Manipulating the Experienced Onset of Intention
after Action Execution
Hakwan C. Lau1,2, Robert D. Rogers2, and Richard E. Passingham1,2
抽象的
& Using transcranial magnetic stimulation (TMS), 我们有
tested the time needed for the perceived onset of spontaneous
motor intention to be fully determined. We found that TMS
applied over the presupplementary motor area after the exe-
cution of a simple spontaneous action shifted the perceived
onset of the motor intention backward in time, and shifted
the perceived time of action execution forward in time. 这
size of the effect was similar regardless of whether TMS was
applied immediately after the action or 200 msec after. 这
results of three control studies suggest that this effect is time-
limited, specific to modality, and also specific to the anatom-
ical site of stimulation. We conclude that the perceived onset
of intention depends, at least in part, on neural activity that
takes place after the execution of action. A model, 这是
based on the mechanism of cue integration under the pres-
ence of noise, is offered to explain the results. The implica-
tions for the conscious control of spontaneous actions are
discussed. &
介绍
We experience a strong sense of conscious control when
generating spontaneous, or self-paced, 运动动作.
This experience has been challenged as ‘‘illusory’’: Wegner
(2002, 2003) has argued that although we perceive our
motor intentions to arise before the execution of actions,
we cannot confidently conclude that the former is caus-
ing the latter; there might be a mere temporal correla-
tion between the two events. 尽管如此, this argument
alone is insufficient to establish that the conscious will is
illusory, as it does not show that motor intentions are in
fact not causing the actions. One strong demonstration
for the case of illusory conscious control would be that
our perceived temporal order of intentions and actions
是, 实际上, 错误的. If intentions, 实际上, arise after actions,
they could not, 原则, be causing the actions.
Using a cross-modal timing method, Libet (1985) 和
Libet, Gleason, 赖特, and Pearl (1983) have reported
that participants start to experience their motor in-
tention at about 200 msec before making a spontane-
ous finger movement. 然而, critics have suggested
that the reported timings given by participants in the
Libet clock paradigm might not be accurate ( Joordens,
van Duijn, & Spalek, 2002; 克莱因, 2002; Trevena &
磨坊主, 2002; Gomes, 1998; Dennett & Kinsbourne,
1995; Dennett, 1991; Libet, 1985). In the context of the
debate of whether conscious intentions cause actions, A
critical possibility is that the reported onset of intentions
is determined by neural activity that takes place after
action execution. This has not been tested before.
1伦敦大学学院, 2牛津大学
Despite its counterintuitiveness, this possibility is
supported by psychophysical research on conscious
意识. Phenomena, such as backward masking
(Breitmeyer 1984) and postdiction (Alais & Burr, 2003;
Eagleman & Sejnowski, 2000), highlight the retrospec-
tive nature of perceptual experience, 那是, the inten-
sity and content of an experience can depend on
information that only becomes available after the sub-
jective time of perception. 尤其,
it has been
shown that the temporal extent of this retrospective
effect can last for as long as 200 毫秒, 正如所证明的
in the auditory modality (Alais & Burr, 2003).
We have therefore tested whether transcranial mag-
netic stimulation (TMS), applied after action execution,
has any effect on the reported onset of intention as
reported by the participants using the method of Libet
等人. (1983). Due to concerns about the absolute
accuracy of the reported timings, 然而, 我们只
assessed the shifts in the reported timings due to TMS,
but not the reported absolute measures. 还, 因为
TMS was only applied in half of the trials in a random
fashion together with sham TMS, participants could not
predict if they were going to be stimulated in a particular
trial until the action was executed.
We have previously performed a functional magnetic
resonance imaging (功能磁共振成像) experiment using Libet et al.’s
范例 (Lau, 罗杰斯, Haggard, & Passingham, 2004),
and we found that when participants were required to
estimate the onset of their intentions, activations were
found in areas that are known to be involved in motor
preparation and attention to action, which suggests that,
in this paradigm, the participants were in fact trying to
D 2007 麻省理工学院
认知神经科学杂志 19:1, PP. 81–90
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
D
H
哦
t
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
r
p
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
米
A
我
r
e
.
d
C
你
哦
米
哦
/
C
n
j
哦
A
C
r
t
n
我
C
/
e
A
–
r
p
t
d
我
C
1
我
9
e
1
–
8
p
1
d
F
1
/
9
1
3
6
9
0
/
6
1
1
/
8
哦
1
C
/
n
1
2
7
0
5
0
6
7
4
1
1
9
4
/
1
j
8
哦
1
C
p
n
d
.
2
乙
0
y
0
G
7
你
.
e
1
s
9
t
.
哦
1
n
.
0
8
8
1
S
.
p
e
d
p
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
.
.
.
.
.
t
F
哦
n
1
8
中号
A
y
2
0
2
1
access information related to the generation of action. 在
addition to activations in the dorsal prefrontal and
parietal cortices that are commonly found in attention-
ally demanding tasks, we found activation in the pre-
supplementary motor area (pre-SMA), which we argued
is likely to reflect the representation of intention. 这
idea that the pre-SMA is particularly important for
spontaneous intention is also supported by previous
学习. 第一的, it has been reported that electrical stimu-
lation of the medial frontal cortex elicits the feeling of
an urge to move (Fried et al., 1991). 第二, 病变
of the medial frontal cortex abolish self-initiated move-
ments in macaque monkeys (Thaler, 陈, Nixon, Stern,
& Passingham, 1995). 最后, we have previously re-
ported activity in the pre-SMA when participants gener-
ate actions of their own free choice (Lau, 罗杰斯, &
Passingham, 2006). The pre-SMA was therefore chosen
as the main site of interest in this study (数字 2). 全部
stimulations conducted in Experiments 1, 2, 和 3 是
targeted at this anatomical region. In Experiment 1, 我们
found that TMS after action execution induces shifts in
the perceived onset of both intentions and movements.
实验 2, 3, 和 4 were set up to rule out alter-
native interpretations.
EXPERIMENT 1: MAIN EXPERIMENT
方法
Six male and four female healthy participants were
tested in this experiment. The task instructions were
explained to them verbally before they received safety
screening and gave informed consent.
