Hemispheric Differences in Frontal and Parietal
Influences on Human Occipital Cortex: Direct
Confirmation with Concurrent TMS–fMRI
Christian C. 拉夫, Felix Blankenburg, Otto Bjoertomt, Sven Bestmann,
Nikolaus Weiskopf, and Jon Driver
D
哦
w
n
我
哦
A
d
e
d
抽象的
& We used concurrent TMS–fMRI to test directly for hemi-
spheric differences in causal
influences of the right or left
fronto-parietal cortex on activity (BOLD signal) in the human
occipital cortex. Clinical data and some behavioral TMS studies
have been taken to suggest right-hemisphere specialization for
top–down modulation of vision in humans, based on deficits
such as spatial neglect or extinction in lesioned patients, 或者
findings that TMS to right (与. 左边) fronto-parietal structures
can elicit stronger effects on visual performance. But prior to
the recent advent of concurrent TMS and neuroimaging, 它
was not possible to directly examine the causal impact of one
(stimulated) brain region upon others in humans. Here we
stimulated the frontal or intraparietal cortex in the left or right
hemisphere with TMS, inside an MR scanner, while measuring
with fMRI any resulting BOLD signal changes in visual areas
V1–V4 and V5/MT+. For both frontal and parietal stimulation,
we found clear differences between effects of right- versus left-
hemisphere TMS on activity in the visual cortex, with all dif-
ferences significant in direct statistical comparisons. Frontal
TMS over either hemisphere elicited similar BOLD decreases
for central visual field representations in V1–V4, but only right
frontal TMS led to BOLD increases for peripheral field repre-
sentations in these regions. Hemispheric differences for effects
of parietal TMS were even more marked: Right parietal TMS
led to strong BOLD changes in V1–V4 and V5/MT+, but left
parietal TMS did not. These data directly confirm that the
human frontal and parietal cortex show right-hemisphere spe-
cialization for causal influences on the visual cortex. &
介绍
It is increasingly recognized that visual processing within
the occipital cortex may be influenced by areas out-
side the conventional visual system. 例如, 这是
often proposed that a putative fronto-parietal ‘‘atten-
tion network’’ may modulate visual processing via back-
projection influences (拉夫 & Driver, 2006; Serences &
Yantis, 2006; 磨坊主 & D’Esposito, 2005; Driver, Eimer,
Macaluso, & van Velzen, 2004; Driver, Vuilleumier, &
Husain, 2004; 科尔贝塔 & 舒尔曼, 2002; Frith, 2001;
Hopfinger, Buonocore, & Mangun, 2000; Kastner &
Ungerleider, 2000; 磨坊主, 2000; Duncan, Humphreys,
& Ward, 1997). This might potentially explain some clin-
ical phenomena in humans, whereby lesions in frontal
and/or parietal areas, well beyond the classical ‘‘visual’’
cortex, can lead to putatively attentional deficits that af-
fect visual abilities, such as neglect or extinction (Milner
& McIntosh, 2005; Mort et al., 2003; Karnath, Milner, &
Vallar, 2002; Vuilleumier & Rafal, 2000; Mesulam, 1999;
Driver & Mattingley, 1998). Such clinical phenomena
are typically more common and pronounced after right-
than left-hemisphere damage. This has been taken to
伦敦大学学院, 英国
suggest that the right frontal and parietal cortex might
normally play special roles in influencing activity in the
visual cortex for humans (例如, see Deco & Zihl, 2004;
Marzi, Girelli, Natale, & Miniussi, 2001; Marzi, Girelli,
Miniussi, Smania, & Maravita, 2000; Mesulam, 1999, 但
see also Barcelo´, Suwazono, & 骑士, 2000), that may
differ from any influences from the left frontal or parie-
tal cortex. 出奇, 然而, this has never been
shown directly to date, due to a lack of methods for
studying and comparing the causal impact of any par-
ticular brain area upon others in humans. 虽然
extensive neuroimaging work on human visual atten-
tion has triggered considerable discussion about whether
right-hemisphere frontal–parietal structures may be
qualitatively or quantitatively special, when compared to
left-hemisphere homologues in attention tasks (例如,
see Serences & Yantis, 2006; Driver, Eimer, 等人。, 2004;
Driver, Vuilleumier, 等人。, 2004; 科尔贝塔 & 舒尔曼,
2002; Kastner & Ungerleider, 2000, for reviews), 标准
neuroimaging studies are not sufficient on their own to
address hemispheric differences in truly causal impacts
upon the visual cortex. 反过来, although stimulation
methods such as transcranial magnetic stimulation (TMS;
见下文) are causal interventions, 直到最近, it has
not been possible to combine such brain stimulation with
D 2008 麻省理工学院
认知神经科学杂志 21:6, PP. 1146–1161
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
1
我
6
e
–
1
p
1
d
4
F
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
哦
4
C
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
0
9
0
7
8
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
.
.
F
.
/
t
哦
n
1
8
中号
A
y
2
0
2
1
neuroimaging in humans in order to study the impact of
stimulating one area upon activity in others.
Recent advances in invasive animal work now allow
for microstimulation (or other manipulations) of a tar-
geted region to be combined with recordings from
another interconnected area, as for example in the pio-
neering work of Armstrong and Moore (2007) 和
Moore and Armstrong (2003), who studied influences
from the frontal eye field (FEF) upon monkey V4 (看
also Winkowski & Knudsen, 2007, for a potentially re-
lated barn–owl study). But animals may not show the
hemispheric asymmetries in function for the frontal and
parietal cortex that have tentatively been attributed to
humans based on clinical evidence (例如, see Wardak, Ibos,
Duhamel, & 奥利维尔, 2006; Wardak, 奥利维尔, & Duhamel,
2004). 而且, 据我们所知, the monkey stud-
ies examining frontal influences on the visual cortex, 到
日期, have typically assessed only the right hemisphere
(Armstrong & 摩尔, 2007; 摩尔 & Armstrong, 2003;
摩尔, personal communication).
因此, here we capitalized on the recent devel-
opment of concurrent TMS–fMRI in humans (例如, 拉夫
等人。, 2006; Baudewig et al., 2001; Bohning et al., 1999;
Shastri, 乔治, & Bohning, 1999), using this as a novel
approach for probing directly whether the right versus
left human frontal (or intraparietal) cortex can have quali-
tatively different causal influences on BOLD signal in the
occipital visual cortex. Intriguingly, several purely behav-
ioral TMS studies already suggest possible hemispheric
differences in the impact of frontal or parietal TMS on
visual processing, typically finding that right frontal or
parietal TMS can have more marked influences on visual
performance than TMS of corresponding left-hemisphere
站点 (例如, Muggleton et al., 2006; Silvanto, Lavie, & Walsh,
2006; Chambers, 佩恩, Stokes, & Mattingley, 2004;
O’Shea, Muggleton, Cowey, & Walsh, 2004; Grosbras &
Paus, 2002, 2003; Muggleton, Juan, Cowey, & Walsh, 2003;
Pourtois, Vandermeeren, 奥利维尔, & de Gelder, 2001).
然而, such purely behavioral TMS effects leave it un-
clear whether the observed differences between left and
right TMS reflect only local processing in the stimulated
区域, or rather the differential physiological
impact of
right versus left frontal or parietal regions in inducing
causal changes in activity of remote visual cortex.
To address this, here we used TMS in combination
with concurrent fMRI of retinotopic visual cortex, 到
characterize any physiological differences between the
impact of right versus left frontal (or parietal) TMS upon
BOLD activity in early visual cortex. We had recently
introduced this combined TMS–fMRI approach, but had
stimulated only right-hemisphere sites (Ruff et al., 2006,
2008; see also Taylor, 高贵, & 拉什沃斯, 2007; Paus
等人。, 1997, for related uses of TMS in combination with
other neuroimaging methods in humans). Here we ran
two new experiments in which we applied the analogous
stimulation protocol in the same participants, but now
applying TMS to corresponding sites in the left hemi-
sphere instead. This allowed us to directly compare, 为了
the first time, the on-line causal effects of left- 相对
right-hemisphere frontal or parietal TMS upon BOLD
activity in retinotopically mapped human occipital cor-
tex. To anticipate the outcome, we observed profound
differences in these influences, indicating specific neural
mechanisms in right-hemisphere fronto-parietal areas for
modulation of the visual cortex. We found that the im-
pact of left frontal (or parietal) TMS upon BOLD signal
in the visual cortex differed significantly from corre-
sponding effects of right fronto-parietal TMS. Our data
may thus offer a new type of explanation, in terms of
remote physiological effects upon the visual cortex, 为了
why TMS (or lesions) to the right-hemisphere fronto-
parietal cortex can often affect performance for visual
tasks in a different (and typically more pronounced) fash-
ion than corresponding left-hemisphere interventions.
方法
Experimental Rationale
In the two new experiments described here, we exam-
ined any influences of left frontal or left parietal TMS on
activity in multiple striate and extrastriate visual areas
of the human brain, as measured concurrently via the
BOLD signal with fMRI. 至关重要的是, these new data al-
lowed direct, well-matched statistical comparisons to
existing data on influences of TMS to corresponding
frontal and parietal sites in the right cortical hemisphere
(originally reported in Ruff et al., 2006, 2008; 见下文).
This allowed us to compare the impact of left or right
frontal/parietal TMS upon the human visual cortex for
the first time. Inside an MR scanner, TMS was applied
at four different intensities over either the left frontal
cortex (at the putative location of the human FEF) 或者
over the left parietal cortex (intraparietal sulcus, IPS).
We concurrently measured with fMRI any BOLD changes
in the occipital visual cortex that covaried with TMS
intensity. Sensitivity for early visual regions (areas V1–
V4 and V5/MT+) was maximized by using fMRI with
an occipital surface coil, in combination with detailed
retinotopic mapping of cortical visual areas for each
individual participant. This allowed us to assess whether
V1 and other retinotopic areas of the human visual
cortex could be affected by TMS stimulation of the
left frontal or left parietal cortex, and to characterize
the retinotopic profile for any such effects. 而且,
we could now formally compare any such effects upon
the visual cortex elicited by stimulation of either left-
hemisphere site against the effects when stimulating
their right-hemisphere homologues (as initially de-
scribed in Ruff et al., 2006, 2008), as the same stimula-
tion protocol was employed in the same participants,
with only the TMS site being varied. 最后, we also
used data acquired for TMS to a vertex control site that
should not be expected to affect activity in the visual
Ruff et al.
1147
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
1
我
6
e
–
1
p
1
d
4
F
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
哦
4
C
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
0
9
0
7
8
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
.
t
F
.
/
.
哦
n
1
8
中号
A
y
2
0
2
1
cortex, except via the potential nonspecific effects of
TMS administration per se (such as the ‘‘clicking’’ sound
or the scalp sensation associated with TMS pulses). 在-
契据, TMS to the vertex was found to have no effect on
activity in the visual cortex (see also Ruff et al., 2006),
which therefore allowed us to directly subtract out any
nonspecific TMS effects (auditory activations, ETC。) 什么时候
considering the results for each ‘‘active’’ TMS site by
本身. For comparison between different ‘‘active’’ TMS
站点, any nonspecific effects should be subtracted out in
any case.
