焦点功能:
Connectivity, 认识, and Consciousness
A natural cortical axis connecting the outside and
inside of the human brain
克劳斯·C. Hilgetag1,2
, Alexandros Goulas1
, and Jean-Pierre Changeux3,4
1计算神经科学研究所, 艾彭多夫大学医学中心, 汉堡大学, 汉堡, 德国
2健康科学系, 波士顿大学, 波士顿, 嘛, 美国
3CNRS UMR 3571, 巴斯德研究所, 巴黎, 法国
4Communications Cellulaires, Collège de France, 巴黎, 法国
关键词: Comparative connectomics, Connectomic hypothesis, Hominization, Brain evolution,
Cortical development
开放访问
杂志
抽象的
What structural and connectivity features of the human brain help to explain the extraordinary
human cognitive abilities? We recently proposed a set of relevant connectomic fundamentals,
some of which arise from the size scaling of the human brain relative to other primate brains,
while others of these fundamentals may be uniquely human. 尤其, we suggested that
the remarkable increase of the size of the human brain due to its prolonged prenatal
development has brought with it an increased sparsification, hierarchical modularization, 作为
well as increased depth and cytoarchitectonic differentiation of brain networks. 这些
characteristic features are complemented by a shift of projection origins to the upper layers of
many cortical areas as well as the significantly prolonged postnatal development and plasticity
of the upper cortical layers. Another fundamental aspect of cortical organization that has
emerged in recent research is the alignment of diverse features of evolution, 发展,
cytoarchitectonics, function, and plasticity along a principal, natural cortical axis from sensory
(“outside”) to association (“inside”) 地区. Here we highlight how this natural axis is integrated
in the characteristic organization of the human brain. 尤其, the human brain displays a
developmental expansion of outside areas and a stretching of the natural axis such that outside
areas are more widely separated from each other and from inside areas than in other species.
We outline some functional implications of this characteristic arrangement.
作者总结
Which brain connectivity features underlie the extraordinary human cognitive abilities? 我们
have proposed a set of relevant connectomic fundamentals, including the increased
sparsification, hierarchical modularization, as well as increased depth and cytoarchitectonic
differentiation of cortical networks due to the prolonged prenatal development and size increase
of the human brain. Such features are complemented by a shift of projection origins to the upper
layers of many cortical areas and significantly prolonged postnatal development of such layers.
Another fundamental aspect of cortical organization is the alignment of diverse features of
evolution, 发展, cytoarchitectonics, function, and plasticity along a natural cortical axis
from sensory (“outside”) to association (“inside”) 地区. Here we highlight how this natural axis is
integrated in the characteristic connectivity organization of the human brain.
引文: 希尔格塔格, C. C。, Goulas, A。, &
尚古克斯, J.-P. (2022). A natural
cortical axis connecting the outside
and inside of the human brain. 网络
神经科学, 6(4), 950–959. https://
doi.org/10.1162/netn_a_00256
DOI:
https://doi.org/10.1162/netn_a_00256
已收到: 9 十一月 2021
公认: 17 可能 2022
利益争夺: 作者有
声明不存在竞争利益
存在.
通讯作者:
克劳斯·C. 希尔格塔格
c.hilgetag@uke.de
处理编辑器:
奥拉夫·斯波恩斯
版权: © 2022
麻省理工学院
在知识共享下发布
归因 4.0 国际的
(抄送 4.0) 执照
麻省理工学院出版社
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
4
9
5
0
2
0
5
6
2
7
0
n
e
n
_
A
_
0
0
2
5
6
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
A natural cortical axis of the human brain
Connectomic fundamentals:
Characteristic connectivity features
underlying computational and
functional properties of the brain.
The connectomic hypothesis:
Suggests that the characteristic
connectivity of the human brain is
responsible for the extraordinary
human cognitive abilities.
Sparsification:
Decreasing density of brain networks
as their size increases.
Modularity of connectivity:
Arises when some regions are more
frequently or densely connected with
each other than with other regions of
a network.
Core:
Network regions that are particularly
densely connected with each other,
more than what would be expected
by the general number of their
连接.
Externopyramidization:
The shift in the relative soma size of
supragranular versus infragranular
pyramidal neurons toward larger
supragranular neurons.
Epigenetic processes:
Phenotype modifications that do not
involve alterations in the DNA
顺序.
Supragranular:
Cortical layers above the granular
layer IV. Infragranular: cortical layers
below the granular layer IV.
CONNECTOMIC FUNDAMENTALS OF THE HUMAN BRAIN
The extraordinary cognitive abilities of modern humans, which have arisen over an evolution-
arily short time span of less than one million years, present an apparent paradox, as there are
no conspicuous differences in the size or the content of the human genome relative to non-
human primates that could readily explain these abilities. 而且, no singular genes have
been identified that are specifically linked to human cognitive abilities such as language,
which rather appear to arise from the interactions of a multitude of genes. As an alternative,
we have recently put forward a connectomic hypothesis for the hominization of the brain
(Changeux et al., 2021). This hypothesis rests on the assumption that primarily small changes
in gene expression, rather than genome content, have affected the development of the human
brain and with it its connectivity, as an intermediate functional phenotype (尚古克斯, 2017).
尤其, many human connectomic features might be accounted for by extended pre-
natal development and the resulting substantial increase in brain size within the global neural
organization of the primate brain (巴克纳 & 克里宁, 2013; Nieuwenhuys & Puelles, 2016).