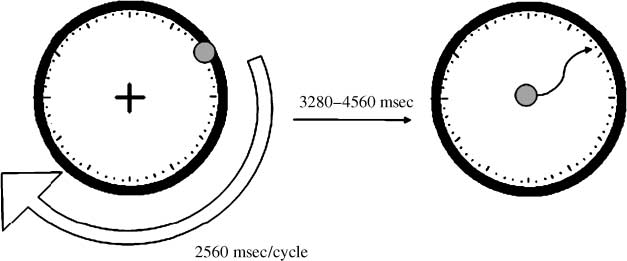
The psychological tasks were based on Libet et al.’s
(1983) clock paradigm (数字 1), and the detailed
procedures were similar to those used in a previous
实验 (Lau, 罗杰斯, Haggard, 等人。, 2004). Partic-
ipants rested their head on a chin rest, and in every trial,
they watched a red dot revolving around a clock face
(diameter (西德:1) 38) on a computer screen placed at about
50 cm away from the chin rest. There was a fixation cross
presented in the center of the clock face, and partic-
ipants were required to maintain fixation while the
red dot was moving. The dot revolved at a speed of
2560 msec per cycle, and after the first revolution in
每次试验, the participants were required to press a
computer mouse button using their left index finger,
at a random time point of their own choice. After the
button press, the dot kept on moving for a period of
1280–2560 msec. There was then a delay of 2000 毫秒,
after which the red dot reappeared at the center of the
clock face, and participants were required to control the
dot as a cursor using another computer mouse held in
their right hand.
In the intention condition, they were required to
move the cursor to where the dot was when they first
felt their intention to press the button. In the movement
状况, they were required to move the cursor to
where the dot was when they actually pressed the
button with their left index finger. After they selected
the location, they clicked the mouse button with their
right hand to finish the trial. The next trial began after an
interval of 1000 毫秒. In half of the trials, TMS was
applied over the pre-SMA. In the remaining trials, 那里
was a sham TMS triggered by another TMS machine with
a coil placed near the back of the head of the partic-
爱普茨, but directed away from the cortex. The stimula-
的 (real or sham) was either presented immediately
after action execution or 200 msec afterwards. 那里
were a total of 240 试验. The two main task conditions
were organized into eight 30-trial blocks. Real and sham
TMS were randomly allocated, and so were early and late
数字 1. Libet’s clock
范例. Participants made a
spontaneous finger movement
while watching a red dot
revolving around a clock face
(左边). After the action, the dot
kept on going for 1/2–1 cycle
before it disappeared. 然后
there was a delay, 后
which the dot reappeared
at the center of the clock.
Participants used a computer
mouse to control the dot
as a cursor and moved it to
the location where the dot
was when they first felt the
intention to move in the
intention condition, 和
to the location where the
dot was when they actually
made the movement in the
movement condition.
82
认知神经科学杂志
体积 19, 数字 1
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
D
H
哦
t
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
r
p
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
米
A
我
r
e
.
d
C
你
哦
米
哦
/
C
n
j
哦
A
C
r
t
n
我
C
/
e
A
–
r
p
t
d
我
C
1
我
9
e
1
–
8
p
1
d
F
1
/
9
1
3
6
9
0
/
6
1
1
/
8
哦
1
C
/
n
1
2
7
0
5
0
6
7
4
1
1
9
4
/
1
j
8
哦
1
C
p
n
d
.
2
乙
0
y
0
G
7
你
.
e
1
s
9
t
.
哦
1
n
.
0
8
8
1
S
.
p
e
d
p
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
.
F
t
.
.
.
.
哦
n
1
8
中号
A
y
2
0
2
1
near the midline. The sham TMS pulses were triggered
by a similar TMS machine, and the intensity was adjusted
individually for each participant to a level that produced
an auditory ‘‘click’’ sound of a volume that was reported
to be similar to that of the real TMS.
实验 2, 3, 和 4 were set up, after obtaining
the results in this experiment, to exclude alternative
解释. 所以, the procedures were similar
to those used in this experiment.
结果
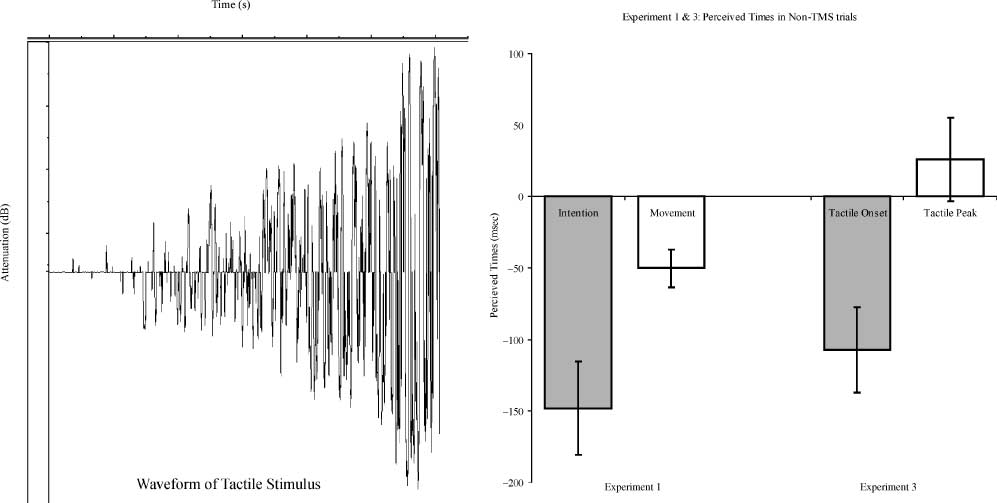
When only sham TMS trials were considered, the group
mean for the judged onset of intention was (西德:2)148 毫秒
(标准差= 103 毫秒) relative to the time of the recorded
button press. The group mean for the judgment of
movement was (西德:2)50 毫秒 (标准差= 42 毫秒). As reported
in previous studies (Lau, 罗杰斯, Haggard, 等人。, 2004;
Sirigu et al., 2004; Haggard & Eimer, 1999), the two
measures differed significantly [one-tailed t test was
used because the test was motivated by evidence ob-
served in previous studies, t(9) = 2.899, p = .009].
The effect of TMS for each individual was assessed by
subtracting the median judgment value for the sham
TMS trials from that of the TMS trials. The group mean
for the TMS effect was (西德:2)9 毫秒 (标准差= 32 毫秒) 为了
the intention condition with 0 msec delayed TMS,
(西德:2)16 毫秒 (标准差= 32 毫秒) for the intention condition
和 200 msec delayed TMS, 14 毫秒 (标准差= 27 毫秒) 为了
the movement condition with 0 msec delayed TMS, 和
9 毫秒 (标准差= 16 毫秒) for the movement condition.
