Neurobiological Systems for Lexical Representation
and Analysis in English
Mirjana Bozic1,2, Lorraine K. Tyler1, Li Su1,2, Cai Wingfield2,
and William D. Marslen-Wilson1,2
抽象的
■ Current research suggests that language comprehension
engages two joint but functionally distinguishable neurobiological
流程: a distributed bilateral system, which supports general
perceptual and interpretative processes underpinning speech
comprehension, and a left hemisphere (LH) frontotemporal
系统, selectively tuned to the processing of combinatorial
grammatical sequences, such as regularly inflected verbs in
英语 [Marslen-Wilson, 瓦. D ., & Tyler, L. K. Morphology, lan-
guage and the brain: The decompositional substrate for language
comprehension. Philosophical Transactions of the Royal Society:
Biological Sciences, 362, 823–836, 2007]. Here we investigated
how English derivationally complex words engage these systems,
asking whether they selectively activate the LH system in the
same way as inflections or whether they primarily engage the
bilateral system that support nondecompositional access. In an
fMRI study, we saw no evidence for selective activation of the
LH frontotemporal system, even for highly transparent forms like
bravely. 反而, a combination of univariate and multivariate
analyses revealed the engagement of a distributed bilateral
系统, modulated by factors of perceptual complexity and
semantic transparency. We discuss the implications for theories
of the processing and representation of English derivational
morphology and highlight the importance of neurobiological
constraints in understanding these processes. ■
介绍
A fundamental property of human language is its genera-
tive capacity, whereby an infinite range of communicative
meanings can be expressed using the combination and
recombination of a finite set of linguistic elements. 这里
we focus on combinatorial processes realized at lexical
and morphological levels, where different morphemes—
typically a stem morpheme (例如, sad, jump) and a bound
grammatical morpheme (例如, -内斯, -编辑 )—combine to
create new surface forms (sadness, jumped), 以及哪里
a basic distinction is made between inflectional and
derivational processes. Inflectional morphology subserves
primarily grammatical functions, whereas derivational
morphology is associated with the creation and representa-
tion of new words in the language. From a novel neuro-
biological perspective, which relates human language
functions to two distinct underlying brain systems (Bozic,
Tyler, Ives, Randall, & Marslen-Wilson, 2010; Marslen-
Wilson & Tyler, 2007), we ask whether derivational
morphology engages a specialized left hemisphere (LH)
系统, in the same way as inflectional morphology,
or whether it engages domain general bilateral systems
associated with nondecompositional access.
1University of Cambridge, 2MRC Cognition and Brain Sciences
Unit, 剑桥
Neurobiological Systems for
Language Comprehension
A substantial body of evidence suggests that spoken word
comprehension engages a network of bilateral temporal
and frontal brain regions (Bozic et al., 2010; Marslen-Wilson
& Tyler, 2007; Beeman, 2005; Tyler, Stamatakis, 邮政,
Randall, & Marslen-Wilson, 2005; Binder et al., 2000). 这
global network can be broken down into two comple-
mentary systems with differential functional roles: a bi-
hemispheric system, which supports sound-to-meaning
mapping and general perceptual demands associated with
word comprehension and an LH frontotemporal system,
supporting grammatical computations (Bozic et al., 2010;
Tyler & Marslen-Wilson, 2008; Marslen-Wilson & Tyler, 2007).
Support for this hypothesis comes from converging
results of neuroimaging and neuropsychological studies.
Contrasted with nonspeech acoustic baselines, spoken
words activate bilateral regions in the superior and middle
temporal lobes, which have long been associated with
access from the speech stream to representations of lexical
form and meaning (例如, 赖特, Stamatakis, & Tyler,
2012; Hickok & Poepell, 2007; Tyler et al., 2005; Binder
等人。, 2000). Consistent with a bilateral distribution of
these functions, patients with damage to left frontal and
superior temporal regions (and spared right hemisphere
equivalents) exhibit a relatively intact ability to recognise
simple spoken words (例如, 狗, 房子) and show semantic
priming effects comparable with those of healthy controls
© 2013 Massachusetts Institute of Technology Published under a Creative
Commons Attribution-NonCommercial 3.0 Unported (CC BY-NC 3.0) 执照
认知神经科学杂志 25:10, PP. 1678–1691
土井:10.1162/jocn_a_00420
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
哦
d
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
哦
9
C
6
n
5
_
4
A
/
_
j
0
哦
0
C
4
n
2
0
_
A
p
_
d
0
0
乙
4
y
2
G
0
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
/
t
.
F
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
(Longworth, Marslen-Wilson, Randall, & Tyler, 2005; Tyler
等人。, 2002). 此外, word recognition supported by
the right hemisphere is maintained even during acute LH
disruption (Hickok et al., 2008), suggesting that it is under-
pinned by existing bilaterally supported capacities and is
not a result of longer-term compensation due to neural
plasticity and functional reorganization. In terms of the
specific functions of this system, Bozic et al. (2010) 展示
that it is sensitive to variations in the perceptual com-
plexity of morphologically simple spoken words. The pre-
sence, 例如, of an onset-embedded stem (例如, clay
in claim) triggers competition and increases the pro-
cessing demands required for selection of the correct
candidate, modifying frontotemporal activity bilaterally.
合在一起, the existing evidence points to a bilateral
system that underpins the general perceptual demands
associated with whole-word, stem-based lexical access.
相比之下, a parallel set of results point to an LH network
selectively tuned to the processing of linguistically complex
序列, whose combinatorial structure requires parsing
and segmentation during language comprehension (比照.
Marslen-Wilson & Tyler, 2007). Neuroimaging data con-
sistently relate LH temporal, inferior frontal, and inferior
parietal regions to the processing of syntactic and mor-
phological complexity, where simple linguistic elements
combine into more complex morphosyntactic structures
(Hagoort, 2005; Shtyrov, Pihko, & Pulvermuller, 2005; Tyler
等人。, 2005; Friederici, Ruschemeyer, Hahne, & Fiebach,
2003). Converging evidence from neuropsychology and
neuroimaging reveals a close relationship between this
LH circuit and regular inflectional morphology. 英语-
speaking patients with left frontotemporal damage typically
exhibit long-term disruption of comprehension and pro-
duction of inflectionally complex words such as played
or jumped (Tyler et al., 2011; Longworth et al., 2005;
古德格拉斯, Christiansen, & 加拉格尔, 1993), where lexi-
cal access requires morpho-phonological parsing that
breaks these forms down into their component morphemes
({玩} + {-编辑}).
The neurocognitive properties of regular inflectional
morphology (although chiefly studied in English) 是
consistent with the defining properties of inflectional
morphology cross-linguistically (Bickel & Nichols, 2007;
Marslen-Wilson, 2007). 第一的, inflectional morphology does
不是, by definition, create new words with new meanings
that require new lexical entries. Inflectional variants like
cat and cats or walk and walked are not listed as separate
headwords in standard dictionaries, and the meaning of
these forms is fully predictable from the meaning of the
stem combined with the grammatical properties of the
inflection. 第二, inflections are responsive, in a regular
and predictable way, to the properties of the grammatical
environment in which they occur (例如, Bickel & Nichols,
2007; 安德森, 1992). This is clear, 例如, 在哪里
inflectional morphemes express agreement—as in the
third person singular {-s} for English verbs (he walks) 或者
when morphological case is used to express the grammat-
ical role of a noun as subject or object in case-marking
语言. The information carried by the inflection is
not just about the stem itself, but about the phrasal and
sentential interpretation to which that stem relates. 这
compositionality and contextual dependence of regularly
inflected forms is consistent with a neurocognitive model
where inflectional morphemes are stripped from their
stems early in processing and where the grammatical in-
formation they carry engages the combinatorial linguistic
mechanisms supported by LH peri-sylvian brain regions
(Marslen-Wilson & Tyler, 2007).
Derivationally complex forms, although also constructed
by combining a stem with a grammatical morpheme,
seem to have quite different linguistic properties. Whereas
inflectional morphemes result in different forms of the
same word, adjusted to the requirements of its current
phrasal and sentential environment, derivational processes
produce new words—in linguistic terms, new lexemes with
new lexical entries (Matthews, 1991)—whose meaning
and syntactic function is much more context independent.
The meaning of these words is less predictable from
the simple combination of the meaning of their constitu-
ent morphemes. This lack of compositionality—and the
range of variation from opaque forms like department
to transparent forms like bravely—has prompted long-
standing controversies in the psycholinguistic literature
as to whether derived words are represented and pro-
cessed with respect to their constituent morphemes or
as whole forms with no underlying representation of their
morphological structure (例如, Baayen, Milin, Filipovic-
Durdevic, Hendrix, & Marelli, 2011; Marslen-Wilson, Bozic,
& Randall, 2008; Longtin & 莫尼耶, 2005; Taft, 2004;
Clahsen, Sonnenstuhl, & Blevins, 2003; Bertram, Schreuder,
& Baayen, 2000; Rastle, 戴维斯, Marslen-Wilson, & Tyler, 2000;
Baayen, Dijkstra, & Schreuder, 1997; Schreuder & Baayen,
1995; Marslen-Wilson, Tyler, Waksler, & Older, 1994).
None of these accounts, 然而, systematically re-
late their hypotheses to the neural systems that underlie
language function more generally. Nor does the current
neuropsychological or neuroimaging literature provide
an easily interpretable or consistent view of derivational
流程, with studies varying in language (英语,
Italian, 德语, Finnish), input modality (visual or audi-
保守党), experimental tasks, and neuroimaging methods,
as well as looking at both comprehension and produc-
的 (例如, Leminen et al., 2011; Meinzer, Lahiri, Flaisch,
Hannemann, & Eulitz, 2009; 汉密尔顿 & Coslett, 2008;
Marangolo & Piras, 2008; Bozic, Marslen-Wilson, Stamatakis,
戴维斯, & Tyler, 2007; Vannest, Polk, & Lewis, 2005; 戴维斯,
莫尼耶, & Marslen-Wilson, 2004; Marslen-Wilson &
Tyler, 1998; Badecker & Caramazza, 1991).
