Mine and Me: Exploring the Neural Basis of
Object Ownership
David J. Turk, Kim van Bussel, Gordon D. Waiter, 和C. Neil Macrae
抽象的
■ Previous research has shown that encoding information in
the context of self-evaluation leads to memory enhancement,
supported by activation in ventromedial pFC. Recent evidence
suggests that similar self-memory advantages can be obtained
under nonevaluative encoding conditions, such as when object
ownership is used to evoke self-reference. Using fMRI, 当前的
study explored the neural correlates of object ownership. 期间
scanning, participants sorted everyday objects into self-owned or
other-owned categories. Replicating previous research, a signifi-
cant self-memory advantage for the objects was observed (IE。,
self-owned > other-owned). 此外, encoding self-owned
items was associated with unique activation in posterior dorso-
medial pFC (dMPFC), left insula, and bilateral supramarginal gyri
(SMG). Subsequent analysis showed that activation in a subset of
these regions (dMPFC and left SMG) correlated with the magni-
tude of the self-memory advantage. Analysis of the time-to-peak
data suggested a temporal model for processing ownership in
which initial activation of dMPFC spreads to SMG and insula.
These results indicate that a self-memory advantage can be elic-
ited by object ownership and that this effect is underpinned by
activity in a neural network that supports attentional, reward,
and motor processing. ■
介绍
Human identity is no longer defined by what one
做, but by what one owns (President Jimmy
Carter, Crisis of Confidence Speech, 15 七月 1979).
Ownership refers to the classification of a physical or
mental object as belonging to self and is a core facet of
human experience ( James, 1929). It pertains to our sense
of owning our bodies and movements and accumulated
material wealth. We can also claim ownership of ideas
and places, as well as other people (IE。, my wife). 的确,
the feeling of ownership extends to just about anything
for which the terms mine, 我的, or ours (for joint owner-
船) can be applied. It can be founded in the laws of
社会 (IE。, legal ownership) or manifest in the feelings
held toward an object that is owned by the individual
(IE。, psychological ownership) without any legal claim to
title (Pierce, Kostova, & Dirks, 2003). Noting this distinc-
的, Etzioni (1991) refers to the feeling of mineness that
is associated with psychological ownership, an experience
we explore in the current investigation. The question
motivating our inquiry is quite straightforward, what hap-
pens in the brain when people acquire arbitrary psycho-
logical ownership over a set of objects?
Ownership and the Self
In an influential article, 加拉格尔 (2000) has explored the
relationship between ownership and self. He states that
University of Aberdeen
this minimal sense of self is grounded in the moment-to-
moment mapping of intentions to act with the sensory
and proprioceptive feedback that accompanies the actions.
因此, we have a sense of body ownership and the ability to
author actions with that body ( Jeannerod, 2003). 虽然
these “minimal” components of self as agent and owner
of actions are generally indistinguishable in voluntary ac-
系统蒸发散, it is possible to experience action ownership without
the accompanying agentic control (Frith, 1992; 范伯格,
1978). Recent neuroimaging studies indicate a functional
role for parietal cortex in the experience of self as the
author of action (Farrer et al., 2008; Shimada, Hiraki, &
Oda, 2005). This bodily self-awareness, manifest in match-
ing intention and sensation, affords a mechanism to dis-
tinguish self from non-self-action. 重要的, agentic
movements may, on occasion, result in self-relevant out-
来了 (both good and bad). One such outcome may be
the acquisition of an object.
Although a minimal self may enable us to form a momen-
tary association with an object, it cannot easily recognize
the self-relevance of this item. For this to occur, Gallagherʼs
(2000) minimal self needs to be extended to include a
memorial component. The development of an extended
autobiographical or narrative self, with the ability to store
self-knowledge, allows us to form associations with self-
相关信息, 物体, 和人 (Damasio, 1999).
因此, object ownership represents the mental synthesis of
object and self in time. The relationships we form with a
childhood comforter, a wedding ring, or a birthday gift
from a child are all important aspects of this extended, 我-
morial self. 实际上, these self–object associations form the
© 2011 麻省理工学院
认知神经科学杂志 23:11, PP. 3657–3668
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
哦
H
w
t
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
r
t
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
r
e
e
r
C
t
C
.
米
H
A
我
e
r
d
.
你
C
哦
哦
米
C
n
/
j
A
哦
r
t
C
我
C
n
e
/
–
A
p
r
d
t
我
2
C
3
我
1
e
1
–
3
p
6
d
5
F
7
/
1
2
9
3
4
/
1
1
8
1
7
/
5
3
6
哦
5
C
7
n
_
/
A
1
_
7
0
7
0
6
0
8
4
4
2
1
p
/
d
j
哦
乙
C
y
n
G
_
你
A
e
_
s
0
t
0
哦
0
n
4
0
2
8
.
S
p
d
e
F
p
e
乙
米
y
乙
e
G
r
你
2
e
0
s
2
t
3
/
j
t
F
/
.
哦
n
1
8
中号
A
y
2
0
2
1
basis for the concrete self-construct developed in early
童年 (Dyl & Wapner, 1996; Montemayor & Eisen,
1977). Loss of material possessions through theft or damage
or the need sell sentimental objects when moving into a
nursing home can be devastating and experienced as a loss
of self (Cram & Paton, 1993; Pierce et al., 2003). Given the
significance of material objects to the development and
maintenance of our sense of self, it is important to under-
stand how these associations are formed and the resultant
impact they exert on cognition and behavior.
As a result of their association with self, owned ob-
jects are believed to enjoy a special psychological status
(Beggan, 1992). 的确, owned objects are viewed as ex-
tensions of self, as Sartre states, “I am what I have…What
is mine is myself” (Sartre, 1943/1969, PP. 591–592; see also
James, 1890). This is further illustrated by the “mere owner-
ship” effect, a tendency for objects arbitrarily assigned
to self (IE。, owned but not chosen by self ) to be imbued
with more positive characteristics (Beggan, 1992; Belk,
1988, 1991) and to be perceived as more valuable (IE。, 这
endowment effect; Kahneman, Knetsch, & Thaler, 1991;
Knetsch & Sinden, 1984) and subsequently more memo-
rable than identical items not assigned to self ( Van den
Bos, 坎宁安, 康威, & Turk, 2010; 坎宁安,
Turk, & Macrae, 2008). This skewing of value and valence
reflects the operation of positivity biases that distort the
evaluation of material possessions that comprise an im-
portant element of self (Beggan, 1992; Belk, 1988, 1991;
Kahneman et al., 1991; Knetsch & Sinden, 1984).
Notwithstanding the observation that we are what we
own, recent neuroimaging investigations have focused on
the effect of explicit, evaluative self-referential encoding
on memory. In the most widely used paradigm (Turk,
坎宁安, & Macrae, 2008; Heatherton et al., 2006;
Macrae, Moran, Heatherton, Banfield, & Kelley, 2004; Kelley
等人。, 2002; 罗杰斯, Kuiper, & Kirker, 1977; see Symons &
约翰逊, 1997, for a review), participants are required
to evaluate personality traits on the degree to which they
describe either self or a familiar other person (例如, 乔治
瓦. 衬套, Angelina Jolie). When memory for the trait words
is subsequently assessed, those encoded in relation to self
are better remembered than those processed in the con-
text of a familiar other. This so-called self-reference effect
is a reliable phenomenon that has been reported in a range
of experimental settings (see Symons & 约翰逊, 1997).
Psychological ownership, 所以, offers an alternative
route to study self through its association with objects.
Cunningham et al. (2008) tested subjects in pairs and told
them to imagine that they each owned one of two colored
shopping baskets that were placed in front of them. 帕-
ticipants were asked to place pictures of items found
in any supermarket (例如, apple, socks, pencil ) 进入
baskets on the basis of a color-sorting task, at the end of
which they each “owned” the items in one of the baskets.
