Both a Nicotinic Single Nucleotide Polymorphism (SNP)
and a Noradrenergic SNP Modulate Working Memory
Performance when Attention is Manipulated
Pamela M. Greenwood1, Ramya Sundararajan2, Ming-Kuan Lin2,
Reshma Kumar1, Karl J. Fryxell2, and Raja Parasuraman1
D
哦
w
n
我
哦
A
d
e
d
抽象的
& We investigated the relation between the two systems of
visuospatial attention and working memory by examining the
effect of normal variation in cholinergic and noradrenergic
genes on working memory performance under attentional
manipulation. We previously reported that working memory
for location was impaired following large location precues, 在-
dicating the scale of visuospatial attention has a role in form-
ing the mental representation of the target. In one of the first
studies to compare effects of two single nucleotide poly-
morphisms (SNPs) on the same cognitive task, we investigated
the neurotransmission systems underlying interactions be-
tween attention and memory. Based on our previous report
that the CHRNA4 rs#1044396 C/T nicotinic receptor SNP
affected visuospatial attention, but not working memory, 和
the DBH rs#1108580 G/A noradrenergic enzyme SNP affected
工作记忆, but not attention, we predicted that both
SNPs would modulate performance when the two systems
interacted and working memory was manipulated by attention.
We found the scale of visuospatial attention deployed around
a target affected memory for location of that target. 记忆
performance was modulated by the two SNPs. CHRNA4 C/C
homozygotes and DBH G allele carriers showed the best mem-
ory performance but also the greatest benefit of visuospatial
attention on memory. 全面的, 然而, the CHRNA4 SNP
exerted a stronger effect than the DBH SNP on memory per-
formance when visuospatial attention was manipulated. 这
evidence of an integrated cholinergic influence on working
memory performance under attentional manipulation is con-
sistent with the view that working memory and visuospatial
attention are separate systems which can interact. &
介绍
Beginning with Cowan (1988), there has been an impe-
tus toward a more comprehensive understanding of the
普通的
interaction between attention and memory.
There is evidence that separate storage and selection
processes exist, but increasing recognition that these
processes interact (Awh, 沃格尔, & 哦, 2006). Newer
theories of working memory argue that the focus of
visuospatial attention plays a central role within working
记忆, although the theories do not agree on the spe-
cifics of that role (考恩, 2001; Engle, Kane, & Tuholski,
1999). One approach has examined how the focus of vi-
suospatial attention influences the limited-capacity store
of working memory (康威, 考恩, & Bunting, 2001;
Baddeley, 1992). Using such an approach, 我们发现
visual working memory for a target location is more
accurate when the target was preceded by smaller com-
pared to larger precues (Greenwood, Lambert, Sunderland,
& Parasuraman, 2005). That finding extended current
views of the relation between visuospatial attention and
working memory by showing that dynamic adjustments
1George Mason University, Fairfax, VA, 2George Mason Univer-
城市, Manassas, VA
of the scale of visuospatial attention play a fundamental
role in controlling the encoded representation in work-
ing memory.
Neuroimaging and electrophysiological studies have
also revealed that the focus of visuospatial attention ex-
erts modulatory effects on posterior working memory
stores. This has been seen in inferior temporal (Ranganath,
2006) as well as striate and extrastriate regions ( 沃格尔,
McCollough, & Machizawa, 2005; Super, Spekreijse, &
瘫痪, 2001). Several authors recently reviewed the
evidence bearing on the role of prefrontal cortex (全氟碳化物)
in working memory (Postle, 2006; Curtis & D’Esposito,
2003), concluding PFC does not provide temporary stor-
age of information, but rather exerts attention-based
control over the posterior stores of working memory.
Consistent with that view is human evidence that manip-
ulations of visuospatial attention induce PFC to modu-
late extrastriate-generated ERPs in the P1 latency range
( Vogel et al., 2005; Yago, Duarte, 黄, Barcelo, &
骑士, 2004; Foxe & 辛普森, 2002). Electrophysiolog-
ical and electromagnetic recordings showed that direct-
ing the focus of attention did not alter the initial striate
response measured in fMRI, but did enhance later striate
活动 (140–250 msec) (Noesselt et al., 2002). 因此,
D 2008 麻省理工学院
认知神经科学杂志 21:11, PP. 2139–2153
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
t
F
r
p
哦
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
H
.
米
A
我
r
e
.
d
C
你
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
1
我
e
1
1
–
p
2
d
1
F
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
哦
3
C
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
8
e
.
s
t
2
哦
1
n
1
6
0
4
8
.
S
p
e
d
p
F
e
米
乙
y
乙
e
r
G
你
2
0
e
2
s
3
t
/
j
.
t
/
F
.
.
哦
n
1
8
中号
A
y
2
0
2
1
there is growing evidence that the attentional focus and
工作记忆, although separate systems mediated
in different brain regions, clearly interact.
Although fMRI and ERP studies have helped identify
the spatio-temporal properties of brain networks under-
lying visuospatial attention and working memory, 这些
techniques cannot speak directly to the innervation of
these networks or to the way gene expression influences
such innervation. Pharmacological and genetic methods
offer another avenue for examining these questions
(Posner, Rothbart, & Sheese, 2007; Fossella & Casey, 2006;
Parasuraman & Greenwood, 2004; Sarter & Bruno, 2004;
Greenwood & Parasuraman, 2003; Everitt & Robbins,
1997). 然而, the specific neurotransmission systems
involved in the influence of attention on working mem-
ory stores are not known. As reviewed below, the nic-
otinic cholinergic system is an important mediator of
visuospatial attention, whereas noradrenergic and dopa-
minergic systems have been shown to be important me-
diators of working memory. There is substantial animal
evidence reporting a role for muscarinic and nicotinic re-
ceptors in cognition generally, including attention, 学习-
英, and memory (莱文, McClernon, & Rezvani, 2006).
然而, some of the paradigms commonly used in
rodent studies of working memory require both atten-
tion and working memory, 例如, the radial-arm
maze. 而且, there is evidence that the influence of
cholinergic agents on delay-period effects on memory
tasks in primates are exerted through visuospatial at-
注意力 (Furey et al., 2000; Voytko, 1996; Robbins et al.,
1989).
Molecular genetics offers one way to investigate the
neurotransmission systems involved in attentional modu-
lation of working memory, viz, by using allelic association
methods in the context of performance on visuospatial
attention and working memory tasks. 这样的做法
does not involve the direct manipulation of neurotrans-
mission systems used in pharmacological studies, 但
rather examines the cognitive consequences of normal
variation in neurotransmission genes on the efficiency
of those systems. Several groups have used this ap-
proach with enzyme and receptor genes (Greenwood,
Fossella, & Parasuraman, 2005; Greenwood, Lambert, 等人。,
2005; Parasuraman, Greenwood, Kumar, & Fossella, 2005;
Goldberg & 温伯格, 2004; Greenwood & Parasuraman,
2003; Fossella et al., 2002).
In a previous investigation, we observed that whereas
working memory was modulated by variation in a nor-
adrenergic gene but not by variation in a nicotinic cho-
linergic gene, shifting (Parasuraman et al., 2005) 和
scaling (Greenwood et al., 2006) of visuospatial atten-
tion were modulated by variation in a nicotinic gene but
not a noradrenergic gene. We hypothesized that if a
processing overlap exists between visuospatial attention
and visual working memory (Greenwood, Lambert, 等人。,
2005; 考恩, 2001; Awh, Jonides, & Reuter-Lorenz, 1998),
then when visuospatial attention is experimentally ma-
nipulated within working memory, memory performance
should be modulated by variation in both cholinergic and
noradrenergic genes. 为了测试这个, we measured the effect
of normal variation in noradrenergic and cholinergic
neurotransmission genes in a working memory task in
which the focus of visuospatial attention was experimen-
tally manipulated.
Selection of genes for studying the mediation of
visuospatial attention and working memory in people
can be guided by what is known of the pharmacology of
those processes. Several groups have reported that
working memory performance is influenced by dopa-
mine (DA) D1 receptor availability (Abi-Dargham et al.,
2002; Castner, 威廉姆斯, & Goldman-Rakic, 2000). 也不-
adrenergic agonists also influence both working mem-
ory and PFC activity. 例如, Avery, Franowicz,
Studholme, van Dyck, and Arnsten (2000) observed that
an alpha-2A adrenoreceptor agonist improved working
memory performance and increased blood flow in pre-
frontal but not temporal cortex in monkeys. With regard
to visuospatial attention, disengagement of attention
from an invalidly cued location is influenced by choliner-
gic agonists. In both monkeys and humans, 管理-
tion of nicotine selectively enhanced the effects of invalid
cues on performance ( Witte, 戴维森, & Marrocco,
1997) and reduced effects of cue validity overall (Thiel,
Zilles, & Fink, 2005). Abstinence from nicotine in smokers
also selectively enhanced effects of invalid cues (Shirtcliff
& Marrocco, 2003).