English Derivational Morphology in Its
Neurobiological Context
Here we ask a specific, neurobiologically framed question
about the processing and representation of derivationally
Bozic et al.
1679
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
哦
d
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
哦
9
C
6
n
5
_
4
A
/
_
j
0
哦
0
C
4
n
2
0
_
A
p
_
d
0
0
乙
4
y
2
G
0
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
t
F
/
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
complex words in English. Do they exhibit the same
neurocognitive characteristics as regular inflected forms,
being processed decompositionally in terms of their com-
ponent morphemes, or are they are primarily processed as
stored whole forms, not requiring morpho-phonological
parsing and segmentation as part of the access process?
Decomposition, Transparency, and Productivity
If decompositional accounts are correct, then the scope of
decomposition is likely to be affected by two variables
thought to influence the representation and processing of
derived words. These are semantic transparency—how
far the meaning of the stem is preserved in the derived
form—and suffix productivity (例如, Bertram et al., 2000;
Marslen-Wilson et al., 1994). Psycholinguistically, 角色
of semantic transparency has been well established since
the work of Marslen-Wilson et al. (1994), showing that
there is strong cross-modal priming between semantically
transparent pairs like bravely/brave, but not between
semantically opaque words like archer/arch. Cross-modal
priming reflects the properties of central lexical represen-
tations and suggests that the representation of semantically
transparent forms preserves their underlying morpholog-
ical structure (Marslen-Wilson, 2007; Clahsen et al., 2003).
The second variable, suffix productivity, is a measure of
whether a suffix is currently in use to create new words in
the language (Plag, 2006; Baayen & Lieber, 1991; Bauer,
1983). Productive suffixes (-是, -内斯) are frequently used
to coin new words in modern English (例如, birther, hub-
内斯), compared with unproductive suffixes (-我知道了, -th),
which have generally fallen out of use. Marslen-Wilson,
Hare, and Older (1993) showed robust cross-modal prim-
ing between word pairs sharing a derivational suffix
(例如, toughness/darkness), but only when the suffix was
productive—indicating that productive suffixes are more
likely to be segmented and have separate, primable repre-
句子. 最近, Ford, 戴维斯, and Marslen-Wilson
(2010) used lexical decision to show that decompositional
effects are only obtained for derived words with produc-
tive suffixes. These considerations suggest that complex
words with productive suffixes are more likely to be stored
and processed in terms of their constituent morphemes.
By manipulating transparency and productivity, 我们可以
both evaluate their role in a neurobiological context and
construct stimulus sets where these factors are appropri-
ately controlled. Our first set of words (桌子 1) 包含
transparent derived forms with productive suffixes (例如,
bravely). These consist of a stem whose meaning is trans-
parently preserved in the whole form, combined with a
highly productive suffix: {-莱}, 例如, applies semi-
paradigmatically to almost all adjectives in the language.
These derivational forms are most similar to regular in-
flectional forms such as played and most likely to be
stored in a decompositional format. These are contrasted
with three other morphologically complex sets varying
in transparency and productivity, ranging from trans-
parent words with nonproductive suffixes (例如, warmth),
opaque words with productive suffixes (例如, archer), 到
opaque words with nonproductive suffixes (例如, breadth).
Two sets of morphologically simple words complete the
six conditions: words with a potential embedded stem
but no suffix (例如, 丑闻, where dal is not a suffix in
英语) and simple words, such as giraffe, with neither a
potential embedded stem nor a suffix.
We test this set of conditions in an fMRI study that
exactly parallels in methods and design the earlier
Bozic et al. (2010) 学习. This showed strongly selective
activation of the left-lateralized decompositional sys-
tem by regularly inflected forms such as played. If deriva-
tionally complex forms in English engage the same LH
系统, then we expect to see a similar pattern here. 如果
the likelihood of decomposition for derived words is tied
to semantic transparency and suffix productivity, 然后
selective LH activation will be seen most strongly for
the transparent and productive bravely set. 然而, 在
桌子 1. Stimulus Properties
Condition (例子)
Stem Properties
Suffix Properties
Embed
Stem
Relatedness
Pretest
COALS
Freq
Ratio
Suffix
生产率
Hapax
1. Transparent/productive (bravely) +S+M+P
2. Transparent/non-productive (warmth) +S+M−P
3. Opaque/productive (archer) +S−M+P
4. Opaque/nonproductive (breadth) +S−M−P
5. Stem only (丑闻) +S−M P0
6. Simple (giraffe) −S M0 P0
是的
是的
是的
是的
是的
不
7.7
7.5
2.2
1.7
1.6
n/a
0.24
0.26
0.06
0.03
0.02
n/a
1.9
2.7
2.1
1.9
1.8
n/a
是的
是的
是的
是的
不
不
Productive
Nonproductive
Productive
Nonproductive
n/a
n/a
0.09
0.02
0.07
0.03
n/a
n/a
+/−S marks presence or absence of an onset-embedded stem; +/−M marks whether the stem is related in meaning to the full form; +/−P marks
whether the suffix is productive. M0 and P0 are used in cases where these descriptions cannot be applied. Relatedness pretest = average pretest scores
of semantic relatedness between the whole word and the embedded stem (1–9); freq ratio = ratio of whole word/stem form log frequencies. COALS =
corpus-based semantic occurence metric; Hapax = corpus-based productivity metric (参见方法).
1680
认知神经科学杂志
体积 25, 数字 10
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
哦
d
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
哦
9
C
6
n
5
_
4
A
/
_
j
0
哦
0
C
4
n
2
0
_
A
p
_
d
0
0
乙
4
y
2
G
0
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
/
F
.
t
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
a stronger decompositional hypothesis, the simple pre-
sence of a potential stem and a suffix may be sufficient to
trigger decompositional processes. Exactly this is seen in
the visual domain, where masked priming reveals early
decomposition for all derived forms that meet the stem +
affix criterion, irrespective of transparency or productiv-
性 (Whiting, Shtryov, & Marslen-Wilson, under review;
Marslen-Wilson et al., 2008; Rastle et al., 2000). This also
holds for regular inflectional morphology in the auditory
domain, where the presence of phonological cues to a
potential past tense inflection generates L peri-sylvian acti-
vation not only for real inflected forms, such as played, 但
also for pseudoregular forms like trade or blend (Bozic
等人。, 2010; Tyler et al., 2005). If this holds for the deriva-
tional morphology, then the four morphologically complex
conditions should all show selective left frontotemporal
effects. This should contrast with morphologically simple
words like scandal, which have an embedded stem (scan)
but no derivational suffix, or like giraffe, with have neither
an embedded stem nor a suffix. Both of these sets should
engage the bilateral system without selective left inferior
frontal gyrus (LIFG) activation.
with stronger competition likely to engage bilateral IFG
(Zhuang, Tyler, Randall, Stamatakis, & Marslen-Wilson,
2012). The degree of semantic relatedness between the
full form and the onset-embedded stem—indexed here
as semantic transparency (+/−M)—could also be expected
to modulate the competition between the two forms
(Raposo, Moss, Stamatakis, & Tyler, 2006).
Data were analyzed using both conventional univariate
approaches and multivariate Representational Similarity
分析 (RSA; Kriegeskorte, 穆尔, & Bandettini, 2008).
The two methods provide complementary information.
Univariate methods allow us to identify how the overall
amount of activation per condition maps onto the lan-
guage processing network and its two systems, 与
activation in each condition reflecting the composite in-
fluences of linguistic, semantic, and perceptual variables.
Multivariate methods such as RSA are sensitive to the in-
formational patterning of neural activity, allowing us to
break down these overall effects and obtain specific data
about the role of different linguistic, semantic, 和每-
ceptual dimensions in the processing of simple and com-
plex spoken forms.
Whole-form Representations and
Perceptual Competition
On a whole-form account of the representation of derived
words in English—of which there are historically many
versions, from Butterworth (1983) to Baayen et al. (2011)—
these forms are neither stored nor accessed as sets of
separate morphemes. For spoken language, this means
that lexical access requires the sequential mapping of
the incoming input onto stored full form representations,
without an intervening process that breaks up the string
into component morphemes. In this respect, derived
stems like darkness will engage the same processing
mechanisms as morphologically simple stems like house,
which do not require preliminary parsing.
This makes two predictions for our six experimental
状况. The first is that all stimulus types, 从
transparent/productive bravely set to the morphologically
simple giraffe set, will engage the bilateral system and in
particular the posterior and middle temporal regions that
support stem-based access. No stimulus types should
selectively engage the left frontotemporal system (LIFG
and STG/MTG) because none of them should trigger de-
compositional processing. The second prediction is that
the relevant processing variables for these word forms will
not be their linguistic properties—whether they have a
productive suffix or not (+/−P)—but rather their percep-
tual processing complexity. Perceptual complexity is in-
creased by the presence of a competing onset-embedded
干 (+/−S), common to all derived and pseudo-derived
形式, as well as to monomorphemic forms like scandal.
Earlier studies show that the degree of perceptual com-
petition can be captured by the relative frequency of the
embedded stem and the whole form (Bozic et al., 2010),
方法
参加者
Eighteen right-handed (11 女性) native speakers
of British English, screened for neurological or devel-
opmental disorders, participated in the study. All gave
informed consent and were paid for their participation.
The study was approved by the Peterborough and Fenland
Ethical Committee.
Stimuli
There were six conditions with 40 words each (桌子 1).
Conditions 1–4 covaried linguistic and semantic variables
that are likely to affect morphological decomposability.
Conditions 5 和 6 provided morphologically simple con-
trol conditions. Condition 5 was made up of words with an
embedded pseudo-stem (例如, 丑闻) but no derivational
suffix, and Condition 6 of words with neither pseudostem
nor suffix (例如, giraffe).
Linguistic cues to decompositionality were based on
suffix productivity. This was initially determined following
Ford et al. (2010), Hay and Plag (2004), Bauer (1983), 和
Marchand (1969). To derive corpus-based values for suffix
生产率, we followed the formula used by Ford et al.
(2010) and Baayen and Lieber (1991). The probability
p that a suffix will be used to create new words is defined
as p = h/N, where h is the number of hapax legomena
(words with a given suffix that occur only once in the
语料库) and N is the total number of tokens for that affix.
