Disposed to Distraction: Genetic Variation in the
Cholinergic System Influences Distractibility
But Not Time-on-Task Effects
Anne S. Berry1, Elise Demeter2, Surya Sabhapathy3, Brett A. English4,
Randy D. Blakely4, Martin Sarter1, and Cindy Lustig1
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
抽象的
■ Both the passage of time and external distraction make it
difficult to keep attention on the task at hand. We tested the
hypothesis that time-on-task and external distraction pose
independent challenges to attention and that the brainʼs cho-
linergic system selectively modulates our ability to resist dis-
牵引力. Participants with a polymorphism limiting cholinergic
容量 (Ile89Val variant [rs1013940] of the choline transporter
gene SLC5A7) and matched controls completed self-report
measures of attention and a laboratory task that measured
decrements in sustained attention with and without distrac-
的. We found evidence that distraction and time-on-task
effects are independent and that the cholinergic system is
strongly linked to greater vulnerability to distraction. Ile89Val
participants reported more distraction during everyday life
than controls, and their task performance was more severely
impacted by the presence of an ecologically valid video dis-
tractor (similar to a television playing in the background).
These results are the first to demonstrate a specific impair-
ment in cognitive control associated with the Ile89Val poly-
morphism and add to behavioral and cognitive neuroscience
studies indicating the cholinergic systemʼs critical role in over-
coming distraction. ■
介绍
“Pay attention!” You may have given yourself this exhor-
tation during a long, boring drive as you realized both
your mind and your car were drifting off the road or
received it from your conversation partner when they
noticed your eyes wandering toward an attractive stimu-
lus walking by. Both extended task periods and external
distractors challenge our ability to remain focused on the
task at hand, and individuals with psychiatric disorders
such as schizophrenia and attention deficit hyperactivity
紊乱 (ADHD) may be especially vulnerable to such
挑战. Effective treatment of these vulnerabilities
will require a better understanding of whether they reflect
general declines in cognitive control or more specific pro-
cessing deficits and of the underlying neural systems that
may be targets for pharmaceutical intervention. Here we
present evidence that performance declines caused by
time-on-task demands and external distraction are inde-
pendent and that the cholinergic system plays an especially
important role in resisting external distraction.
直观地, one might expect that any demand on
cognitive control, regardless of its source, would reduce
表现, and that compounding such demands
would have an especially detrimental effect. 那是,
1密歇根大学, 2杜克大学, 3University of Michi-
gan Medical School, 4Vanderbilt University School of Medicine
© 2014 麻省理工学院
external distractors might be especially tempting when
one is already having difficulty maintaining attention on
an attended task. 反过来, competition from external dis-
tractors may make it especially difficult to maintain focus,
causing steeper time-on-task declines. 然而, the reality
is much more complex. Vulnerability to external distraction
is typically increased by cognitive load but decreased by
perceptual load (Lavie, 2010; Lavie, Hirst, de Fockert, &
Viding, 2004). Time-on-task performance declines have
been attributed by some to overloads of attention and by
others to underloads of attention (see discussion by Pattyn,
Neyt, Heridericlcx, & Soetens, 2008), suggesting opposite
predictions for the effects of adding an additional cognitive
load in the form of external distraction.
Previous attempts to test potential interactions between
distraction and time-on-task effects have yielded ambigu-
ous results. Rosenberg, Noonan, DeGutis, and Esterman
(2013) used a continuous performance test (detect female
target faces vs. male nontarget faces) that showed sig-
nificant declines in both accuracy and RT stability over
a 12-min period and manipulated whether or not the
background consisted of distracting scenes. This manip-
ulation did not affect performance overall or the slope of
time-on-task declines, but did alter individual differences
因素: Only in the distraction condition did mind-
fulness scores predict time-on-task declines. In another
学习, Breckel, Giessing, and Thiel (2011) added motion
distractors (moving vs. stationary dots) to a simple signal
认知神经科学杂志 26:9, PP. 1981–1991
土井:10.1162/jocn_a_00607
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
6
/
9
2
6
1
/
9
9
8
/
1
1
1
9
9
8
4
1
7
/
5
1
7
7
9
8
哦
1
C
5
n
9
_
2
A
/
_
j
0
哦
0
C
6
n
0
7
_
A
p
_
d
0
0
乙
6
y
0
G
7
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
.
/
F
t
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
检测任务. 然而, these “distractors” did not affect
time-on-task declines on the minute scale and para-
doxically alleviated performance declines associated with
long intertarget intervals on no distractor (stationary)
试验. The distraction manipulation also had no effect on
brain activity (measured using fMRI) related to time-on-
task effects and intertarget interval. Looking in the other
方向, activity related to processing the motion dis-
tractors was not affected by time-on-task, and there were
mixed results in regions involved in motion processing
(some areas increasing, others decreasing) as a function
of intertarget interval.
简而言之, the bulk of the evidence from these studies
suggests that time-on-task and external distractor effects
are independent. 然而, the lack of an overall perfor-
mance decrement as a result of the nominal distractor
manipulations in these studies reduces the support for
this conclusion. 那是, without a reliable main effect
of distraction, it is hard to know whether distractor effects
might interact with time-on-task effects. The neuro-
imaging data are also somewhat ambiguous, as it is hard
to distinguish which changes represent the engagement
of cognitive control to inhibit distractor processing from
those that may represent increased distractor process-
ing because such inhibition has failed (see also Demeter,
Hernandez-Garcia, Sarter, & Lustig, 2011).
We therefore took a different approach, applying a
manipulation that creates robust distractor effects to a
task with rapid time-on-task performance declines and
asked whether time-on-task and distractor effects might
be related to different neuromodulatory systems. 这
approach also has a translational advantage: Developing
effective drug treatments for specific aspects of atten-
tional function will require understanding their links to
the neuromodulator systems targeted by such treatments.
Time-on-task effects have been linked to genetic variation
in the dopaminergic system (Lim et al., 2012); here we
asked whether distraction effects might be linked to
genetic variation in the cholinergic system.
The basal forebrain cholinergic system projects through-
out neocortex and modulates several neurocognitive func-
系统蒸发散, including both perceptual and goal-driven aspects of
注意力 (例如, Rokem, Landau, Garg, Prinzmetal, & Silver,
2010; Silver, Shenhav, & DʼEsposito, 2008; see Demeter &
Sarter, 2013, for a recent review). Although the cholinergic
system has been traditionally described as a diffuse neuro-
modulatory system contributing to arousal, plasticity, 和
improved signal-to-noise ratio (see Picciotto, Higley, &
Mineur, 2012, for a recent review of this evidence), current
models suggest that acetylcholine is also capable of medi-
ating specific cognitive operations through spatially re-
stricted signaling (Hasselmo & Sarter, 2011).
尤其, rodent studies indicate that right-lateralized
frontoparietal cholinergic circuitry mediates the ability to
resist distraction (英石. Peters, Demeter, Lustig, Bruno, &
Sarter, 2011; Broussard, Karelina, Sarter, & Givens, 2009;
Gill, Sarter, & Givens, 2000). When humans are tested in
the same paradigm, they show increased right prefron-
tal activation during the distraction condition, parallel-
ing rodentsʼ increase in right prefrontal acetylcholine
级别. Translational studies using human clinical popu-
lations (例如, Demeter, 格思里, 泰勒, Sarter, & Lustig,
2013; Pa et al., 2013) and animal models (see reviews by
Lustig, Kozak, Sarter, Young, & Robbins, 2012; Sarter,
Martinez, & Kozak, 2009) also suggest that cholinergic
pathways play a critical role in the increased vulnerabil-
ity to distraction seen in neurological disorders includ-
ing schizophrenia and mild cognitive impairment.
