方法
Empirical evaluation of human fetal fMRI
preprocessing steps
Lanxin Ji1
, Cassandra L. Hendrix1, and Moriah E. Thomason1,2,3
1Department of Child and Adolescent Psychiatry, New York University School of Medicine, 纽约, 纽约, 美国
2Department of Population Health, New York University School of Medicine, 纽约, 纽约, 美国
3Neuroscience Institute, New York University School of Medicine, 纽约, 纽约, 美国
关键词: Fetal fMRI, 预处理, Normalization, Denoising, 平滑, Functional
连接性
开放访问
杂志
抽象的
Increased study and methodological innovation have led to growth in the field of fetal brain
功能磁共振成像. An important gap yet to be addressed is optimization of fetal fMRI preprocessing. Rapid
developmental changes, imaged within the maternal compartment using an abdominal coil,
introduce novel constraints that challenge established methods used in adult fMRI. This study
evaluates the impact of (1) normalization to a group mean-age template versus normalization
to an age-matched template; (2) independent components analysis (ICA) denoising at two
criterion thresholds; 和 (3) smoothing using three kernel sizes. Data were collected from 121
fetuses (25–39 weeks, 43.8% 女性). Results indicate that the mean age template is superior
in older fetuses, but less optimal in younger fetuses. ICA denoising at a more stringent
threshold is superior to less stringent denoising. A larger smoothing kernel can enhance cross-
hemisphere functional connectivity. 全面的, this study provides improved understanding of
the impact of specific steps on fetal image quality. Findings can be used to inform a common
set of best practices for fetal fMRI preprocessing.
介绍
Understanding of human brain development has grown rapidly with the introduction of fetal
resting-state functional connectivity (RSFC) (van den Heuvel & Thomason, 2016). 在 2011,
Veronica Schöpf and colleagues published the first fetal RSFC study (Schöpf et al., 2011),
demonstrating that it was possible to noninvasively image whole-brain functional systems
prior to birth by using MRI. Before this time, very few studies had measured fetal brain activity
(安德森 & Thomason, 2013). 的确, most of what was known about prenatal brain devel-
opment was the product of histological or structural analytic approaches in postmortem or
clinical samples (Chi et al., 1977; Dobbing & Sands, 1973), or was inferred from RSFC studies
conducted in preterm neonates (Doria et al., 2010; Fransson et al., 2007).
Fetal RSFC has enabled us to begin describing properties of typical development as well as
the role of the environment in shaping neural network development. Studies of typical devel-
opment have revealed that network connectivity patterns in utero precede and may guide
functional selectivity of certain brain regions, such as the fusiform face area (van den Heuvel
等人。, 2018), and that macroscale characteristics of fetal functional networks share significant
overlap with adult networks (Turk et al., 2019). In addition to shedding light on the origins of
引文: 吉, L。, Hendrix, C. L。, &
Thomason, 中号. 乙. (2022). Empirical
evaluation of human fetal fMRI
preprocessing steps. 网络
神经科学, 6(3), 702–721. https://土井
.org/10.1162/netn_a_00254
DOI:
https://doi.org/10.1162/netn_a_00254
支持信息:
https://doi.org/10.1162/netn_a_00254
已收到: 30 十一月 2021
公认: 9 可能 2022
利益争夺: 作者有
声明不存在竞争利益
存在.
通讯作者:
Moriah E. Thomason
moriah.thomason@nyulangone.org
处理编辑器:
Michael Cole
版权: © 2022
麻省理工学院
在知识共享下发布
归因 4.0 国际的
(抄送 4.0) 执照
麻省理工学院出版社
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
A
_
0
0
2
5
4
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Evaluation of fetal fMRI preprocessing steps
typical neural development, fetal RSFC studies also inform our understanding of health risk.
例如, exposures like prenatal stress (Thomason et al., 2021A), cannabis (Thomason
等人。, 2021乙), and lead (Thomason et al., 2019) have been linked to altered fetal neuro-
发展, which has important implications for policy and intervention. 合在一起,
this work highlights that fetal MRI is a crucial tool for understanding typical and atypical
human neurodevelopment and for uncovering the earliest origins of disease risk.
There is need to optimize fetal RSFC analytic pipelines so that this important work can be
conducted in a rigorous and reproducible manner. A number of methodological studies have
highlighted vulnerabilities in the processing and analysis of RSFC data in adults. 例如,
data-driven approaches have revealed that traditional denoising techniques using linear
modeling may incorrectly classify intrinsic neural signal as noise (Bright et al., 2017). 更远-
更多的, interlab variation in fMRI processing choices can lead to disparate results, even when
labs are analyzing the same data (Botvinik-Nezer et al., 2020). Because of the potent impact of
analytic choices on fMRI outcomes, there have been several efforts to create and distribute
集中, robust preprocessing pipelines for adult fMRI data such as fMRI PREP (Esteban
等人。, 2019) and the Human Connectome Pipeline (Glasser et al., 2013). 然而, these pre-
processing pipelines were not developed to manage the unique challenges inherent to imaging
the brain in utero, including high motion, encasement within the maternal compartment, 两个都
fetal and maternal sources of noise, and unique geometry of the large field of view and abdom-
inal coil array. There is need for development of fMRI processing pipelines suited to the devel-
opmental and methodological considerations specific to the fetus (Rajagopalan et al., 2021).
We elected to focus on three preprocessing steps that require particular attention in the
developing brain: normalization to standard space, 去噪, and smoothing. 中的一个
largest challenges in fetal fMRI is excessive motion as introduced by both the fetus and by
the mother (例如, breathing). Discarding periods of high motion or excluding subjects whose
motion exceeds a stringent threshold is often the first attempt to tackle the problem, 但它
invariably leads to significant data loss. The balance between maximizing amount of data
and maximizing data quality is challenging and highly varied across datasets with different
motion profiles. As an additional step, the regression-based motion artifact removal strategy
is widely used to control the secondary intravolume effects induced by motion, such as arti-
facts related to partial voluming and magnetic field inhomogeneities (Friston et al., 1996;
Pruim et al., 2015). Typical regression models include 6 到 24 motion covariates derived from
the volume realignment (Friston et al., 1996; Yan et al., 2013), yet these covariates are highly
reliant on the algorithm used for the realignment and, furthermore, the underlying intravolume
effects cannot be captured by the realignment parameters. Beyond motion parameter-based
型号, spatial independent component analysis (ICA) provides a powerful tool to separate
neural-related signal from different sources of noise, including the motion-related artifacts.
Applied to fMRI data, ICA decomposes data into a set of spatial independent components
and associated time courses (贝克曼 & 史密斯, 2004). Components presenting noise fea-
tures can subsequently be regressed out of the data. ICA-based denoising is well established
as a method for removing motion artifacts in adult imaging, but has yet been evaluated in fetal
成像. 因此, this study examines ICA-based data denoising in a large collection of fetal fMRI
scans. Another challenge of the fetal brain is its unparalleled, rapid development across
gestation, which complicates the normalization process. 例如, it is unclear whether
normalizing to a fetal template from a particular stage in gestation (例如, 32 weeks) is adequate,
or if instead it is necessary to normalize to a template that is closely age matched to the fetus
(例如, within a week). 最后, the smaller size of fetal brains compared to adults may require
different recommendations regarding smoothing kernel size, which may influence the
Denoising:
A critical step of preprocessing to
remove noise and nonneuronal
contributions, such as motion-related
and physiological noise.
Normalization:
A step transforming brains to a
template, to ensure that each voxel
for each subject corresponds to same
brain parts.
Realignment:
A step of preprocessing to correct
head movements, by co-registering
all volumes in a time series to a
reference volume.
Independent component
分析 (ICA):
A data-driven approach to
decompose fMRI data into a set of
statistically independent spatial maps
together with associated time
courses.
网络神经科学
703
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
A
_
0
0
2
5
4
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Evaluation of fetal fMRI preprocessing steps
likelihood of identifying significant associations (Botvinik-Nezer et al., 2020). The present
study addresses the effect of these key processing decisions during the preparation of fetal fMRI
data for second-level analyses in a large fetal dataset.
材料和方法
参加者
Healthy mothers were recruited during routine obstetrical appointments at Hutzel Women’s
Hospital in Detroit, Michigan. Inclusionary criteria included maternal age ≥18 years old,
native English speaking, singleton pregnancy, and normal fetal brain anatomy as assessed
by ultrasound and MRI examination. MRI visits occurred when fetuses were between 22
和 39 weeks gestational age (遗传算法). This study included data from second- and third-trimester
fetuses from a larger ongoing project on fetal brain development who had manually seg-
mented and quality assured raw resting-state fMRI data available at the time of this analysis
(N= 165). Development of automated processes for fetal brain segmentation is an active area
of study (Rutherford et al., 2021), 但, 现在, manual tracing of the brain is the most pre-
cise approach. Additional exclusions were applied for fetuses subsequently born very preterm
or with low birth weight (<33 weeks GA, <1,800 g; n = 14), those scanned before 25 weeks
GA (n = 9), and with fewer than 100 low-motion volumes or high segment-weighted
average motion (1.5 mm max excursion, 0.5 mean; rotational>2°, rotation mean
>1°, n = 21), resulting in a final sample of 121 fetuses (68 male; 53 女性). Included fetuses
had a mean GA of 32.89 weeks at scanning (range = 25.86–39.57; 标准差= 3.75) and were born,
一般, 在 39.08 (标准差= 1.49) weeks gestation. More detailed characteristics of the sample
are provided in Table 1. Motion parameters were not correlated with demographic variables
桌子 1.
Sample demographic characteristics (N= 121)
Maternal age, 年
Race/ethnicity, 氮 (%)
Caucasian
African American
Latina
Asian American
Biracial
Not disclosed
Fetal sex, 氮 (%)
女性
男性
Gestational age at scan, weeks
Gestational age at birth, weeks
Birth weight, 克
网络神经科学
Mean ± SD
25.34 ± 4.56
10 (8.26%)
99 (81.82%)
1 (0.83%)
1 (0.83%)
5 (4.13%)
5 (4.13%)
53 (43.80%)
68 (56.20%)
32.89 ± 3.75
39.08 ± 1.49
3,237.22 ± 510.73
704
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
A
_
0
0
2
5
4
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Evaluation of fetal fMRI preprocessing steps
including scan age and sex in the final sample (see Supporting Information, Figure S1). 全部
study procedures were approved by the Wayne State University Human Investigation
委员会.
Data Acquisition
Fetal MRI data were acquired on a Siemens Verio 70-cm open-bore 3T MR system using a
550 g abdominal 4-Channel Siemens Flex Coil (Siemens, 慕尼黑, 德国). Twelve minutes
of fetal resting-state fMRI data were acquired using the following gradient echo planar imaging
顺序: TR/TE 2,000/30 多发性硬化症, flip angle 80°, 360 frames, axial 4-mm-slice thickness, voxel
尺寸 3.4 × 3.4 × 4 mm3. The sequence was repeated when time permitted.