The task was operationalized, such that action (IE。, mov-
ing the item) was equally shared between self-owned and
other-owned items. At the end of this sorting task, a recog-
nition memory task revealed a significant memorial ad-
vantage for items owned by self, regardless of who acted
upon the object.
What Cunningham et al.ʼs (2008) findings reveal is that
self-item associations forged through psychological owner-
ship yield a similar mnemonic advantage to that generated
through the explicit, evaluative encoding of trait adjectives
(Turk et al., 2008; Symons & 约翰逊, 1997). This then
raises an interesting question: To what extent are owner-
ship effects supported by activation of the neural systems
associated with self-referential processing?
The Neural Basis of Self-referential Encoding
A recent meta-analysis of functional neuroimaging studies
suggests that self-referential processing is supported by
activity in a network of cortical midline structures (CMS), 在
addition to task-related lateral brain areas (Northoff et al.,
2006). The CMS areas engaged in core aspects of self-
referential cognition include ventromedial pFC (vMPFC),
dorsomedial pFC (dMPFC), posterior cingulate, and parietal
cortices (Amodio & Frith, 2006; Heatherton et al., 2006;
Macrae et al., 2004; Kelley et al., 2002). Northoff et al.
(2006) argue that these commonly activated regions have
specific functional roles in the instantiation of a core men-
tal self. 具体来说, the vMPFC functions as a polymodal
convergence zone between exteroceptive sensory areas
(例如, amygdala, BG including the striatum and nucleus
accumbens) and interoceptive areas in the midbrain and
brainstem. 此外, the dorsal part of the medial pFC
(including the anterior cingulate) is densely connected to
lateral pFC (including the insula). These regions are be-
lieved to play an important role in affective processing and
the reappraisal and evaluation of stimuli with regard to self
(and “mind reading” ;Frith & Frith, 2003). 最后, the pos-
terior regions (composed of cingulate, retrosplenial, 和
parietal cortices) are densely connected to the hippocampus
and may, 所以, reflect activity associated with autobio-
graphical aspects of the self.
In the case of the trait–adjective paradigm (Kelley et al.,
2002), the coactivation of ventromedial and posterior CMS
areas reflects a need to evaluate external cues against inter-
nal representations of self in memory (例如, “am I happy?”).
But what of object ownership, how may it be supported
in the brain? In addition to the aforementioned regions, 它
may be expected that ownership would also activate areas
associated with affective processing, as acquiring an object
is a potentially rewarding experience. 在人类中, the neural
basis of reward has been studied in the context of taking
addictive substances (David et al., 2005; Stein et al., 1998;
Breiter et al., 1997), monetary gains and losses (Chiu,
Holmes, & Pizzagalli, 2008; 刘等人。, 2007; Elliott, 弗里斯顿,
& Dolan, 2000; Koepp et al., 1998; Thut et al., 1997), 风险-
taking behavior (Dreher, 2007; Bechara, Damasio, Damasio,
& 安德森, 1994; Damasio, 1994), listening to music
(Menon & Levitin, 2005), and sexual intercourse (Ortigue,
Grafton, & Bianchi-Demicheli, 2007). Brain areas associated
3658
认知神经科学杂志
体积 23, 数字 11
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
F
r
哦
米
D
哦
H
w
t
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
r
t
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
r
e
e
r
C
t
C
.
米
H
A
我
e
r
d
.
你
C
哦
哦
米
C
n
/
j
A
哦
r
t
C
我
C
n
e
/
–
A
p
r
d
t
我
2
C
3
我
1
e
1
–
3
p
6
d
5
F
7
/
1
2
9
3
4
/
1
1
8
1
7
/
5
3
6
哦
5
C
7
n
_
/
A
1
_
7
0
7
0
6
0
8
4
4
2
1
p
/
d
j
哦
乙
C
y
n
G
_
你
A
e
_
s
0
t
0
哦
0
n
4
0
2
8
.
S
p
d
e
F
p
e
乙
米
y
乙
e
G
r
你
2
e
0
s
2
t
3
/
j
/
t
F
.
哦
n
1
8
中号
A
y
2
0
2
1
with reward and hedonic experience include the striatum,
midbrain, thalamus, orbito-frontal cortices, limbic areas (在-
cluding the amygdala and insula), and medial pFC (刘等人。,
2007; 克林格尔巴赫 & Rolls, 2004; Damasio, 1996). We could,
所以, expect that in addition to CMS activation as-
sociated with self, psychological ownership may recruit a
subset these affective brain regions reflecting the hedonic
importance of self–object associations.
The Current Study
To determine the neural correlates of psychological owner-
船, we used fMRI to measure brain activity while par-
ticipants performed a simple sorting task similar to that
employed by Cunningham et al. (2008). Participants were
required to allocate shopping items to one of two baskets
(one of which was owned by self ) by means of a button
press. Following this sorting phase, participants undertook
a surprise recognition test to assess the impact of owner-
ship on memory. Because object possession reflects a
combination of self-referential and affective processing
(Pierce et al., 2003; Beggan, 1992; Belk, 1991), we expected
cortical midline areas as well as regions associated with
positive reward to underpin psychological ownership.
方法
参加者
Nineteen participants (12 女性) recruited from the
University of Aberdeen undergraduate community took
part in the study. All participants were right-handed, 本国的
English speakers with no history of neurological problems.
All gave informed consent according to the procedures
approved by the Grampian Region Ethics Committee.
Design and Stimulus Materials
The experiment included an encoding (IE。, sorting) phase
and a recognition test phase. Both were carried out while
participants were lying in the magnet bore, but only brain
activity at encoding was recorded. Before scanning com-
menced, participants were informed that the experiment
was designed to measure the neural activity associated with
sorting shopping items according to a color cue. 期间
the encoding phase, images were presented in two func-
tional runs. Each run contained 72 trials of interest and
38 rest trials. During the encoding phase images of two
colored shopping baskets were presented in the top left
(blue basket) and top right quadrants (red basket) 的
visual field. Participants were informed that one basket
belonged to them, whereas the other belonged to the ex-
perimenter in the control room. The color of the basket
associated with self or experimenter was counterbalanced
across participants. The task was to place items into the
correct basket by matching a color patch presented di-
rectly above the item with the color of the basket.
The stimulus set comprised 216 full color photographic
images of items available for purchase in a large super-
市场 (例如, 食物, 衣服, electrical items) sized to 400 ×
400 pixels at a resolution of 72 dpi. They were divided
into three equal sets (matched for item type, word length
and number of syllables, and broadly on purchase price).
One of the sets was paired with a red color patch, 一
was paired with a blue color patch, and the third was used
as foils in a surprise recognition memory test that followed
the encoding phase. In this phase, 全部 216 items were pre-
sented individually, and participants made an old/new rec-
ognition judgment.
During the encoding phase, a single item was presented
为了 2 秒. Following a 500-msec delay interval, a circular
color patch (50 pixels in diameter) was presented to de-
note the location into which the item should be placed.
Participants were then required to make a response with
either the left or right index finger to place the item into
the appropriate colored basket. There was then a 500-msec
intertrial interval in which only the two colored baskets
remained on the screen. In addition to the 144 encoding
试验, data from a further 76 randomly interleaved “jittered”
rest trials were also collected. On these rest trials, only the
basket image remained on the screen for the full 2.5-sec
TR period. During the test phase, 全部 216 stimuli were used.
Each was centrally presented and remained on the screen
while participants made an old/new recognition memory
判断.
Image Acquisition
Image acquisition was undertaken on a 1.5-T whole body
scanner (GE Healthcare) with a standard head coil. Cush-
ions were used to minimize head movement. Anatomical
images were acquired using a high-resolution 3-D spoiled
gradient recalled echo sequence (124 sagittal slices, =
3.2 毫秒, TR = 8 毫秒, flip angle = 15°, voxel size = 1 ×
1 × 1.6 毫米). Functional images were collected in runs,
each comprising 110 volumes using a gradient spin-echo,
echo-planar sequence sensitive to BOLD contrast (TR =
2500 毫秒, = 40 毫秒, flip angle = 90°, 3.75 × 3.75 在-
plane resolution). For each volume, 30 axial slices, 5-毫米
slice thickness and 0-mm skip between slices, were ac-
quired allowing complete brain coverage.