We tested the hypothesis that increased vulnerability
to distraction would be associated with genetic variation
affecting the high-affinity choline transporter (CHT),
which is responsible for transporting choline into the
nerve terminal for the synthesis of acetylcholine (ACh).
Mice with reduced CHT expression have normal ACh
release and choline clearance at baseline but are impaired
on both these measures in response to task demands
(Parikh, 英石. Peters, Blakely, & Sarter, 2013). 在人类中,
the Ile89Val variant (rs1013940) of the CHT gene SLC5A7
reduces the rate of choline transport by approximately
40–60% compared with the major allele (Okuda, Okamura,
Kaitsuka, Haga, & Gurwitz, 2002). The frequency of
the Ile89Val variant in normal White participants in the
美国, ∼6% (English et al., 2009), is equivalent to
a prevalence of over 10 million individuals. This variant
occurs more often in ADHD patients than controls and
has been linked to greater symptom severity in depressed
患者 (English et al., 2009; Hahn et al., 2008).
The link to depression suggests that individuals with
the Ile89Val allele might be more likely to engage in
rumination and mind-wandering but based on previous
human and animal data linking the cholinergic system
to distraction, we expected an increased susceptibility
to distraction to be their major difference from control
participants without the allele. To preview our results,
Ile89Val participants showed an increased vulnerability
to distraction on both self-report measures and labora-
tory task performance but were spared on other mea-
sures of attentional control, including those involved in
maintaining performance over time. These results indi-
cate that time-on-task demands and external distraction
tax at least partially dissociable components of cognitive
control and that the cholinergic system plays a particu-
larly important role in resisting distraction.
EXPERIMENT 1
方法
参加者
Six hundred seventeen individuals recruited from the
greater Ann Arbor community completed the Poor Atten-
tional Control questionnaire (Huba, 歌手, Aneshensel,
& Antrobus, 1982; see description below) and contrib-
uted saliva samples for genotyping. From this larger pool,
1982
认知神经科学杂志
体积 26, 数字 9
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
6
/
9
2
6
1
/
9
9
8
/
1
1
1
9
9
8
4
1
7
/
5
1
7
7
9
8
哦
1
C
5
n
9
_
2
A
/
_
j
0
哦
0
C
6
n
0
7
_
A
p
_
d
0
0
乙
6
y
0
G
7
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
F
/
t
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
67 Ile89Val heterozygotes were identified and compared
with age, 性别, and education-matched controls. 看
桌子 1 for demographics.
have reasonable discriminant validity (average correlation
between subscale total and items not in that subscale all
r < .49 compared with items in that subscale all r > .72).
Genotyping Methods
Results and Discussion
CHT SNP genotyping was carried out using the proce-
dures described in English et al. (2009). Briefly, DNA was
extracted from saliva samples using a commercial DNA
isolation kit (Gentra Systems, 明尼阿波利斯, 明尼苏达州) as pre-
viously described (Mazei-Robinson, Couch, Shelton,
斯坦因, & Blakely, 2005). An allelic discrimination assay
was performed in the Vanderbilt Center for Human
Genetics Research DNA Resources Core using TaqMan
SNP Genotyping Assay reagents (Applied Biosystems,
公司, Foster City, CA). Four nanograms of DNA were used
as template in reactions containing 1× TaqMan Universal
PCR Master Mix and 900 nM forward (50-TGTACCAGGT-
TATGGCCTAGCTT-30) and reverse (50-ACTGAGATTTGC-
ACTTTCACTTACCT-30) amplification primers, 200 nM VIC
(50-CAGGCACCAATTGGATA-30) and FAM (50-AGGCACC-
AGTTGGATA-30) dye-labeled probes. Thermal cycling
(95°C for 10 min, 其次是 50 cycles of 92°C for 15 秒
and 60°C for 1 min) and product detection were accom-
plished using the ABI 7900HT Real-Time PCR System
(ABI). Quality control analyses showed that the results
were consistent with recommended guidelines (例如,
Edenberg & 刘, 2009). The call rate (rate of at which
samples could be successfully assigned a genotype) 在
our sample was >95% with TaqMan, 和 100% consis-
tency when TaqMan was compared with gel-based geno-
typing. No-call TaqMan samples were reanalyzed using
gel-based genotyping.
Attention Questionnaire Measure
Trait attention. Participants completed 36 items from
the Imaginal Processes Inventory (歌手 & Antrobus,
1970). Each item consisted of a statement about cog-
nitive function in everyday life (例如. “I find it difficult to
concentrate when the TV or radio is on”), and partici-
pants rated the degree to which they identified with each
statement on a scale from 1 到 5. Our analyses focus on
这 15 items that make up the Poor Attentional Control
(PAC) subscale identified in a later factor analysis (Huba
等人。, 1982).
The PAC has good internal consistency (coefficient
alpha = .83) and test–retest reliability (r = .73; see also
Tanaka & Huba, 1985/1986). It can be subdivided into
subscales (five questions each) of distractibility, 头脑-
wandering, and boredom. Although Huba et al. (1982)
do not provide psychometric data on these subscales,
analyses of a large data set from our laboratory (N=
510; see Berry, 李, 林, & Lustig, 2014) indicate good
internal consistency within subscales (mind-wandering
coefficient alpha = .84, distraction coefficient alpha =
.79, boredom coefficient alpha = .77). The subscales also
As illustrated in Table 1, Ile89Val participants reported
experiencing more distractibility and mind-wandering in
everyday life than did controls, but not more boredom.
The groupsʼ equivalence on boredom is important, as it
indicates that Ile89Val participants were not simply inter-
preting the scale differently and marking all items higher
than controls.
The Ile89Val participantsʼ higher ratings for distraction
were in line with our predictions based on the cholin-
ergic systemʼs role in resisting distraction as described in
the Introduction. The higher ratings for mind-wandering
were somewhat less expected. 然而, in everyday life,
it may be difficult to disentangle the subjective experi-
ences of mind-wandering and distraction. 例如,
in factor analyses “I notice all the other things around
me I could be doing” groups with the distraction fac-
tor and involves external distractors whereas “thoughts
unrelated to my work always creep in” groups with the
mind-wandering factor and involves only internal experi-
恩塞斯. Despite these distinctions on a principled basis
(external vs. 内部的) and in large-scale factor analyses,
it is easy to see how they might overlap or become con-
fused in individual participantsʼ self-reports.
桌子 1. Demographics and Self-reported Everyday Attention
Function (PAC Measures; Huba et al., 1982) for Ile89Val
Participants and Controls
控制
Ile89Val
t Test
Effect Size
(Cohenʼs d)
年龄 (年)
中号
标清
Edu (年)
中号
标清
42.49
17.20
16.08
2.50
Distractibility
中号
标清
13.43
4.04
Mind-wandering
中号
标清
Boredom
中号
标清
12.93
3.46
12.64
3.26
42.33
17.33
16.50
3.49
15.16
4.43
14.45
4.77
12.97
4.12
t < 1
p = .96
t < 1
p = .43
t = 2.36
p = .02*
t = 2.16
p = .05*
t < 1
p = .61
d = .01
d = .14
d = .41
d = .36
d = .09
Each group included 67 participants (41 women, 26 men).