Preprocessing Pipelines
A full preprocessing workflow is shown in Figure 1. Time frames in the raw fMRI data corre-
sponding to periods of significant head motion were identified using FSL image viewer (FSL,
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
A
_
0
0
2
5
4
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
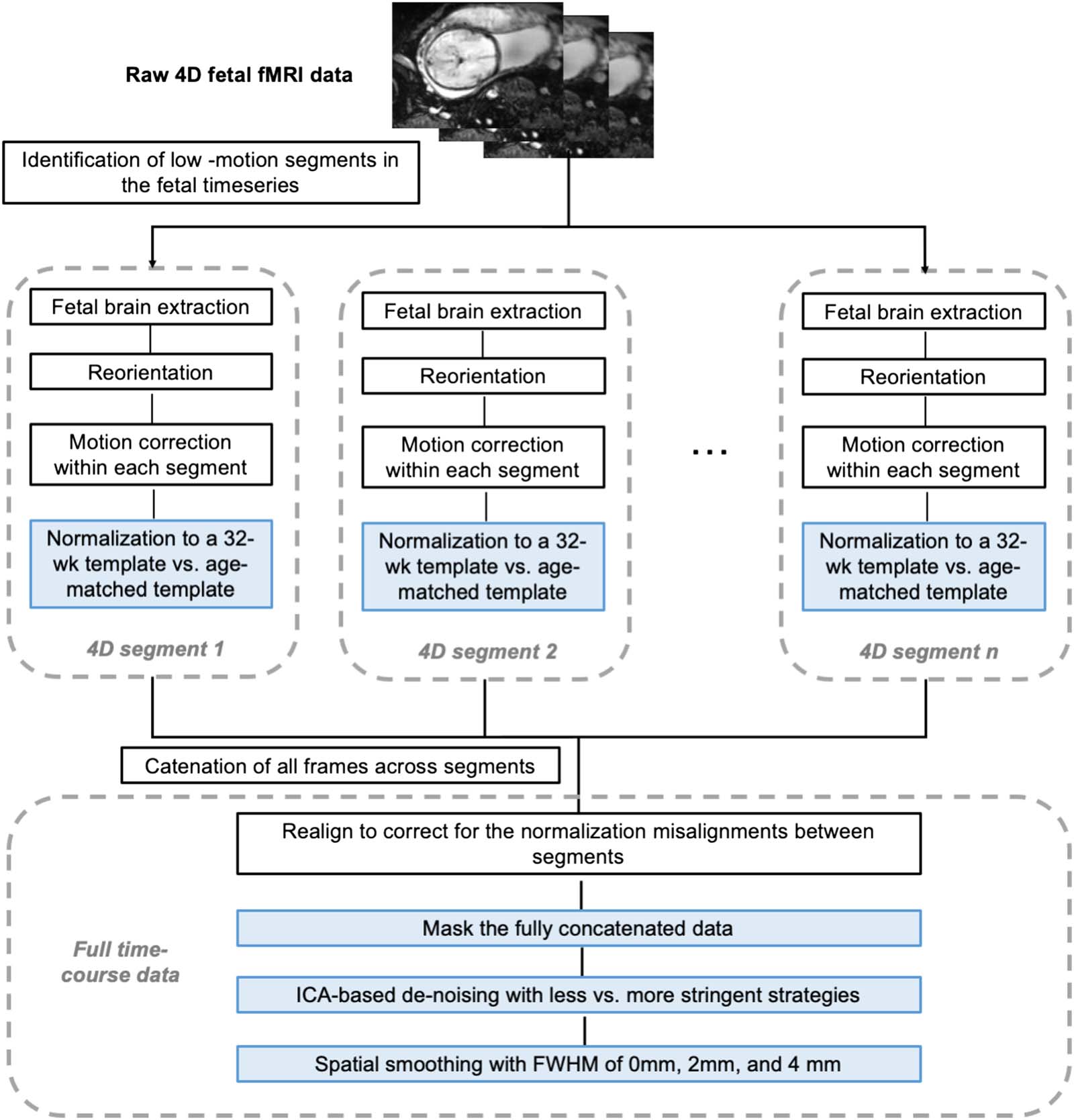
数字 1. Workflow of the fetal fMRI preprocessing pipeline. Key steps validated in this study are colored by blue boxes.
网络神经科学
705
Evaluation of fetal fMRI preprocessing steps
fMRI preprocessing:
A set of image processing steps to
clean and standardize fMRI data
before statistical analysis.
2018) and excluded. Brainsuite (Shattuck & Leahy, 2002) was used to manually generate fetal
brain masks around single reference images that were then applied to each resulting contin-
uous, low-motion 4D segment. After implementing volume-to-volume motion correction using
SPM’s ‘Realign’ function within each segment, we then evaluated the effects of different strat-
egies in several key fMRI preprocessing steps:
1. Normalization to age-matched versus 32-week template. To assess the influence of
different templates on the quality of normalization, we tested normalization from the
functional data directly to the standard template of a 32-week GA fetus (mean age for
the group) versus to the nearest week-specific template for a given subject (测距
从 25 到 37 weeks GA). Serag’s 2012 templates were used (Serag et al., 2012),
and normalization was conducted in SPM using nonlinear warping. The warping met-
rics were estimated with the first volume of each segment and were then applied to
remaining volumes within that segment. Two metrics (Calhoun et al., 2017) were used
for comparison: (1) voxel-wise variability of the normalized images across subjects; (2)
mean and maximum absolute frame-to-frame displacement derived from performing
volume-to-volume realignment, a second time, across the full normalized,
concatenated time series. Voxel-wise variability provides a measure for mismatch
between fMRI data and the template across subjects. If a given voxel is on the edge
and varying constantly between being “in” and “out” of the brain, this voxel will tend
to have a high standard deviation. 具体来说, the first normalized volume of subjects
at the same gestational age were concatenated along the fourth dimension to create a
single image file. We calculated the standard deviation of this file along the subject
dimension using Image Calculator of DPABI toolbox (Yan et al., 2016) 实施的
in MATLAB. Measurements of absolute displacement of the brain from the original
position included total translational movement (maximum and mean difference in
position in millimeters) and total head rotation (maximum difference in rotation in
degrees; Van Dijk et al., 2012). Scans with more accurate normalization across seg-
ments are expected to show lower intersubject displacement (Calhoun et al., 2017).
最后, to explore possible effects resulting from the choice of normalization templates
用过的, we additionally evaluated normalization to alternative age-specific fetal tem-
plates (Gholipour et al., 2017). This was a secondary analysis and was thus performed
for one representative subject from each gestational age.
After normalization, segments were concatenated within each scan and potential
misalignments between segments were corrected using SPM’s realignment function.
For the following processing step comparisons, data resulting from normalization to
the 32-week template were used. One subject was excluded here due to low usable
frames (n = 120 for the following analyses).
2. Masking the full concatenated data. To repress background spurious signals, a next
step evaluated the utility of applying a dilated brain mask. We tested whether masking
at this step improved downstream processing.
3. Denoising at two stringency thresholds, based on ICA. Data were decomposed into
independent components using FSL’s MELODIC (multivariate exploratory linear opti-
mized decomposition into independent components; 贝克曼 & 史密斯, 2004). 这
number of components was automatically estimated by MELODIC. Noise components
were manually labeled twice, once in a less stringent way and once in a more stringent
方式 (Griffanti et al., 2017). With the less stringent method, components showing
nonbiological spatial banding patterns, ring-like patterns on edges of the brain, AND
high-frequency peaks were labeled as noise; with the more stringent method,
网络神经科学
706
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
A
_
0
0
2
5
4
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Evaluation of fetal fMRI preprocessing steps
components showing banding patterns, ring-like patterns on edges of the brain OR
clusters mainly located in the white matter or cerebrospinal fluid OR time series with
sudden jumps (caused by segment concatenation) OR significant changes in oscillation
patterns OR high-frequency peaks were labeled as noise. 一般来说, the main differ-
ence between the two thresholds is whether noise is defined on the basis of temporal
or spatial features, 独自的, or in combination of both. In this study, components showed
abnormal temporal features due to segment concatenation, such as sudden jumps
(图2A) and alterations of oscillation patterns (Figure 2E), are unique to fetal
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
A
_
0
0
2
5
4
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
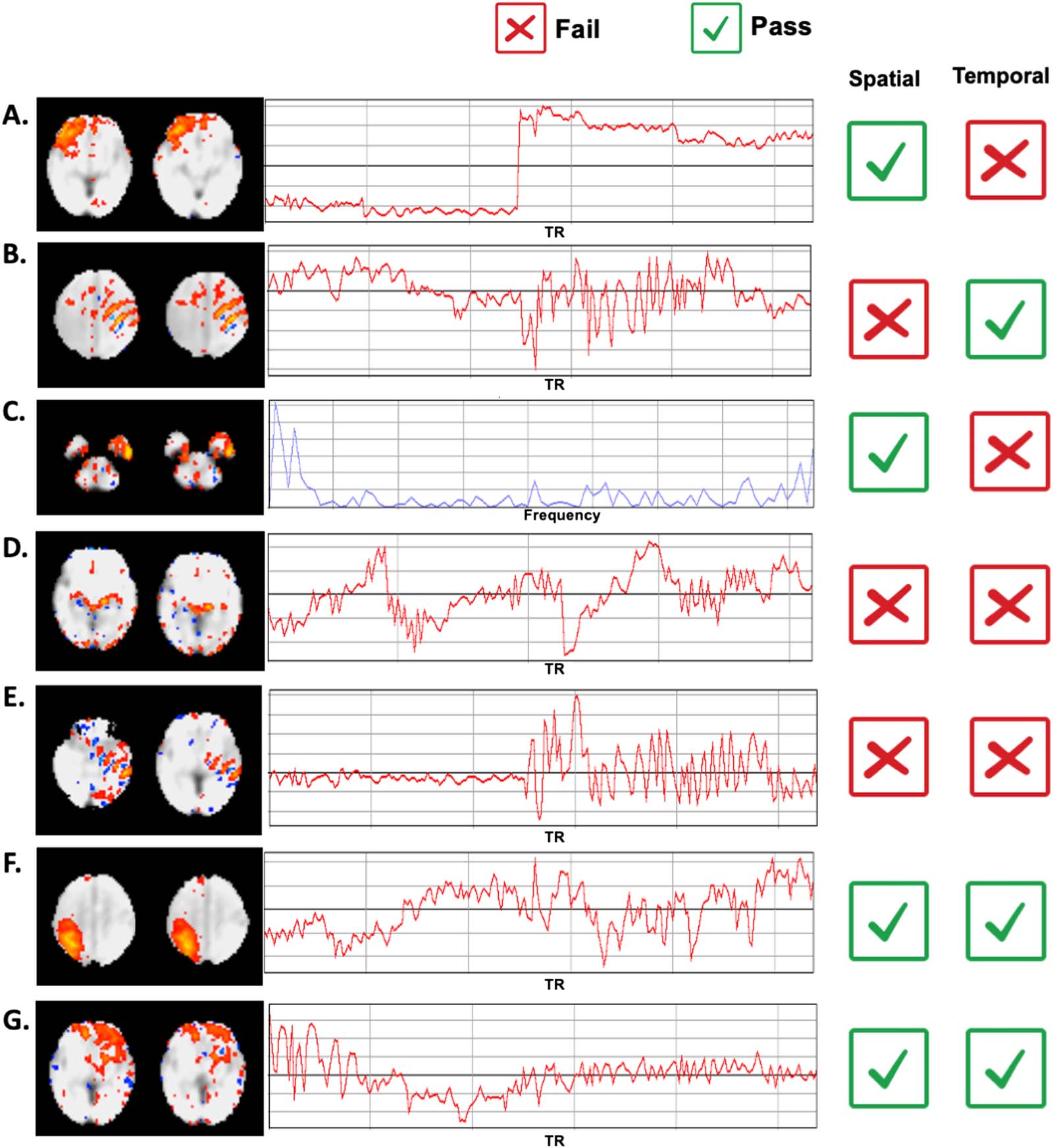
数字 2. Strategy used for less and more stringent ICA elimination. Exemplar components observed in the fetal dataset are presented above.
Observation of a single failure in spatial, 颞, or frequency domains results in elimination of the component, but only at the more strin-
gent level. Less stringent correction only eliminates components if more than one failure is observed, 例如, in both spatial and temporal
域. Pass and fail examples are provided here, depicted with checkbox and cross-out, 分别. As examples of single failures, com-
ponent B shows nonbiological banding patterns (positive/negative stripes), but shows acceptable time course, and component C shows a
typical spatial pattern, but shows high-frequency peaks, indicative of scanner-related artifacts. Examples A–C, were eliminated only at the
more stringent threshold. D and E show failures in two domains, and F and G pass both spatial and temporal analysis.
网络神经科学
707
Evaluation of fetal fMRI preprocessing steps
Spatial smoothing:
A step of preprocessing aiming to
enhance the signal-to-noise ratio, 经过
averaging data points with their
neighbors.
Voxel-mirrored homotopic
连接性 ( VMHC):
A voxel-wise measure of functional
connectivity between hemispheres
by computing correlations between
images and their left-right mirror
version.
Seed-based functional connectivity:
A voxel-wise correlation analysis
between a predefined region, 那是,
the seed, and all other voxels in the
脑.
成像. Criteria that formed the basis of each exclusion level are depicted in Figure 2.
Noise components were removed using the fsl_regfilt function. As outlined in the
following section, we evaluated the effect of ICA denoising on RSFC measures resulting
from each approach.
4. Smoothing with different kernel sizes. We tested the effect of smoothing kernels of
0 毫米 (no smoothing), 2 毫米, 和 4 mm full-width at half maximum (FWHM) 和
SPM. The chosen kernels equal 1 或者 2 times of our voxel size. We quantified the effects
of spatial smoothing by evaluating mean cross-hemisphere functional connectivity
strength across different kernel sizes.