Such increased brain size, 反过来, leads to a larger number of cortical neurons as well as areas,
and the sparsification and increased hierarchical (encapsulated) modularity of cortical connec-
系统蒸发散. The combination of these features with the developmental expansion of upper cortical
layers, the increased laminar differentiation of cortical projections, as well as a prolonged post-
natal brain development, which provides a substantially expanded exposure to nongenetic
interactions with the physical, 社会的, and cultural environment, gives rise to categorically
human-specific cognitive abilities and particularly language.
In this hypothesis, we distinguish characteristic aspects of two kinds, 具体来说, features of
the human brain that are scaled within a primate envelope of brain organization versus fea-
tures that are uniquely human. Concretely, we consider as scaled features the developmental
expansion of the human brain with the resulting sparsification and hierarchical modularization
of its connectivity as well as expanded depth and cytoarchitectonic specialization of cortical
processing stages, 尤其, the cytoarchitectonic differentiation between the core and
periphery of cortical connectivity. 相比之下, singular features may be formed by a shift of
laminar projection origins (“externopyramidization”) in many cortical areas from lower to
upper layers (Goulas et al., 2018; Sanides, 1962, 1970), a postnatal dendritic expansion of
associative projection layer IIIc pyramidal cells, 例如, in the prefrontal cortex (Petanjek
等人。, 2019), as well as more generally a prolonged period of postnatal development with
considerable involvement of epigenetic processes of synapse selection and connectomic reor-
ganization (尚古克斯, 2022; Changeux et al., 2021). We have suggested that these features
may be controlled through specific gene regulatory events, although the concrete mechanisms
are presently unknown.
Recent findings by several groups support and extend this hypothesis. 例如, Hendy
等人. (2020) identified genes that are enriched specifically in the supragranular layers of the
human cortex compared to mice. These genes show a relatively protracted expression in the
人脑, corresponding to an extended duration of cortical connectivity development,
and they may contribute to the characteristically expanded network of frontal cortico-cortical
projections in the human brain. 而且, Heyer et al. (2022) observed a selective expansion
of supragranular layers II and III of left temporal cortex (specifically Brodmann area 21) in sub-
jects with higher verbal and general IQ. This expansion was associated with larger cell body
size of pyramidal neurons, which can support faster propagation of action potentials and may
serve to improve information processing. Such observations directly confirm predictions of
our connectomic hypothesis and demonstrate that variations of gene expression, 特别
网络神经科学
951
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
4
9
5
0
2
0
5
6
2
7
0
n
e
n
_
A
_
0
0
2
5
6
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
A natural cortical axis of the human brain
in upper cortical layers, may have affected developmental trajectories of the human brain,
resulting in characteristically expanded and modified connectivity supporting enhanced
cognitive functioning.
A NATURAL AXIS OF OUTSIDE-TO-INSIDE CORTICAL ORGANIZATION
In addition to the features mentioned above, there is another fundamental aspect of the orga-
nization of mammalian brains in general, and the human cerebral cortex in particular, 这
alignment of several structural and functional features along a principal gradient, or natural
轴, of cortical organization (Goulas et al., 2019, 2021; Sydnor et al., 2021).
The heterogeneity of the structure and function of the cerebral cortex was already appre-
ciated in classical studies (例如, Gall & Spurzheim, 1810; Broca, 1861). At least since the
cytoarchitectonic studies of von Economo and Koskinas (von Economo, 1927; von Economo
& Koskinas, 1925), who described an ordered system of structural cortical types, 它变成了
clear that this structural heterogeneity is not arranged randomly or in a mosaic fashion, 但
organized along spatial gradients of the cerebral cortex. Examples of graded features include
the laminar differentiation (IE。, how well different cortical layers, and particularly the granular
layer IV, can be observed in a cortical region), the density of cells and the distribution of
different cell types and morphologies across cortical layers (Dombrowski et al., 2001; Goulas
等人。, 2018), the degree of myelination, as well as fine-grained cellular morphology (Elston,
2002), forming a graded spectrum of cortical structural organization (John et al., 2021) (看
数字 1).
Later studies unified the concept of cortical gradients, by demonstrating that multiple such
graded features may be spatially aligned. An important insight was the observation by Pandya
and Sanides (1973) that cytoarchitectonic similarity and connectivity may be associated. 这
idea was formalized and expanded into the so-called Structural Model of Connections by
Barbas and colleagues, which summarizes how the existence and laminar projection patterns
of connections are aligned with the graded cytoarchitecture of primate cortical regions
(Barbas, 1986; Barbas & Rempel-Clower, 1997; García-Cabezas et al., 2019). A further con-
ceptualization of these fundamental interrelations, the Architectonic Type Principle (希尔格塔格
等人。, 2019), also describes how microscopic morphological features of neurons as well as
macroscopic features of cortical connectivity and topology are related to the structural types
of cortical areas, as captured by their neuron density and laminar differentiation, 以及如何
these interrelations may arise from the ordered spatial-temporal ontogeny of the cerebral
cortex (Beul et al., 2018; Beul & 希尔格塔格, 2020). 所以, the spatially ordered gradient
of structural types of cortical areas has emerged as a fundamental feature to which many other
microscopic and macroscopic features of cortical organization are related, presumably orga-
nized by interactions during joined development. This includes patterns of gene expression;
gross morphological features, such as cortical thickness and areal expansion during postnatal
发展; or functional aspects such as energy consumption or the time scale or complex-
ity of physiological responses (Huntenburg et al., 2018; Murray et al., 2014; Sydnor et al.,
2021; Wagstyl et al., 2015). 最近, we demonstrated that this gradient of structural cortical
types is also aligned with the organization of different neurotransmitter receptors (Zilles &
Palomero-Gallagher, 2017), forming a unified natural cortical axis (Goulas et al., 2021).