These data are plotted in Figure 3. They were entered
into an analysis of variance (ANOVA) with task (Inten-
tion vs. Movement) and time (0 msec or 200 毫秒)
considered as experimental factors, and it was found
that task was a significant factor [F(1,9) = 24.089,
p = .001] but time was not [F(1,9) = 0.485, p = .504).
There was no significant interaction between time and
任务 [F(1,9) = 0.009, p = .926].
When the data for the different time points of TMS
were considered together, it could be shown that the
effect of TMS on the judgment of intention is signifi-
cantly negative to zero[two-tailed t test, t(9) = (西德:2)2.6,
p = .029] and the effect on the judgment of movement
是, although weaker, significantly positive to zero [二-
tailed t test, t(9) = 2.35, p = .045].
EXPERIMENT 2: TIME SPECIFICITY
方法
Seven male and three female healthy participants were
tested in this experiment where the effects of TMS
applied at other time points were evaluated. The meth-
odology and procedures were similar to those used in
实验 1, except that the time points for TMS were
不同的. Instead of applying a delay of either 0 毫秒
Lau, 罗杰斯, and Passingham
83
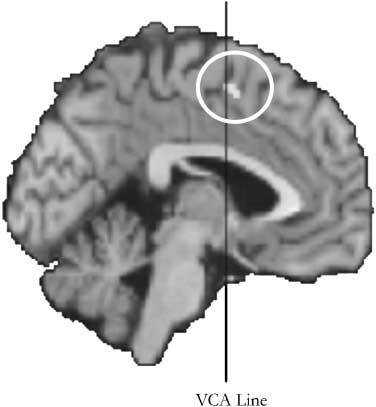
数字 2. The pre-SMA. This area was the target of stimulation in
实验 1 到 3. The coordinates (X, y, z = 2, 4, 54) were obtained
from a previous fMRI study (Lau, 罗杰斯, Haggard, 等人。, 2004), 和
figure is also adapted from the report of that study. The VCA line
vertically passes through the anterior commissure, and is the border
between the pre-SMA and the SMA proper.
stimulations. 所以, from the point of view of the
参与者, they did not know whether they would be
stimulated, or at which point would the pulse be
triggered, until after they have pressed the button.
The anatomical site of stimulation was the pre-SMA
(数字 2). The localization for stimulation here was
based on the coordinates obtained from the imaging
study described in the previous section (X, y, z, = 2, 4,
54). The relationship between the brain of each indi-
vidual and the standard Talairach space was computed
by applying spatial normalization (FLIRT version 5.0,
www.fmrib.ox.ac.uk/fsl/flirt/index.html) to the previous-
ly acquired MRI scans of each participant, using the
Montreal Neurological Institute canonical single-subject,
high-resolution MRI image as the template as in the
previous fMRI study. Using the Brainsight Frameless
系统 (version 1.5B3, www.rogue-research.com/), 这
location of the pre-SMA was then marked on each in-
dividual’s MRI scan, which was then registered with
the actual brain in 3-D space. Because the location
of the intended site of stimulation is quite far away from
the surface of the scalp, a double cone coil (Magstim
公司, Whitland, South West Wales, 英国) was used in
this experiment. The hotspot of the coil was placed
directly above the marked position for stimulation on
the MRI scan as presented on a computer screen by the
Brainsight software. The stimulation pulses were trig-
gered by a Magstim Rapid Rate TMS machine (Magstim
公司), and the intensity of stimulation was set at 5%
above the active motor threshold for a noticeable foot
twitch, tested over the foot area of the motor cortex
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
D
H
哦
t
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
r
p
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
米
A
我
r
e
.
d
C
你
哦
米
哦
/
C
n
j
哦
A
C
r
t
n
我
C
/
e
A
–
r
p
t
d
我
C
1
我
9
e
1
–
8
p
1
d
F
1
/
9
1
3
6
9
0
/
6
1
1
/
8
哦
1
C
/
n
1
2
7
0
5
0
6
7
4
1
1
9
4
/
1
j
8
哦
1
C
p
n
d
.
2
乙
0
y
0
G
7
你
.
e
1
s
9
t
.
哦
1
n
.
0
8
8
1
S
.
p
e
d
p
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
.
F
.
.
.
.
t
哦
n
1
8
中号
A
y
2
0
2
1
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
D
H
哦
t
w
t
p
n
:
哦
/
A
/
d
米
e
我
d
t
F
r
p
哦
r
米
C
.
H
s
我
p
我
v
d
e
我
r
r
e
C
C
H
t
.
米
A
我
r
e
.
d
C
你
哦
米
哦
/
C
n
j
哦
A
C
r
t
n
我
C
/
e
A
–
r
p
t
d
我
C
1
我
9
e
1
–
8
p
1
d
F
1
/
9
1
3
6
9
0
/
6
1
1
/
8
哦
1
C
/
n
1
2
7
0
5
0
6
7
4
1
1
9
4
/
1
j
8
哦
1
C
p
n
d
.
2
乙
0
y
0
G
7
你
.
e
1
s
9
t
.
哦
1
n
.
0
8
8
1
S
.
p
e
d
p
F
e
米
乙
乙
y
e
r
G
2
你
0
e
2
s
3
t
/
j
.
.
.
.
t
.
F
哦
n
1
8
中号
A
y
2
0
2
1
数字 3. The effects of TMS on the perceived times. TMS produced a forward shift in the reported onset of intention and a backward shift
in the reported onset of movement, regardless of whether TMS was administered immediately after the action (button press) 或者 200 毫秒
afterwards (实验 1, top left). This effect of exaggerating the difference between the two reported onsets was significant ( p < .001,
main effect of task in the ANOVA; see also Figure 4). Experiments 2, 3, and 4 were control studies set up to rule out alternative explanations.
In Experiment 2 (top right), TMS was administered either 500 msec after the action or immediately before the subjects were required to
report the experienced onsets. In Experiment 3 (bottom left), the subjects were required to judge the onset and offset of a slowly ramping-up
tactile stimuli (see Figure 5), instead of the onsets of intention and movements; they were not required to make a spontaneous action.
