Musician Children Detect Pitch Violations
in Both Music and Language Better
than Nonmusician Children:
Behavioral and Electrophysiological Approaches
Cyrille Magne1,2, Daniele Scho¨n1,2, and Mireille Besson1,2
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
抽象的
& The idea that extensive musical training can influence
processing in cognitive domains other than music has re-
ceived considerable attention from the educational system
and the media. Here we analyzed behavioral data and re-
corded event-related brain potentials (ERPs) from 8-year-old
children to test the hypothesis that musical training facili-
tates pitch processing not only in music but also in language.
We used a parametric manipulation of pitch so that the final
notes or words of musical phrases or sentences were con-
gruous, weakly incongruous, or strongly incongruous. Musi-
cian children outperformed nonmusician children in the
detection of the weak incongruity in both music and lan-
规格. 而且, the greatest differences in the ERPs of
musician and nonmusician children were also found for the
weak incongruity: whereas for musician children, early neg-
ative components developed in music and late positive com-
ponents in language, no such components were found for
nonmusician children. 最后, comparison of these results
with previous ones from adults suggests that some aspects of
pitch processing are in effect earlier in music than in lan-
规格. 因此, the present results reveal positive transfer ef-
fects between cognitive domains and shed light on the time
course and neural basis of the development of prosodic and
melodic processing. &
介绍
Many results in the rapidly evolving field of the neuro-
science of music demonstrate that musical practice has
important consequences on the anatomo-functional or-
ganization of the brain. From an anatomical perspective,
magnetic resonance imaging, 例如, has revealed
morphological differences between musicians and non-
musicians in auditory (including Heschl’s gyrus and sec-
ondary auditory cortex), motor (central), and visuospatial
(顶叶) brain areas (Gaser et al., 2003; 施耐德
等人。, 2002), as well as in the size of the corpus callo-
sum and planum temporale (Schlaug, Jancke, 黄,
& Steinmetz, 1995; Schlaug, Jancke, 黄, Staiger, &
Steinmetz, 1995). Such anatomical differences have func-
tional implications. 的确, research using functional
magnetic resonance imaging and magnetoencephalog-
raphy has shown increased activity in Heschl’s gyrus of
professional and amateur musicians compared with non-
musicians (Schneider et al., 2002), increased somatosen-
sory and motor representations with musical practice
(Pantev et al., 1998; Elbert, Pantev, Wienbruch, Rockstroh,
& Taub, 1995), and larger bilateral activation of planum
1Institut de Neurosciences Cognitives de la Me´diterrane´e,
2Universite´ de la Me´diterrane´e
temporale for musicians than nonmusicians (Ohnishi
等人。, 2001).
有趣的是, although these different regions may
fulfill different musical functions, such as the encoding
of auditory information (Heschl’s gyrus and secondary
auditory cortex), transcoding visual notation into motor
陈述, and playing an instrument (visuospatial,
somatosensory, 和运动脑区), they are not
necessarily specific to music. 相当, these different
brain structures have also been shown to be activated
by other cognitive functions. 例如, Heschl’s
gyrus, the secondary auditory cortex, and planum tem-
porale are typically involved in different aspects of
语言处理 (迈耶, Alter, Angela, Lohmann,
& von Cramon, 2002; Tzourio et al., 1997). 此外,
visuospatial areas in the parietal lobes have been shown
to be activated by approximate calculation in arithmetic
(Culham & Kanwisher, 2001; 德阿内, 游戏, Pinel,
Stanescu, & Tsivkin, 1999). 反过来, recent results
obtained with the magnetoencephalography meth-
od have demonstrated that Broca’s area is not as
language-specific as believed for almost a century. 在-
契据, this brain area was activated not only by syntactic
processing of linguistic phrases, but also by syntactic
processing of musical phrases (Maess, Koelsch, Gunter,
& Friederici, 2001).
D 2006 麻省理工学院
认知神经科学杂志 18:2, PP. 199–211
我
我
/
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
D
H
哦
t
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
r
p
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
米
A
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
t
n
我
C
/
e
A
–
r
p
t
d
我
C
1
我
8
e
2
–
1
p
9
d
9
F
/
1
1
9
8
3
5
/
7
2
0
/
3
1
9
哦
9
C
/
n
1
2
7
0
5
0
6
6
0
1
3
8
3
/
2
j
1
哦
9
C
9
n
p
.
d
2
0
乙
0
y
6
G
.
你
1
e
8
s
.
t
2
哦
.
n
1
0
9
8
9
S
.
p
e
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
.
F
.
t
.
.
.
哦
n
1
8
中号
A
y
2
0
2
1
合在一起, these results show that musical prac-
tice has consequences on the anatomo-functional orga-
nization of brain regions that are not necessarily specific
to music. The idea that we wanted to test in the present
experiment is that musical practice, by favoring the
development and functional efficiency of specific brain
地区, may not only benefit different aspects of music
加工, but may also favor positive transfers in other
domains of cognition.
Positive transfer due to extended musical practice has
been described at the behavioral level, in both adults
and children, in domains that are not directly linked
to music, such as mathematical abilities (Bilhartz,
Bruhn, & 奥尔森, 2000; Costa-Giomi, 1999; Graziano
等人。, 1999; Gardiner, 狐狸, Knowles, & 杰弗里, 1996),
精神的
imagery (Aleman, Nieuwenstein, Bo¨cker, &
Hann, 2000), symbolic and spatio-temporal reasoning
(Gromko & Poorman, 1998; Rauscher et al., 1997), visuo-
spatial abilities (Brochard, Dufour, & Despre`s, 2004;
Cupchick, Philips, & 爬坡道, 2001; Hetland, 2000), verbal
记忆 (Ho, 张, & Chan, 2004; Chan, Ho, &
张, 1998), self-esteem (Costa-Giomi, 2004), 和
智力
very recently for measures of general
(Schellenberg, 2004). 然而, as noted by Thompson,
Schellenberg, and Husain (2004), although most of the
studies reported above were successful
in showing
positive correlations between music and other cognitive
域, very few studies have aimed at testing specific
hypotheses regarding the causal links underlying these
effects. 清楚地, such causal links would be easier to test
by studying positive transfer between music and other
cognitive domains that involve, at least partially, a similar
set of computations. One such candidate is language.
的确, several authors have emphasized the similarities
between language and music processing (see Koelsch,
2005; Patel, 2003A, 2003乙; Zatorre et al., 2002; Besson &
Scho¨n, 2001, for reviews).
Although a number of experiments have aimed at
comparing aspects of music and language processing
that are presumably quite different, such as syntax and
harmony or semantic and melody (Patel, 吉布森, Ratner,
Besson, & Holcomb, 1998; Besson & Faı¨ta, 1995), 仅有的
few recent studies have compared two aspects that are
objectively more similar, melody and prosody, 音乐
of speech. Prosody has both a linguistic and an emo-
function and can broadly be defined at the
的
抽象的, phonological
等级, as the patterns of stress
and intonation in a spoken language, and at the con-
crete, acoustic level, by the same parameters that define
melody (IE。, the rhythmic succession of pitches in
音乐), 那是, fundamental frequency (F0), intensity,
duration, and spectral characteristics. Based on these
similarities, Thompson et al. (2004) tackled the emo-
tional function of prosody. They were able to show that
adult musicians outperformed adult nonmusicians at
identifying emotions (例如, sadness, fear) conveyed by
spoken sentences and by tone sequences that mimicked
the utterances’ prosody. Most importantly, 他们也
showed that 6-year-olds, tested after a year of musical
训练, were better than nonmusician children at iden-
tifying anger or fear.
