Communicated by John Wyatt
A Silicon Model Of Auditory Localization
John Lazzaro
Carver A. Mead
Department of Computer Science, California Institute of Technology,
MS 256-80, Pasadena, CA 91125, 美国
The barn owl accurately localizes sounds in the azimuthal plane, 我们-
ing interaural time difference as a cue. The time-coding pathway in
the owl’s brainstem encodes a neural map of azimuth, by processing
interaural timing information. We have built a silicon model of the
time-coding pathway of the owl. The integrated circuit models the
structure as well as the function of the pathway; most subcircuits in
the chip have an anatomical correlate. The chip computes all outputs
in real time, using analog, continuous-time processing.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
1 介绍
The principles of organization of neural systems arose from the combi-
nation of the performance requirements for survival and the physics of
neural elements. From this perspective, the extraction of time-domain
information from auditory data is a challenging computation; 系统
must detect changes in the data which occur in tens of microseconds,
using neurons which can fire only once per several milliseconds. 新-
ral approaches to this problem succeed by closely coupling algorithms
and implementation. We are building silicon models of the auditory lo-
calization system of the barn owl, to explore the general computational
principles of time-domain processing in neural systems.
The barn owl (Tyto alba) uses hearing to locate and catch small rodents
in total darkness. The owl localizes the rustles of the prey to within one
to two degrees in azimuth and elevation (Knudsen et al. 1979). The owl
uses different binaural cues to determine azimuth and elevation. 这
elevational cue for the owl is interaural intensity difference (IID). 这
cue is a result of a vertical asymmetry in the placement of the owl’s ear
openings, as well as a slight asymmetry in the left and right halves of
the owl’s facial ruff (Knudsen and Konishi 1979). The azimuthal cue is
interaural time difference (ITD). The ITDs are in the microsecond range,
and vary as a function of azimuthal angle of the sound source (Moiseff
and Konishi 1981). The external nucleus of the owl’s inferior colliculus
(ICx) contains the neural substrate of sound localization, a map of au-
ditory space (Knudsen and Konishi 1978). Neurons in the ICx respond
Neurul Computation 1,47-57 (1989) @ 1989 Massachusetts Institute of Technology
/
e
d
你
n
e
C
哦
A
r
t
我
C
e
–
p
d
/
我
F
/
/
/
/
/
1
1
4
7
8
1
1
8
2
3
n
e
C
哦
1
9
8
9
1
1
4
7
p
d
.
.
.
.
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
48
John Lazzaro and Carver A. Mead
maximally to stimuli located in a small area in space, 对应于
a specific combination of IID and ITD.
There are several stages of neural processing between the cochlea
and the computed map of space in the ICx. Each primary auditory fiber
initially divides into two distinct pathways. One pathway processes in-
tensity information, encoding elevation cues, whereas the other pathway
processes timing information, encoding azimuthal cues. The time-coding
and intensity-coding pathways recombine in the ICx, producing a com-
plete map of space (Takahashi and Konishi 1988).
2 A Silicon Model of the Time-Coding Pathway
We have built an integrated circuit that models the time-coding path-
way of the barn owl, using analog, continuous-time processing. 数字
1 shows the floorplan of the chip. The chip receives two inputs, 科尔-
responding to the sound pressure at each ear of the owl. Each input
connects to a silicon model of the cochlea, the organ that converts the
sound energy present at the eardrum into the first neural representation
of the auditory system. In the cochlea, sound is coupled into a travel-
ing wave structure, the basilar membrane, which converts time-domain
information into spatially-encoded information, by spreading out signals
in space according to their time scale (or frequency). The cochlea circuit
is a one-dimensional physical model of this traveling wave structure; 在
engineering terms, the model is a cascade of second-order sections, 和
exponentially scaled time constants (Lyon and Mead 1988).
In the owl, inner hair cells contact the basilar membrane at discrete
intervals, converting basilar-membrane movement into a graded, half-
wave rectified electrical signal. Spiral ganglion neurons connect to each
inner hair cell, producing action potentials in response to inner-hair-cell
electrical activity. The temporal pattern of action potentials encodes the
shape of the sound waveform at each basilar-membrane position. Spi-
ral ganglion neurons also reflect the properties of the cochlea; a spiral
ganglion neuron is most sensitive to tones of a specific frequency, 这
neuron’s characteristic frequency.
In our chip, inner hair cell circuits connect to taps at discrete inter-
vals along the basilar-membrane model. These circuits compute signal
processing operations (half-wave rectification and nonlinear amplitude
compression) that occur during inner hair cell transduction. Each in-
ner hair cell circuit connects to a spiral ganglion neuron circuit. 这
integrate-to-threshold neuron circuit converts the analog output of the
inner-hair-cell model into fixed-width, fixed-height pulses. Timing infor-
mation is preserved by greatly increasing the probability of firing near
the zero crossings of the derivative of the neuron’s input.