This measure reflects the readiness with which a suffix
enters into new combinations and is thought to reflect its
synchronic productivity.
Bozic et al.
1681
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
哦
d
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
哦
9
C
6
n
5
_
4
A
/
_
j
0
哦
0
C
4
n
2
0
_
A
p
_
d
0
0
乙
4
y
2
G
0
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
/
t
F
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
语义学, stem-based aspects of decompositionality
were captured by manipulating the semantic transparency
of the relationship between the embedded stem and the
derived full form. Transparency was initially determined
by pretests, where participants rated word pairs on a
9-point scale, 和 1 being not related at all in meaning
和 9 being very related. Semantically unrelated pairs were
rated between 1 和 3, whereas related pairs were rated
之间 6 和 9. We also used a less subjective corpus-
based distance metric based on co-occurrence measures.
The semantic relatedness between the derived word and
its embedded stem (例如, bravely and brave) was esti-
mated using COALS co-occurrence statistics. This is a
vector-space method for deriving word meanings (Rohde,
Gonnerman, & 普劳特, 2005), where a large corpus of text is
used to derive the statistics of word co-occurrence, 哪个
are expressed as high-dimensional semantic vectors. 这
pairwise correlations between these vectors provide in-
formation about the degree to which the meaning of the
embedded stem is preserved in the derived form.
To estimate the perceptual competition between
the embedded stem and the whole form during lexical
使用权, we calculated the ratio of the log frequencies of
the whole form and the embedded stem. The average
value of perceptual competition did not differ significantly
across Conditions 1–5 ( p = .12).
All words and their embedded stems (where present)
were matched on word frequency ( WF) and lemma fre-
quency (LF), syllable number (SN), and sound file length
(SFL) [all ps > .1; average condition values in the order
bravely, warmth, archer, breadth, 丑闻, giraffe: 日志
WF = 0.65, 0.52, 0.59, 0.54, 0.70, 0.67; log LF = 0.64, 0.51,
0.57, 0.54, 0.67, 0.65; log stem WF = 1.07, 1.13, 0.95, 0.88,
0.95, n/a; log stem LF = 1, 1.08, 0.91, 0.86, 0.91, n/a; SN =
2.13, 2.18, 2.10, 2.10, 2.13, 2.10; SFL msec = 678, 679,
675, 694, 678, 650]. Each condition contained a mix of
grammatical word categories. 这 240 test words were
mixed with 80 filler words, 240 acoustic baseline trials,
和 160 silence trials.
The acoustic baseline trials were constructed to share the
complex auditory properties of speech without triggering
phonetic interpretation. Musical rain (MuR) achieves this
by extracting temporal energy envelopes for each of the
speech tokens, which are then filled by jittered fragments
of synthesized speech (Uppenkamp, Johnsrude, Norris,
Marslen-Wilson, & 帕特森, 2006). The resulting envelope-
shaped MuR stimuli are matched in RMS level and long-term
spectrotemporal distribution of energy to the correspond-
ing speech stimuli while triggering no speech percept.
Fourier transforms and spectrographic analyses of the
speech tokens and their corresponding MuR tokens show
that their spectra are well matched (Bozic et al., 2010).
程序
To maintain the close parallel with our earlier study of
inflectional processing (Bozic et al., 2010), we used the
same gap detection task. This is a nonlinguistic task that
engages lexical processing (Gaskell & Dumay, 2003; Mattys
& 克拉克, 2002) while keeping task requirements constant
across speech and nonspeech stimuli. Short silent gaps
(400 毫秒) were inserted in approximately 20% 的考验
(60 filler words and 60 MuR trials), and participants
were asked to decide as quickly and accurately as possible
whether or not words and MuR sounds contained a silent
gap. Participants responded using a button box. Only gap-
absent trials were subsequently analyzed.
The words were recorded in a sound-proof room by a
female native speaker of British English onto a DAT re-
corder. They were then digitized at a sampling rate of
22 kHz with 16 bit conversion and stored as separate files
using CoolEdit. CoolEdit was also used for gap insertion.
Items were presented using in-house software, and par-
ticipants heard the stimuli binaurally over Etymotic R-30
plastic tube phones. Sounds were preemphasized to
compensate for distortions imposed by the tube phone
presentation.
There were a total of 720 试验, which were pseudo-
randomized with respect to their type (测试, filler, 基线,
null) and presence or absence of gaps, and presented in
four blocks of 180 items each. There were five items at
the beginning of each block to allow the signal to reach
equilibrium. The experiment started with a short practice
outside the scanner, where participants were given feed-
back on their performance. Participants were asked to
keep their eyes closed during scanning.
Scanning was performed on a 3T Trio Siemens Scanner
at the MRC-CBU, 剑桥, using a fast sparse imaging
协议. Each trial consisted of a 1.4-sec silent period
and a 2-sec acquisition, with sounds played within the si-
lent periods to minimize the interference of scanner noise
with auditory processing (gradient-echo EPI sequence, rep-
etition time = 3.4 秒, acquisition time (饰面) = 2 秒, echo
time = 30 毫秒, flip angle = 78°, 矩阵大小= 64 × 64,
field of view = 192 × 192 毫米, 32 oblique slices 3 毫米
thick, 0.75 mm gap). MPRAGE T1-weighted scans were
acquired for anatomical localization.
For both univariate and multivariate analyses, pre-
processing was performed in SPM5 using the automatic analy-
sis library (github.com/rhodricusack/automaticanalysis).
For the univariate analyses, preprocessing involved image
realignment to correct for movement, segmentation, 和
spatial normalization of functional images to the MNI refer-
ence brain and smoothing with a 10-mm isotropic Gaussian
kernel. The data for each subject were then analyzed using
the general linear model, with four blocks and 11 事件
类型 (six test conditions, fillers, MuR, fillers with gap,
MuR with gap, silence). Motion regressors were included
as covariates of no interest to account for any residual
movement artifacts. A high-pass filter with a 200-sec cut-
off was applied to remove low-frequency noise. The BOLD
response for each event was modeled with the canonical
HRF and its derivatives. Contrast images were combined
into a group random effects analysis. At the group level,
1682
认知神经科学杂志
体积 25, 数字 10
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
哦
d
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
哦
9
C
6
n
5
_
4
A
/
_
j
0
哦
0
C
4
n
2
0
_
A
p
_
d
0
0
乙
4
y
2
G
0
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
t
.
F
/
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
effects across conditions were tested in a repeated mea-
sures ANOVA, implemented as a flexible factorial analysis
with the six test conditions and subject-specific effects to
remove the between subject variability. Differential effects
across conditions were tested by F-test effects of interest,
followed up by a series of t tests. The reported results are
significant at FDR p < .05 level, corrected for multiple
comparisons.
For the multivariate RSA, general linear models were
constructed using unsmoothed native space images that
had been realigned and coregistered to the subjectʼs
MPRAGE. Data were then extracted for each participant
individually using a “sphere of information” searchlight
approach (Kriegeskorte, Goebel, & Bandettini, 2006).
Here, a roaming spherical spotlight with a standard 5-mm
radius (Kriegeskorte et al., 2006) was moved through the
gray matter volume, centering onto each voxel in turn and
creating a continuous map of local activation patterns. The
signal from all voxels in a spotlight is combined to show
how well that pattern differentiates between the experi-
mental conditions and therefore reveals the information
contained in the pattern. For any given location, this in-
formation can be expressed as a representational dissimi-
larity matrix (RDM), which shows the correlation (1 − r
Pearson correlation) between activation elicited by differ-
ent conditions across voxels. Finally, we tested whether
these data RDMs correlate with theoretical models, ex-
pressed as model RDMs, which are generated to test the
role of linguistic, semantic, and perceptual variables (see
Results for details). This yielded maps of r values for each
participant and each model, which were normalized onto
the MNI template and statistically evaluated in a second-
level analysis using random effects statistics in SPM.
RESULTS
Behavioral Results
All errors (incorrect gap identifications, 1.5%) and time-
outs (RT > 3000 毫秒, 0.1%) were removed, and data
were inverse transformed (拉特克利夫, 1993). An ANCOVA
on the inverse transformed RTs with Condition as a fixed
factor and Speech File Duration as a covariate showed
significant differences between conditions by subjects,
but not by items [F1(5, 85) = 6.38, p < .01; F2(5, 233) =
1.43, p > .1]. There were no statistically significant differ-
ences in error rates [F1(1, 85) = .63, p > .1, F2(5, 233) =
.80, p > .1].
Imaging Results
On the basis of previous research (Bozic et al., 2010;
Tyler & Marslen-Wilson, 2008; Beeman, 2005; Binder
等人。, 1997, 2000) and our predictions, we selected bi-
lateral frontotemporoparietal regions as the volume of
interest for the analyses. Using WFU Pickatlas, a mask
was constructed, consisting of bilateral temporal lobes
(superior, 中间, and inferior temporal gyri and angu-
lar gyrus), IFG (pars orbitalis, pars opercularis, pars tri-
angularis, precentral gyrus) and the anterior cingulate.
Univariate Analyses
Using classic univariate analyses, we first established the
network that supports lower-level acoustic processing.
