Masking Disrupts Reentrant Processing
in Human Visual Cortex
J. J. Fahrenfort1, H. S. Scholte1, 和V. A. F. Lamme1,2
抽象的
& In masking, a stimulus is rendered invisible through the
presentation of a second stimulus shortly after the first. 超过
这些年, authors have typically explained masking by postu-
lating some early disruption process. In these feedforward-
type explanations, the mask somehow ‘‘catches up’’ with the
target stimulus, disrupting its processing either through lat-
eral or interchannel inhibition. 然而, studies from recent
years indicate that visual perception—and most notably vi-
sual awareness itself—may depend strongly on cortico-cortical
feedback connections from higher to lower visual areas. 这
has led some researchers to propose that masking derives its
effectiveness from selectively interrupting these reentrant pro-
过程. In this experiment, we used electroencephalogram mea-
surements to determine what happens in the human visual
cortex during detection of a texture-defined square under
nonmasked (见过) and masked (unseen) 状况. Electro-
encephalogram derivatives that are typically associated with
reentrant processing turn out to be absent in the masked
状况. 而且, extrastriate visual areas are still acti-
vated early on by both seen and unseen stimuli, as shown
by scalp surface Laplacian current source-density maps. 这
conclusively shows that feedforward processing is preserved,
even when subject performance is at chance as determined
by objective measures. From these results, we conclude that
masking derives its effectiveness, at least partly, from disrupt-
ing reentrant processing, thereby interfering with the neural
mechanisms of figure–ground segmentation and visual aware-
ness itself. &
介绍
In backward masking, a target stimulus is rendered less
perceptible or even invisible through the presentation of
a second stimulus, the mask. By now, a respectable
number of studies have investigated the neural basis of
visual masking, both in its own right and as a corollary to
studying visual perception and awareness. In explaining
a variety of masking phenomena, some have empha-
sized low-level lateral inhibitory mechanisms (Macknik
& Livingstone, 1998; Francis, 1997), whereas others
have postulated target disruption at the precategorical
level through interchannel inhibition (Breitmeyer, Ro, &
Ogmen, 2004; Ogmen, Breitmeyer, & Melvin, 2003).
然而, early studies, as well as more recent imaging
studies on masked priming, show that processing of
masked targets may continue well beyond the precate-
gorical level (Naccache et al., 2005; Dehaene et al., 1998,
2001; Eimer & Schlaghecken, 1998; Greenwald, Klinger,
& 刘, 1989). These results suggest that masking does
not disrupt target processing at an early stage. This is in
line with various neurophysiological reports in which
early transient responses to undetected masked stimuli
1University of Amsterdam, 2Netherlands Institute for Neurosci-
恩斯, 阿姆斯特丹
were measured in high visual areas such as the infero-
temporal cortex, and even in visual neurons of the fron-
tal eye field (汤普森 & 沙尔, 1999; Kovacs, Vogels,
& Orban, 1995; Rolls & Tovee, 1994).
This has resulted in a number of researchers propos-
ing that masking disrupts reentrant cortical signals,
while largely leaving signals related to feedforward pro-
cessing intact (Ro, Breitmeyer, Burton, Singhal, & Lane,
2003; 瘫痪, Zipser, & Spekreijse, 2002; Di Lollo, Enns,
& Rensink, 2000; Bridgeman, 1980). These proposals
build on the notion that besides feedforward connec-
tions from low to higher visual areas, roughly equal
numbers of recurrent or reentrant connections exist
going back from higher to lower visual areas (Felleman
& Van Essen, 1991). Such feedback connections have
been suggested to play an integral role in a range of
流程, such as figure–ground segmentation, atten-
的, 意识, predictive coding, as well as perception
of visual detail (Spratling & 约翰逊, 2004; Hochstein &
Ahissar, 2002; Kastner & Ungerleider, 2000; 瘫痪 &
罗尔夫塞玛, 2000; 饶 & Ballard, 1999; Luck, Chelazzi,
Hillyard, & 德西莫内, 1997; 瘫痪, 1995). 它不是
unlikely that masking derives its effectiveness (至少在
部分) from disrupting reentrant signals, while leaving
feedforward signals intact, thereby interfering with
mechanisms thought to mediate visual awareness itself
(瘫痪 & 罗尔夫塞玛, 2000).
D 2007 麻省理工学院
认知神经科学杂志 19:9, PP. 1488–1497
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
t
n
我
C
/
A
e
–
r
p
t
d
我
C
1
我
9
e
9
–
1
p
4
d
8
F
8
/
1
1
9
9
/
3
6
9
5
/
2
1
8
4
8
哦
8
C
/
n
1
2
7
0
5
0
6
7
8
1
6
9
4
/
9
j
1
哦
4
C
8
n
8
.
p
2
d
0
0
乙
7
y
.
G
1
你
9
e
.
s
9
t
.
哦
1
n
4
0
8
7
8
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
.
.
.
F
哦
n
1
8
中号
A
y
2
0
2
1
然而, many masking studies do not show a rela-
tion between reentrant processing and masking. 这
may be caused by the fact that many such studies make
use of luminance or color contrast targets and metacon-
trast masks. In such metacontrast masking paradigms,
the mask tightly fits the preceding target, 和
common contour has a contrast of opposite polarity
(Becker & Anstis, 2004). It is quite likely that low-level
lateral
inhibitory mechanisms come into play when
processing these types of stimuli, at least at sufficiently
short stimulus onset asynchronies (Macknik & Martinez-
Conde, 2004; Macknik & Livingstone, 1998).
Contrast sensitivity is fundamental to vision and con-
trast is greatly enhanced by lateral inhibition at many
levels of visual processing, starting in the retina (Hartline,
1949). 所以, the use of (meta)contrast stimuli with-
out appropriately controlling for the influence of contrast
on the signal exaggerates the influence of low-level lateral
inhibition, while obscuring the influence of other higher-
level processes, such as reentrant processing. The stimuli
used in this experiment—in combination with a subtrac-
tion method (detailed below)—eliminate the influence of
luminance contrast on the signal, while at the same time
controlling for other low-level influences.
Aside from the contrast issue, visibility of masked tar-
gets is often assessed using an identification task. 如何-
曾经, a detection task is more fundamentally connected
to the notion of visual awareness, as many studies dem-
onstrate that residual identification in the absence of
awareness exists. This is evident not only from research
into the phenomenon of blindsight ( Weiskrantz, Barbur,
& Sahraie, 1995; Weiskrantz, Warrington, Sanders, &
马歇尔, 1974), but selective behavior in the absence
of awareness has also been shown in normal subjects
(博耶, 哈里森, & Ro, 2005; VanRullen & 科赫, 2003)
as well as in studies using masked priming (例如, Vorberg,
Mattler, Heinecke, 施密特, & Schwarzbach, 2003; Eimer
& Schlaghecken, 1998). Selectivity thus seems an inap-
propriate indicator of awareness. 所以, the pres-
ent experiment employs an objective detection task to
assess visibility.