Based on this evidence of nicotinic cholinergic medi-
ation of visuospatial attention and noradrenergic and
dopaminergic mediation of working memory, we previ-
ously tested hypotheses concerning the effect of normal
variation of genes in the noradrenergic and cholinergic
neurotransmission pathways on working memory and
visuospatial attention, 分别. We reported that
normal variation in a gene that encodes dopamine beta
hydroxylase (DBH), an enzyme that converts DA to nor-
epinephrine (NE) in synaptic vesicles, modulated work-
ing memory for location. The G allele of the 444 G/A SNP
(rs#1108580) in the DBH gene was previously associat-
ed with higher DBH enzyme activity levels in serum
(Cubells et al., 1998) and reduced risk of paranoid
ideation in depressed patients ( Wood, Joyce, 磨坊主,
Mulder, & 肯尼迪, 2002). We found that the G allele
was associated with superior spatial working memory
performance but did not modulate visuospatial atten-
的, either attentional shifting (Parasuraman et al., 2005)
or attentional scaling (Greenwood, Fossella, 等人。, 2005).
With regard to genes with a role in attention, we re-
ported that the ability to shift visuospatial attention was
modulated by normal variation in a nicotinic receptor
gene. The alpha4/beta2 nicotinic cholinergic receptor is
the most widely distributed nicotinic receptor in the CNS
(Flores, 罗杰斯, Pabreza, 沃尔夫, & Kellar, 1992). 这
CHRNA4 gene codes the alpha4 subunit of the nicotinic
cholinergic receptor and has an SNP (termed 1545 C/T,
2140
认知神经科学杂志
体积 21, 数字 11
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
t
F
r
p
哦
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
H
.
米
A
我
r
e
.
d
C
你
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
1
我
e
1
1
–
p
2
d
1
F
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
哦
3
C
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
8
e
.
s
t
2
哦
1
n
1
6
0
4
8
.
S
p
e
d
p
F
e
米
乙
y
乙
e
r
G
你
2
0
e
2
s
3
t
/
j
F
t
.
/
.
.
哦
n
1
8
中号
A
y
2
0
2
1
rs#1044396) previously found to be associated with nic-
otine addiction (Feng et al., 2004) but not with attention
deficit hyperactivity disorder (Kent et al., 2001). Shifting
the focus of visuospatial attention was modulated by
the CHRNA4 rs#1044396 C/T SNP but not by the DBH
rs#1108580 G/A SNP (Parasuraman et al., 2005). Adjust-
ing the size (规模) of the focus of visuospatial attention
(Greenwood, Sunderland, Putnam, 征收, & Parasuraman,
2005; Greenwood & Parasuraman, 2004) was modulated
by the same CHRNA4 SNP but not by the DBH SNP
(Greenwood, Lambert, 等人。, 2005).
There is also evidence that the cholinergic system
does influence some aspects of memory. 一般来说, nic-
otinic agonists benefit memory and nicotinic antagonists
impair it. 然而, it appears that only certain types of
memory are influenced and only under certain condi-
系统蒸发散 (Dani & Bertrand, 2007). The work of Levin and
西蒙 (1998) in rodents shows that such influences
occur most often when the task is difficult or the animal
cognitively impaired. 而且, the 8-arm radial maze
used in most rodent studies of working memory may
require the effective use of attention as well as working
记忆. Consistent with that interpretation, Voytko
等人. (1994) found that lesions of the nucleus basalis of
Meynert (nbM) in monkeys did not impair performance
on several working memory tasks (delayed nonmatch-
to-sample, delayed response) but did impair the ability
to shift visuospatial attention. Such findings led Voytko
(1996) to conclude that cholinergic dysfunction in the
primate basal forebrain cholinergic system leads to deficits
of attention rather than memory. Everitt and Robbins
(1997) and Robbins et al. (1989) have also argued that
apparent cholinergic modulation of delay-period effects
on performance may be due to cholinergic modulation of
visuospatial attention rather than of working memory.
Understanding interactions between cognitive systems
can be advanced by converging evidence from different
methodologies. The interaction between visuospatial
attention and working memory has been investigated be-
haviorally (例如, Awh & Jonides, 1998), electrophysiolog-
ically (例如, Vogel et al., 2005; 沃格尔 & Machizawa, 2004),
and pharmacologically (例如, Everitt & Robbins, 1997;
Voytko, 1996). In the present study, we investigate the
question using genetics. 在之前的工作中, we have argued
that evidence of genetic dissociation on cognitive perfor-
mance indicates dissociation in the underlying innervation
(Greenwood, Fossella, 等人。, 2005). In the present study,
we apply that logic to investigate the interaction between
visuospatial attention and working memory. We know
from our previous observation of a double dissociation
that CHRNA4 rs#1044396 modulates visuospatial atten-
的, but not working memory, whereas DBH rs#1108580
modulates working memory, but not visuospatial atten-
的 (Parasuraman et al., 2005). What has not been in-
vestigated previously is the effect of these SNPs when
working memory is manipulated by attention. Such an
investigation allows us to move beyond cataloging the
separate effects of neurotransmission SNPs on compo-
nent processes of cognitive functions to greater under-
standing of the normal interaction between component
流程. Based on our previously reported evidence of
specificity in the effect of neurotransmission genes on
visuospatial attention and working memory, we predicted
that both cholinergic and noradrenergic neurotransmis-
sion SNPs would modulate performance when working
memory was manipulated by attention. We manipulated
visuospatial attention in the size—hence, the precision—
of precues to target location and measured the accuracy
of memory for that location after 3 秒 (数字 1). 我们也
manipulated working memory difficulty by varying the
distance between the target location and the test location
when the two did not match.
方法
参加者
Participants were recruited from Catholic University,
George Mason University, and the Washington, DC com-
社区. All procedures were approved by the insti-
tutional review boards of each university and were
performed in accordance with the 1964 Declaration of
Helsinki. All persons gave informed consent prior to
their inclusion in the study. Informed consent was ob-
tained and vision was tested to ensure at least 20/30 六-
锡安 (after correction,
如果需要的话) on a Rosenbaum
pocket screener. Participants included both young and
old but the sample was not large enough to allow
analysis by both age group and genotype. All participants
were screened by questionnaire for neurologic and psy-
chiatric illness. All participants were cognitively screened
by means of the Wechsler WAIS Vocabulary subtest
( Wechsler, 1981) and the Wechsler Memory Scale—
Revised Logical Memory subtest (Wechsler, 1987). 到
eliminate individuals with a dementing illness, partici-
pants aged 65 years and older were additionally screened
with the Mini-Mental State Exam. Demographic informa-
tion for each SNP and for young and old groups is
provided in Table 1. There were no significant differences
in performance on these standardized neuropsychologi-
cal tests whether individuals were grouped by CHRNA4
rs#1044396 C/T or DBH rs#1108580 G/A genotype.
Genotyping
Buccal (cheek) swabs were obtained from each partici-
pant. Genomic DNA was purified from these swabs as di-
rected by the manufacturer (MasterAMP TMBuccal Swab
DNA Extraction Kit, Epicentre Technologies, 麦迪逊, WI).
Participants were genotyped (double-blind) for a nicotin-
ic receptor SNP (rs#1044396, originally termed CHRNA4
1545 C/T by Steinlein et al., 1997) and a noradrenergic
enzyme SNP (rs#1108580, originally termed DBH 444 G/A;
Cubells et al., 1998) 如下所述.
Greenwood et al.
2141
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
t
F
r
p
哦
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
H
.
米
A
我
r
e
.
d
C
你
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
1
我
e
1
1
–
p
2
d
1
F
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
哦
3
C
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
8
e
.
s
t
2
哦
1
n
1
6
0
4
8
.
S
p
e
d
p
F
e
米
乙
y
乙
e
r
G
你
2
0
e
2
s
3
t
/
j
.
/
.
.