Analyzing both the behavioral measures and variations
in brain electrical activity time-locked to events of
兴趣 (IE。, event-related brain potentials, or ERPs),
Scho¨n, Magne, and Besson (2004) designed an experi-
ment to directly compare pitch processing in music and
语言 (F0). Short musical and linguistic phrases were
aurally presented, and the final word/note was melodi-
cally/prosodically congruous or incongruous. Incongru-
ities were built by increasing the pitch of the final notes
or the F0 of the final words by one fifth of a tone and
35%, 分别, for the weak incongruities and by half
of a tone and 120%, 分别, for the strong incon-
gruities. The general hypothesis is that if similar pro-
cesses underlie the perception of pitch in language and
音乐, then improved pitch perception in music, due to
musical expertise, may extend to pitch perception in
语言. 最后, musicians should perceive
pitch deviations better than nonmusicians not only in
音乐, but also in language. 的确, results showed that
adult musicians not only detected variations of pitch in
melodic phrases better than nonmusicians, but that they
also detected variations of fundamental frequency in
句子 (linguistic prosody) better than nonmusicians.
而且, detailed analysis of the ERPs revealed that the
latency of the positive components elicited by the weak
and strong incongruities in both music and language was
shorter for musicians than for nonmusicians. 最后,
analysis of the amplitude and scalp distribution of early
negative components also revealed evidence for positive
transfer between music and language.
Based on these results, the aim of the present exper-
iment is twofold. 第一的, we wanted to determine whether
such positive transfer effects between pitch processing
in music and language would also be found in 8-year-old
孩子们. 换句话说, 会 3 到 4 years of extended
musical practice be sufficient for musician children to
outperform nonmusician children in the detection of
pitch violations in both music and language, as was
shown for adults with an average of 15 years of musical
训练 (Scho¨n et al., 2004)? Based on the provocative
results by Thompson et al. (2004), 证明
1 year of musical training has a strong influence on the
identification of emotional prosody, we also expected to
find positive evidence for linguistic prosody. 而且,
by using a parametric manipulation of pitch in both
language and music as in our previous study (Scho¨n
等人。, 2004), we were able to make specific predictions
regarding the effects of musical training. 因此, 我们
expected no differences between musician and nonmu-
sician children in the detection of congruous endings,
because they match the expectations derived from the
previous linguistic or musical contexts. 相似地, 我们
expected no differences between the two groups in
200
认知神经科学杂志
体积 18, 数字 2
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
D
H
哦
t
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
r
p
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
米
A
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
t
n
我
C
/
e
A
–
r
p
t
d
我
C
1
我
8
e
2
–
1
p
9
d
9
F
/
1
1
9
8
3
5
/
7
2
0
/
3
1
9
哦
9
C
/
n
1
2
7
0
5
0
6
6
0
1
3
8
3
/
2
j
1
哦
9
C
9
n
p
.
d
2
0
乙
0
y
6
G
.
你
1
e
8
s
.
t
2
哦
.
n
1
0
9
8
9
S
.
p
e
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
F
t
.
.
.
.
.
哦
n
1
8
中号
A
y
2
0
2
1
the detection of the strong incongruity, because in both
language and music, this deviation was constructed in
such a way as to be obvious. 相比之下, we expected
differences between musicians and nonmusician chil-
dren in the detection of the weak incongruity because
this deviation was subtle and should require a musical
ear to be detected.
The second aim was to study the neurophysiological
basis of positive transfer using a developmental ap-
普罗奇. 的确, one further reason to test 8-year-olds
is that previous results, based on the analysis of the
auditory evoked potentials, have shown that the audi-
tory cortex is not completely mature at this age (Pang &
泰勒, 2000; Ponton, Eggermont, Kwong, & 大学教师, 2000).
通常, the amplitude of the P1, N1b, and P2 compo-
nents of the auditory evoked potential increases until
the age of 10–12 years and remains stable (N1b and P2)
or decreases (P1) during adulthood. 而且, while P1
and N1 latencies typically decrease, P2 latency remains
stable and N2 latency increases as a function of age.
因此, it was of interest to compare the ERP effects found
in children during the critical period of development of
the auditory cortex with those previously found in
adults.
结果
Behavioral Data
Results of a three-way analysis of variance (ANOVA)
[expertise (two levels), 材料 (two levels), 和骗局-
gruity (三个级别)] on the transformed percentages of
error showed main effects of expertise [F(1,18) = 16.59,
p < .001], material [F(1,18) = 30.53, p < .001], and
congruity [F(2,36) = 36.05, p < .001]. Clearly, nonmu-
sician children (27%) made overall more errors than
musician children (12%), and both made more errors
with the musical (27%) than linguistic materials (12%).
Moreover, the error rate was highest for the weak
incongruity (see Figure 1). Most importantly, and as
predicted, musician children detected the weak incon-
gruity better than nonmusician children, not only in
music, but in language as well [Expertise (cid:1) Congruity
interaction: F(2,36) = 4.47, p = .01, with no Expertise (cid:1)
Material (cid:1) Congruity interaction, p < .38].
Electrophysiological Data
Mean amplitude ERPs to final note/word were measured
in several latency bands (100–200, 200–400, and 400–
700 msec) determined from both visual inspection and
based on previous results. Results were analyzed sepa-
rately for musicians and nonmusicians and for the
linguistic and musical materials,1 using ANOVAs that
included congruity (three levels: congruous, weakly
incongruous, and strongly incongruous) and electrodes
(four levels: Fz, Cz, Pz, and Oz) as within-subject factors
for midline analyses. ANOVAs were also computed for
lateral electrodes, using six regions of interest (ROIs):
left and right fronto-central (F3, F7, Fc5, and F4, F8, Fc6,
respectively), left and right temporal (C3, T3, Cp5, and
C4, T4, Cp6, respectively), and left and right temporo-
parietal (Cp1, P3, T5, and Cp2, P4, T6, respectively).
ANOVAs were computed for lateral electrodes using
congruity (three levels), hemispheres (two levels: left
and right),
fronto-central,
temporal, and temporo-parietal), and electrodes (three
for each ROI, as described above) for lateral analyses.
All p values were adjusted with the Greenhouse–Geisser
epsilon correction for nonsphericity when necessary.
When the factor congruity was significant or interacted
with other factors, planned comparisons between pairs
of conditions were computed. To simplify the presen-
tation of the results, outcomes of the main ANOVAs
in the different latency ranges are reported in Tables 1
and 2. When the main effects or interactions are signifi-
cant, results of two by two comparisons are presented
in text.
localization (three levels:
Music
For musician children, the ERPs associated to the final
notes clearly differ as a function of congruity in all
latency bands (100–200, 200–400, and 400–700 msec)
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
2
-
1
p
9
d
9
f
/
1
1
9
8
3
5
/
7
2
0
/
3
1
9
o
9
c
/
n
1
2
7
0
5
0
6
6
0
1
3
8
3
/
2
j
1
o
9
c
9
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
2
o
.
n
1
0
9
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
f
.
t
.