We investigated the effect of pattern masking on
the detection of an orientation-defined square in a tex-
真实 (数字 1), while recording electroencephalogram
(EEG) in human subjects. Figure and no-figure trials were
either strongly masked by a pattern mask (masked)
leading to chance detection performance, or weakly
masked by a uniform gray screen leading to near-perfect
detection performance (unmasked). We used a subtrac-
tion procedure to control for various confounding influ-
ences on the EEG. By subtracting the figure trials from
the no-figure trials (数字 2), two goals were achieved:
(1) because both figure and no-figure trials were followed
by a mask in the masked condition, any direct influence
of the mask on the EEG was subtracted out. 相同
holds in the unmasked condition. (2) Because both figure
and no-figure trials were created using the same set of
数字 1. Schematic description of a trial. Subjects were required
to indicate whether a figure was present in a texture or not. Half
of the trials contained a figure. Half of the trials only contained a
homogenous texture. A trial could either be strongly masked (masked)
or weakly masked (unmasked). If subjects could not see anything due
to masking, they were instructed to guess (forced-choice detection).
oriented line elements,
figure and no-figure trials
contained exactly equal amounts of contrast and orienta-
的. 因此, any direct influence of low-level processes
such as contrast detection and orientation tuning is sub-
tracted out (Zipser, 瘫痪, & Schiller, 1996). This sub-
traction procedure would not work for metacontrast
target–mask combinations, as the net result of the sub-
traction of target-present and target-absent trials would
inherently leave a net result of contrast.
The spatio-temporal dynamics of the target-present
and target-absent subtraction signal was studied at an
extremely short SOA (16.6 毫秒) where targets were
either easily perceived or remained totally undetected as
determined by objective measures. We observed marked
activation in extrastriate regions in both the visible and
invisible condition as a result of feedforward process-
英. Later and more posterior activation resulting from
reentrant processing was observed only in the visible
状况. From the results, we infer that reentrant
processing is abolished as detection performance drops
to chance, even though feedforward processing is en-
tirely preserved.
方法
参加者
Twenty-one psychology students took part in the exper-
iment in partial fulfillment of first-year course require-
评论. All subjects (mean age 21.4, ±1.9) had normal or
corrected-to-normal vision and 71% were right-handed.
Each subject provided written informed consent before
继续行驶, 斯科尔特, and Lamme
1489
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
t
n
我
C
/
A
e
–
r
p
t
d
我
C
1
我
9
e
9
–
1
p
4
d
8
F
8
/
1
1
9
9
/
3
6
9
5
/
2
1
8
4
8
哦
8
C
/
n
1
2
7
0
5
0
6
7
8
1
6
9
4
/
9
j
1
哦
4
C
8
n
8
.
p
2
d
0
0
乙
7
y
.
G
1
你
9
e
.
s
9
t
.
哦
1
n
4
0
8
7
8
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
.
.
.
F
哦
n
1
8
中号
A
y
2
0
2
1
数字 2. (A) Examples of
stimuli and the schematic
orientations scheme.
Each gray value represents
an orientation; white is
isoluminant gray. (乙)
Representation of subtractions
leading to correlates of global
加工. Influences of
local line elements as well as
direct mask contributions are
canceled out by ensuring that
each orientation is present
equally often at each side of
the minus sign. Color scheme
is the same as in (A).
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
t
n
我
C
/
A
e
–
r
p
t
d
我
C
1
我
9
e
9
–
1
p
4
d
8
F
8
/
1
1
9
9
/
3
6
9
5
/
2
1
8
4
8
哦
8
C
/
n
1
2
7
0
5
0
6
7
8
1
6
9
4
/
9
j
1
哦
4
C
8
n
8
.
p
2
d
0
0
乙
7
y
.
G
1
你
9
e
.
s
9
t
.
哦
1
n
4
0
8
7
8
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
F
.
.
.
哦
n
1
8
中号
A
y
2
0
2
1
the start of the experiment. All procedures were ap-
proved by the ethical committee of the Psychology
Department of the University of Amsterdam. Two sub-
jects scored more than two standard deviations below
the mean in the unmasked task,
indicating difficulty
performing the task properly. These were excluded from
further analyses. Three subjects scored significantly
above chance in the masked condition as ascertained
by binomial tests ( p < .05), indicating an ability (how-
ever slight) to detect the masked figure at this masking
interval. These subjects were excluded because it cannot
be ruled out that they were still consciously processing
these stimuli. In total, five subjects were excluded.
Stimuli and Task
Without prior training, subjects had to detect the pres-
ence or absence of an orientation-defined square in a
texture pattern, which was either followed by a mask or
by an isoluminant gray screen (all stimuli had a lumi-
nance of 72.7 cd/m2). Each of the four resulting trial
types was presented an equal number of 300 times in
random order throughout the experiment. A trial started
with 300 msec fixation, followed by a centrally located
target stimulus for 16 msec, immediately followed by
either a mask or an isoluminant gray screen for 500 msec
(Figure 1). Target stimuli and mask consisted of a
texture of oriented line elements (approx. 0.078 thick
and 0.48 long in visual angle). All
line elements in a
target either had the same orientation (homogenous
trials) or contained a centrally located square with line
elements orthogonal to the background (figure trials).
The square subtended 2.738 of visual angle. In the
masked condition, targets were immediately followed
by a texture-defined pattern mask (4.548 visual angle)
using orientations not used in the preceding target. In
the unmasked condition, targets were followed by an
isoluminant gray screen (no texture). A total of four
1490
Journal of Cognitive Neuroscience
Volume 19, Number 9
orientations of line segments were used for creating fig-
ure, homogenous, and mask stimuli (22.58, 67.58, 112.58,
and 157.58; also see Figure 2) so that different orienta-
tions for target and mask could always be used within
any one trial. All orientations were completely counter-
balanced over trials so figure and homogenous trials did
not differ from each other with respect to local stimu-
lation (for a similar procedure, see Scholte, Witteveen,
Spekreijse, & Lamme, 2006; Caputo & Casco, 1999;
Lamme, Van Dijk, & Spekreijse, 1992). Stimuli were
created using Matlab (The MathWorks, Inc., Natick,
MA, USA). Stimuli were presented using Presentation
(Neurobehavioral Systems, Inc., Albany, CA, USA).
Subjects were instructed to fixate throughout the ex-
periment. The fixation dot would turn green 500 msec
after target offset to indicate that a response was re-
quired, after which the subject had a maximum of
1000 msec to respond before the experiment would
continue. On half of the trials, subjects were required
to press the left button to indicate square detection and
the right button for a homogenous trial. In the other
half of the trials, subjects had to switch so that a square
was associated with a left button press and a homo-
genous trial was associated with a right button press.
This response scheme was counterbalanced across sub-
jects, half of the subjects starting out square detection
with their right hand, and half of the subjects starting
out using their left hand. Switching hands posed no
difficulty as the high percentages correct on unmasked
trials show. Subjects were encouraged to guess if they
were unable to see whether a figure was presented,
effectively turning the task into a forced-choice task.