F
t
哦
n
1
8
中号
A
y
2
0
2
1
桌子 1. Demographics of Participant Groups (Means and
Standard Deviations)
CHRNA4 rs#1044396 Genotype Groups
Old
Sample size
年龄
性别 (F/M)
WAIS-R Vocabularya
Logical memoryb
Immediate
Delayed
C/C
C/T
T/T
26
39
31
70.0 (4.7)
71.5 (5.1)
71.7 (5.2)
11/15
27/12
15/16
50.1 (8.1)
58.3 (7.4)
59.2 (7.4)
10.2 (3.8)
10.7 (3.8)
10.6 (4.4)
8.2 (4.2)
8.4 (3.7)
8.6 (5.3)
Mini-Mental State Examc
29.0 (1.2)
28.6 (1.3)
27.8 (2.1)
Young
Sample size
年龄
性别 (F/M)
15
50
17
20.0 (2.1)
19.6 (1.7)
20.1 (2.1)
12/3
37/13
10/7
WAIS-R Vocabularya
45.1 (11.9)
47.3 (10.2)
48.3 (6.6)
Logical memory (WMS-R)乙
Immediate
Delayed
11.4 (3.7)
11.1 (3.9)
12.6 (3.6)
10.1 (3.5)
9.9 (3.8)
11.2 (3.7)
DBH rs#1108580 Genotype Groups
Old
Sample size
年龄
性别 (F/M)
A/A
A/G
G/G
37
60
23
71.9 (4.4)
71.0 (5.5)
70.5 (4.8)
14/23
36/24
15/8
WAIS-R Vocabularya
59.6 (7.8)
59.6 (7.3)
57.4 (7.2)
Logical memory (WMS-R)乙
Immediate
Delayed
10.3 (3.9)
11.1 (4.2)
10.1 (3.9)
8.7 (4.3)
8.6 (4.7)
8.0 (3.6)
Mini-Mental State Examc
27.8 (2.0)
28.7 (1.8)
28.5 (1.2)
Young
Sample size
年龄
性别 (F/M)
45
55
33
19.9 (2.1)
19.9 (2.0)
19.5 (1.7)
30/15
35/20
25/8
WAIS-R Vocabularya
50.4 (10.9)
48.0 (8.4)
48.8 (8.8)
Logical memory (WMS-R)乙
Immediate
Delayed
aWechsler (1981).
bWechsler (1987).
11.3 (3.1)
12.4 (3.9)
10.8 (4.4)
10.2 (3.0)
11.2 (4.0)
9.9 (4.6)
cAdministered only to those older than 65 年 (Folstein, Folstein, & McHugh, 1975).
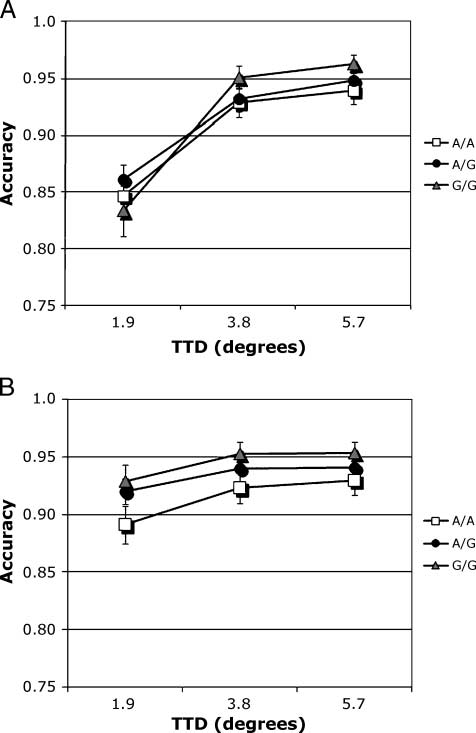
数字 1. Illustration of task manipulating the scale of visuospatial
attention at the location to be remembered over 3 秒. Circular
precues varying in size (1.68, 5.28, 和 8.18) preceded a black target dot
(100 msec duration) with an SOA of 500 毫秒. 有一个
requirement to remember dot location over a 3-sec delay. After the
delay, a decision was required indicating whether the location of a red
test dot matched that of the target dot. (A) Match trial, with test
location matching target location. (乙) Nonmatch trial illustrated with
test dot at one of three distances (1.98, 3.88, 5.78) from target location.
In order to analyze the CHRNA4 rs#1044396 C/T SNP,
a PCR fragment 309 bp in length was amplified from
genomic DNA in a reaction volume of 50 铝, using PTC-
100 or PTC-200 thermal cyclers (MJ Research). The am-
plified PCR fragment was purified on AMPure magnetic
beads (Agencourt) and the DNA sequence of this PCR
fragment was determined by cycle sequencing with BigDye
terminators on an ABI 310 capillary sequencer. We and
others have previously shown that this method allows
reliable determination of both homozygous and hetero-
zygous genotypes (Hare & Palumbi, 1999).
In order to analyze the DBH rs#1108580 G/A SNP, 我们
chose an alternative method that combined nested PCR,
plus allele-specific Tm-shift primers and automated melt-
ing curve analysis ( 王等人。, 2005). First an ‘‘external’’
PCR fragment was preamplified for 20 cycles in a reac-
tion volume of 50 Al in PTC-100 or PTC-200 thermal
cyclers (MJ Research). Then a 1-Al aliquot of the first-
round PCR was reamplified with two or three ‘‘internal’’
primers (which primed inside the original PCR frag-
蒙特), in a reaction volume of 15 Al on a Bio-Rad iCycler.
The internal primers included two allele-specific Tm-shift
primers, which were designed as described (王等人。,
2005). The allele-specific Tm-shift primers were used
both separately and together (IE。, in three PCR reac-
系统蒸发散), all of which also contained a common reverse
primer. DBH alleles were scored by automated analysis
of the melting curves of the PCR products (using Bio-
Rad iCycler software). The assay is designed so that the
2142
认知神经科学杂志
体积 21, 数字 11
D
哦
w
n
我
哦
A
d
e
d
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
F
r
哦
米
D
H
哦
t
w
t
n
p
哦
:
A
/
d
/
e
米
d
我
t
F
r
p
哦
米
r
C
H
.
s
p
我
我
d
v
我
e
r
e
r
C
C
t
H
.
米
A
我
r
e
.
d
C
你
哦
哦
米
C
/
n
j
A
哦
r
C
t
我
n
C
/
e
A
–
p
r
d
t
我
2
C
1
我
e
1
1
–
p
2
d
1
F
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
哦
3
C
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
哦
p
C
d
n
.
乙
y
2
0
G
0
你
8
e
.
s
t
2
哦
1
n
1
6
0
4
8
.
S
p
e
d
p
F
e
米
乙
y
乙
e
r
G
你
2
0
e
2
s
3
t
/
j
/
.
t
.
F
.
哦
n
1
8
中号
A
y
2
0
2
1
two possible PCR products (incorporating one or the
other allele-specific Tm-shift primer) differ in melting tem-
perature because of a GC-tail that is included in the 50 结尾
of one of the allele-specific primers (王等人。, 2005).
Nine participants were excluded for low accuracy (perfor-
mance at a level less than chance). Seven of the excluded
were over 65 年龄. For CHRNA4 rs#1044396 C/T,
there were 41 C/C homozygotes, 89 C/T heterozygotes,
48 T/T homozygotes. For DBH rs#1108580 G/A, there were
82 A/A homozygotes, 115 A/G heterozygotes, 和 56 G/G
homozygotes. SNPs in both samples were in Hardy–
Weinberg equilibrium ( p > .05 in respective x2 tests).
Stimuli and Procedures
A cued working memory task was used (数字 1). 这
task was designed to manipulate the accuracy of mem-
ory for location by varying the precision of location
precues to that location. Following a 1-sec duration
fixation cross, a circular cue appeared for 500 msec in
1 的 12 randomly selected locations on the screen and in
1 的 3 of visual angles in size (1.68, 5.28, 和 8.18). At cue
抵消, one black target dot (0.678 in diameter) appeared
centered within the cue for 100 毫秒. 在 10% 的考验,
the cues were invalid and the target appeared outside
the cue. This condition was included to reduce predict-
ability of the cue. Data from these trials were not ana-
lyzed, as there were insufficient numbers. When cues
were valid, the target was always centered in the cue. 在
target offset, a 3-sec delay began during which time only
the fixation cross was visible. After the delay, 屏幕
cleared and a red test dot appeared either at the same
location as the target dot (match trial) or at a different
地点 (nonmatch trial). This trial type manipulation
(match/nonmatch) provided a memory load manipula-
的. On nonmatch trials, the distance between the tar-
get location and the test dot varied (target–test distance
[TTD]), 存在 1.98, 3.88, 或者 5.78 apart. The closest dis-
tance required the most accurate memory. The red test
dot remained visible for 2 sec during which a same/
different decision was required. Both accuracy and re-
action time (RT) were measured but the instructions
emphasized accuracy.
In an additional sample of participants not yet geno-
打字的, the same task was administered, but with a no-
cue condition added to the other conditions. 这是
done to confirm the beneficial nature of the cue on task
表现. The duration of the fixation was length-
ened on no-cue trials to preserve the timing between
fixation and target onset.