Subtracting null events from the MuR baseline showed
strong bilateral activity in Heschlʼs gyrus and surrounding
superior temporal regions (BA 41/BA 42, BA 22, peaks at
−52 −26 2 和 52 −26 8), consistent with results from
the literature (Uppenkamp et al., 2006; 戴维斯 & Johnsrude,
2003; Binder et al., 1997, 2000). This activity is shown in
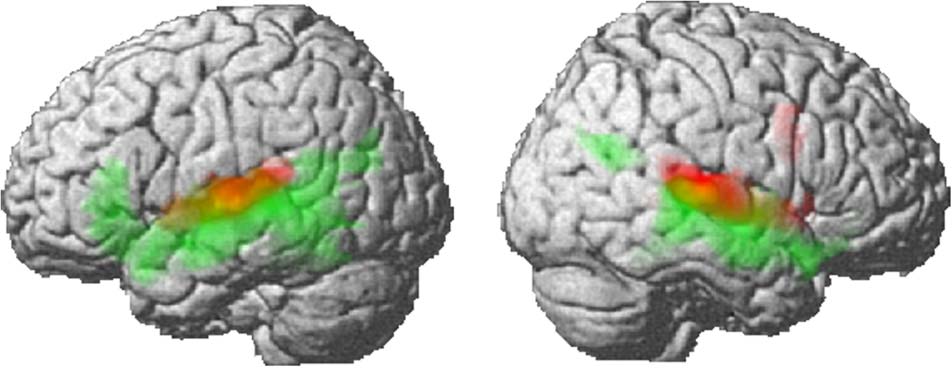
red in Figure 1. To extract the activity specifically related
to lexical processes, we contrasted all words with the
MuR baseline. This comparison showed that lexical pro-
cesses activated regions anterior and ventro-lateral to the
activity observed for lower-level auditory processing, 在-
marily in bilateral middle, superior, and inferior temporal
gyri (BA 21, BA 22, BA 38), spreading to left IFG (BA 45/
BA 47), left angular gyrus (BA 39), bilateral fusiform gyrus
(BA 37), and anterior cingulate (BA 32; 数字 1, 显示
in green and Table 2).
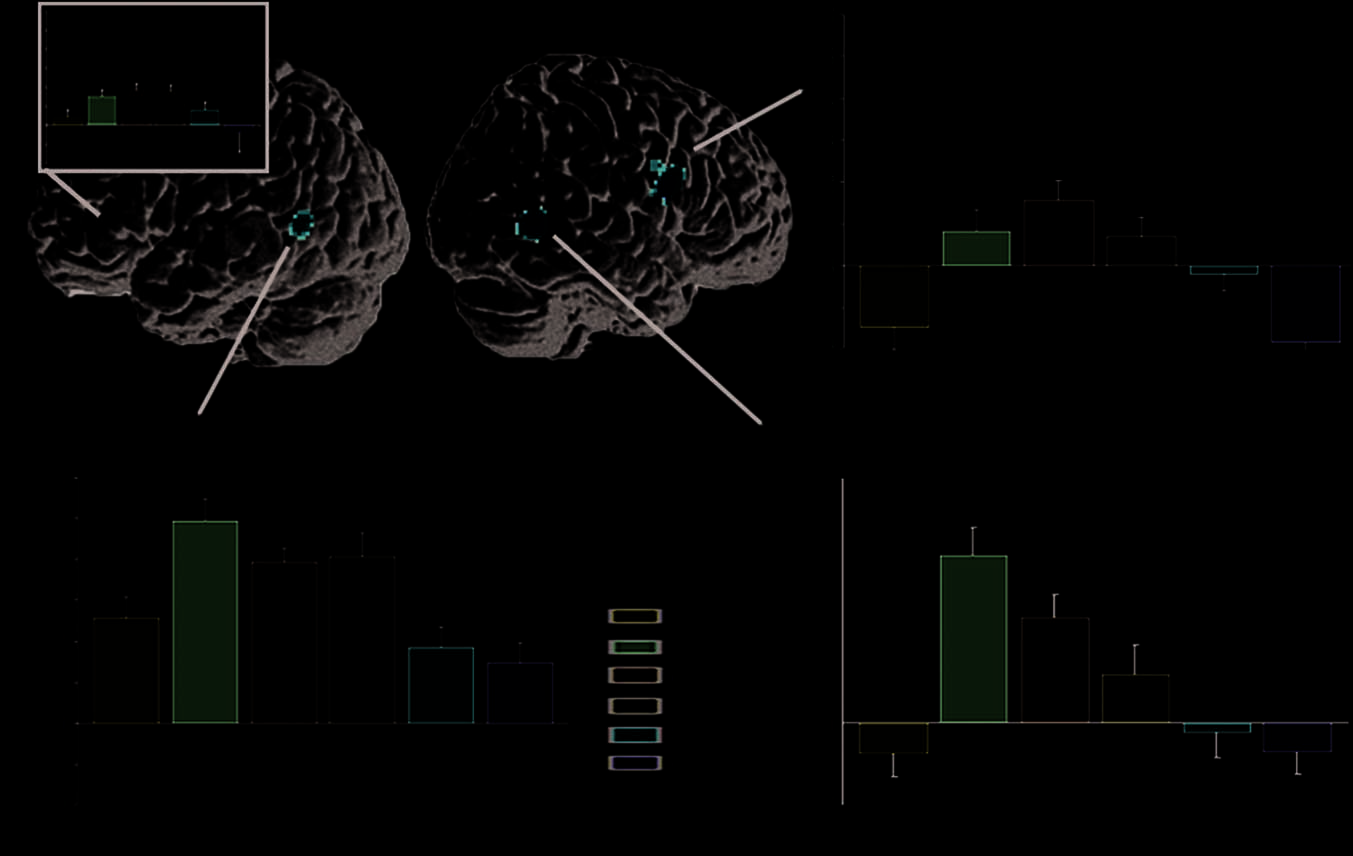
Next we focused on the processing of derivational
complexity in the frontotemporal network, performing a
repeated-measures ANOVA with added subject-specific
effects to compare lexical processing (words minus MuR
基线) across the six conditions. This analysis revealed
significant differential activation in bilateral middle tem-
poral gyrus (MTG) and in RIFG (数字 2 and Table 3A).
Signal intensity plots for each condition show that all three
clusters exhibit a similar activation pattern, 哪里的
semantically transparent words with productive suffixes
(bravely) pattern with simple words (giraffe) 生产
less activation than either semantically opaque words
(archer, breadth) or semantically transparent words with
nonproductive suffixes (warmth). No significant differ-
ences between conditions emerged in the left inferior
frontal areas. When we examined LIFG activations at a
lower threshold (see inset signal plot for LIFG in Fig-
乌尔 2), we saw the same pattern as seen for RIFG and
for MTG bilaterally. There was no evidence for selective
数字 1. Activation for complex acoustic processing (红色的) and lexical
加工 (绿色的) rendered onto the surface of a canonical brain.
All results are significant at FDR p < .05.
Bozic et al.
1683
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
o
9
c
6
n
5
_
4
a
/
_
j
0
o
0
c
4
n
2
0
_
a
p
_
d
0
0
b
4
y
2
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 2. Regions Supporting Speech-driven Lexical Processing
Regions
Words – MR
Cluster Extent
Voxel Z
R superior temporal gyrus (BA 22)
3939
R MTG (BA 21)
RIFG (BA 47)
L MTG (BA 21)
L MTG (BA 21)
L MTG (BA 21)
L inferior temporal gyrus (BA 37)*
LIFG (BA 45)*
L angular gyrus (BA 39)*
R parahippocampal region
R fusiform
R hippocampus
Anterior cingulate (BA 32)
Anterior cingulate (BA 32)
Medial frontal area (BA 10)
R MTG (BA 21)
7721
285
340
497
6.29
5.57
5.37
6.04
5.92
5.52
5.07
4.92
3.88
5.00
4.49
4.39
4.34
4.14
3.66
4.11
Coordinates
y
0
−4
24
−12
−34
−26
−60
34
−68
−26
−36
−18
32
34
58
−60
x
62
60
36
−54
−64
−58
−48
−46
−48
30
36
26
−6
12
2
50
z
−2
−20
−20
−6
4
−2
−14
0
16
−14
−16
−12
−10
−8
2
20
All results are significant at FDR p < .05 threshold. The three most significant peaks for each cluster are shown. The left temporal cluster also extends
into inferior frontal, angular, and inferior temporal areas. The most significant peaks in these regions are marked with an asterisk.
activation of the LH network, even for the highly com-
positional bravely condition. Instead, the bravely forms
seem to elicit lower levels of activation than the other
complex forms, uniformly across the brain.
We further explored this pattern of results in a series of
post hoc t tests between individual conditions. Compared
with simple words (giraffe, −S M0 P0), all conditions with
embedded stems or pseudostems (+S ), apart from the
Figure 2. Univariate results
showing areas differentially
activated by the six conditions,
tested by a repeated-measures
ANOVA. All results are
significant at FDR p < .05.
Signal plots from the peak
voxels of the three clusters
show a comparable inverted
U-shaped pattern across
conditions (the curve shows
the global average over
clusters). The additional analysis
for LIFG (at a lower threshold) is
shown as an inset at top left.
1684
Journal of Cognitive Neuroscience
Volume 25, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
o
9
c
6
n
5
_
4
a
/
_
j
0
o
0
c
4
n
2
0
_
a
p
_
d
0
0
b
4
y
2
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 3. Brain Areas Differentially Activated by the Six Conditions, Revealed by a Repeated Measures ANOVA Follow-up
Comparisons between Complex and Simple Forms
Regions
Cluster Extent
Voxel Z
(A) Brain Areas Differentially Activated by the Six Conditions
R MTG (BA 21)
RIFG (BA 44)
RIFG (BA 44)
R precentral (BA 6)
L MTG (BA 21)
229
252
83
4.73
4.01
3.69
3.41
3.57
(B) Complex Forms ( Warmth, Archer, Breadth, Scandal) vs. Simple Forms (Giraffe)
RIFG (BA 44)
LIFG (BA 44/6)
R MTG (BA 21/22)
R superior temporal gyrus (BA 22)
L superior temporal gyrus (BA 22)
L MTG (BA 21)
All results are significant at FDR p < .05.
388
82
97
78
63
4.23
4.08
3.68
3.26
3.42
3.40
Coordinates
y
−54
14
8
4
−56
10
8
−38
−32
−28
−50
x
58
60
50
52
−52
56
−58
58
62
−44
−54
z
2
22
16
30
2
18
20
14
20
8
4
transparent productive condition (bravely, +S +M +P),
showed a consistent pattern of increased activation in
bilateral frontotemporal regions. Because not all of the
clusters from these individual t tests reached statistical
significance (due to the limited power of t tests between
individual conditions), we grouped the relevant +S con-
ditions together and report the results of this combined
analysis (Table 3B). This comparison picks up signifi-
cant effects in the LIFG as well as in RIFG and bilateral
temporal regions, suggesting that the processing of
onset-embedded forms is indeed handled by a bilaterally
distributed system.