Functional Connectivity Analysis
Cross-hemisphere functional connectivity. We examined RSFC between homotopic voxels in the
brain by using the voxel-mirrored homotopic connectivity ( VMHC) 技术 (Zuo et al.,
2010), which is a voxel-wise correlation analysis between the images and their left-right mirror
version. Preprocessing pipelines were compared on the basis of resultant summary measures
of homotopic functional connectivity for each fetus. As an additional means of evaluating the
above preprocessing pipeline, global mean VMHC was tested for correlation with the number
of frames and head motion parameters derived from the entire time course across subjects
using R software (version 4.0.5). Adequate removal of noise in preprocessing steps should
be reflected in lack of association of RSFC with frame count and motion.
Seed-based functional connectivity. Seeds were selected to represent regions both distal and
proximal to the midline. Seeds were defined manually as spheres with a 3-mm radius
(179 voxels), centered on MNI coordinates: (−20.6, 19.8, −8.6), (−7.7, −18.9, −26.6),
(−8.6, 13.7, −0.8), (−5.2, −21.4, 8.6), (−5.2, 33.5, −8.6), (−9.5, −5.2, 22.3), (−8.6, −5.2,
−4.3), 和 (−3.4, −39.5, −4.3). These were constructed using Mango Multi-image Analysis
软件 (https://ric.uthscsa.edu/mango/mango.html). Locations approximate the anterior
insular, cerebellum, putamen, precuneus, medial prefrontal cortex, supplementary motor
区域, thalamus, and the visual cortex, 分别, in the 32-week fetal template (Serag
等人。, 2012). Seeds were selected to approximate locations used in prior research in preterm
and term newborns (Smyser et al., 2010) and because functional neural networks related to
these seeds are evidenced to be sensitive to early brain development (Thomason et al.,
2015). These left hemisphere masks were duplicated for the right hemisphere, resulting
in a total of 16 seed regions. Seed regions of interest (ROIs) are represented in Figure S2
的
the Supporting Information and files themselves are available online at www
.brainnexus.com.
Seed-to-voxel whole-brain analyses were performed on the denoised data in DPABI tool-
盒子. For each subject, the mean time course was extracted from each seed region and corre-
lated with the time course of each voxel throughout the whole brain, yielding individual RSFC
maps for each seed region. All RSFC maps were converted to z-scores for post hoc analyses.
Seed-based RSFC maps of less versus more stringently denoised data were compared using
paired two-sample t test. Clusters were corrected for whole-brain multiple comparisons by
using false discovery rate (FDR) p < 0.05. Group mean RSFC maps were estimated by one-
sample t tests testing the z-transformed values against 0, with threshold at p < 0.00001 FDR
corrected.
In addition to the above voxel-wise
Group-level ICA as a validation of the proposed pipeline.
and seed-based functional connectivity analyses used for evaluation of preprocessing
Network Neuroscience
708
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
a
_
0
0
2
5
4
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Evaluation of fetal fMRI preprocessing steps
strategies, we conducted a brain network analysis. To extract group-level intrinsic connec-
tivity networks, we performed spatial ICA implemented in group ICA of Functional MRI
Toolbox (GIFT v3.0b, https://trendscenter.org/software/gift/). Optimally preprocessed
fMRI data (more stringently denoised, 4-mm FWHM smoothed) were decomposed into 35
spatial components, each of which exhibited a unique time course profile based on the Info-
max algorithm. The number of components was estimated based on the image quality by
using a minimum description length approach (Rissanen, 1978). A higher order ICA
approach was applied to improve functional parcellation (Kiviniemi et al., 2009). Reliability
and stability of the algorithm was ensured using ICASSO by repeating the component esti-
mation 20 times (Himberg et al., 2004). Subject-specific spatial maps and time courses were
obtained using the back-reconstruction approach (GICA; Calhoun et al., 2004) and con-
verted to z-scores.
RESULTS
Assessment of Spatial Normalization to Age-Matched Versus 32-Week Template
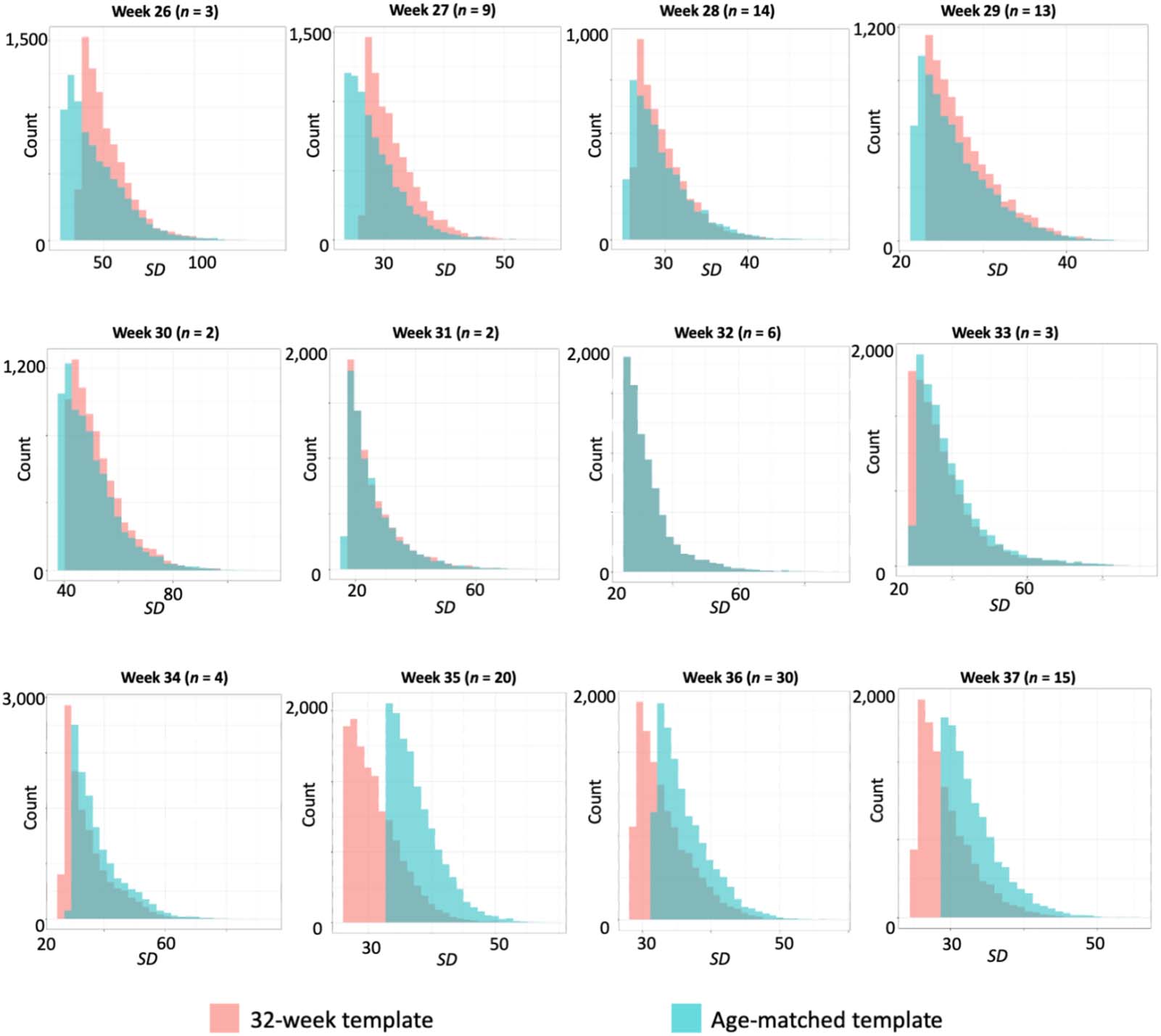
When plotting the distribution of 8,000 voxels with the highest standard deviation (Figure 3),
we observe that intersubject alignment was improved (SD reduced) when using the
32-week template for fetuses older than 32 weeks. However, the reverse was true for
younger fetuses; there, an age-matched template was associated with reduced intersubject
alignment and better normalization performance. We also observe that the areas of greatest
variability are identified at the edges of the brain (see Figure S3 in Supporting Information).
This finding may reflect a combination of greater displacement associated with distance
from origin and also stronger BOLD signal in cortex compared to white matter and cere-
brospinal fluid. We did not find a significant difference in mean and maximum subject-to-
subject displacement between the templates at any fetal age (see Figure S4 in Supporting
Information). Furthermore, we did not observe a marked difference in normalization per-
formance when an alternate fetal anatomical
template set was used (see Figure S5 in
Supporting Information).
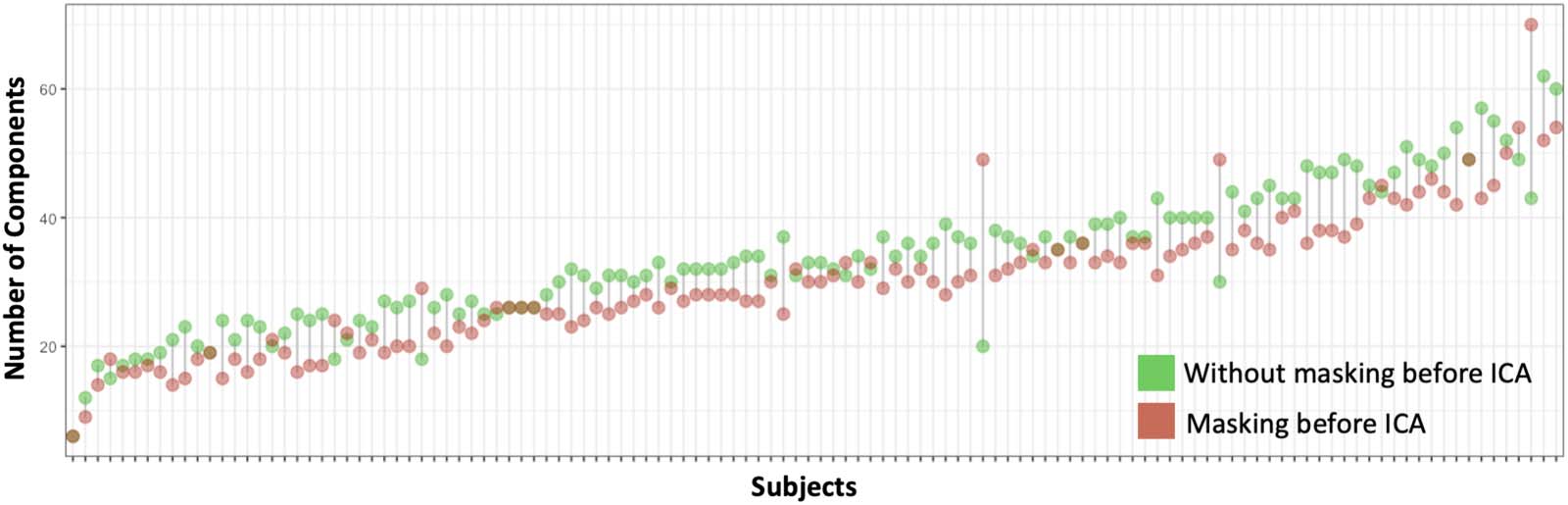
Assessment of Individual-Level Masking
With unmasked data, we detected a number of noise components located outside of the brain.
We found that masking before denoising reduced the number of ICA-derived components
(Figure 4) for most fetal subjects; the total number of independent components across all
subjects decreased from 4,043 to 3,623 after reapplying a brain mask. The individual-level
masking removed most of the ring-like noise components in the following ICA analysis. This
reduction of components alleviates the workload of manual inspection, which is currently
needed for fetal data. The dilated mask is shown in Figure S6 of the Supporting Information,
and is available online at https://www.brainnexus.com/.
Assessment of ICA Denoising
The less stringent approach to labeling noise components resulted in the identification of 5%
of all ICA-derived components as noise. In contrast, the more stringent approach resulted in
60% of components being labeled as noise. More stringent denoising led to improvement by
visual inspection; in particular, we observed reductions in spatial banding patterns, intensity
inhomogeneity, and abnormal signal oscillations caused by rapid motion or segment
Network Neuroscience
709
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
a
_
0
0
2
5
4
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Evaluation of fetal fMRI preprocessing steps
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
a
_
0
0
2
5
4
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Distributions of standard deviation values after normalization, by fetal age and by template used. Fetuses of different ages were
normalized either to a 32-week template (mean for the group) or to a same-age template. Voxels on the edge of the brain have lower
standard deviation if they are consistently characterized the same way. The 8,000 voxels with the highest standard deviation are plotted
here. Review of observed distributions suggest that the 32-week template performs more optimally for fetuses older than 32 weeks, seen in a
leftward shift of 32-week values. The reverse is noted for fetuses younger than 32 weeks, where the age-matched template corresponds to a
leftward shift.