*Significant difference between groups.
Berry et al.
1983
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
9
2
6
1
/
9
9
8
/
1
1
1
9
9
8
4
1
7
/
5
1
7
7
9
8
o
1
c
5
n
9
_
2
a
/
_
j
0
o
0
c
6
n
0
7
_
a
p
_
d
0
0
b
6
y
0
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
We therefore conducted a second experiment with a
clearer operationalization of external distraction versus
other, presumably internal challenges to attentional
control and performance, including those that result
from extended time on task. In particular, when asked
about distractibility, Experiment 1 participants regardless
of genotype gave high ratings to items such as “Faced
with a tedious job, I notice all the other things around
me I could be doing” and “I find it difficult to concentrate
when the TV or radio is on.” These responses guided the
design of the experimental task we used in Experiment 2.
Specifically, we added an external distractor to the
Continuous Temporal Expectancy Test (CTET; OʼConnell
et al., 2009) and examined its effect on overall perfor-
mance and time-on-task declines. The CTET is a duration
judgment task: On most trials, a grid pattern rotates
after a standard duration (800 msec), but on target trials
it takes slightly longer (1070 msec). Because the grid
pattern is the same for target and nontarget trials, there
is no bottom–up perceptual change signaling the pre-
sence of a target. Instead, duration judgments require
continuous, focused attention to time and are exqui-
sitely sensitive to fluctuations in attention; in some cases,
performance declines may occur in as few as four trials
(Lustig & Meck, 2005; Penney, Gibbon, & Meck, 2000;
Zakay & Block, 1997). OʼConnell et al. found that the
ability to detect the target declined linearly over 3 min
and that failures to detect the target were predicted up
to 20 sec beforehand by trends in alpha-band activity
(measured using EEG) thought to index mind-wandering.
The CTET thus constituted the “tedious task” described
in the self-report data. To mimic the distraction caused
by the TV or radio, next to the main task computer, we
placed a laptop playing a series of distracting video
clips. This compelling distractor, similar to the situa-
tions that participants described as most distracting in
everyday life, allowed us to test for potential inter-
actions between distraction and time-on-task demands
with greater sensitivity than previous studies that did
not find overall distractor effects. To preview our results,
we found that both distraction and time-on-task had
robust but independent effects on performance and
that Ile89Val participants were especially susceptible to
distraction.
EXPERIMENT 2
Methods
Participants
Thirty-two Ile89Val heterozygotes agreed to return for
additional testing including the CTET with distractor
and more questionnaire measures. The control group
consisted of 32 individuals homozygous for the major
allele and matched with the heterozygotes on age, gen-
der, education, and PAC scores (Table 2). We matched
participants on PAC score to reduce potential concerns
that a finding of increased distractibility by Ile89Val par-
ticipants on the laboratory task might be an artifact
of selection bias. That is, if we had not matched the
samples for this experiment on PAC score, there might
have been concerns that we happened to pick low-
distractibility participants from the control population
and high-distractibility participants from the Ile89Val
population and thereby inflated our chances of finding
a group difference on the laboratory task. Instead, by
matching the groups on PAC score, we conducted a
conservative test, as any selection bias created using
this method would be against our preferred hypothesis
(i.e., we have likely picked control participants relatively
high in the distractibility distribution for their pop-
ulation and Ile89Val participants relatively low in the
distractibility distribution for their population). Our re-
sults may thus underestimate the size of the group dif-
ference in the distraction effect. Ile89Val has been
associated with ADHD and depression, so to maintain
sample size, we matched participants as closely as pos-
sible on history of diagnosis rather than excluding on
this basis. (Analyses excluding these participants gave
similar results with slightly reduced power.)
Additional Screening Measures
Depression. Hahn et al. (2008) found that Ile89Val was
associated with depression severity (though not inci-
dence) in a clinical population. We therefore matched
participants on depression ratings using the Patient
Health Questionnaire-9 (PHQ-9; Kroenke, Spitzer, &
Williams, 2001). Participants respond to each of the nine
symptom questions using a scale of 0–3. Possible scores
range from 0 to 27, with 0 indicating no self-reported
experience of depression and 27 reflecting severe de-
pression symptoms.
Sleep and other health measures. Because the choli-
nergic system is also involved in the sleep/wake cycle
and sleep quality can affect attention, we wanted to en-
sure that our groups were also matched on this front.
The Pittsburgh Sleep Quality Index (Buysse, Reynolds,
Monk, Berman, & Kupfer, 1989) consists of 10 multi-
component questions. Scores range from 0 to 21, with 0
indicating the best sleep quality. The Berlin Sleep
Questionnaire (Netzer, Stoohs, Netzer, Clark, & Strohl,
1999) consists of 10 questions relating to snoring, sleep
problems, and blood pressure, as well as age and body
mass index. Participants are considered at high risk
for sleep apnea if they have a positive score (more than
2 points) in two categories. Participants also completed
the standard health, demographics, and screening form
used in our laboratory, which asks questions about med-
ications and medical or psychiatric conditions that could
affect performance.
1984
Journal of Cognitive Neuroscience
Volume 26, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
9
2
6
1
/
9
9
8
/
1
1
1
9
9
8
4
1
7
/
5
1
7
7
9
8
o
1
c
5
n
9
_
2
a
/
_
j
0
o
0
c
6
n
0
7
_
a
p
_
d
0
0
b
6
y
0
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 2. Demographic and PAC Questionnaire Measures for Participants Completing the CTET with Distraction Task
Control
Ile89Val
t Test
Effect Size
(Cohenʼs d)
Demographic and Health Measures
Age (years)
M
SD
Education (years)
M
SD
History of ADHD/depression/anxiety (n)
Psychotropic medications (n)
PHQ-9 depression score
M
SD
Pittsburgh Sleep Quality Index
M
SD
Self-report Everyday Attention Measures (PAC)
Distractibility
M
SD
Mind-wandering
M
SD
Boredom
M
SD
45.13
16.85
16.73
2.57
6
6
3.50
3.97
5.91
4.32
13.50
4.21
12.91
3.77
12.94
3.30
44.66
16.99
16.53
3.59
7
5
3.47
4.12
4.75
3.54
14.59
4.45
13.38
4.47
12.25
4.21
t < 1
p = .91
t < 1
p = .80
–
–
t < 1
p = .98
t = 1.17
p = .25
t = 1.01
p = .32
t < 1
p = .65
t < 1
p = .47
d = .03
d = .06
–
–
d = .01
d = .29
d = .25
d = .11
d = .18
t test and Cohenʼs d refer to the comparison between control and lle89Val participants. Each group included 19 women and 13 men, for a total n =
32 per group.
CTET with Video Distractor
CTET procedures generally followed those used by
OʼConnell et al. (2009), with some modifications (e.g.,
target and standard durations, response window) based on
pilot testing to make it feasible for participants with a wide
range of ability. The task was presented on a Dell PC using
Presentation software (Neurobehavioral Systems, Inc.,
Berkeley, CA). On each trial, participants were presented
with a black and white 10 × 10 grid of square tiles (15 mm2
each) divided diagonally into black and white halves. On
standard trials, the grid randomly changed rotation (90°,
180°, or 270°) after 800 msec; on target trials it rotated
after 1070 msec (Figure 1). Participants were instructed
to press the spacebar as soon as they detected the target.