In all experiments, participants had to fixate centrally,
with no other task during scanning, to ensure that any
remote physiological influences of TMS upon activity in
the visual cortex could not possibly be contaminated by
TMS-induced changes in behavior. We administered
TMS either while subjects passively viewed a blank dis-
玩, or while they were presented with bilateral moving/
changing visual stimuli designed to activate many visual
地区 (see Figure 1B, C). This was done to allow a test
for whether any TMS influences on activity in the visual
cortex (as measured via the BOLD signal) might depend
on the level of bottom–up activation via visual inputs.
参加者
The same four male, right-handed participants (age =
26–35 years) as in our previous studies (Ruff et al., 2006,
2008) took part in the present experiments. They had
normal vision, good health, and no history of neurolog-
ical or psychiatric illness. All gave written informed con-
sent in accord with local ethics.
TMS Sites
We used the Brainsight Frameless Stereotaxy System
(Rogue Research, 蒙特利尔, 加拿大), together with in-
dividual T1-weighted anatomical MR images, to deter-
mine the scalp coordinates for placing the TMS probe
over the different stimulation sites. The same strategies
as for our previous studies (Ruff et al., 2006, 2008) 是
used to determine the individual stimulation sites. 这
coordinates for left frontal stimulation (over putative
human FEF) were determined on the basis of anatomical
criteria (Blanke et al., 2000; Tehovnik, 索默, 周,
Slocum, & Schiller, 2000) in conjunction with activa-
tions during a 5-min fMRI session of interleaved rest and
auditorily paced voluntary saccades in total darkness.
This strategy resulted in a chosen cortical surface site
for the left FEF with mean Montreal Neurological In-
研究所 (MNI) coordinates x, y, z = (西德:1)27, (西德:1)1, 57 (斯坦-
dard errors: 0.72, 0.53, 2.44), corresponding well with
TMS coordinates used in other human FEF studies
(例如, Grosbras, Laird, & Paus, 2005; O’Shea et al.,
2004; Muggleton et al., 2003; Ro, Cheifet, Ingle, Shoup,
& Rafal, 1999; Paus, 1996). For the left IPS TMS site, 我们
adopted a normalized MNI coordinate (X, y, z = (西德:1)36,
(西德:1)48, 45) based on the mean coordinates of published
activation peaks in the IPS during covert shifts of atten-
tion and/or eye movement planning and execution (taken
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
1
我
6
e
–
1
p
1
d
4
F
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
哦
4
C
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
0
9
0
7
8
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
F
.
t
.
.
/
哦
n
1
8
中号
A
y
2
0
2
1
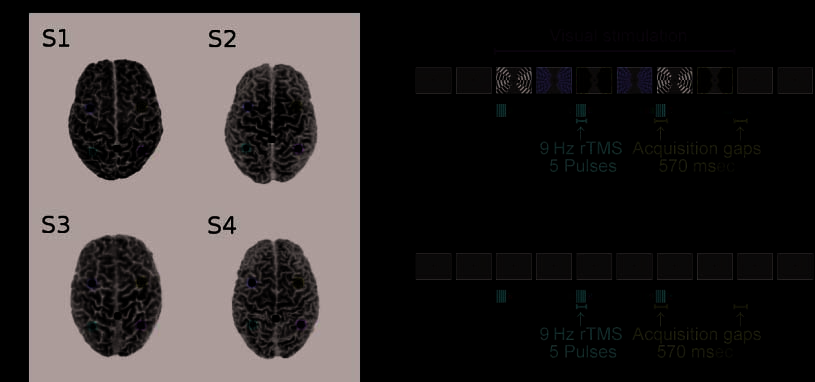
数字 1. TMS sites in individual participants and experimental design. (A) Three-dimensional images of the individual structural scans of our
参与者 (all brains viewed from above; S1 = Subject 1, ETC。). Colored dots mark the position of the five TMS sites studied here, 包括
the two new left-hemisphere sites (yellow for left FEF, red for left IPS) and three sites from previous work by Ruff et al. (2006, 2008; 蓝色的
for right FEF, green for right IPS, and white for the vertex control site). (B and C) A schematic time course (with time running left to right,
and successive rectangles indicating successive screen displays) of a single block of interleaved TMS–fMRI: (乙) with visual stimuli on the
screen during TMS, 或者 (C) without visual stimuli other than the constant central fixation point on a blank gray screen. For each block, 三
TMS trains were delivered in the 570-msec gaps between the acquisitions of subsequent image volumes, at one of the four intensities used
(参见方法). Seven rest scans were included between successive blocks. Visual stimuli (when present, as in six of the illustration panels for B)
remained visible during all three TMS trains and during the acquisition of the three image volumes following the TMS trains.
1148
认知神经科学杂志
体积 21, 数字 6
from Brown et al., 2004; Curtis, 饶, & D’Esposito, 2004;
Connolly, Goodale, Menon, & Munoz, 2002; Connolly,
Goodale, DeSouza, Menon, & Vilis, 2000; Perry & Zeki,
2000; Corbetta et al., 1998). Perhaps the most important
point is that the selection procedure for right and left
homologue sites was thus equivalent, allowing for mean-
ingful comparisons between the effects arising in the vi-
sual cortex when stimulating these sites.
fMRI Procedures
The experiments described here used the same setup,
scanners, and fMRI sequences as our other recent TMS–
fMRI studies (Ruff et al., 2006, 2008), to enable the novel
direct statistical comparison of the impact of left frontal
or parietal TMS against the impact of TMS to comparable
right-hemisphere sites, for activity in the visual cortex
as assessed with fMRI. A 3-T head scanner (Magnetom
Allegra, Siemens Medical, Erlangen, 德国) was used
to acquire T1-weighted structural anatomical images and
the fMRI data used for retinotopic mapping of visual
地区. A 1.5-T whole-body scanner (Magnetom Sonata,
Siemens Medical) was used for acquisition of the sac-
cade localizers (with the standard Siemens CP head coil)
and of the functional data for the critical TMS sessions.
For the TMS sessions, we employed a custom-built vi-
sual surface MR coil (Nova Medical, 波士顿, 嘛, 美国)
with maximum sensitivity over the occipital cortices, 作为
the questions we sought to address here all concerned
possible activity changes in the visual cortex.
All experimental TMS datasets were acquired with
an identical multislice gradient-echo EPI sequence (27
oblique axial slices, 64 (西德:2) 64 矩阵, in-plane resolution:
3 (西德:2) 3 mm2, 2.5 mm slice thickness, 1.25 mm spatial
gap between adjacent slices, = 50 毫秒, slice TR
90 毫秒, 2298 Hz/pixel receiver bandwidth, echo spac-
ing = 500 Asec). A 570-msec gap (see Figure 1B, C) 曾是
included between the acquisitions of subsequent vol-
umes to allow for enough time to apply TMS pulses
within the scanner during this gap, without influencing
MR image acquisition. To shift any possible residual
Nyquist ghost in the direct vicinity of the TMS probe
outside the brain image, 50% oversampling was imple-
mented in the phase encoding direction. For each TMS
实验, 606 image volumes were recorded, lasting
30 min and 18 秒.
TMS Procedures
TMS was employed inside the MR scanner using a Magstim
Super Rapid stimulator and custom-built, figure-of-eight,
MR-compatible nonferrous coils (from The MAGSTIM
公司, Dyfed, 英国; same as used in Ruff et al., 2006,
2008). To eliminate potential interference with image
acquisition from RF noise generated by the TMS device,
the stimulator box was housed in an RF-shielded metal
cabinet and connected with the TMS coil through a
custom filter box (The MAGSTIM Company) and fur-
ther ferrite sleeves ( Wuerth Elektronik, Waldenburg,
德国). The stimulator was remotely controlled by
the same MATLAB script that was also used to deliver
the visual stimuli (见下文).
Inside the scanner, the participant’s head was fixed
with a standard vacuum-suction cushion (Siemens Medi-
卡尔). A nonferromagnetic custom holder with several
degrees of freedom in each direction was used to firmly
position the TMS coil tangentially over the left FEF or
left IPS site (see Figure 1A). The initial flow of the in-
duced current was either in posterior–anterior (FEF) 或者
in anterior–posterior (IPS) 方向, but biphasic pulses
were applied in all experiments. On each trial, 三
equal-intensity trains of five TMS pulses (在 9 赫兹, 在-
tensity either at 85%, 70%, 55%, 或者 40% of stimulator
输出) were applied in the 570-msec temporal gap
between acquisitions of three subsequent image vol-
umes. The maximum stimulation intensity (85%) 用过的
during scanning only corresponded to 118% (±14%) 的
resting motor threshold for our subjects when applied
over the motor cortex, due to the custom nonferrous
TMS coil used and the resistive properties of the MR-
compatible connecting cable. We confirmed by piloting,
by visual
inspection during the experiment, 并由
participant report that this TMS protocol did not induce
any muscle twitches, as expected given the TMS sites
涉及. Six stimulation trials were administered for
each of the eight conditions (four TMS intensity levels,
each with peripheral visual stimulation present or absent)
at each TMS site. Our protocol thus contained a total of
48 TMS stimulation blocks (720 pulses in total) per TMS
site, complying with published safety limits for repetitive
TMS (Wassermann, 1998). Each experiment also con-
泰内德 12 control trials without any TMS, 在。。。期间
visual stimuli could be present or absent also. All trials
were separated by a constant intertrial interval (ITI) 的
seven image volumes without any stimulation. The order
of conditions for each TMS site was randomly deter-
mined by the program used to deliver all experimen-
tal stimulation. This program was implemented in the
MATLAB (The Mathworks, Natick, 嘛) stimulus presenta-
tion toolbox COGENT (www.vislab.ucl.ac.uk/Cogent2000/
index.html).
Visual Stimulation and Eye Tracking
On half of the trials, we projected dynamic visual pat-
terns onto a frosted screen (308 (西德:2) 228 visual angle, gray
background, 0.58 (西德:2) 0.58 central fixation cross always
展示) mounted at the rear end of the scanner bore.
Participants viewed this screen via a mirror system sitting
on top of the MR surface coil. As in Ruff et al. (2006,
2008), the stimuli were patterns that spared the fovea
and the vertical meridian, that randomly changed form
and color every 500 毫秒, and that randomly moved on
Ruff et al.
1149
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
1
我
6
e
–
1
p
1
d
4
F
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
哦
4
C
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
0
9
0
7
8
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
.
t
/
F
.
.
哦
n
1
8
中号
A
y
2
0
2
1
each frame (whole-pattern movement, maximum trans-
lation in both horizontal and vertical direction of 0.38
per 16-msec frame). These patterns, when present, 是
visible on the screen throughout the three TMS trains
per trial and the associated consecutive MR image vol-
umes (see Figure 1B). This manipulation of visual stim-
计算 (present or absent) was implemented to assess
whether any influences of frontal or parietal TMS upon
activity in the visual cortex would depend upon current
visual context.