结果
Image Analysis
Preprocessing and analysis of the imaging data were per-
formed using SPM2 (Wellcome Department of Cognitive
Neurology, 伦敦, 英国). 第一的, functional data were time-
corrected for differences in acquisition time between slices
for each whole-brain volume and realigned to the first vol-
ume to minimize the effects of head movements on data
分析. Functional data were then transformed into a stan-
dard anatomical space (2-mm 各向同性体素) 基于
Turk et al.
3659
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
哦
H
w
t
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
r
t
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
r
e
e
r
C
t
C
.
米
H
A
我
e
r
d
.
你
C
哦
哦
米
C
n
/
j
A
哦
r
t
C
我
C
n
e
/
–
A
p
r
d
t
我
2
C
3
我
1
e
1
–
3
p
6
d
5
F
7
/
1
2
9
3
4
/
1
1
8
1
7
/
5
3
6
哦
5
C
7
n
_
/
A
1
_
7
0
7
0
6
0
8
4
4
2
1
p
/
d
j
哦
乙
C
y
n
G
_
你
A
e
_
s
0
t
0
哦
0
n
4
0
2
8
.
S
p
d
e
F
p
e
乙
米
y
乙
e
G
r
你
2
e
0
s
2
t
3
/
j
.
t
F
/
哦
n
1
8
中号
A
y
2
0
2
1
the ICBM 152 brain template (MNI). Normalized data were
then spatially smoothed (6 mm FWHM) using a Gaussian
kernel. Statistical analyses were performed using the gen-
eral linear model. An event-related design was modeled
using a canonical hemodynamic response function and
its temporal derivative. The model also included regressors
for additional covariates of no interest (例如, linear trends
for each functional run). This analysis was performed indi-
vidually for each participant, and resulting contrast im-
ages were subsequently entered in a second-level analysis,
treating participants as a random effect. To minimize false-
positive results, we ran a Monte Carlo simulation (看
Slotnick, Moo, Segal, & 哈特, 2003) to determine the mini-
mum cluster size necessary to enforce an a priori threshold
of p < .05 (corrected for multiple comparisons). This simu-
lation effects were considered statistically significant using
a criterion of 27 or more contiguous resampled voxels at a
voxelwise threshold of p < .0001.
For each functional run, data were preprocessed to re-
move sources of noise and artifact. Functional data were
corrected for differences in acquisition time between slices
for each whole-brain volume, realigned within and across
runs to correct for head movement, and coregistered with
each participantʼs anatomical data. Functional data were
then transformed into a standard anatomical space (3 mm
isotropic voxels) based on the ICBM 152 brain template
(MNI), which approximates Talairach and Tournouxʼs atlas
space. Normalized data were then spatially smoothed using
a Gaussian kernel (6 mm FWHM). For each participant, a
general linear model specifying task effects (modeled with
a function for the hemodynamic response) and runs (mod-
eled as constants) was used to compute parameter esti-
mates (β) and t contrast images for each comparison at
each voxel. These individual contrast images were then
submitted to a second-level, random-effects analysis to ob-
tain mean t images. A direct contrast between the two con-
ditions of interest (thresholded at p < .0001, uncorrected;
K > 27; see Slotnick et al., 2003) revealed a network of
brain areas more active for owned than not-owned objects.
These regions are reported in Talairach atlas space.
Event-related fMRI Latency
To determine the latency or time to peak for each condition
(IE。, self-owned vs. other-owned), functional activity was
modeled using one regressor for each condition. 那里-
gressors were obtained by convolving the vector of onsets
of each condition with a canonical hemodynamic response
function, as described by SPM2. This resulted in two pre-
dictors of brain activity for each experimental run. 这
model was fitted to the fMRI time course data on a voxel-
by-voxel basis. The fitted hemodynamic response for each
condition and each participant in a number of ROIs (的-
termined from the self-owned > other-owned contrast)
was extracted. The individual fitted hemodynamic re-
sponse curves were then averaged to produce a grand
average hemodynamic response curve. The latency of each
condition in each ROI was therefore the time to peak of
this grand average response curve (IE。, the time after stim-
ulus onset where the maximum fitted signal amplitude
occurred). This calculation affords a mechanism to de-
termine the time at which each brain region reached its
peak level of activation (Sun, 磨坊主, & DʼEsposito, 2005).
This analysis can therefore be used to explore the temporal
relationship in activity across regions of the ownership
网络.
Behavioral Data
Overall recognition rates were reasonable for this type of
任务 (Hits Self = .55, Hits Other = .42, False Alarms =
.21). Each participantʼs recognition score was corrected
for baseline false alarm rate by subtracting the propor-
tion of old responses to foils from the proportion of old
responses to previously presented items. Participantsʼ cor-
rected scores were submitted to a single factor (所有权:
self-owned or other-owned) paired t test. The analysis re-
vealed a significant effect of ownership [t(18) = 3.944, p <
.001, two-tailed], such that more self-owned than other-
owned items were correctly recognized (mean = 0.34
(SD = 0.14) and mean = 0.21 (SD = 0.11), respectively).
fMRI Analysis
To explore differences in brain activation to owned and
not-owned objects, two specific contrasts were conducted.
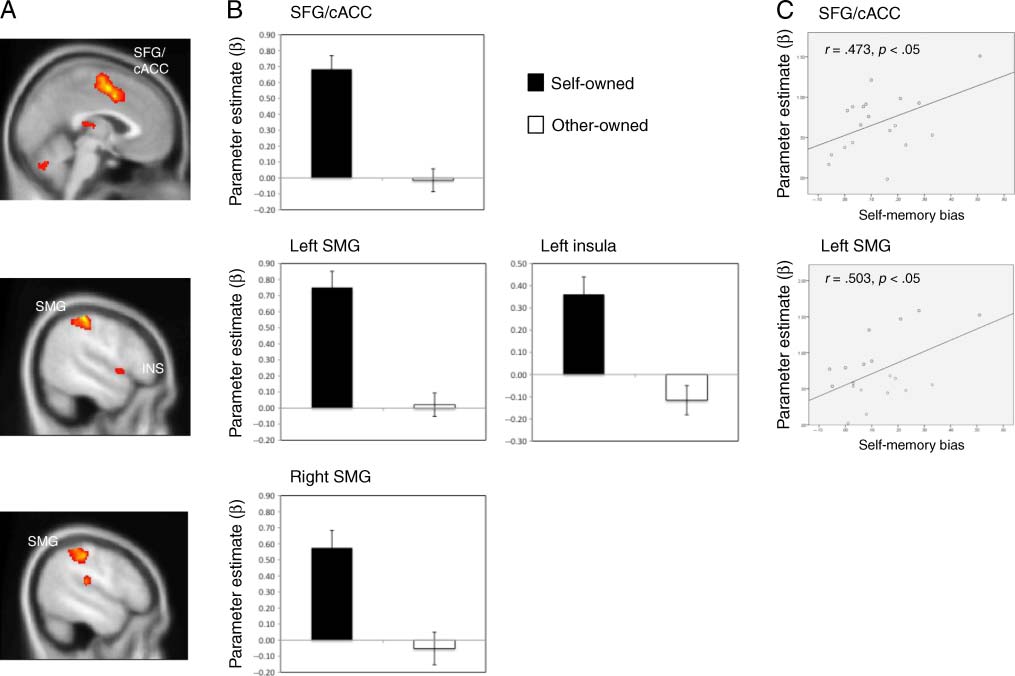
First, we explored brain regions, in which BOLD signal was
greater for owned than not-owned objects. This contrast
revealed a network of areas including a large cluster of
voxels on the medial surface of the superior frontal gyrus
(SFG; BA 6) extending dorsally to a caudal region of the
ACC (cACC; BA 24/32). In addition, activation was also ob-
served in left insula (including frontal operculum), bilateral
regions of the anterior inferior parietal lobe (BA 2, 40), and
right superior temporal cortex (BA 22) (see Figure 1A and
B). Significant BOLD increases were also observed in the
cerebellum and in subcortical structures (e.g., bilateral thal-
amus, left putamen, left globus pallidus; see Table 1, top).