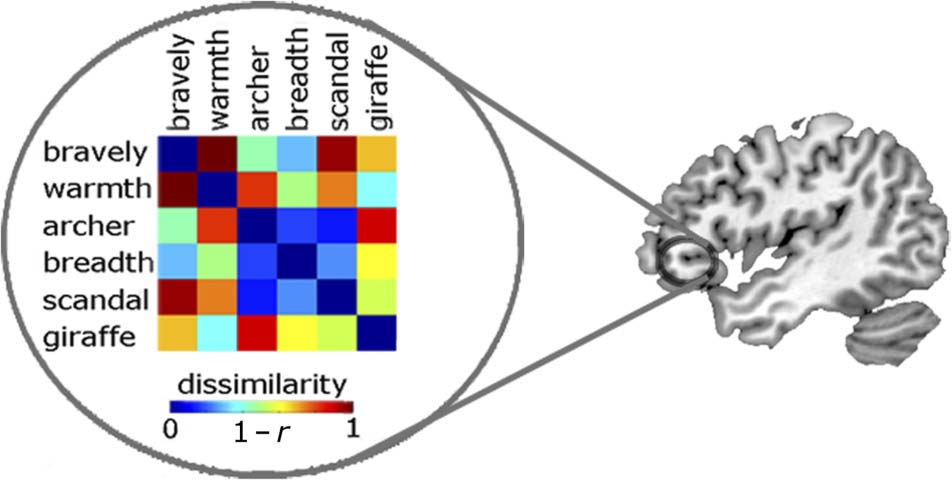
tion patterns for each pair of conditions. Figure 3 illustrates
the roaming searchlight approach and an example RDM.
To make inferences about the processing and representa-
tional properties of these data RDMs, they were compared
at each location against theoretical models, expressed as
model RDMs.
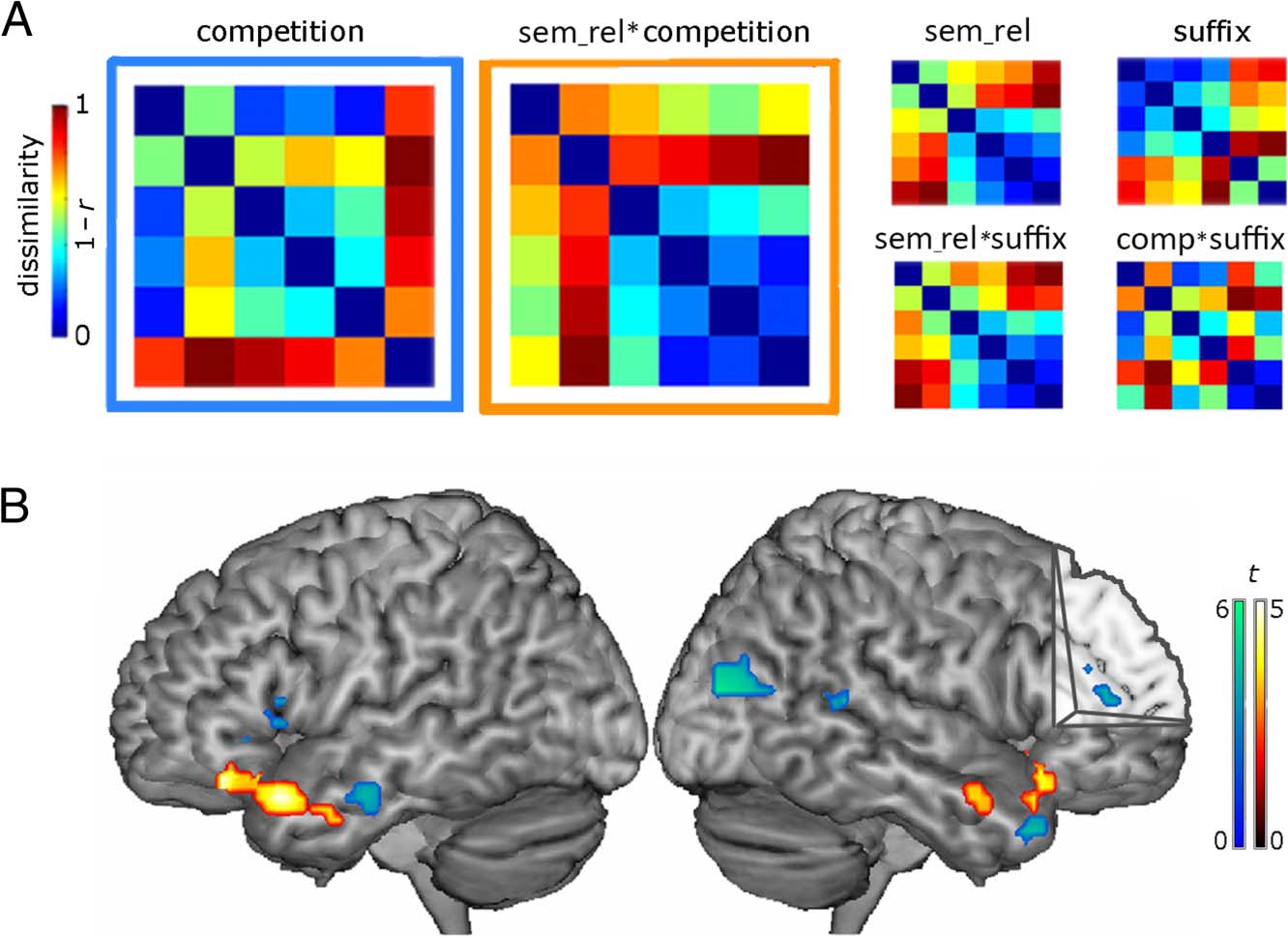
Six theoretical RDM models (see Figure 4) were devel-
oped to test hypotheses about the critical processing
dimensions underlying the brainʼs response to the experi-
mental stimuli and to assess the relative contributions of
linguistic, semantic, and perceptual variables to these re-
sponses. For the “linguistic” model RDM, we calculated
Multivariate Analyses
Here we explore the information encoded in the pat-
terns of neural activity within the frontotemporal language
processing network in order to assess the role of the dif-
ferent linguistic, semantic, and perceptual variables that
are diagnostic of the engagement of the two processing
systems. The multivariate analyses were conducted using
RSA combined with a roaming searchlight algorithm (Su
et al., 2010; Kriegeskorte et al., 2008). Using a 5-mm radius
spherical searchlight, a continuous voxel-by-voxel map
of activation patterns was generated for each condition
in each participant. These maps are subjected to a multi-
variate RSA analysis to reveal the correlations between con-
ditions (1 − r Pearson correlation across voxels). At each
searchlight location, these correlations are expressed as
RDMs (data RDMs) to show the distances between activa-
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 3. An example RDM extracted at a specific location by the
RSA roaming searchlight. The 6 × 6 matrix reflects the number
of experimental conditions and displays the correlation distance
(1 − r) between the activation patterns for each pair of conditions
at a given location.
Bozic et al.
1685
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
o
9
c
6
n
5
_
4
a
/
_
j
0
o
0
c
4
n
2
0
_
a
p
_
d
0
0
b
4
y
2
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
Figure 4. (A) The six RSA
models developed to capture
the processing of linguistic,
semantic, and perceptual
dimensions and their
interactions. Only the first two
models (perceptual competition
and Semantic Relatedness ×
Competition interaction)
significantly correlated with
the activation patterns in the
frontotemporal language
network. (B) Areas of significant
correlation for the competition
main effect (blue) and for
the Semantic Relatedness ×
Competition interaction
(orange). All results are
significant at FDR p < .05.
the productivity for each suffix using the Baayen and
Lieber (1991) hapax legomena procedure, extracting a
quantitative measure of productivity suitable for use in
the RSA context. The information whether or not a deriva-
tional suffix was present, and whether it was productive or
not (+/−P), suffix model, was expected to interact with lin-
guistically specific processes in left-lateralized peri-sylvian
areas. To construct the model RDM that tested for effects
of semantic transparency (+/−M), sem_rel, we used the
COALS metric (Rohde et al., 2005), which provides a corpus-
based estimate of the semantic relatedness between a
word and its embedded stem. To the extent that the
semantic transparency of a derived form contributes to its
decomposition, the effects were expected to emerge in
left-lateralized peri-sylvian areas. If, on the other hand,
semantic relatedness between the word and its embedded
stem primarily modulates the discriminability of these two
competing forms, this variable was expected to engage the
bihemispheric system. For the perceptual competition
RDM, the estimated degree of competition between a word
and its embedded stem (+/−S), competition model, was
expressed as the ratio of the logged form frequencies of
the derived word and the stem (or pseudostem). This
model was expected to reflect general perceptual analysis
and decision processes in the bihemispheric system. Three
further models (Suffix × Semantic relatedness, Suffix ×
Competition, Competition × Semantic relatedness) were
created to test the interactions between these variables.
For each of the continuous variables (suffix productivity,
semantic relatedness, perceptual competition, and their
respective interactions), we calculated a value for each of
the 240 test words (6 × 40 words per condition).1 These
variables were converted into model RDMs by creating
a 240 × 240 matrix across all the items (one matrix per
variable) and then calculating the average distance be-
tween each pair of conditions. This gave us a measure of
correlation distances between conditions for each of the
variables and their interactions (Figure 4A). The six model
RDMs were correlated with the activation RDMs at each
location and tested for significance using random effects
statistics in SPM. Figure 4B shows maps of significant
correlations in the bilateral frontotemporal network.
The results revealed significant bilateral activation for
perceptual competition between a word and its embedded
stem (+/−S), with the competition model correlating with
activation patterns in a distributed set of bilateral temporal
areas, left frontal gyrus and bilateral anterior cingulate (AC).
A separate cluster of activations in bilateral inferior frontal
regions (BA 47) and anterior MTG reflected the interaction
of competition with semantic relatedness (sem_rel × com-
petition model). No significant correlations were observed
for the linguistically related suffix model (+/−P) or any
of its interactions. There was a trend of correlations
between the semantic relatedness model and activation
patterns in bilateral frontal regions, but these were below
threshold. A more detailed listing of the pattern of results
is given in Table 4.