To confirm whether participants kept fixation through-
out the experiment, we measured eye position, pupil
diameter, and any blinks at 60 Hz during scanning with
an ASL 504 remote optics infrared eye tracker (Applied
Science Laboratories, Bedford, 嘛). Eye signals were
obtained with the same mirror used for visual stimu-
lus viewing. Raw eye position data were filtered for
blinks (continuous losses of pupil signal for more than
80 毫秒) and transformed to degrees of visual angle be-
fore analysis.
Image Processing and Analyses
Data from the left FEF and left IPS experiments under-
went exactly the same SPM2 (www.fil.ion.ucl.ac.uk/spm)
analyses as the data for the right-hemisphere sites. Func-
tional images were reconstructed off-line, and the first
six images of each run were discarded to account for T1
equilibration effects. Images were realigned to the first
of the series and corrected for movement-induced im-
age distortions (安德森, Hutton, Ashburner, 车工,
& 弗里斯顿, 2001). Any slices containing TMS capacitor-
induced artifacts (少于 1%) were identified as out-
lier changes in the slice signal by more than 3 SD of the
mean slice difference in the time series between con-
secutive volumes, and were replaced by the mean of
the spatially equivalent slices from the previous and the
subsequent image volume. For analyses in stereotactic
空间 (which were further complemented by individual
retinotopic analyses, 见下文), images were normal-
ized to the MNI anatomical standard space and spa-
tially smoothed with a three-dimensional 6-mm FWHM
Gaussian kernel, in accord with the SPM approach.
Voxelwise effects of each experimental condition per
TMS site were estimated by multiple regression of the
voxel time series onto a composite model with 10 共-
variates of interest per session (four TMS stimulation
intensities and no TMS, each with and without visual
stimulation). These covariates were derived by convolv-
ing appropriately placed series of delta functions with
the canonical hemodynamic response function em-
ployed in SPM2. The model additionally contained one
regressor representing eye blinks and another regressor
for mean pupil diameter per scan. This multiple regres-
sion approach ensured that any variance in brain activity
shared by two regressors (例如, activity that might cor-
relate with both TMS intensity and eye blinks) was not
included in our fMRI results (Friston et al., 1995). 这
model removed low-frequency drifts and short-term
temporal autocorrelation of scans by means of a high-
pass filter (128 sec cutoff ) and an AR(1) 过程, 关于-
spectively (Friston et al., 2002). After model estimation,
linear contrasts were used to assess and compare the
effects associated with the different experimental con-
版本. Correlations of BOLD with TMS intensity were
modeled as the corresponding weighted linear combi-
nation of the four covariates representing the different
TMS intensities. For all analyses, the statistical threshold
was set to t > 3 and a cluster threshold of p < .05,
corrected for multiple comparisons across the image
volume. All reported peak voxel coordinates correspond
to the MNI space employed in SPM2.
In addition to standard SPM group analyses in stereo-
tactic space, individually defined retinotopic visual areas
V1–V4 and area V5/MT+ were analyzed for TMS-induced
activity changes. For all these analyses, mean BOLD
signal estimates during the different conditions were ex-
tracted from the individually defined regions (see be-
low) in the same fashion for each TMS site, and were
directly compared by means of repeated measures
ANOVAs and subsequent paired t tests for planned com-
parisons. This repeated measures approach was ap-
propriate given our use of the same subjects in each
experiment for all conditions. Moreover, although differ-
ent TMS sites were used in different scanning sessions,
we did not just compare overall session effects per se,
but rather the effects of specific manipulations within
sessions (TMS intensity, crossed with presence and ab-
sence of visual stimuli) for the different TMS sites.
Retinotopic areas V1–V4 were determined for each par-
ticipant individually by a standard retinotopic meridian
mapping localizer, consisting of a 5-min fMRI session of
subjects viewing flickering checkerboards presented in
an alternating fashion either along the horizontal or ver-
tical meridian. The unsmoothed data from this session
were modeled voxelwise using a general linear model
that included the two meridian conditions. The borders
of visual areas V1–V4 (Sereno et al., 1995) were then
plotted onto cortical flatmaps derived by segmentation
and cortical flattening in MrGray ( Wandell, Chial, &
Backus, 2000; Teo, Sapiro, & Wandell, 1997). These flat-
maps and region definitions were used to inspect the
SPM(t)s quantifying the correlation of TMS intensity
and BOLD signal from the main experiments. For these
retinotopic analyses of the impact from each active TMS
site, we always directly controlled for any possible non-
specific effects of TMS (e.g., due to the ‘‘click-sound’’
or scalp sensation associated with TMS pulses) on early
retinotopic visual cortex. This was done by subtracting
out the (null) effects of TMS to the vertex control site
when characterizing the activation patterns for each of
the two new active sites of interest in isolation; or by
directly comparing effects for two active TMS sites.
1150
Journal of Cognitive Neuroscience
Volume 21, Number 6
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
6
e
-
1
p
1
d
4
f
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
o
4
c
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
f
.
t
.
o
n
1
8
M
a
y
2
0
2
1
As for our previous studies, which had found system-
atically different effects of right FEF TMS upon repre-
sentations of the central versus peripheral visual field in
early visual cortex, the V1–V4 data were analyzed for
TMS effects in representations of different visual eccen-
tricity. Each area was divided into four different eccen-
tricity ‘‘sectors,’’ moving progressively from more to less
foveal (see also Ruff et al., 2006, 2008; Schwartz et al.,
2005). The correlation of BOLD signal with TMS inten-
sity was quantified as t value in relation to voxelwise
noise, and averaged across the voxels contained in each
sector. This statistic-based approach ensured that TMS-
induced effects could be compared across different eccen-
tricity sectors, and different experiments, without being
confounded by voxel- or session-specific noise. Moreover,
averaging TMS effects across all voxels for particular
eccentricity sectors of the retinotopic cortex (rather than
the less conservative strategy of selecting the peak voxels
displaying the maximum effects), allowed us to compare
effects between sectors, regions, and experiments in a
spatially unbiased manner.
Visual area V5/MT+ was defined in each participant
by means of a separate 5-min fMRI session with alter-
nating presentations of moving or static starfields. These
stimuli spared the fovea by 28 to each side. A voxelwise
general linear model with two conditions was applied to
the unsmoothed data to determine the cortical region
in the lateral occipital cortex maximally driven by mov-
ing relative to static starfield stimuli, corresponding to
the putative anatomical location of V5/MT+ (see e.g.,
Rees, Friston, & Koch, 2000; Watson et al., 1993). TMS
intensity-dependent effects in this region during the
main experiment were then assessed by extracting mean
signals per condition (SPM betas scaled for each voxel
as percent of the session mean) from spherical regions
of interest (V5/MT+ ROIs, 6 mm radius) centered at
the individual peak of activations elicited by the mo-
tion localizer in this area. We compared the two highest
TMS intensities (85% and 70% total output) versus the
two lowest (55% and 40% total output) when con-
sidering each specific visual area, doing so separately
for trials with and without visual stimuli present on the
screen.
RESULTS
We compared effects of TMS over the left versus right
frontal sites, or left versus right parietal sites, in two
complementary sets of analyses. Initially, we used a stan-
dard group-analysis approach for each of the four active
TMS sites to identify regions in stereotactic space that
showed systematic relationships of BOLD with TMS in-
tensity. The particular patterns of influence for left- or
right-hemisphere TMS sites upon specific visual areas
were then characterized in more detail, and compared
directly, by means of individual retinotopic analyses in
conjunction with cortical flattening for V1–V4, and via
ROI analysis for V5/MT+.
Frontal TMS: Group Analyses in Stereotactic Space
We had previously observed (Ruff et al., 2006) that fron-
tal TMS over the right FEF leads to systematic activity
increases in the bilateral cuneus (representing the pe-
ripheral visual field), but leads to bilateral activity de-
creases instead in the occipital poles (representing
the central visual field). We now performed the corre-
sponding group analysis for the new left frontal TMS
data. This revealed very similar activity decreases in the
occipital poles of both hemispheres (Figure 2A) as a
function of increasing intensity of TMS administered
over the left FEF. The locations of these TMS-elicited
activity decreases were virtually identical to those pre-
viously found for the right frontal TMS experiment (see
lower part of Figure 2 for overlay of the regions found
in both experiments). However, we now did not find
any region that showed activity increases as a function
of increasing intensity of TMS over the left FEF, in con-
trast to the strong activations found for this comparison
in the right FEF TMS dataset (see Figure 2B). Finally, the
effects of TMS within either frontal TMS experiment
were very similar when visual stimuli were present or
absent; no regions were found that displayed any in-
teraction of frontal TMS intensity with visual stimulus
presence/absence. These apparent similarities (activity
decreases for the occipital poles/central visual field; and
independence of concurrent visual stimulation) and
differences (activity increases for the cuneus regions/
peripheral visual field only after right frontal TMS) in
the effects of left versus right frontal TMS were con-
firmed and further specified in the individual retinotopic
analyses described below.
Frontal TMS: Individual Analyses of Specific
Visual Areas
We further characterized the spatial topography of left
versus right frontal TMS effects on specific visual areas
by means of individual analyses. To this end, we created
flattened representations of each participant’s visual
cortices, determined the borders of visual areas V1–V4
on these flatmaps by means of standard retinotopy pro-
cedures, and divided each of these areas into four
eccentricity sectors coding the central through to more
peripheral eccentricities in the visual field (see Meth-
ods). For each region, we could then derive the inter-
subject mean correlation of BOLD with TMS intensity
in each of these eccentricity sectors, to directly charac-
terize and compare the spatial topography of effects of
left versus right FEF TMS.
Figure 3A shows the effects of TMS to the left FEF (left
histogram) or the right FEF (right histogram) on BOLD
Ruff et al.
1151
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
6
e
-
1
p
1
d
4
f
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
o
4
c
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
.
t
.
f
o
n
1
8
M
a
y
2
0
2
1
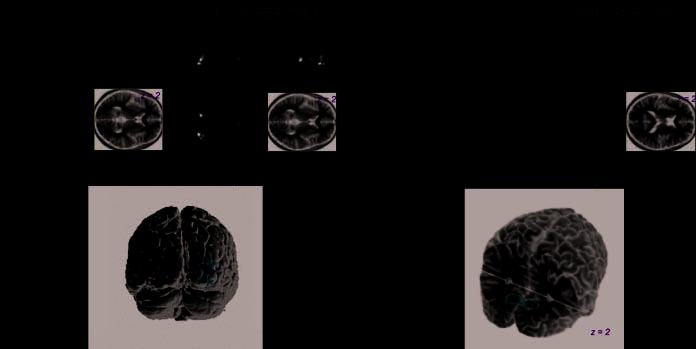
Figure 2. Group analyses of
frontal TMS data: Occipital
regions displaying activity
changes correlating with
intensity of TMS over the left
or right FEF. (A) All occipital
regions that displayed reliable
BOLD signal decreases with
increased intensity of frontal
TMS over the left or right FEF.