The second contrast examined brain regions showing
greater BOLD signal for other-owned relative to self-owned
items. This analysis revealed a number of brain regions
more active during the encoding of other–object relations
(see Table 1, bottom). Of particular interest, regions lo-
cated along the cortical midline (see Figure 2C), previously
shown to be important in self-referential processing, were
more active during other than self-trials. Kelley et al.
(2002) reported task-related decreases in posterior cingu-
late (BA 23/31) and vMPFC (BA 10) during explicit, eval-
uative encoding. The contrast BOLD difference in these
regions was characterized by significantly greater deactiva-
tion on trials in which other-referential processing occurred
relative to self-related activity. Task-related deactivation
relative to rest in anterior and posterior cortical midline
3660
Journal of Cognitive Neuroscience
Volume 23, Number 11
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
1
-
3
p
6
d
5
f
7
/
1
2
9
3
4
/
1
1
8
1
7
/
5
3
6
o
5
c
7
n
_
/
a
1
_
7
0
7
0
6
0
8
4
4
2
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
4
0
2
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
.
/
f
o
n
1
8
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
1
-
3
p
6
d
5
f
7
/
1
2
9
3
4
/
1
1
8
1
7
/
5
3
6
o
5
c
7
n
_
/
a
1
_
7
0
7
0
6
0
8
4
4
2
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
4
0
2
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
.
t
Figure 1. (A and B) Brain regions more active for self-owned than other-owned trials and their corresponding parameter estimates (calculated
as the average for all voxels in the ROI). Error bars indicate SEM. (C) Areas that show a significant correlation between ROI parameter estimate
and self-memory bias (self-other memory).
Table 1. Group Activations Associated with Ownership
Coordinates
Brain Region
Self Owned > Other Owned
Frontal cortex
Medial surface of SFG
Extending to cACC
Left insula/frontal operculum
Parietal cortex
Right precentral gyrus
Right precentral gyrus
Right supramarginal/postcentral gyrus
Left SMG
Extending to postcentral gyrus
Left postcentral gyrus
Right postcentral gyrus
X
4
−2
−42
36
42
51
−46
−50
−65
60
y
−3
8
−3
−9
−13
−26
−35
−27
−20
−20
z
50
38
9
59
56
53
46
46
19
18
t
BA
13.39
9.39
9.26
10.83
8.42
10.24
9.84
8.80
7.95
6.26
哦
n
1
8
中号
A
y
2
0
2
1
6
24/32
4/6
4
2/40
40
2/40
2
42/2
Turk et al.
3661
桌子 1. (continued )
Brain Region
Temporal cortex
Right superior temporal gyrus
小脑
小脑
小脑
小脑
小脑
小脑
Subcortical
Left putamen
Left medial globus pallidus
Right thalamus
Other Owned > Self Owned
Frontal cortex
Left middle frontal gyrus
Left middle frontal gyrus
Right middle frontal gyrus
Anterior cingulate
Cingulate/calloso SMG
Medial/SFG
dMPFC
vMPFC
vMPFC
Parietal cortex
Right angular gyrus
Left posterior cingulate
Left precuneus extending to PCC
Right postcentral gyrus
Temporal cortex
Right middle temporal gyrus
Left superior occipital gyrus
Extending to
Left superior occipital gyrus
Left superior temporal gyrus
Coordinates
y
6
10
−69
−68
−67
−63
−52
2
0
−19
8
29
29
33
25
31
46
49
54
54
57
−70
−55
−44
−63
−11
−69
−51
−65
−59
X
48
55
0
4
−28
24
42
−28
−10
2
−44
−24
28
24
36
6
10
18
10
−6
−4
50
−12
−4
−2
42
53
−44
−42
−57
z
0
−4
−22
−10
−22
−22
−21
0
4
10
49
41
32
44
39
6
−7
10
27
−6
5
29
21
43
14
23
11
25
25
23
t
6.15
5.84
6.85
6.14
11.47
5.54
8.22
8.11
6.94
6.36
8.17
9.10
6.22
6.01
5.73
4.94
5.7
8.06
7.26
6.25
6.68
6.63
8.43
9.04
8.08
5.77
6.46
8.01
7.65
6.81
BA
22
21
6
8
9
8/9
8
24
32/10
10
9
10
10
39
31/23
31/7
31
43/41
39
19
19
39
3662
认知神经科学杂志
体积 23, 数字 11
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
F
r
哦
米
D
哦
H
w
t
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
r
t
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
r
e
e
r
C
t
C
.
米
H
A
我
e
r
d
.
你
C
哦
哦
米
C
n
/
j
A
哦
r
t
C
我
C
n
e
/
–
A
p
r
d
t
我
2
C
3
我
1
e
1
–
3
p
6
d
5
F
7
/
1
2
9
3
4
/
1
1
8
1
7
/
5
3
6
哦
5
C
7
n
_
/
A
1
_
7
0
7
0
6
0
8
4
4
2
1
p
/
d
j
哦
乙
C
y
n
G
_
你
A
e
_
s
0
t
0
哦
0
n
4
0
2
8
.
S
p
d
e
F
p
e
乙
米
y
乙
e
G
r
你
2
e
0
s
2
t
3
/
j
F
.
/
t
哦
n
1
8
中号
A
y
2
0
2
1
桌子 1. (continued )
Brain Region
Right superior temporal gyrus
Left middle temporal gyrus
Right hippocampal gyrus
Left middle temporal gyrus
Left lingual gyrus
Left middle temporal gyrus
Right superior temporal gyrus
Right superior temporal gyrus
Left superior temporal gyrus
Subcortical
Left anterior commissure/hippocampus
Coordinates
y
−59
−49
−49
−46
−45
−27
−14
−6
5
−9
z
23
1
1
4
−4
−2
−11
10
−14
−16
X
55
−53
30
−61
−22
−61
51
57
−40
−34
BA
37
21
19
21
19
21
41/42
42
38
t
7.43
6.96
6.25
7.35
6.77
5.99
6.39
5.54
8.39
6.3
Activations determined to be significant are listed along with the best estimate of their location. BA = approximate Brodmannʼs area. Coordinates are
from the Talairach and Tournoux (1988) atlas. Locations of the activations are determined based on the functional responses superimposed on averaged
anatomical MRI images and are referenced to the Talairach and Tournoux atlas. Minimum cluster size = 27 voxels ( p < .0001, uncorrected).
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
1
-
3
p
6
d
5
f
7
/
1
2
9
3
4
/
1
1
8
1
7
/
5
3
6
o
5
c
7
n
_
/
a
1
_
7
0
7
0
6
0
8
4
4
2
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
4
0
2
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
f
.