Figure 4. Number of components estimated in FSL’s MELODIC by subjects with and without masking.
Network Neuroscience
710
Evaluation of fetal fMRI preprocessing steps
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
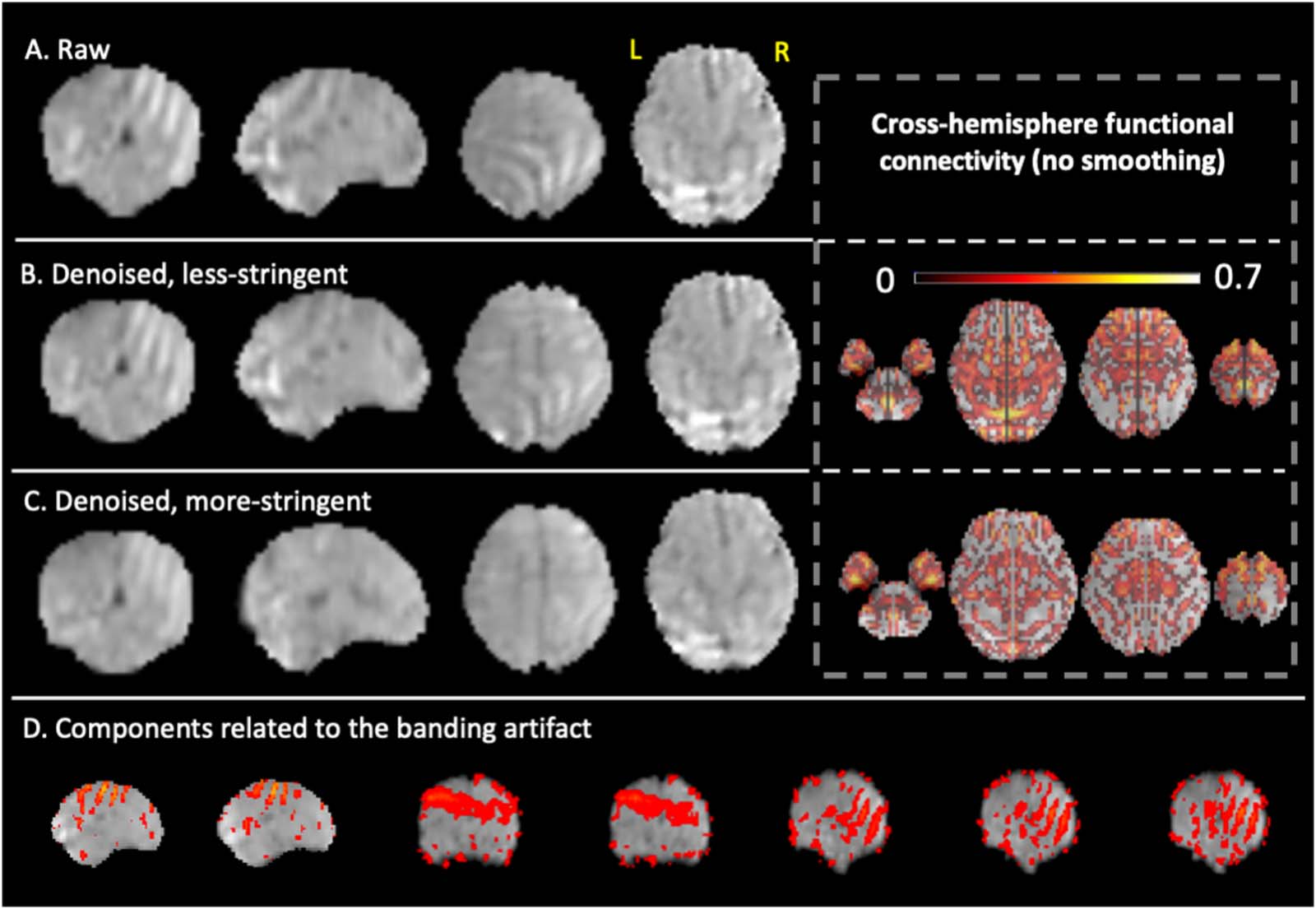
Figure 5. Comparison of less stringent versus more stringent ICA denoising in a representative subject, Case 1. A single volume is shown for a
case (35 weeks GA) presenting severe nonbiological banding patterns. (A) Raw data with different planes. (B) Less stringently denoised data
with different planes (left), and corresponding cross-hemisphere RSFC (right). (C) More stringently denoised data with different planes (left) and
corresponding cross-hemisphere RSFC (right). (D) Examples of ICA noise components related to the banding artifact.
concatenation. Comparison of the two strategies in three representative subjects are shown
below for qualitative inspection, following by group-level RSFC comparisons.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
a
_
0
0
2
5
4
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Case 1: A representative subject with severe nonbiological banding patterns in the posterior
part of the brain was selected as Case 1. As shown in Figure 5, the nonbiological
banding patterns were detected with ICA (bottom row) and were slightly lessened
with a less stringent denoising approach. The presence of this nonphysiological band-
ing pattern is usually related to the MRI sequence (e.g., EPI susceptibility or multiband
acceleration) or hardware artifacts (e.g., RF interference) or interactions of the acqui-
sition with head motion (e.g., interleaved slice acquisitions) (Griffanti et al., 2017). In
contrast, the more stringent denoising further improved homogeneity within the brain
and was associated with reduced cross-hemisphere functional connectivity. It is pos-
sible that banding patterns remain at the less stringent level of denoising, because
even though the spatial pattern is atypical, the time series falls within the normal
range.
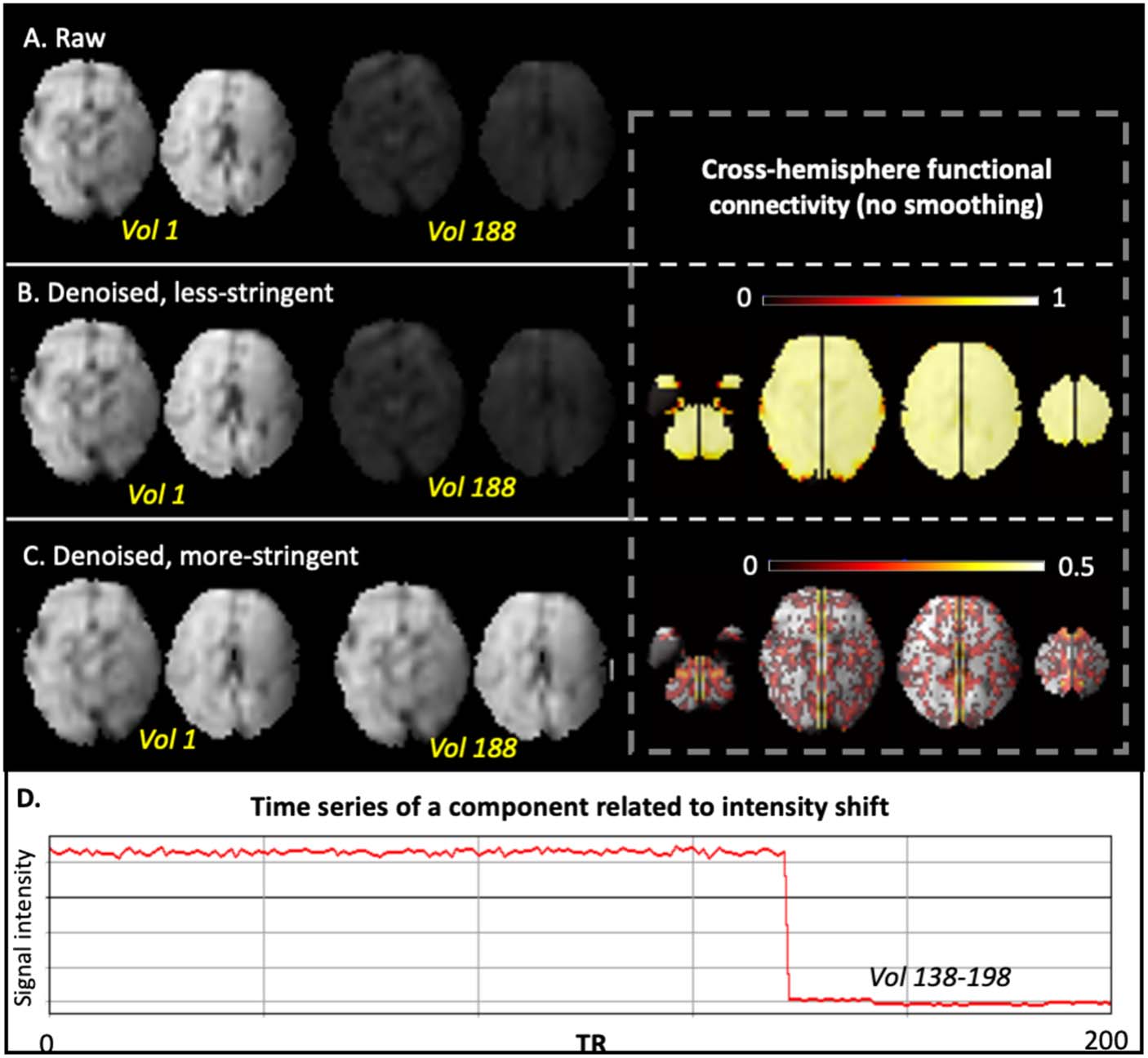
Case 2: When checking components, we noticed an unusual case with massive whole-
brain intensity shift during the scan, which was likely due to an issue in the coil or
other electronics. With the more stringent strategy, we labeled all components as
noise for this subject, as all components shared the same time course that contained
a sudden jump. The intensity shift was completely corrected in this approach. As
could be expected, without this correction, the cross-hemisphere functional connec-
tivity is biologically implausible across the whole brain (Figure 6).