The axis can be considered natural in the sense that it does not strictly follow a prescribed
spatial dimension, such as posterior to anterior. 反而, the axis has been proposed to stretch
more generally from “sensory” to “association” or “transmodal” areas. A further conceptual
simplification of this already simple picture would be to describe the axis as a progression from
网络神经科学
952
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
4
9
5
0
2
0
5
6
2
7
0
n
e
n
_
A
_
0
0
2
5
6
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
A natural cortical axis of the human brain
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
4
9
5
0
2
0
5
6
2
7
0
n
e
n
_
A
_
0
0
2
5
6
p
d
t
.
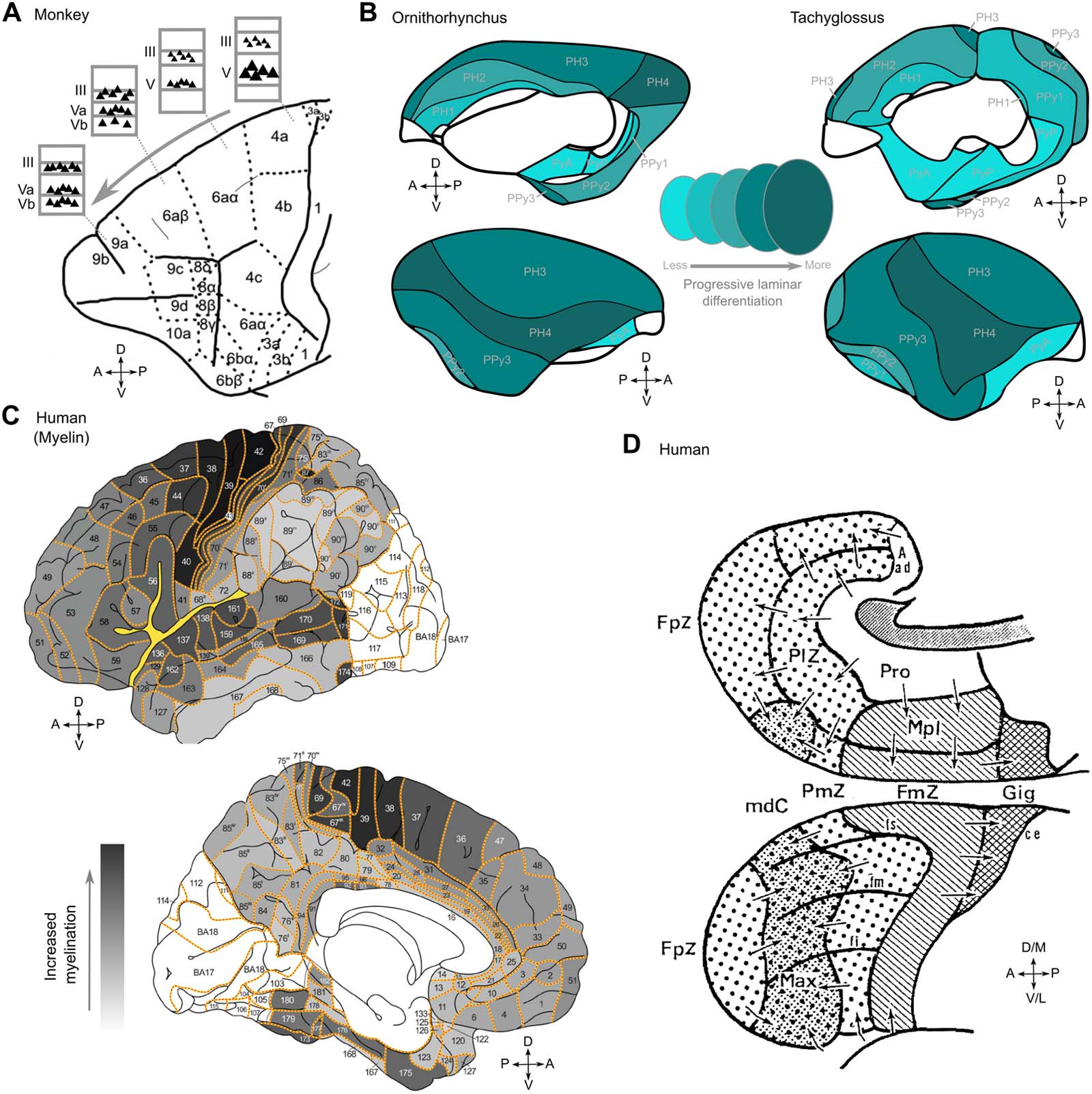
数字 1. Gradation principle of the cerebral cortex of human and nonhuman animals. (A) Gradation along the caudal to rostral axis in the
monkey frontal cortex based on its cytoarchitecture (Vogt & Vogt, 1919) showing a shift of large pyramidal cells from the deep to the upper
cortical layers. Note that the very large projection neurons in deep layer V of primary motor cortex, 区域 4, are not necessarily an indication of
the “inside” nature of this area (Barbas & García-Cabezas, 2015; Shipp, 2005). Apart from these Betz cells, providing direct and strong output
to the spinal cord, the largest pyramidal neuron bodies in area 4 are located in layer IIIc (García-Cabezas et al., 2020). (乙) Cytoarchitecture-
基于, whole-cortex gradients in two monotremes, ornithorynchus and tachyglossus (Abbie, 1940). (C) Cytoarchitectonic gradients in the
human frontal lobe (Sanides, 1970). (D) Myeloarchitectonic gradients in the human cerebral cortex. Visual rendering from Nieuwenhuys
and Broere (2017) using observations from Hopf (1955, 1956) and Hopf and Vitzthum (1957). Darker shading indicates stronger myelination.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
“outside” to “inside” areas of the cortex. 尤其, the labels of outside versus inside areas are
meant as a characterization of the endpoints of the axis, rather than as a dichotomous classi-
fication of all cortical areas into just two groups. Key features of outside versus inside areas are
listed in Table 1, and a schematic depiction of the progressive arrangement of the cortical
spectrum from outside to inside areas is given in Figure 2.