All upper panels in (A) are
SPM(t) images corresponding
to the negative correlation
of BOLD signal with the
intensity of TMS, rendered
either on sagittal, coronal,
or transverse views of a
transparent version (so that
no effects are obscured) of
the MNI brain template, or onto a transversal slice of the template structural image. The threshold is set to t = 3 and a cluster-level p < .05,
corrected across the brain volume. In the bottom panel, a rendering of these SPM(t)s onto a standard 3-D brain template shows the clear
similarity between the effects of left and right frontal TMS: Both lead to bilateral BOLD signal decreases at the occipital poles as TMS intensity
increases (shown in green for left frontal TMS, in red for right frontal, and in yellow for overlapping BOLD effects from either frontal TMS site).
(B) The outcome when testing instead for BOLD signal increases with increased intensity of frontal TMS over the left or right FEF. Whereas
the medial occipital cortex (representing the peripheral visual field) showed such an inf luence during right frontal TMS over the FEF (shown
in the cutaway 3-D rendering on a standard brain template at bottom of B), there were no such effects of corresponding left frontal TMS.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
6
e
-
1
p
1
d
4
f
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
o
4
c
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
.
.
.
t
f
o
n
1
8
M
a
y
2
0
2
1
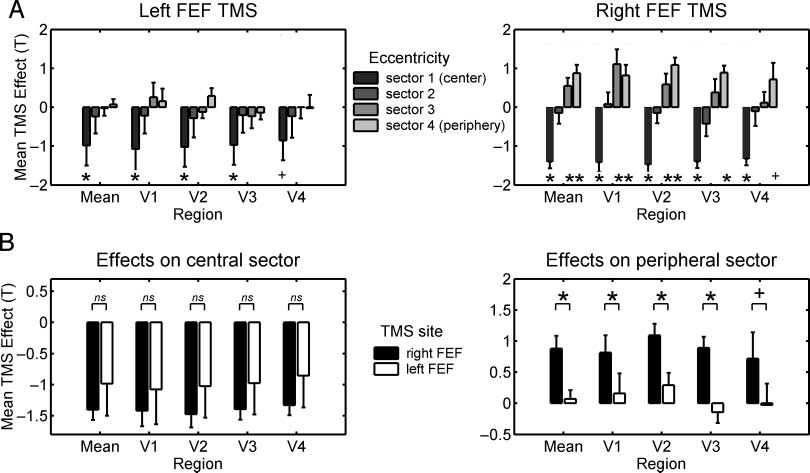
Figure 3. Retinotopic analyses of areas V1–V4: Higher intensity of left or right frontal TMS elicits similar BOLD decreases for the central visual
field, but only right frontal TMS leads to BOLD increases for the peripheral visual field. (A) The pattern of effects of TMS to the left or right
FEF upon BOLD signal in each of four different eccentricity sectors within visual areas V1–V4. See main text for how the eccentricity sectors
were derived, but note that the first along the x-axis for each visual area in A corresponds to the representation of the central visual field, with
the next three successive sectors (further to the right along the x-axis, for each visual area) corresponding to increasingly eccentric visual field
representations. The correlation of TMS intensity with BOLD (quantified as t value, as in Ruff et al., 2006, 2008) was averaged across flatmaps
and voxels within each eccentricity sector of areas V1–V4. These average correlations (±SEM ) are displayed here either averaged across visual
areas V1–V4 (‘‘mean’’; shown in leftmost group of bars of each histogram in A), or separately for each area V1 through to V4, pooling across
the dorsal and ventral cuneus. In all these retinotopic visual areas, increased TMS intensity over the right FEF produced activity increases for
the peripheral sectors and activity decreases for the central sector, whereas left FEF TMS only elicited activity decreases for the central sector
in all visual areas (*p < .05, +p < .1 in simple t tests). (B) Direct comparisons of the TMS effects of either frontal stimulation site on the most
central sector (left histogram) or most peripheral sector (right histogram). These plots confirm for each visual area that the TMS-induced
activity decreases in the most central sector did not differ for left versus right frontal TMS (ns = not significant), whereas the TMS-induced
activity increases in the peripheral sector were consistently stronger during right than left frontal TMS (*p < .05, +p < .1 in paired t tests).
1152
Journal of Cognitive Neuroscience
Volume 21, Number 6
signal
in each eccentricity sector (four sectors, going
from most central to most peripheral) of visual re-
gions V1–V4, plotted as t values to convey the robust-
ness of all effects relative to noise. Increased intensity of
either left or right frontal TMS elicited similar activity
decreases for representations of the central visual field
(darkest bars in the histograms of Figure 3A), in good
accord with the similar activity decreases that had been
observed for the occipital poles in the group stereotactic
analyses (cf. Figure 2A). However, strong (and signifi-
cant) differences in the effects of left versus right FEF
TMS were observed for activity changes in peripheral vi-
sual field representations: Whereas right frontal TMS led
to reliable activity increases in these sectors for all early
retinotopic visual areas (see pale bars in right histogram
of Figure 3A), no reliable effects on BOLD signal were
observed in these sectors for the left frontal TMS data
(see left histogram in Figure 3A). Moreover, this was not
simply a null result, as the impact of left frontal versus
right frontal TMS on peripheral visual field representa-
tions for V1–V4 differed significantly between the two
TMS sites (see below).
This pattern of some similarities (activity decreases for
the central visual field), but also some differences (con-
cerning activity increases in representations of the pe-
ripheral visual field), for effects of left versus right frontal
TMS was confirmed in direct statistical comparisons. We
calculated a 2 (TMS over left vs. right FEF) (cid:2) 2 (most
central vs. most peripheral sector) (cid:2) 2 (visual stimuli
absent vs. present) repeated measures ANOVA on the
TMS-intensity effects (correlations of BOLD with TMS
intensity, quantified as t value), pooled across cortical
hemisphere and the dorsal and ventral parts of V1–V4
(there were no reliable differences due to those factors).
This analysis showed a significant main effect of eccen-
tricity sector [F(1, 56) = 35.6, p < .000001] and a signifi-
cant interaction of eccentricity sector with stimulation
site [F(1, 56) = 7.41, p < .01], arising because left or
right frontal TMS-intensity effects differed for peripheral
but not for central visual field representations (see be-
low). The factor of visual stimulation did not modulate
the impact of frontal TMS intensity (all p > .05). 这
confirms the initial findings from the group SPM analy-
ses above, which had also shown that the influences of
left and right FEF TMS were each similar during either
the presence or absence of concurrent visual input. 笔记
that a similar pattern was found when each retinotopic
visual area was considered separately.
Figure 3B displays TMS effects for the most central
and most peripheral eccentricity sector in detail for each
retinotopic visual area, and marks the significance of
planned comparisons between effects of the two fron-
tal stimulation sites (IE。, over the left vs. right FEF, 现在
plotted as white or black bars, 分别). These plots
show that in each retinotopic visual area, the activity
decreases in the eccentricity sector representing the cen-
tral visual field (left histogram in Figure 3B) were com-
parable in magnitude for left or right frontal TMS. 经过
对比, the activity increases observed in the most pe-
ripheral sector, as a consequence of right frontal TMS,
were significantly stronger than during left frontal TMS in
each visual area (albeit only at trend level for V4; see right
histogram in Figure 3B).
最后, we also conducted ROI analyses to examine
BOLD signal in visual area V5/MT+ (参见方法), 哪个
we had been found to be unaffected by the intensity of
TMS to the right FEF (see Ruff et al., 2008). The new left
FEF data similarly showed no significant effect of TMS
intensity on BOLD in V5/MT+, neither during the pres-
ence nor absence of concurrent visual stimuli.
Parietal TMS: Group Analyses
We recently reported that right parietal TMS (超过
IPS) led to activity changes in the occipital visual cortex
that differed qualitatively from those due to right fron-
tal TMS (see Ruff et al., 2008). Our new dataset now
allowed a direct comparison of the impact of left versus
right parietal TMS on BOLD signal in the occipital visual
cortex (analogous to the comparison of left vs. 正确的
frontal TMS data presented above). These new analyses
revealed clear hemispheric differences: Whereas right
parietal TMS elicited BOLD signal changes in the visual
cortex that depended strongly on the current visual con-
文本 (IE。, the presence or absence of visual input), 这
new left parietal TMS data showed no influence of left
IPS TMS on the occipital cortex. 重要的, this new
finding was not just a null result, as the patterns found in
the visual cortex for right versus left parietal TMS were
significantly different in direct statistical comparisons.
具体来说, for the right parietal site, TMS-induced
activity increases arose in the medial cuneus only in the
absence of concurrent visual input (see Figure 4A, 蓝色的
line plots with star), whereas activity decreases due to
right parietal TMS were found in the lateral occipital cor-
tex (corresponding to V5/MT+ as confirmed further be-
低的), only when the moving visual stimuli were present
(see Figure 4B, blue line plots with star). In the new left
IPS experiment, 相比之下, we did not find any region
in the recorded image volume that showed systematic
activity changes as a function of TMS intensity (see left
panels in Figure 4A and B), neither during the absence
nor during the presence of the visual stimuli. 同样地,
no region was found to display an interaction of left
parietal TMS intensity with presence versus absence of
visual stimuli.
We confirmed a significant difference in outcome be-
tween left versus right parietal TMS, by direct statistical
comparisons between the two parietal experiments. 为了
individually defined area V5/MT+ (as determined by a
motion localizer; 参见方法), we extracted the mean
BOLD signals in the different experimental conditions
and compared the TMS-intensity effects (two highest
TMS intensities minus two lowest; see Methods and also
Ruff et al.
1153
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
1
我
6
e
–
1
p
1
d
4
F
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
哦
4
C
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
0
9
0
7
8
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
.
.
F
/
t
.
哦
n
1
8
中号
A
y
2
0
2
1
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
F
t
r
哦
p
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
.
H
米
A
我
r
e
.
d
你
C
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
1
我
6
e
–
1
p
1
d
4
F
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
哦
4
C
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
9
e
.
s
t
2
哦
1
n
0
9
0
7
8
.
S
p
e
d
p
F
e
米
乙
y
乙
e
G
r
你
2
0
e
2
s
3
t
/
j
F
.
.
/
t
.