Experiment 4 was identical to Experiment 1, except that the motor cortex, instead of the pre-SMA, was targeted in TMS. None of the control
studies showed any significant result in the statistical analyses, and the patterns of the effects were clearly different from those observed in
Experiment 1. The error bars represent standard errors across participants.
or 200 msec, the delay in this experiment was either
500 msec, or between 3280 and 4560 msec, so that the
TMS pulse (shame or real) was triggered at the point
when the cursor appeared at the middle of the clock face,
prompting the participants to report the estimated tim-
ings. These times were chosen to test whether the effect
obtained in Experiment 1 was actually due to memory or
responding, rather than the experienced onset itself.
Results
When only the sham TMS trials were considered, on
average, the participants judged the onset of intention
to be (cid:2)110 msec (SD = 82 msec) relative to the times
for recorded button press. The average for the judgment
of movement was (cid:2)7 msec (SD = 69 msec). The two
measures differed significantly [one-tailed t test was
used as in Experiment 1 because the test was motivated
by evidence observed in previous studies, t(9) = 4.964,
p = .0005].
The group mean for the TMS effect was 9 msec (SD =
28 msec) for the intention condition with 500 msec
delayed TMS, 0 msec (SD = 26 msec) for the inten-
tion condition with 3280–4560 msec delayed TMS,
5 msec (SD = 21 msec) for the movement condition
with 500 msec delayed TMS, and 5 msec (SD = 32 msec)
for the movement condition with 3280–4560 msec de-
layed TMS. These data are plotted in Figure 3.
As in Experiment 1, the effects of TMS for each task
condition were entered into an ANOVA with task (In-
tention vs. Movement) and time (500 msec or 3280–
4560 msec) considered as experimental factors, and it
was found that neither task nor time was a significant
factor [F(1,9) = 0.001, p = .974 and F(1,9) = 0.208,
p = .659, respectively]. There was also no significant
interaction between time and task [F(1,9) = 0.549,
p = .478]. The main effect of task was of special interest
as this was found to be significant in Experiment 1.
However, from the p value it is clear that this effect
would not have been significant even if a one-tailed t test
had been applied. The size of this effect (i.e., the ef-
fect of TMS for intention minus the effect of TMS for
movement) is plotted in Figure 4 to allow a comparison
across all experiments reported here. Also, the effect of
84
Journal of Cognitive Neuroscience
Volume 19, Number 1
up that is characteristic of the electrophysiological
measures of the readiness potential (Deecke, Scheid,
& Kornhuber, 1969; Kornhuber & Deecke, 1965). In-
stead of estimating the onset of intention and move-
ment, participants were required to estimate the timing
of either the onset or the peak of the tactile stimulus
using the Libet clock. A small tactile stimulator (100-(cid:1)
bone conduction vibrators; Oticon, Hamilton, Scotland)
was connected to the audio output from the sound card
of a PC, where a wave file containing white noise was
played, to give the tactile sensation. The intensity of the
stimulation was set at a low level, only slightly beyond
detection threshold for each participant, such that the
earliest part of the stimulation would not be felt. The
tactile stimulator and the left index finger of the partic-
ipants were inserted into a piece-insulating sponge so
as to minimize the auditory noise associated with the
tactile stimulus. TMS was triggered at either 0 msec or
200 msec after the offset of the tactile stimulus.
Results
When only the sham TMS trials were considered, on
average, the participants judged the onset of the tactile
stimulus to be (cid:2)107 msec (SD = 95 msec) relative to the
offset of the stimulus. The average for the judgment of
the peak of the stimulus was 26 msec (SD = 92 msec)
relative to the actual offset of the stimulus. The two
measures differed significantly [one-tailed t test used as
in Experiment 1, t(9) = 4.764, p = .0005], and the
magnitude of this difference was similar to the difference
between the intention judgment and movement judg-
ment in Experiment 1 (see Figure 5).
The group mean for the TMS effect was (cid:2)18 msec
(SD = 46 msec) for the onset timing condition with
0 msec delayed TMS, (cid:2)7 msec (SD = 41 msec) for the
onset timing condition with 200 msec delayed TMS,
(cid:2)17 msec (SD = 31 msec) for the peak timing condition
with 0 msec delayed TMS, and 2 msec (SD = 22 msec)
for the peak timing condition with 200 msec delayed
TMS. These data are plotted in Figure 3.
As in Experiment 1, the effects of TMS for each task
condition were entered into an ANOVA with task (On-
set Timing vs. Offset Timing) and time (0 msec or
200 msec) considered as experimental factors, and it
was found that neither task nor time was a significant
factor [F(1,9) = 0.222, p = .649 and F(1,9) = 1.716,
p = .223, respectively]. There was also no significant
interaction between the two factors [F(1,9) = 0.159,
p = .699]. The main effect of task was of special inter-
est as this was found to be significant in Experiment 1.
However, from the p value it is clear that this effect
would not have been significant even if a one-tailed
t test was applied. The size of this effect (the effect of
TMS for onset timing minus the effect of TMS for peak
timing) is plotted in Figure 4 to allow a comparison
across all experiments reported here.
Lau, Rogers, and Passingham
85
Figure 4. The exaggeration of the difference between the two
reported onsets as induced by TMS. Plotted on the vertical scale
is the size of the main effect of task, that is, the difference between
the effects of TMS on the reported onset of intention and the
reported onset of movement (for Experiments 1, 2, and 4) or
between the effects of TMS on the reported onset of the tactile
stimulus and the reported offset of the stimulus (for Experiment 3).
This effect was clearly not present for the control experiments,
Experiments 2, 3, and 4 ( p = .974, .649, and .493, respectively).
The error bars represent standard errors across participants.
TMS (TMS vs. sham) was not significant on either the
intention task [two-tailed t test, t(9) = 0.720, p = .435]
or the movement task alone [two-tailed t test, t(9) =
0.818, p = .490].
The effect of task was also assessed when the trials
for the different time points of TMS were considered
separately. One-tailed t test was used based on the
results obtained in Experiment 1 to maximize statistical
power. The effect of task was not significant for either
the 500 msec trials [direction of effect opposite to
prediction based on Experiment 1, one-tailed t test,
t(9) = (cid:2)0.694, p = ns] or the 3280–4560 msec trials
[one-tailed t test, t(9) = 0.331, p = .374].
EXPERIMENT 3: MODALITY SPECIFICITY
Methods
Eight male and two female healthy participants were
tested in this experiment, which was set up to test if the
effect in Experiment 1 was actually due to the general
mechanism of cross-modal timing using the clock face.