/
o
n
1
8
M
a
y
2
0
2
1
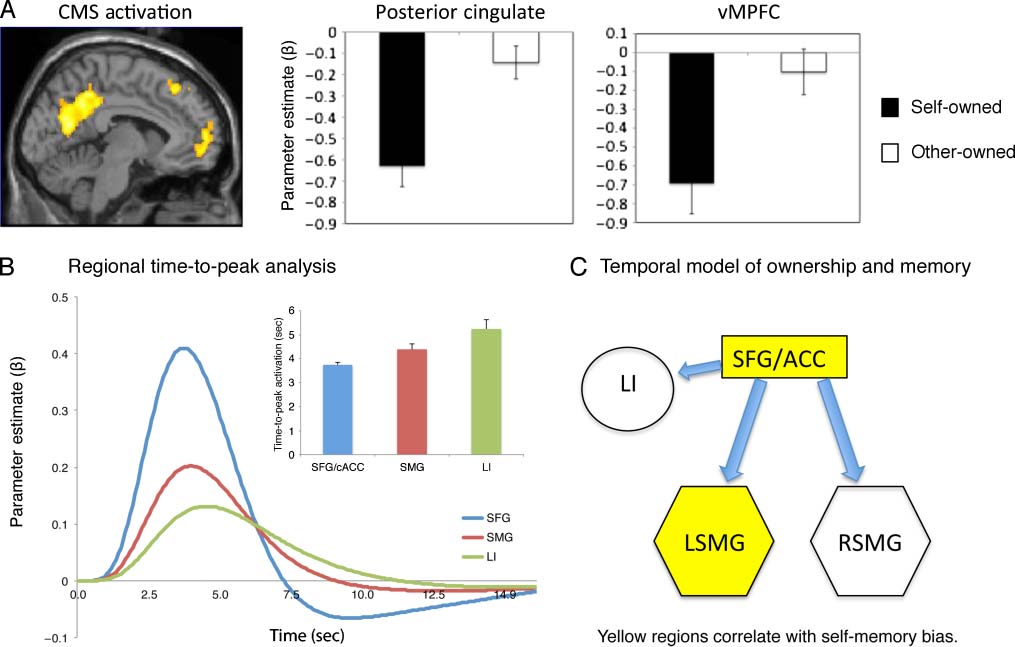
Figure 2. (A) Time-to-peak analysis of signal change across SFG/cACC (SFG), supramarginal cortex (SMG), and left insula (LI). (B) Temporal
neural model of ownership. Activation in regions highlighted in yellow also predicted self-memory bias. (C) Specific regions of cortical midline
(CMS) previously shown to be functionally important in explicit, evaluative self-referential encoding and memory (posterior cingulate and
vMPFC). These regions show decreased activation on trials in which objects were owned by self.
Turk et al.
3663
sites has been also observed in previous studies exploring
default brain state (Raichle & Snyder, 2007; Gusnard &
Raichle, 2001; Raichle et al., 2001). What makes the current
pattern of cortical midline activity particularly interesting
is that in the deactivation observed was significantly greater
for self-owned events. This finding is therefore in direct
opposition to the pattern of CMS activation observed in
previous studies investigating the neural correlates of self-
referential processing (e.g., Kelley et al., 2002).
Self-memory Bias and the Ownership Network
To investigate the relationship between memory perfor-
mance and the brain activity that accompanied self-owned
trials, we examined the correlation between the BOLD
response in the ownership network and self-memory bias
(i.e., the difference in memory for self-owned vs. other-
owned objects). This revealed a positive relationship be-
tween self-memory bias and brain activation in the SFG/
cACC [r(19) = .473, p < .05] and left supramarginal cortex
[r(19) = .503, p < .05] (see Figure 1C). However, there
was no significant correlation between BOLD response
and memory bias in right supramarginal cortex [r(19) =
.357, p = .13] or in left insula [r(19) = .312, p = .19].
A Temporal Pattern of Activation in the
Ownership Network
In addition to exploring the spatial extent of the ownership
network, we also performed an analysis of the temporal
aspects of the BOLD signal when processing self-owned
objects. This analysis examined the time-to-peak latency in
medial pFC (including SFG and cACC), averaged across
bilateral parietal cortex (supramarginal and postcentral
gyri) and left insular cortex (including frontal operculum).
In this analysis, differences in the latency of the peak
hemodynamic response function were used to build a tem-
poral model of activation during self–object association.
The fitted hemodynamic response functions are presented
in Figure 2A, along with a simplified temporal model of
maximal neural activation in each region (see Figure 2B).
For analysis of the time-to-peak data, we explored temporal
differences in peak activation across dorsomedial frontal,
parietal and fronto-temporal clusters. We submitted the
time-to-peak data to a single factor (region: SFG/cACC,
bilateral supramarginal gyri [SMG], left insula) repeated-
measures ANOVA. This revealed a significant main effect
of Region F(1.29, 39.6) = 16.362, p < .001. Post hoc pair-
wise t tests exploring the temporal relationship between
these three brain regions demonstrated that time-to-peak
was significantly faster in medial prefrontal than SMG re-
gions [t(18) = -3.191, p < .005] and left insular cortex
[t(18) = −4.303, p < .001]. The difference in time-to-peak
between parietal cortex and insula also approached sta-
tistical significance [t(18) = −1.96, p = .066 ns]. These
data suggest a temporal model in which initial activation
in dorsal midline areas promulgates secondary activity in
lateral posterior and frontal brain regions.
DISCUSSION
Whereas previous research exploring the self-reference
effect in memory and its neural basis has tended to utilize
a directed approach to the formation of self-item associa-
tions (Macrae et al., 2004; Kelley et al., 2002; Rogers et al.,
1977; see Symons & Johnson, 1997, for a full review),
the current study supports the notion that self-memory
biases can be generated under less evaluative encoding
conditions such as object ownership (Van den Bos et al.,
2010; Cunningham et al., 2008), unconstrained choice
(Cloutier & Macrae, 2008), or the incidental presentation
of the perceiverʼs own name or face with the task-relevant
information (Turk et al., 2008). The present investigation
explored the neural correlates of self–object associations
formed through temporary ownership.
A Network of Brain Regions for Ownership
Brain regions exhibiting increased BOLD response to
objects owned by self included posterior dMPFC extend-
ing ventrally to cACC (BA 6/24/32), bilateral areas in ante-
rior inferior parietal cortex, including the supramarginal
and postcentral gyri (BA 40/2), left insula (including the
frontal operculum), and right superior temporal gyrus. In
addition, BOLD signal increases were also observed
bilaterally in the thalamus and in the left medial globus
pallidus and putamen. In comparison with rest, activity in
these ownership brain areas is characterized by increased
BOLD signal, whereas for other-owned objects there ap-
pears to be no difference in evoked neural response. This
suggests that activation in this network is specific to self–
object associations. In addition, this ownership network
was characterized by a distinct pattern of temporal onsets
where early medial prefrontal activity was followed by ac-
tivation in both posterior parietal cortex and insula. This
suggests that multiple processes across a range of brain
areas support temporary object ownership.
Dorsomedial SFG and cACC
Initially, self-ownership is characterized by increased activ-
ity in cACC and medial SFG. This region has been defined
as functionally important in modulating attention to salient
stimuli (Chiu et al., 2008; Carretié, Hinojosa, Martin-
Loeches, Mercado, & Tapia, 2004), as well as in the sig-
naling of positive reward (Liu et al., 2007). Activation of
the cACC region has also been observed in studies involv-
ing simple cued responses ( Winterer, Adams, Jones, &
Knutson, 2002) attributable to motivational, volitional,
and effortful task requirements (Winterer et al., 2000) and
subsequent information processing and memory. In the
current study, signal change in this region correlated with
the ownership bias in memory. These dual processes of
3664
Journal of Cognitive Neuroscience
Volume 23, Number 11
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
1
-
3
p
6
d
5
f
7
/
1
2
9
3
4
/
1
1
8
1
7
/
5
3
6
o
5
c
7
n
_
/
a
1
_
7
0
7
0
6
0
8
4
4
2
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
4
0
2
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
.
f
/
o
n
1
8
M
a
y
2
0
2
1
attentional modulation and the experiencing of positive
reward can be observed in temporal patterns of activity
within the ownership network in which activation from
this region occurs before activity in anterior inferior pa-
rietal cortex and insular cortex that have also been impli-
cated in attentional and affective processing, respectively.
nal change in this region predicted the magnitude of the
observed memorial advantage. Because our sample group
was composed of right-handed participants, this finding
suggests a possible link between activation of motor afford-
ances associated with object use (by the dominant right
hand) and self-memory effects.