In the owl, the spiral ganglion neurons project to the nucleus magno-
cellularis (NM), the first nucleus of the time-coding pathway. The NM
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
n
e
C
哦
A
r
t
我
C
e
–
p
d
/
我
F
/
/
/
/
/
1
1
4
7
8
1
1
8
2
3
n
e
C
哦
1
9
8
9
1
1
4
7
p
d
.
.
.
.
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
A Silicon Model Of Auditory Localization
49
I1
Left Ear Input
Nonllnur Inhlbitlon Clrcult
(170 inpurr)
Right Ear Input
Time-Multiplexing Scanne.
Output Map
的
Interaural Time Delay
数字 1: Floorplan of the silicon model of the time-coding pathway of the
owl. Sounds for the left ear and right ear enter the respective silicon cochleas
at the lower left and lower right of the figure. Inner hair cell circuits tap each
silicon cochlea at 62 equally-spaced locations; each inner hair cell circuit con-
nects directly to a spiral ganglion neuron circuit. The square box marked with
a pulse represents both the inner hair cell circuit and spiral ganglion neuron
circuit. Each spiral ganglion neuron circuit generates action potentials; 这些
signals travel down silicon axons, which propagate from left to right for spiral
ganglion neuron circuits from the left cochlea, and from right to left for spiral
ganglion circuits from the right cochlea. The rows of small rectangular boxes,
marked with the symbol At, represent the silicon axons. 170 NL neuron circuits,
represented by small circles, lie between each pair of antiparallel silicon axons.
Each NL neuron circuit connects directly to both axons, and responds maxi-
mally when action potentials present in both axons reach that particular neuron
同时. 这样, ITDs map into a neural place code. Each vertical
wire which spans the array combines the response of all NL neuron circuits
which correspond to a specific ITD. 这些 170 vertical wires form a temporally
smoothed map of ITD, which responds to a wide range of input sound frequen-
化学系. The nonlinear inhibition circuit near the bottom of the figure increases the
selectivity of this map. The time-multiplexing scanner transforms this map into
a signal suitable for display on an oscilloscope.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
n
e
C
哦
A
r
t
我
C
e
–
p
d
/
我
F
/
/
/
/
/
1
1
4
7
8
1
1
8
2
3
n
e
C
哦
1
9
8
9
1
1
4
7
p
d
.
.
.
.
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
50
John Lazzaro and Carver A. Mead
acts as a specialized relay station; neurons in the NM preserve timing in-
形成, and project bilaterally to the nucleus laminaris (NL), the first
nucleus in the time-coding pathway that receives inputs from both ears.
For simplicity, our chip does not model the NM; each spiral ganglion
neuron circuit directly connects to a silicon NL.
Neurons in the NL are most sensitive to binaural sounds with a spe-
cific ITD. 在 1948, Jeffress proposed a model to explain the encoding of
ITD in neural circuits (Jeffress 1948). In the Jeffress model applied to
the owl, axons from the ipsilateral and contralateral NM, with similar
characteristic frequencies, enter the NL from opposite surfaces. The ax-
ons travel antiparallel, and action potentials counterpropagate across the
NL; the axons act as neural delay lines. NL neurons are adjacent to both
axons. Each NL neuron receives synaptic connections from both axons,
and fires maximally when action potentials present in both axons reach
that particular neuron at the same time. In this way, ITD is mapped
into a neural place coding; the ITD that maximally excites an NL neuron
depends on the position of the neuron in the NL. Anatomical and physi-
ological evidence in the barn owl supports this theory (Carr and Konishi
1988).
The chip models the anatomy of the NL directly (如图. 我). Two sil-
icon cochleas lie at opposite ends of the chip; spiral ganglion neuron
circuits from each cochlea, with similar characteristic frequencies, 项目
to separate axon circuits, which travel antiparallel across the chip. 这
axon circuit is a discrete neural delay line; for each action potential at the
axon’s input, a fixed-width, fixed-height pulse travels through the axon,
section by section, at a controllable velocity (Mead 1989). NL neuron cir-
cuits lie between each pair of antiparallel axons at every discrete section,
and connect directly to both axons. Simultaneous action potentials at
both inputs excite the NL neuron circuit; if only one input is active, 这
neuron generates no output. For each pair of antiparallel axons, 有
a row of 170 NL neuron circuits across the chip. These neurons form a
place encoding of ITD.
Our silicon NL differs from the owl’s NL in several ways. The silicon
NL neurons are perfect coincidence detectors; in the owl, NL neurons
also respond, with reduced intensity, to monaural input. In the owl,
many axons from each side converge on an NL neuron; in the chip, 仅有的
two silicon axons converge on each silicon NL neuron. 最后, 大脑-
stem of the owl contains two NLs, symmetric about the midline; each
NL primarily encodes one half of the azimuthal plane. For simplicity,
our integrated circuit has only one copy of the NL, which encodes all
azimuthal angles.