Measurements and Analyses
Event-related potentials (ERPs) were recorded from
the scalp using a BioSemi ActiveTwo 48-channel active
EEG system (BioSemi, Amsterdam, the Netherlands) at
256 Hz. Forty-eight scalp electrodes were measured (ref-
erenced to Fz), as well as two electrodes for horizontal
and two for vertical eye movements (each referenced to
its opposite counterpart). The data were filtered using a
high-pass filter of 1 Hz and a low-pass filter of 20 Hz, as
well as a 50-Hz notch filter. Before ocular correction,
automatic artifact rejection was applied by removing seg-
ments containing voltage steps of more than 50 AV, re-
moving any segments falling outside the (cid:1)200 to 200 AV
range, and removing those segments containing larger
than 300 AV differences within the segment. Ocular cor-
rection was applied on the basis of the horizontal and
vertical electrooculograms (Gratton, Coles, & Donchin,
1983). After ocular correction, artifact rejection was ap-
plied again by removing all segments outside the (cid:1)75
to 75 AV range. Linear local direct current detrending
was applied. This is a procedure to remove current drift
by subtracting a linear function from each segment.
Baseline correction was applied in the (cid:1)300 msec to
0 msec interval. All preprocessing steps were conducted
using Brain Vision Analyzer (Brain Products, Munich,
Germany).
All analyses were conducted on difference waves.
These were obtained by subtracting average no-figure
trials from figure trials, separately for masked and un-
masked trials. Spline Laplacian distribution maps were
calculated by interpolating difference waves using spher-
ical splines and approximating current source densities
(Perrin, Pernier, Bertrand, & Echallier, 1989). The result-
ing maps are spatial second-order derivatives of the scalp
potentials lending greater weight to local contributions of
cortical generators, filtering out deep sources, as well as
being reference free (Nunez & Srinivasan, 2006).
In order to test at which time points the differences
between figure and no-figure trials for the masked and
unmasked condition significantly deflect from chance, as
well as test the difference between masked and un-
masked trials, a random effects analysis was performed
by employing a paired two-tailed t test at each time
point, treating the average of each subject at that time
point as an observation. Correction for multiple com-
parisons with respect to the number of time points
being tested was done by limiting the false discovery
rate (FDR), a method in which the p value, at which
significance is evaluated, is corrected for the number of
tests being performed (Benjamini & Hochberg, 1995).
The method fixes the expected proportion of
false
positives (the FDR) in contrast to Bonferroni correction,
which controls the chance of any false positive among
all tests. The FDR is an often used method in a wide
variety of scientific fields (for an explanation of how it
is used in the field of neuroimaging, see Genovese,
Lazar, & Nichols, 2002). Statistical analyses and visuali-
zation of the timecourses were done using Matlab (The
MathWorks, Inc.).
RESULTS
The difference between figure and no-figure trials was
easily detected when no mask was present, as expressed
in an average detection rate of 96.5% (±2.5). For the
masked trials, the detection rate was at chance at 51.4%
(±4.1). Detection rates are expressed as the average
perfect observer score, which is an objective and subject
bias-free measure of a subjects’ ability to detect a signal
(Wickens, 2002). None of the included subjects scored
significantly above chance in the masked condition
(binomial tests, significance evaluated at .05).
Visual evoked potential (VEP) averages were calculat-
ed from the EEG data for each condition. Difference
waves of these averages were computed by subtracting
no-figure from figure trials in order to isolate activity
related to processing of global organization of the target
stimulus and discard processing related to processing of
line elements (Figure 2; see also Scholte et al.,
local
Fahrenfort, Scholte, and Lamme
1491
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
a
e
-
r
p
t
d
i
c
1
l
9
e
9
-
1
p
4
d
8
f
8
/
1
1
9
9
/
3
6
9
5
/
2
1
8
4
8
o
8
c
/
n
1
2
7
0
5
0
6
7
8
1
6
9
4
/
9
j
1
o
4
c
8
n
8
.
p
2
d
0
0
b
7
y
.
g
1
u
9
e
.
s
9
t
.
o
1
n
4
0
8
7
8
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
2006; Caputo & Casco, 1999; Lamme et al., 1992). This
was done separately for the masked and unmasked con-
dition, as to subtract out direct contributions of the
mask to the VEPs and enable comparison of masked
and unmasked trials. All VEP difference waves were
converted to spline Laplacian (see Methods) to be able
to better isolate local generators of any observed ef-
fects and filter out deep sources (Nunez & Srinivasan,
2006).
A split-half procedure of odd and even trials was used
to evaluate the data (Figure 3). Even trials were used for
a visualization of instructive time segments using spline
Laplacian maps. These were used to select relevant elec-
trodes for pooling. Odd trials were used to evaluate sig-
nificant deflections of the average current source density
at these electrodes over time. Using odd trials for the
spatial map and even trials for the timecourse values
ensures complete independence of data used for inspec-
tion and data used for evaluation of significance. This
gives an enormous boost to the reliability of the effects
that were found, while precluding coincidental signifi-
cance of sources that were identified visually. It is im-
portant to note that, although for brevity not all time
points in the 0–305 msec range have been visualized
using scalp distribution maps in Figure 3, electrode
pooling was done on the basis of all visually identifiable
sources in this domain, whereas none were opportunis-
tically left out. Pooling the data to a limited number of
electrodes in this way circumvents the multiple compar-
isons problem of having to evaluate large numbers of
electrodes. Pooling relevant electrodes furthermore in-
creases the signal-to-noise ratio.