结果
All statistical tests were performed at the .05 level of
significance. Repeated measures F values were corrected
for violations from sphericity. The task was designed to
assess effects of genotype on accuracy of memory for
target location after a delay, under an attentional manip-
计算. 所以, percent correct is the most useful
措施. 此外, some analyses of the RT data were
carried out.
Reaction Time Analyses
Median RTs were analyzed in an omnibus ANOVA for
each SNP. To allow comparisons between match and
nonmatch conditions, data were collapsed across TTD.
When the data were grouped by CHRNA4, there was no
main effect of genotype. Median RTs were slower on
nonmatch compared to match trials [F(1, 174) = 4.05].
RT was speeded slightly but significantly [F(2, 348) =
5.04] as the size of the cues increased (667, 661, 654 毫秒
for sizes 1.68, 5.28, 和 8.18, 分别). Cue size inter-
acted with CHRNA4 genotype [F(4, 348) = 2.49; 如图-
ure 2A]. There was also a nonsignificant trend in the
interaction of Trial type (西德:2) Cue size (西德:2) Age group [F(4,
348) = 1.91, p < .11]. There were no other significant
effects. Nonmatch RT data were analyzed over levels of
TTD, revealing no effect of CHRNA4 genotype.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
1
-
p
2
d
1
f
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
o
3
c
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
1
n
1
6
0
4
8
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
f
.
.
.
t
/
o
n
1
8
M
a
y
2
0
2
1
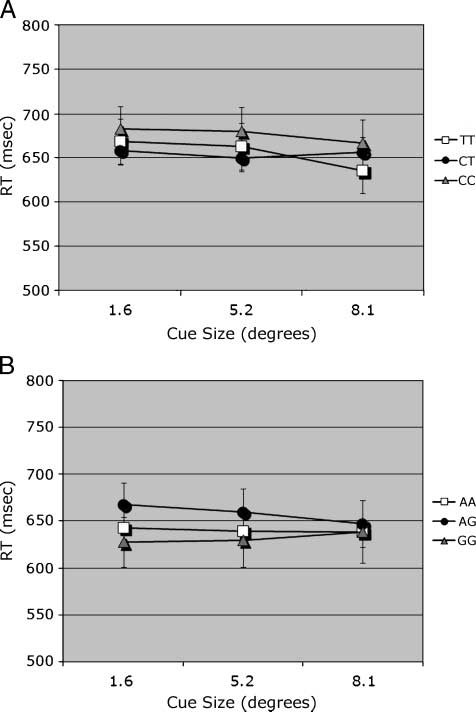
Figure 2. RT measure. (A) Interaction of Cue size (cid:2) CHRNA4 1044396
in an omnibus analysis (collapsed across TTD). (B) Interaction of
Cue size (cid:2) DBH rs#1108580 (cid:2) Trial type; match trial plotted.
Greenwood et al.
2143
When the data were grouped by DBH genotype, there
was no effect of DBH. There was a significant main effect
of cue size [F(2, 500) = 4.88], with RT decreasing as cue
size increased (645, 641, and 631 msec for sizes 1.68,
5.28, and 8.18, respectively). There were significant inter-
actions of Trial type (cid:2) Cue size [F(2, 500) = 3.51] and
DBH (cid:2) Trial type (cid:2) Cue size [F(4, 500) = 2.41]. The
latter interactions can be attributed to a genotype effect
on match trials, which decreased with cue size (Figure 2B).
Nonmatch RT data were analyzed over levels of TTD,
revealing no effect of DBH genotype.
Accuracy Analyses
Accuracy ratios (defined as number correct/number pre-
sented) were subjected to repeated measures ANOVA
with factors as described below.
Age Effects
Because the sample was heterogeneous with regard to
age, effect of age group was initially assessed. An omni-
bus ANOVA compared performance of young and old
(between-subjects factor of age group) on the within-
subjects factor of trial type (match, nonmatch). As only
nonmatch trials had levels of TTD, data were averaged
over those distances on nonmatch trials. The age groups
did not differ on working memory accuracy overall, but
there was a significant interaction of Trial type (cid:2) Age
group [F(1, 252) = 3.96]. The old group was slightly
more accurate than the young on match trials (means of
0.882 for old, 0.867 for young), but slightly less accurate
on nonmatch trials (means of 0.919 for old and 0.924
for young). There were no other significant interactions
with age group. Accuracy was lower on match trials over-
all [task type: F(1, 252) = 83.03] and highest following
the medium-sized cue [cue size: F(2, 504) = 3.87].
These effects interacted [F(2, 504) = 4.78]. Despite the
absence of age effects, it remains possible that age might
drive some of the genotype effects. To assess that
possibility, effects of age group were analyzed for each
SNP. There were no main effects of age group and no
interactions involving age group and genotype. The
criterion used for this analysis was p < .10. In light of
the weak effects of age, and the low numbers in some
cells when the sample was divided by both genotype and
age, subsequent analyses were conducted without age
group as a factor.
CHRNA4 C/T (rs#1044396)
Because TTD could be varied only under nonmatch con-
ditions, an omnibus ANOVA compared trial type (match,
nonmatch) by averaging across TTD on nonmatch trials.
The within-subjects factor was trial type (match, nonmatch).
The between-subjects factor was CHRNA4 rs#1044396
genotype (T/T, T/C, C/C).
The omnibus analysis required collapsing across TTD
to allow comparisons between match and nonmatch con-
ditions. This analysis revealed significant main effects of
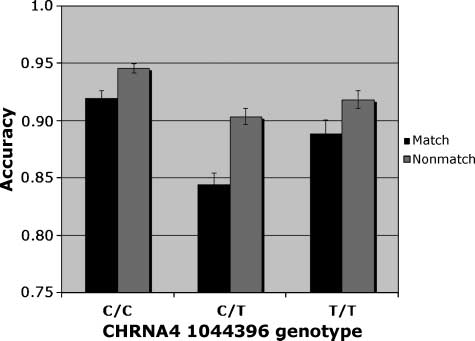
CHRNA4 genotype [F(2, 174) = 4.15, h2 = .05], trial type
[F(1, 174) = 37.40], and cue size [F(2, 348) = 7.29].
Accuracy was highest in the C/C homozygotes (mean =
0.93). Both the Trial type (cid:2) Cue size interaction [F(2,
348) = 5.23] and the Trial type (cid:2) CHRNA4 genotype
interaction [F(2, 174) = 3.21, h2 = .01] were significant.
The three-way interaction was not significant. The C/C
homozygotes were most accurate and the C/T hetero-
zygotes were least accurate, particularly on match trials.
Although heterozygotes are expected to perform at
an intermediate level between homozygote groups (as
we have reported previously; Greenwood, Fossella,
et al., 2005), we do not invariably observe that result
(e.g., Greenwood, Lambert, et al., 2005). Based on
the Trial type (cid:2) CHRNA4 interaction (Figure 3), simple
main effects of genotype were calculated at each level of
trial type.
On match trials, there was a significant effect of CHRNA4
genotype [F(2, 174) = 4.76, h2 = .055], with accuracy
highest in C/C homozygotes. Simple effects of CHRNA4
reveal that, after using a Bonferroni adjustment for mul-
tiple comparisons, only the C/C genotype differed sig-
nificantly from the C/T genotype (mean difference =
0.095). Using a formula for unequal samples (Cohen,
1988), the size of the main effect of CHRNA4 was
calculated to be 0.25. In light of the concern about false
positives in candidate gene studies, it can be noted that
this effect size is close to the value of 0.29 recently
reported in a meta-analysis of the effect of the COMT
val158met polymorphism on the Wisconsin Card Sort
Task, a test of executive function (Barnett, Jones, Robbins,
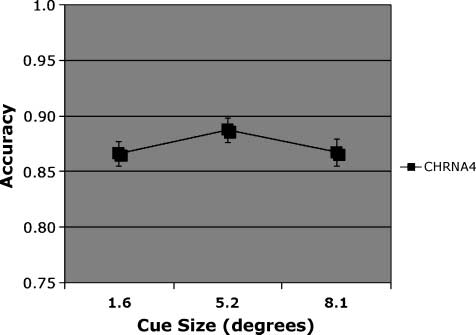
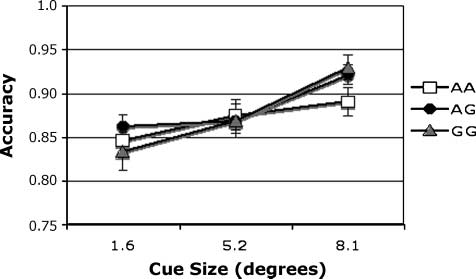
& Muller, 2007). With regard to cue size, accuracy was
highest following the medium-sized cue [Figure 4; F(2,
348) = 4.07], consistent with our previous report on this
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
1
-
p
2
d
1
f
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
o
3
c
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
1
n
1
6
0
4
8
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
/
.