Anterior Inferior Parietal Cortex—Supramarginal
and Postcentral Gyri
Activation in parietal cortex is generally associated with
attentional processes (Milner & Goodale, 1995). Although
posterior regions of the inferior and superior parietal lobe
have been identified in spatial aspects of attentional orien-
tation (Hopfinger, Buonocore, & Mangun, 2000; Corbetta,
Miezin, Shulman, & Petersen, 1993; see also Corbetta &
Shulman, 2002, for a review), the current study identified
anterior aspects of the inferior parietal lobe as functionally
important in self–object associations. Specifically, bilateral
regions of the SMG (BA 40) and postgentral gyrus (BA 2)
showed increased BOLD signal toward self-owned com-
pared with other-owned objects. Several studies have
also recorded activity in anterior inferior parietal cortex
in perceiver-object associations. For example, Handy,
Grafton, Shroff, Ketay, and Gazzaniga (2003) suggest that
when the potential for acting upon objects is recognized
(without specific instruction to do so) this signals increased
activation in inferior parietal cortex, predominantly on
the left. When compared with viewing or naming houses,
faces, animals, or abstract shapes, viewing tools lead to
increased activity in identical regions of supramarginal
cortex (Chao & Martin, 2000). It has, therefore, been sug-
gested that, in contrast to nonmanipulable objects, the per-
ception of graspable items is accompanied by activation
of the motor affordances associated with those objects
(Martin, Wiggs, Ungerleider, & Haxby, 1996; Martin, Haxby,
Lalonde, Wiggs, & Ungerleider, 1995) or in the simulation
of actions associated with object use (Ruby & Decety,
2001) in anterior inferior parietal lobe.
In the current study, participants responded to objects
found in any major supermarket (e.g., apple, iPod, beer).
Although these items might not be considered as tools,
they are all manipulable objects and as such have actions
associated with them. It is these motor affordances that
are represented in anterior inferior parietal cortex. Of note
in the current study is that both self- and other-owned
objects have similar action affordances (e.g., apple vs. pear)
and yet, compared with rest, BOLD increases in action-
related perceptual areas was uniquely associated with
self-owned items. This may be because self-ownership
signals the motor affordance of objects, whereas objects
that belong to others should generally not be touched
or used by self without prior approval. In this case, the po-
tential for action upon graspable objects owned by others
is not activated, and thus, concomitant perceptuo-motor
action representations may be suppressed. Subsequent
analysis of activity in SMG showed that left lateralized sig-
Insula/ Frontal Operculum
Previous research has suggested that the insula is involved
in a diverse set of functions. When directly stimulated,
some regions of the insula give rise to visceral, somes-
thetic, and gustatory responses (Penfield & Faulk, 1955).
The insula appears to be important for emotional sensa-
tion, and projections to limbic regions suggest it plays
an important function in the integration of emotion and
behavior (Dupont, Bouilleret, Hasboun, Semah, & Baulac,
2003) and the signaling of reward (Liu et al., 2007; see also
Phan, Wager, Taylor, & Liberzon, 2002, for a review). Acti-
vation in a similar region of the insula was reported by Liu
et al. (2007) to reflect the processing of positive reward
resulting from gain in a gambling task. Although BOLD sig-
nal in left insula does differentiate self-owned from other-
owned items, it does not predict subsequent memory bias.
According to Ferraro, Escalas, and Bettman (2011), the
monetary value of possessions may influence the per-
ceived importance of those items but does not affect the
strength of self-possession associations likely to support
memory performance.
CMS and Ownership
We hypothesized that a network of cortical midline brain
areas previously associated with self-referential encod-
ing (see Northoff et al., 2006) as well as regions that have
been shown to support hedonic aspects of item owner-
ship might play an important role in forming associations
between self and owned objects. Although dorsomedial
aspects of the cortical midline did show increased BOLD
signal for self–object associations, other previously identi-
fied brain areas associated with self-referential processing
did not. Specifically, regions in vMPFC (Heatherton et al.,
2006; Macrae et al., 2004; Johnson et al., 2002; Kelley
et al., 2002; see also Northoff et al., 2006, for a review)
and posterior cingulate that have been shown to predict
self-referential encoding and memory (Macrae et al.,
2004) showed greater neural responses to other-owned
objects, characterized by smaller decreases from baseline.
Specifically, vMPFC and posterior cingulate regions re-
ported by Kelley et al. (2002) during self-referential encod-
ing appear to be associated with other–object encoding in
the present study. So why in this case do these putative self
areas respond more to objects associated with others?
Sustained activity in cortical midline regions in posterior
and anterior frontal cortex has been observed during per-
iods of sustained rest (Shulman et al., 1997). Gusnard and
Raichle (2001) suggest that this default state represents
Turk et al.
3665
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
1
-
3
p
6
d
5
f
7
/
1
2
9
3
4
/
1
1
8
1
7
/
5
3
6
o
5
c
7
n
_
/
a
1
_
7
0
7
0
6
0
8
4
4
2
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
4
0
2
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
t
.
f
o
n
1
8
M
a
y
2
0
2
1
“a stable, unified perspective of the organism relative to its
environment (a self )” (p. 692). Activity is tonically high
during rest, as this area may be ready to interpret, respond
to, and perhaps predict future environmental events
(Raichle & Snyder, 2007). Only when individuals are re-
quired to engage in specified cognitive tasks does meta-
bolic activity decrease.
These regions (in addition to lateral parietal areas) are
also characterized by their associative functions. In line
with this preparatory or predictive function, Bar, Aminoff,
Mason, and Fenske (2007) noted that tasks that manipu-
late the degree of association between stimuli also activate
the default network. That is, highly associated items
showed higher BOLD signal relatively to weakly associated
items. They argue that this is because of unconstrained
or stimulus-independent thoughts (SITs) that propagate
such associations. These SITs are also referred to as mind
wandering (Mason et al., 2007). The greater the associa-
tion, the more mind wandering, the higher the metabolic
rate recorded in posterior and anterior cortical midline.
Mason et al. (2007) explored the impact of SITs on default
activity by giving participants novel and practiced tasks to
undertake. In addition, they probed participants during
task performance to see if they were currently on task or
mind wandering (i.e., having SITs). Because a practiced
task requires less effort, it affords a greater opportunity
for unconstrained thought and incidents of SITs. This level
of mind wandering positively correlated with the magni-
tude of the BOLD response in areas of the default network.
Conversely, stimulus-driven cognitive processes reduce
BOLD signal in these networks (Burgess, Dumontheil, &
Gilbert, 2007). Thus, it may be the nature of the thought
process (i.e., unconstrained and spontaneous vs. con-
strained and task related) that governs activation in the
default network rather than the referent to whom they
are directed.
In the current study, there was a greater decrease in
BOLD signal in posterior and anterior midline areas for
self-owned relative to other-owned trials. This can be in-
terpreted in the context of increased SITs during other–
object associations than to self–object associations. As we
have previously indicated, self-owned events appear to
increase arousal, affective state and attentional processes.
As a result, one might infer that such states are accompa-
nied by increased stimulus-driven processes (e.g., “I own
the iPod. I could listen to music on the way to work.”).
Other–object associations do not result in changes in at-
tentional processes nor do they offer a change in reward
state. As such, it is possible that these events therefore
lead to increased SITs and, therefore, sustained BOLD
signal in default areas.
Conclusion
The current investigation explored the neural basis of
nonevaluative self-referential encoding through the use of
a novel temporary ownership paradigm. This revealed a
network of brain regions that appear to respond specifi-
cally to owned objects in a distinct temporal sequence.
Given the functional specificity of these brain areas and
the temporal order in which they appear to reach maximal
activation, it is tempting to speculate upon a functional
temporal model of object ownership, with the initial de-
tection of object salience in caudal medial pFC followed
by activation of motor affordances in parietal cortex and
processing of reward in insula. Future research might prof-
itably seek to expand upon this speculative model by de-
termining the extent to which factors pertaining to the
objects (e.g., value/valence or usability) or to self (e.g.,
whether or not self was the agent or mere recipient during
the acquisition of the item or the degree to which self has
prior association with the items) might modulate the ac-
tivity in this ownership network. That these effects were
obtained from temporary ownership of ordinary, low-value,
common objects speaks to the potential usefulness of such
a methodology in elucidating the mechanisms that under-
pin the nature of self reflected in material possessions.
