connec-
tivity from resting-state fMRI data. 神经影像, 208, 116367.
https://doi.org/10.1016/j.neuroimage.2019.116367, 考研:
31812714
Quiroga, 右. Q., Arnhold, J。, & Grassberger, 磷. (2000). 学习
driver-response relationships from synchronization patterns.
网络神经科学
996
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
The arrow of time of brain signals in cognition
Physical Review E: Statistical Physics, Plasmas, Fluids, 和
Related Interdisciplinary Topics, 61(5 Pt A), 5142–5148. https://
doi.org/10.1103/PhysRevE.61.5142, 考研: 11031559
Quiroga, 右. Q., Kreuz, T。, & Grassberger, 磷. (2002). Event synchro-
nization: A simple and fast method to measure synchronicity and
time delay patterns. Physical Review E: Statistical, Nonlinear, 和
Soft Matter Physics, 66(4 Pt 1), 041904. https://doi.org/10.1103
/PhysRevE.66.041904, 考研: 12443232
Raichle, 中号. 乙. (2006). The brainʼs dark energy. 科学, 314(5803),
1249–1250. https://doi.org/10.1126/science.1134405, 考研:
17124311
Raichle, 中号. 乙. (2010). Two views of brain function. 趋势
认知科学, 14(4), 180–190. https://doi.org/10.1016/j
.tics.2010.01.008, 考研: 20206576
Raichle, 中号. E., MacLeod, A. M。, 斯奈德, A. Z。, 权力, 瓦. J。,
Gusnard, D. A。, & 舒尔曼, G. L. (2001). A default mode of
brain function. 美国国家科学院院刊
美利坚合众国, 98(2), 676–682. https://doi.org
/10.1073/pnas.98.2.676, 考研: 11209064
Salimi-Khorshidi, G。, Douaud, G。, 贝克曼, C. F。, Glasser, 中号. F。,
Griffanti, L。, & 史密斯, S. 中号. (2014). Automatic denoising of
functional MRI data: Combining independent component
analysis and hierarchical fusion of classifiers. 神经影像, 90,
449–468. https://doi.org/10.1016/j.neuroimage.2013.11.046,
考研: 24389422
Sanz Perl, Y。, Bocaccio, H。, Pérez-Ipiña, 我。, Laureys, S。, Laufs, H。,
克林格尔巴赫, 中号. L。, 德科, G。, & Tagliazucchi, 乙. (2021). Non-
equilibrium brain dynamics as a signature of consciousness.
Physical Review E, 104(1-1), 014411. https://doi.org/10.1103
/PhysRevE.104.014411, 考研: 34412335
Schaefer, A。, 孔, R。, Gordon, 乙. M。, 劳曼, 时间. 奥。, Zuo,
X. N。, Holmes, A. J。, Eickhoff, S. B., & 杨, 乙. 时间. 时间. (2018).
Local-Global Parcellation of the Human Cerebral Cortex from
Intrinsic Functional Connectivity MRI. 大脑皮层, 28(9),
3095–3114. https://doi.org/10.1093/cercor/ bhx179, 考研:
28981612
Schrödinger, 乙. (1944). What is life? The physical aspect of the
living cell. 剑桥, 英国: 剑桥大学出版社.
Seif, A。, Hafezi, M。, & Jarzynski, C. (2021). Machine learning the
thermodynamic arrow of time. Nature Physics, 17, 105–113.
https://doi.org/10.1038/s41567-020-1018-2
Sejnowski, 时间. J. (2018). The deep learning revolution. 剑桥,
嘛: 与新闻界. https://doi.org/10.7551/mitpress/11474.001
.0001
Sejnowski, 时间. J. (2020). The unreasonable effectiveness of deep
learning in artificial intelligence. 国家会议录
美国科学院, 117(48),
30033–30038. https://doi.org/10.1073/pnas.1907373117,
考研: 31992643
赛斯, A. K., Chorley, P。, & Barnett, L. C. (2013). Granger causality
analysis of fMRI BOLD signals is invariant to hemodynamic con-
volution but not downsampling. 神经影像, 65, 540–555.
https://doi.org/10.1016/j.neuroimage.2012.09.049, 考研:
23036449
Sheremet, A。, Qin, Y。, 肯尼迪, J. P。, 周, Y。, & Maurer, A. 磷. (2019).
Wave turbulence and energy cascade in the hippocampus. Fron-
tiers in Systems Neuroscience, 12, 62. https://doi.org/10.3389
/fnsys.2018.00062, 考研: 30662397
Sherrington, C. (1906). The integrative action of the nervous system.
新天堂, CT: 耶鲁大学出版社.