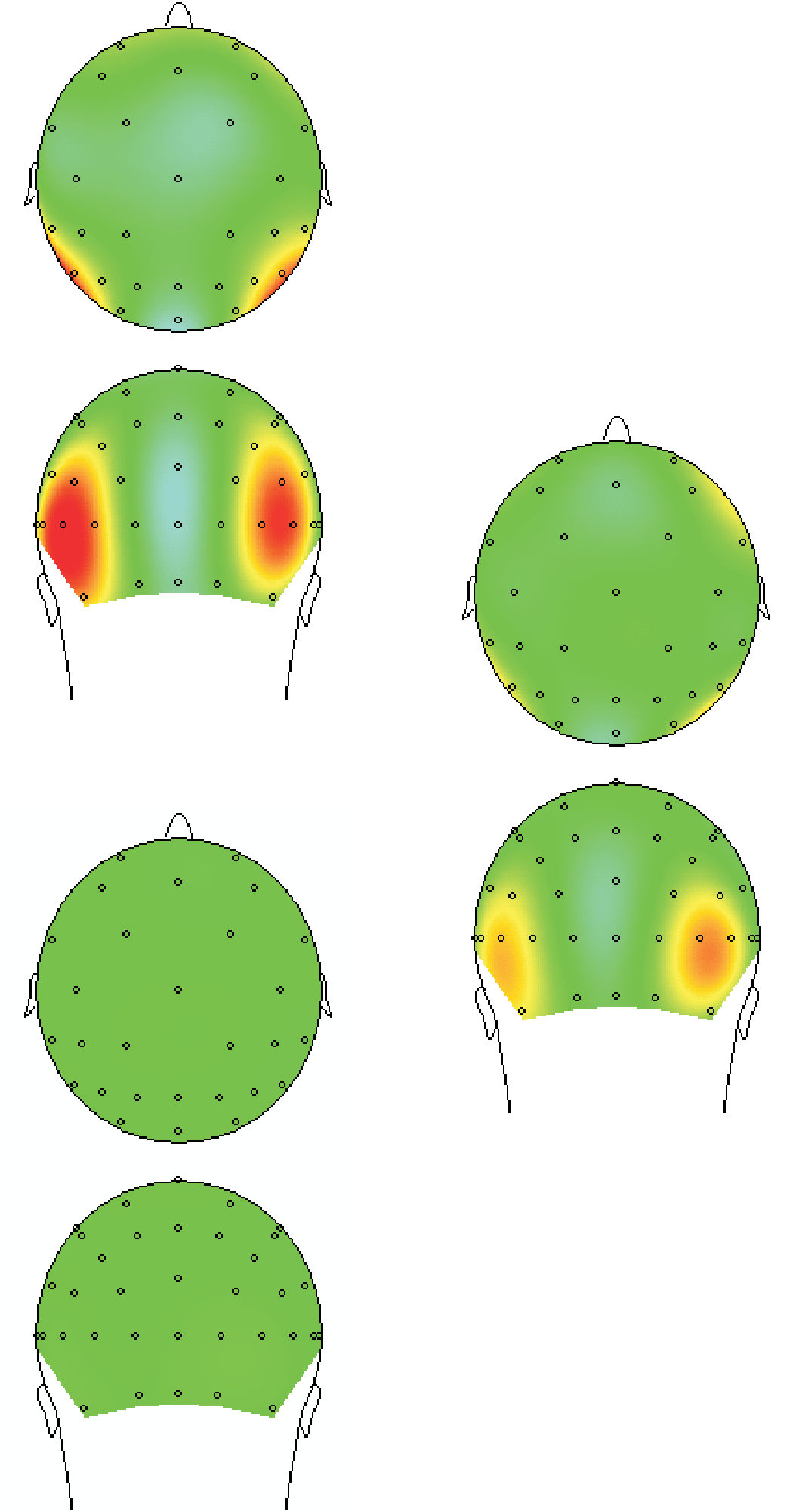
Figure 3A shows the course of neural processing in the
case of unmasked trials. Three stages can be distinguished:
1. A pre-110 msec stage with bilateral anterior occipito-
temporal neural generators consistent with feedforward
processing toward extrastriate visual areas and beyond
(Foxe & Simpson, 2002).
2. A post-110 msec stage with a clear posterior occi-
pital generator consistent with reentrant processing to-
ward early visual areas, possibly V1.
3. A strong recurring bilateral generator in the 200–
300 msec range, plausibly extrastriate and beyond.
All generators show significant deflections as ascer-
tained by paired t tests between figure and no-figure
trials. These were performed on each time point in the
time segment, subsequently corrected for multiple com-
parisons by fixing the FDR at .05 (see Methods).
Figure 3B shows the same three stages for the masked
trials. Stage 1 again shows a significant bilateral extrastri-
ate neural generator. Stages 2 and 3 are largely absent and
show no significant deflections. A direct comparison of
the masked and unmasked difference waves was also done
(see Figure 4). In accordance with the other results, these
showed significant differences in Stage 2 and Stage 3, but
not in Stage 1. Taken together, these results are consis-
tent with an interpretation where masking results in an
intact initial feedforward activation of extrastriate areas,
followed by a disruption of reentrant processing, possibly
affecting both striate and extrastriate visual cortex.
Because the resolution of ERPs is rather limited, some
may object to the idea that reentrant processing can be
pinpointed using ERPs. Yet, in our view, only a relatively
simple claim needs to be substantiated: (1) there is early
activation found anterior on the scalp; (2) there is
distinct later activation posterior on the scalp. In com-
bination with current source densities, and provided that
the generators are clearly distinct, this simple anterior/
posterior claim is, in our view, sufficient to claim reen-
trant processing.
DISCUSSION
The most salient aspect of these results is that even
when subjects are unable to perceive anything by objec-
tive measures, widespread bilateral occipito-temporal
activation still takes place, showing a clear ability of
the brain to pick up differences that the subject cannot
express. This early activation is almost indistinguishable
from early activity in the visible situation and seems
insufficient to generate visual awareness. Some addition-
al process appears necessary in order to achieve con-
scious perception and detection. The earliest difference
between the visible and nonvisible condition arises in
the >110-msec timeframe, and occurs more posterior
than the early bilateral activation. This is most consistent
with an interpretation where masking disrupts reentrant
processing between high and low visual areas, 尽管
leaving feedforward activation intact.
It is unclear how masking accounts resting solely
on lateral inhibition could explain these data. The stron-
gest evidence for the influence of lateral
inhibition
in masking comes from studies employing first-order
metacontrast stimuli (例如, Macknik & Livingstone, 1998).
First-order (luminance-defined) stimuli are obvious can-
didates for spatio-temporal lateral inhibitory influences,
as one can see how a metacontrasting stimulus laterally
inhibits a first-order contrast stimulus of opposite polar-
性. 然而, the present study makes use of second-
order target stimuli and pattern masking. As target and
mask do not bear a metacontrasting relation to each
inhibitory influences are less
其他,
likely. 而且, any remains of low-level inhibitory in-
fluences that may still exist are subtracted out using the
figure minus no-figure subtraction.
low-level
lateral
然而, one could still postulate that high-level lat-
eral inhibitory mechanisms operate between complex
representations—as suggested by Macknik and Martinez-
Conde (2004). If this were the case, one would pre-
dict the pre-110 msec activation to be disrupted in the
masked scenario as a result of these inhibitory influ-
1492
认知神经科学杂志
体积 19, 数字 9
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
t
n
我
C
/
A
e
–
r
p
t
d
我
C
1
我
9
e
9
–
1
p
4
d
8
F
8
/
1
1
9
9
/
3
6
9
5
/
2
1
8
4
8
哦
8
C
/
n
1
2
7
0
5
0
6
7
8
1
6
9
4
/
9
j
1
哦
4
C
8
n
8
.
p
2
d
0
0
乙
7
y
.
G
1
你
9
e
.
s
9
t
.
哦
1
n
4
0
8
7
8
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
F
.
.
.
哦
n
1
8
中号
A
y
2
0
2
1
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
t
n
我
C
/
A
e
–
r
p
t
d
我
C
1
我
9
e
9
–
1
p
4
d
8
F
8
/
1
1
9
9
/
3
6
9
5
/
2
1
8
4
8
哦
8
C
/
n
1
2
7
0
5
0
6
7
8
1
6
9
4
/
9
j
1
哦
4
C
8
n
8
.
p
2
d
0
0
乙
7
y
.
G
1
你
9
e
.
s
9
t
.
哦
1
n
4
0
8
7
8
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
F
.
.
.
.
哦
n
1
8
中号
A
y
2
0
2
1
数字 3. 结果. Spline Laplacian maps over three time segments: (1) 78–109 msec, (2) 109–141 msec, 和 (3) 180–305 msec, and event-related
averages of relevant electrodes (pooled occipitally and occipito-temporally). Even trials were used for data inspection (the maps) to determine
sensible electrode poolings. Odd trials were used for the averages and evaluation of statistical significance. (A) Unmasked condition; (乙) Masked
状况. Both masked and unmasked show strong significant temporal bilateral activation reflecting feedforward activation in Stage 1. 更多的
posterior occipital activation reflecting reentrant processing can only be seen in the unmasked condition in Stage 2, but is abolished by masking.