.
Figure 1. Percentage of error
rates for congruous (Cong)
final notes or words and for
weak and strong incongruities
in music and language are
presented for musicians and
nonmusicians. Clearly, in
both music and language, the
percentage of errors to weak
incongruities was significantly
higher for nonmusicians than
for musicians.
o
n
1
8
M
a
y
2
0
2
1
Magne, Scho¨n, and Besson
201
Table 1. Results of Main ANOVAs for Music
Latency Bands
Electrodes
Factors
Musicians
Nonmusicians
100–200 msec
200–400 msec
400–700 msec
Midlines
Laterals
Midlines
Laterals
Midlines
Laterals
C
C
C
C
C
F(2,18) = 10.44, p = .001
F(2,18) = 11.83, p < .001
F(2,18) = 16.50, p < .001
F(2,18) = 5.12, p = .019
ns
ns
ns
ns
F(2,18) = 10.93, p = .001
F(2,18) = 4.59, p = .026
C (cid:1) L
F(4,36) = 4.50, p = .010
F(4,36) = 5.13, p = .014
C = Congruity, L = localization (three regions of interest: fronto-central, temporal, and temporo-parietal).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
considered for analysis (see Table 1 for results of main
ANOVAs). Compared with congruous notes, weak in-
congruities elicited a larger early negative component,
between 200 and 400 msec, with maximum amplitude
around 340 msec [midlines: F(1,9) = 23.20, p < .001;
laterals: F(1,9) = 6.92, p < .027; see Figures 2 and 3]. This
negative effect was well distributed over the scalp, as
suggested by the absence of any significant Congruity (cid:1)
Localization interactions at lateral electrodes.
Strong incongruities also elicited a larger early nega-
tive component than congruous notes, with maximum
amplitude around 210 msec. This effect was significant
earlier, between 100 and 200 msec, than for the weak in-
congruities and was broadly distributed across scalp sites
[midlines: F(1,9) = 22.56, p < .001; laterals: F(1,9) =
19.23, p < .001; no Congruity (cid:1) Localization interaction
at lateral electrodes; see Figure 2]. Moreover, this early
negative component was followed by an increased pos-
itivity that differed from the ERPs to congruous notes
as early as 200–400 msec at midline sites [F(1,9) = 6.06,
p = .036]. This effect extended in the 400- to 700-msec
range and was significant at both midlines [F(1,9) =
19.67, p < .001] and lateral electrodes [Congruity (cid:1)
Localization interaction: F(2,18) = 8.01, p = .003], with a
temporo-parietal distribution [F(1,9) = 11.89, p = .007;
see Figure 3].
In contrast to musician children, the ERPs to weak
incongruities in nonmusicians did not differ from con-
gruous notes in any of the latency bands considered for
analysis (see Figure 2). However, strong incongruities
elicited an early negative component, peaking around
250 msec. This effect was significant later (in the 200- to
400-msec latency band) than in musicians and was larger
over the right hemisphere [Congruity (cid:1) Hemisphere
interaction: F(1,9) = 4.97, p = .05, see Figures 2 and 3].
This early negative component was also followed by
an increased positivity compared with congruous note,
but that started later, in the 400- to 700-msec range, than
for musician children [midlines: F(1,9) = 7.82, p = .02;
laterals: Congruity (cid:1) Localization interaction, F(2,18) =
13.71, p = .003]. This positive effect was localized over
the temporal and temporo-parietal sites bilaterally
[temporal: F(1,9) = 17.77, p = .002; temporo-parietal:
F(1,9) = 22.24, p = .001, see Figures 2 and 3].
Language
For musician children, the main effect of congruity was
significant in both the 200- to 400-msec and the 400- to
700-msec ranges (see Table 2). Both weak and strong
prosodic incongruities elicited larger positivities than
congruous endings between 200 and 700 msec (see
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
2
-
1
p
9
d
9
f
/
1
1
9
8
3
5
/
7
2
0
/
3
1
9
o
9
c
/
n
1
2
7
0
5
0
6
6
0
1
3
8
3
/
2
j
1
o
9
c
9
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
2
o
.
n
1
0
9
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
f
.
.
.
.
t
Table 2. Results of Main ANOVAs for Language
Latency Bands
Electrodes
Factors
Musicians
Nonmusicians
100–200 msec
200–400 msec
400–700 msec
Midlines
Laterals
Midlines
Laterals
Midlines
Laterals
C
C
C
ns
ns
ns
ns
F(2,18) = 10.88, p = .001
F(2,18) = 3.86, p = .041
C (cid:1) L
F(4,36) = 4.65, p = .022
ns
C
C
F(2,18) = 11.84, p < .001
F(2,18) = 4.27, p = .032
F(2,18) = 5.67, p = .015
ns
C (cid:1) L
F(4,36) = 6.69, p = .006
F(4,36) = 3.90, p = .043
C = Congruity, L = localization (three regions of interest: fronto-central, temporal, and temporo-parietal).
o
n
1
8
M
a
y
2
0
2
1
202
Journal of Cognitive Neuroscience
Volume 18, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
2
-
1
p
9
d
9
f
/
1
1
9
8
3
5
/
7
2
0
/
3
1
9
o
9
c
/
n
1
2
7
0
5
0
6
6
0
1
3
8
3
/
2
j
1
o
9
c
9
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
2
o
.
n
1
0
9
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
t
.
f
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Illustration of the variations in brain electrical activity time-locked to final note onset and elicited by congruous endings,
weak incongruities, or strong incongruities. Each trace represents an average of electrophysiological data recorded from 10 musician and
10 nonmusician 8-year-old children. EEG was recorded from 28 electrodes; selected traces from 9 electrodes are presented. In this figure,
as in the following ones, the amplitude (in microvolts) is plotted on the ordinate (negative up) and the time (in milliseconds) is on the
abscissa. White arrows point to the effects that are present for both musician and nonmusician children, whereas black arrows show effects
that are present for musicians only.
Figure 4). This positive effect was largest over the
midline sites for the weak incongruity [200–400 msec:
F(1,9) = 9.42, p = .013; 400–700 msec: F(1,9) = 8.90,
p = .015; see Figure 5] and was broadly distributed over
the scalp, with a bilateral temporo-parietal distribution
for the strong incongruities (Congruity (cid:1) Localization
interaction, 200–400 msec: F(2,18) = 8.18, p = .015,
400–700 msec: F(2,18) = 16.65, p < .001; results of post
hoc comparisons in the temporal and temporo-parietal
ROIs always revealed significant differences at p < .05).