Network Neuroscience
711
Evaluation of fetal fMRI preprocessing steps
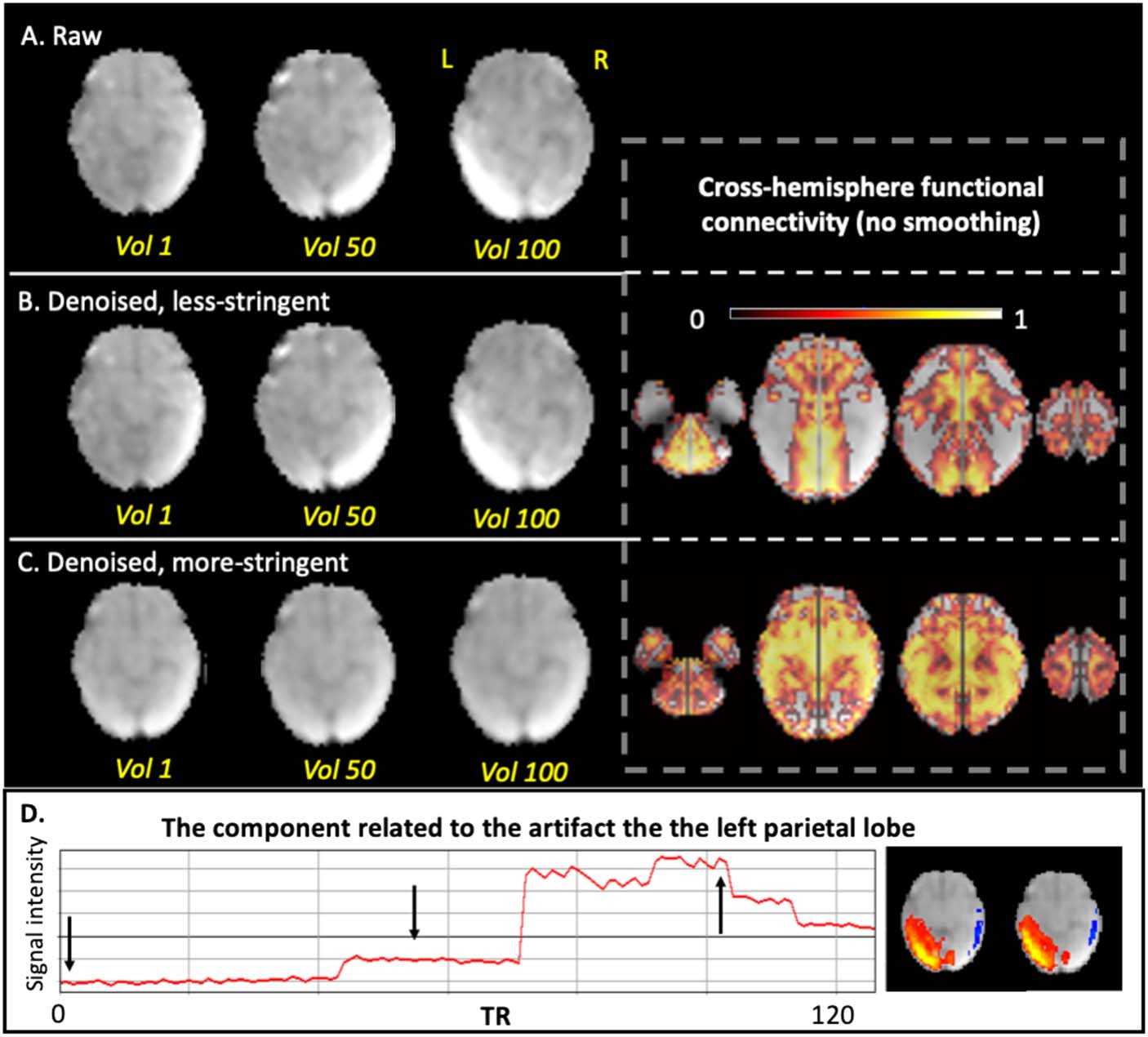
Case 3: In the third subject, there was a sharp increase in intensity in the left parietal
cortex in the middle of the scan that corresponded to one ICA-derived component
(the bottom row of Figure 7). With more stringent ICA denoising, artifacts were
removed (Figure 7) and cross-hemisphere functional connectivity increased. Strong
intensity changes in the time course at the joint of two segments, observed in
Figure 7D, results from both (1) normalization misalignment between segments
and/or (2) fetal repositioning. These signal “jumps” thus reflect inconsistent
segment-to-segment spatial alignment (imprecise normalization) or can reflect siz-
able fetal repositioning with potential to change field geometry or interactions at
tissue interfaces.
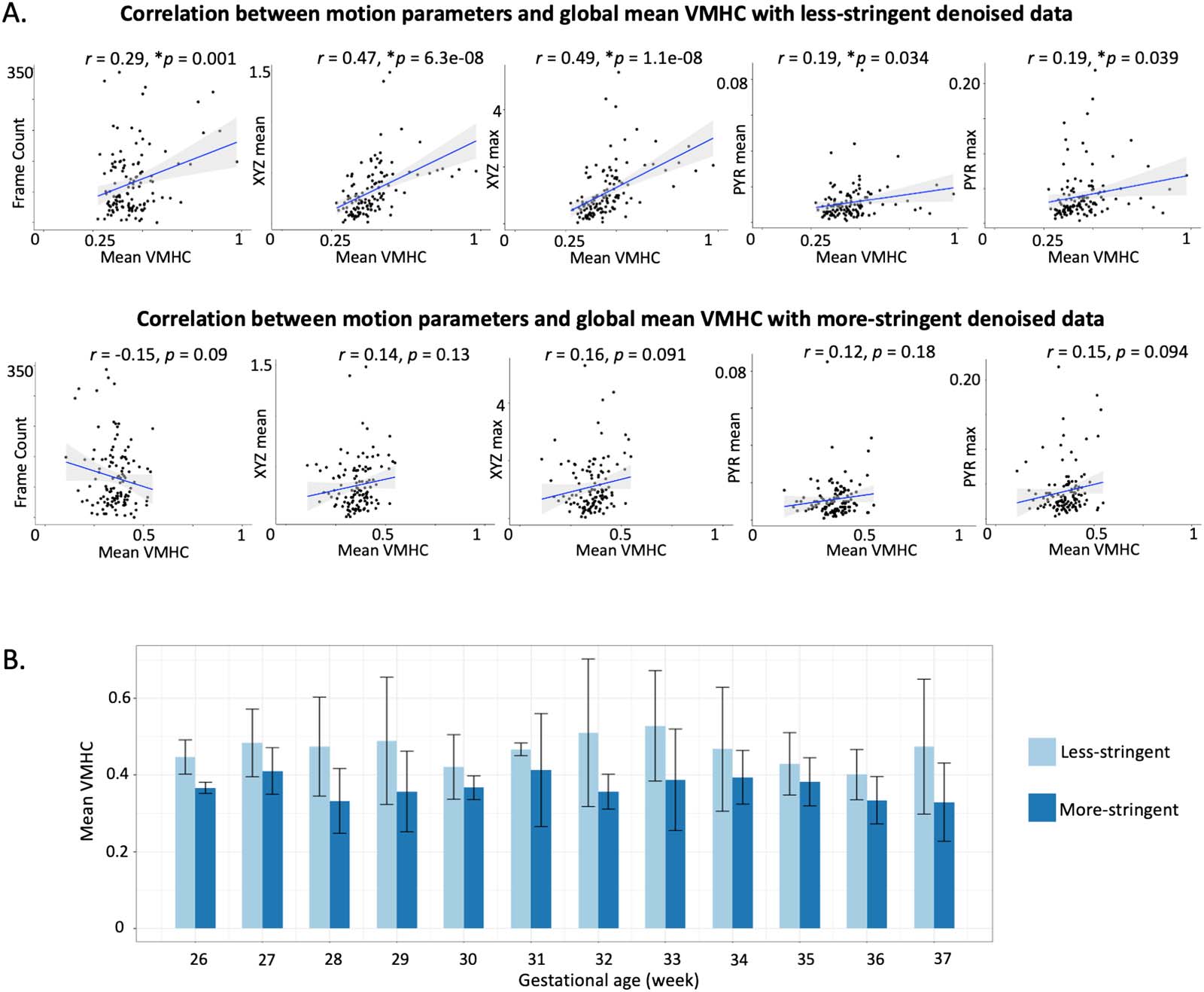
Denoising effects on group-level cross-hemisphere functional connectivity. Global mean cross-
hemisphere RSFC was correlated with number of frames (r = 0.29, p = 0.001) and with motion
parameters (mean translational movements: r = 0.47, p = 6.3e−08; maximum translational
movements: r = 0.49, p = 1.1e−08; mean rotations: r = 0.19, p = 0.03; maximum rotations:
r = 0.19, p = 0.039) following less stringent denoising (top row of Figure 8A). After regressing
out noise components by using the more stringent approach, the associations between RSFC,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
a
_
0
0
2
5
4
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Comparison of less stringent versus more stringent ICA denoising in a representative sub-
ject, Case 2. A single volume is shown for a case with noted intensity shift. (A) Raw data of two axial
slices at the 1st and 188th volumes. (B) Less stringently denoised data at the 1st and 188th volumes
(left) and corresponding cross-hemisphere RSFC (right). (C) More stringently denoised data at the 1st
and 188th volumes (left) and corresponding cross-hemisphere RSFC (right). (D) Time series of an
example noise component.
Network Neuroscience
712
Evaluation of fetal fMRI preprocessing steps
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
a
_
0
0
2
5
4
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 7. Comparison of less stringent versus more stringent ICA denoising in a representative sub-
ject, Case 3. A single volume is shown for a case with high residual motion. (A) Raw data of one
slice at the 1st, 50th, and the 100th volumes. (B) Less stringently denoised data at the 1st, 50th, and
the 100th volumes (left) and corresponding cross-hemisphere RSFC (right). (C) More stringently
denoised data at the 1st, 50th, and the 100th volumes (left) and corresponding cross-hemisphere
RSFC (right). (D) The time course and the spatial map of the noise component corresponding to the
artifact in left parietal cortex. Arrows indicate the volumes we showed in the above rows.
frame count, and motion were no longer significant (bottom row of Figure 8A). Overall, a more
stringent denoising strategy corresponded to a reduction in cross-hemisphere RSFC (Figure 8B).
However, it is notable that decreased cross-hemispheric connectivity was not a ubiquitous
feature of denoising; individual cases showed increases in cross-hemispheric connectivity
when applying a more stringent ICA. For example, in Case 3 (Figure 7), after removing left-
lateralized artifact, we observe increased cross-hemisphere RSFC, suggestive of unmasking
underlying connectivity effects.
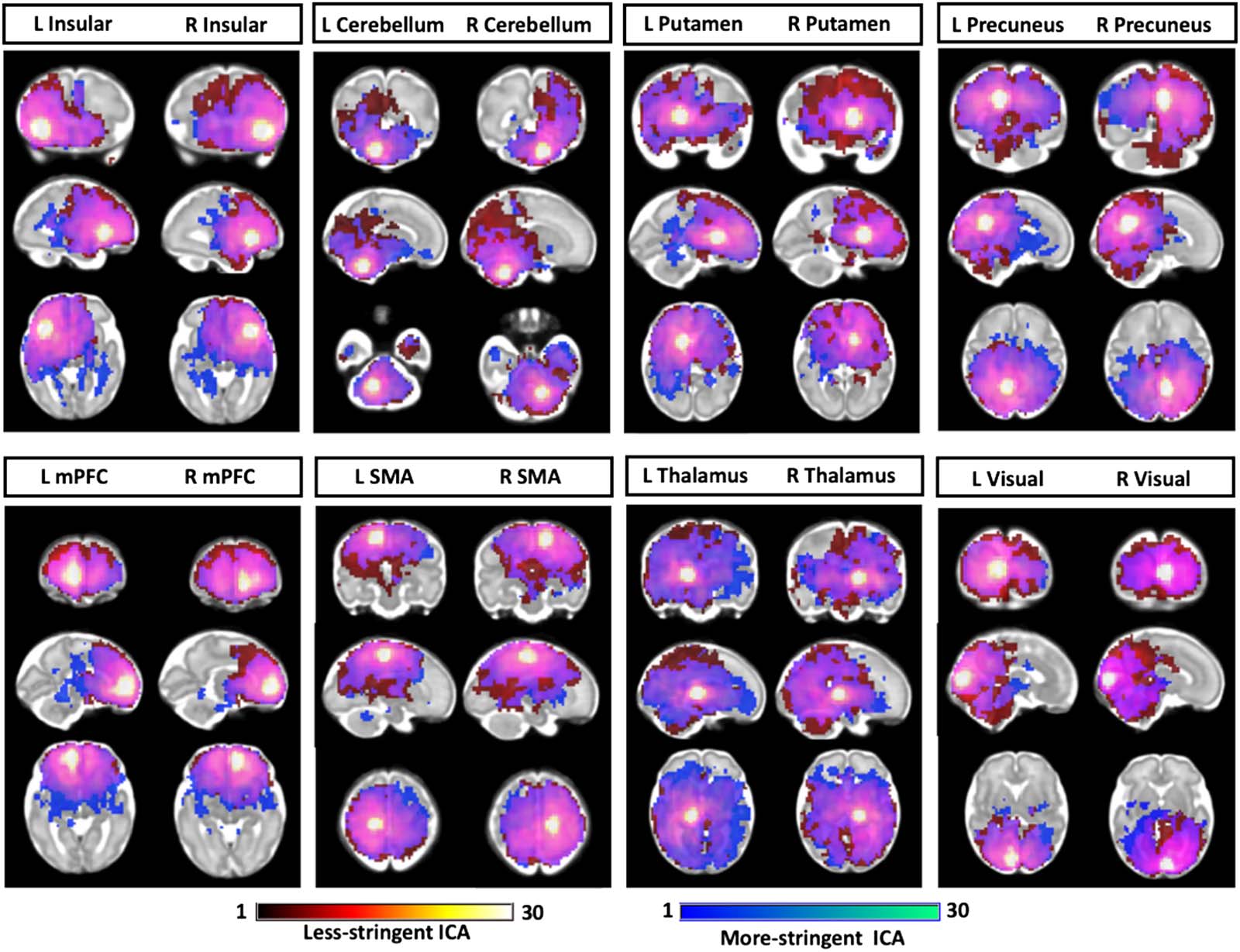
Denoising effects on group-level seed-based functional connectivity. Use of different denoising
levels was associated with changes in the pattern of RSFC across ROIs, as shown in
Figure 9. Select regions showed increased or decreased connectivity at each denoising thresh-
old. Overall, the pattern was such that less stringent denoising was associated with greater
overall RSFC, demonstrated in Figure S7 of the Supporting Information.