DISCUSSION
In the context of more general questions about how the
key combinatorial properties of language are handled in
the neurocognitive language system, we asked how a
major combinatorial language function—derivational word
formation—interfaced with the dual language systems
we have proposed in earlier research (Bozic et al., 2010;
Marslen-Wilson & Tyler, 2007).
1686
Journal of Cognitive Neuroscience
Volume 25, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
o
9
c
6
n
5
_
4
a
/
_
j
0
o
0
c
4
n
2
0
_
a
p
_
d
0
0
b
4
y
2
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 4. Regions Significantly Correlated with the Perceptual Competition Model and the Semantic Relatedness × Competition
Interaction
Coordinates
Regions
Cluster Extent
Voxel Z
Perceptual Competition Model
R MTG (BA 21)
R MTG (BA 21/19)
R MTG (BA 21/19)
R middle temporal pole (BA38)
L anterior cingulate (BA32)
L anterior cingulate (BA32)
L insula
LIFG (BA 45)
LIFG (BA 44)
R anterior cingulate (BA32)
L MTG (BA 21)
L MTG (BA 21)
Semantics × Competition Interaction
RIFG (BA 47)
RIFG (BA 47)
R insula
L superior temporal pole (BA 38)
LIFG (BA 47)
LIFG (BA 47)
R MTG (BA 21)
22
74
19
13
44
16
27
93
143
22
4.42
4.17
3.71
3.72
3.63
3.20
3.60
3.57
3.25
3.57
3.45
3.23
4.32
3.75
3.49
4.21
4.17
3.70
3.37
x
48
42
27
36
−3
−9
−42
−36
−45
9
−54
−48
21
39
30
−42
−36
−24
48
y
−42
−84
−72
18
51
39
18
27
18
33
−12
−3
15
21
6
18
36
18
3
z
11
15
22
−34
11
19
4
0
11
22
−22
−22
−19
−19
−15
−22
−15
−19
−22
Results are significant at FDR p < .05 threshold, corrected for multiple comparisons. Clusters of 12 or more contiguous voxels are reported.
We investigated two sets of hypotheses: If derivation-
ally complex forms are stored and processed with respect
to their constituent morphemes, as suggested by decom-
positional accounts, we expected to observe selective
activation of the LH decompositional system, similar to
regular inflectional morphology. On a strong decomposi-
tional hypothesis, this activation would be seen for all
items that contain a stem and a suffix. On a weaker hy-
pothesis, the likelihood of decomposition is modulated
by semantic transparency and suffix productivity, so that
the strongest selective activation of the LH system is
predicted for transparent productive words like bravely.
On other hand, if derived forms are neither stored nor
accessed as sets of separate morphemes, as suggested
by whole-form accounts, then we expect no selective
engagement of the left frontotemporal system for any
of the word sets. Instead, derived words should engage
the bilateral system, which supports stem-based, whole-
form lexical access. The activation of this system should
primarily reflect the perceptual processing complexity
of the words being heard—how easily can they be sepa-
rated from any competing lexical candidates. We consider
first whether or not derived words in English selectively
activate the LH decompositional system.
For English regular inflectional morphology, the neural
response has two distinctive properties. It is highly left-
lateralized, and it contrasts strongly with the bilateral pat-
tern of activation associated with increased nonlinguistic
processing demands (Bozic et al., 2010). Testing for
these properties in derived words using exactly parallel
methods and design and viewing the results from com-
bined univariate and multivariate perspectives, we see no
evidence for selective engagement of the LH system. The
univariate analyses (Tables 2 and 3 and Figures 1 and 2)
show no differential left-lateralized activation associated
with the morphologically complex stimuli relative to the
other stimulus classes. An ANOVA testing for differential
effects across conditions revealed processing differences
Bozic et al.
1687
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
o
9
c
6
n
5
_
4
a
/
_
j
0
o
0
c
4
n
2
0
_
a
p
_
d
0
0
b
4
y
2
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
in bilateral MTG, as well as right inferior frontal regions.
The profile of these differential effects (Figure 2) suggests
that they arise from the increased engagement of these re-
gions (as well as LIFG) by the set of perceptually complex
but less compositional words (warmth, archer, breadth,
scandal), compared with more compositional (bravely)
or simple words ( giraffe). Strikingly, the bravely stimuli
seem to pattern throughout with the simple words rather
than the complex forms. But there is no evidence that
this is because the bravely class falls into a distinctively dif-
ferent left-lateralized processing category. Whatever effects
are at work here, they seem to apply across the entire
bilateral frontotemporal system.
A complementary set of results emerged from the multi-
variate RSA analyses, which provide a more precise spec-
ification of the dimensions that drive the processing
of derived words. The RSA searchlight showed that the
patterns of voxel activity across frontotemporal regions
strongly correlate with levels of perceptual competition
(+/−P) and with the modulation of these processes by
semantic relatedness (+/−M). These are both dimensions
that we associate with the perceptual analysis of whole
forms as a function of their cohort competitor environ-
ment. No significant correlations emerged for the suffix
productivity model (+/−P) nor any of its interactions.
Moreover and consistent with the univariate results, the
competition-related effects were distributed bilaterally
across the regions that support the processing demands
associated with mapping from sound to meaning. These
analyses suggest that the comprehension of derived words
predominantly engages general perceptual processes in the
bihemispheric system, rather than specifically linguistic
decompositional processes supported by the LH.
Competition and Selection in Bilateral IFG
The two successful RSA models may be capturing, we
suggest, two related but separable general purpose pro-
cessing mechanisms that have been observed across a
broad range of stimulus domains and are not restricted
to language. The perceptual competition model reflects
the simultaneous activation of multiple lexical candidates
in words with an embedded stem, which requires addi-
tional processing to select the correct candidate. The RSA
searchlight shows that this correlates with activity patterns
in anterior cingulate, left inferior frontal areas BA 44/BA 45,
and bilateral middle temporal regions. Activation in the
anterior cingulate has long been associated with competi-
tion in general and more specifically with the detection
and signaling of the occurrence of conflict in stimulus pro-
cessing (Botvinick, Cohen, & Carter, 2004; Barch, Braver,
Sabb, & Noll, 2000). Similarly, more dorsal IFG (BA 44/
BA 45) has been consistently related to selection between
automatically retrieved competitors (Zhuang et al., 2012;
Zhuang, Randall, Stamatakis, Marslen-Wilson, & Tyler,
2011; Badre, Poldrack, Paré-Blagoev, Insler, & Wagner,
2005; Thompson-Schill, DʼEsposito, & Kan, 1999), often in
the context of a larger network of functionally coupled
temporal regions (e.g., Bokde, Tagamets, Friedman, &
Horwitz, 2001). In previous research, we have seen a com-
parable network of activation in the anterior cingulate,
left frontal and bilateral temporal regions associated with
the detection of increased processing demands in spoken
language processing (Stamatakis, Marslen-Wilson, Tyler, &
Fletcher, 2005).
In contrast to the perceptual competition model,
which seems to be primarily driven by the relative acti-
vation strength of an onset-embedded competitor, the
Semantics × Competition interaction reflects in addition
the semantic distance between the competitors, where
greater semantic similarity may affect the discriminability
of separate lexemes such as warm and warmth (Raposo
et al., 2006). This model correlated with a more ventral
set of bilateral regions, specifically the inferior frontal area
BA 47, as well as the temporal pole and anterior MTG. The
anterior parts of the IFG have been previously related to
lexical semantics (Hagoort, 2005) and to controlled se-
mantic retrieval and the associative strength between
words (Badre et al., 2005; Wagner, Paré-Blagoev, Clark, &
Poldrack, 2001). Several studies have suggested a role for
the temporal poles in semantic processing (e.g., Patterson,
Nestor, & Rogers, 2007). Competition-related activation in
bilateral BA 47 was also observed in our previous study
(Bozic et al., 2010), where semantic relatedness between
the competitors was not explicitly manipulated, but was sig-
nificantly correlated with the competition variable. Zhuang
et al. (2012) report cohort competition effects in similar
ventral IFG regions. In summary, the multivariate RSA
analyses not only confirm that the processing of derived
words predominantly engages perceptual processes in
a bihemispheric system but also allow a more fine-
grained specification of the joint but separable variables
that underlie this process.
Derivational Morphology in
Neurobiological Context
These results have significant implications for an account
of the processing and representation of derivationally com-
plex forms in English. First, they contrast strongly with the
distinctive left-lateralized decompositional processes seen
for English regular inflection. As discussed earlier, both
neurocognitive and linguistic properties of regular in-
flections converge to suggest that they undergo an obli-
gatory decomposition in the recognition process (e.g.,
Bozic et al., 2010; Bickel & Nichols, 2007; Marslen-Wilson,
2007). Left frontotemporal systems dynamically separate
inflected forms into stems, which are the primary carriers
of lexical meaning, and into inflectional suffixes, which
convey grammatical information but do not interact with
the meaning of the stem.
In the current experiment, we saw no selective engage-
ment of the LH system that supports core grammatical
computations and no evidence for processing specifically
1688
Journal of Cognitive Neuroscience
Volume 25, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
o
9
c
6
n
5
_
4
a
/
_
j
0
o
0
c
4
n
2
0
_
a
p
_
d
0
0
b
4
y
2
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
driven by the properties of derivational suffixes. This is
likely to reflect differences in the functional roles of inflec-
tional and derivational morphemes. Unlike inflectional
morphemes, derivational morphemes interact with the
meaning of the stem they attach to, forming a tight lexical-
ized connection and creating a new lexical entry in the
mental lexicon. These lexical entries vary in their com-
positionality and predictability, so that the meaning
of derived words cannot be reliably computed from the
meaning of the stem combined with the meaning of the
suffix. Word meaning must instead be represented as
a property of the whole form. In such cases, the pro-
cessing focus will be on the derived form as a whole, and
the main processing signatures are likely to reflect the
general processing demands associated with separating it
from its cohort competitors.
This is a pattern that we expected to see most strongly
for the opaque archer and breadth sets, where the mean-
ing of the whole form was not related to the meaning
of the embedded stem or pseudo-stem, and which we
also saw for the warmth transparent nonproductive set.