.
.
t
f
o
n
1
8
M
a
y
2
0
2
1
Figure 3. Accuracy. Interaction of CHRNA4 genotype (cid:2) Trial type
in an omnibus analysis (collapsed across TTD) of individuals genotyped
for CHRNA4 rs#1044396 C/T.
2144
Journal of Cognitive Neuroscience
Volume 21, Number 11
genotype groups. This suggests that the C/C genotype
obtained greater benefits from the large cue when the
discrimination was difficult.
DBH rs#1108580 G/A
Omnibus
Collapsing across TTD allowed comparisons between
match and nonmatch conditions (trial type) in one
omnibus analysis. That analysis revealed no main effect
of DBH genotype. There were significant main effects of
trial type [F(1, 250) = 82.43] and cue size [F(2, 500) =
3.67], which interacted [Trial type (cid:2) Cue size: F(2,
500) = 5.17]. Trial type also interacted with DBH [F(2,
250) = 4.58, h2 = .013]. Based on the interaction of
Trial type (cid:2) DBH, separate analyses were carried out for
DBH at each level of trial type (match and nonmatch).
On match trials, when the test stimulus appeared at
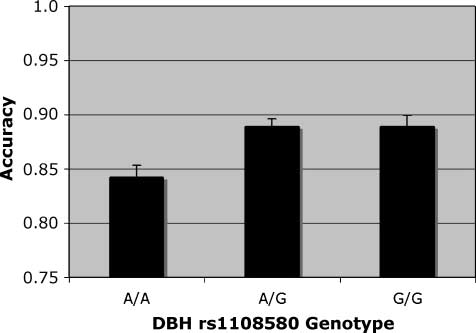
the target location, accuracy was higher in the DBH
rs#1108580 G carriers and lower in the A/A homozy-
gotes [Figure 6; F(2, 250) = 3.55]. Simple main effects
analysis revealed that only A/A and A/G differed signifi-
cantly. Using a formula for unequal samples (Cohen,
1988), the effect size ( f ) was calculated to be .18, pro-
ducing power of .75 for the sample. As with CHRNA4
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
1
-
p
2
d
1
f
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
o
3
c
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
1
n
1
6
0
4
8
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
/
.
t
.
.
f
o
n
1
8
M
a
y
2
0
2
1
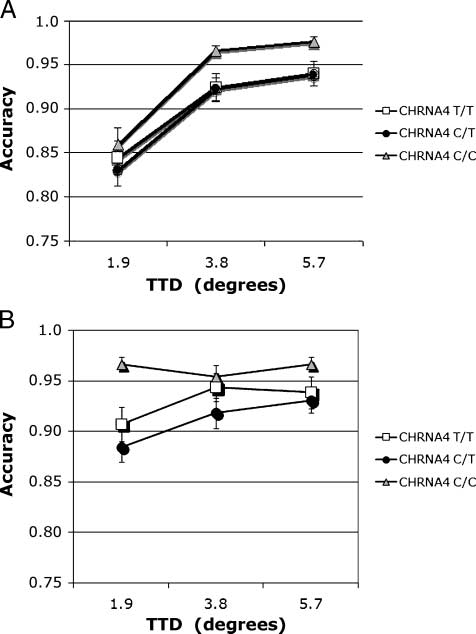
Figure 5. (A, B) Accuracy. Nonmatch condition, interaction of Cue
size (cid:2) TTD in individuals genotyped for CHRNA4 rs#1044396 C/T.
(A) Cue size 1 = 1.68. (B) Cue size 3 = 8.18.
Greenwood et al.
2145
Figure 4. Accuracy. Match condition, main effect of cue size on
accuracy in individuals genotyped for CHRNA4 rs#104439 C/T.
task from a different population (Greenwood, Lambert,
et al., 2005). These two factors did not interact.
On nonmatch trials, CHRNA4 C/C homozygotes were
again the most accurate but that was only a nonsignif-
icant trend ( p = .07). Accuracy increased with cue size
[F(2, 348) = 14.19]. A full analysis of nonmatch trials was
also conducted, without the constraints imposed by the
omnibus analysis.
Full Analysis of Nonmatch Trials
A full analysis was carried out on nonmatch trials, with
CHRNA4 genotype as the between-subjects factor and
cue size and TTD as within-subjects factors. Memory
accuracy was highest in the C/C homozygotes but that
was only a nonsignificant trend ( p = .07). Accuracy
increased with cue size [F(2, 348) = 14.19], but that
effect was strongest when target and test stimuli loca-
tions were close together, making the match/nonmatch
judgment difficult. These results were reflected in a
main effect of TTD [F(2, 348) = 106.39] and a TTD (cid:2)
Cue size interaction [F(4, 696) = 25.41]. Overall, per-
formance improved as cue size increased and as TTD
increased. These factors interacted with CHRNA4 geno-
type [Cue size (cid:2) TTD (cid:2) CHRNA4: F(8, 696) = 1.96, h2 =
.01). This interaction was analyzed using simple main
effects. Simple main effects of TTD and genotype at each
level of cue size were conducted to determine the
source of that interaction. Results from the smallest
cue are plotted in Figure 5A and from the largest cue
in Figure 5B. The two smaller cue size conditions did not
yield significant effects. At the largest cue size, main
effects of CHRNA4 [F(2, 175) = 3.56], TTD [F(2, 350) =
8.61], and their interaction were significant [Figure 5A
and B; F(4, 350) = 3.56]. When adjusted for the number
of comparisons, the interaction remained significant.
Following the large cue, C/C homozygotes were more
accurate at the closest 1.98 TTD compared to the other
that all participants had reduced accuracy when test and
target were very close. However, this effect was reduced
when the cue was large. To further understand this
interaction, we calculated the simple effects of DBH (cid:2)
Cue size at the shortest target–test difference. Accuracy
increased with cue size [F(2, 500) = 43.96] and G allele
carriers benefited more from the largest cue than the
A/A group [Figure 8; F(4, 500) = 3.18]. This result did
survive Bonferroni correction.
Direct Comparison of Both SNPs
An exploratory analysis was carried out on people geno-
typed for both SNPs which allows comparison of the two
SNPs in one analysis. This was considered exploratory
because of the small size of some of the cells. The largest
group (DBH A/G and CHRNA4 C/T) had 37 individuals,
whereas the smallest group (DBH G/G, CHRNA4 T/T)
had only 7 individuals. In an omnibus analysis involving
both SNPs and collapsed across TTD, there was a main
effect of CHRNA4 [F(2, 170) = 4.37, h2 = .05], but no
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
1
-
p
2
d
1
f
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
o
3
c
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
1
n
1
6
0
4
8
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
f
.
t
.
/
o
n
1
8
M
a
y
2
0
2
1
Figure 7. (A. B) Accuracy. Nonmatch condition, interaction of Cue
size (cid:2) TTD (cid:2) Genotype in individuals genotyped for DBH rs#1108580
SNP. (A) Cue size 1 = 1.68. (B) Cue size 3 = 8.18.
Figure 6. Accuracy. Match condition, main effect of DBH rs#1108580
genotype on accuracy in simple effects analysis.
analyses, the best performance was seen with the medium-
sized cue, but that effect was not significant and there was
no interaction.
On nonmatch trials (collapsed across TTD), simple
effects of DBH revealed that the only significant effect
was due to an increase in accuracy with cue size [F(2,
250) = 12.47].