Assessment of Spatial Smoothing
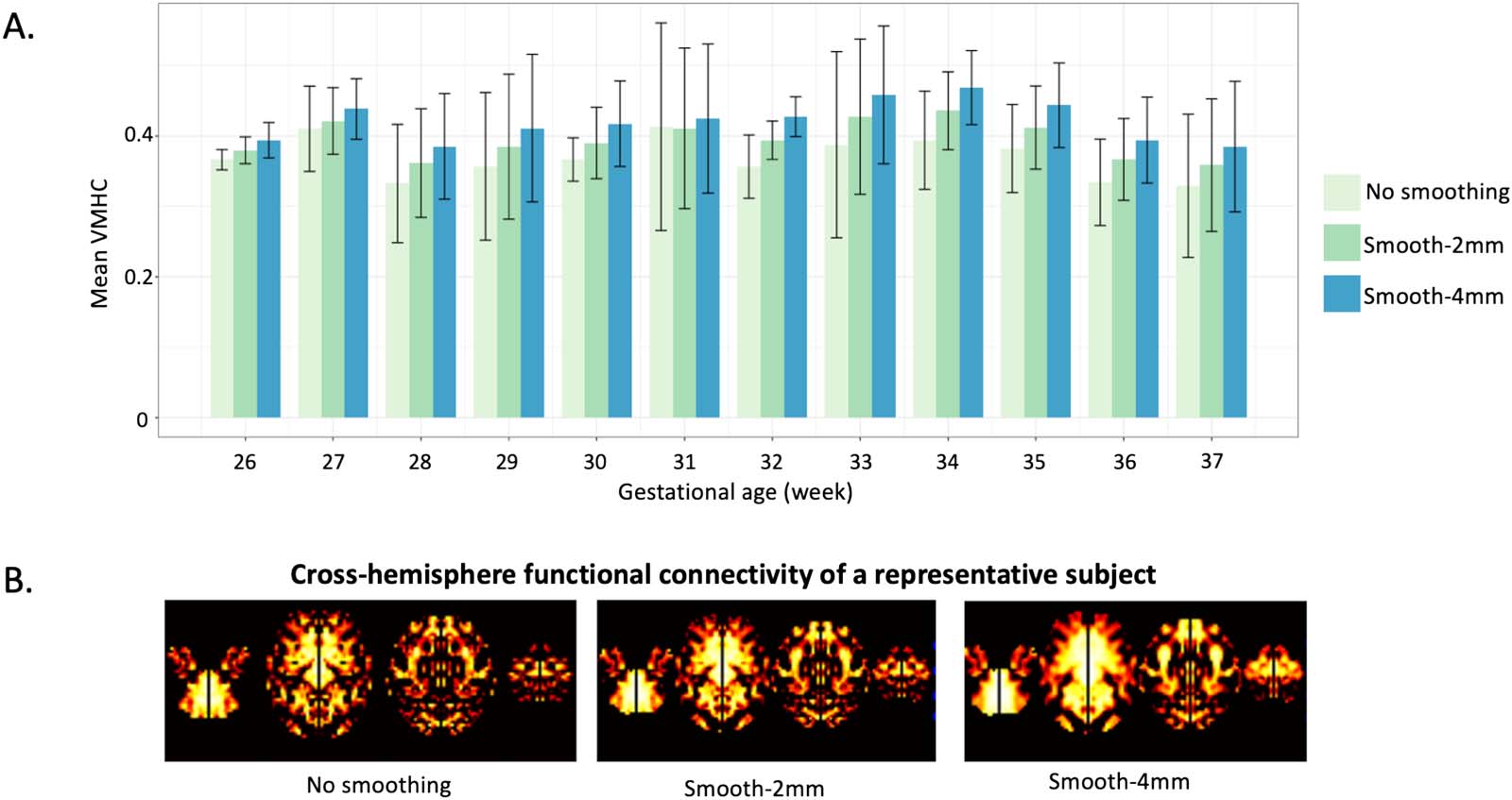
Generally, we observed a dose-dependent relationship between smoothing kernel size and
cross-hemisphere functional connectivity, with a 4-mm kernel resulting in the greatest
Network Neuroscience
713
Evaluation of fetal fMRI preprocessing steps
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
Figure 8. Group-wise comparison of different ICA denoising strategies. (A) Correlations of global mean voxel-mirrored homotopic connec-
tivity ( VMHC) with frame counts and motion parameters (XYZ mean and XYZ max for translational movements; PYR mean and PYR max for
rotations). (Top row) Less stringently denoised data; (bottom row) more stringently denoised data. (B) Group mean VMHC by age group with
less versus more stringent denoising methods. Asterisks (*) in front of p values indicate significant correlations.
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
a
_
0
0
2
5
4
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
cross-hemisphere functional connectivity. We also note that this effect does not interact
with age (Figure 10), suggesting the impact of smoothing kernel size does not vary with
fetal age.
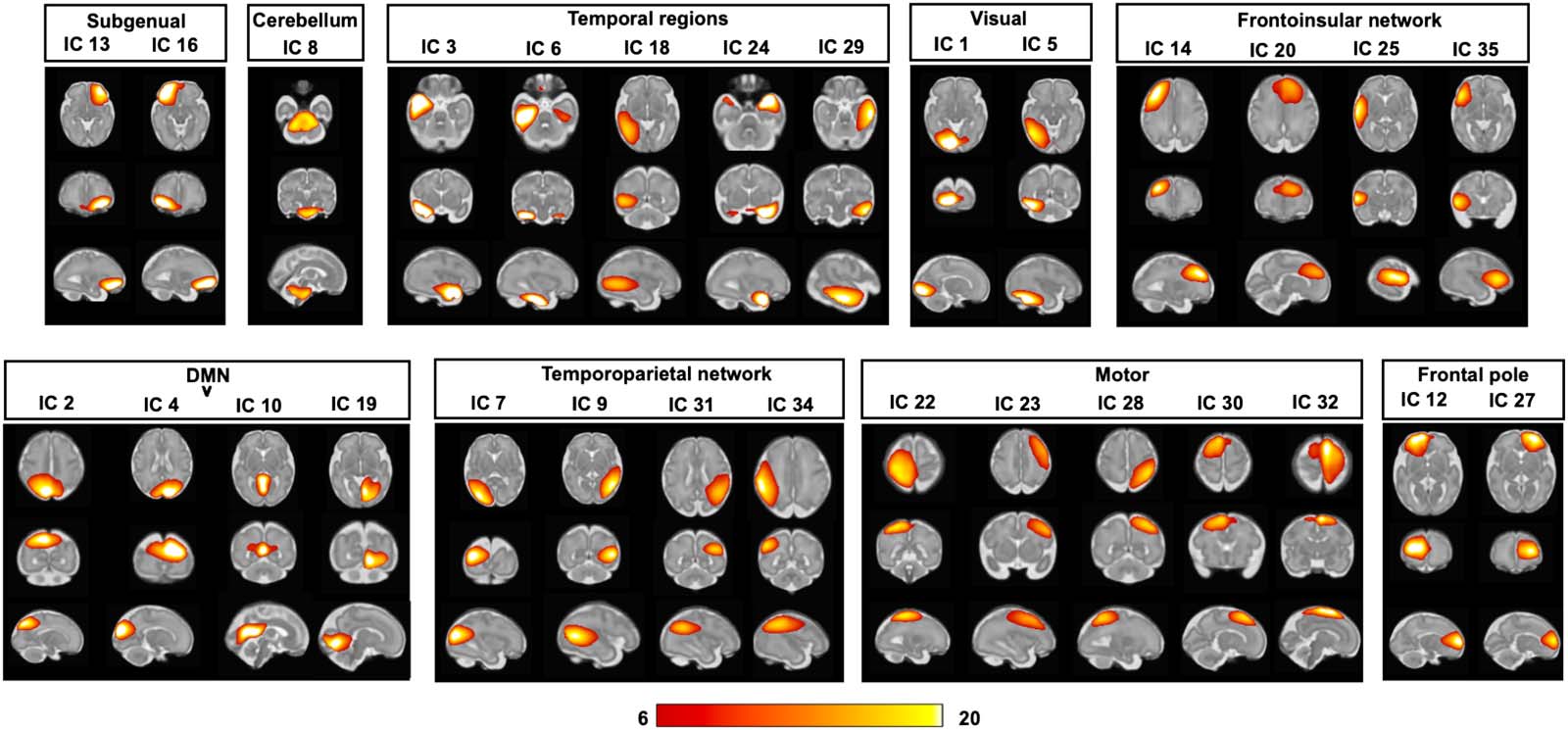
Resting-State Functional Networks in the Fetal Brain
In an exploratory validation analysis, we examined the presence of fetal resting-state net-
works in more stringent denoised ICA components (4-mm FWHM smoothed). Thirty-one of
the 35 components (available online at www.brainnexus.com) were identified as signal com-
ponents because their peak coordinates were located primarily in gray matter and their time
courses were dominated by low-frequency fluctuations (Allen et al., 2011). We organized the
signal components into nine functional networks based on the temporal correlation between
the components and the anatomical locations, including the subgenual area, cerebellum,
temporal regions, visual network, frontoinsular network, default mode network, temporopar-
ietal network, motor network, and frontal pole areas (Figure 11). Examples of group-level ICA
noise are shown in Figure S8 of the Supporting Information. The identified components spa-
tially resembled those previously described in preterm neonates (Smyser et al., 2010) and
fetuses (Thomason et al., 2015).
Network Neuroscience
714
Evaluation of fetal fMRI preprocessing steps
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
Figure 9. Comparison of seed-based functional connectivity in data analyzed with more or less stringent denoising. One-sample t test
was sued to compare more stringent ICA (blue) and less stringent ICA (red) (p < 0.00001, FDR corrected). Overlapping regions are shown
in purple.
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
a
_
0
0
2
5
4
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 10. Group-wise comparison of different smoothing kernels. (A) Global mean VMHC changes without or with smoothing kernels of
2 mm and 4 mm. (B) Voxel-wise VMHC of a representative fetus of 37 weeks.
Network Neuroscience
715
Evaluation of fetal fMRI preprocessing steps
Figure 11. Fetal brain networks derived from ICA. Positive t maps threshold at t value > 6 are shown.
讨论
Our analyses confirm that different approaches to normalization, masking, 去噪, 和
smoothing during fetal fMRI preprocessing have notable impacts on data quality. Specific ana-
lytic choices during preprocessing impact connectivity metrics derived from BOLD images at
the subject level, with implications for reliability and reproducibility of group-level effects.
Results indicate that choice of template relates to normalization variability in an age-
dependent manner. 具体来说, using a 32-week template resulted in greater normalization
accuracy compared to using age-matched templates for fetuses 32 weeks or older. 反过来,
using an age-matched template results in greater normalization accuracy for fetuses under
32 weeks gestational age. It is possible that either maturational changes within the brain,
variation in size and shape, or some combination of these contribute to this effect. 考试用-
普莱, wide difference in size between the source and template images leads to more scaling
transformations and increased interpolation. These data also show that when a large fetal
age range is studied, fetuses at extreme edges of the range will be most impacted by choice
of normalization template. 全面的, these observations suggest that template choice may best
be determined by research objectives and characteristics of the sample. 例如, 如果
age-related development is not the goal of the study, it may be advisable to include gestational
age at scan as a nuisance covariate in second-level models and/or to exclude cases to restrict
the age range being studied. If the study objective(s) includes age-related development, 意思是
voxel-wise standard deviation can be considered as a regressor to correct for normalization
differences across age. In future work it would be valuable to evaluate alternative registration
algorithms that are not reliant on off-the-shelf tools.