Acknowledgments
D. J. T. was supported by grants from the BBSRC (RGA1149) and
the European Research Council (202893). C. N. M. was supported
by a Royal Society Wolfson Fellowship.
Reprint requests should be sent to David J. Turk, School of Psy-
chology, University of Aberdeen, Kingʼs College, Aberdeen, UK,
AB24 3FX, or via e-mail: david.j.turk@abdn.ac.uk.
REFERENCES
Amodio, D. M., & Frith, C. D. (2006). Meeting of minds: The
medial frontal cortex and social cognition. Nature Reviews
Neuroscience, 7, 268–277.
Bar, M., Aminoff, E., Mason, M., & Fenske, M. (2007). The
units of thought. Hippocampus, 17, 420–428.
Bechara, A., Damasio, A. R., Damasio, H., & Anderson, S. W.
(1994). Insensitivity to future consequences following
damage to human prefrontal cortex. Cognition, 50, 7–15.
Beggan, J. K. (1992). On the social nature of nonsocial
perception—The mere ownership effect. Journal of
Personality and Social Psychology, 62, 229–237.
Belk, R. W. (1988). Possessions and the extended self.
Journal of Consumer Research, 15, 139–168.
Belk, R. W. (1991). The ineluctable mysteries of possessions.
Journal of Social Behavior and Personality, 6, 17–55.
Breiter, H. C., Gollub, R. L., Weisskoff, R. M., Kennedy,
D. N., Makris, N., Berke, J. D., et al. (1997). Acute effects
of cocaine on human brain activity and emotion. Neuron,
19, 591–611.
Burgess, P. W., Dumontheil, I., & Gilbert, S. J. (2007). The
gateway hypothesis of rostral prefrontal cortex (area 10)
function. Trends in Cognitive Sciences, 11, 290–298.
Carretié, L., Hinojosa, J. A., Martin-Loeches, M., Mercado, F.,
& Tapia, M. (2004). Automatic attention to emotional
stimuli: Neural correlates. Human Brain Mapping, 22,
290–299.
Chao, L. L., & Martin, A. (2000). Representation of manipulable
man-made objects in the dorsal stream. Neuroimage, 12,
478–484.
3666
Journal of Cognitive Neuroscience
Volume 23, Number 11
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
1
-
3
p
6
d
5
f
7
/
1
2
9
3
4
/
1
1
8
1
7
/
5
3
6
o
5
c
7
n
_
/
a
1
_
7
0
7
0
6
0
8
4
4
2
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
4
0
2
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
.
/
f
o
n
1
8
M
a
y
2
0
2
1
Chiu, P. H., Holmes, A. J., & Pizzagalli, D. A. (2008). Dissociable
recruitment of rostral anterior cingulate and inferior frontal
cortex in emotional response inhibition. Neuroimage, 42,
988–997.
Gusnard, D. A., & Raichle, M. E. (2001). Searching for a
baseline: Functional imaging and the resting human
brain. Nature Reviews Neuroscience, 2, 685–694.
Handy, T. C., Grafton, S. T., Shroff, N. M., Ketay, S., &
Cloutier, J., & Macrae, C. N. (2008). Self-involvement and
the cognitive status of things past. Consciousness &
Cognition, 17, 125–135.
Corbetta, M., Miezin, F. M., Shulman, G. L., & Petersen,
S. E. (1993). A PET study of visual attention. Journal of
Neuroscience, 13, 1202–1226.
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed
and stimulus-driven attention in the brain. Nature Reviews
Neuroscience, 3, 201–215.
Cram, F., & Paton, H. (1993). Personal possessions and
self-identity: The experiences of elderly women in three
residential settings. Australian Journal on Aging, 12,
19–24.
Cunningham, S. J., Turk, D. J., & Macrae, C. N. (2008). Yours
or mine? Ownership and memory. Consciousness &
Cognition, 17, 312–318.
Gazzaniga, M. S. (2003). Graspable objects grab attention
when the potential for action is recognized. Nature
Neuroscience, 6, 421–427.
Heatherton, T. F., Wyland, C. L., Macrae, C. N., Demos, K. E.,
Denny, B. T., & Kelley, W. M. (2006). Medial prefrontal
activity differentiates self from close others. Social
Cognitive & Affective Neuroscience, 1, 18–25.
Hopfinger, J. B., Buonocore, M. H., & Mangun, G. R. (2000).
The neural mechanisms of top–down attentional control.
Nature Neuroscience, 3, 284–291.
James, W. (1890). Principles of psychology. New York:
Holt, Rinehart and Winston.
James, W. (1929). The varieties of religious experience:
A study in human nature. Being the Gifford Lectures
on Natural Religion Delivered at Edinburgh in 1901-2.
Toronto: Random House.
Damasio, A. (1999). The feeling of what happens: Body and
Jeannerod, M. (2003). Agency and self awareness. In J. Roessler
emotion in the making of consciousness. New York:
Harcourt Brace.
Damasio, A. R. (1994). Descartesʼ error: Emotion, reason,
and the human brain. New York: Grosset/Putnam.
Damasio, A. R. (1996). The somatic marker hypothesis and the
possible functions of the prefrontal cortex. Philosophical
Transactions of the Royal Society of London, Series B,
Biological Sciences, 351, 1413–1420.
David, S. P., Munafo, M. R., Johansen-Berg, H., Smith, S. M.,
Rogers, R. D., Matthews, P. M., et al. (2005). Ventral striatum/
nucleus accumbens activation to smoking-related pictorial
cues in smokers and nonsmokers: A functional magnetic
resonance imaging study. Biological Psychiatry, 58, 488–494.
Dreher, J. C. (2007). Sensitivity of the brain to loss aversion
during risky gambles. Trends in Cognitive Sciences, 11,
270–272.
Dupont, S., Bouilleret, V., Hasboun, D., Semah, F., & Baulac, M.
(2003). Functional anatomy of the insula: New insights
from imaging. Surgical & Radiologic Anatomy, 25, 113–119.
Dyl, J., & Wapner, S. (1996). Age and gender differences in
the nature, meaning, and function of cherished possessions
for children and adolescents. Journal of Experimental
Child Psychology, 62, 340–377.
Elliott, R., Friston, K. J., & Dolan, R. J. (2000). Dissociable
neural responses in human reward systems. Journal of
Neuroscience, 20, 6159–6165.
Etzioni, A. (1991). The socioeconomics of property. Journal
of Social Behavior & Personality, 6, 465–468.
Farrer, C., Frey, S. H., Van Horn, J. D., Tunik, E., Turk, D.,
Inati, S., et al. (2008). The angular gyrus computes action
awareness representations. Cerebral Cortex, 18, 254–261.
Feinberg, I. (1978). Efference copy and corollary discharge:
Implications for thinking and its disorders. Schizophrenia
Bulletin, 4, 636–640.
Ferraro, R., Escalas, J. E., & Bettman, J. R. (2011). Our
possessions, our selves: Domains of self-worth and the
self-possession link. Journal of Consumer Psychology, 21,
169–177.
Frith, C. (1992). The cognitive neuropsychology of
schizophrenia. London: Erlbaum.
Frith, U., & Frith, C. D. (2003). Development and
neurophysiology of mentalizing. Philosophical Transactions
of the Royal Society of London, Series B, Biological
Sciences, 358, 459–473.
Gallagher, S. (2000). Philosophical conceptions of the self:
Implications for cognitive science. Trends in Cognitive
Sciences, 4, 14–21.
& N. Eilan (Eds.), Issues in philosophy and psychology
(pp. 128–149). Oxford: Oxford University Press.