继续行驶, 斯科尔特, and Lamme
1493
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
t
n
我
C
/
A
e
–
r
p
t
d
我
C
1
我
9
e
9
–
1
p
4
d
8
F
8
/
1
1
9
9
/
3
6
9
5
/
2
1
8
4
8
哦
8
C
/
n
1
2
7
0
5
0
6
7
8
1
6
9
4
/
9
j
1
哦
4
C
8
n
8
.
p
2
d
0
0
乙
7
y
.
G
1
你
9
e
.
s
9
t
.
哦
1
n
4
0
8
7
8
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
F
.
.
.
.
.
哦
n
1
8
中号
A
y
2
0
2
1
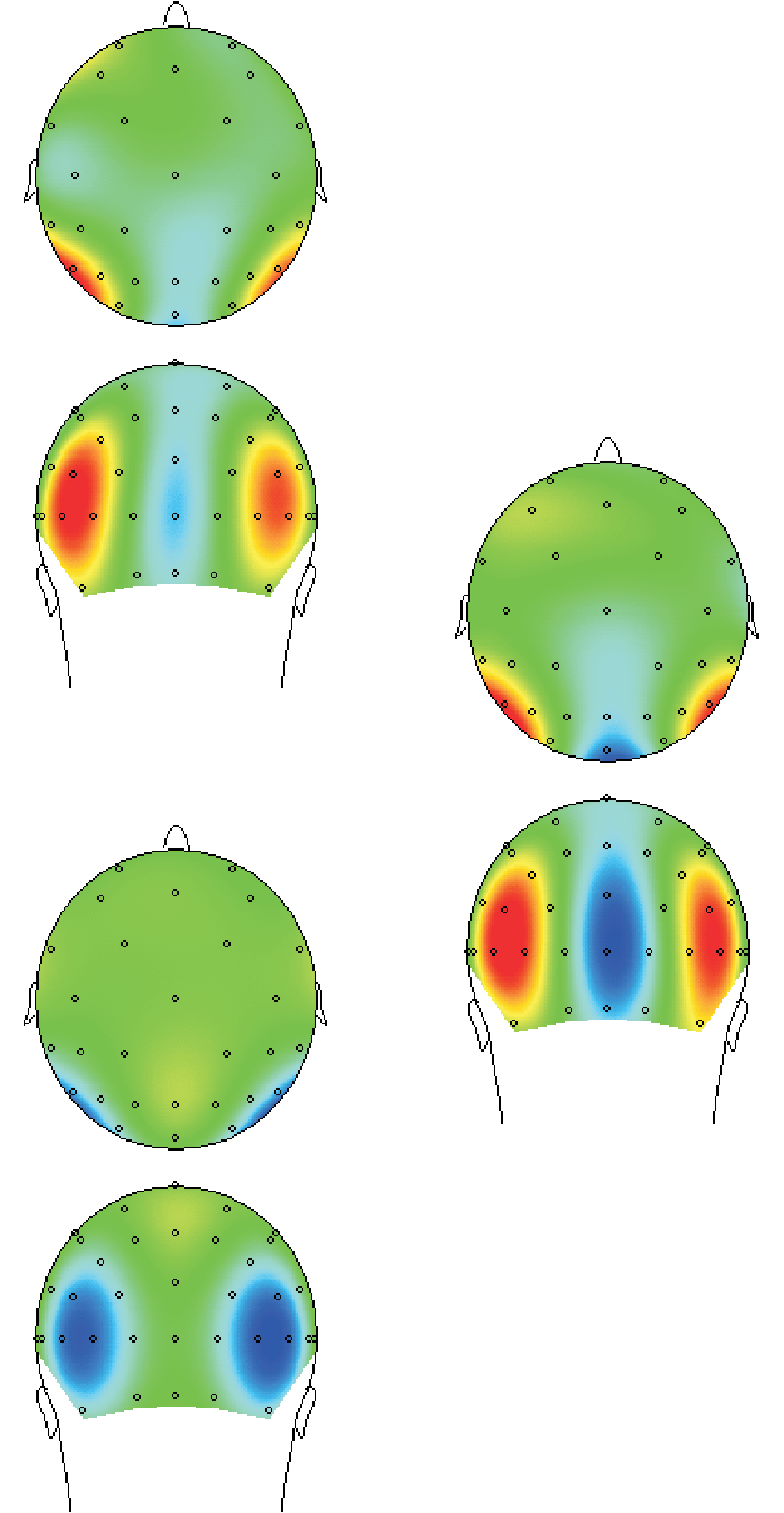
数字 4. A difference wave of the difference waves outlined in Figure 3 was calculated so as to directly test at which intervals the masked
and the unmasked figures differ from each other. This figure clearly shows that masked and unmasked trials do not significantly differ from each
other in the early 78–109 msec occipito-temporal stage related to feedforward processing (1), but do differ significantly from each other in the
109–141 msec occipital stage (2), which is plausibly related to reentrant processing as well as the later 180–305 msec occipito-temporal stage (3).
恩塞斯. 然而, the locus of interruption in this exper-
iment is later in time and more posterior. This is not to
say that lateral inhibitory influences do not play a role in
many types of masking, only that the view of ubiquitous
lateral inhibition as an explanation for our results is not
tenable.
Interchannel inhibition accounts of masking presume
the existence of two channels in visual processing, a fast
and a slow one. Masking is presumably caused by the
mask in a fast-acting channel inhibited by the processing
of the target in a slow-acting channel. Such accounts
cannot be ruled out entirely by these data, 像这样
theories do not specify at which level inhibition takes
地方. It could be argued that the feedforward activity
found in this study is caused by target activity in a fast-
acting channel, and that the posterior >110-msec ac-
活力, which we call reentrant activity, is caused by a
slow-acting channel. In the masked scenario, this activity
would be inhibited by the mask through subsequent
activation of the fast-acting channel (这会有
to be projecting downward to inhibit the slow-acting
渠道). 然而, aside from the fact that the poste-
rior activity we observed is not very ‘‘sustained’’ as such
models propose, there is little neurophysiological evi-
dence that interchannel inhibition actually exists (还
see Enns & Di Lollo, 2000). This makes an explanation
built on proven notions of reentrant processing much
more parsimonious in this context.
Aside from the present study, a number of other
studies have inferred the influence of masking on re-
entrant processing in human subjects, most notably stud-
ies employing object substitution masking (Weidner,
Shah, & Fink, 2006; Woodman & Luck, 2003; Di Lollo
等人。, 2000). In object substitution masking (also referred
to as four-dot masking), a four-dot mask is presented
surrounding a nonattended target. The subsequent dis-
crimination of this target is unimpaired when target and
mask coterminate, but strongly impaired when the dots
remain visible after target offset. This is explained by
postulating that the four dots (after offset of the target)
create a mismatch between the cortical representation
of the four dots in early visual areas and the cortical
representation of the target in higher areas. Although we
believe that the results of these studies have significant
implications for the influence of reentrant processing on
洞察力, they differ from our results in important ways.