Although the main effect of congruity was also sig-
nificant for nonmusician children in both the 200- to
400-msec and the 400- to 700-msec ranges (see Table 2)
results of 2 (cid:1) 2 comparisons showed that only the ERPs
associated to strong incongruities elicited larger positiv-
ities than congruous endings (see Figure 4). This effect
was significant between 200 and 700 msec at midline sites
[200–400 msec: F(1,9) = 9.57, p = .012; 400–700 msec:
F(1,9) = 11.27, p = .008] and between 400 and 700 msec
at lateral sites [Congruity (cid:1) Localization interaction:
F(2,18) = 12.32, p < .001], with a bilateral temporo-
parietal maximum [F(1,9) = 15.56, p = .003; see Figure 5].
Finally, results of ANOVAs performed in successive
25-msec latency bands between 200 and 400 msec over
the midline sites revealed that the positive differences
between strong incongruities and congruous endings
started earlier for musician (275–300 msec, p < .01) than
nonmusician children (350–375 msec, p < .01; see
Table 3).
DISCUSSION
In line with our hypotheses, error rate analyses showed
that musician children outperformed nonmusician chil-
dren in the detection of weak incongruities, not only in
music, but also in language, thereby pointing to a
Magne, Scho¨n, and Besson
203
Figure 3. Topographic maps
of the weak incongruity effect
(mean amplitude difference
between weak incongruity and
congruous ending) and strong
incongruity effect (mean
amplitude difference between
strong incongruity and
congruous ending) in music
for musicians (top) and
nonmusicians (bottom). In
the three latency windows
considered for analyses
(100–200, 200–400, and
400–700 msec), only significant
effects are represented.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
2
-
1
p
9
d
9
f
/
1
1
9
8
3
5
/
7
2
0
/
3
1
9
o
9
c
/
n
1
2
7
0
5
0
6
6
0
1
3
8
3
/
2
j
1
o
9
c
9
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
2
o
.
n
1
0
9
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
t
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
common pitch processing mechanism in language and
music perception. In line with these behavioral data,
ERPs analyses also showed greatest differences between
the two groups of children for the weak incongruity. In
this case, both an early negative component in music
and a late positive component in language were found
only for musician children. By contrast, early negative
and late positive components were elicited by strong
incongruities in both groups, although with some quan-
titative differences. These results are considered in turn
in the following discussion.
Effects of Musical Training on the Detection
of Pitch Changes in Music and Language
Behavioral data clearly showed that the overall level of
performance in the pitch detection task was higher for
musician than nonmusician children. This difference was
expected in the music task because musician children
had 4 years of musical training on average, and previous
reports have highlighted the positive effect of musical
expertise on music perception in both adults and chil-
dren (Scho¨n et al., 2004; Thompson, Schellenberg, &
Husain, 2003, 2004; Besson & Faı¨ta, 1995, but see also
Bigand, Parncutt, & Lerdahl, 1996, for evidence in adults
that suggests otherwise). What is most striking is that
musicians’ performance was also better in language.
Because Schellenberg (2004) recently showed that 1 year
of musical training significantly improved IQ, one could
argue that general nonspecific processes are at play,
which explains why musician children outperformed
nonmusician children. In this case, however, one would
expect differences between the two groups of children
in the three experimental conditions. The present re-
sults show that this is not the case: The only significant
difference between the two groups was found for the
weak incongruity, which is clearly the most difficult to
detect. In this condition, for both music and language,
the level of performance of musician children was
twice as high as for nonmusician children. Therefore,
intelli-
although music training may improve general
gence (Schellenberg, 2004), it also seems to exert spe-
cific beneficial influences on both music and language
perception. Although positive transfer effects between
music and other cognitive domains have already been
reported in the literature, as mentioned in the Intro-
duction, the causal links underlying these effects have
not been directly tested. Here, we provide evidence that
music training, by increasing sensitivity to a specific basic
acoustic parameter, pitch, which is equally important for
music and speech prosody, does enhance children’s
ability to detect pitch changes not only in music, but
also in language.
The present results also extend those recently re-
ported by Thompson et al. (2004), which showed that
1 year of musical training allowed 6-year-olds to identify
emotional prosody in utterances better than the non-
musician control group. Thus, evidence for positive
transfer effects between music and language is increas-
ingly being shown when basic acoustic parameters,
such as pitch, intensity, or duration, are manipulated
in both domains.
204
Journal of Cognitive Neuroscience
Volume 18, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
2
-
1
p
9
d
9
f
/
1
1
9
8
3
5
/
7
2
0
/
3
1
9
o
9
c
/
n
1
2
7
0
5
0
6
6
0
1
3
8
3
/
2
j
1
o
9
c
9
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
2
o
.
n
1
0
9
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
f
.
.
.
t
.
o
n
1
8
M
a
y
2
0
2
1
Figure 4. Illustration of the variations in brain electrical activity time-locked to final word onset and elicited by congruous endings,
weak incongruities, or strong incongruities. Each trace represents an average of electrophysiological data recorded from 10 musician and
10 nonmusician children.
Neurophysiological Basis of Positive
Transfer Effects
In line with the behavioral data, ERPs analyses showed
that the differences between musician and nonmusician
children were larger for the weak incongruity than for
both congruous endings and strong incongruities. In-
deed, although for musician children, weak incongrui-
ties elicited a larger early negativity than congruous
notes in music and a larger late positivity than congru-
ous words in language, no such differences were found
for nonmusician children in either music or language
(see Figures 3 and 5). Therefore, when pitch violations
are most difficult to detect, different processes seem to
be involved as a function of musical expertise.
By contrast, similar ERP patterns were elicited by the
strong incongruity for both musician and nonmusician
children. In music, early negativities were followed by
late positivities in both groups. Importantly, however,
precise analyses of their time course and scalp distribu-
tion also revealed some quantitative differences. First,
the onset of the early negative effect was 100 msec
shorter (significant between 100 and 200 msec for musi-
cians and between 200 and 400 msec for nonmusicians)
and the onset of the late positive effect was 200 msec
shorter (significant between 200 and 400 msec for
musicians and between 400 and 700 msec for nonmu-
sicians) for musician than nonmusician children. Sec-
ond, although the early negative effect was broadly
distributed over the scalp for musician children, it was
localized over the right hemisphere for nonmusician
children. Although right lateralization for pitch process-
ing is in line with some results in the literature (see
Zatorre et al., 2002), the lateralized distribution reported
here may result from an overlap of the early negative
components by subsequent later positivities that devel-
oped over left fronto-central regions, thereby reducing
the negativity over the left hemisphere (see Figure 3).
Magne, Scho¨n, and Besson
205
Figure 5. Topographic maps
of the weak incongruity effect
(mean amplitude difference
between weak incongruity and
congruous ending) and strong
incongruity effect (mean
amplitude difference between
strong incongruity and
congruous ending) in language
for musicians (top) and
nonmusicians (bottom).
Only significant effects are
represented.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
2
-
1
p
9
d
9
f
/
1
1
9
8
3
5
/
7
2
0
/
3
1
9
o
9
c
/
n
1
2
7
0
5
0
6
6
0
1
3
8
3
/
2
j
1
o
9
c
9
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
2
o
.
n
1
0
9
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
f
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
In language, the strong incongruity elicited a larger
late positive component than the congruous word for
both musician and nonmusician children. The precise
analysis of the time course of this positive effect revealed
that it started 75 msec earlier and was larger for the
musician than nonmusician groups, although with a
similar temporo-parietal scalp distribution.