The distraction manipulation was implemented using
a laptop oriented 32° to the left of the CTET task and 65 cm
from the participant. In the No Distractor condition, the
laptop was silent and displayed a gray screen. In the Dis-
tractor condition, the laptop played a series of 30-sec video
clips from various sources (e.g., cartoons, movies, sports)
with sound presented via headphones. None of the videos
contained music or other obviously rhythmic content or
overtly violent or sexual content.
Responses were recorded as correct (hits) up to 1.5 sec
following target offset. Responses outside this window
were coded as false alarms (FAs). Participants received
feedback at the end of each run. Before beginning the
experiment, participants were given six practice runs that
contained three targets each. For the first practice run,
the duration difference between target and standard trials
was exaggerated to ensure participants understood the
task rules (standard: 800 msec; target: 1600 msec). Par-
ticipants were informed that the timing parameters of
Berry et al.
1985
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
9
2
6
1
/
9
9
8
/
1
1
1
9
9
8
4
1
7
/
5
1
7
7
9
8
o
1
c
5
n
9
_
2
a
/
_
j
0
o
0
c
6
n
0
7
_
a
p
_
d
0
0
b
6
y
0
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 1. CTET with video
distractor. As shown in (A) each
trial consisted of a black and
white grid made up of squares
divided into triangles. At the
end of the trial, the triangles
rotated (90°, 180°, or 270°,
chosen randomly) to start
the next trial. The participantʼs
task was to press the spacebar
when they realized that the grid
had taken longer than usual
(1070 msec rather than the
standard 800 msec) to rotate.
(B) The distractor manipulation
was implemented using a laptop
oriented 32° to the left of the
main task computer. In the No
Distractor condition, the laptop
was silent and displayed a gray
screen. In the Distractor
condition, it played video clips
with sound.
the subsequent five practice runs would be more challeng-
ing and identical to the test runs (standard: 800 msec;
target: 1070 msec). To ensure participants were able to
discriminate between standard and target durations, they
were required to continue practice until they achieved
100% (3/3 targets detected) if they had not already
reached this criterion in the mandated five practice runs.
Participants then performed nine task runs (4 min
each) with 24 targets per run.1 Stimuli were pseudo-
randomly intermixed, with four to eight target trials pre-
sented per minute and 7–14 standard duration stimuli
presented between each target. Participants took a 1-min
break between each experimental run. For each par-
ticipant, four runs were presented in the No Distractor
condition, and four runs were presented in the Distrac-
tor condition. Distractor and No Distractor runs alter-
nated, and their order was counterbalanced across
participants. Participants completed one additional “Boring
Distractor” run for piloting purposes that is not included
in the present analysis. The single Boring Distractor run
was inserted as either the first, third, sixth, or ninth run.
Post Experiment Questionnaire
Distractor video content. After the CTET, participants
completed a surprise 15-item multiple-choice quiz assess-
ing memory for the video content. Similar procedures
have revealed relationships between memory for the
distracting stimuli and the impact of distraction on per-
formance, as well as experimental group differences
(Berry et al., 2014; Biss, Ngo, Hasher, Campbell, & Rowe,
2013; Clapp & Gazzaley, 2012; Gazzaley, Cooney, Rissman,
& DʼEsposito, 2005).
the level of distractibility, mind-wandering, and bore-
dom they experienced during the task. These questions
were intended to be similar in form and content to items
from the PAC trait attention measure. Participants were
given five statements and asked to rate the degree to
which they identified with each statement on a scale
from 1 to 5 (see Table 3). Questions 1, 2, and 4 measured
mind-wandering, Question 3 measured boredom, and
Question 5 measured distractibility. We emphasized
mind-wandering (three questions) when constructing
the questionnaire as that had been the focus of the orig-
inal OʼConnell et al. (2009) article. However, subsequent
analyses pooling across CTET experiments in our labora-
tory show that Question 4 consistently shows the highest
correlations with the PAC mind-wandering score and
with CTET performance. For that reason and so that an
equal number of items are used for the mind-wandering,
distractibility, and boredom scores, as in prior work (Berry
et al., 2014), we focus on Question 4 as the “state” measure
of mind-wandering.
Results and Discussion
Consistent with previous articles (Berry et al., 2014;
OʼConnell et al., 2009), analyses of CTET performance
focused on hits (correct target detections); analyses of
0 measures yield similar results. Greenhouse–Geisser
d
sphericity correction was applied as needed for reporting
p values, but degrees of freedom are reported as integers
in the text for easier reading. Effect sizes are reported
using η2
G (Bakeman, 2005), which gives smaller values
than the frequently used η2
P but is preferable as it
reduces error when comparing across studies (Fritz,
Morris, & Richler, 2012).
State attention. After completing the CTET and the
quiz for memory of the distractors, participants rated
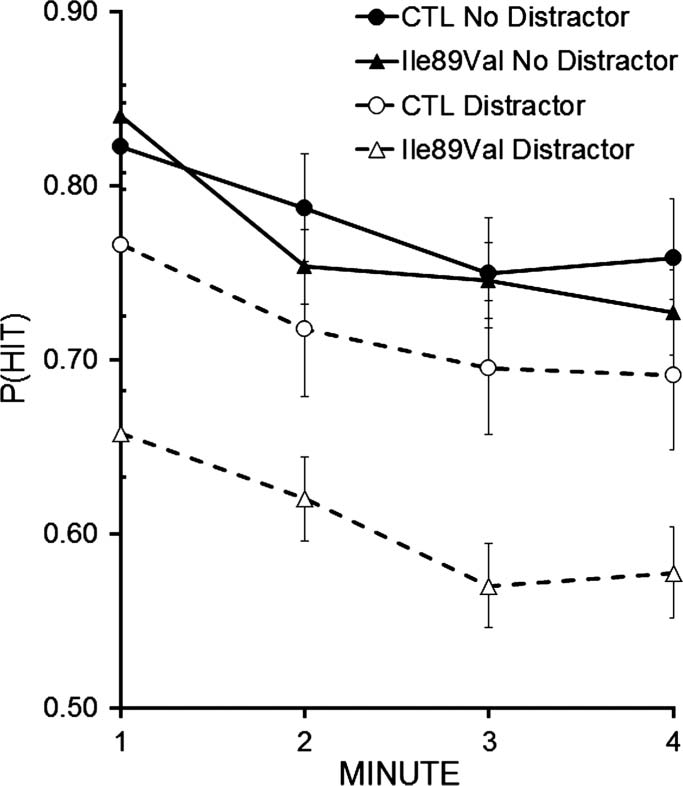
As seen in Figure 2, external distraction and time-on-
task effects were independent, and heterozygotes showed
1986
Journal of Cognitive Neuroscience
Volume 26, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
9
2
6
1
/
9
9
8
/
1
1
1
9
9
8
4
1
7
/
5
1
7
7
9
8
o
1
c
5
n
9
_
2
a
/
_
j
0
o
0
c
6
n
0
7
_
a
p
_
d
0
0
b
6
y
0
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 3. Post Experiment Questionnaire Measures for
Participants Completing the CTET with Distraction Task
1. At times of this task, it was hard for me to keep my mind from
wandering.
M
SD
2.63
1.21
3.53
1.19
t = 3.02
p = .004*
d = .75
2. (Reverse scored) During the task, my thoughts seldom
shifted from the subject in front of me.