The concentric arrangement in Figure 2 is based on a gradient of neural density and
cytoarchitecture, which is fundamental for other structural features of mammalian organiza-
的, such as connectivity profiles, cellular morphological features, or cortical thickness (Beul
& 希尔格塔格, 2019). As this structural spectrum closely aligns with several other structural, 骗局-
nectional, and functional aspects of cortical organization, the diagram effectively unifies many
网络神经科学
953
A natural cortical axis of the human brain
桌子 1.
Typical features of outside versus inside areas of the cerebral cortex
Features
Cyto-architecture
“Outside” areas
Dense (more cells), high laminar differentiation
(eulaminate), larger pyramidal projection
neurons in upper layers
“Inside” areas
Sparse (more neuropil), low laminar differentiation
(agranular or dysgranular types), larger pyramidal
projection neurons in deep layers
Connectivity
More locally connected, projections originating
Widely connected, projections originating mostly
mostly from upper layers
from deep layers
Neurotransmitter
Specific, more inhibitory, ionotropic
Diverse, more excitable, metabotropic
receptors
Maturation
Functions
Late prenatal formation, early myelination,
Early prenatal formation, late myelination,
truncated plasticity
prolonged plasticity
Sensorimotor representations, 高的
spatiotemporal resolution
Abstract representations, coarse spatiotemporal
resolution
笔记. These descriptions reflect characteristic features at the extreme end points of the cortical spectrum. Intermediate areas possess the features in a more
graded manner.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
4
9
5
0
2
0
5
6
2
7
0
n
e
n
_
A
_
0
0
2
5
6
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
数字 2. A multidimensional axis of mammalian and human cortical cytoarchitecture and connectivity. Cortical areas of the human brain
are arranged by their cytoarchitectonic spectrum, from dense areas on the outside to more sparsely populated areas on the inside. 这些
cellular densities are well correlated with other macroscopic and microscopic morphological features of cortical areas, such as laminar
differentiation, cortical thickness, soma size, and spine density of pyramidal neurons (例如, Beul & 希尔格塔格, 2019; John et al., 2021; van
den Heuvel et al., 2015); compare Table 1. 而且, as depicted in the lower panel, the existence or absence of connections is related to
the similarity of cellular densities of the areas, as described by the Structural Model of Connections and Architectonic Type Principle (García-
Cabezas et al., 2019; Hilgetag et al., 2019), with connections predominantly linking areas of a similar structure type, and the areas on the
inside forming a densely connected core. The structural gradient also relates to the laminar organization of projection origins and terminations.
具体来说, outside areas project to inside areas predominantly from the supragranular (upper) cortical layers. 反过来, inside areas project
to outside areas predominantly from infragranular (深的) cortical layers, while areas of a similar type show a more balanced bilaminar pattern
of projection origins. Compared to such arrangements for other mammalian species, such as the mouse and macaque monkey, the human
arrangement possesses an expanded cytoarchitectonic gradient, as is apparent in the larger number of concentric circles than in other species
(比照. Goulas et al., 2019; Hilgetag et al., 2019). 而且, it shows a clear cytoarchitectonic differentiation between the core and the periphery
of the cortical connectome. Figure adapted from Goulas et al. (2019), Changeux et al. (2021).
网络神经科学
954
A natural cortical axis of the human brain
of the different kinds of cortical connection “hierarchies” described by Hilgetag and Goulas
(2020), including feature gradients, projection sequences, orderings of directed “feedforward”
and “feedback” projections, as well as hierarchical modular connectivity. The latter feature is
represented by the core network module formed by the inside areas, which possess many
long-distance projections. The present diagram also aligns with the sequence of synaptic steps
from the outside to the inside of the brain, similar to the influential diagram of Mesulam (1998),
providing an intuitive yet multidimensional representation of the natural axis of external to
internal cortical organization.
It is important to note that the alignment between the various structural and functional fea-
ture gradients is not perfect. 的确, Sydnor et al. (2021) presented associations among 10
cortical features, including tissue histology, evolutionary expansion, metabolism, gene expres-
锡安, and function, that were widely differing in strength. Disagreements between feature gra-
dients may arise from experimental noise, the linkage of features through indirect mechanisms,
or unknown common factors rather than by direct relations, from misinterpretation, or by
actual divergence, such as suggested by Paquola et al. (2019) for the increasing divergence
between histological and functional gradients in transmodal cortices.
因此, we do not think that all these cortical features are synonymous or directly related. A
better understanding of their interrelations will be achieved by investigating and establishing
the actual mechanisms that link the different aspects. Of particular relevance in this context is
research into principles of cortical development and plasticity that will have to clarify how
these features are mechanistically related, and will thus move beyond correlational studies.