哦
n
1
8
中号
A
y
2
0
2
1
数字 4. Group analyses of parietal TMS data: Right but not left parietal TMS elicits BOLD changes in the visual cortex that depend on
current visual context. (A–B) Two sets of occipital regions where increased intensity of right parietal TMS elicited BOLD effects that depended
on visual context (see also Ruff et al., 2007), whereas no such effects were found for left parietal TMS. The images in both panels show
SPM(t)s (thresholded at t > 3 和 p < .05, cluster-level corrected across the image volume) quantifying (A) positive correlations of BOLD
with parietal TMS intensity, specifically during the absence of visual stimuli; or (B) negative correlations of BOLD with parietal TMS intensity,
now during the presence of visual stimuli. Within both (A) and (B), SPMs for the new left parietal TMS experiment are shown on the left,
whereas the right parietal TMS data are shown on the right. The line plots displayed at the bottom of (A) show the mean signal intensity
(±SEM ) during the different experimental conditions, as extracted from a spherical ROI (6 mm radius) centered in the peak medical cuneus
voxel of the SPM(t) for the right parietal TMS data shown above. Note that significant BOLD decreases ( p < .05 in paired t tests, marked
by a star) during the absence of visual stimuli were only found in this medial occipital region for right parietal TMS, but not for left parietal
TMS (ns = not significant). In both experiments, the medical cuneus was, of course, more active during visual stimulation (red lines) than
in its absence (blue lines), but the TMS effect was only found in the absence of visual stimuli, and only for right parietal TMS (see star). The line
plots displayed at the bottom of (B) show the mean BOLD signal intensity (±SEM ) during left or right parietal TMS in area V5/MT+, as determined
for each subject with an fMRI motion localizer (see Methods, signal is pooled over hemispheres as all effects were symmetric and bilateral).
Increased intensity of right parietal TMS led to significant ( p < .05 in paired t tests, marked by a star) activity decreases in V5/MT+ only
when the moving visual stimuli were present (green line in right plot), whereas no such effects were found for left parietal TMS, with this outcome
differing significantly between the left and right parietal TMS sites (see main text).
Ruff et al., 2008) in a 2 (cid:2) 2 repeated measures ANOVA
(Left or right parietal TMS (cid:2) Visual stimulus present
or absent). A significant interaction [F(1, 31) = 8.05,
p < .01] arose between these two factors because right
parietal TMS elicited activity decreases in V5/MT+ only
during the presence of moving visual stimuli, whereas
left parietal TMS had no impact. This was confirmed
by planned comparisons, which showed significant TMS
effects on V5/MT+ due to right parietal TMS only when
in the presence of visual stimuli [t(1, 7) = 2.59, p < .05],
but no effects for left parietal TMS [t(1, 7) = 0.26, ns; see
Figure 4B, bottom].
Left versus right parietal TMS effects for the early
visual cortex (corresponding to the medial cuneus as
shown in Figure 4A) were considered further in indi-
vidual analyses of retinotopic early visual areas, as de-
scribed below.
Parietal TMS: Retinotopic Analysis
The different impacts of left or right parietal TMS on
the medial occipital cortex (see Figure 4A) were further
characterized by direct statistical comparisons of the
BOLD signal changes in individually mapped retinotopic
visual areas V1–V4. As for the corresponding analyses of
the frontal TMS data (see above), we extracted the mean
TMS effect from each eccentricity sector in each retino-
topic visual area (see Figure 5). This revealed that left
1154
Journal of Cognitive Neuroscience
Volume 21, Number 6
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
6
e
-
1
p
1
d
4
f
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
o
4
c
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
.
/
t
.
o
n
1
8
M
a
y
2
0
2
1
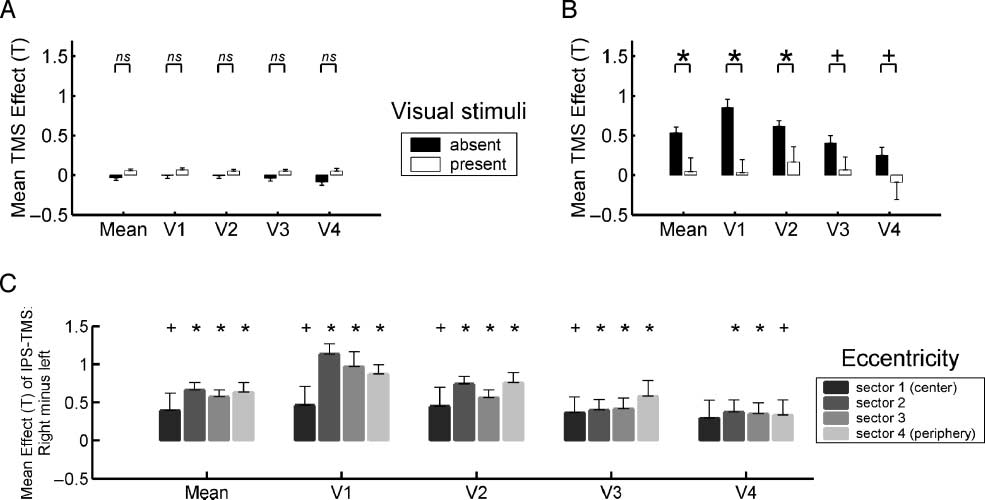
Figure 5. Retinotopic analyses of parietal TMS influences on visual areas V1–V4: Right but not left parietal TMS elicits BOLD increases in
V1–V4 during the absence of visual stimuli, for all eccentricity sectors. The upper panels show the mean effects (±SEM ) of (A) left or (B)
right parietal TMS upon BOLD signal in retinotopic visual areas V1–V4, during the presence (white bars) or absence (black bars) of visual
input. (A) Increased intensity of left parietal TMS did not lead to activity changes in any of these areas, in neither of the two visual conditions.
By contrast, (B) shows that activity increases were found in visual areas V1–V4 for increased intensity of right parietal TMS, but only during
the absence of visual stimuli; these activity increases were stronger than during the presence of visual stimuli (*p < .05, +p < .1 in paired
t tests). (C) The direct comparison of TMS effects for right minus left parietal TMS, separately for the four eccentricity sectors, either averaged
across V1–V4 (‘‘mean,’’ at left of histogram) or separately for each area. See main text for how the eccentricity sectors were derived, but note
that eccentricity sector number 1 (the first along the x-axis for each visual area) corresponds to the representation of the central visual field,
with increasing sector numbers (further to the right along the x-axis, for each visual area) corresponding to increasingly eccentric visual field
representations. The basic findings here are that effects of right parietal TMS on V1–V4 depended on the absence of current visual input,
and were similar for different eccentricity sectors; whereas left parietal TMS differed strikingly in having no effect on areas V1–V4, confirming
a significant difference between the impact of right versus left parietal TMS on the visual cortex.
parietal TMS had no effect on any retinotopic area
(Figure 5A), whereas right parietal TMS affected retino-
topic regions only in the absence of visual stimuli (Fig-
ure 5B). This difference in impact between right parietal
versus left parietal TMS on areas V1–V4 was comparable
for all eccentricity sectors (Figure 5C).
These conclusions were confirmed by directly com-
paring the two parietal experiments in a 2 (left or right
parietal TMS) (cid:2) 2 (most central vs. most peripheral sec-
tor) (cid:2) 2 (visual stimuli present vs. present) repeated
measures ANOVA of the TMS-intensity effects (i.e.,
correlation of BOLD with TMS intensity, quantified by
t values as also in Ruff et al., 2006, 2008). A significant
[F(1, 56) = 7.38, p < .01] interaction between right
versus left parietal TMS and absence versus presence of
visual stimulation indicated that right parietal TMS af-
fected the retinotopic visual cortex in the absence but
not in the presence of visual input, whereas left parietal
TMS had no impact on retinotopic visual areas. This
interaction pattern was found in each retinotopic visual
area [all F(1, 56) > 4.56, 所有p < .05], reflecting activity
increases due to right parietal TMS found only during
the absence of visual stimuli. We confirmed with pairwise
tests that these right parietal TMS-induced increases
were indeed reliably larger during the absence than dur-
ing the presence of visual stimuli (see Figure 5B), and
critically also that left parietal TMS did not induce such
effects on V1–V4 (all p > .88; see Figure 5A).
Figure 5C displays, for every eccentricity sector and
visual area, the difference between right-minus-left pa-
rietal TMS effects during the absence of visual stimuli.
These plots show that, in contrast to the frontal TMS
data considered earlier above, the right parietal TMS
data did not show any significant effect involving the
factor of eccentricity [all F(1, 56) > 2.81], and also that
right parietal TMS effects were reliably larger than the
left parietal TMS effects across all eccentricity sectors.
因此, the data for the new left parietal TMS site do not
just reveal a null result, but instead represent a signifi-
cant difference to the effects of right parietal TMS on the
视觉皮层.
Eye-data Analyses
Eye position, blinks, and pupil diameter were measured
throughout all the fMRI experiments considered here.
Ruff et al.
1155
It seems implausible that any of these factors could
account for our specific fMRI results, given the reliable
differences found here for TMS to homologous regions
in opposite hemispheres, the differences in the specific
retinotopic patterns (in relation to eccentricity sectors),
and the dependence versus independence of current
visual context for the parietal and frontal TMS effects.
We nevertheless took great care to ensure that potential
nonspecific effects of TMS upon eye position, blinks, 或者
pupil dilation could not account for the activity changes
observed in the visual cortex. As in our previous pub-
lications for right-hemisphere TMS (Ruff et al., 2006,
2008), we included blinks and pupil dilations as in-
dependent regressors in all of our statistical models of
the hemodynamic responses changes elicited by TMS.
This ensured that any effect correlating with these eye
variables could not contribute to the reported effects.
而且, we also confirmed with formal statistics that
high-intensity TMS, low-intensity TMS, and no-TMS blocks
did not differ in mean horizontal and vertical eye position
[all F(2, 237) < 1.46], nor in the standard deviation of
those position signals [all F(2, 237) < 1.52], nor in pupil
diameter [both F(2, 237) < 2.8]. We also confirmed that
blinks occurred equally often during trials with high-
intensity, low-intensity, or no TMS [x2(2) = 5.38].
DISCUSSION
Clinical observations of brain-damaged patients (Becker
& Karnath, 2007; Milner & McIntosh, 2005; Mort et al.,
2003; Karnath et al., 2002; Mesulam, 1999) and some
purely behavioral TMS studies of neurologically healthy
people (e.g., Muggleton et al., 2003, 2006; Silvanto
et al., 2006; Chambers et al., 2004; O’Shea et al., 2004;
Grosbras & Paus, 2002, 2003; Pourtois et al., 2001)
have often been taken to suggest that right fronto-
parietal areas may play a more predominant role in visual
function than comparable left-hemisphere structures (al-
though see Barcelo´ et al., 2000). However, such findings
have left it unclear whether any such hemispheric asym-
metries may, at least in part, reflect the specific capacity
of right-hemisphere fronto-parietal structures to exert
remote influences upon the visual cortex, rather than
solely reflecting local processing within frontal–parietal
areas themselves. Moreover, standard fMRI studies of
visual attention have not always agreed on whether the
putative fronto-parietal ‘‘attention network’’ is strongly
right-lateralized in humans, or might be essentially (or
partially) bilateral/symmetric, as often suspected for non-
human primates (e.g., see Serences & Yantis, 2006;
Driver, Eimer, et al., 2004; Driver, Vuilleumier, et al.,
2004; Corbetta & Shulman, 2002; Kastner & Ungerleider,
2000, for reviews). In any case, neuroimaging studies, or
purely behavioral patient studies, on their own typically
cannot identify causal
influences between brain areas,
which was the central issue considered here when asking
whether the impact of right frontal or parietal TMS upon
activity in the visual cortex may differ qualitatively from
that for comparable left frontal or parietal TMS.