Taken together, these results clearly show both qual-
itative (scalp distribution) and quantitative (latency
differences) differences between musician and nonmu-
sician children. It is also interesting to note that the late
positivity is overall larger and lasts longer for nonmusi-
cians than musicians, which may reflect the fact that
nonmusicians need more processing resources to per-
Table 3. Timing of the Strong Incongruity Effect in Language
Latencies (msec)
Musicians
Nonmusicians
200–225
225–250
250–275
275–300
300–325
325–350
350–375
375–400
* p < .01.
** p < .001.
–
–
–
*
*
**
**
**
–
–
–
–
–
–
*
**
form the tasks and that processing takes longer than
for musicians.
Developmental Perspective
The functional significance of the results reported above
is now considered in light of previous results found with
adults performing the same tasks with the same materi-
als (Scho¨n et al., 2004). Considering first the differences,
the overall ERP amplitude was larger and the latency of
the ERP components was longer in children than in
adults. Consider, for instance, the negative component
to strong musical incongruity at the electrode T4 where
it is clearly defined. The mean amplitude of this nega-
tivity was (cid:2)7.35 AV, and its peak latency 255 msec for
children (musicians and nonmusicians), whereas it was
(cid:2)4.10 AV and 165 msec, for adults. These results are in
line with a large literature showing decreased ampli-
tude and shortened latency of ERP components as age
progresses (see Taylor, 1995, for a review). Decreases in
amplitude are thought to depend upon the number of
pyramidal cell synapses contributing to postsynaptic
potentials (Ponton et al., 2000) and are interpreted as
reflecting the automation of the underlying processes
that thereby require fewer and fewer neurons (Batty &
Taylor, 2002). Decreases in latency may result from
increased speed of nervous transmission, due to axon
myelinization, as well as to the maturation of synaptic
connections, due to the repeated synchronization of
specific neuronal populations (Batty & Itier, 2004;
Taylor, 1995; Courchesne, 1990; Eggermont, 1988). In
sum, the overall decreased amplitude and shortened
206
Journal of Cognitive Neuroscience
Volume 18, Number 2
latency of ERPs with age may reflect an enhanced effi-
ciency of cognitive processing over the course of devel-
opment. Moreover, the differences between musicians
and nonmusician children in the amplitude and latency
of the early negative components elicited by the weak
and strong incongruities in music are in line with recent
results by Shahin, Roberts, and Trainor (2004) showing
overall enhanced amplitude of the early ERP compo-
nents with musical practice in 4- to 5-year-old children.
Interestingly, these results also showed that the increase
in amplitude of the N1 and P2 components was specific
to the instrument played.
Regarding the present series of experiments, results
revealed differences in the early negative components
between the adults tested by Scho¨n et al. (2004) and the
children tested here when they perform the same
explicit task (pitch congruity judgment) on the same
materials. In adults, early negative components were
elicited, between 50 and 200 msec, by strong incongru-
ities both in music and in language. In music, they were
distributed over the right temporal regions, whereas
they were distributed over the temporal regions bilater-
ally in language. By contrast, for children, they were only
found in music.
The functional interpretation of these early negativ-
ities is still a matter of debate. Previous results in adults
have shown that both harmonic (Koelsch, Gunter,
Friederici, & Schro¨ger, 2000; Patel et al., 1998) and
melodic incongruities (Scho¨n et al., 2004) elicit an early
negative component over right frontal sites between 200
and 400 msec. Moreover, results of a study with 5- and
9-year-old nonmusician children have also shown that
harmonic violations elicited early negative components
(Koelsch et al., 2003). Finally, the finding that these early
negativities were typically elicited in musical contexts
and were larger for participants with than without
formal musical training led the authors to propose that
this early negativity may reflect specific musical expec-
tancies (Koelsch, Schmidt, & Kansok, 2002). However,
many results in the literature have also demonstrated
that unexpected changes in the basic acoustic properties
of sounds, such as frequency,
intensity, or duration,
elicit an early automatic brain response, the mismatch
negativity (Na¨a¨ta¨nen, 1992). Therefore, the issue of
whether the early negativity reflects specific musical
expectancies or a domain general mismatch detection
process remains an open question (Koelsch, Maess,
Grossmann, & Friederici, 2002; Koelsch, Schro¨ger, &
Gunter, 2002). Because early negative components were
found in response to pitch violations in both language
and music in our previous experiment with adults
(Scho¨n et al., 2004), we favor the interpretation fol-
lowing which they reflect automatic aspects of pitch
processing in both domains. However, how can we
reconcile such an interpretation with the present results
with children showing an early negativity to pitch devia-
tions in music, but no such component in language? This
matter raises the intriguing possibility that automatic
detection of pitch changes in music may be functional
earlier on (as early as 5–8 years old) than in language.
Several authors have emphasized the importance of
melodic elements in infant-directed speech and for
language acquisition (Trehub, 2003; Papousek, 1996;
Jusczyk & Krumhansl, 1993). Thus, the development of
the early negativity in both music and language needs to
be tested in further experiments using a longitudinal
approach with children ages 4, 6, 8, and 10 years.
Turning to the similarities, results with children
showed that, as was previously found with adults, strong
incongruities elicited late positive components with a
centro-parietal distribution in both music and language.
Therefore, in contrast with the processes underlying the
negative components, the processes underlying the
occurrence of these late positivities seem to be present
already at age 8 years in both music and language. Based
on numerous results in the ERP literature, the occur-
rence of these late positivities (P3b component) is
generally considered as being related to the processing
of surprising and task-relevant events (Picton, 1992;
Duncan-Johnson & Donchin, 1977; see Donchin &
Coles, 1988, for a review). Moreover, the latency of
these positive components often varies with the diffi-
culty of the categorization task (Kutas, MacCarthy, &
Donchin, 1977), which is in line with our previous and
present results showing shorter latencies for the strong
than weak incongruities.
Conclusions
The most important conclusion to be drawn from these
results is that we found behavioral evidence for a
common pitch processing mechanism in language and
music perception. Moreover, by showing qualitative and
quantitative differences in the ERPs recorded from mu-
sician and nonmusician children, we were able to un-
cover some of the neurophysiological processes that
may underlie positive transfer effects between music
and language. The occurrence of an early negative
component to the weak incongruity in music for musi-
cian children only may indeed reflect a greater sensitivity
to pitch processing. Such enhanced pitch sensitivity in
musician children would also be reflected by the larger
late positivity to weak incongruities than congruous
words in language that was not found in nonmusician
children. Although these findings may reflect the facili-
tation, due to musical training, of domain-general pitch
mismatch detection processes common to both music
and language, further experiments are needed to specify
the relationships between the early negative and late
positive components and why early negative compo-
nents were elicited by strong incongruities in both
musician and nonmusician children in music but not
in language.
Magne, Scho¨n, and Besson
207
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
2
-
1
p
9
d
9
f
/
1
1
9
8
3
5
/
7
2
0
/
3
1
9
o
9
c
/
n
1
2
7
0
5
0
6
6
0
1
3
8
3
/
2
j
1
o
9
c
9
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
2
o
.
n
1
0
9
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
.