In the owl, the NL projects to a subdivision of the central nucleus
of the inferior colliculus (ICc), which in turn projects to the ICx. 这
ICx integrates information from the time-coding pathway and from the
amplitude-coding pathway to produce a complete map of auditory space.
The final output of our integrated circuit models the responses of ICx
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
n
e
C
哦
A
r
t
我
C
e
–
p
d
/
我
F
/
/
/
/
/
1
1
4
7
8
1
1
8
2
3
n
e
C
哦
1
9
8
9
1
1
4
7
p
d
.
.
.
.
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
A Silicon Model Of Auditory Localization
51
neurons to ITDs. In response to ITDs, ICx neurons act differently from
NL neurons. Experiments suggest mechanisms for these differences; 我们的
integrated circuit implements several of these mechanisms to produce a
neural map of ITD.
Neurons in the NL and ICc respond to all ITDs that result in the
same interaural phase difference (IPD) of the neuron’s characteristic fre-
quency; neurons in the ICx respond to only the one true ITD. This behav-
ior suggests that ICx neurons combine information from many frequency
channels in the ICc, to disambiguate ITDs from IPDs; 的确, 神经元
in the NL and ICc reflect the frequency characteristics of spiral ganglion
神经元, whereas ICx neurons respond equally to a wide range of fre-