In particular, we wanted to know whether TMS simply
exaggerates the perceived temporal difference between
early and late events. This was tested by presenting a
tactile stimulus to the subjects and requiring them to
judge the timing of this stimulus using the clock. The
methodology and procedures were similar to those used
in Experiment 1. Instead of requiring the participants to
make a self-paced left index finger movement, a slowly
ramping up tactile stimulus (duration = 600 msec) was
applied to the tip of this finger at a random time point
(2960–7980 msec) in every trial. The tactile stimulus
used here had the same property of slowly ramping
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
n
j
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
1
-
8
p
1
d
f
1
/
9
1
3
6
9
0
/
6
1
1
/
8
o
1
c
/
n
1
2
7
0
5
0
6
7
4
1
1
9
4
/
1
j
8
o
1
c
p
n
d
.
2
b
0
y
0
g
7
u
.
e
1
s
9
t
.
o
1
n
.
0
8
8
1
S
.
p
e
d
p
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
t
.
.
f
.
.
o
n
1
8
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
n
j
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
1
-
8
p
1
d
f
1
/
9
1
3
6
9
0
/
6
1
1
/
8
o
1
c
/
n
1
2
7
0
5
0
6
7
4
1
1
9
4
/
1
j
8
o
1
c
p
n
d
.
2
b
0
y
0
g
7
u
.
e
1
s
9
t
.
o
1
n
.
0
8
8
1
S
.
p
e
d
p
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
.
.
t
f
o
n
1
8
M
a
y
2
0
2
1
Figure 5. The temporal profile of the tactile stimulus used in Experiment 3. The tactile stimulus used in Experiment 3 took a slowly
ramping-up form (left), and was presented at a low intensity such that the early part of the stimulus was not felt. This was set up to mimic
the form of the readiness potential preceding spontaneous actions (Deecke et al., 1969; Kornhuber & Deecke, 1965). The magnitude of the
difference between the perceived onset of the tactile stimulus and the perceived time of the peak of the stimulus in Experiment 3 was very
similar to the magnitude of the difference between the perceived onset of intention and the perceived time of movement in Experiment 1
(right).
Also, the effect of TMS (TMS vs. sham) was not sig-
nificant on either the onset judgments [two-tailed t test,
t(9) = (cid:2)1.197, p = .262] or the offset judgments alone
[two-tailed t test, t(9) = (cid:2)1.185, p = .266].
Despite the fact that task was not found to be a sig-
nificant factor for the effect of TMS in this experiment,
the magnitude of the effect of TMS on the onset judg-
ments was somewhat similar to that of the effect of TMS
on intention in Experiment 1. However, this nonsignif-
icant effect was not specific to onset judgment alone but
was also there in offset judgments, which means that,
unlike in Experiment 1, this effect was not specific to
task. It is likely that because TMS produces a tactile
sensation to the scalp, it produced a nonspecific inter-
ference with the tactile judgments in general. That
would fit with the fact that this effect was stronger for
the TMS applied closer to the stimuli. In any case, it is
clear that TMS did not exaggerate the perceived tempo-
ral difference between early and late events as observed
in Experiment 1.
EXPERIMENT 4: ANATOMICAL SPECIFICITY
Methods
Seven male and three female healthy participants were
tested in this experiment in which we try to assess if the
effect obtained in the experiment was simply because
TMS added noise to the motor system. The methodol-
ogy and procedures were similar to those used in
Experiment 1, except that the site of TMS was the right
primary motor cortex instead of the pre-SMA. Because
of the ease of accessibility of the motor cortex as shown
in previous studies, a normal figure-of-eight shaped
coil (Double 70mm Coil, Magstim, www.magstim.com/
Products.html) was used. The localization of the primary
motor cortex was done by checking the intensity of
TMS-induced motor twitch for the first dorsal interos-
seus in the right hand of the participants while moving
the coil around the expected region of the scalp. Once
the most sensitive spot was identified, the threshold
for stimulation was set at 5% above the active motor
threshold for a noticeable hand twitch.
Results
When only the sham TMS trials were considered, the
group mean for the judged onset of
intention was
(cid:2)99 msec (SD = 79 msec) relative to the time of the
recorded button press. The group mean for the judg-
ment of movement was (cid:2)13 msec (SD = 76 msec). The
two measures differed significantly [one-tailed t test as in
Experiment 1 because the test was motivated by evidence
observed in previous studies, t(9) = 3.733, p = .002].
The group mean for the TMS effect was (cid:2)8 msec
(SD = 30 msec) for the intention condition with 0 msec
delayed TMS, 16 msec (SD = 28 msec) for the intention
condition with 200 msec delayed TMS, (cid:2)4 msec (SD =
86
Journal of Cognitive Neuroscience
Volume 19, Number 1
23 msec) the movement condition with 0 msec delayed
TMS, and (cid:2)2 msec (SD = 26 msec) for the movement
condition with 200 msec delayed TMS. These data are
plotted in Figure 3.
As in Experiment 1, the effects of TMS for each task
condition were entered into an ANOVA with task (Inten-
tion vs. Movement) and time (0 msec or 200 msec) con-
sidered as experimental factors, and it was found that
neither task nor time was a significant factor [F(1,9) =
0.510, p = .493 and F(1,9) = 2.418, p = .154, respective-
ly]. There was also no significant interaction between
time and task [F(1,9) = 2.052, p = .186]. The main effect
of task was of special interest as this was found to be
significant in Experiment 1. However, from the p value
it is clear that this effect would not have been signifi-
cant even if a one-tailed t test was applied. The size of this
effect (i.e., the effect of TMS for intention minus the ef-
fect of TMS for movement) is plotted in Figure 4 to allow
a comparison across all experiments reported here.
Also, the effect of TMS (TMS vs. sham) was not
significant on either the intention task [two-tailed t test,
t(9) = 0.686, p = .510] or the movement task alone
[two-tailed t test, t(9) = (cid:2)0.456, p = .659].
DISCUSSION
Summary of Findings
To summarize the results, Experiment 1 showed that
there was a retrospective effect for TMS over the pre-
SMA on the perceived onset of intention as well as for
perceived timing of the movement itself. TMS shifted the
perceived onset of intention backward in time and
shifted the perceived timing of the movement forward
in time. The main effect of task, that is, the differential
effect on intention and movement, is about 24 msec.
This difference is not driven by the TMS effect on the
movement condition alone, as the effect of TMS in the
intention condition is also significant on its own. This
differential effect was found for both stimulations ap-
plied at 0 msec and 200 msec after action execution.