Full Analysis of Nonmatch Trials
A full analysis was carried out on nonmatch trials, when
the test stimulus appeared in a different location from
the target. DBH genotype was the between-subjects
factor, whereas cue size and TTD were within-subjects
factors. There was no main effect of DBH on nonmatch
trials. As with the CHRNA4 SNP, accuracy increased with
cue size [F(2, 500) = 12.47]. There was also a main effect
of TTD [F(2, 500) = 161.24]. The effects of cue size were
strongest under difficult task conditions when target and
test were close together [Cue size (cid:2) TTD: F(4, 1000) =
31.30]. Under those conditions, the large precue was
associated with better performance. This effect inter-
acted with DBH genotype [DBH (cid:2) Cue size (cid:2) TTD: F(8,
1000) = 2.55, h2 = .006]. The interaction was analyzed
by simple effects of TTD and DBH at each level of cue
size. On nonmatch trials when the cue was smallest,
accuracy increased with TTD [Figure 7A; F(2, 500) =
136.18], being highest in G allele carriers at longer dis-
tances [DBH (cid:2) TTD: F(4, 500) = 2.50]. For the inter-
mediate cue size, accuracy increased with TTD [F(2,
500) = 105.44]. For the large cue size, the only signif-
icant effect was of TTD [F(2, 500) = 18.94]. Under that
condition, both G allele carrier groups (G/G and A/G)
performed better than the A/A homozygotes (Figure 7B),
but that effect was not significant. The DBH (cid:2) TTD in-
teraction at the smallest cue size did not remain signif-
icant after Bonferroni correction. These plots suggest
2146
Journal of Cognitive Neuroscience
Volume 21, Number 11
DISCUSSION
We investigated effects of normal variation in nicotinic
and noradrenergic neurotransmission genes on atten-
tionally modulated working memory by comparing ef-
fects of two SNPs on memory performance in the same
task. We previously reported that working memory was
modulated by a noradrenergic but not by a nicotinic SNP
(Parasuraman et al., 2005), whereas the focus of visuo-
spatial attention was modulated by the same nicotinic
but not by the noradrenergic SNP (Greenwood, Fossella,
et al., 2005; Greenwood, Lambert, et al., 2005; Greenwood,
Sunderland, et al., 2005; Parasuraman et al., 2005). These
findings are consistent with separate lines of evidence
from humans showing working memory is modulated by
dopaminergic and noradrenergic systems, whereas vis-
uospatial attention is modulated by cholinergic, specif-
ically nicotinic, systems (reviewed below). Based on this,
we hypothesized that both neurotransmission systems
would modulate working memory performance when
the two systems were required to interact by manipu-
lation of working memory by visuospatial attention.
The results supported this prediction, although more
strongly for CHRNA4 than for DBH. Among people
genotyped for CHRNA4, the C/C homozygotes showed
the best performance on match trials and following large
cues on nonmatch trials. Among people genotyped for
DBH, the best memory performance was seen in DBH G
allele carriers, although that reached significance only
on match trials and on nonmatch trials in an interaction
with TTD and cue size. Moreover, a preliminary analysis
does not indicate that the effects of those SNPs inter-
acted. Although the finding that both nicotinic and
noradrenergic SNPs modulate working memory under
attentional manipulation may seem predictable in hind-
sight, it should be noted that studies using chemical and
lesion methods to manipulate cholinergic and noradren-
ergic systems have obtained variable results, with evi-
dence of both synergism (Hasselmo, Linster, Patil, Ma, &
Cekic, 1997) and antagonism between the two systems
(Ammassari-Teule, Maho, & Sara, 1991).
Consistent with our hypothesis, visuospatial attention
did alter the influence of both SNPs on working memory
performance, although the effect of variation in CHRNA4
was stronger than that of DBH. On Match trials, neither
SNP altered the effect of manipulation of the focus of
attention by cue size. On nonmatch trials, working mem-
ory accuracy increased with both cue size and TTD.
When the cue was small and the test stimulus close to
target location (hardest discrimination), accuracy was
poor in all groups. However, performance improved as
the scale of visuospatial attention increased with cue
size, but mainly in individuals with the CHRNA4 C/C
genotype (Figure 5) or who were DBH G allele carriers
(G/G and A/G, Figure 8). Thus, effect of the beneficial
genotypes was most evident when the task was hard
and the cue size optimal for nonmatch conditions.
Greenwood et al.
2147
Figure 8. Nonmatch condition, interaction of Cue size (cid:2) TTD (cid:2)
genotype in individuals genotyped for DBH rs#1108580 SNP.
Data from the shortest TTD (1.98) plotted as a function of cue size
and genotype.
effect of DBH and no interaction between the two
genotypes. As in the above analyses, there were main
effects of cue size [F(2, 340) = 5.86] and trial type [F(1,
170) = 28.35] and an interaction between them [F(2,
340) = 5.17]. The Trial type (cid:2) CHRNA4 [F(2, 170) =
3.54, h2 = .014] interaction was also significant. The
latter interaction justified separate analysis of the full
nonmatch conditions and both SNPs. This revealed
again a main effect of CHRNA4 [F(2, 170) = 2.93] but
not of DBH. All other main effects were significant [cue
size: F(2, 340) = 13.35; TTD: F(2, 340) = 97.05; their
interaction: F(4, 680) = 22.24]. There were no interac-
tions with either SNP, although the interaction of Cue
size (cid:2) TTD (cid:2) DBH was a nonsignificant trend ( p = .09).
Because the original design did not include a ‘‘no-
cue’’ condition, an analysis was carried out on a separate
sample of people who were administered the same task
with a no-cue condition added (described in Methods).
This new sample of 54 individuals (42 young, 12 old) has
not as yet been genotyped, but is included here to
demonstrate the overall benefits of the cue for working
memory performance. Under match conditions, accura-
cy was lowest in the no-cue condition (mean = 0.753)
and highest in the 5.28 (middle) cue size condition, the
latter being similar to cue size effects plotted in Figure 3.
Under nonmatch conditions, accuracy was highest in the
no-cue condition (mean = 0.89), and lowest in the small
cue (1.68) condition [mean = 0.85, F(3, 159) = 4.37]. As
in the analyses above, accuracy was lowest when test and
target were closest [F(2, 106) = 98.59]. Those factors
interacted [F(6, 318) = 12.96], such that the lowest
accuracy overall was seen under no-cue conditions when
TTD was smallest (mean = 0.791) and the highest
accuracy overall was seen following the largest cue when
TTD was greatest (mean = 0.896). Thus, precues do
benefit working memory performance. Small cues ben-
efit memory on match trials and large cues benefit
memory on nonmatch trials when test and target loca-
tions are widely separated.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
1
-
p
2
d
1
f
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
o
3
c
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
1
n
1
6
0
4
8
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
t
.
/
.
.
f
o
n
1
8
M
a
y
2
0
2
1
Although both SNPs exerted this influence on working
memory accuracy on nonmatch trials, CHRNA4 had the
stronger effect. The significant effects of CHRNA4 and
TTD at each level of cue size on nonmatch trials survived
correction for multiple comparisons. The only test of DBH
simple main effects that survived correction showed that
the G/G and A/G groups were more accurate than the A/A
group under the hardest condition when the cue size was
large (Figure 8).
There was a weak relation between RT and accuracy.
For CHRNA4, RT to the comparison stimulus was slow-
est in C/C homozygotes, which showed the highest
accuracies, although the main effect was not significant.
The difference between C/C and T/T homozygote groups
was 14 msec at the smallest cue and 31 msec at the
largest cue. However, this was due to speeding up of
the T/T group rather than slowing of the C/C group
(Figure 2A). The significant interaction is due to this
crossover of the C/T and T/T groups from the interme-
diate to the largest cue. For DBH, Figure 2B shows that
the G/G genotype group was fastest at the smallest
cue (where the highest accuracies were seen), but only
on match trials (three-way interaction of DBH (cid:2) Trial
type (cid:2) Cue size). There was no main effect of DBH
genotype on RT. Therefore, it is acknowledged that the
results for CHRNA4 are weakly consistent with a speed–
accuracy tradeoff. Nevertheless, that concept may be
most relevant to the initial processing of a stimulus, not,
as here, to processing the mental representation of a
stimulus. It is unlikely that comparison of a stimulus
with a mental representation after the 3-sec delay would
be subject to such a tradeoff. Participants are not making
a perceptual decision, they are comparing a percept
with a stored mental representation.
The present results extend our understanding of the
neurotransmission systems important in working mem-
ory. Our previous study found strong noradrenergic but
no cholinergic modulation of a working memory task
without an attentional manipulation (Parasuraman et al.,
2005), whereas the present study shows cholinergic and
weaker noradrenergic modulation of a working memory
task with an attentional manipulation. (There was a pow-
er of 0.75 to detect a main effect of DBH rs#1108580
genotype in the match analysis in the present study.)
The previous task and the present task used the same
stimuli and delays. However, the previous task required
retention of one, two, or three locations, whereas the
present task required retention of just one location. We
argue that the important factor in determining whether
the nicotinic SNP modulated working memory was the
presence of an attentional manipulation. Nevertheless, it
is possible—although somewhat harder to explain—that
the stronger manipulation of working memory load in
the previous task somehow suppressed the effect of the
nicotinic SNPs in that study.