M
SD
2.81
1.12
3.31
1.23
t = 1.7
p = .09
3. I was easily bored during this task.
M
SD
2.47
1.19
2.63
1.43
t < 1
p = .64
d = .43
d = .12
4. I had difficulty in keeping my attention focused on this long,
tedious task.
M
SD
2.59
0.98
3.09
1.28
t = 1.76
p = .08
d = .44
5. No matter how hard I tried to concentrate, I felt easily dis-
tracted by the videos playing.
M
SD
2.47
1.16
3.09
1.20
t = 2.11
p = .04*
d = .53
Recognition memory for distractor content (% correct)
M
SD
0.60
0.26
0.71
0.22
t = 0.193
p = .06
d = .46
t test and Cohenʼs d refer to the comparison between control and
lle89Val participants.
*Significant difference between groups.
a specific sensitivity to distraction. ANOVA results with
within-participant factors Distraction (No Distractor, Dis-
tractor), Time (Minutes 1, 2, 3, 4), and between-participant
factor Genotype (Control, Ile89Val) revealed main effects
of Distraction, F(1, 62) = 59.27, p < .0005, η2
G = .07,
and Time-on-task, F(3, 186) = 21.35, p < .0005, η2
G =
.03, that did not interact, F < 1 (Figure 2). Of primary
interest, heterozygotes were more sensitive than controls
to distraction, F(1, 62) = 11.65, p = .001, η2
G = .01,
but not time-on-task, F < 1.2 The three-way interaction
between Distraction, Time-on-task, and Genotype did
not approach significance, F < 1. Although groups were
matched on PAC distractibility, the CTET with distraction
was sensitive to group differences in distractor vulnerabil-
ity, suggesting that the PAC and laboratory performance
measures captured at least partially distinct aspects of
distractibility.
FA rates were low and did not differ across groups. An
ANOVA with within-participant factors Distraction (No
Distractor, Distractor) and Time (Minutes 1, 2, 3, 4) and
between-participant factor Genotype (Control, Ile89Val)
revealed only a main effect of Distraction, F(1, 62) =
5.10, p = .03, η2
G < 0.01, such that FA rates were higher
during distraction (No Distractor M = 0.94%, SD = 1.7;
Distractor M = 1.15%, SD = 2.04). There was no effect
of Time or Genotype and no interactions (all Fs < 1).
Post experiment ratings of subjective experience gener-
ally followed the patterns seen for ratings of everyday at-
tentional function (PAC scores) in Experiment 1: Ile89Val
participants tended to give higher ratings for items
concerning mind-wandering and distractibility, but not
boredom (Table 3). Although these effects are small, they
are notable given that, in selecting our participants, we
had matched Ile89Val and control participants as closely
as possibly in their PAC subjective ratings of everyday
attention.
Thus, although the Ile89Val participants tested in this
experiment were selected to give similar ratings as con-
trols for attentional function in everyday life, they still
tended to show an increased subjective as well as objec-
tive susceptibility to distraction during the experimental
task. In addition, Ile89Val participants tended to have bet-
ter memory for the video contents. Together with the
CTET performance data, these patterns suggest that
Ile89Val individuals are more susceptible to distraction
but may benefit from better memory for nominally irrel-
evant information.
To determine how task performance related to subjec-
tive measures of attention, we examined how CTET per-
formance correlated with the self-report measures of
attention in everyday life and state of attention during
the task. To reduce the number of comparisons and
ensure replication across data sets, unless otherwise
noted we restrict our analyses to those of theoretical
interest and also examined in a previous experiment
using an undergraduate sample (Berry et al., 2014).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
9
2
6
1
/
9
9
8
/
1
1
1
9
9
8
4
1
7
/
5
1
7
7
9
8
o
1
c
5
n
9
_
2
a
/
_
j
0
o
0
c
6
n
0
7
_
a
p
_
d
0
0
b
6
y
0
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. CTET performance with and without video distractor.
Markers represent mean proportion of hits for each minute; error bars
represent SEM. Both groups exhibit time-on-task and distraction effects
that do not interact, and the distractor effect is larger for IIeVal89
participants.
Berry et al.
1987
In the text, we note the consistencies and discrepan-
cies compared with Berry et al. so that the reader can
weigh results accordingly. Full correlation tables for the
combined group and each subgroup are available upon
request.
We first asked how performance correlated with the
PAC “trait” measures of attention, with particular interest
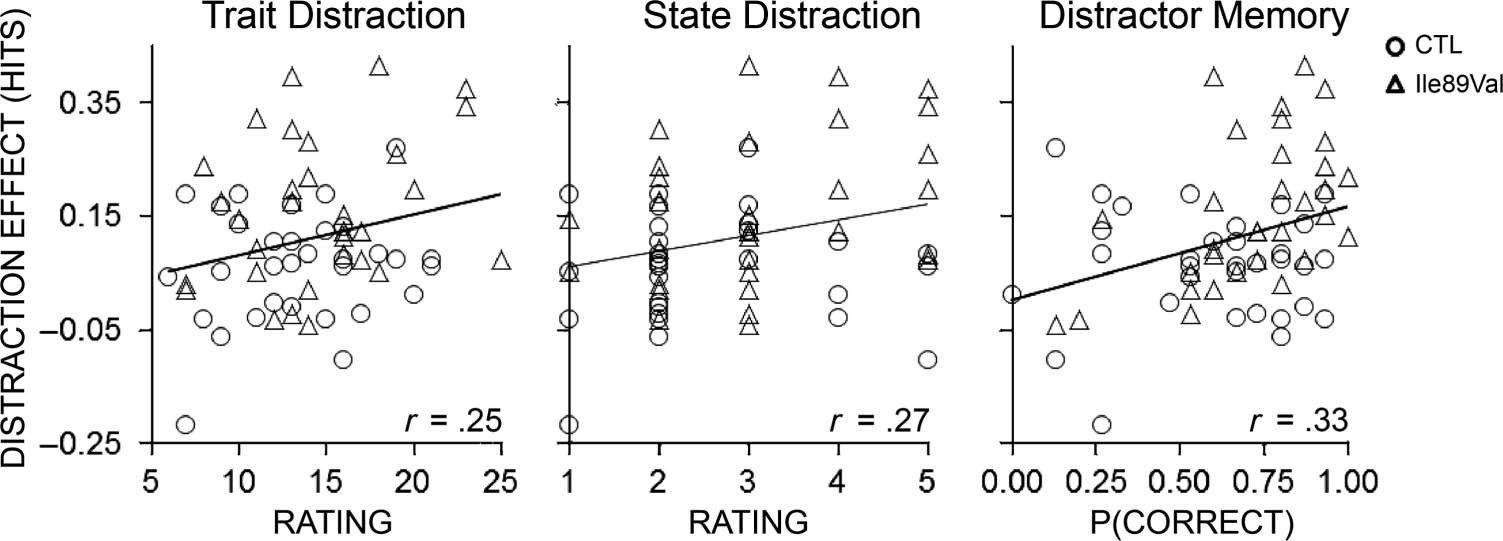
in those measures related to distraction (Figure 3). Across
both groups, there was a small-to-moderate correlation
such that participants who reported more distractibility in
everyday life also showed a greater distractor-related drop
in performance on the CTET, r = .25, p = .05. In a pre-
vious experiment using an undergraduate sample (Berry
et al., 2014), this correlation was in the same direction
but did not reach significance. The greater variance in
the distraction effect in this study (due to Ile89Val par-
ticipantsʼ distractor vulnerability) may have contributed
to this difference. In our undergraduate sample, self-
reported mind-wandering negatively correlated with over-
all performance in the No Distractor condition, but this
relationship did not reach significance in the current sam-
ple (r = −.19, p = .13). Thus, across studies, correlations
between performance and self-reported everyday atten-
tional function are of modest size but in the expected
direction.