One could argue that the TMS effect we have observed
was not genuinely retrospective, but rather, it simply af-
fected the memory of timing, or the reporting of the
timing, which took place after the TMS. However, Exper-
iment 2 showed that the effect observed in Experiment 1,
that is, the exaggeration of the difference of the judg-
ments for the onsets of intention and movement, was
not found when TMS was applied at 500 msec after the
action or right before the participants made their reports
of their time estimates. One could argue that this only
shows that there is a critical time window within which
memory is susceptible to manipulation. However, this
interpretation would require the ad hoc assumption that
an experience could be first generated and then misre-
membered as something else, unbeknownst to the sub-
ject. If this assumption is allowed, one could as well
argue against the existence of well-established phenom-
ena, such as backward masking (Breitmeyer, 1984) and
flash-lag (Nijhawan, 1994, 2002; Eagleman & Sejnowski,
2000), by reinterpreting them as results of failure or
change of memory. Although philosophers have pointed
out that we cannot unequivocally reject these alternative
interpretations (Dennett & Kinsbourne, 1995; Dennett,
1991), one generally prefers parsimonious explanations
that do not require ad hoc assumptions.
Neither did we observe the same effect when we tested
this for the tactile modality, as shown in Experiment 3
when the same timing method was used. This suggests
that TMS did not simply affect the visual processing re-
quired to make accurate judgments in the Libet task. In
particular, TMS did not simply exaggerate the perceived
temporal difference between early and late events. The
tactile task in Experiment 3 was similar in structure to
the task in Experiment 1. When only the control sham
TMS trials where considered, the differences between the
perceived onset of the tactile stimulus and the perceived
time for its peak were very similar to the differences
between the perceived onset of intention and perceived
time of movement (see Figure 4). Yet, no significant effect
was observed in Experiment 3.
The targeted anatomical site of stimulation of Experi-
ments 1 to 3 was the pre-SMA. This anatomical localiza-
tion was based on the fMRI results of our previous study
(Lau, Rogers, Haggard, et al., 2004), and each individual
participant’s structural MRI scan. However, we could not
be sure that only the pre-SMA was stimulated, as it was
not clear if TMS afforded such high spatial specificity,
especially given the potential spread of the effect and
interaction between different areas. Also, the lateral
premotor areas might have been stimulated by the
magnetic fields generated by the wings of the double-
cone coil used in this experiment, too. Nonetheless, the
conclusion of the study does not depend on the precision
of the stimulation, as the main question is about whether
the experience of intention is fully determined before
action execution, but not about where it is represented in
the brain. Experiment 4 further showed that the effect
observed in Experiment 1 is, in fact, reasonably specific, in
that even when another area in the motor system (the
primary motor cortex) is stimulated, the same effect is
not observed. This suggests that the effect was not simply
due to the fact that TMS added noise to the motor system
or that participants were simply startled.
Effect Size and Reliability of the Data
One could argue that the temporal shifts produced by
TMS are small, and thus, unimpressive. Due to consid-
erations about the safety and comfort of the partici-
pants, we did not apply the stimulations at a higher
intensity or in a rapid-rate, repetitive fashion. This might
be one reason why the size of the effect did not appear
to be very large. However, the size of the temporal shifts
Lau, Rogers, and Passingham
87
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
n
j
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
1
-
8
p
1
d
f
1
/
9
1
3
6
9
0
/
6
1
1
/
8
o
1
c
/
n
1
2
7
0
5
0
6
7
4
1
1
9
4
/
1
j
8
o
1
c
p
n
d
.
2
b
0
y
0
g
7
u
.
e
1
s
9
t
.
o
1
n
.
0
8
8
1
S
.
p
e
d
p
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
t
.
f
.
.
o
n
1
8
M
a
y
2
0
2
1
cannot be directly compared with the results obtained
in most other cognitive TMS experiments ( Walsh &
Pascual-Leone, 2003; Stewart, Ellison, Walsh, & Cowey,
2001), as the common measures in previous experi-
ments are normally a change in reaction times or error
rates, which are very different from subjective onset
estimates. One study (Haggard, Clark, & Kalogeras,
2002), which used the same paradigm to investigate
the retrospective effect of external stimuli on the per-
ceived onset of movement (but not intention), re-
vealed a temporal shift of only 15 msec. Another study
(Haggard & Magno, 1999) used the same paradigm to
investigate the prospective effect of TMS on the per-
ceived onset of movement (but not intention), and one
of the targeted areas of stimulation was close to the pre-
SMA (FCz). It was found that TMS applied to this area
prior to action execution can only shift the perceived
onset of movement by 54 msec, even when the actual
shift of movement execution was as large as 113 msec.
Taken together, it seems that experimentally induced
temporal shifts as measured by the Libet method are, in
general, not very large. Nonetheless, this does not mean
that the measures are uninformative, as the principled
way to assess the significance of an effect and compare it
with other studies that use different measures is to
characterize it in terms of the variance of the data (i.e.,
to look at the statistical effect size). The eta-squared
value, which is a standard estimate of statistical effect
size, is .728 for the main effect of task in Experiment 1;
the corresponding p value is as small as .001. This is a
relatively high level of significance as far as cognitive
TMS experiments are concerned.
Despite this high level of statistical significance, and
the fact that each experiment involved 10 subjects (even
for each control studies), there might still be concerns
about the reliability of the presented data. In particular,
the baseline values of the reported onsets (i.e., values
for sham TMS trials) fluctuated across Experiment 1 and
the control studies. This could be due to the fact that
some of the control studies differed from Experiment 1
in various aspects: Experiment 2 presented sham TMS at
later time points; Experiment 3 required subjects to
judge the timing of a tactile stimulus instead of a self-
generated action; Experiment 4 presented the sham
TMS using a different coil
in order to match for the
sound of the TMS administered in that experiment.
These suggest that subtle factors, such as the auditory
level and the timing of the sham TMS, could, in fact,
affect the reported onsets in this difficult timing task.
Also, the variability of the effect of TMS was also a
source of concern. Although the statistical analyses
revealed no significant effect in all control studies, there
were sometimes weak trends observed in unpredicted
directions, as shown in Figure 3. We emphasize that TMS
exaggerated the difference between the reported onsets
of intention and movement in a robust fashion, and this
effect was clearly not present in the control studies, as
shown in Figure 4. This is the main effect of interest in
the article. Nonetheless, one major limitation could be
that the control studies involved different experimental
sessions and did not always involve the same subjects.