In all three of these conditions, we saw dominant effects
of competition-related variables in the bilateral fronto-
temporal system that supports sound-to-meaning mapping
for whole-word, stem-based lexical access. In each case,
there is likely to be cohort-based competition between
the separate lexical entries corresponding to the whole
derived form and its embedded stem, with this com-
petition made more complex in the warmth case by the
semantic similarity between stem and derived form.
This pattern of results across the majority of morpho-
logically complex conditions is consistent with the view
that English-derived words are stored as full forms, rather
than decompositionally in terms of their component mor-
phemes. However, this cannot be the whole story for two
reasons. The first is the results for the bravely category.
These words, although clearly containing an embedded
stem, do not behave as if this generates additional com-
petition-based processing load. The activation profiles in
the univariate analysis are much closer to those for the
simple words than for complex forms like warmth, sug-
gesting a different underlying representation, possibly sen-
sitive to linguistic factors (the synchronic productivity of
the suffix) as well as to semantic variables. The second
reason is the substantial evidence that semantically compo-
sitional words with productive suffixes such as bravely or
joyful robustly facilitate the processing of their stems in
overt priming tasks (Longtin, Segui, & Hallé, 2003; Meunier
& Segui, 2002; Marslen-Wilson et al., 1994). Overt priming
(unlike masked priming) reflects the properties of central
lexical representations and suggests that representations
for this type of derived word contain a preserved trace of
the underlying morphological structure to which the pro-
cessing system is responsive. Semantic transparency and
suffix productivity have also been shown to influence the
processing of derived forms in nonpriming studies (e.g.,
Bertram et al., 2000).
Given this body of evidence, Clahsen et al. (2003) pro-
posed that derivations are stored as separate lexemes,
but with preserved marking of morphological structure
for semantically transparent forms with productive suf-
fixes (see also Marslen-Wilson, 2007). Such an account
may be related to the processing similarity we saw be-
tween the transparent productive bravely forms and the
simple forms like giraffe. In the auditory modality—where
the input gradually unfolds over time and lexical access
requires the sequential mapping of the incoming input
onto stored representations—the preserved morphologi-
cal structure of bravely and its conceptual closeness to
the competing stem brave would make the access to this
representation minimally taxing for the comprehension
system, similarly to the processing of words that do not
have onset-embedded competitors. The presence of
morphological marking in the underlying representation
of derived forms like bravely arguably reflects the lin-
guistic experience of the user, and it is also possible—
although not tested here—that it will be influenced by
the salience of stem/affix structure in their orthographic
input, as there is good evidence that stems and affixes
are obligatorily identified in the early stages of visual word
recognition (e.g., Marslen-Wilson et al., 2008; Rastle et al.,
2000).
In conclusion, the current experiment investigated the
processing of English derivational complexity in a neuro-
biological context. We asked whether derivationally com-
plex words show a similar neurocognitive distribution to
regularly inflected words. The results were clear: We saw
no selective engagement of the LH systems that support
core grammatical computations and no evidence for left-
lateralized processes driven by the properties of deriva-
tional suffixes. Instead, we observed robust activation
in a distributed bihemispheric system, which supports
the processing demands associated with mapping from
sound to meaning. Responses were primarily driven by
the properties of the derived form as a whole, with pro-
cessing signatures that reflect the demands associated
with separating the word from competing alternatives.
At the same time, the results for the bravely condition
suggest that the bilateral system is sensitive to decompo-
sitional aspects of lexical representations, with a reduced
processing load for semantically and linguistically trans-
parent derived forms. The results have important impli-
cations for theories of the processing and representation
of English derivational morphology and highlight the
fact that the underlying neural substrates provide an
essential framework for understanding these processes.
Acknowledgments
This work was supported by a European Research Council
Advanced Grant (230570 Neurolex) and Medical Research Council
Cognition and Brain Sciences Unit funding to W. D. M.-W.
(U.1055.04.002.00001.01). We thank D. Timothy Ives for his help
with the musical rain baseline.
Bozic et al.
1689
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
o
9
c
6
n
5
_
4
a
/
_
j
0
o
0
c
4
n
2
0
_
a
p
_
d
0
0
b
4
y
2
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Reprint requests should be sent to Dr. Mirjana Bozic or William
D. Marslen-Wilson, Department of Psychology, University of
Cambridge, Downing Street, Cambridge CB2 3EB, UK, or via
e-mail: mb383@cam.ac.uk, wdm10@cam.ac.uk.
Note
1. To capture the absence of semantic and competition values
for the words without an embedded stem ( giraffe), a value of
0 was assigned. This was entered into the sem_rel and competi-
tion models as a constrained normal distribution of random
numbers around the mean of 0. The same procedure was
applied to nonsuffixed words ( giraffe, scandal ) in the suffix.
REFERENCES
Anderson, S. R. (1992). A-morphous morphology. Cambridge:
Cambridge University Press.
Baayen, H., & Lieber, R. (1991). Productivity and English
derivation: A corpus-based study. Linguistics, 29, 801–844.
Baayen, R. H., Dijkstra, T., & Schreuder, R. (1997). Singulars
and plurals in Dutch: Evidence for a parallel dual-route
model. Journal of Memory and Language, 36, 94–117.
Baayen, R. H., Milin, P., Filipovic-Durdevic, D., Hendrix, P., &
Marelli, M. (2011). An amorphous model for morphological
processing in visual comprehension based on naive
discriminative learning. Psychological Review, 118,
428–481.
Badecker, W., & Caramazza, A. (1991). Morphological
composition in the lexical output system. Cognitive
Neuropsychology, 8, 335–321.
Badre, D., Poldrack, R. A., Paré-Blagoev, E. J., Insler, R. Z.,
& Wagner, A. D. (2005). Dissociable controlled retrieval
and generalized selection mechanisms in ventrolateral
prefrontal cortex. Neuron, 47, 907–918.
Barch, D. M., Braver, T. S., Sabb, F. W., & Noll, D. C. (2000).
Anterior cingulate and the monitoring of response conflict:
Evidence from an fMRI study of overt verb generation.
Journal of Cognitive Neuroscience, 12, 298–309.
Bauer, L. (1983). English word formation. Cambridge:
Cambridge University Press.
Beeman, M. (2005). Bilateral brain processes for
comprehending natural language. Trends in Cognitive
Sciences, 9, 512–518.
Bertram, R., Schreuder, R., & Baayen, R. H. (2000). The
balance of storage and computation in morphological
processing: The role of word formation type, affixal
homonymy, and productivity. Journal of Experimental
Psychology: Memory, Learning, and Cognition, 26,
419–511.
Bickel, B., & Nichols, J. (2007). Inflectional morphology.
In T. Shopen (Ed.), Language typology and syntactic
description, Vol. 3: Grammatical categories and the lexicon
(pp. 169–240). Cambridge: Cambridge University Press.
Binder, J. R., Frost, J. A., Hammeke, T. A., Bellgowan, P. S. F.,
Springer, J. A., & Kaufman, J. N. (2000). Human temporal
lobe activation by speech and nonspeech sounds. Cerebral
Cortex, 10, 512–528.
Binder, J. R., Frost, J. A., Hammeke, T. A., Cox, R. W., Rao, S. M.,
& Prieto, T. (1997). Human brain language areas identified
by functional magnetic resonance imaging. The Journal
of Neuroscience, 17, 353–362.
Bokde, A. L. W., Tagamets, M.-A., Friedman, R. B., & Horwitz, B.
(2001). Functional interactions of the inferior frontal cortex
during the processing of words and word-like stimuli.
Neuron, 30, 609–617.
Botvinick, M., Cohen, J. D., & Carter, C. S. (2004). Conflict
monitoring and anterior cingulate cortex: An update.
Trends in Cognitive Sciences, 8, 539–546.
Bozic, M., Marslen-Wilson, W. D., Stamatakis, E. A., Davis, M. H.,
& Tyler, L. K. (2007). Differentiating morphology, form, and
meaning: Neural correlates of morphological complexity.
Journal of Cognitive Neuroscience, 19, 1464–1475.
Bozic, M., Tyler, L. K., Ives, D. T., Randall, B., & Marslen-Wilson,
W. D. (2010). Bi-hemispheric foundations for human speech
comprehension. Proceedings of the National Academy
of Sciences, U.S.A., 107, 17439–17444.
Butterworth, B. (1983). Lexical representation. In B. Butterworth
(Ed.), Language production ( Vol. 2, pp. 257–294).
San Diego, CA: Academic Press.
Clahsen, H., Sonnenstuhl, I., & Blevins, J. P. (2003). Derivational
morphology in the German mental lexicon: A dual mechanism
account. In R. H. Baayen & R. Schreuder (Eds.), Morphological
structure in language processing (pp. 125–155). Berlin:
Mouton de Gruyter.
Davis, M. D., Meunier, F., & Marslen-Wilson, W. D. (2004).
Neural responses to morphological, syntactic, and semantic
properties of single words: An fMRI study. Brain and
Language, 89, 439–449.
Davis, M. H., & Johnsrude, I. S. (2003). Hierarchical processing in
spoken language comprehension. Journal of Neuroscience,
23, 3423–3431.
Ford, M. A., Davis, M. H., & Marslen-Wilson, W. (2010).
Derivational morphology and base morpheme frequency.
Journal of Memory and Language, 63, 117–130.
Friederici, A. D., Ruschemeyer, S. A., Hahne, A., & Fiebach, C. J.
(2003). The role of left inferior frontal and superior temporal
cortex in sentence comprehension: Localizing syntactic
and semantic processes. Cerebral Cortex, 13, 170–177.
Gaskell, G. M., & Dumay, N. (2003). Lexical competition and
the acquisition of novel words. Cognition, 89, 105–132.
Goodglass, H., Christiansen, J., & Gallagher, R. (1993). Comparison
of morphology and syntax in free narrative and structured
tests: Fluent versus nonfluent aphasics. Cortex, 29, 377–407.
Hagoort, P. (2005). On Broca, brain, and binding: A new
framework. Trends in Cognitive Sciences, 9, 416–423.