Shirts, 中号. R。, Bair, E., Hooker, G。, & Pande, V. S. (2003). Equilib-
rium free energies from nonequilibrium measurements using
maximum-likelihood methods. 物理评论快报, 91(14),
140601. https://doi.org/10.1103/ PhysRevLett.91.140601,
考研: 14611511
舒尔曼, G. L。, 科尔贝塔, M。, Fiez, J. A。, 巴克纳, 右. L。, 米津,
F. M。, Raichle, 中号. E., & 彼得森, S. 乙. (1997). Searching for
activations that generalize over tasks. 人脑图谱,
5(4), 317–322. https://doi.org/10.1002/(SICI)1097-0193(1997)
5:4<317::AID-HBM19>3.0.一氧化碳;2-A, 考研: 20408235
Silver, D ., Hubert, T。, Schrittwieser, J。, Antonoglou, 我。, Lai, M。, Guez,
A。, Lanctot, M。, Sifre, L。, Kumaran, D ., & Graepel, 时间. (2018). A
general reinforcement learning algorithm that masters chess, shogi,
and Go through self-play. 科学, 362(6419), 1140–1144. https://
doi.org/10.1126/science.aar6404, 考研: 30523106
Smallwood, J。, Bernhardt, 乙. C。, 水蛭, R。, Bzdok, D ., Jefferies, E.,
& 马古利斯, D. S. (2021). The default mode network in cogni-
的: A topographical perspective. 自然评论神经科学,
22(8), 503–513. https://doi.org/10.1038/s41583-021-00474-4,
考研: 34226715
史密斯, S. M。, 贝克曼, C. F。, 安德森, J。, 奥尔巴赫, 乙. J。,
Bijsterbosch, J。, Douaud, G。, Duff, E., 范伯格, D. A。, Griffanti,
L。, Harms, 中号. P。, 凯莉, M。, 劳曼, T。, 磨坊主, K. L。, Moeller, S。,
彼得森, S。, 力量, J。, Salimi-Khorshidi, G。, 斯奈德, A. Z。, Vu,
A. T。, 伍尔里奇, 中号. W., 徐, J。, Yacoub, E., Ugurbil, K., Van Essen,
D. C。, & Glasser, 中号. F。, for the WU-Minn HCP Consortium.
(2013). Resting-state fMRI in the Human Connectome Project.
N e u ro I m a g e , 8 0 , 1 4 4 – 1 6 8 . h t t p s : / / d o i . o rg / 1 0 . 1 0 1 6 / j
.neuroimage.2013.05.039, 考研: 23702415
Strogatz, S. H. (2018). Nonlinear dynamics and chaos with student
solutions manual: With applications to physics, 生物学, chemis-
尝试, 和工程. Boca Raton, FL: CRC Press. https://doi.org
/10.1201/9780429399640
Tagliazucchi, E., Siniatchkin, M。, Laufs, H。, & Chialvo, D. 右. (2016).
The voxel-wise functional connectome can be efficiently derived
from co-activations in a sparse spatio-temporal point-process.
Frontiers in Neuroscience, 10, 381. https://doi.org/10.3389/fnins
.2016.00381, 考研: 27601975
Turnbull, A。, 王, H. T。, 墨菲, C。, Ho, 氮. S. P。, 王, X。,
Sormaz, M。, Karapanagiotidis, T。, 水蛭, 右. M。, Bernhardt, B.,
马古利斯, D. S。, Vatansever, D ., Jefferies, E., & Smallwood, J.
(2019). Left dorsolateral prefrontal cortex supports context-
dependent prioritisation of off-task thought. Nature Communica-
系统蒸发散, 10(1), 3816. https://doi.org/10.1038/s41467-019-11764-y,
考研: 31444333
van den Heuvel, 中号. P。, & 假如, A. (2014). Brain networks in
schizophrenia. Neuropsychology Review, 24(1), 32–48. https://
doi.org/10.1007/s11065-014-9248-7, 考研: 24500505
van Vugt, B., Dagnino, B., Vartak, D ., Safaai, H。, Panzeri, S。,
德阿内, S。, & 罗尔夫塞玛, 磷. 右. (2018). The threshold for con-
scious report: Signal loss and response bias in visual and frontal
cortex. 科学, 360(6388), 537–542. https://doi.org/10.1126
/science.aar7186, 考研: 29567809
哪个, G. R。, & 王, X. J. (2020). Artificial neural networks for neu-
roscientists: A primer. 神经元, 107(6), 1048–1070. https://土井
.org/10.1016/j.neuron.2020.09.005, 考研: 32970997
网络神经科学
997
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
The arrow of time of brain signals in cognition
杨, 乙. T。, 克里宁, F. M。, 墓, J。, 肥皂, 中号. R。, 军队,
D ., 霍林斯黑德, M。, 罗夫曼, J. L。, 斯莫勒, J. W., 佐莱伊, L。,
波利梅尼, J. R。, 菲舍尔, B., 刘, H。, & 巴克纳, 右. L. (2011). 这
organization of the human cerebral cortex estimated by intrinsic
功能连接. 神经生理学杂志, 106(3),
1125–1165. https://doi.org/10.1152/jn.00338.2011, 考研:
21653723
Yuste, R。, MacLean, J. N。, 史密斯, J。, & Lansner, A. (2005). 皮质
as a central pattern generator. 自然评论神经科学,
6(6), 477–483. https://doi.org/10.1038/nrn1686, 考研:
15928717
Zanin, M。, Güntekin, B., Aktürk, T。, Hanoğlu, L。, & Papo, D. (2019).
Time irreversibility of resting-state activity in the healthy brain
and pathology. Frontiers in Physiology, 10, 1619. https://doi.org
/10.3389/fphys.2019.01619, 考研: 32038297
张, D ., & Raichle, 中号. 乙. (2010). Disease and the brain’s dark
活力. Nature Reviews Neurology, 6(1), 15–28. https://doi.org
/10.1038/nrneurol.2009.198, 考研: 20057496
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
7
3
9
6
6
2
1
5
4
8
1
9
n
e
n
_
A
_
0
0
3
0
0
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
网络神经科学
998