.
f
t
o
n
1
8
M
a
y
2
0
2
1
To summarize, these results add to the body of
cognitive neuroscience literature on the beneficial ef-
fects of musical education; in particular, the present
findings highlight the positive effects of music lessons
for linguistic abilities in children. Therefore, these find-
ings argue in favor of music classes being an intrinsic and
important part of the educational programs in public
schools and in all the institutions that aim at improving
children’s perceptive and cognitive abilities. Finally, the
present study also confirms that the ERP method is
particularly well adapted for the exploration of positive
transfer effects between music processing and other
cognitive domains. Further research is also needed to
determine the extent of these transfers, as well as their
existence between music cognition and nonauditory
processes such as visuospatial reasoning.
METHODS
Participants
Twenty-six children (14 girls and 12 boys; age 8 ±
1 years), 13 musicians and 13 nonmusicians, participated
in the experiment, which lasted for about 2 hr. The
musician children had 4 ± 1 years of musical training on
average. All children were right-handed, had normal
hearing, and were native speakers of French. Most
importantly, all the children came from the same ele-
mentary school and had similar socioeconomic back-
grounds (e.g., a t test on the mean family incomes
revealed no significant differences between the two
groups, p = .74). All musician children played an in-
strument (violin = 5, guitar = 2, flute = 1, clarinet = 2,
harp = 1, piano = 2), which they regularly practiced
everyday for around 20 to 30 min. They also took music
lessons twice a week for a half an hour. Thus, these
children played music for about 3–4 hr per week. All
nonmusician children also had regular extracurricular
activities (judo = 2, swimming = 2, cycling = 2, tennis =
1, rugby = 1, rollerblading = 1, circus training = 1,
gymnastics = 1, horseback riding = 1, soccer = 1). Six of
the participants (three musicians and three nonmusi-
cians) were not included in the analyses because of
technical problems or too many artifacts during the
electroencephalogram (EEG) recording session. Chil-
dren were given presents at the end of the recording
session. All parents gave informed consent for their
children to participate in the experiment.
Stimuli
Stimuli comprised 96 French-spoken declarative sen-
tences taken from children’s books and ending with
bisyllabic words (e.g., ‘‘Dans la barque se tient l’enemi
de Peter Pan, le terrible pirate’’/‘‘In the boat is the en-
emy of Peter Pan, the terrible pirate’’). Sentences were
spoken at a normal speech rate by a native French
female speaker, recorded in a soundproof room using
a digital audiotape (sampling at 44.1 kHz), and synthe-
sized using the software Winpitch (Martin, 1996). The
mean duration of the sentence was 3.97 ± 0.7 sec.
A total of 96 melodies were also presented in the
experiment. Half were selected from the repertoire of
children’s music (e.g. ‘‘Happy Birthday’’), and half were
composed for the experiment by a professional musi-
cian, following the same rules of composition as for
familiar melodies. Tunes were converted into MIDI files
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
2
-
1
p
9
d
9
f
/
1
1
9
8
3
5
/
7
2
0
/
3
1
9
o
9
c
/
n
1
2
7
0
5
0
6
6
0
1
3
8
3
/
2
j
1
o
9
c
9
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
2
o
.
n
1
0
9
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
.
.
f
t
o
n
1
8
M
a
y
2
0
2
1
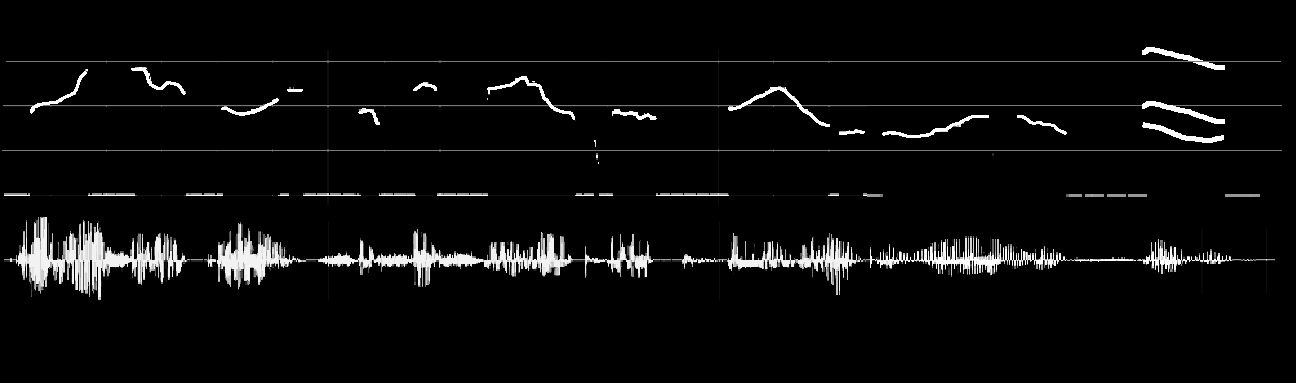
Figure 6. Examples of stimuli used in the experiment. (A) The speech signal is illustrated for the sentence: ‘‘Un loup solitaire se faufile entre
les troncs de la grande foreˆt’’ [literal translation: ‘‘A lonely wolf worked his way through the trees of the big forest’’]. (B) The musical notation
is illustrated for the song ‘‘Happy Birthday.’’
208
Journal of Cognitive Neuroscience
Volume 18, Number 2
using the synthetic sound of a piano (KORG XDR5,
Tokyo, Japan). The mean duration of the melodies was
10.3 ± 2.44 sec.
An equal number of sentences/melodies (32) were
presented in each of the three following experimental
conditions, thus leading to a total of 192 stimuli with 96
sentences and 96 melodies: The final word or note was
prosodically or melodically congruous, weakly incongru-
ous, or strongly incongruous (see Figure 6A). Based
upon results of pretests of a preliminary version of this
material with both adults and children, the F0 of the
last word was increased, using the software WinPitch,
by 35% for the weak incongruity and by 120% for the
strong incongruity (without changing the original pitch
contour). In the musical material, the last note was
increased by one fifth of a tone for the weak incon-
gruity and by half of a tone for the strong incongruities
using the sound file editor software Wavelab, Hamburg,
Germany (see Figure 6B).
Procedure
In eight separate blocks of trials, children were required
to listen attentively, through headphones, either to the
melodies (four blocks) or the sentences (four blocks).
Within each block of trials, stimuli were presented in a
pseudorandom order, and children were asked to de-
cide whether the last word or note seemed normal or
strange (i.e., something was wrong), by pressing one of
two response keys as quickly and as accurately as
possible. The hand of response and the order of pre-
sentation (musical or prosodic materials first) were
counterbalanced across children.
Event-related Brain Potential Recordings
EEG was recorded for 2200 msec starting 150 msec
before the onset of the last word/note, from 28 scalp
electrodes, mounted on a child-sized elastic cap and
located according to the International 10/20 system.