Hamilton, A. C., & Coslett, H. B. (2008). Role of inflectional
regularity and semantic transparency in reading
morphologically complex words: Evidence from acquired
dyslexia. Neurocase, 14, 347–368.
Hay, J., & Plag, I. (2004). What constrains possible suffix
combinations? On the interaction of grammatical and
processing restrictions in derivational morphology.
Natural Language and Linguistic Theory, 22, 565–596.
Hickok, G., Okada, K., Barr, W., Pa, J., Rogalsky, C., Donnelly, K.,
et al. (2008). Bilateral capacity for speech sound processing
in auditory comprehension: Evidence from Wada
procedures. Brain and Language, 107, 179–184.
Hickok, G., & Poepell, D. (2007). The cortical organization
of speech processing. Nature Reviews Neuroscience, 8,
393–402.
Kriegeskorte, N., Goebel, R., & Bandettini, P. (2006).
Information-based functional brain mapping.
Proceedings of the National Academy of Sciences,
U.S.A., 103, 3863–3868.
Kriegeskorte, N., Mur, M., & Bandettini, P. A. (2008).
Representational similarity analysis—Connecting the
branches of systems neuroscience. Frontiers in Systems
Neuroscience, 2, 1–28.
Leminen, A., Leminen, M., Lehtonen, M., Nevalainen, P.,
Ylinen, S., Kimppa, L., et al. (2011). Spatiotemporal
dynamics of the processing of spoken inflected and
derived words: A combined EEG and MEG study.
Frontiers in Human Neuroscience, 5, 1–14.
1690
Journal of Cognitive Neuroscience
Volume 25, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
o
9
c
6
n
5
_
4
a
/
_
j
0
o
0
c
4
n
2
0
_
a
p
_
d
0
0
b
4
y
2
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Longtin, C. M., & Meunier, F. (2005). Morphological
decomposition in early visual word processing. Journal
of Memory and Language, 53, 26–41.
Longtin, C. M., Segui, J., & Hallé, P. A. (2003). Morphological
priming without morphological relationship. Language
and Cognitive Processes, 18, 313–334.
Longworth, C., Marslen-Wilson, W. D., Randall, B., & Tyler, L. K.
(2005). Getting to the meaning of the regular past tense:
Evidence from neuropsychology. Journal of Cognitive
Neuroscience, 17, 1087–1097.
Marangolo, P., & Piras, F. (2008). Dissociations in processing
derivational morphology: The right basal ganglia
involvement. Neuropsychologia, 46, 196–205.
Marchand, H. (1969). The categories and types of present-day
English word-formation. Munich: Beck.
Marslen-Wilson, W. D. (2007). Morphological processes
in language comprehension. In G. Gaskell (Ed.), Oxford
handbook of psycholinguistics (pp. 175–193). Oxford:
Oxford University Press.
Marslen-Wilson, W. D., Bozic, M., & Randall, B. (2008). Early
decomposition in visual word recognition: Dissociating
morphology, form, and meaning. Language and Cognitive
Processes, 23, 394–421.
Marslen-Wilson, W. D., Hare, M., & Older, L. (1993). Inflectional
morphology and phonological regularity in the English
mental lexicon. Proceedings of the 15th Annual Meeting
of the Cognitive Science Society. Princeton, NJ: Erlbaum.
Marslen-Wilson, W. D., & Tyler, L. K. (1998). Rules,
representations, and the English past tense. Trends in
Cognitive Sciences, 2, 428–435.
Marslen-Wilson, W. D., & Tyler, L. K. (2007). Morphology,
language and the brain: The decompositional substrate
for language comprehension. Philosophical Transactions
of the Royal Society: Biological Sciences, 362, 823–836.
Marslen-Wilson, W. D., Tyler, L. K., Waksler, R., & Older, L.
(1994). Morphology and meaning in the English mental
lexicon. Psychological Review, 101, 3–33.
Matthews, P. H. (1991). Morphology (2nd ed.). Cambridge:
Cambridge University Press.
Mattys, S. L., & Clark, J. H. (2002). Lexical activity in speech
processing: Evidence from pause detection. Journal of
Memory and Language, 47, 343–359.
Meinzer, M., Lahiri, A., Flaisch, T., Hannemann, R., & Eulitz, C.
(2009). Opaque for the reader but transparent for the
brain: Neural signatures of morphological complexity.
Neuropsychologia, 47, 1964–1971.
Meunier, F., & Segui, J. (2002). Cross-modal morphological
priming in French. Brain and Language, 83, 89–102.
Patterson, K., Nestor, P. J., & Rogers, T. T. (2007). Where do
you know what you know? The representation of semantic
knowledge in the human brain. Nature Reviews
Neuroscience, 8, 976–987.
Plag, I. (2006). Productivity. In B. Aarts & A. McMahon
(Eds.), The handbook of English linguistics (pp. 537–556).
Oxford: Blackwell.
∼plaut/papers/abstracts/RohdeGonnermanPlautSUB-CogSci.
COALS.html.
Schreuder, R., & Baayen, H. (1995). Modeling morphological
processing. In L. Feldman (Ed.), Morphological aspects
of language processing (pp. 131–154). Hillsdale, NJ:
Lawrence Erlbaum Associates.
Shtyrov, Y., Pihko, E., & Pulvermuller, F. (2005). Determinants
of dominance: Is language laterality determined by physical
or linguistic features of speech? Neuroimage, 27, 37–47.
Stamatakis, E. A., Marslen-Wilson, W. D., Tyler, L. K., &
Fletcher, P. C. (2005). Cingulate control of frontotemporal
integration reflects linguistic demands: A three-way interaction
in functional connectivity. Neuroimage, 28, 115–121.
Su, L., Wingfield, C., Bozic, M., Fonteneau, E., Marslen-Wilson,
W. D., & Kriegeskorte, N. (2010). A multimodal approach
to representational similarity analysis. Poster presented
at 16th Annual Meeting of the Organization for Human
Brain Mapping.
Taft, M. (2004). Morphological decomposition and the
reverse base frequency effect. The Quarterly Journal
of Experimental Psychology, 57A, 745–765.
Thompson-Schill, S. L., DʼEsposito, M., & Kan, I. P. (1999). Effects
of repetition and competition on activity in left prefrontal
cortex during word generation. Neuron, 23, 513–522.
Tyler, L. K., de Mornay-Davies, P., Anokhina, R., Longworth, C.,
Randall, B., & Marslen-Wilson, W. D. (2002). Dissociations
in processing past tense morphology: Neuropathology and
behavioural studies. Journal of Cognitive Neuroscience,
14, 79–94.
Tyler, L. K., & Marslen-Wilson, W. D. (2008). Frontotemporal
brain systems supporting spoken language comprehension.
Philosophical Transactions of the Royal Society of London,
Series B, Biological Sciences, 363, 1037–1054.
Tyler, L. K., Marslen-Wilson, W. D., Randall, B., Wright, P.,
Devereux, B. J., Zhuang, J., et al. (2011). Left inferior frontal
cortex and syntax: Function, structure and behaviour in
left-hemisphere damaged patients. Brain, 134, 415–431.
Tyler, L. K., Stamatakis, E. A., Post, B., Randall, B., &
Marslen-Wilson, W. D. (2005). Temporal and frontal
systems in speech comprehension: An fMRI study of past
tense processing. Neuropsychologia, 43, 1963–1974.
Uppenkamp, S., Johnsrude, I. S., Norris, D., Marslen-Wilson,
W. D., & Patterson, R. D. (2006). Locating the initial stages
of speech-sound processing in human temporal cortex.
Neuroimage, 31, 1284–1296.
Vannest, J., Polk, T. A., & Lewis, R. L. (2005). Dual-route
processing of complex words: New fMRI evidence
from derivational suffixation. Cognitive, Affective, and
Behavioral Neuroscience, 5, 67–76.
Wagner, A. D., Paré-Blagoev, E. J., Clark, J., & Poldrack, R. A.
(2001). Recovering meaning: Left prefrontal cortex guides
controlled semantic retrieval. Neuron, 31, 329–338.
Whiting, C., Shtryov, Y., & Marslen-Wilson, W. D. (under
review). Spatiotemporal dynamics of visual word
recognition: Morphology, form, and meaning.
Raposo, A., Moss, H. E., Stamatakis, E. A., & Tyler, L. K.
Wright, P., Stamatakis, E. A., & Tyler, L. K. (2012). Differentiating
(2006). Repetition suppression and semantic enhancement:
An investigation of the neural correlates of priming.
Neuropsychologia, 44, 2284–2295.
Rastle, K., Davis, M. D., Marslen-Wilson, W. D., & Tyler, L. K.
(2000). Morphological and semantic effects in visual word
recognition: A time-course study. Language and Cognitive
Processes, 15, 507–537.
Ratcliff, R. (1993). Methods for dealing with reaction time
outliers. Psychological Bulletin, 114, 510–532.
Rohde, D. L. T., Gonnerman, L., & Plaut, D. C. (2005). An
improved model of semantic similarity based on lexical
co-occurrence. Retrieved from www.cnbc.cmu.edu/
hemispheric contributions to syntax and semantics in
patients with left-hemisphere lesions. Journal of
Neuroscience, 32, 8149–8157.
Zhuang, J., Randall, B., Stamatakis, E. A., Marslen-Wilson,
W. D., & Tyler, L. K. (2011). The interaction of lexical
semantics and cohort competition in spoken word
recognition: An fMRI study. Journal of Cognitive
Neuroscience, 23, 3778–3790.
Zhuang, J., Tyler, L. K., Randall, B., Stamatakis, E. A., &
Marslen-Wilson, W. D. (2012). Optimally efficient neural
systems for processing spoken language. Cerebral Cortex,
doi:10.1093/cercor/bhs366.
Bozic et al.
1691
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
1
2
0
5
/
1
1
6
0
7
/
8
1
1
6
9
7
4
8
5
/
8
1
3
9
7
7
o
9
c
6
n
5
_
4
a
/
_
j
0
o
0
c
4
n
2
0
_
a
p
_
d
0
0
b
4
y
2
g
0
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1