Future studies would benefit from replicating these
results in the same sessions by, for instance, testing
more different time points of TMS in each session,
thereby combining and replicating the results of Experi-
ments 1 and 2, and possibly exploring other critical time
points such as those before action execution as well.
Direction of Temporal Shifts
In Experiment 1, it was found that retrospective TMS
shifted the perceived onset of intention backward in
time, and shifted the perceived timing of action execu-
tion forward in time. The directions of these effects were
not predicted a priori. Here we offer a tentative model
to account for this pattern of temporal shifts, which we
hope could be tested in future experiments.
When performing difficult perceptual tasks, we often
try to combine information from different sources, such
as cues from different sensory modalities. A natural and
optimal method to combine these different sources of
information is to perform Bayesian cue integration (Knill
& Pouget, 2004). Assuming that the noise or uncertainty
in the information can be characterized as Gaussian in
form, this method amounts to taking a weighted average
of the information from the different sources, where the
weights are inversely proportional to the amount of
noise in each respective source.
The readiness potential (Deecke et al., 1969; Kornhuber
& Deecke, 1965), measurable by EEG from the scalp,
reflects the slowly ramping-up neural activity that pre-
cedes spontaneous movements. It has been argued that
the readiness potential comprises different components
that are functionally distinct (Deecke, 1987). The earliest
onset of the readiness potential is around 1 to 1.5 sec.
Despite the lack of an uncontroversial method to mea-
sure accurately the onset of the experience of intention,
it seems plausible that the experienced onset is not as
early as the onset of the readiness potential. One
possibility is that due to the difficult nature of the task
to estimate the onset of intention, the brain combines
the information reflected by both the early and the late
components of the readiness potential. The late compo-
nents of the readiness potential might indicate that the
onset of intention is much closer to the time of action
execution, and because the signal of the early compo-
nent of the readiness potential is weak, the weighting
for the representation reflected by early component is
likely to be low. That might explain why we typically
experience the onset of the intention as much later than
the onset of the readiness potential.
It has been suggested that the best way to character-
ize the effect of TMS is to view it as adding neural noise
to the targeted cortical area ( Walsh & Pascual-Leone,
88
Journal of Cognitive Neuroscience
Volume 19, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
n
j
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
1
-
8
p
1
d
f
1
/
9
1
3
6
9
0
/
6
1
1
/
8
o
1
c
/
n
1
2
7
0
5
0
6
7
4
1
1
9
4
/
1
j
8
o
1
c
p
n
d
.
2
b
0
y
0
g
7
u
.
e
1
s
9
t
.
o
1
n
.
0
8
8
1
S
.
p
e
d
p
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
f
.
.
.
o
n
1
8
M
a
y
2
0
2
1
2003). The pre-SMA is likely to be one of the sources of
the readiness potential (Cunnington, Windischberger,
Deecke, & Moser, 2002). Because TMS is applied after
action execution in the experiments described in this
article, it is reasonable to assume that noise is mostly
added to the late components of the readiness potential,
but not the early ones. If the brain combines the timing
information of intention in a way that is similar to
optimal Bayesian cue integration, increasing the uncer-
tainty of the representations reflected by the late com-
ponents of the readiness potential would decrease the
weight assign to these representations, and thus,
in-
crease the relative weight for the early components. The
result would be that we estimate the onset of intention
to be earlier.
Similarly, when estimating the timing of action execu-
tion, the brain might combine the information from
proprioceptive sensory feedback as well as from the
representations reflected by the late components of
the readiness potential. The late components of the
readiness potential may indicate an earlier time of action
execution than does the sensory feedback, as the former
is a preparatory signal. According to the model, adding
noise to the late component would lead to a forward
temporal shift of the resulted estimation, as the relative
weight for the sensory feedback is increased.
To explain the present data, the model need not be
fully Bayesian in nature, in the sense that it explicitly
represents the probability distribution functions for the
different cues. Neither does it require the assumption
that noise has a Gaussian structure or is normally dis-
tributed. The foregoing explanation could be derived
so long as the system weighs the different cues accord-
ing to the noise associated with each cue, so that the
higher the noise, the smaller the weight would be
ascribed to that cue.
Conclusions
The main conclusion of this article does not depend on
the aforementioned model. Regardless of what mecha-
nism is used in the brain, the results suggest that the
perceived onset of intention depends on neural activity
that can be manipulated by TMS at as late as 200 msec
after the execution of a spontaneous action. The mech-
anism for this retrospective effect is unclear. One possi-
bility is that TMS over the medial frontal area interferes
with feedback processes that confirm the execution of
the action. The present data also do not determine
whether neural activity that takes place before the
execution of the action is sufficient to yield some form
of experience of intention. It could be the case that
some weaker form of experience of intention is suffi-
ciently determined by neural activity that takes place
before the execution of the action, and such experience
might have some causal impact on the control of the
action. One way to examine this possibility would be to
apply TMS before the execution of action, as suggested
above, and compare the effect with those obtained in
this study. Although the onsets of spontaneous actions
cannot be easily predicted, TMS could be delivered at
random time points. Given sufficient number of trials,
one could identify, after the experiment, the trials where
TMS are delivered just before action onsets, and perform
the analysis on these trials. Before this experiment is
conducted, however, one cannot draw the strong con-
clusion that the experience of having conscious control
of a simple motor action is entirely illusory.
Nonetheless, the current results throw doubt on the
commonsensical view that the experience of intention,
including the experienced onset, is completely deter-
mined before an action. The commonsensical view is
attractive when we assume that the main function of
experience of intention is for the conscious control of
action, but it cannot account for the data presented
here. The data suggest that the perceived onset of inten-
tion depends at least in part on neural activity that takes
place after the execution of action, which could not, in
principle, have any causal impact on the action itself.
An alternative view that is compatible with the data is
that one function of the experience of intention might
be to help clarify the ownership of actions ( Wegner,
2002, 2003), which can help to guide future actions.
This process could take place immediately after action
execution.
Acknowledgments
This work was supported by the Wellcome Trust (R. E. P.) and
a Rhodes Scholarship (H. C. L.). We thank Jon Driver for his
critical comments on part of the results represented.
Reprint requests should be sent to Hakwan Lau, Functional Im-
aging Laboratory, 12 Queen Square, London, WC1N 3BG UK,
or via e-mail: h.lau@fil.ion.ucl.ac.uk.