Johnson, S. C., Baxter, L. C., Wilder, L. S., Pipe, J. G., Heiserman,
J. E., & Prigatano, G. P. (2002). Neural correlates of
self-reflection. Brain, 125, 1808–1814.
Kahneman, D., Knetsch, J. L., & Thaler, R. (1991). The
endowment effect, loss aversion and status quo bias.
Journal of Economic Perspectives, 5, 193–206.
Kelley, W. M., Macrae, C. N., Wyland, C. L., Caglar, S., Inati, S.,
& Heatherton, T. F. (2002). Finding the self? An event-related
fMRI study. Journal of Cognitive Neuroscience, 14, 785–794.
Knetsch, J. L., & Sinden, J. A. (1984). Willingness to pay and
compensation demanded: Experimental evidence of an
unexpected disparity in measures of value. Quarterly
Journal of Economics, 99, 507–521.
Koepp, M. J., Gunn, R. N., Lawrence, A. D., Cunningham,
V. J., Dagher, A., Jones, T., et al. (1998). Evidence for
striatal dopamine release during a video game. Nature,
393, 266–268.
Kringelbach, M. L., & Rolls, E. T. (2004). The functional
neuroanatomy of the human orbito-frontal cortex:
Evidence from neuroimaging and neuropsychology.
Progress in Neurobiology, 72, 341–372.
Liu, X., Powell, D. K., Wang, H. B., Gold, B. T., Corbly, C. R.,
& Joseph, J. E. (2007). Functional dissociation in frontal
and striatal areas for processing of positive and negative
reward information. Journal of Neuroscience, 27,
4587–4597.
Macrae, C. N., Moran, J. M., Heatherton, T. F., Banfield, J. F.,
& Kelley, W. M. (2004). Medial prefrontal activity predicts
memory for self. Cerebral Cortex, 14, 647–654.
Martin, A., Haxby, J. V., Lalonde, F. M., Wiggs, C. L., &
Ungerleider, L. G. (1995). Discrete cortical regions
associated with knowledge of color and knowledge of
action. Science, 270, 102–105.
Martin, A., Wiggs, C. L., Ungerleider, L. G., & Haxby, J. V. (1996).
Neural correlates of category-specific knowledge. Nature,
379, 649–652.
Mason, M. F., Norton, M. I., Van Horn, J. D., Wegner, D. M.,
Grafton, S. T., & Macrae, C. N. (2007). Wandering minds:
The default network and stimulus-independent thought.
Science, 315, 393–395.
Menon, V., & Levitin, D. J. (2005). The rewards of music
listening: Response and physiological connectivity of
the mesolimbic system. Neuroimage, 28, 175–184.
Milner, A. D., & Goodale, M. A. (1995). The visual brain
in action. Oxford: Oxford University Press.
Turk et al.
3667
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
1
-
3
p
6
d
5
f
7
/
1
2
9
3
4
/
1
1
8
1
7
/
5
3
6
o
5
c
7
n
_
/
a
1
_
7
0
7
0
6
0
8
4
4
2
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
4
0
2
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
/
.
o
n
1
8
M
a
y
2
0
2
1
Montemayor, R., & Eisen, M. (1977). The development of
Shulman, G. L., Fiez, J. A., Corbetta, M., Buckner, R. L.,
self-conceptions from childhood to adolescence.
Developmental Psychology, 13, 314–319.
Northoff, G., Heinzel, A., Greck, M., Bennpohl, F., Dobrowolny,
H., & Panksepp, J. (2006). Self-referential processing in
our brain—A meta-analysis of imaging studies on the self.
Neuroimage, 31, 440–457.
Ortigue, S., Grafton, S. T., & Bianchi-Demicheli, F. (2007).
Correlation between insula activation and self-reported
quality of orgasm in women. Neuroimage, 37, 551–560.
Penfield, W., & Faulk, M. E. (1955). The insula: Further
observations on its function. Brain, 78, 445–470.
Phan, K. L., Wager, T., Taylor, S. F., & Liberzon, I. (2002).
Functional neuroanatomy of emotion: A meta-analysis of
emotion activation studies in PET and fMRI. Neuroimage,
16, 331–348.
Pierce, J. L., Kostova, T., & Dirks, K. T. (2003). The state of
psychological ownership: Integrating and extending a century
of research. Review of General Psychology, 7, 84–107.
Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J.,
Gusnard, D. A., & Shulman, G. L. (2001). A default mode
of brain function. Proceedings of the National Academy
of Sciences, U.S.A., 98, 676–682.
Raichle, M. E., & Snyder, A. Z. (2007). A default mode of
brain function: A brief history of an evolving idea.
Neuroimage, 37, 1083–1090.
Rogers, T. B., Kuiper, N. A., & Kirker, W. S. (1977).
Self-reference and the encoding of personal information.
Journal of Personality & Social Psychology, 35, 677–688.
Ruby, P., & Decety, J. (2001). Effect of the subjective
perspective taking during simulation of action: A PET
investigation of agency. Nature Neuroscience, 4, 546–550.
Sartre, J. P. (1969). Being and nothingness : An essay on
phenomenological ontology (H. E. Barnes, Trans.).
London: Methuen. (Original work published in 1943.)
Shimada, S., Hirakia, K., & Oda, I. (2005). The parietal role
in the sense of self-ownership with temporal discrepancy
between visual and proprioceptive feedbacks. Neuroimage,
24, 1225–1232.
Miezin, F. M., Raichle, M. E., et al. (1997). Common blood
flow changes across visual tasks: 2. Decreases in cerebral
cortex. Journal of Cognitive Neuroscience, 9, 648–663.
Slotnick, S. D., Moo, L. R., Segal, J. B., & Hart, J. (2003). Distinct
prefrontal cortex activity associated with item memory
and source memory for visual shapes. Cognitive Brain
Research, 17, 75–82.
Stein, E. A., Pankiewicz, J., Harsch, H. H., Cho, J. K., Fuller,
S. A., Hoffmann, R. G., et al. (1998). Nicotine-induced
limbic cortical activation in the human brain: A functional
MRI study. American Journal of Psychiatry, 155, 1009–1015.
Sun, F. T., Miller, L. M., & DʼEsposito, M. (2005). Measuring
temporal dynamics of functional networks using phase
spectrum of fMRI data. Neuroimage, 28, 227–237.
Symons, C. S., & Johnson, B. T. (1997). The self-reference
effect in memory: A meta-analysis. Psychological Bulletin,
121, 371–394.
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic
atlas of the human brain. New York: Thieme.
Thut, G., Schultz, W., Roelcke, U., Nienhusmeier, M., Missimer, J.,
Maguire, R. P., et al. (1997). Activation of the human brain
by monetary reward. NeuroReport, 8, 1225–1228.
Turk, D. J., Cunningham, S. J., & Macrae, C. N. (2008).
Self-memory biases in explicit and incidental encoding of
trait adjectives. Consciousness & Cognition, 17, 1040–1045.
Van den Bos, M., Cunningham, S. J., Conway, M. A., & Turk, D. J.
(2010). Mine to remember: The impact of ownership on
recollective experience. Quarterly Journal of Experimental
Psychology, 63, 1065–1071.
Winterer, G., Adams, C. M., Jones, D. W., & Knutson, B.
(2002). Volition to action-an event-related fMRI study.
Neuroimage, 17, 851–858.
Winterer, G., Ziller, M., Dorn, H., Frick, K., Mulert, C.,
Wuebben, Y., et al. (2000). Frontal dysfunction in
schizophrenia: A new electrophysiological classifier
for research and clinical applications. European
Archives of Psychiatry & Clinical Neuroscience, 250,
207–214.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
1
-
3
p
6
d
5
f
7
/
1
2
9
3
4
/
1
1
8
1
7
/
5
3
6
o
5
c
7
n
_
/
a
1
_
7
0
7
0
6
0
8
4
4
2
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
4
0
2
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
f
/
.
o
n
1
8
M
a
y
2
0
2
1
3668
Journal of Cognitive Neuroscience
Volume 23, Number 11