We also confirmed that the optimal scale of visuospa-
tial attention for influencing working memory varies
with task demands, being smaller when test and target
are at the same location on match trials, but larger when
test and target appear at different locations. Our finding
that the cue size manipulation produced the best per-
formance following the medium-sized cue on match
trials is consistent with our previous findings from a
different population (Greenwood, Lambert, et al., 2005)
but stands in contrast to findings from visual search
paradigms in which the best performance follows the
smallest cue (Greenwood & Parasuraman, 1999, 2004;
Eriksen & St James, 1986). That the optimal scale of the
attentional focus is larger when encoding and retention
are required compared to when only search is required,
indicates that attentional scaling may control the form of
the encoded mental representation, perhaps by encod-
ing a portion of ground along with the figure.
It is of interest that the CHRNA4 C/C genotype asso-
ciated in our previous work with the strongest effects of
spatial cueing in visual search (Greenwood, Fossella, et al.,
2005; Greenwood, Lambert, et al., 2005; Greenwood,
Sunderland, et al., 2005) is the genotype associated with
more accurate working memory accuracy in the present
study. This finding links dependence on spatial cues with
better working memory. Bleckley, Durso, Crutchfield,
Engle, and Khanna (2003) reported that people with high
working memory capacity show more flexible allocation
of the focus of visuospatial attention compared to those
with low working memory capacity. The present findings
are consistent with that finding.
The present design cannot determine whether the
effect of visuospatial attention was exerted on encoding
or retention of location, or both. Presumably, discrimi-
nation accuracy in this task reflects the result of com-
paring the stored representation of target location with
the observed test location. That this measure is sensitive
to the size of precues presented some 3.6 sec earlier in
the trial suggests that the stored representation of the
target could have been enhanced at encoding or during
the retention interval (maintenance). Recent studies find
that both encoding and maintenance stages of spatial
working memory benefit from visuospatial attention
(Matsukura, Luck, & Vecera, 2007; Griffin & Nobre, 2003).
Our previous and present work adds to this nascent
literature by showing: (a) memory performance is affected
by the scale of visuospatial attention; (b) the optimal
scale of attention for working memory is small when
stimuli are predictably located but large when stimuli are
unpredictably located; (c) an optimally sized attentional
scale at encoding improves later memory for location;
(d) normal variation in neurotransmission genes influ-
ences the strength of the effect of visuospatial attention
on working memory accuracy, perhaps by heightening
the perceptual benefit accruing to an attended target
when the focus of attention is optimally scaled. The
present work also emphasizes the particular importance
of cholinergic neurotransmission for attention-based con-
trol of the encoded representation in working memory.
2148
Journal of Cognitive Neuroscience
Volume 21, Number 11
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
1
-
p
2
d
1
f
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
o
3
c
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
1
n
1
6
0
4
8
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
t
.
.
.
f
/
o
n
1
8
M
a
y
2
0
2
1
We can consider our present findings in the context of
pharmacological evidence that acetylcholine (ACh) and
NE can work synergistically (Gu, 2002; Hasselmo et al.,
1997), showing both neurotransmission systems are in-
volved when tasks require deployment of visuospatial
attention as well as memory. We review these different
strands of evidence and argue (a) that normal genetic
variation in neurotransmission genes has specific behav-
ioral effects and (b) that the relative importance of cho-
linergic or noradrenergic efficiency on a given task varies
with the relative demands made by that task on pro-
cesses of attention and memory.
There is considerable evidence bearing on the neuro-
transmission systems involved in processes of working
memory and visuospatial attention. Work in rodents in-
dicates that nicotinic agents influence both memory
(Levin & Simon, 1998) and sustained attention (Rezvani,
Bushnell, & Levin, 2002). Levin, Kaplan, and Boardman
(1997) argue from their work in rodents that nicotine
boosts the performance of working memory specifically.
However, other investigators have argued that in pri-
mates the basal forebrain cholinergic system influences
visuospatial attention but not memory (Everitt & Robbins,
1997; Voytko et al., 1994). Consistent with that view,
there is human evidence that noradrenergic agents mod-
ulate working memory, whereas cholinergic agents mod-
ulate visuospatial attention. DA D1 receptor availability
has been found to be correlated with working memory
performance in humans and monkeys (Abi-Dargham et al.,
2002; Castner et al., 2000) and alpha-2A adrenorecep-
tor agonists modulate both working memory perfor-
mance and blood flow in dorsolateral PFC in monkeys
(Franowicz & Arnsten, 2002; Avery et al., 2000; Mao,
Arnsten, & Li, 1999). Suggesting specificity, a noradren-
ergic agonist has been shown to modulate an alerting
effect but not an attentional cue validity effect in mon-
keys (Witte & Marrocco, 1997).
In primates,
including humans, cholinergic effects
appear to be exerted selectively on visuospatial attention
and not on memory. First, there is evidence of nicotinic
cholinergic mediation of attention. Administration of nic-
otine to both humans and monkeys slowed responses
on invalidly cued trials but not on validly cued trials
( Witte et al., 1997). In human smokers, effects of cue
validity increased with days of abstinence and were
inversely related to salivary levels of the nicotine metab-
olite cotinine (Shirtcliff & Marrocco, 2003). Thiel et al.
(2005) have also found selective effects of nicotine on
discrimination following invalid cues in nonsmokers
which were accompanied by reduced fMRI activation
in the intraparietal sulcus and precuneus. Secondly, cho-
linergic effects appear to be exerted selectively on visuo-
spatial attention. Neurotoxic inactivation of the nbM in
the monkey basal
forebrain, the major subcortical
source of cortical ACh, disrupted performance in a cued
visuospatial attention task but had no effect on several
tasks of memory (Voytko, 1996; Voytko et al., 1994). More-
over, the attentional deficit observed was qualitatively
similar to one observed in patients with Alzheimer’s Dis-
ease (Parasuraman, Greenwood, Haxby, & Grady, 1992),
known to have marked cortical cholinergic depletion
due to progressive degeneration of the nbM. Nicotine
patches have been found to improve visuospatial atten-
tion, but not memory, in older people with memory
impairment (White & Levin, 2004) and in AD patients
(Sahakian, Jones, Levy, Gray, & Warburton, 1989). Thus,
in humans and monkeys, there is evidence that working
memory is selectively modulated by noradrenergic and
dopaminergic systems, whereas visuospatial attention is
selectively modulated by cholinergic systems.
Moreover, there is increasing evidence that choliner-
gic influences on working memory are exerted mainly
under attentional load—that is, when the two systems
interact. In a working memory task, Furey et al. (2000)
found that during a 3-sec encoding period, the choliner-
gic agonist physostigmine enhanced the extrastriate re-
sponse to faces and decreased the response to control
stimuli. The differential effect of physostigmine was not
seen during memory maintenance. The authors argued
that the enhanced cholinergic activity improved working
memory performance by selectively heightening percep-
tual processing of the relevant stimuli. In a subsequent
study, Furey et al. presented overlapping stimuli of faces
and houses and used a cue to direct the focus of visuo-
spatial attention in advance to one of the two overlap-
ping stimuli. Physostigmine speeded RT on trials after
the shift in category, but not on the trial with the shift
in category. Scopolamine had the opposite effect. Thus,
enhancement or inhibition of cholinergic activity influ-
enced ability to maintain selective attention on the cued
category of target, but did not alter the ability to shift
category (Furey, Pietrini, Haxby, & Drevets, 2008). This
suggests that the cholinergic system has a large role in
selective attention. Consistent with the work of Furey
et al., scopolamine has been reported to cause deficien-
cies in delayed match-to-sample performance, but only
when heightened attention was required at the longest
delay (Robbins et al., 1997).
Considered as a whole, this work is consistent with an
interpretation that the role of cholinergic systems in
working memory is attentional. To test this directly,
Robbins et al. developed a task which allowed separate
assessment of attention and working memory. Scopol-
amine infused into PFC was found to impair memory
but not attention in intact rats. Similarly, animals with
selective Ig-G saporin lesions of the nucleus basalis
magnocellularis (no scopolamine administered) showed
impaired memory under high attentional load but not
under low attentional load (Chudasama, Dalley, Nathwani,
Bouger, & Robbins, 2004). These data indicate a role for
basal cholinergic input to PFC in memory function when
attention is required. Providing a possible explanation at
the cellular level, a recent study showed that nicotinic
receptors affect processes of long-term potentiation in
Greenwood et al.
2149
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
1
-
p
2
d
1
f
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
o
3
c
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
1
n
1
6
0
4
8
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
t
.
.
/
.
f
o
n
1
8
M
a
y
2
0
2
1
PFC (Couey et al., 2007). Consistent with that finding
are the increasing indications that human PFC plays
a role in attentional modulation of working memory
(Postle, 2006; Curtis & D’Esposito, 2003). The evidence
of nicotine-driven enhancement of signal-to-noise ratio
during PFC information processing (Couey et al., 2007)
is consistent with the present finding of nicotinic recep-
tor modulation of working memory under attentional
load in humans. Nonetheless, further work will be needed
to disentangle the cholinergic and noradrenergic influ-
ences on the interactions between visuospatial attention
and working memory.