THE NATURAL AXIS OF THE HUMAN BRAIN
A natural axis of organization is present to a varying extent across the cerebral cortex of
different mammalian species (Goulas et al., 2019). The natural axis of the human brain differs
from that of other species in that it is expanded toward the outside, and these expanded
outside regions possess a more elaborate cytoarchitecture and projection systems specifically
in the upper cortical layers (数字 2).
Compared to other primate species, the human brain forms along a developmental trajec-
tory of prolonged pre- and postnatal development. This results in an expansion of late-
developing structures, which concerns mostly “outside” areas of the dorso-lateral surface (爬坡道
等人。, 2010), as well as their late-developing laminar compartments, 那是, particularly their
upper layers. These areas and layers become large and very densely populated with neurons,
also leading to a high laminar differentiation. Due to the association of these fundamental
architectonic features with other structural, connectional, and functional aspects along the
natural axis, the elongation of the human axis has direct consequences for the organization
of the human connectome and ultimately human brain function.
一般来说, human cortical organization becomes more multistaged and more differentiated
at each of the stages. This also results in a deeper organization of cortical networks. 而且,
the expansion of outside areas is associated with a shift of large pyramidal projection neurons
to the upper layers of these areas, forming an enhanced system for processing signals from the
expanded sensory surface (Goulas et al., 2018). 同时, outside areas become con-
nectionally segregated from each other, due to expanded brain size and the increasing sparsity
and localization of connections in the human brain (Changeux et al., 2021). 所以, 这些
regions are not directly connected with each other as they are in other species, but commu-
nicate with each other via the densely connected network core (数字 2).
网络神经科学
955
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
4
9
5
0
2
0
5
6
2
7
0
n
e
n
_
A
_
0
0
2
5
6
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
A natural cortical axis of the human brain
Deep learning networks:
Artificial neuronal networks with
more than one intermediate layer
between inputs and outputs.
Global Neuronal Workspace:
A distributed core-periphery network
architecture for computations
underlying conscious and
unconscious cognitive functions.
FUNCTIONAL IMPLICATIONS
The concept of a unified cortical axis can be used to explain some of the special structural and
functional features of the human brain. As a central feature, there is an expansion of this nat-
ural axis in the human brain due to prolonged brain development. This extension corresponds
to the existence of further distinct stages, in terms of architecture, 连接性, and function, 在
the human brain compared to the brains of other species (比照. Hilgetag et al., 2019). Such an
expansion of the cortical axis has a number of functional implications.
Foremost, due to the addition of areas, the human brain connectome possesses more pro-
cessing stages, generally resulting in an increased depth of processing. Functionally, this likely
results in increased representational depth, more accurate representations as well as more
abstract representations at the stage of inside areas, similar to what is observed in artificial
神经网络, particularly deep learning networks (LeCun et al., 2015; Wyss et al., 2006).
There are also more specialized stages (due to the greater architectonic differentiation), 哪个
can form the basis of elaborate multiscale representations.
The elongation also results in greater separation of the different processing stages, and partic-
ularly in the separation of outside areas representing different sensory modalities, 比如视觉上的
and auditory, that are no longer directly connected, but only via the network core of inside areas.
This means that separate representations can be kept apart more stably, with less interference. 这
segregation implies reduced perturbation and greater stability of representations of the different
sensory modalities, and may particularly support cognitive functions such as working memory
(Rodriguez et al., 2019). The segregation is balanced by hierarchical modular connectivity that
facilitates recursive integration and scaling (Moretti & Muñoz, 2013). 尤其, 中央
modules in this hierarchical modular connectivity are formed by the widely connected network
core of inside areas. The clear architectonic differentiation of the core-periphery network structure
in the human brain likely provides the basis of specialized representations of the sensory and
motor interface with the world that is being integrated through the core of inside areas.
The prolonged developmental expansion of the outside areas results in more detailed sen-
sory representations as well as a more intricate motor interface, which is essential for increased
dexterity and writing as well as speaking. 此外, these outside areas possess an elaborate
system of enlarged projection neurons in the upper cortical layers that can serve to support fast
sensorimotor interactions with the outside world as well as underlie higher cognitive functions
such as language and intelligence (Heyer et al., 2022). These enhanced outside-to-inside pro-
jections are balanced by a system of long-range connectivity formed by the network core of
inside areas, which is hypothesized to be engaged as an important anatomical component of
the Global Neuronal Workspace for conscious access (Dehaene et al., 1998).
Further studies of this kind will need to determine the relevance of individual and combined
connection features for brain function. 尤其, computational studies may provide an
avenue for isolating the influence of individual connectomic fundamentals and studying them
in selected combinations in a more accessible way than is possible in experimental research.
总之, the concept of the natural axis of human cortical organization provides a flex-
ible framework for integrating a multitude of structural, 发展性的, and evolutionary
aspects and relating them to the extraordinary cognitive abilities of the human brain.
作者贡献
克劳斯·C. 希尔格塔格: 概念化; 资金获取; Writing – original draft; Writing –
review & 编辑. Alexandros Goulas: 概念化; 可视化; 写作——原创
网络神经科学
956
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
4
9
5
0
2
0
5
6
2
7
0
n
e
n
_
A
_
0
0
2
5
6
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
A natural cortical axis of the human brain
草稿; 写作——复习 & 编辑. Jean-Pierre Changeux: 概念化; Funding acquisi-
的; Writing – original draft; 写作——复习 & 编辑.