In ICA-based denoising using FSL’s MELODIC, a high proportion of noise components is
usually expected in adults. With standard sequences at 3T, 大约 70% (Rummel et al., 2013)
和 88% (Griffanti et al., 2014) of components may be reported as noise. In our fetal dataset,
even using a more stringent strategy, 仅有的 60% of components were identified as noise, 大多数
which were motion related. Observing a lower proportion of noise components in fetal fMRI
data may be the result of either developmental processes, such as changes in cerebrovascular
结构 (Reilly & Gutierrez, 2021), or altered noise characteristics unique to this context. 作为
examples of the latter, abdominal versus head coil geometry or field inhomogeneity
网络神经科学
716
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
A
_
0
0
2
5
4
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Evaluation of fetal fMRI preprocessing steps
introduced by large field of view may contribute to differential effects. 另一种可能性是
that a smaller number of components arises simply from larger motion-induced artifacts wash-
ing out smaller, more subtle artifacts. Because ICA denoising may perform differently in fetal
数据, it is advisable that comparisons between fetal and postnatal datasets take this possibility
into account.
We also observed that more stringent denoising removed the correlation between all
motion parameters and global cross-hemisphere RSFC and resulted in different seed-based
RSFC maps. These findings highlight the sensitivity of voxel-to-voxel and seed-to-voxel RSFC
to head motion. 然而, we also want to acknowledge that the more stringent denoising
does not guarantee “better” estimates of functional connectivity, considering the risk of poten-
tial inadvertent removal of signal. Selection of cross-hemispheric connectivity was the refer-
ence analysis used in the present study because it has been evidenced in previous studies a
sensitive measure of fetal development (Thomason et al., 2013) and because cerebral homo-
topy is a fundamental principle of brain organization (Toga & 汤普森, 2003). 然而, 它
should be noted that this is one of many strategies that could be used to test the effects of
denoising on resultant RSFC. This method was useful in confirming that after denoising, 一个
expected pattern in brain organization was observed and was no longer correlated with
motion parameters.
In fetal fMRI, motion-related artifact is a significant challenge; motion artifacts can be intro-
duced both by frequent and large-scale changes in fetal position in utero and by maternal res-
piration (Thomason, 2020). One previous attempt to remove motion-related artifacts in fetal
fMRI employed a combined approach of slice to volume registration and scattered data interpo-
lation with bias field and spin history corrections on a small sample of eight fetuses (Ferrazzi
等人。, 2014). This approach avoids discarding frames, but requires additional scans to assist reg-
istration and estimations of field inhomogeneity. 相比之下, ICA denoising can be implemented
without additional scans. At the single-subject level, ICA-based denoising has proven to be a
powerful tool for separating neural-related signal from different sources of noise, 包括
movement artifacts (Griffanti et al., 2017). This is the first study to verify the efficacy of ICA
denoising in fetal imaging in a large fetal cohort. 此外, the components manually labeled
here can be used as a training set for future automatic signal/noise classifiers in fetal imaging
数据. Given ICA does not guarantee a uniform reconstruction of all frames, especially for
high-motion periods, combination of censoring, ICA-based denoising, and covariate regression
(例如, motion, fame count, ETC。) may be advisable for mitigating noise in future fetal fMRI studies.
Spatial smoothing can improve signal to noise and reduce the effects of spatial normaliza-
tion misalignment (Lowe & Sorenson, 1997) at the expense of decreasing resolution. 以前的
studies on adult brains suggest that the kernel size should be at least twice the size of the fMRI
acquisition voxel (Alahmadi, 2021). 然而, best practices for smoothing parameters in fetal
fMRI remain to be addressed. Given the significantly smaller size of fetal brains compared to
adult brains, the smoothing kernel recommended in adult imaging may not be appropriate. 我们
observed that using a larger smoothing kernel resulted in enhanced cross-hemisphere func-
tional connectivity in fetuses, fitting with likely improvement in signal to noise. 然而,
given the trade-off between sensitivity and spatial specificity (IE。, the resolution) of findings,
general advice in fetal RSFC could be to use a moderate value of ×1 to ×2 voxel size. 最好的
practices in kernel selection will necessarily vary with attributes of the data and with the ana-
lytic approach being applied.
While the present study examines key steps in preprocessing fetal fMRI data, 这很重要
to note that many questions remain to be addressed. Fetal data present novel challenges,
网络神经科学
717
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
A
_
0
0
2
5
4
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Evaluation of fetal fMRI preprocessing steps
especially higher and more complex motion and unique image artifacts (van den Heuvel &
Thomason, 2016). Methods tested in this article draw from parameters that are most standard
in published reports in the literature (Jakab et al., 2014; Thomason et al., 2019; Thomason
等人。, 2021A; Turk et al., 2019). 然而, there are alternative emergent approaches,
such as that presented by Scheinost and colleagues that perform automatic censoring of
low-quality frames and aim to correct for both large and small motion, that are important to
explore with advancement of this field of study (Scheinost et al., 2018). It will be valuable for
future works to additionally evaluate the efficacy of alternative denoising strategies, 例如
CompCorr (Behzadi et al., 2007), confound regression and band-pass filtering (Yan et al.,
2013), and advanced slice level reconstruction (Ferrazzi et al., 2014), and to do so across
datasets with variable noise profiles and motion thresholds. Future studies will also benefit
from considering interaction between preprocessing steps, such as whether the choice of
normalization target would affect the ICA performance. Additional areas for future work are
to test the generalizability of preprocessing steps in data collected from different
scanners/vendors, and across in different populations, including clinical samples. 此外,
it will be useful to evaluate alternative fMRI acquisition techniques, such as multiecho–echo
planar imaging (ME-EPI), which lends itself to empirically informed strategies for denoising
data during postprocessing (Kundu et al., 2012). 此外, an additional acquisition
mapping the field may help to address the distortions of the image. 最后, there is great prom-
ise in using deep learning algorithms to automate manual steps involved in fetal fMRI data
加工, such as brain segmentation (Rutherford et al., 2021). One can imagine further
development of these, even for purposes of automated identification of noise and signal com-
ponents following ICA. 全面的, fetal research MRI represents an extraordinary opportunity for
basic and clinical science, but it does require continued investment toward optimization and
transition to the mainstream. This is elaborated further in a recent commentary (Rajagopalan
等人。, 2021). The present study addresses common decision points in fMRI data processing
and provides empirical comparisons of outputs achieved when applying different methods
at each step.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
A
_
0
0
2
5
4
p
d
.
t
致谢
The authors thank Jasmine Hect and Pavan Jella for their assistance in data acquisition and
thank Ava Palopoli and Amyn Majbri for assistance with data management and quality assur-
安斯. 重要的, the authors thank participant families who generously shared their time and
expressed interest in helping future babies to achieve their best possible health outcomes.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00254.
The data and code used in this study will be made available via https://ndar.nih.gov/ and/or
accessed upon direct request to M. 乙. Thomason (数据) or L. 吉 (代码).
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
作者贡献
Lanxin Ji: 概念化; 数据管理; 形式分析; 方法; 软件;
可视化; Writing – original draft; 写作——复习 & 编辑. Cassandra L. Hendrix:
概念化; 可视化; Writing – original draft; 写作——复习 & 编辑. Moriah
乙. Thomason: 概念化; 资金获取; 资源; 监督; Writing –
review & 编辑.
网络神经科学
718
Evaluation of fetal fMRI preprocessing steps
资金信息
Moriah E. Thomason, Foundation for the National Institutes of Health (https://dx.doi.org/10
.13039/100000009), 奖项ID: MH110793. Moriah E. Thomason, Foundation for the
美国国立卫生研究院 (https://dx.doi.org/10.13039/100000009), 奖项ID:
DA050287. Moriah E. Thomason, Foundation for the National Institutes of Health (https://dx
.doi.org/10.13039/100000009), 奖项ID: MH122447. Moriah E. Thomason, Foundation for
the National Institutes of Health (https://dx.doi.org/10.13039/100000009), 奖项ID:
ES032294.
参考
Alahmadi, A. A. (2021). Effects of different smoothing on global and
regional resting functional connectivity. Neuroradiology, 63(1),
99–109. https://doi.org/10.1007/s00234-020-02523-8, 考研:
32840683
艾伦, 乙. A。, Erhardt, 乙. B., Damaraju, E., Gruner, W., Segall, J. M。,
席尔瓦, 右. F。, … Kalyanam, 右. (2011). A baseline for the multi-
variate comparison of resting-state networks. Frontiers in Systems
神经科学, 5, 2. https://doi.org/10.3389/fnsys.2011.00002,
考研: 21442040
安德森, A. L。, & Thomason, 中号. 乙. (2013). Functional plasticity
before the cradle: A review of neural functional imaging in the
human fetus. 神经科学 & Biobehavioral Reviews, 37(9),
2220–2232. https://doi.org/10.1016/j.neubiorev.2013.03.013,
考研: 23542738
贝克曼, C. F。, & 史密斯, S. 中号. (2004). Probabilistic independent
component analysis for functional magnetic resonance imaging.
IEEE Transactions on Medical Imaging, 23(2), 137–152. https://
doi.org/10.1109/TMI.2003.822821, 考研: 14964560
Behzadi, Y。, Restom, K., Liau, J。, & 刘, 时间. 时间. (2007). A component
based noise correction method (CompCor) for BOLD and perfu-
sion based fMRI. 神经影像, 37(1), 90–101. https://doi.org/10
.1016/j.neuroimage.2007.04.042, 考研: 17560126
Botvinik-Nezer, R。, Holzmeister, F。, Camerer, C. F。, Dreber, A。,
Huber, J。, Johannesson, M。, … Adcock, 右. A. (2020). Variability
in the analysis of a single neuroimaging dataset by many teams.
自然, 582(7810), 84–88. https://doi.org/10.1038/s41586-020
-2314-9, 考研: 32483374
Bright, 中号. G。, Tench, C. R。, & 墨菲, K. (2017). Potential pitfalls
when denoising resting state fMRI data using nuisance regres-
锡安. 神经影像, 154, 159–168. https://doi.org/10.1016/j
.neuroimage.2016.12.027, 考研: 28025128
Calhoun, V. D ., Adalı, T。, & Pekar, J. J. (2004). A method for com-
paring group fMRI data using independent component analysis:
Application to visual, motor and visuomotor tasks. Magnetic
Resonance Imaging, 22(9), 1181–1191. https://doi.org/10.1016/j
.mri.2004.09.004, 考研: 15607089
Calhoun, V. D ., Wager, 时间. D ., Krishnan, A。, Rosch, K. S。, Seymour,
K. E., Nebel, 中号. B., … Kiehl, K. (2017). The impact of T1 versus
EPI spatial normalization templates for fMRI data analyses.
人脑图谱, 38(11), 5331–5342. https://doi.org/10
.1002/hbm.23737, 考研: 28745021
志, J. G。, Dooling, 乙. C。, & 吉尔斯, F. H. (1977). Gyral development
of the human brain. Annals of Neurology, 1(1), 86–93. https://土井
.org/10.1002/ana.410010109, 考研: 560818
Dobbing, J。, & Sands, J. (1973). Quantitative growth and develop-
ment of human brain. Archives of Disease in Childhood, 48(10),
757–767. https://doi.org/10.1136/adc.48.10.757, 考研:
4796010
Doria, 五、, 贝克曼, C. F。, Arichi, T。, Merchant, N。, Groppo, M。,
Turkheimer, F. E., … Nunes, 右. G. (2010). Emergence of resting
state networks in the preterm human brain. 诉讼程序
美国国家科学院, 107(46), 20015–20020. https://
doi.org/10.1073/pnas.1007921107, 考研: 21041625
Esteban, 奥。, Markiewicz, C. J。, 布莱尔, 右. W., Moodie, C. A。, Isik,
A. 我。, Erramuzpe, A。, … Snyder, 中号. (2019). fMRIPrep: A robust
preprocessing pipeline for functional MRI. Nature Methods, 16(1),
111–116. https://doi.org/10.1038/s41592-018-0235-4, 考研:
30532080
Ferrazzi, G。, Murgasova, 中号. K., Arichi, T。, Malamateniou, C。, 狐狸,
中号. J。, Makropoulos, A。, … Aljabar, 磷. (2014). Resting State fMRI in
the moving fetus: A robust framework for motion, bias field and
spin history correction. 神经影像, 101, 555–568. https://土井
.org/10.1016/j.neuroimage.2014.06.074, 考研: 25008959
Fransson, P。, Skiöld, B., Horsch, S。, Nordell, A。, Blennow, M。,
Lagercrantz, H。, & Åden, U. (2007). Resting-state networks in
the infant brain. 美国国家科学院院刊
科学, 104(39), 15531–15536. https://doi.org/10.1073/pnas
.0704380104, 考研: 17878310
弗里斯顿, K. J。, 威廉姆斯, S。, 霍华德, R。, Frackowiak, 右. S。, & 车工, 右.