quencies.
In our chip, all NL neuron outputs corresponding to a particular ITD
are summed to produce a single output value. NL neuron outputs are
current pulses; a single wire acts as a dendritic tree to perform the sum-
运动. 这样, a two-dimensional matrix of NL neurons reduces to
a single vector; this vector is a map of ITD, for all frequencies. In the owl,
inhibitory circuits between neurons tuned to the same ITD may also be
展示, before summation across frequency channels. Our model does
not include these circuits.
Neurons in the ICc are more selective to ITDs than are neurons in the
NL; 反过来, ICx neurons are more selective to ITDs than are ICc neurons,
for low frequency sounds. At least two separate mechanisms join to
increase selectivity. The selectivity of ICc and ICx neurons increases
with the duration of a sound, for sounds lasting less than 5 毫秒,
implying that the ICc and perhaps the ICx may use temporal integration
to increase selectivity (Wagner and Konishi, in preparation). Our chip
temporally integrates the vector that represents ITD; the time constant of
integration is adjustable.
Nonlinear inhibitory connections between neurons tuned to different
ITDs in the ICc and ICx also increase sensitivity to ITDs; 应用
of an inhibitory blocker to either the ICc or ICx decreases sensitivity to
ITD (Fujita and Konishi, in preparation). In our chip, a global shunt-
ing inhibition circuit (Lazzaro et al. 1988) processes the temporally in-
tegrated vector that represents ITD. This nonlinear circuit performs a
winner-take-all function, producing a more selective map of ITD. 这
chip time-multiplexes this output map on a single wire for display on an
oscilloscope.
3 Comparison of Responses
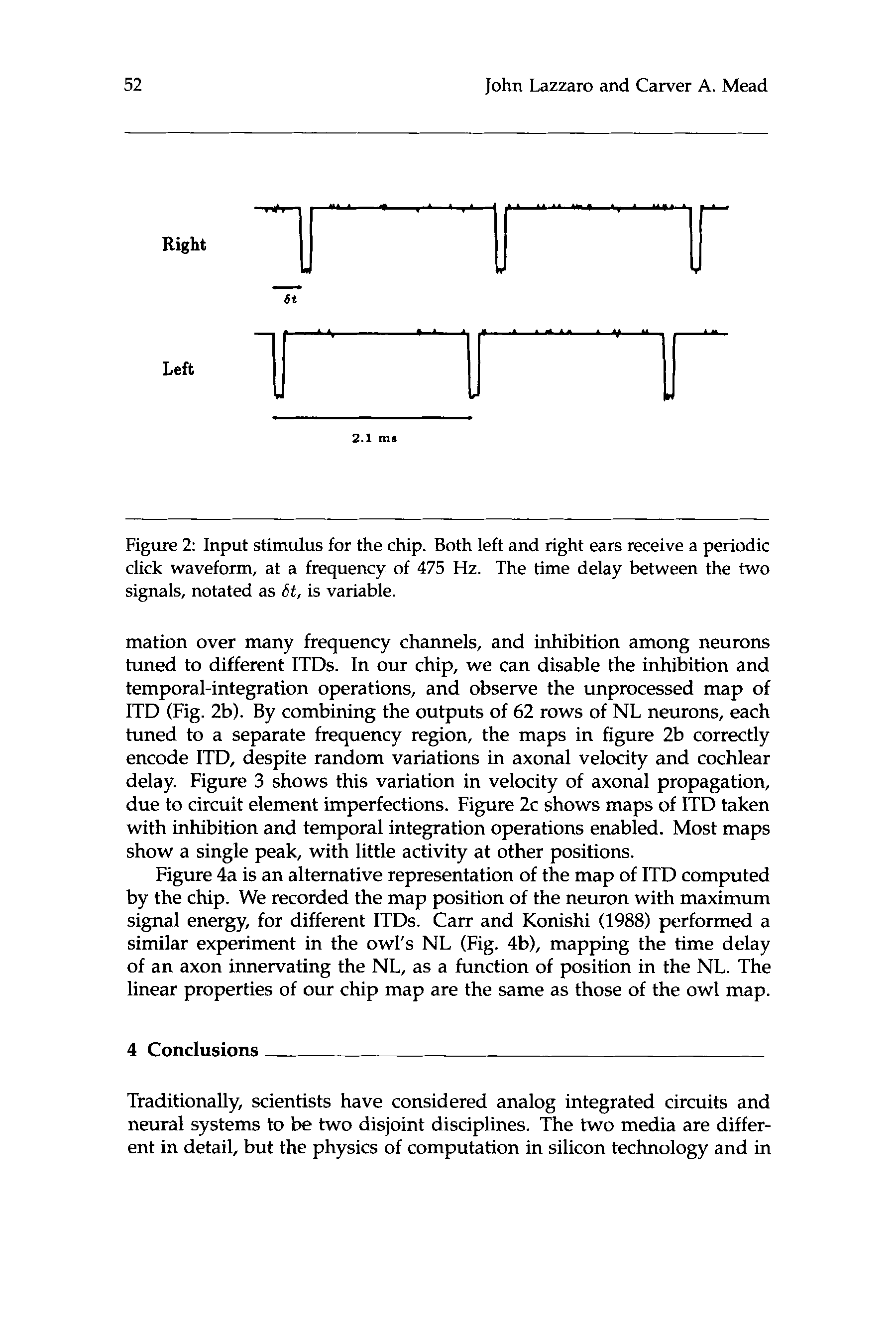
We presented periodic click stimuli to the chip (如图. 2A), and recorded
the final output of the chip, a map of ITD. Three signal-processing op-
erations, computed in the ICx and ICc of the owl, improve the original
encoding of ITDs in the NL: temporal integration, integration of infor-
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
n
e
C
哦
A
r
t
我
C
e
–
p
d
/
我
F
/
/
/
/
/
1
1
4
7
8
1
1
8
2
3
n
e
C
哦
1
9
8
9
1
1
4
7
p
d
.
.
.
.
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
52
John Lazzaro and Carver A. Mead
Right w
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
n
e
C
哦
A
r
t
我
C
e
–
p
d
/
我
F
/
/
/
/
/
1
1
4
7
8
1
1
8
2
3
n
e
C
哦
1
9
8
9
1
1
4
7
p
d
.
.
.
.
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
2.1 多发性硬化症
数字 2: Input stimulus for the chip. Both left and right ears receive a periodic