资金信息
克劳斯·C. 希尔格塔格, Human Brain Project, 奖项ID: SGA2, SGA3. Jean-Pierre Changeux, 人类
Brain Project, 奖项ID: SGA2, SGA3. Alexandros Goulas, 德国研究基金会
(https://dx.doi.org/10.13039/501100001659), 奖项ID: SPP 2041. 克劳斯·C. 希尔格塔格,
德国研究基金会 (https://dx.doi.org/10.13039/501100001659), 奖项ID:
SPP 2041. 克劳斯·C. 希尔格塔格, 德国研究基金会 (https://dx.doi.org/10.13039
/501100001659), 奖项ID: SFB 936/A1. 克劳斯·C. 希尔格塔格, Deutsche Forschungsge-
meinschaft (https://dx.doi.org/10.13039/501100001659), 奖项ID: TRR 169/A2. 克劳斯·C.
希尔格塔格, 德国研究基金会 (https://dx.doi.org/10.13039/501100001659),
奖项ID: SFB 1461/A4.
参考
Abbie, A. A. (1940). Cortical lamination in the Monotremata. 这
Journal of Comparative Neurology, 72(3), 429–467. https://土井
.org/10.1002/cne.900720302
Barbas, H. (1986). Pattern in the laminar origin of corticocortical
连接. The Journal of Comparative Neurology, 252(3),
415–422. https://doi.org/10.1002/cne.902520310, 考研:
3793985
Barbas, H。, & García-Cabezas, 中号. Á. (2015). Motor cortex layer 4:
Less is more. Trends in Neurosciences, 38(5), 259–261. https://
doi.org/10.1016/j.tins.2015.03.005, 考研: 25868984
Barbas, H。, & Rempel-Clower, 氮. (1997). Cortical structure predicts
the pattern of corticocortical connections. 大脑皮层, 7(7),
635–646. https://doi.org/10.1093/cercor/7.7.635, 考研:
9373019
Beul, S. F。, Goulas, A。, & 希尔格塔格, C. C. (2018). Comprehensive
computational modelling of the development of mammalian
cortical connectivity underlying an architectonic type principle.
公共科学图书馆计算生物学, 14(11), e1006550. https://doi.org
/10.1371/journal.pcbi.1006550, 考研: 30475798
Beul, S. F。, & 希尔格塔格, C. C. (2019). Neuron density fundamentally
relates to architecture and connectivity of the primate cerebral
cortex. 神经影像, 189, 777–792. https://doi.org/10.1016/j
.neuroimage.2019.01.010, 考研: 30677500
Beul, S. F。, & 希尔格塔格, C. C. (2020). Systematic modelling of the
development of laminar projection origins in the cerebral cortex:
Interactions of spatio-temporal patterns of neurogenesis and
cellular heterogeneity. 公共科学图书馆计算生物学, 16(10),
e1007991. https://doi.org/10.1371/journal.pcbi.1007991,
考研: 33048930
Broca, 中号. 磷. (1861). Remarques sur le siège de la faculté du langage
articulé, suivies d’une observation d’aphémie (perte de la parole).
Bulletin de la Société Anatomique, 6, 330–357.
巴克纳, 右. L。, & 克里宁, F. 中号. (2013). The evolution of distributed
association networks in the human brain. Trends in Cognitive
科学, 17(12), 648–665. https://doi.org/10.1016/j.tics.2013
.09.017, 考研: 24210963
尚古克斯, J.-P. (2017). Climbing brain levels of organisation from
genes to consciousness. 认知科学的趋势, 21(3),
168–181. https://doi.org/10.1016/j.tics.2017.01.004, 考研:
28161289
尚古克斯, J.-P. (2022). Epigenesis, synapse selection, 文化
imprints, and brain development: From molecules to cognition.
In The Cambridge handbook of cognitive development.
剑桥, 英国: 剑桥大学出版社. https://doi.org/10
.1017/9781108399838.004
尚古克斯, J.-P., Goulas, A。, & 希尔格塔格, C. C. (2021). A Connec-
tomic hypothesis for the hominization of the brain. Cerebral
Cortex, 31(5), 2425–2449. https://doi.org/10.1093/cercor
/bhaa365, 考研: 33367521
德阿内, S。, Kerszberg, M。, & 尚古克斯, J.-P. (1998). A neuronal
model of a global workspace in effortful cognitive tasks. Proceed-
ings of the National Academy of Sciences of the United States of
美国, 95(24), 14529–14534. https://doi.org/10.1073/pnas.95
.24.14529, 考研: 9826734
Dombrowski, S. M。, 希尔格塔格, C. C。, & Barbas, H. (2001). Quanti-
tative architecture distinguishes prefrontal cortical systems in the
rhesus monkey. 大脑皮层, 11(10), 975–988. https://doi.org
/10.1093/cercor/11.10.975, 考研: 11549620
Elston, G. 氮. (2002). Cortical heterogeneity: Implications for visual
processing and polysensory integration. Journal of Neurocytol-
奥吉, 31, 317–335. https://doi.org/10.1023/A:1024182228103,
考研: 12815250
Gall, F. J。, & Spurzheim, J. G. (1810). Anatomie et physiologie du
système nerveux en général, et du cerveau en particulier, avec
des observations sur la possibilité de reconnoître plusieurs
dispositions intellectuelles et morales de l’homme et des animaux
par la configuration de leurs têtes (卷. 1–4). 巴黎, 法国: F.
Schoell.