Four-dot masking depends on attention being direct-
ed elsewhere and increases with increasing set size
(Enns, 2004; Di Lollo et al., 2000). In our pattern
masking study, attention is fully directed on the target,
and set size is just one. 而且, uninterrupted pro-
cessing of masked targets using object substitution con-
tinues well into the 200–300 msec domain ( Woodman
& Luck, 2003), whereas our results show that processing
is disrupted at 110 毫秒. 的确, a recent fMRI study
comparing pattern and object substitution masking
shows that differences between the two can be found
in the left middle frontal gyrus, in the precentral gyrus of
the right hemisphere, and in the medial superior frontal
gyrus (Weidner et al., 2006), whereas our study shows
strictly occipital disruption due to pattern masking.
What could account for these differences? What hap-
pens during object substitution masking seems to be
more akin to what happens during other attentional
操纵, such as the attentional blink. Many at-
tentional blink studies have shown that processing of
unseen target stimuli goes almost completely uninter-
rupted up to 300 msec and that differences between
1494
认知神经科学杂志
体积 19, 数字 9
reported and unreported targets are to be found in
frontal and parietal cortices (Kranczioch, Debener,
Schwarzbach, Goebel, & 恩格尔, 2005; 军士, 巴耶莱特,
& 德阿内, 2005; Luck, 沃格尔, & 夏皮罗, 1996). 这
is reminiscent of the response profile found in object
substitution masking. As other authors have noted, 这
failure of subjects to report stimuli in such instances may
reflect a loss of information at a postperceptual stage,
where information is replaced before it can be trans-
ferred to working memory and accessible awareness
(例如, Woodman & Luck, 2003; Luck et al., 1996).
It may still be reentrant processing that is interrupted,
but at a later stage and between visual and higher brain
地区 (possibly frontal and parietal), whereas early reen-
trant processing within visual areas goes uninterrupted.
Recent models of consciousness proposed by Dehaene,
尚古克斯, 纳克卡什, 萨克尔, and Sergent (2006) 和
瘫痪 (2003, 2006) outline similar ideas in more detail.
The notion of several reentrant or recurrent loops at
multiple levels is highly likely given the temporal dy-
namics of cortical processing (例如, Foxe & 辛普森,
2002). Because of the +110-msec timing and location
observed in this experiment, we conclude that reentrant
processing was disrupted at perceptual levels early on,
comparable to what has been shown in other studies
using different stimuli (Haynes, Driver, & Rees, 2005;
Breitmeyer et al., 2004; Ro et al., 2003).
The function of reentrant processing within occipital
cortex is a somewhat open question, although strong evi-
dence exists that it is of importance for figure–ground
segmentation (例如, 瘫痪, Rodriguez-Rodriguez, &
Spekreijse, 1999; Hupe et al., 1998; 瘫痪, 1995). 在
such studies, it is typically shown that a neuron in V1
starts responding differentially after about 100 msec when
its classical receptive field (病例报告表) is located on the center
of a figure compared to when it is on a background,
despite local stimulation being exactly equal (比照. 数字 2).
These extra-CRF effects are usually referred to as contex-
tual modulation, as they are modulated by context infor-
mation outside their CRF. The presence and timing
(>100 msec) of contextual modulation strongly suggests
that reentrant neural processing takes place going back
from higher areas containing larger receptive fields to
lower areas such as V1. 还, when extrastriate regions are
lesioned, contextual modulation disappears, leaving re-
ceptive field tuning properties intact and conclusively
showing the role of reentrant processing in contextual
modulation (瘫痪, Super, & Spekreijse, 1998).
A study in which the effect of target–mask SOA on
contextual modulation in macaque V1 was investigated
showed a strong relation between SOA, contextual mod-
计算, and the monkey’s ability to detect a texture-
defined figure (Lamme et al., 2002). 而且, 任何
influence of SOA on the CRF tuning properties of these
neurons was almost completely absent, showing that
feedforward activation was largely preserved. 然而,
它是
because measurements were done only in V1,
unclear to what extent feedforward activation would
continue to higher areas and to what extent the same
would hold for humans.
The present study clearly demonstrates that in humans,
undetected and strongly masked texture stimuli are
processed far beyond the striate cortex (见图 3),
showing that feedforward processing is preserved, 在哪里-
as reentrant processing is selectively interrupted. A neu-
ral network model of figure–ground segmentation by
罗尔夫塞玛, 瘫痪, Spekreijse, and Bosch (2002),
在
which texture segregation in macaques is modeled in
terms of feedforward and reentrant processing, serves
extremely well in explaining the spatio-temporal profile
of such results.
Related models propose that reentrant processing is
more intimately related to visual awareness (例如, Ro
等人。, 2003; 瘫痪, 2001). These views on reentrant
processing are not necessarily contradicting, as visual
awareness might simply be the way surface segregation
and related processes express themselves phenomeno-
logically. The view that reentrant activity in the visual
cortex correlates with visual awareness is now supported
by converging evidence from monkey physiology (例如,
瘫痪, Super, Landman, 罗尔夫塞玛, & Spekreijse,
2000), EEG (the present study), transcranial magnetic
stimulation (Pascual-Leone & Walsh, 2001), and fMRI
(例如, Haynes et al., 2005).
结论
We argue and found evidence for at least three stages
in early visual processing: (1) a completely unconscious
pre-110 msec feedforward stage, which is probably in-
volved in boundary detection but goes uninterrupted
by masking; (2) a second stage, which operates in the
110–140 msec range reflecting reentrant processing in
视觉皮层, plausibly encapsulating processes such as
figure–ground segmentation and phenomenal aware-
内斯; 和 (3) a stage in the 200–300 msec range, 哪个
is contingent upon earlier stages. 具体来说, the sec-
ond stage was interrupted by backward masking, 骗局-
firming that pre-110 msec activation proceeds outside
意识, whereas more posterior >110 msec activity
是 (至少) a necessary condition for awareness to arise.
最后, we reject explanations of backward masking as a
unitary phenomenon, in agreement with other authors
such as Bachmann, Luiga, and Poder (2005). Any expla-
nation of the masking phenomenon should carefully
take into account physical stimulus characteristics and
how these are processed by the various components of
the visual system.
致谢
We thank IJsbrand Attema for his help in collecting the data for
this experiment, as well as the anonymous reviewers for their
helpful comments.
继续行驶, 斯科尔特, and Lamme
1495
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
t
n
我
C
/
A
e
–
r
p
t
d
我
C
1
我
9
e
9
–
1
p
4
d
8
F
8
/
1
1
9
9
/
3
6
9
5
/
2
1
8
4
8
哦
8
C
/
n
1
2
7
0
5
0
6
7
8
1
6
9
4
/
9
j
1
哦
4
C
8
n
8
.
p
2
d
0
0
乙
7
y
.
G
1
你
9
e
.
s
9
t
.
哦
1
n
4
0
8
7
8
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
F
.
.
.