click waveform, at a frequency of 475 赫兹. The time delay between the two
signals, notated as 6t, is variable.
mation over many frequency channels, and inhibition among neurons
tuned to different ITDs. In our chip, we can disable the inhibition and
temporal-integration operations, and observe the unprocessed map of
ITD (如图. 2乙). By combining the outputs of 62 rows of NL neurons, each
tuned to a separate frequency region, the maps in figure 2b correctly
encode ITD, despite random variations in axonal velocity and cochlear
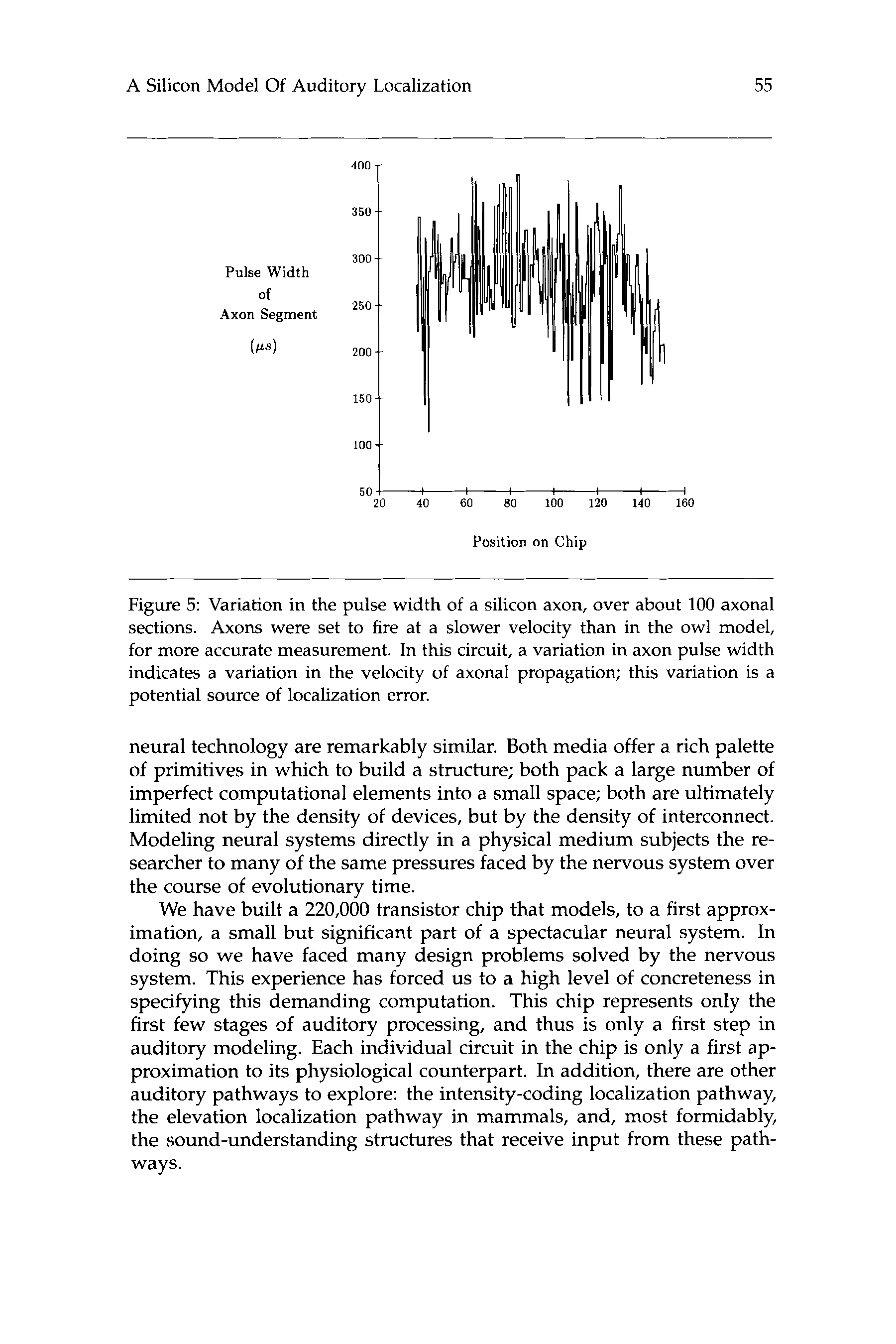
delay. 数字 3 shows this variation in velocity of axonal propagation,
due to circuit element imperfections. Figure 2c shows maps of ITD taken
with inhibition and temporal integration operations enabled. Most maps
show a single peak, with little activity at other positions.
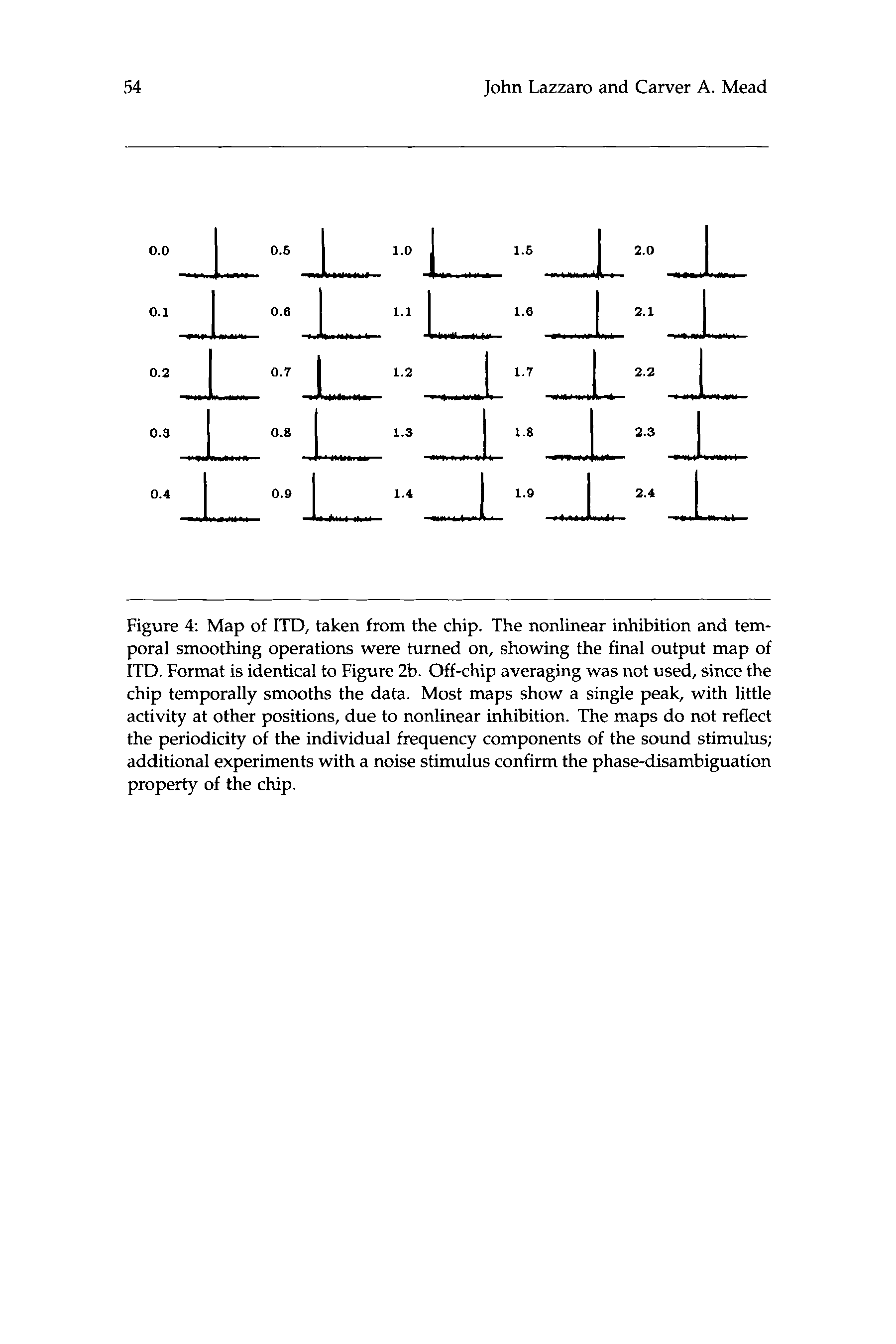
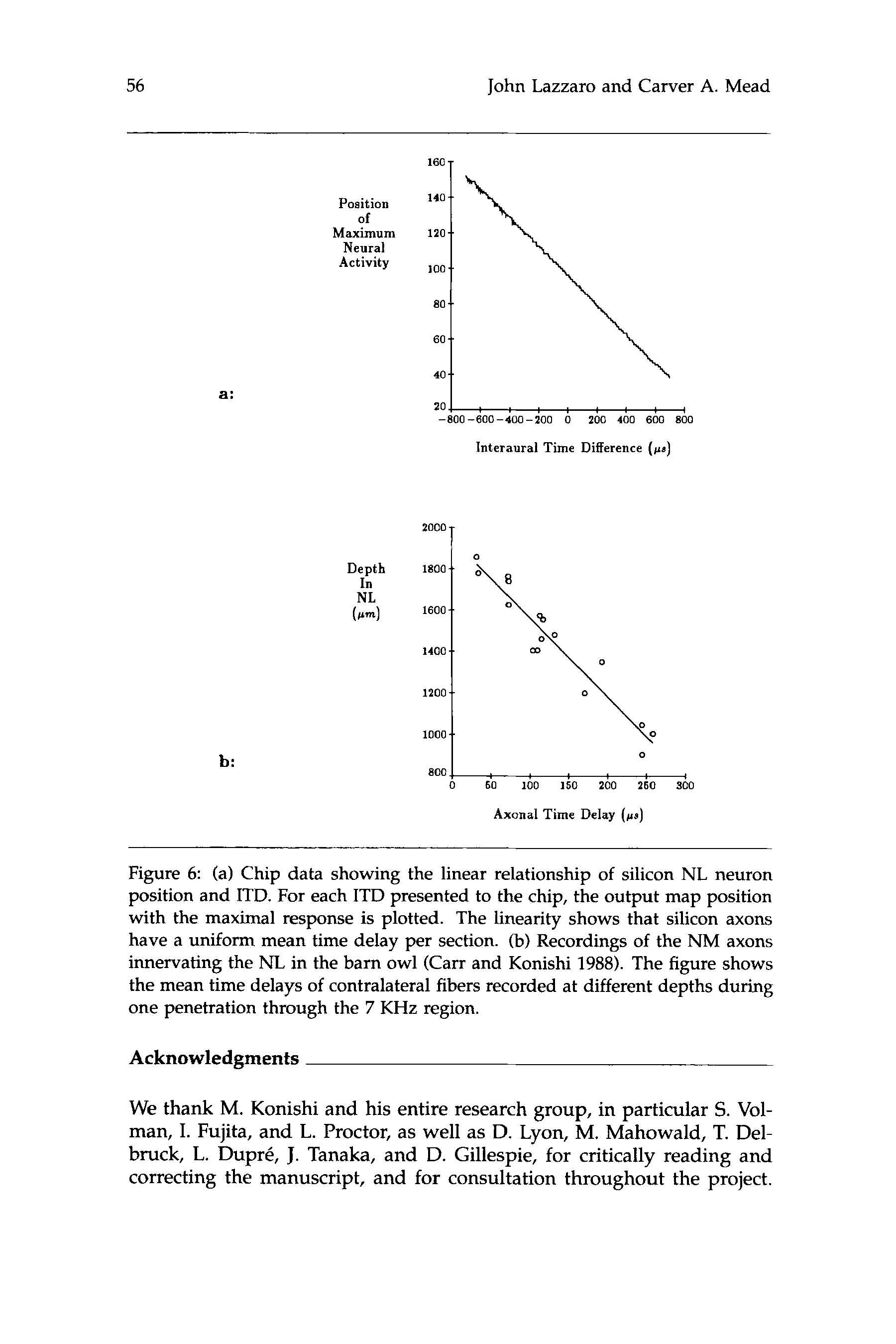
Figure 4a is an alternative representation of the map of ITD computed
by the chip. We recorded the map position of the neuron with maximum
signal energy, for different ITDs. Carr and Konishi (1988) performed a
similar experiment in the owl’s NL (如图. 4乙), mapping the time delay
of an axon innervating the NL, as a function of position in the NL. 这
linear properties of our chip map are the same as those of the owl map.