(1996). Movement-related effects in fMRI time-series. Magnetic
Resonance in Medicine, 35(3), 346–355. https://doi.org/10.1002
/mrm.1910350312, 考研: 8699946
Gholipour, A。, Rollins, C. K., Velasco-Annis, C。, Ouaalam, A。,
Akhondi-Asl, A。, Afacan, 奥。, … Yang, 乙. (2017). A normative
spatiotemporal MRI atlas of the fetal brain for automatic segmen-
tation and analysis of early brain growth. Scientific Reports, 7(1),
1–13. https://doi.org/10.1038/s41598-017-00525-w, 考研:
28352082
Glasser, 中号. F。, 索蒂罗普洛斯, S. N。, Wilson, J. A。, Coalson, 时间. S。,
菲舍尔, B., 安德森, J. L。, … Polimeni, J. 右. (2013). The minimal
preprocessing pipelines for the Human Connectome Project.
N e u ro I m a g e , 8 0 , 1 0 5 – 1 2 4 . h t t p s : / / d o i . o rg / 1 0 . 1 0 1 6 / j
.neuroimage.2013.04.127, 考研: 23668970
Griffanti, L。, Douaud, G。, Bijsterbosch, J。, Evangelisti, S。, Alfaro-
Almagro, F。, Glasser, 中号. F。, … Carone, D. (2017). Hand classifica-
tion of fMRI ICA noise components. 神经影像, 154, 188–205.
https://doi.org/10.1016/j.neuroimage.2016.12.036, 考研:
27989777
网络神经科学
719
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
A
_
0
0
2
5
4
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Evaluation of fetal fMRI preprocessing steps
Griffanti, L。, Salimi-Khorshidi, G。, 贝克曼, C. F。, 奥尔巴赫, 乙. J。,
Douaud, G。, Sexton, C. E., … Mackay, C. 乙. (2014). ICA-based
artefact removal and accelerated fMRI acquisition for improved
resting state network imaging. 神经影像, 95, 232–247. https://
doi.org/10.1016/j.neuroimage.2014.03.034 , 考研:
24657355
Himberg, J。, Hyvärinen, A。, & Esposito, F. (2004). Validating the
independent components of neuroimaging time series via cluster-
ing and visualization. 神经影像, 22(3), 1214–1222. https://土井
.org/10.1016/j.neuroimage.2004.03.027, 考研: 15219593
Jakab, A。, 施瓦茨, E., Kasprian, G。, Gruber, G. M。, Prayer, D .,
Schöpf, 五、, & Langs, G. (2014). Fetal functional imaging portrays
heterogeneous development of emerging human brain networks.
Frontiers in Human Neuroscience, 8, 852. https://doi.org/10
.3389/fnhum.2014.00852, 考研: 25374531
Kiviniemi, 五、, Starck, T。, 雷梅斯, J。, 长的, X。, Nikkinen, J。, Haapea,
M。, … Zang, 是. F. (2009). Functional segmentation of the brain
cortex using high model order group PICA. 人脑
测绘, 30(12), 3865–3886. https://doi.org/10.1002/ hbm
.20813, 考研: 19507160
Kundu, P。, Inati, S. J。, 埃文斯, J. W., Luh, W.-M., & Bandettini, 磷. A.
(2012). Differentiating BOLD and non-BOLD signals in fMRI time
series using multi-echo EPI. 神经影像, 60(3), 1759–1770.
https://doi.org/10.1016/j.neuroimage.2011.12.028, 考研:
22209809
Lowe, 中号. J。, & Sorenson, J. A. (1997). Spatially filtering functional
magnetic resonance imaging data. Magnetic Resonance in
药品, 37(5), 723–729. https://doi.org/10.1002/mrm
.1910370514, 考研: 9126946
Pruim, 右. H。, Mennes, M。, van Rooij, D ., Llera, A。, Buitelaar, J. K., &
贝克曼, C. F. (2015). ICA-AROMA: A robust ICA-based
strategy for removing motion artifacts from fMRI data. Neuro-
图像, 112, 267–277. https://doi.org/10.1016/j.neuroimage
.2015.02.064, 考研: 25770991
Rajagopalan, 五、, Deoni, S。, Panigrahy, A。, & Thomason, 中号. 乙.
(2021). Is fetal MRI ready for neuroimaging prime time? 一个
examination of progress and remaining areas for development.
Developmental Cognitive Neuroscience, 51, 100999. https://土井
.org/10.1016/j.dcn.2021.100999, 考研: 34391003
Reilly, K., & Gutierrez, J. (2021). The embryological development
of the cerebrovascular system. In Pediatric vascular neurosurgery
(PP. 1–5). 占婆, 瑞士: 施普林格. https://doi.org/10.1007
/978-3-030-74749-7_1
Rissanen, J. (1978). Modeling by shortest data description. Automa-
tica, 14(5), 465–471. https://doi.org/10.1016/0005-1098(78)
90005-5
Rummel, C。, Verma, 右. K., Schöpf, 五、, Hauf, M。, Abela, E., Zapata
Berruecos, J. F。, & Wiest, 右. (2013). Time course based artifact
identification for independent components of resting-state fMRI.
Frontiers in Human Neuroscience, 7, 214. https://doi.org/10
.3389/fnhum.2013.00214, 考研: 23734119
Rutherford, S。, Sturmfels, P。, Angstadt, M。, Hect, J。, Wiens, J。, van
den Heuvel, 中号. 我。, … Thomason, 中号. (2021). Automated brain
masking of fetal functional MRI with open data. Neuroinfor-
数学. https://doi.org/10.1007/s12021-021-09528-5, 考研:
34129169
Scheinost, D ., Onofrey, J. A。, 权, S. H。, 叉, S. N。, Sze, G。,
Ment, L. R。, & Papademetris, X. (2018, April 4–7). A fetal fMRI
specific motion correction algorithm using 2nd order edge
特征 [Paper presentation]. 2018 IEEE 15th International
Symposium on Biomedical Imaging. https://doi.org/10.1109
/ISBI.2018.8363807
Schöpf, 五、, Kasprian, G。, & Prayer, D. (2011). Functional imaging
in the fetus. Topics in Magnetic Resonance Imaging, 22(3),
113–118. https://doi.org/10.1097/ RMR.0b013e3182699283,
考研: 23558466
Serag, A。, Kyriakopoulou, 五、, Rutherford, 中号. A。, 爱德华兹, A. D .,
Hajnal, J. 五、, Aljabar, P。, … Rueckert, D. (2012). A multi-channel
4D probabilistic atlas of the developing brain: Application to
fetuses and neonates. Annals of the BMVA, 2012(3), 1–14.
Shattuck, D. W., & Leahy, 右. 中号. (2002). BrainSuite: An automated
cortical surface identification tool. Medical Image Analysis, 6(2),
129–142. https://doi.org/10.1016/S1361-8415(02)00054-3,
考研: 12045000
Smyser, C. D ., Inder, 时间. E., Shimony, J. S。, 爬坡道, J. E., Degnan, A. J。,
斯奈德, A. Z。, & Neil, J. J. (2010). Longitudinal analysis of neural
network development in preterm infants. 大脑皮层, 20(12),
2852–2862. https://doi.org/10.1093/cercor/ bhq035, 考研:
20237243
Thomason, 中号. 乙. (2020). Development of brain networks in utero:
Relevance for common neural disorders. Biological Psychiatry,
88(1), 40–50. https://doi.org/10.1016/j.biopsych.2020.02.007,
考研: 32305217
Thomason, 中号. E., Dassanayake, 中号. T。, 沉, S。, Katkuri, Y。, Alexis,
M。, 安德森, A. L。, … Romero, 右. (2013). Cross-hemispheric
functional connectivity in the human fetal brain. Science Trans-
lational Medicine, 5(173), 173ra124. https://doi.org/10.1126
/scitranslmed.3004978, 考研: 23427244
Thomason, 中号. E., Grove, L. E., Lozon Jr., 时间. A。, Vila, A. M。, 叶, Y。,
Nye, 中号. J。, … Yeo, L. (2015). Age-related increases in long-range
connectivity in fetal functional neural connectivity networks in
utero. Developmental Cognitive Neuroscience, 11, 96–104.
https://doi.org/10.1016/j.dcn.2014.09.001, 考研: 25284273
Thomason, 中号. E., Hect, J. L。, Rauh, V. A。, Trentacosta, C。,
Wheelock, 中号. D ., Eggebrecht, A. T。, … Burt, S. A. (2019). 预-
natal lead exposure impacts cross-hemispheric and long-range
connectivity in the human fetal brain. 神经影像, 191, 186–192.
https://doi.org/10.1016/j.neuroimage.2019.02.017, 考研:
30739062
Thomason, 中号. E., Hect, J. L。, Waller, R。, & Curtin, 磷. (2021A). 国际米兰-
active relations between maternal prenatal stress, fetal brain
连接性, and gestational age at delivery. Neuropsychophar-
macology, 46(10), 1839–1847. https://doi.org/10.1038/s41386
-021-01066-7, 考研: 34188185
Thomason, 中号. E., Palopoli, A. C。, Jariwala, 氮. N。, 韦尔坎, D. M。,
陈, A。, Adhikari, S。, … Trentacosta, C. J. (2021乙). Miswiring
大脑: Human prenatal Δ9-tetrahydrocannabinol use associ-
ated with altered fetal hippocampal brain network connectivity.
Developmental Cognitive Neuroscience, 51, 101000. https://土井
.org/10.1016/j.dcn.2021.101000, 考研: 34388638
Toga, A. W., & 汤普森, 磷. 中号. (2003). Mapping brain asymmetry.
自然评论神经科学, 4(1), 37–48. https://doi.org/10
.1038/nrn1009, 考研: 12511860
Turk, E., van den Heuvel, 中号. 我。, Benders, 中号. J。, de Heus, R。, Franx,
A。, 曼宁, J. H。, … van den Heuvel, 中号. 磷. (2019). Functional
connectome of the fetal brain. 神经科学杂志, 39(49),
网络神经科学
720
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
A
_
0
0
2
5
4
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
Evaluation of fetal fMRI preprocessing steps
9716–9724. https://doi.org/10.1523/JNEUROSCI.2891-18.2019,
考研: 31685648
van den Heuvel, 中号. 我。, & Thomason, 中号. 乙. (2016). Functional
connectivity of the human brain in utero. Trends in Cognitive
科学, 20(12), 931–939. https://doi.org/10.1016/j.tics.2016
.10.001, 考研: 27825537
van den Heuvel, 中号. 我。, Turk, E., 曼宁, J. H。, Hect, J。, Hernandez-
Andrade, E., Hassan, S. S。, … Thomason, 中号. 乙. (2018). Hubs in
the human fetal brain network. Developmental Cognitive Neuro-
科学, 30, 108–115. https://doi.org/10.1016/j.dcn.2018.02
.001, 考研: 29448128
Van Dijk, K. R。, 肥皂, 中号. R。, & 巴克纳, 右. L. (2012). 这
influence of head motion on intrinsic functional connectivity
MRI. 神经影像, 59(1), 431–438. https://doi.org/10.1016/j
.neuroimage.2011.07.044, 考研: 21810475
严, C.-G., 张, B., 凯莉, C。, Colcombe, S。, Craddock, 右. C。, 从
Martino, A。, … Milham, 中号. 磷. (2013). A comprehensive assessment
of regional variation in the impact of head micromovements on
functional connectomics. 神经影像, 76, 183–201. https://土井
.org/10.1016/j.neuroimage.2013.03.004, 考研: 23499792
严, C.-G., 王, X.-D., Zuo, X.-N., & Zang, Y.-F. (2016). DPABI:
Data processing and analysis for (resting-state) brain imaging.
神经信息学, 14(3), 339–351. https://doi.org/10.1007
/s12021-016-9299-4, 考研: 27075850
Zuo, X.-N., 凯莉, C。, Di Martino, A。, Mennes, M。, 马古利斯, D. S。,
Bangaru, S。, … Castellanos, F. X. (2010). Growing together and
growing apart: Regional and sex differences in the lifespan
developmental trajectories of functional homotopy. 杂志
神经科学, 30(45), 15034–15043. https://doi.org/10.1523
/JNEUROSCI.2612-10.2010, 考研: 21068309
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
3
7
0
2
2
0
3
6
0
3
2
n
e
n
_
A
_
0
0
2
5
4
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
网络神经科学
721