To address this directly, we used concurrent TMS–fMRI
to assess with direct statistical comparisons how apply-
ing TMS to the left frontal or parietal cortex may differ
from effects of corresponding right TMS, in terms of the
impact on BOLD activity in human visual areas. We ob-
served marked and statistically significant differences in
these influences, indicating that right-hemisphere fronto-
parietal areas may play special roles in the network that
can modulate the visual cortex. Our data may thus pro-
vide a new possible explanation, in terms of remote phys-
iological effects on the visual cortex, for the frequent
finding that performance in perceptual tasks is more often
affected by TMS to the right (rather than left) fronto-
parietal cortex in humans (e.g., Muggleton et al., 2003,
2006; Silvanto et al., 2006; Chambers et al., 2004; O’Shea
et al., 2004; Grosbras & Paus, 2002, 2003; Pourtois et al.,
2001).
Although increased intensity of TMS over both the left
and right frontal cortex elicited strikingly similar activity
decreases for central visual field representations in V1–
V4, only right frontal TMS led to strong BOLD increases
for peripheral field representations in these regions. This
suggests right-hemisphere predominance for frontal in-
fluences upon (bilateral) visual cortex representations of
the periphery (cf. Mesulam, 1999, for original clinical pro-
posals, based on brain-damaged patients, that some
right-hemisphere structures might deal with both sides
of space). For the parietal TMS sites, we found even
clearer hemispheric differences. Left parietal TMS did
not affect BOLD signal in any area of the occipital visual
cortex, in contrast to right parietal TMS, which led to
BOLD signal changes in V1–V4 and V5/MT+, which were
strongly dependent on current visual context (and thus,
quite unlike the frontal TMS effects). All these differences
in the effects of right- versus left-hemisphere TMS were
significant in direct statistical comparisons, providing new
causal evidence that right-hemisphere frontal and parietal
regions of the human brain can influence activity in the
occipital visual cortex in a more pronounced and distinct
fashion than homologous left-hemisphere fronto-parietal
regions.
The only significant effect of left-hemisphere TMS on
the occipital cortex found here consisted of BOLD signal
decreases for representations of the central visual field
in areas V1–V4 during left frontal stimulation. These par-
ticular effects were remarkably similar for both left- and
right-frontal TMS, in terms of their spatial extent and
magnitude (see Figure 2A). Moreover, all effects of TMS
at both these frontal sites applied regardless of the pres-
ence versus absence of visual input. This latter finding
supports the general notion that frontal influences on
processing in the visual cortex may apply in a ‘‘top–
down’’ fashion (e.g., see Buschman & Miller, 2007,
for a recent example) that can operate regardless of
1156
Journal of Cognitive Neuroscience
Volume 21, Number 6
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
6
e
-
1
p
1
d
4
f
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
o
4
c
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
f
.
.
.
t
o
n
1
8
M
a
y
2
0
2
1
current ‘‘bottom–up’’ visual input. But we also found
clear hemispheric differences in the effects of frontal
TMS on the visual cortex. Only right frontal TMS led to
BOLD increases for more peripheral visual field repre-
sentations in V1–V4 (see Figures 2B and 3), whereas
left frontal TMS did not. From a functional point of view,
this dissociation of common BOLD decreases (for cen-
tral visual field) and distinct BOLD increases (for periph-
eral field only with right frontal TMS) might indicate
possible functional specialization of the right frontal cor-
tex for enhancing processing of the peripheral visual
field. This might potentially relate to the putative role
of the frontal cortex in controlling covert spatial atten-
tion to the periphery (e.g., Juan, Shorter-Jacobi, & Schall,
2004; Tehovnik et al., 2000; Mesulam, 1999), and our
findings clearly suggest some right-hemisphere domi-
nance for such functional contributions in humans.
From an anatomical perspective, our findings suggest
that separable neural tracts may link the human frontal
cortex with occipital representations of the central ver-
sus the more peripheral visual field. Anatomical separa-
tion of FEF–occipital connections by visual eccentricity
(central vs. more peripheral) has already been suggested
by anatomical tracing studies in the macaque brain
(Bullier, Schall, & Morel, 1996; Schall, Morel, King, &
Bullier, 1995; Blatt, Andersen, & Stoner, 1990). However,
to our knowledge, no study to date has systematically
examined possible differences between the anatomical
or functional connectivity of the FEFs in the two dif-
ferent cortical hemispheres, neither in humans nor in
monkeys. In the monkey brain, FEFs in either hemi-
sphere are generally considered to be symmetric homo-
logues, each with a primarily contralateral preference
for visual stimuli (‘‘receptive field’’) and for eye move-
ments (‘‘motor fields’’) (e.g., Wardak et al., 2006; Tehovnik
et al., 2000; Schall & Thompson, 1999). Some contralater-
ality has also now been suggested for human FEF with
fMRI (e.g., Serences & Yantis, 2007; Hagler & Sereno,
2006). Nevertheless, this contralaterality may be relative
rather than absolute in humans. Moreover, such studies
have, to date, concentrated on activity profiles just within
the FEF, whereas here we were concerned instead with
causal influences of stimulating the frontal cortex (with
TMS) upon activity in the remote but potentially intercon-
nected visual cortex.
Our new findings here provide the first evidence that
‘‘effective connectivity’’ of the human left and right fron-
tal cortex with the occipital visual cortex might be similar
for representations of the central visual field, yet show
right-hemisphere dominance for the more peripheral vi-
sual field. Whether this may relate to different anatom-
layout and connectivity of the left versus right
ical
frontal cortex might be examined in future studies, such
as perhaps with diffusion tensor imaging in humans or
anatomical tracing techniques in monkeys. For the ex-
periments described here, we had used a surface MR coil
centered over the occipital cortex, which allowed us to
characterize and compare TMS effects on the retinotopic
visual cortex with high sensitivity, but conversely did
not record signal from more anterior regions.
It is important to stress, however, that the TMS effects
on the visual cortex described here are unlikely to just
reflect fixed effects of monosynaptic anatomical connec-
tions of the stimulated sites with the visual cortex. For
instance, the finding that BOLD changes due to unilateral
frontal stimulation arose in the visual cortex bilaterally,
and even as early as area V1, suggests the involvement of
polysynaptic pathways via intervening cortical or subcor-
tical brain regions (for more extensive discussion, see
Ruff et al., 2006, 2008). Moreover, the visual-context-
dependence of effects for right-parietal TMS, as described
below, suggests that remote effects of TMS may not re-
flect only fixed anatomical connections, but may rather
indicate functional coupling between areas that can
change with current context, that is, with the functional
state of the neuronal circuitry at the time when TMS is
applied (see also Bestmann et al., 2007; Massimini et al.,
2005; Friston, 2002; Munchau, Bloem, Irlbacher, Trimble,
& Rothwell, 2002; McIntosh, 2000).
Our results for TMS to the parietal sites revealed even
more striking evidence that right-hemisphere TMS can
result in influences upon the visual cortex that compa-
rable left-hemisphere TMS does not produce. Right pa-
rietal TMS elicited strong BOLD increases in areas V1–V4
when no visual stimuli were presented, so that the visual
cortex was not activated by external input. When visual
stimuli were present, by contrast, right IPS TMS spe-
cifically affected activity only in visual area V5/MT+,
leading to strong decreases in BOLD response to the
moving visual stimuli in this region. This change of ‘‘ef-
fective connectivity’’ between right parietal cortex and
areas in the visual cortex, as a function of visual context,
may fit the emerging view that neural signals in parietal
regions may be more visually driven than those in fron-
tal areas (e.g., Buschman & Miller, 2007), and may
possibly relate to flexible, context-dependent coding of
the environment (e.g., Wardak et al., 2006; Macaluso &
Driver, 2005; Shulman et al., 2003; Culham, Cavanagh, &
Kanwisher, 2001; Kastner, Pinsk, De Weerd, Desimone,
& Ungerleider, 1999). Context-dependence of effective
connectivity between the right IPS and visual areas
might explain why right parietal TMS affected activity
in V5/MT+ only in the presence of moving visual stimuli
and, by the same token, why functional connections be-
tween the right parietal and retinotopic visual cortex
(V1–V4) here appeared less responsive to any TMS-driven
‘‘feedback’’
influences when TMS was applied in the
presence of strong driving visual input, which may have
dominated connections between visual cortex and IPS in
a ‘‘feedforward’’ manner. Future studies might test this
conjecture in more detail; for instance, by examining how
varying the strength (contrast) of concurrent visual stim-
ulation may affect the modulatory influence of right pa-
rietal TMS upon activity in the early visual cortex.
Ruff et al.
1157
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
6
e
-
1
p
1
d
4
f
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
o
4
c
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
.
f
/
t
o
n
1
8
M
a
y
2
0
2
1
The new data presented here now show that left pa-
rietal TMS had drastically different effects, as it did not
affect BOLD signal in any visual cortical area, in either
visual context (i.e., with or without visual stimuli). More-
over, this was more than just an uninformative null
outcome, as the outcome for left parietal TMS differed
significantly from that for right parietal TMS. This pat-
tern of marked right-hemisphere dominance for the ef-
fects of parietal TMS seems to accord well with clinical
observations because deficits such as neglect or ex-
tinction are most often reported after lesions of right-
hemisphere brain areas in and around the parietal
cortex and the IPS, whereas lesions of corresponding
parietal regions in the left hemisphere rarely have such
effects (Becker & Karnath, 2007; Milner & McIntosh,
2005; Mort et al., 2003; Karnath et al., 2002; Mesulam,
1999). Our findings provide a possible new interpreta-
tion of such clinical lesion data, namely, that the marked
effects of lesions to right-hemisphere regions of the
human parietal cortex may relate to the specific capac-
ity of these regions for functional influences upon the
visual cortex. This interpretation could also fit with the
fact that some of the clinical sequelae of right parietal
damage (e.g., extinction) likewise depend on current
visual context (see e.g., Driver, Vuilleumier, et al., 2004;
Marzi et al., 2001; Vuilleumier & Rafal, 2000; Mesulam,
1999), as established also for the right parietal TMS
effects upon the visual cortex here.
Our new TMS–fMRI findings here may also shed new
light on the possible neural mechanisms underlying
purely behavioral TMS effects. Several behavioral TMS
studies have reported more marked impacts of right-
than left-hemisphere frontal or parietal TMS upon visual
performance (e.g., Muggleton et al., 2003, 2006; Chambers
et al., 2004; O’Shea et al., 2004; Grosbras & Paus, 2002,
2003; Pourtois et al., 2001). But it has so far remained
unclear whether such behavioral TMS effects reflect
hemispheric differences only in the local function of the
stimulated cortical sites, or instead, in their functional
interactions with the remote visual cortex. Our new re-
sults here provide direct evidence that the visual cortex
itself can be more strongly affected when frontal or pa-
rietal TMS is administered over the right rather than over
the left hemisphere. Such remote influences upon the
visual cortex may plausibly underlie some of the marked
behavioral effects of right-hemisphere frontal/parietal
TMS on performance on visual tasks. This point may also
be noteworthy from a purely methodological perspective,
as our findings imply that TMS effects on behavior may
not solely reflect influences on neural processing directly
under the stimulation coil, but also the impact upon
other areas in the network of remote but interconnected
brain regions participating in a given cognitive function
(see also Bestmann, Ruff, Driver, & Blankenburg, 2008;
Sack et al., 2007).