4 结论
传统上, scientists have considered analog integrated circuits and
neural systems to be two disjoint disciplines. The two media are differ-
ent in detail, but the physics of computation in silicon technology and in
A Silicon Model Of Auditory Localization
53
2.0
]Re*p0nse
0.0 A 0.6 & 1.0 JJkJlc 1.5
0.6 & 1.1
0.1
0.2 A 0.7
0.8 & 1.3
0.0 & 1.4 & 1.9
0.3
0.4
– Position
1.6 & 2.1
1.2 4 1.7
1.8 A 2.3
2.2 Jwl)JwL
2.4
数字 3: Map of ITD, taken from the chip. The nonlinear inhibition and tem-
poral smoothing operations were turned off, showing the unprocessed map of
ITD. The vertical axis of each map corresponds to neural activity level, 然而
the horizontal axis of each map corresponds to linear position within the map.
The stimulus for each plot is the periodic click waveforms of Figure 2a; 6t is
shown in the upper left corner of each plot, measured in milliseconds. 每个
map is an average of several maps recorded at 100 millisecond intervals; averag-
ing is necessary to capture a representation of the quickly changing, temporally
unsmoothed response. The encoding of ITD is present in the maps, but false
correlations add unwanted noise to the desired signal. Since we are using a
periodic stimulus, large time delays are interpreted as negative delays, 和
map response wraps from one side to the other at an ITD of 1.2 毫秒.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
n
e
C
哦
A
r
t
我
C
e
–
p
d
/
我
F
/
/
/
/
/
1
1
4
7
8
1
1
8
2
3
n
e
C
哦
1
9
8
9
1
1
4
7
p
d
.
.
.
.
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
54
John Lazzaro and Carver A. Mead
1.0 L , ,
1.6 1 2.0 1
0.0 1 0.5
0.1 1 0.6 1 1.1 我 1.6 A 2.1
0.2 __I 0.7 我 1.2 1 1.7 我 2.2 1
0.3 我 0.8 我 1.3 1 1.8
0.9 我 1.4 1 1.9
2.4 1 _ ,
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
2.3
d
我
r
e
C
t
.
0.4
米
:
/
/
我
t
.
p
t
t
我
数字 4: Map of ITD, taken from the chip. The nonlinear inhibition and tem-
poral smoothing operations were turned on, showing the final output map of
ITD. Format is identical to Figure 2b. Off-chip averaging was not used, 自从
chip temporally smooths the data. Most maps show a single peak, with little
activity at other positions, due to nonlinear inhibition. The maps do not reflect
the periodicity of the individual frequency components of the sound stimulus;
additional experiments with a noise stimulus confirm the phase-disambiguation
property of the chip.
/
e
d
你
n
e
C
哦
A
r
t
我
C
e
–
p
d
/
我
F
/
/
/
/
/
1
1
4
7
8
1
1
8
2
3
n
e
C
哦
1
9
8
9
1
1
4
7
p
d
.
.
.
.
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
A Silicon Model Of Auditory Localization
55
400 时间
—
350
—
300
-.
250
.-
200
150–
Pulse Width
的
Axon Segment
( P S )
loot,,,,,
50 20
40
60
80
100
120
140
160
Position on Chip
数字 5: Variation in the pulse width of a silicon axon, over about 100 axonal
sections. Axons were set to fire at a slower velocity than in the owl model,
for more accurate measurement. In this circuit, a variation in axon pulse width
indicates a variation in the velocity of axonal propagation; this variation is a
potential source of localization error.
neural technology are remarkably similar. Both media offer a rich palette
of primitives in which to build a structure; both pack a large number of
imperfect computational elements into a small space; both are ultimately
limited not by the density of devices, but by the density of interconnect.
Modeling neural systems directly in a physical medium subjects the re-
searcher to many of the same pressures faced by the nervous system over
the course of evolutionary time.