These recording sites plus an electrode placed on the
right mastoid were referenced to the left mastoid elec-
trode. The data were then rereferenced offline to the
algebraic average of the left and right mastoids. Imped-
ances of the electrodes never exceeded 3 k(cid:1). To detect
blinks and vertical eye movements, the horizontal elec-
trooculogram (EOG ) was recorded from electrodes
placed 1 cm to the left and right of the external canthi,
and the vertical EOG was recorded from an electrode
beneath the right eye, referenced to the left mastoid.
Trials containing ocular or movement artifacts, or am-
plifier saturation, were excluded from the averaged ERP
waveforms. The EEG and EOG were amplified by an SA
Instrumentation amplifier with a bandpass of 0.01–30 Hz
and were digitized at 250 Hz by a PC-compatible micro-
computer (Compaq Prosignia 486, Hewlett-Packard Co.,
Palo Alto, CA).
Acknowledgments
This research was first supported by a grant from the In-
ternational Foundation for Music Research (IFRM: RA 194) and
later by a grant from the Human Frontier Science Program to
Mireille Besson (HSFP: RGP0053). Cyrille Magne benefited
from a research fellowship from the Cognitive Program of
French Ministry of Research, and Daniele Scho¨n was a post-
doctorate student supported by the HFSP grant. The authors
acknowledge Monique Chiambretto and Reyna Leigh Gordon
for their technical assistance.
Reprint requests should be sent to Cyrille Magne, Center for
Complex Systems and Brain Sciences, Florida Atlantic Univer-
sity, 777 Glades Road, Boca Raton, FL 33431, USA, or via e-mail:
magne@ccs.fau.edu.
Note
1. Because latency, amplitude, and scalp distribution differ-
ences were found between musician and nonmusician children
and between the linguistic and musical materials, results of a
general ANOVA including expertise and materials as factors are
less informative than computing the analyses for each group
and each material separately.
REFERENCES
Aleman, A., Nieuwenstein, M. R., Bo¨cker, K. B. E., & Hann,
E. H. F. (2000). Music training and mental imagery ability.
Neuropsychologia, 38, 1664–1668.
Batty, B., & Itier, R. J. (2004). Les modifications des
potentiels e´voque´s cognitifs au cours du de´veloppement.
In B. Renault (Ed.), L’imagerie fonctionnelle e´lectrique
(EEG) et magne´tique (MEG): Ses applications en sciences
cognitives (pp. 217–234). Paris: Herme´s.
Batty, M., & Taylor, M. J. (2002). Visual categorization during
childhood: An ERP study. Psychophysiology, 39, 1–9.
Besson, M., & Faı¨ta, F. (1995). An event-related potential (ERP)
study of musical expectancy: Comparison of musicians with
non-musicians. Journal of Experimental Psychology:
Human Perception and Performance, 21, 1278–1296.
Besson, M., & Scho¨n, D. (2001). Comparison between
language and music. Annals of the New York Academy of
Sciences, 930, 232–259.
Bigand, E., Parncutt, R., & Lerdahl, F. (1996). Perception of
musical tension in short chord sequences: The influence of
harmonic function, sensory dissonance, horizontal motion,
and musical training. Perception and Psychophysics, 58,
125–141.
Bilhartz, T. D., Bruhn, R. A., & Olson, J. E. (2000). The effect
of early music training on child cognitive development.
Journal of Applied Developmental Psychology, 20,
615–636.
Brochard, R., Dufour, A., & Despre`s, O. (2004). Effect of
musical expertise on visuospatial abilities: Evidence from
reaction times and mental imagery. Brain & Cognition, 54,
103–109.
Chan, A. S, Ho, Y. C., & Cheung, M. C. (1998). Music training
improves verbal memory. Nature, 396, 128.
Costa-Giomi, E. (2004). Effects of three years of piano
instruction on children’s academic achievement, school
performance and self-esteem. Psychology of Music, 32,
139–152.
Costa-Giomi, E. (1999). The effects of three years of piano
instruction on children’s cognitive development. Journal of
Research in Music Education, 47, 198–212.
Magne, Scho¨n, and Besson
209
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
2
-
1
p
9
d
9
f
/
1
1
9
8
3
5
/
7
2
0
/
3
1
9
o
9
c
/
n
1
2
7
0
5
0
6
6
0
1
3
8
3
/
2
j
1
o
9
c
9
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
2
o
.
n
1
0
9
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
f
t
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Courchesne, E. (1990). Chronology of postnatal human brain
development: Event-related potentials, positron emission
tomography, myelogenesis, and synaptogenesis studies.
In J. W. Rohrbaugh, R. Parasuraman, & R. Johnson (Eds.),
Event-related brain potentials (pp. 210–241). Oxford:
Oxford University Press.
Culham, J. C., & Kanwisher, N. G. (2001). Neuroimaging of
cognitive functions in human parietal cortex. Current
Opinion in Neurobiology, 11, 157–163.
Cupchick, G. C., Philips, K., & Hill, D. S. (2001). Shared
processes in spatial rotation and musical permutation. Brain
& Cognition, 46, 373–382.
Dehaene, S., Spelke, E., Pinel, P., Stanescu, R., & Tsivkin, S.
(1999). Sources of mathematical thinking: Behavioral and
brain-imaging evidence. Science, 284, 970–974.
Donchin, E., & Coles, M. G. H. (1988). Is the P300 component
a manifestation of context-updating? Behavioral and
Brain Science, 11, 355–372.
Duncan-Johnson, C., & Donchin, E. (1977). On quantifying
surprise, the variation of event-related potentials with
subjective probability. Psychophysiology, 14, 456–467.
Eggermont, J. J. (1988). On the maturation of sensory
evoked potentials. Electroencephalography and Clinical
Neurophysiology, 70, 293–305.
Elbert, T., Pantev, C., Wienbruch, C., Rockstroh, B., & Taub,
E. (1995). Increased cortical representation of the
fingers of the left hand in string players. Science, 270,
305–307.
Gardiner, M. F., Fox, A., Knowles, F., & Jeffrey, D. (1996).
Learning improved by arts training. Nature, 381, 284.
Gaser, C., & Schlaug, G. (2003). Brain structures differ
between musicians and non-musicians. Journal of
Neuroscience, 23, 9240–9245.
Graziano, A. B., Peterson, M., & Shaw, G. L. (1999). Enhanced
learning of proportional math through music training and
spatial-temporal training. Neurological Research, 21,
139–152.
Koelsch, S., Schro¨ger, E., & Gunter, T. (2002). Music matters:
Preattentive musicality of the human brain.
Psychophysiology, 39, 1–11.
Kutas, M., McCarthy, G., & Donchin, E. (1977). Augmenting
mental chronometry: The P300 as a measure of stimulus
evaluation time. Science, 197, 792–795.
Maess, B., Koelsch, S., Gunter, T. C., & Friederici, A. D. (2001).
Musical syntax is processed in Broca’s area: An MEG study.
Nature Neuroscience, 4, 540–545.
Martin, P. (1996). WinPitch: Un logiciel d’analyse temps re´el de
la fre´quence fondamentale fonctionnant sous Windows.
Actes des XXIV Journe´es d’Etude sur la Parole, 224–227.
Avignon, France.