Likewise consistent with the Berry et al. (2014) under-
graduate sample, in this community sample stronger cor-
relations were found between task performance and post
experiment ratings (Table 3; Figure 3). Target detection
in the No Distractor condition was negatively related to
difficulty keeping attention focused (r = −.40, p = .001).
Greater performance declines as a result of the video
distractor correlated with greater self-rated distraction by
the videos, r = .27, p = .03, and to better memory for
the videosʼ content, r = .33, p = .008.
A reviewer noted that the mean group differences in
the size of the distractor effect on performance might
have inflated the size of the cross-group correlations with
this effect. To examine this possibility, we calculated the
correlations with this factor for each group. For the cor-
relation with trait distractibility as measured by the PAC
subscale, the difference between the groups was rela-
tively small (r = .16 for controls, r = .27 for Ile89Val par-
ticipants), perhaps because we had matched the groups
on PAC scores. However, for both self-rated distraction
(r = −.01 for controls, r = .34 for Ile89Val participants)
and memory for the videosʼ content (r = −.004 for con-
trols, r = .53 for Ile89Val participants), correlations with
the distractor effect on performance were largely driven
by Ile89Val participants. In other words, in addition to
being more vulnerable to distractionʼs detrimental effects
on performance, Ile89Val participants were also more sen-
sitive to its subjective effects and its potential benefits to
later memory.
GENERAL DISCUSSION
The present results reveal three major findings. First,
although both distraction and time-on-task reduce per-
formance, their effects are independent. This seems
inconsistent with simple “underload” or “overload” re-
source explanations of vigilance decrements. Instead,
these different types of demand appear to tap disso-
ciable attention control processes. Although neuro-
imaging studies suggest a core network involved in
stable task control (e.g., Dosenbach et al., 2006), recent
studies focused on time-on-task or fatigue effects show
substantial variance in the neural correlates of such effects
indicating fatigue of specific task-dependent processes
(e.g., Persson, Larsson, & Reuter-Lorenz, 2013; Lim
et al., 2010). The task used here may provide a useful
tool for examining these processes in clinical and indivi-
dual differences research.
Second, in a community-based sample, genetic varia-
tion in cholinergic function was related to self-reported
everyday (trait) distractibility and mind-wandering, but
not boredom. There are legitimate concerns about the
degree to which a single nucleotide polymorphism (SNP)
can be linked to cognitive processes, as genes are not
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
9
2
6
1
/
9
9
8
/
1
1
1
9
9
8
4
1
7
/
5
1
7
7
9
8
o
1
c
5
n
9
_
2
a
/
_
j
0
o
0
c
6
n
0
7
_
a
p
_
d
0
0
b
6
y
0
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
Figure 3. Correlations
between distractor-related
performance declines in the
CTET, self-rated “trait”
distractibility measured using
the PAC, self-rated “state”
distractibility measured
using the post experiment
questionnaire and performance
on the post experiment
quiz for memory of the
content of the distracting
videos. Larger numbers
indicate greater distractibility.
If the data from the control
participant in the lower left corner are excluded, the correlations remain in the same direction as indicated in the main text at reduced effect size
(r = .20 for trait distraction, r = .22 for state distraction, and r = .29 for distractor memory).
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
1988
Journal of Cognitive Neuroscience
Volume 26, Number 9
solitary mediators of behavior. Instead, gene–behavior
relationships likely take the form of complex interactions
that include other genes and the environment (Thomas,
2010). However, the pattern found here, especially in-
creased distractibility, was predicted a priori from prior
research and can be interpreted in the context of evi-
dence about its neurobiological impact. When first describ-
ing the Ile89Val SNP, Okuda et al. (2002) noted that the
effects of reduced choline transport might be especially
evident under demanding conditions, consistent with
rodent studies that show that responding to the demands
imposed by distraction depends critically on the right
basal forebrain cholinergic system (St. Peters et al., 2011;
Gill et al., 2000). These findings, as well as parallel human
neuroimaging studies showing increased right prefrontal
activation in the same conditions that produce increased
right prefrontal ACh in rodents, converge to predict the
present association between Ile89Val and distractibility.
Third, even after matching participants on self-report
measures of distraction in everyday life, Ile89Val partici-
pants showed a specific vulnerability to distraction on a
laboratory task (the CTET). A distraction score of 12%
discriminated the groups with 63% sensitivity and 72%
specificity. Conceptually replicating the group differences
on the trait measures, when asked about their attentional
state during the task, Ile89Val participants gave higher
ratings on items concerning distractibility and mind-
wandering, but not boredom. The equivalent state and
trait boredom scores, No Distractor task performance,
and time-on-task declines, as well as superior perfor-
mance on the memory test, indicate that Ile89Val par-
ticipants are not generally impaired but have a particular
vulnerability to distraction.
To summarize, the association between Ile89Val and
distractibility is predicted from molecular, systems, and
cognitive neuroscience and replicates across self-report
measures of everyday attention, laboratory task perfor-
mance, and self-report ratings of attention during the
task. In contrast, the link to mind-wandering is more ten-
uous. It is not clear how mind-wandering relates to the
mechanism (attenuated cholinergic response to high task
demands) indicated by molecular and cellular studies,
and although Ile89Val participants consistently indicated
higher rates of mind-wandering on the self-report mea-
sures, they did not show worse task performance overall
or steeper time-on-task declines. A potential reason for
the discrepancy between the self-report and performance
measures is that, although the former ask about off-track
thoughts, those thoughts may not always disrupt per-
formance. Esterman, Noonan, Rosenberg, and DeGutis
(2013) recently reported different neural correlates and
timescales for vigilance declines, off-track thought, and
performance lapses. Of particular interest, default net-
work activity putatively related to off-track thought had
a nonlinear relationship to performance lapses depend-
ing on the stability of goal-directed attention. Future
investigations using experience-sampling techniques and
neural measures with Ile89Val participants may resolve
this issue.
The more robust link between Ile89Val and distract-
ibility found here is to our knowledge the first evidence
linking this polymorphism to a specific cognitive process
and the first indicating its role in normal cognitive varia-
tion beyond clinical conditions such as ADHD. Besides
their vulnerability to the distractorʼs detrimental effects,
Ile89Val participantsʼ better memory for the distractor is
reminiscent of findings from older adults showing that
increased processing of nominally irrelevant information
provides an advantage if that information becomes use-
ful later on (Biss et al., 2013; Clapp & Gazzaley, 2012;
Gazzaley et al., 2005). Increased processing of irrelevant
information has been linked to creativity in ADHD (White
& Shah, 2011); an interesting question for future studies
is whether this advantage extends to Ile89Val.
As noted above, understanding the role that any SNP,
including Ile89Val, plays in normal cognition and cogni-
tive disorders requires consideration of its interaction
with other genetic and environmental factors. For exam-
ple, variation at a nearby SNP (G to T substitution at
CHT 30 UTR) has been linked to corticolimbic reactivity
(Neumann et al., 2006), and this variation paired with
Ile89Val was especially common in the combined subtype
of ADHD (English et al., 2009). Recent evidence from a
mouse model suggests that reduced CHT function in
combination with dopamine depletion may explain why
some Parkinsonʼs patients show pronounced cognitive
declines (Zurkovsky et al., 2013) and links between the
amyloid precursor protein and CHT indicate a potential
role in Alzheimerʼs disease (Wang, Yang, Wang, & Zheng,
2007).