We have built a 220,000 transistor chip that models, to a first approx-
imation, a small but significant part of a spectacular neural system. 在
doing so we have faced many design problems solved by the nervous
系统. This experience has forced us to a high level of concreteness in
specifying this demanding computation. This chip represents only the
first few stages of auditory processing, and thus is only a first step in
auditory modeling. Each individual circuit in the chip is only a first ap-
proximation to its physiological counterpart. 此外, there are other
auditory pathways to explore: the intensity-coding localization pathway,
the elevation localization pathway in mammals, 和, most formidably,
the sound-understanding structures that receive input from these path-
方法.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
n
e
C
哦
A
r
t
我
C
e
–
p
d
/
我
F
/
/
/
/
/
1
1
4
7
8
1
1
8
2
3
n
e
C
哦
1
9
8
9
1
1
4
7
p
d
.
.
.
.
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
56
John Lazzaro and Carver A. Mead
Position
的
Maximum
Neural
Activity
20 – -800-600-400-200 0 200 400 600 800
D
哦
w
n
哦
A
d
e
d
Interaural Time Difference ( p a )
F
r
哦
米
H
p
t
t
我
:
/
/
d
我
r
e
C
t
.
米
我
t
.
Depth
在
NL
( r m )
时间
1800-.
1600-
1400-
1200.-
..
1000
800
0
0
60
100 150
200 260
Axonal Time Delay ( p s )
我
300
/
e
d
你
n
e
C
哦
A
r
t
我
C
e
–
p
d
/
我
F
/
/
/
/
/
1
1
4
7
8
1
1
8
2
3
n
e
C
哦
1
9
8
9
1
1
4
7
p
d
.
.
.
.
.
A:
乙:
数字 6: (A) Chip data showing the linear relationship of silicon NL neuron
position and ITD. For each ITD presented to the chip, the output map position
with the maximal response is plotted. The linearity shows that silicon axons
have a uniform mean time delay per section. (乙) Recordings of the NM axons
innervating the NL in the barn owl (Carr and Konishi 1988). The figure shows
the mean time delays of contralateral fibers recorded at different depths during
one penetration through the 7 KHz region.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
We thank M. Konishi and his entire research group, in particular S. 卷-
男人, 我. Fujita, 和L. Proctor, as well as D. Lyon, 中号. Mahowald, 时间. Del-
bruck, L. Dupr6, J. Tanaka, and D. Gillespie, for critically reading and
correcting the manuscript, and for consultation throughout the project.
A Silicon Model Of Auditory Localization
57
We thank Hewlett-Packard for computing support, a n d DARPA a n d MO-
SIS for chip fabrication. This work w a s sponsored by the Office of Naval
Research and the System Development Foundation.
参考
Carr, C.E. 和M. Konishi. 1988. Axonal Delay Lines for Time Measurement in
the Owl’s Brainstem. Proc. Nat. Acad. Sci. 85, 8311-8315.
Fujita, 我. 和M. Konishi. In preparation.
Jeffress, L.A. 1948. A Place Theory of Sound Localization. J. Cornp. Physiol.
Pyschol. 41, 35-39.
Knudsen, E.I., G.G. Blasdel, 和M. Konishi. 1979. Sound Localization by the
Barn Owl Measured with the Search Coil Technique. 1. 比较. Physiol. 133,
1-11.
Knudsen, E.I. 和M. Konishi. 1979. Mechanisms of Sound Localization in the
Barn Owl (Tyto alba). J. Cornp. Physiol. 133, 13-21.
. 1978. A Neural Map of Auditory Space in the Owl. 科学 200, 795-
797.
Lazzaro, J.P., S. Ryckebusch, 嘛. Mahowald, and C.A. Mead. 1988. Winner-
Take-All Networks of O(n) Complexity. Proc. I E E E Conf. Neural lnforrnation
Processing Systems, 丹佛, 一氧化碳.
Lyon, R.F. and C. Mead. 1988. An Analog Electronic Cochlea. IEEE Trans.
Acoust., Speech, Signal Processing 36, 1119-1134.
Mead, C.A. 1989. Analog V L S l and Neural Systems. Reading, 嘛: 艾迪生-
韦斯利.
Moiseff, A. 和M. Konishi. 1981. Neuronal and behavioral sensitivity to bin-
aural time differences in the owl. J. Neurosci. 1, 4048.
Takahashi, T.T. 和M. Konishi. 1988. Projections of the Nucleus Angularis and
Nucleus Laminaris to the Lateral Lemniscal Nuclear Complex of the Barn
Owl. J. Cornpar. Neurol. 274, 221-238.
瓦格纳, H. 和M. Konishi. In preparation.
已收到 26 十月; accepted 9 十一月 1988.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
n
e
C
哦
A
r
t
我
C
e
–
p
d
/
我
F
/
/
/
/
/
1
1
4
7
8
1
1
8
2
3
n
e
C
哦
1
9
8
9
1
1
4
7
p
d
.
.
.
.
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3