REFERENCES
Alais, D., & Burr, D. (2003). The ‘‘Flash-Lag’’ effect occurs
in audition and cross-modally. Current Biology, 13, 59–63.
Breitmeyer, B. G. (1984). Visual masking: An integrative
approach. Oxford: Oxford University Press.
Cunnington, R., Windischberger, C., Deecke, L., & Moser, E.
(2002). The preparation and execution of self-initiated and
externally-triggered movement: A study of event-related
fMRI. Neuroimage, 15, 373–385.
Deecke, L. (1987). Bereitschaftspotential as an indicator
of movement preparation in supplementary motor area
and motor cortex. Ciba Foundation Symposium, 132,
231–250.
Deecke, L., Scheid, P., & Kornhuber, H. H. (1969). Distribution
of readiness potential, pre-motion positivity, and motor
potential of the human cerebral cortex preceding voluntary
finger movements. Experimental Brain Research, 7,
158–168.
Dennett, D. C. (1991). Consciousness explained (1st ed.).
Boston: Little Brown and Co.
Dennett, D. C., & Kinsbourne, M. (1995). Time and the
Lau, Rogers, and Passingham
89
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
n
j
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
1
-
8
p
1
d
f
1
/
9
1
3
6
9
0
/
6
1
1
/
8
o
1
c
/
n
1
2
7
0
5
0
6
7
4
1
1
9
4
/
1
j
8
o
1
c
p
n
d
.
2
b
0
y
0
g
7
u
.
e
1
s
9
t
.
o
1
n
.
0
8
8
1
S
.
p
e
d
p
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
f
t
.
.
o
n
1
8
M
a
y
2
0
2
1
observer: The where and when of consciousness in
the brain. Behavioral and Brain Sciences, 15, 183–247.
Eagleman, D. M., & Sejnowski, T. J. (2000). Motion
integration and postdiction in visual awareness. Science,
287, 2036–2038.
Fried, I., Katz, A., McCarthy, G., Sass, K. J., Williamson, P.,
Spencer, S. S., et al. (1991). Functional organization of
human supplementary motor cortex studied by electrical
stimulation. Journal of Neuroscience, 11, 3656–3666.
Gomes, G. (1998). The timing of conscious experience: A
critical review and reinterpretation of Libet’s research.
Consciousness and Cognition, 7, 559–595.
Haggard, P., Clark, S., & Kalogeras, J. (2002). Voluntary
action and conscious awareness. Nature Neuroscience,
5, 382–385.
Haggard, P., & Eimer, M. (1999). On the relation between brain
potentials and the awareness of voluntary movements.
Experimental Brain Research, 126, 128–133.
Haggard, P., & Magno, E. (1999). Localising awareness of
action with transcranial magnetic stimulation. Experimental
Brain Research, 127, 102–107.
Joordens, S., van Duijn, M., & Spalek, T. M. (2002). When
timing the mind one should also mind the timing: Biases
in the measurement of voluntary actions. Consciousness
and Cognition, 11, 231–240.
Klein, S. (2002). Libet’s research on the timing of conscious
intention to act: A commentary. Consciousness and
Cognition, 11, 273–279.
Knill, D. C., & Pouget, A. (2004). The Bayesian brain: The
role of uncertainty in neural coding and computation.
Trends in Neurosciences, 27, 712–719.
Kornhuber, H. H., & Deecke, L. (1965). Changes in the brain
potential in voluntary movements and passive movements
in man: Readiness potential and reafferent potentials.
Pflu¨gers Archiv fu¨r die Gesamte Physiologie des Menschen
und der Tiere, 284, 1–17.
Lau, H. C., Rogers, R. D., Haggard, P., & Passingham, R. E.
(2004). Attention to intention. Science, 303, 1208–1210.
Lau, H., Rogers, R. D., & Passingham, R. E. (2006). Dissociating
response selection and conflict in the medial frontal
surface. Neuroimage, 29, 446–451.
Libet, B. (1985). Unconscious cerebral initiative and the role
of conscious will in voluntary action. Behavioral and
Brain Sciences, 8, 529–566.
Libet, B., Gleason, C. A., Wright, E. W., & Pearl, D. K.
(1983). Time of conscious intention to act in relation
to onset of cerebral activity (readiness-potential). The
unconscious initiation of a freely voluntary act. Brain,
106, 623–642.
Nijhawan, R. (1994). Motion extrapolation in catching.
Nature, 370, 256–257.
Nijhawan, R. (2002). Neural delays, visual motion and the
flash-lag effect. Trends in Cognitive Sciences, 6, 387.
Sirigu, A., Daprati, E., Ciancia, S., Giraux, P., Nighoghossian, N.,
Posada, A., et al. (2004). Altered awareness of voluntary
action after damage to the parietal cortex. Nature
Neuroscience, 7, 80–84.
Stewart, L., Ellison, A., Walsh, V., & Cowey, A. (2001). The
role of transcranial magnetic stimulation (TMS) in studies
of vision, attention and cognition. Acta Psychologica, 107,
275–291.
Thaler, D., Chen, Y. C., Nixon, P. D., Stern, C. E., & Passingham,
R. E. (1995). The functions of the medial premotor cortex:
I. Simple learned movements. Experimental Brain
Research, 102, 445–460.
Trevena, J. A., & Miller, J. (2002). Cortical movement
preparation before and after a conscious decision to
move. Consciousness and Cognition, 11, 162–190.
Walsh, V., & Pascual-Leone, A. (2003). Transcranial magnetic
stimulation: A neurochronometrics of mind. Cambridge:
MIT Press.
Wegner, D. M. (2002). The illusion of conscious will.
Cambridge: MIT Press.
Wegner, D. M. (2003). The mind’s best trick: How we
experience conscious will. Trends in Cognitive Sciences,
7, 65–69.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
n
j
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
1
-
8
p
1
d
f
1
/
9
1
3
6
9
0
/
6
1
1
/
8
o
1
c
/
n
1
2
7
0
5
0
6
7
4
1
1
9
4
/
1
j
8
o
1
c
p
n
d
.
2
b
0
y
0
g
7
u
.
e
1
s
9
t
.
o
1
n
.
0
8
8
1
S
.
p
e
d
p
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
.
.
t
f
.
o
n
1
8
M
a
y
2
0
2
1
90
Journal of Cognitive Neuroscience
Volume 19, Number 1