García-Cabezas, 中号. Á., Hacker, J. L。, & Zikopoulos, 乙. (2020). A
protocol for cortical type analysis of the human neocortex
applied on histological samples, the atlas of Von Economo and
Koskinas, and magnetic resonance imaging. Frontiers in Neuro-
anatomy, 14, 576015. https://doi.org/10.3389/fnana.2020
.576015, 考研: 33364924
García-Cabezas, 中号. Á., Zikopoulos, B., & Barbas, H. (2019). 这
structural model: A theory linking connections, plasticity,
网络神经科学
957
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
t
/
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
4
9
5
0
2
0
5
6
2
7
0
n
e
n
_
A
_
0
0
2
5
6
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
A natural cortical axis of the human brain
pathology, development and evolution of the cerebral cortex.
Brain Structure and Function, 224(3), 985–1008. https://doi.org
/10.1007/s00429-019-01841-9, 考研: 30739157
Goulas, A。, 尚古克斯, J.-P., Wagstyl, K., Amunts, K., Palomero-
加拉格尔, N。, & 希尔格塔格, C. C. (2021). The natural axis of
transmitter receptor distribution in the human cerebral cortex.
Proceedings of the National Academy of Sciences of the United
States of America, 118(3), e2020574118. https://doi.org/10.1073
/pnas.2020574118, 考研: 33452137
Goulas, A。, Majka, P。, Rosa, 中号. G. P。, & 希尔格塔格, C. C. (2019). A
blueprint of mammalian cortical connectomes. 公共科学图书馆生物学,
17(3), e2005346. https://doi.org/10.1371/journal.pbio.2005346,
考研: 30901324
Goulas, A。, Zilles, K., & 希尔格塔格, C. C. (2018). Cortical gradients
and laminar projections in mammals. Trends in Neurosciences,
41(11), 775–788. https://doi.org/10.1016/j.tins.2018.06.003,
考研: 29980393
Hendy, J. P。, Takahashi, E., 范德库韦, A. J。, & Charvet, C. J.
(2020). Brain wiring and supragranular-enriched genes linked to
protracted human frontal cortex development. 大脑皮层,
30(11), 5654–5666. https://doi.org/10.1093/cercor/ bhaa135,
考研: 32537628
Heyer, D. B., Wilbers, R。, Galakhova, A. A。, Hartsema, E., Braak, S。,
打猎, S。, … Goriounova, 氮. A. (2022). Verbal and general IQ
associate with supragranular layer thickness and cell properties
of the left temporal cortex. 大脑皮层, 32(11), 2343–2357.
https://doi.org/10.1093/cercor/bhab330, 考研: 34550325
希尔格塔格, C. C。, Beul, S. F。, van Albada, S. J。, & Goulas, A. (2019).
An architectonic type principle integrates macroscopic cortico-
cortical connections with intrinsic cortical circuits of the primate
脑. 网络神经科学, 3(4), 905–923. https://doi.org/10
.1162/netn_a_00100, 考研: 31637331
希尔格塔格, C. C。, & Goulas, A. (2020). ‘Hierarchy’ in the organiza-
tion of brain networks. Philosophical Transactions of the Royal
Society B: Biological Sciences, 375(1796). https://doi.org/10
.1098/rstb.2019.0319, 考研: 32089116
爬坡道, J。, Inder, T。, Neil, J。, Dierker, D ., Harwell, J。, & Van Essen, D.
(2010). Similar patterns of cortical expansion during human
development and evolution. Proceedings of the National Acad-
emy of Sciences of the United States of America, 107(29),
13135–13140. https://doi.org/10.1073/pnas.1001229107,
考研: 20624964
Hopf, A. (1955). Über die Verteilung myeloarchitektonischer Merk-
male in der isokortikalen Schläfenlappenrinde beim Menschen.
Journal für Hirnforsch, 2, 36–54.
Hopf, A. (1956). Über die Verteilung myeloarchitektonischer
Merkmale in der Stirnhirnrinde beim Menschen. Journal für
Hirnforsch, 2, 311–333.
Hopf, A。, & Vitzthum, H. G. (1957). Über die Verteilung myeloarch-
itektonischer Merkmale in der Scheitellappenrinde beim
Menschen. Journal für Hirnforsch, 3, 79–104. https://doi.org/10
.1515/9783112522387-001
Huntenburg, J. M。, Bazin, P.-L., & 马古利斯, D. S. (2018). 大的-
scale gradients in human cortical organization. Trends in Cogni-
tive Sciences, 22(1), 21–31. https://doi.org/10.1016/j.tics.2017
.11.002, 考研: 29203085
约翰, 是. J。, Zikopoulos, B., García-Cabezas, 中号. Á., & Barbas, H.
(2021). The Cortical Spectrum: A robust structural continuum
in primate cerebral cortex revealed by histological staining and
magnetic resonance imaging. bioRxiv. https://doi.org/10.1101
/2021.09.09.459678
乐存, Y。, 本吉奥, Y。, & 欣顿, G. (2015). 深度学习. 自然,
521(7553), 436–444. https://doi.org/10.1038/nature14539,
考研: 26017442
Mesulam, 中号. 中号. (1998). From sensation to cognition. Brain, 121(Pt 6),
1013–1052. https://doi.org/10.1093/brain/121.6.1013, 考研:
9648540
Moretti, P。, & Muñoz, 中号. A. (2013). Griffiths phases and the stretching
of criticality in brain networks. Nature Communications, 4, 2521.