哦
n
1
8
中号
A
y
2
0
2
1
Reprint requests should be sent to J. J. 继续行驶, Department
心理学系, University of Amsterdam, Roetersstraat 15, 1018
WB Amsterdam, 荷兰人, or via e-mail: j.j.fahrenfort@
uva.nl.
Francis, G. (1997). Cortical dynamics of lateral inhibition:
Metacontrast masking. 心理评论, 104,
572–594.
Genovese, C. R。, Lazar, 氮. A。, & Nichols, 时间. (2002). Thresholding
参考
Bachmann, T。, Luiga, 我。, & Poder, 乙. (2005). Variations in
backward masking with different masking stimuli: 二.
The effects of spatially quantised masks in the light
of local contour interaction, interchannel inhibition,
perceptual retouch, and substitution theories. 洞察力,
34, 139–153.
Becker, 中号. W., & Anstis, S. (2004). Metacontrast masking
is specific to luminance polarity. 视觉研究, 44,
2537–2543.
Benjamini, Y。, & Hochberg, 是. (1995). Controlling the false
discovery rate—A practical and powerful approach to
multiple testing. Journal of the Royal Statistical Society,
Series B, Methodological, 57, 289–300.
博耶, J. L。, 哈里森, S。, & Ro, 时间. (2005). Unconscious
processing of orientation and color without primary
视觉皮层. 美国国家科学院院刊
科学, 美国。, 102, 16875–16879.
Breitmeyer, 乙. G。, Ro, T。, & Ogmen, H. (2004). A comparison
of masking by visual and transcranial magnetic stimulation:
Implications for the study of conscious and unconscious
visual processing. Consciousness and Cognition, 13,
829–843.
Bridgeman, 乙. (1980). Temporal response characteristics of
cells in monkey striate cortex measured with metacontrast
masking and brightness discrimination. 脑研究,
196, 347–364.
Caputo, G。, & Casco, C. (1999). A visual evoked potential
correlate of global figure–ground segmentation. Vision
研究, 39, 1597–1610.
德阿内, S。, 尚古克斯, J. P。, 纳克卡什, L。, 萨克尔, J。,
& 军士, C. (2006). 有意识的, 前意识, 和
subliminal processing: A testable taxonomy. Trends
in Cognitive Sciences, 10, 204–211.
德阿内, S。, 纳克卡什, L。, 科恩, L。, Le Bihan, D ., 曼金,
J. F。, 波琳娜, J. B., 等人. (2001). Cerebral mechanisms of
word masking and unconscious repetition priming. 自然
神经科学, 4, 752–758.
德阿内, S。, 纳克卡什, L。, Le Clec’H, G。, Koechlin, E.,
Mueller, M。, Dehaene Lambertz, G。, 等人. (1998). Imaging
unconscious semantic priming. 自然, 395, 597–600.
Di Lollo, 五、, Enns, J. T。, & Rensink, 右. A. (2000). Competition
for consciousness among visual events: The psychophysics
of reentrant visual processes. 实验杂志
心理学: General, 129, 481–507.
Eimer, M。, & Schlaghecken, F. (1998). Effects of masked stimuli
on motor activation: Behavioral and electrophysiological
证据. 实验心理学杂志: 人类
Perception and Performance, 24, 1737–1747.
Enns, J. 时间. (2004). Object substitution and its relation to
other forms of visual masking. 视觉研究, 44,
1321–1331.
Enns, J. T。, & Di Lollo, V. (2000). What’s new in visual
masking? 认知科学的趋势, 4, 345–352.
Felleman, D. J。, & Van Essen, D. C. (1991). Distributed
hierarchical processing in the primate cerebral cortex.
大脑皮层, 1, 1–47.
Foxe, J. J。, & 辛普森, G. V. (2002). Flow of activation from
V1 to frontal cortex in humans—A framework for defining
‘‘early’’ visual processing. Experimental Brain Research,
142, 139–150.
of statistical maps in functional neuroimaging using the
false discovery rate. Neuroimage, 15, 870–878.
Gratton, G。, Coles, 中号. G。, & Donchin, 乙. (1983). A new
method for off-line removal of ocular artifact.
Electroencephalography and Clinical Neurophysiology,
55, 468–484.
Greenwald, A. G。, Klinger, 中号. R。, & 刘, 时间. J. (1989).
Unconscious processing of dichoptically masked
字. 记忆 & 认识, 17, 35–47.
Hartline, H. K. (1949). Inhibition of activity of visual
receptors by illuminating nearby retinal areas in the
Limulus eye. Federation Proceedings, 8, 69.
Haynes, J. D ., Driver, J。, & Rees, G. (2005). Visibility reflects
dynamic changes of effective connectivity between V1
and fusiform cortex. 神经元, 46, 811–821.
Hochstein, S。, & Ahissar, 中号. (2002). View from the top:
Hierarchies and reverse hierarchies in the visual system.
神经元, 36, 791–804.
Hupe, J. M。, James, A. C。, 佩恩, 乙. R。, Lomber, S. G。,
Girard, P。, & Bullier, J. (1998). Cortical feedback improves
discrimination between figure and background by V1,
V2 and V3 neurons. 自然, 394, 784–787.
Kastner, S。, & Ungerleider, L. G. (2000). Mechanisms of
visual attention in the human cortex. Annual Review
of Neuroscience, 23, 315–341.
Kovacs, G。, Vogels, R。, & Orban, G. A. (1995). Cortical
correlate of pattern backward-masking. 会议记录
the National Academy of Sciences, 美国。, 92, 5587–5591.
Kranczioch, C。, Debener, S。, Schwarzbach, J。, Goebel, R。,
& 恩格尔, A. K. (2005). Neural correlates of conscious
perception in the attentional blink. Neuroimage, 24,
704–714.
瘫痪, V. A. F. (1995). The neurophysiology of figure
ground segregation in primary visual-cortex. 杂志
of Neuroscience, 15, 1605–1615.
瘫痪, V. A. F. (2001). Blindsight: The role of feedforward and
feedback corticocortical connections. 心理学学报,
107, 209–228.
瘫痪, V. A. F. (2003). Why visual attention and awareness
are different. 认知科学的趋势, 7, 12–18.
瘫痪, V. A. F. (2006). Towards a true neural stance on
consciousness. 认知科学的趋势, 10, 494–501.
瘫痪, V. A. F。, Rodriguez-Rodriguez, 五、, & Spekreijse, H.
(1999). Separate processing dynamics for texture elements,
boundaries and surfaces in primary visual cortex of the
macaque monkey. 大脑皮层, 9, 406–413.
瘫痪, V. A. F。, & 罗尔夫塞玛, 磷. 右. (2000). The distinct
modes of vision offered by feedforward and recurrent
加工. Trends in Neurosciences, 23, 571–579.
瘫痪, V. A. F。, Super, H。, Landman, R。, 罗尔夫塞玛, 磷. R。,
& Spekreijse, H. (2000). The role of primary visual
cortex (V1) in visual awareness. 视觉研究, 40,
1507–1521.
瘫痪, V. A. F。, Super, H。, & Spekreijse, H. (1998).