Meyer, M., Alter, K., Angela, A. D., Lohmann, G., & von
Cramon, D. Y. (2002). fMRI reveals brain regions mediating
slow prosodic modulations in spoken sentences. Human
Brain Mapping, 17, 73–88.
Na¨a¨ta¨nen, R. (1992). Attention and brain function. Hillsdale,
NJ: Erlbaum.
Ohnishi, T., Matsuda, H., Asada, T., Aruga, M., Hirakata, M.,
Nishikawa, M., Katoh, A., & Imabayashi, E. (2001). Functional
anatomy of musical perception in musicians. Cerebral
Cortex, 11, 754–760.
Pang, E. W., & Taylor, M. J. (2000). Tracking the development
of the N1 from age 3 to adulthood: An examination of
speech and non-speech stimuli. Clinical Neurophysiology,
111, 388–397.
Pantev, C., Oostenveld, R., Engelien, A., Ross, B., Roberts, L. E.,
& Hoke, M. (1998). Increased auditory cortical
representation in musicians. Nature, 392, 811–814.
Papousek, M. (1996). Intuitive parenting: A hidden source of
musical stimulation in infancy. In I. Deliege & J. Sloboda
(Eds.), Musical beginnings: Origins and development of
musical competence (pp. 88–112). Oxford, New York.
Patel, A. D. (2003a). Rhythm in language and music: Parallels
and differences. Annals of the New York Academy of
Sciences, 999, 140–143.
Gromko, J. E., & Poorman, A. (1998). The effect of music
Patel, A. D. (2003b). Language, music, syntax and the brain.
training on preschoolers’ spatial temporal task performance.
Journal of Research in Music Education, 46, 173–181.
Hetland, L. (2000). Learning to make music enhances spatial
reasoning. Journal of Aesthetic Education, 34, 179–238.
Ho, Y.-C., Cheung, M.-C., & Chan, A. S. (2003). Music
training improves verbal but not visual memory:
Cross-sectional and longitudinal explorations in children.
Neuropsychology, 17, 439–450.
Nature Neuroscience, 6, 674–681.
Patel, A., Gibson, E., Ratner, J., Besson, M., & Holcomb,
P. (1998). Processing syntactic relations in language and
music: An event-related potential study. Journal of
Cognitive Neuroscience, 10, 717–733.
Picton, T. W. (1992). The P300 wave of the human event-
related potential. Journal of Clinical Neurophysiology, 9,
456–479.
Jusczyk, P. W., & Krumhansl, C. L. (1993). Pitch and rhythmic
Ponton, C. W., Eggermont, J. J., Kwong, B., & Don, M. (2000).
patterns affecting infants’ sensitivity to musical phrase
structure. Journal of Experimental Psychology: Human
Perception and Performance, 19, 627–640.
Koelsch, S. (2005). Neural substrates of processing syntax and
semantics in music. Current Opinion in Neurobiology, 15,
207–212.
Koelsch, S., Grossmann, T., Gunter, T. C., Hahne, A., Schro¨ger,
E., & Friederici, A. D. (2003). Children processing music:
Electric brain responses reveal musical competence and
gender differences. Journal of Cognitive Neuroscience, 15,
683–693.
Koelsch, S., Gunter, T., Friederici, A. D., & Schro¨ger, E. (2000).
Brain indices of music processing: ‘‘Non-musicians’’
are musical. Journal of Cognitive Neuroscience, 12,
520–541.
Koelsch, S., Maess, B., Grossmann, T., & Friederici, A. (2002).
Sex difference in music-syntactic processing. NeuroReport,
14, 709–712.
Koelsch, S., Schmidt, B., & Kansok, J. (2002). Influences of
musical expertise on the ERAN: An ERP-study.
Psychophysiology, 39, 657–663.
Maturation of human central auditory system activity:
Evidence from multi-channel evoked potentials. Clinical
Neurophysiology, 111, 220–236.
Rauscher, F. H., Shaw, G. L., Levine, L. J., Wright, E. L.,
Dennis, W. R., & Newcomb, R. (1997). Music training
causes long-term enhancement of pre-school children’s
spatial–temporal reasoning. Neurological Research,
19, 2–8.
Schellenberg, E. G. (2004). Music lessons enhance HQ.
Psychological Science, 15, 511–514.
Schlaug, G., Jancke, L., Huang, Y., & Steinmetz, H. (1995).
In vivo evidence of structural brain asymmetry in
musicians. Science, 267, 699–701.
Schlaug, G., Jancke, L., Huang, Y., Staiger, J. F., & Steinmetz,
H. (1995). Increased corpus callosum size in musicians.
Neuropsychologia, 33, 1047–1055.
Schneider, P., Scherg, M., Dosch, H. G., Specht, H. J.,
Gutschalk, A., & Rupp, A. (2002). Morphology of
Heschl’s gyrus reflects enhanced activation in the
auditory cortex of musicians. Nature Neuroscience, 5,
688–694.
210
Journal of Cognitive Neuroscience
Volume 18, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
2
-
1
p
9
d
9
f
/
1
1
9
8
3
5
/
7
2
0
/
3
1
9
o
9
c
/
n
1
2
7
0
5
0
6
6
0
1
3
8
3
/
2
j
1
o
9
c
9
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
2
o
.
n
1
0
9
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
.
.
f
t
o
n
1
8
M
a
y
2
0
2
1
Scho¨n, D., Magne, C., & Besson, M. (2004). The music
of speech: Electrophysiological study of pitch perception
in language and music. Psychophysiology, 41, 341–349.
Shahin, A., Roberts, L. E., & Trainor, L. J. (2004). Enhancement
of auditory cortical development by musical experience in
children. NeuroReport, 15, 1917–1921.
Taylor, M. J. (1995). The role of event-related potentials in the
study of normal and abnormal cognitive development. In F.
Boller & J. Grafman (Eds.), Handbook of neuropsychology
(pp. 187–211). Amsterdam: Elsevier.
Thompson, W. F., Schellenberg, E. G., & Husain, G. (2003).
Perceiving prosody in speech: Effects of music lessons.
Annals of the New York Academy of Sciences, 999,
530–532.
Trehub, S. E. (2003). The developmental origins of musicality.
Nature Neuroscience, 6, 669–673.
Tzourio, N., Massioui, F. E., Crivello, F., Joliot, M., Renault, B.,
& Mazoyer, B. (1997). Functional anatomy of human
auditory attention studied with PET. Neuroimage, 5,
63–77.
Thompson, W. F., Schellenberg, E. G., & Husain, G. (2004).
Zatorre, R. J. (2002). Structure and function of auditory
Decoding speech prosody: Do music lessons help? Emotion,
4, 46–64.
cortex: Music and speech. Trends in Cognitive Science, 6,
37–46.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
8
e
2
-
1
p
9
d
9
f
/
1
1
9
8
3
5
/
7
2
0
/
3
1
9
o
9
c
/
n
1
2
7
0
5
0
6
6
0
1
3
8
3
/
2
j
1
o
9
c
9
n
p
.
d
2
0
b
0
y
6
g
.
u
1
e
8
s
.
t
2
o
.
n
1
0
9
8
9
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
f
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Magne, Scho¨n, and Besson
211