The present results suggest that the cholinergic system
plays an important and specific role in susceptibility to
distraction in everyday life (Experiment 1) and on a
laboratory task (Experiment 2). They add to a growing
body of research that integrates findings from several
areas of psychology and neuroscience to clarify links be-
tween cognitive functions and the neural systems that
underlie them (see also Howe et al., 2013). This integ-
rative approach holds promise for improving our under-
standing of how genes influence cognition and behavior
and for developing more effective, precisely targeted
treatments when these processes are disordered (Insel
et al., 2010).
Acknowledgments
We thank Mina Raj for assistance in creating the stimuli for
Experiment 2, Patricia J. Deldin for advising on depression
and sleep screening measures and serving as a clinical contact
for the depression screen, and John Jonides for comments on
the manuscript. We are also grateful to the participants, espe-
cially those who returned for Experiment 2, and the many
research assistants who facilitated data collection. The study was
funded by NIH Grant 1R01MH086530. A. B. and E. D. were both
funded by National Science Foundation Graduate Fellowships.
Berry et al.
1989
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
9
2
6
1
/
9
9
8
/
1
1
1
9
9
8
4
1
7
/
5
1
7
7
9
8
o
1
c
5
n
9
_
2
a
/
_
j
0
o
0
c
6
n
0
7
_
a
p
_
d
0
0
b
6
y
0
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Reprint requests should be sent to Cindy Lustig, Psychology
Department, Neuroscience Program, University of Michigan,
4016 East Hall, 530 Church Street, Ann Arbor, MI 49109-1043,
or via e-mail: clustig@umich.edu.
Notes
1.
In this and other studies, time-on-task effects occur within
runs; there are no systematic differences between runs (i.e.,
performance is not worse in Run 5 than in Run 1; see also Berry
et al., 2014; OʼConnell et al., 2009).
2. As a check of reliability, we split each group in half (even/
odd pairs) and reran the analysis on the subsamples; effects
were in the same direction and of similar effect size as those
reported here.
REFERENCES
Bakeman, R. (2005). Recommended effect size statistics for
repeated measures designs. Behavior Research Methods, 37,
379–384.
Berry, A. S., Li, X., Lin, Z., & Lustig, C. (2014). Shared and
distinct factors driving attention and temporal processing
across modalities. Acta Psychologica, 147, 42–50.
Biss, R. K., Ngo, J. K. W., Hasher, L., Campbell, K. L., & Rowe, G.
(2013). Distraction can reduce age-related forgetting.
Psychological Science, 24, 448–455.
Breckel, T. P. K., Giessing, C., & Thiel, C. M. (2011). Impact of
brain networks involved in vigilance on processing irrelevant
visual motion. Neuroimage, 55, 1754–1762.
Broussard, J. I., Karelina, K., Sarter, M., & Givens, B. (2009).
Cholinergic optimization of cue-evoked parietal activity
during challenged attentional performance. European
Journal of Neuroscience, 29, 1711–1722.
Buysse, D. J., Reynolds, C. F., III, Monk, T. H., Berman, S. R., &
Kupfer, D. J. (1989). The Pittsburgh Sleep Quality Index:
A new instrument for psychiatric practice and research.
Psychiatry Research, 28, 193–213.
Clapp, W. C., & Gazzaley, A. (2012). Distinct mechanisms for
the impact of distraction and interruption on working
memory in aging. Neurobiology of Aging, 33, 134–148.
Demeter, E., Guthrie, S. K., Taylor, S. F., Sarter, M., & Lustig, C.
(2013). Increased distractor vulnerability but preserved
vigilance in patients with schizophrenia: Evidence from a
translational sustained attention task. Schizophrenia
Research, 144, 136–141.
Demeter, E., Hernandez-Garcia, L., Sarter, M., & Lustig, C.
(2011). Challenges to attention: A continuous arterial spin
labeling (ASL) study of the effects of distraction on sustained
attention. Neuroimage, 54, 1518–1529.
Demeter, E., & Sarter, M. (2013). Leveraging the cortical
cholinergic system to enhance attention.
Neuropharmacology, 64, 294–304.
Dosenbach, N. U. F., Visscher, K. M., Palmer, E. D., Miezin, F. M.,
Wenger, K. K., Kang, H. S. C., et al. (2006). A core system for
the implementation of task sets. Neuron, 50, 799–812.
Edenberg, H. J., & Liu, Y. (2009). Laboratory methods for
high-throughput genoyping. Cold Spring Harbor Protocols,
2009, pdb top62.
English, B. A., Hahn, M. K., Gizer, I. R., Mazei-Robison, M.,
Steele, A., Kurnik, D. M., et al. (2009). Choline transporter
gene variation is associated with attention-deficit hyperactivity
disorder. Journal of Neurodevelopmental Disorders, 1,
252–263.
Esterman, M., Noonan, S. K., Rosenberg, M., & DeGutis, J.
(2013). In the zone or zoning out? Tracking behavioral
and neural fluctuations during sustained attention.
Cerebral Cortex, 23, 2712–2723.
Fritz, C. O., Morris, P. E., & Richler, J. J. (2012). Effect size
estimates: Current use, calculations, and interpretation.
Journal of Experimental Psychology: General, 141, 2–18.
Gazzaley, A., Cooney, J. W., Rissman, J., & DʼEsposito, M.
(2005). Top-down suppression deficit underlies working
memory impairment in normal aging. Nature Neuroscience,
8, 1298–1300.
Gill, T. M., Sarter, M., & Givens, B. (2000). Sustained visual
attention performance-associated prefrontal neuronal
activity: Evidence for cholinergic modulation. Journal of
Neuroscience, 20, 4745–4757.
Hahn, M. K., Blackford, J. U., Haman, K., Mazei-Robison, M.,
English, B. A., Prasad, H. C., et al. (2008). Multivariate
permutation analysis associates multiple polymorphisms
with subphenotypes of major depression. Genes Brain and
Behavior, 7, 487–495.
Hasselmo, M. E., & Sarter, M. (2011). Modes and models of
forebrain cholinergic neuromodulation of cognition.
Neuropsychopharmacology: Official Publication of the
American College of Neuropsychopharmacology, 36, 52–73.
Howe, W. M., Berry, A. S., Francois, J., Gilmour, G., Carp, J. M.,
Tricklebank, M., et al. (2013). Prefrontal cholinergic
mechanisms instigating shifts from monitoring for cues to
cue-guided performance: Converging electrochemical and
fMRI evidence from rats and humans. Journal of Neuroscience,
33, 8742–8752.
Huba, G. J., Singer, J. L., Aneshensel, C. S., & Antrobus, J. S.
(1982). Short Imaginal Processes Inventory. Port Huron, MI:
Research Psychologists Press.
Insel, T., Cuthbert, B., Garvey, M., Heinssen, R., Pine, D. S.,
Quinn, K., et al. (2010). Research domain criteria (RDoC):
Toward a new classification framework for research on
mental disorders. American Journal of Psychiatry, 167,
748–751.
Kroenke, K., Spitzer, R. L., & Williams, J. B. (2001). The PHQ-9:
Validity of a brief depression severity measure. Journal of
General Internal Medicine, 16, 606–613.