Feedforward, 水平的, and feedback processing in
the visual cortex. 神经生物学的当前观点, 8,
529–535.
瘫痪, V. A. F。, Van Dijk, 乙. W., & Spekreijse, H. (1992).
Texture segregation is processed by primary visual cortex
in man and monkey. Evidence from VEP experiments.
视觉研究, 32, 797–807.
瘫痪, V. A. F。, Zipser, K., & Spekreijse, H. (2002).
Masking interrupts figure–ground signals in V1. 杂志
of Cognitive Neuroscience, 14, 1044–1053.
1496
认知神经科学杂志
体积 19, 数字 9
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
t
n
我
C
/
A
e
–
r
p
t
d
我
C
1
我
9
e
9
–
1
p
4
d
8
F
8
/
1
1
9
9
/
3
6
9
5
/
2
1
8
4
8
哦
8
C
/
n
1
2
7
0
5
0
6
7
8
1
6
9
4
/
9
j
1
哦
4
C
8
n
8
.
p
2
d
0
0
乙
7
y
.
G
1
你
9
e
.
s
9
t
.
哦
1
n
4
0
8
7
8
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
.
.
.
F
哦
n
1
8
中号
A
y
2
0
2
1
Luck, S. J。, Chelazzi, L。, Hillyard, S. A。, & 德西莫内, 右.
Rolls, 乙. T。, & Tovee, 中号. J. (1994). Processing speed in the
(1997). Neural mechanisms of spatial selective attention
in areas V1, V2, and V4 of macaque visual cortex. 杂志
of Neurophysiology, 77, 24–42.
Luck, S. J。, 沃格尔, 乙. K., & 夏皮罗, K. L. (1996). Word
meanings can be accessed but not reported during
the attentional blink. 自然, 383, 616–618.
Macknik, S. L。, & Livingstone, 中号. S. (1998). Neuronal correlates
of visibility and invisibility in the primate visual system.
自然神经科学, 1, 144–149.
Macknik, S. L。, & Martinez-Conde, S. (2004). Dichoptic visual
masking reveals that early binocular neurons exhibit weak
interocular suppression: Implications for binocular vision
and visual awareness. 认知神经科学杂志,
16, 1049–1059.
纳克卡什, L。, 盖拉德, R。, 亚当, C。, 哈斯本, D .,
克列孟梭, S。, 鲍拉克, M。, 等人. (2005). A direct
intracranial record of emotions evoked by subliminal
字. 美国国家科学院院刊,
美国。, 102, 7713–7717.
Nunez, P。, & 斯里尼瓦桑, 右. (2006). Electric fields of the
脑. 牛津: 牛津大学出版社.
Ogmen, H。, Breitmeyer, 乙. G。, & Melvin, 右. (2003). 这
what and where in visual masking. 视觉研究, 43,
1337–1350.
Pascual-Leone, A。, & Walsh, V. (2001). Fast backprojections
from the motion to the primary visual area necessary for
visual awareness. 科学, 292, 510–512.
Perrin, F。, Pernier, J。, Bertrand, 奥。, & Echallier, J. F. (1989).
Spherical splines for scalp potential and current-density
映射. Electroencephalography and Clinical
Neurophysiology, 72, 184–187.
饶, 右. 磷. N。, & Ballard, D. H. (1999). Predictive coding
in the visual cortex: A functional interpretation of some
extra-classical receptive-field effects. 自然神经科学,
2, 79–87.
Ro, T。, Breitmeyer, B., Burton, P。, Singhal, 氮. S。, & Lane, D.
(2003). Feedback contributions to visual awareness in
human occipital cortex. 现代生物学, 13, 1038–1041.
罗尔夫塞玛, 磷. R。, 瘫痪, V. A. F。, Spekreijse, H。, & Bosch, H.
(2002). Figure–ground segregation in a recurrent network
建筑学. 认知神经科学杂志, 14,
525–537.
cerebral-cortex and the neurophysiology of visual masking.
Proceedings of the Royal Society of London, Series B,
Biological Sciences, 257, 9–15.
斯科尔特, H. S。, Witteveen, S. C。, Spekreijse, H。, & 瘫痪,
V. A. F. (2006). The influence of inattention on the neural
correlates of scene segmentation. 脑研究, 1076,
106–115.
军士, C。, 巴耶莱特, S。, & 德阿内, S. (2005). Timing of the
brain events underlying access to consciousness during
the attentional blink. 自然神经科学, 8, 1391–1400.
Spratling, 中号. W., & 约翰逊, 中号. H. (2004). A feedback model
of visual attention. 认知神经科学杂志, 16,
219–237.
汤普森, K. G。, & 沙尔, J. D. (1999). The detection of
visual signals by macaque frontal eye field during masking.
自然神经科学, 2, 283–288.
VanRullen, R。, & 科赫, C. (2003). Visual selective behavior
can be triggered by a feed-forward process. 杂志
Cognitive Neuroscience, 15, 209–217.
Vorberg, D ., Mattler, U。, Heinecke, A。, 施密特, T。, &
Schwarzbach, J. (2003). Different time courses for visual
perception and action priming. 诉讼程序
美国国家科学院, 美国。, 100, 6275–6280.
Weidner, R。, Shah, 氮. J。, & Fink, G. 右. (2006). The neural
basis of perceptual hypothesis generation and testing.
认知神经科学杂志, 18, 258–266.
Weiskrantz, L。, Barbur, J. L。, & Sahraie, A. (1995). Parameters
affecting conscious versus unconscious visual-discrimination
with damage to the visual-cortex (V1). 诉讼程序
美国国家科学院, 美国。, 92, 6122–6126.
Weiskrantz, L。, Warrington, 乙. K., Sanders, 中号. D ., & 马歇尔, J.
(1974). Visual capacity in the hemianopic field following
a restricted occipital ablation. Brain, 97, 709–728.
Wickens, 时间. D. (2002). Elementary signal detection theory.
牛津: 牛津大学出版社.
Woodman, G. F。, & Luck, S. J. (2003). Dissociations
among attention, 洞察力, and awareness during
object-substitution masking. 心理科学, 14,
605–611.
Zipser, K., 瘫痪, V. A. F。, & Schiller, 磷. H. (1996).
Contextual modulation in primary visual cortex. 杂志
of Neuroscience, 16, 7376–7389.
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
t
哦
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
p
r
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
A
米
我
r
e
.
d
C
你
哦
米
哦
/
C
j
n
哦
A
C
r
t
n
我
C
/
A
e
–
r
p
t
d
我
C
1
我
9
e
9
–
1
p
4
d
8
F
8
/
1
1
9
9
/
3
6
9
5
/
2
1
8
4
8
哦
8
C
/
n
1
2
7
0
5
0
6
7
8
1
6
9
4
/
9
j
1
哦
4
C
8
n
8
.
p
2
d
0
0
乙
7
y
.
G
1
你
9
e
.
s
9
t
.
哦
1
n
4
0
8
7
8
S
.
e
p
p
d
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
t
.
.
.
.
.
F
哦
n
1
8
中号
A
y
2
0
2
1
继续行驶, 斯科尔特, and Lamme
1497