In contrast to lesion and TMS investigations in hu-
mans, studies in nonhuman primates have, to date, typi-
cally not reported (but also rarely sought) hemispheric
differences for the perceptual effects of interventions
such as lesions, microstimulation, cooling, or chemical in-
activation of frontal or parietal brain areas (e.g., Wardak
et al., 2004, 2006; Moore & Fallah, 2004). Hemispheric
differences in effects of TMS on perceptual behavior,
and for remote impacts upon visual cortex as described
here, may reflect some aspects of hemispheric speciali-
zation that are potentially unique to the human brain
(Sun & Walsh, 2006; Davidson & Hugdahl, 1995; Hellige,
1990). Such right-hemisphere lateralization of perceptual
control might complement, or even relate in an evolu-
tionary sense, to other hemispheric specializations of
the human brain, such as typical left-hemisphere domi-
nance for language processing (e.g., Josse & Tzourio-
Mazoyer, 2004). In future work with the new approach
developed here, it might now become possible to study
with concurrent TMS–fMRI whether regions related to,
say, language processing might also show hemispheric
differences in terms of their functional connectivity with
remote but interconnected brain areas, perhaps with left-
hemisphere predominance for remote effects of TMS in
a language context.
In conclusion, we found that TMS to right-hemisphere
frontal and parietal regions of the human brain could
have distinct influences on BOLD signal in visual areas
V1–V4 and V5/MT+, which were absent (and signifi-
cantly different) during TMS over the corresponding
left-hemisphere sites. These data directly confirm right-
hemisphere predominance for fronto-parietal causal in-
fluences upon processing in the human visual cortex,
which may, in turn, relate to the stronger perceptual–
attentional disruptions typically found after lesions or
TMS interventions concerning right-hemisphere human
fronto-parietal regions. More generally, our results il-
lustrate how concurrent TMS–fMRI can provide a new
approach to the long-standing question of functional lat-
eralization in the human brain, now in relation to causal
interactions between remote but interconnected brain
regions.
Acknowledgments
Supported by the Wellcome Trust, the Medical Research Coun-
cil UK, and the European Commission, 7th Framework Pro-
gram (BrainSync: HEALTH-F2-2008-200728). J. D. holds a Royal
Society-Leverhulme Trust Senior Research Fellowship.
Reprint requests should be sent to Christian C. Ruff, UCL Institute
of Cognitive Neuroscience, University College London, 17 Queen
Square, London, WC1 3AR UK, or via e-mail: c.ruff@ucl.ac.uk.
REFERENCES
Andersson, J. L., Hutton, C., Ashburner, J., Turner, R., &
Friston, K. (2001). Modeling geometric deformations
in EPI time series. Neuroimage, 13, 903–919.
Armstrong, K. M., & Moore, T. (2007). Rapid enhancement of
visual cortical response discriminability by microstimulation
1158
Journal of Cognitive Neuroscience
Volume 21, Number 6
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
6
e
-
1
p
1
d
4
f
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
o
4
c
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
t
f
/
.
o
n
1
8
M
a
y
2
0
2
1
of the frontal eye field. Proceedings of the National
Academy of Sciences, U.S.A, 104, 9499–9504.
of spatial and motor codes during oculomotor delayed
response tasks. Journal of Neuroscience, 24, 3944–3952.
Barcelo´, F., Suwazono, S., & Knight, R. T. (2000). Prefrontal
Davidson, R. A., & Hugdahl, K. (1995). Brain asymmetry.
modulation of visual processing in humans. Nature
Neuroscience, 3, 399–403.
Baudewig, J., Siebner, H. R., Bestmann, S., Tergau, F.,
Tings, T., Paulus, W., et al. (2001). Functional MRI of
cortical activations induced by transcranial magnetic
stimulation (TMS). NeuroReport, 12, 3543–3548.
Becker, E., & Karnath, H. O. (2007). Incidence of visual
extinction after left versus right hemisphere stroke.
Stroke, 38, 3172–3174.
Bestmann, S., Ruff, C. C., Driver, J., & Blankenburg, F.
(2008). Concurrent TMS and functional magnetic resonance
imaging: Methods and current advances. In E. Wassermann,
C. Epstein, U. Ziemann, V. Walsh, T. Paus, & S. Lisanby
(Eds.), Oxford handbook of transcranial stimulation
(pp. 569–592). Oxford: Oxford University Press.
Cambridge: MIT Press.
Deco, G., & Zihl, J. (2004). A biased competition based
neurodynamical model of visual neglect. Medical
Engineering Physics, 26, 733–743.
Driver, J., Eimer, M., Macaluso, E., & van Velzen, J. (2004). The
neurobiology of human spatial attention. In N. Kanwisher
& J. Duncan (Eds.), Functional neuroimaging of visual
cognition: Attention and performance XX (pp. 267–300).
Oxford: Oxford University Press.
Driver, J., & Mattingley, J. B. (1998). Parietal neglect and
visual awareness. Nature Neuroscience, 1, 17–22.
Driver, J., Vuilleumier, P., & Husain, M. (2004). Spatial
neglect and extinction. In M. J. Gazzaniga (Ed.), The
new cognitive neurosciences (pp. 589–606). Cambridge,
MA: MIT Press.
Bestmann, S., Swayne, O., Blankenburg, F., Ruff, C. C.,
Duncan, J., Humphreys, G., & Ward, R. (1997). Competitive
Haggard, P., Weiskopf, N., et al. (2007). Dorsal premotor
cortex exerts state-dependent causal influences on activity
in contralateral primary motor and dorsal premotor
cortex. Cerebral Cortex, 26, 7452–7459.
Blanke, O., Spinelli, L., Thut, G., Michel, C. M., Perrig, S.,
Landis, T., et al. (2000). Location of the human frontal
eye field as defined by electrical cortical stimulation:
Anatomical, functional and electrophysiological
characteristics. NeuroReport, 11, 1907–1913.
Blatt, G. J., Andersen, R. A., & Stoner, G. R. (1990). Visual
receptive field organization and cortico-cortical connections
of the lateral intraparietal area (area LIP) in the macaque.
Journal of Comparative Neurology, 299, 421–445.
Bohning, D. E., Shastri, A., McConnell, K. A., Nahas, Z.,
Lorberbaum, J. P., Roberts, D. R., et al. (1999). A combined
TMS/fMRI study of intensity-dependent TMS over motor
cortex. Biological Psychiatry, 45, 385–394.
Brown, M. R., DeSouza, J. F., Goltz, H. C., Ford, K., Menon,
R. S., Goodale, M. A., et al. (2004). Comparison of
memory- and visually guided saccades using event-related
fMRI. Journal of Neurophysiology, 91, 873–889.
Bullier, J., Schall, J. D., & Morel, A. (1996). Functional
streams in occipito-frontal connections in the monkey.
Behavioral Brain Research, 76, 89–97.
Buschman, T. J., & Miller, E. K. (2007). Top–down versus
bottom–up control of attention in the prefrontal and
posterior parietal cortices. Science, 315, 1860–1862.
Chambers, C. D., Payne, J. M., Stokes, M. G., & Mattingley,
J. B. (2004). Fast and slow parietal pathways mediate
spatial attention. Nature Neuroscience, 7, 217–218.
Connolly, J. D., Goodale, M. A., DeSouza, J. F., Menon, R. S.,
& Vilis, T. (2000). A comparison of frontoparietal fMRI
activation during anti-saccades and anti-pointing. Journal
of Neurophysiology, 84, 1645–1655.
Connolly, J. D., Goodale, M. A., Menon, R. S., & Munoz, D. P.
(2002). Human fMRI evidence for the neural correlates of
preparatory set. Nature Neuroscience, 5, 1345–1352.
Corbetta, M., Akbudak, E., Conturo, T. E., Snyder, A. Z.,
Ollinger, J. M., Drury, H. A., et al. (1998). A common
network of functional areas for attention and eye
movements. Neuron, 21, 761–773.
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed
and stimulus-driven attention in the brain. Nature
Reviews Neuroscience, 3, 201–215.
Culham, J. C., Cavanagh, P., & Kanwisher, N. G. (2001).
Attention response functions: Characterizing brain areas
using fMRI activation during parametric variations of
attentional load. Neuron, 32, 737–745.
Curtis, C. E., Rao, V. Y., & D’Esposito, M. (2004). Maintenance
brain activity in visual attention. Current Opinion in
Neurobiology, 7, 255–261.
Friston, K. (2002). Beyond phrenology: What can
neuroimaging tell us about distributed circuitry?
Annual Review of Neuroscience, 25, 221–250.
Friston, K. J., Holmes, A. P., Worsley, K. J., Poline, J. B.,
Frith, C. D., & Frackowiak, R. S. J. (1995). Statistical
parametric maps in functional imaging: A general
linear approach. Human Brain Mapping, 2, 189–210.
Friston, K. J., Penny, W., Phillips, C., Kiebel, S., Hinton, G.,
& Ashburner, J. (2002). Classical and Bayesian inference
in neuroimaging: Theory. Neuroimage, 16, 465–483.
Frith, C. (2001). A framework for studying the neural basis
of attention. Neuropsychologia, 39, 1367–1371.
Grosbras, M. H., Laird, A. R., & Paus, T. (2005). Cortical regions
involved in eye movements, shifts of attention, and gaze
perception. Human Brain Mapping, 25, 140–154.
Grosbras, M. H., & Paus, T. (2002). Transcranial magnetic
stimulation of the human frontal eye field: Effects on
visual perception and attention. Journal of Cognitive
Neuroscience, 14, 1109–1120.
Grosbras, M. H., & Paus, T. (2003). Transcranial magnetic
stimulation of the human frontal eye field facilitates
visual awareness. European Journal of Neuroscience,
18, 3121–3126.
Hagler, D. J., Jr., & Sereno, M. I. (2006). Spatial maps in
frontal and prefrontal cortex. Neuroimage, 29, 567–577.
Hellige, J. B. (1990). Hemispheric asymmetry. Annual
Review of Psychology, 41, 55–80.
Hopfinger, J. B., Buonocore, M. H., & Mangun, G. R. (2000).
The neural mechanisms of top–down attentional control.
Nature Neuroscience, 3, 284–291.
Josse, G., & Tzourio-Mazoyer, N. (2004). Hemispheric
specialization for language. Brain Research, Brain
Research Review, 44, 1–12.
Juan, C. H., Shorter-Jacobi, S. M., & Schall, J. D. (2004).
Dissociation of spatial attention and saccade preparation.
Proceedings of the National Academy of Sciences, U.S.A.,
101, 15541–15544.
Karnath, H.-O., Milner, A. D., & Vallar, G. (2002). The
cognitive and neural bases of spatial neglect. Oxford:
Oxford University Press.