Lavie, N. (2010). Attention, distraction, and cognitive control
under load. Current Directions in Psychological Science,
19, 143–148.
Lavie, N., Hirst, A., de Fockert, J. W., & Viding, E. (2004).
Load theory of selective attention and cognitive control.
Journal of Experimental Psychology: General, 133, 339–354.
Lim, J., Ebstein, R., Tse, C. Y., Monakhov, M., Lai, P. S., Dinges,
D. F., et al. (2012). Dopaminergic polymorphisms associated
with time-on-task declines and fatigue in the psychomotor
vigilance test. PloS One, 7, e33767.
Lim, J., Wu, W. C., Wang, J. J., Detre, J. A., Dinges, D. F., & Rao,
H. Y. (2010). Imaging brain fatigue from sustained mental
workload: An ASL perfusion study of the time-on-task effect.
Neuroimage, 49, 3426–3435.
Lustig, C., Kozak, R., Sarter, M., Young, J. W., & Robbins, T. W.
(2012). CNTRICS final animal model task selection: Control
of attention. Neuroscience and Biobehavioral Reviews, 37,
2099–2110.
Lustig, C., & Meck, W. H. (2005). Chronic treatment with
haloperidol induces deficits in working memory and feedback
effects of interval timing. Brain and Cognition, 58, 9–16.
Mazei-Robinson, M. S., Couch, R. S., Shelton, R. C., Stein, M. A.,
& Blakely, R. D. (2005). Sequence variation in the human
dopamine transporter gene in children with attention deficit
hyperactivity disorder. Neuropharmacology, 49, 724–736.
Netzer, N. C., Stoohs, R. A., Netzer, C. M., Clark, K., & Strohl,
K. P. (1999). Using the Berlin Questionnaire to identify
patients at risk for the sleep apnea syndrome. Annals of
Internal Medicine, 131, 485–491.
1990
Journal of Cognitive Neuroscience
Volume 26, Number 9
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
9
2
6
1
/
9
9
8
/
1
1
1
9
9
8
4
1
7
/
5
1
7
7
9
8
o
1
c
5
n
9
_
2
a
/
_
j
0
o
0
c
6
n
0
7
_
a
p
_
d
0
0
b
6
y
0
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Neumann, S. A., Brown, S. M., Ferrell, R. E., Flory, J. D., Manuck,
S. B., & Hariri, A. R. (2006). Human choline transporter gene
variation is associated with corticolimbic reactivity and
autonomic-cholinergic function. Biological Psychiatry, 60,
1155–1162.
OʼConnell, R. G., Dockree, P. M., Robertson, I. H., Bellgrove,
M. A., Foxe, J. J., & Kelly, S. P. (2009). Uncovering the neural
signature of lapsing attention: Electrophysiological signals
predict errors up to 20 s before they occur. Journal of
Neuroscience, 29, 8604–8611.
Okuda, T., Okamura, M., Kaitsuka, C., Haga, T., & Gurwitz, D.
(2002). Single nucleotide polymorphism of the human high
affinity choline transporter alters transport rate. Journal of
Biological Chemistry, 277, 45315–45322.
Pa, J., Berry, A. S., Compagnone, M., Boccanfuso, J., Greenhouse,
I., Rubens, M. T., et al. (2013). Cholinergic enhancement of
functional networks in older adults with mild cognitive
impairment. Annals of Neurology, 73, 762–773.
Parikh, V., St. Peters, M., Blakely, R. D., & Sarter, M. (2013).
The presynaptic choline transporter imposes limits on
sustained cortical acetylcholine release and attention.
Journal of Neuroscience, 33, 2326–2337.
Pattyn, N., Neyt, X., Heridericlcx, D., & Soetens, E. (2008).
Psychophysiological investigation of vigilance decrement:
Boredom or cognitive fatigue? Physiology & Behavior, 93,
369–378.
Penney, T. B., Gibbon, J., & Meck, W. H. (2000). Differential
effects of auditory and visual signals on clock speed and
temporal memory. Journal of Experimental Psychology:
Human Perception and Performance, 26, 1770–1787.
Persson, J., Larsson, A., & Reuter-Lorenz, P. A. (2013). Imaging
fatigue of interference control reveals the neural basis of
executive resource depletion. Journal of Cognitive
Neuroscience, 25, 338–351.
Picciotto, M. R., Higley, M. J., & Mineur, Y. S. (2012).
Acetylcholine as a neuromodulator: Cholinergic signaling
shapes nervous system function and behavior. Neuron, 76,
116–129.
Rokem, A., Landau, A. N., Garg, D., Prinzmetal, W., & Silver,
M. A. (2010). Cholinergic enhancement increases the effects
of voluntary attention but does not affect involuntary attention.
Neuropsychopharmacology: Official Publication of the
American College of Neuropsychopharmacology, 35,
2538–2544.
Rosenberg, M., Noonan, S., DeGutis, J., & Esterman, M. (2013).
Sustaining visual attention in the face of distraction: A novel
gradual-onset continuous performance task. Attention,
Perception & Psychophysics, 75, 426–439.
Sarter, M., Martinez, V., & Kozak, R. (2009). A neurocognitive
animal model dissociating between acute illness and
remission periods of schizophrenia. Psychopharmacology,
202, 237–258.
Silver, M. A., Shenhav, A., & DʼEsposito, M. (2008). Cholinergic
enhancement reduces spatial spread of visual responses in
human early visual cortex. Neuron, 60, 904–914.
Singer, J. L., & Antrobus, J. S. (1970). Manual for the Imaginal
Processes Inventory. Princeton, NJ: Educational Testing
Service.
St. Peters, M., Demeter, E., Lustig, C., Bruno, J. P., & Sarter, M.
(2011). Enhanced control of attention by stimulating
mesolimbic-corticopetal cholinergic circuitry. Journal of
Neuroscience, 31, 9760–9771.
Tanaka, J. S., & Huba, G. J. (1985/1986). Longitudinal stability of
three second-order daydreaming factors. Imagination,
Cognition and Personality, 5, 181–206.
Thomas, D. (2010). Gene-environment-wide association studies:
Emerging approaches. Nature Reviews Genetics, 11, 259–272.
Wang, B., Yang, L., Wang, Z., & Zheng, H. (2007). Amyolid
precursor protein mediates presynaptic localization and
activity of the high-affinity choline transporter. Proceedings
of the National Academy of Sciences, U.S.A., 104,
14140–14145.
White, H. A., & Shah, P. (2011). Creative style and achievement
in adults with attention-deficit/hyperactivity disorder.
Personality and Individual Differences, 50, 673–677.
Zakay, D., & Block, R. A. (1997). Temporal cognition. Current
Directions in Psychological Science, 6, 12–16.
Zurkovsky, L., Bychkov, E., Tsakem, E. L., Siedlecki, C., Blakely,
R. D., & Gurevich, E. V. (2013). Cognitive effects of dopamine
depletion in the context of diminished acetylcholine
signaling capacity in mice. Disease Models & Mechanisms, 6,
171–183.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
9
2
6
1
/
9
9
8
/
1
1
1
9
9
8
4
1
7
/
5
1
7
7
9
8
o
1
c
5
n
9
_
2
a
/
_
j
0
o
0
c
6
n
0
7
_
a
p
_
d
0
0
b
6
y
0
g
7
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Berry et al.
1991