https://doi.org/10.1038/ncomms3521, 考研: 24088740
穆雷, J. D ., Bernacchia, A。, 弗里德曼, D. J。, 罗莫, R。, Wallis,
J. D ., Cai, X。, Padoa-Schioppa, C。, Pasternak, T。, Seo, H。, 李, D .,
& 王, X.-J. (2014). A hierarchy of intrinsic timescales across
primate cortex. 自然神经科学, 17(12), 1661–1663. https://
doi.org/10.1038/nn.3862, 考研: 25383900
Nieuwenhuys, R。, & Broere, C. A. J. (2017). A map of the human
neocortex showing the estimated overall myelin content of the
individual architectonic areas based on the studies of Adolf
Hopf. Brain Structure and Function, 222(1), 465–480. https://
doi.org/10.1007/s00429-016-1228-7, 考研: 27138385
Nieuwenhuys, R。, & Puelles, L. (2016). Towards a new neuromor-
phology. 占婆, 瑞士: Springer International Publishing.
https://doi.org/10.1007/978-3-319-25693-1
Pandya, D. N。, & Sanides, F. (1973). Architectonic parcellation of
the temporal operculum in rhesus monkey and its projection
pattern. Zeitschrift Für Anatomie Und Entwicklungsgeschichte,
139(2), 127–161. https://doi.org/10.1007/ BF00523634,
考研: 4197942
Paquola, C。, Vos De Wael, R。, Wagstyl, K., Bethlehem, 右. A. 我。,
洪, S.-J., Seidlitz, J。, 布莫尔, 乙. T。, 埃文斯, A. C。, Misic, B.,
马古利斯, D. S。, Smallwood, J。, & Bernhardt, 乙. C. (2019).
Microstructural and functional gradients are increasingly dissoci-
ated in transmodal cortices. 公共科学图书馆生物学, 17(5), e3000284.
https://doi.org/10.1371/journal.pbio.3000284, 考研:
31107870
Petanjek, Z。, Sedmak, D ., Džaja, D ., Hladnik, A。, Rašin, 中号. R。, &
Jovanov-Milosevic, 氮. (2019). The protracted maturation of
associative layer IIIC pyramidal neurons in the human prefrontal
cortex during childhood: A major role in cognitive development
and selective alteration in autism. Frontiers in Psychiatry, 10, 122.
https://doi.org/10.3389/fpsyt.2019.00122, 考研: 30923504
罗德里格斯, N。, Izquierdo, E., & Ahn, Y.-Y. (2019). Optimal modularity
and memory capacity of neural reservoirs. 网络神经科学,
3(2), 551–566. https://doi.org/10.1162/netn_a_00082, 考研:
31089484
Sanides, F. (1962). Die Architektonik des Menschlichen Stirnhirns.
柏林, 德国: 施普林格. https://doi.org/10.1007/978-3-642
-86210-6
Sanides, F. (1970). Functional architecture of motor and sensory
cortices in primates in the light of a new concept of neocortex
evolution.
In The primate brain: Advances in primatology
(PP. 137–208). 纽约, 纽约: Appleton-Century-Crofts.
Shipp, S. (2005). The importance of being agranular: A comparative
account of visual and motor cortex. Philosophical Transactions
乙, 360(1456), 797–814. https://doi.org/10.1098/rstb.2005.1630,
考研: 15937013
网络神经科学
958
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
/
t
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
4
9
5
0
2
0
5
6
2
7
0
n
e
n
_
A
_
0
0
2
5
6
p
d
t
.
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
A natural cortical axis of the human brain
Sydnor, V. J。, Larsen, B., Bassett, D. S。, Alexander-Bloch, A。, Fair,
D. A。, Liston, C。, … Satterthwaite, 时间. D. (2021). Neurodevelopment
of the association cortices: Patterns, mechanisms, and implica-
tions for psychopathology. 神经元, 109(18), 2820–2846. https://
doi.org/10.1016/j.neuron.2021.06.016, 考研: 34270921
van den Heuvel, 中号. P。, Scholtens, L. H。, Feldman Barrett, L。, 希尔格塔格,
C. C。, & Reus, 中号. A. (2015). Bridging cytoarchitectonics and
connectomics in human cerebral cortex. 神经科学杂志,
35(41), 13943–13948. https://doi.org/10.1523/JNEUROSCI.2630
-15.2015, 考研: 26468195
Vogt, C。, & Vogt, 氧. (1919). Allgemeine Ergebnisse unserer Hirn-
forschung. 莱比锡, 德国: J. A. Barth.
von Economo, C. F. (1927). Zellaufbau der Grosshirnrinde des
Menschen. 柏林, 德国: 施普林格.
von Economo, C。, & Koskinas, G. 氮. (1925). Die Cytoarchitektonik der
Hirnrinde des erwachsenen Menschen. 柏林, 德国: 施普林格.
Wagstyl, K., Ronan, L。, Goodyer, 我. M。, & 弗莱彻, 磷. C. (2015).
Cortical thickness gradients in structural hierarchies. Neuro-
图像, 111, 241–250. https://doi.org/10.1016/j.neuroimage
.2015.02.036, 考研: 25725468
维斯, R。, König, P。, & Verschure, 磷. F. 中号. J. (2006). A model of the
ventral visual system based on temporal stability and local mem-
奥里. 公共科学图书馆生物学, 4(5), e120. https://doi.org/10.1371/journal
.pbio.0040120, 考研: 16605306
Zilles, K., & Palomero-Gallagher, 氮. (2017). Multiple transmitter
receptors in regions and layers of the human cerebral cortex.
Frontiers in Neuroanatomy, 11, 78. https://doi.org/10.3389/fnana
.2017.00078, 考研: 28970785
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
t
/
e
d
你
n
e
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
/
6
4
9
5
0
2
0
5
6
2
7
0
n
e
n
_
A
_
0
0
2
5
6
p
d
.
t
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
网络神经科学
959