Kastner, S., Pinsk, M. A., De Weerd, P., Desimone, R., &
Ungerleider, L. G. (1999). Increased activity in human
visual cortex during directed attention in the absence
of visual stimulation. Neuron, 22, 751–761.
Kastner, S., & Ungerleider, L. G. (2000). Mechanisms of
visual attention in the human cortex. Annual Review
of Neuroscience, 23, 315–341.
Ruff et al.
1159
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
6
e
-
1
p
1
d
4
f
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
o
4
c
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
f
/
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Macaluso, E., & Driver, J. (2005). Multisensory spatial
Pourtois, G., Vandermeeren, Y., Olivier, E., & de Gelder, B.
interactions: A window onto functional integration in the
human brain. Trends in Neurosciences, 28, 264–271.
Marzi, C. A., Girelli, M., Miniussi, C., Smania, N., & Maravita, A.
(2000). Electrophysiological correlates of conscious vision:
Evidence from unilateral extinction. Journal of Cognitive
Neuroscience, 12, 869–877.
Marzi, C. A., Girelli, M., Natale, E., & Miniussi, C. (2001).
What exactly is extinguished in unilateral visual extinction?
Neurophysiological evidence. Neuropsychologia, 39,
1354–1366.
Massimini, M., Ferrarelli, F., Huber, R., Esser, S. K., Singh, H.,
& Tononi, G. (2005). Breakdown of cortical effective
connectivity during sleep. Science, 309, 2228–2232.
McIntosh, A. R. (2000). Towards a network theory of
cognition. Neural Networks, 13, 861–870.
Mesulam, M. M. (1999). Spatial attention and neglect:
Parietal, frontal and cingulate contributions to the
mental representation and attentional targeting of salient
extrapersonal events. Philosophical Transactions of the
Royal Society of London, Series B, Biological Sciences,
354, 1325–1346.
Miller, B. T., & D’Esposito, M. (2005). Searching for ‘‘the
top’’ in top–down control. Neuron, 48, 535–538.
Miller, E. K. (2000). The neural basis of the top–down
control of visual attention in the prefrontal cortex.
In S. Monsell & J. Driver (Eds.), Control of cognitive
processes: Attention and performance XVIII (pp. 511–534).
Cambridge, MA: MIT Press.
Milner, A. D., & McIntosh, R. D. (2005). The neurological
basis of visual neglect. Current Opinion in Neurology,
18, 748–753.
Moore, T., & Armstrong, K. M. (2003). Selective gating of
visual signals by microstimulation of frontal cortex.
Nature, 421, 370–373.
Moore, T., & Fallah, M. (2004). Microstimulation of the
frontal eye field and its effects on covert spatial attention.
Journal of Neurophysiology, 91, 152–162.
Mort, D. J., Malhotra, P., Mannan, S. K., Rorden, C., Pambakian,
A., Kennard, C., et al. (2003). The anatomy of visual
neglect. Brain, 126, 1986–1997.
Muggleton, N. G., Juan, C. H., Cowey, A., & Walsh, V. (2003).
Human frontal eye fields and visual search. Journal of
Neurophysiology, 89, 3340–3343.
Muggleton, N. G., Postma, P., Moutsopoulou, K.,
Nimmo-Smith, I., Marcel, A., & Walsh, V. (2006). TMS
over right posterior parietal cortex induces neglect in a
scene-based frame of reference. Neuropsychologia, 44,
1222–1229.
Munchau, A., Bloem, B. R., Irlbacher, K., Trimble, M. R., &
Rothwell, J. C. (2002). Functional connectivity of human
premotor and motor cortex explored with repetitive
transcranial magnetic stimulation. Journal of Neuroscience,
22, 554–561.
O’Shea, J., Muggleton, N. G., Cowey, A., & Walsh, V. (2004).
Timing of target discrimination in human frontal eye
fields. Journal of Cognitive Neuroscience, 16, 1060–1067.
Paus, T. (1996). Location and function of the human
frontal eye-field: A selective review. Neuropsychologia, 34,
475–483.
Paus, T., Jech, R., Thompson, C. J., Comeau, R., Peters, T.,
& Evans, A. C. (1997). Transcranial magnetic stimulation
during positron emission tomography: A new method
for studying connectivity of the human cerebral cortex.
Journal of Neuroscience, 17, 3178–3184.
Perry, R. J., & Zeki, S. (2000). The neurology of saccades
and covert shifts in spatial attention: An event-related
fMRI study. Brain, 123, 2273–2288.
(2001). Event-related TMS over the right posterior
parietal cortex induces ipsilateral visuo-spatial
interference. NeuroReport, 12, 2369–2374.
Rees, G., Friston, K., & Koch, C. (2000). A direct quantitative
relationship between the functional properties of human
and macaque V5. Nature Neuroscience, 3, 716–723.
Ro, T., Cheifet, S., Ingle, H., Shoup, R., & Rafal, R. (1999).
Localization of the human frontal eye fields and motor
hand area with transcranial magnetic stimulation and
magnetic resonance imaging. Neuropsychologia, 37,
225–231.
Ruff, C. C., Bestmann, S., Blankenburg, F., Bjoertomt, O.,
Josephs, O., Weiskopf, N., et al. (2008). Distinct causal
influences of parietal versus frontal areas on human
visual cortex: Evidence from concurrent TMS fMRI.
Cerebral Cortex, 18, 817–827.
Ruff, C. C., Blankenburg, F., Bjoertomt, O., Bestmann, S.,
Freeman, E., Haynes, J. D., et al. (2006). Concurrent
TMS–fMRI and psychophysics reveal frontal influences
on human retinotopic visual cortex. Current Biology,
16, 1479–1488.
Ruff, C. C., & Driver, J. (2006). Attentional preparation for a
lateralized visual distractor: Behavioral and fMRI evidence.
Journal of Cognitive Neuroscience, 18, 522–538.
Sack, A. T., Kohler, A., Bestmann, S., Linden, D. E., Dechent, P.,
Goebel, R., et al. (2007). Imaging the brain activity changes
underlying impaired visuospatial judgments: Simultaneous
fMRI, TMS, and behavioral studies. Cerebral Cortex, 17,
2841–2852.
Schall, J. D., Morel, A., King, D. J., & Bullier, J. (1995).
Topography of visual cortex connections with frontal
eye field in macaque: Convergence and segregation of
processing streams. Journal of Neuroscience, 15,
4464–4487.
Schall, J. D., & Thompson, K. G. (1999). Neural selection
and control of visually guided eye movements. Annual
Review of Neuroscience, 22, 241–259.
Schwartz, S., Vuilleumier, P., Hutton, C., Maravita, A., Dolan,
R. J., & Driver, J. (2005). Attentional load and sensory
competition in human vision: Modulation of fMRI
responses by load at fixation during task-irrelevant
stimulation in the peripheral visual field. Cerebral Cortex,
15, 770–786.
Serences, J. T., & Yantis, S. (2006). Selective visual attention
and perceptual coherence. Trends in Cognitive Sciences,
10, 38–45.
Serences, J. T., & Yantis, S. (2007). Spatially selective
representations of voluntary and stimulus-driven
attentional priority in human occipital, parietal, and
frontal cortex. Cerebral Cortex, 17, 284–293.
Sereno, M. I., Dale, A. M., Reppas, J. B., Kwong, K. K.,
Belliveau, J. W., Brady, T. J., et al. (1995). Borders of
multiple visual areas in humans revealed by functional
magnetic resonance imaging. Science, 268, 889–893.
Shastri, A., George, M. S., & Bohning, D. E. (1999).
Performance of a system for interleaving transcranial
magnetic stimulation with steady-state magnetic
resonance imaging. Electroencephalography and
Clinical Neurophysiology, 51, 55–64.
Shulman, G. L., McAvoy, M. P., Cowan, M. C., Astafiev, S. V.,
Tansy, A. P., d’Avossa, G., et al. (2003). Quantitative
analysis of attention and detection signals during visual
search. Journal of Neurophysiology, 90, 3384–3397.
Silvanto, J., Lavie, N., & Walsh, V. (2006). Stimulation of
human frontal eye fields modulates sensitivity of
extrastriate visual cortex. Journal of Neurophysiology, 96,
941–945.
1160
Journal of Cognitive Neuroscience
Volume 21, Number 6
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
6
e
-
1
p
1
d
4
f
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
o
4
c
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
t
.
.
f
.
o
n
1
8
M
a
y
2
0
2
1
Sun, T., & Walsh, C. A. (2006). Molecular approaches to
brain asymmetry and handedness. Nature Reviews
Neuroscience, 7, 655–662.
Taylor, P. C., Nobre, A. C., & Rushworth, M. F. (2007). FEF
TMS affects visual cortical activity. Cerebral Cortex, 17,
391–399.
Tehovnik, E. J., Sommer, M. A., Chou, I. H., Slocum, W. M.,
& Schiller, P. H. (2000). Eye fields in the frontal lobes of
primates. Brain Research, Brain Research Reviews, 32,
413–448.
Teo, P. C., Sapiro, G., & Wandell, B. A. (1997). Creating
connected representations of cortical gray matter for
functional MRI visualization. IEEE Transactions in
Medical Imaging, 16, 852–863.
Vuilleumier, P. O., & Rafal, R. D. (2000). A systematic study
of visual extinction. Between- and within-field deficits of
attention in hemispatial neglect. Brain, 123, 1263–1279.
Wandell, B. A., Chial, S., & Backus, B. T. (2000). Visualization
and measurement of the cortical surface. Journal of
Cognitive Neuroscience, 12, 739–752.
Wardak, C., Ibos, G., Duhamel, J. R., & Olivier, E. (2006).
Contribution of the monkey frontal eye field to covert
visual attention. Journal of Neuroscience, 26, 4228–4235.
Wardak, C., Olivier, E., & Duhamel, J. R. (2004). A deficit in
covert attention after parietal cortex inactivation in the
monkey. Neuron, 42, 501–508.
Wassermann, E. M. (1998). Risk and safety of repetitive
transcranial magnetic stimulation: Report and suggested
guidelines from the International Workshop on the
Safety of Repetitive Transcranial Magnetic Stimulation,
June 5–7, 1996. Electroencephalography and Clinical
Neurophysiology, 108, 1–16.
Watson, J. D., Myers, R., Frackowiak, R. S., Hajnal, J. V.,
Woods, R. P., Mazziotta, J. C., et al. (1993). Area V5 of
the human brain: Evidence from a combined study using
positron emission tomography and magnetic resonance
imaging. Cerebral Cortex, 3, 79–94.
Winkowski, D. E., & Knudsen, E. I. (2007). Top–down
control of multimodal sensitivity in the barn owl optic
tectum. Journal of Neuroscience, 27, 13279–13291.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
6
e
-
1
p
1
d
4
f
6
/
1
2
9
1
3
/
7
6
6
/
7
1
3
1
o
4
c
6
n
/
1
2
0
7
0
6
9
0
1
2
5
1
5
0
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
7
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
/
.
.
t
o
n
1
8
M
a
y
2
0
2
1
Ruff et al.
1161