What is the physiological basis for the apparent
influence of cholinergic and noradrenergic systems on
working memory when attention is manipulated? In
several studies, Hasselmo et al. (1997) has shown that
when perfused directly into cortex, the effects of ACh
and NE combine in an additive manner to produce dose-
dependent suppression of synaptic potentials. In contrast
to this synergism in ACh–NE physiological interactions,
Dyon-Laurent, Herve, and Sara (1994), Ammassari-Teule
et al. (1991), and Sara (1989) have reported antagonistic
interactions in a set of learning and memory experiments.
Learning and memory deficits, which developed subse-
quent to cholinergic lesions, were reduced by the admin-
istration of a noradrenergic alpha-2 agonist. From this and
other evidence, Yu and Dayan (2005) have argued that
ACh is involved in ‘‘expected uncertainty,’’ whereas NE is
involved in ‘‘unexpected uncertainty.’’ They interpret
evidence that cue validity effects vary inversely with level
of nicotine or ACh in attentional cueing tasks (Phillips,
McAlonan, Robb, & Brown, 2000; Witte et al., 1997; Voytko
et al., 1994) as reflecting cholinergic suppression of cue-
ing effects. On the other hand, they interpret findings
that NE modulates performance on tasks with unpredict-
able changes in cue–target relationship (Devauges & Sara,
1990) as reflecting noradrenergic-induced change in cue–
target associations. The model advanced by Yu and Dayan
would predict a role for cholinergic neuromodulation in
any task using attentional cueing, consistent with the pres-
ent findings.
These results can also be viewed in the context of the
role of visuospatial attention in working memory. Sev-
eral theorists have argued that the focus of visuospatial
attention is integral to working memory (Cowan, 2001;
Awh et al., 1998). Elsewhere, we have reported that dy-
namic adjustments of the scale of visuospatial attention
in response to task demands (Greenwood & Parasuraman,
2004) have a role in controlling the encoded represen-
tation in working memory (Greenwood, Lambert, et al.,
2005). Using two independent samples, we have shown
that the scale of visuospatial attention deployed around
a target affects memory for location of that target
stimulus. The present study shows that variation in the
gene controlling a subunit of the most common nico-
tinic receptor—the alpha4/beta 2 nicotinic ACh receptor
(Flores, DeCamp, Kilo, Rogers, & Hargreaves, 1996)—
modulates that effect of attention on memory. This indi-
cates the importance of cholinergic neurotransmission
for attention-based control of the encoded representa-
tion in working memory. As such, the present results are
consistent with theories that argue for a relation between
visuospatial attention and working memory (Cowan,
2001; Engle et al., 1999). The present results go beyond
those theories by showing that cholinergic mediation of
working memory is particularly important when working
memory is explicitly modulated by the focus of visuo-
spatial attention. This indicates that visuospatial atten-
tion and working memory are separate systems which
can interact.
In light of the concern about false positives in candi-
date gene studies, it can be noted that the effect size we
report for CHRNA4 C/T of .25 is close to the effect size of
.29 recently reported in a meta-analysis of the effect of
the well-studied COMT val158met polymorphism on the
Wisconsin Card Sort Task (Barnett et al., 2007). We have
now observed effects of the CHRNA4 rs#1044396 SNP
on visuospatial attention in two large and independent
samples (Espeseth et al., 2006; Greenwood, Fossella,
et al., 2005; Parasuraman et al., 2005). Effects of this SNP
have been reported by other investigators on fMRI mea-
sures of an attentional manipulation (Winterer et al.,
2007). Auditory and visual ERPs likewise reflect polymor-
phic variation in this SNP (Espeseth, Endestad, Rootwelt,
& Reinvang, 2007).
There are some limitations of the present study. The
sample size, although large for the psychological litera-
ture, allowed only a preliminary full comparison of the
two SNPs in one analysis and did not allow analysis of
combined age and genotype effects. Also, although our
interpretations are supported in the human literature,
we acknowledge that our findings cannot completely
rule out cholinergic modulation of working memory in
the absence of attentional manipulations. Although the
present findings will need to be replicated with larger
samples, they are notable for showing that the effect of
visuospatial attention on working memory is modulated
by normal genetic variation in separate neurotransmis-
sion systems.
Acknowledgments
This work was supported by NIA grant AG19653 to R. P. and
the Virginia Center on Aging grant ARDRAF 06-2 to P. M. G. We
thank Ryan McGarry and Ruchi Kapani for assistance with data
collection and management.
Reprint requests should be sent to P. M. Greenwood, George
Mason University, David King Hall 2060, MSN 3F5, 4400 Uni-
versity Drive, Fairfax, VA 22030-4444, or via e-mail: Pgreenw1@
gmu.edu.
REFERENCES
Abi-Dargham, A., Mawlawi, O., Lombardo, I., Gil, R., Martinez,
D., Huang, Y., et al. (2002). Prefrontal dopamine D1
2150
Journal of Cognitive Neuroscience
Volume 21, Number 11
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
1
-
p
2
d
1
f
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
o
3
c
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
1
n
1
6
0
4
8
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
f
t
/
.
.
o
n
1
8
M
a
y
2
0
2
1
receptors and working memory in schizophrenia. Journal of
Neuroscience, 22, 3708–3719.
Ammassari-Teule, M., Maho, C., & Sara, S. J. (1991). Clonidine
reverses spatial learning deficits and reinstates theta
frequencies in rats with partial fornix section. Behavioural
Brain Research, 45, 1–8.
Avery, R. A., Franowicz, J. S., Studholme, C., van Dyck, C. H., &
Arnsten, A. F. (2000). The alpha-2A-adrenoceptor agonist,
guanfacine, increases regional cerebral blood flow in
dorsolateral prefrontal cortex of monkeys performing a
spatial working memory task. Neuropsychopharmacology,
23, 240–249.
Awh, E., & Jonides, J. (1998). Spatial working memory and
spatial selective attention. In R. Parasuraman (Ed.), The
attentive brain (pp. 353–380). Cambridge, MA: MIT Press.
Awh, E., Jonides, J., & Reuter-Lorenz, P. A. (1998). Rehearsal in
spatial working memory. Journal of Experimental
Psychology: Human Perception and Performance, 24,
780–790.
Awh, E., Vogel, E. K., & Oh, S. H. (2006). Interactions between
attention and working memory. Neuroscience, 139,
201–208.
Baddeley, A. (1992). Working memory. Science, 255, 556–559.
Barnett, J. H., Jones, P. B., Robbins, T. W., & Muller, U.
(2007). Effects of the catechol-O-methyltransferase
Val158Met polymorphism on executive function: A
meta-analysis of the Wisconsin Card Sort Test in
schizophrenia and healthy controls. Molecular Psychiatry,
12, 502–509.
Bleckley, M. K., Durso, F. T., Crutchfield, J. M., Engle, R. W.,
& Khanna, M. M. (2003). Individual differences in working
memory capacity predict visual attention allocation.
Psychonomic Bulletin & Review, 10, 884–889.
Castner, S. A., Williams, G. V., & Goldman-Rakic, P. S. (2000).
Reversal of antipsychotic-induced working memory deficits
by short-term dopamine D1 receptor stimulation. Science,
287, 2020–2022.
Chudasama, Y., Dalley, J. W., Nathwani, F., Bouger, P., &
Robbins, T. W. (2004). Cholinergic modulation of visual
attention and working memory: Dissociable effects of
basal forebrain 192-IgG-saporin lesions and intraprefrontal
infusions of scopolamine. Learning and Memory, 11,
78–86.
Cohen, J. (1988). Statistical power analysis for the behavioral
sciences (2nd ed.). Hillsdale, NJ: Erlbaum.
Conway, A. R., Cowan, N., & Bunting, M. F. (2001). The cocktail
party phenomenon revisited: The importance of working
memory capacity. Psychonomic Bulletin & Review, 8,
331–335.
Couey, J. J., Meredith, R. M., Spijker, S., Poorthuis, R. B.,
Smit, A. B., Brussaard, A. B., et al. (2007). Distributed
network actions by nicotine increase the threshold for
spike-timing-dependent plasticity in prefrontal cortex.
Neuron, 54, 73–87.
Cowan, N. (1988). Evolving conceptions of memory storage,
selective attention, and their mutual constraints within
the human information-processing system. Psychological
Bulletin, 104, 163–191.
Cowan, N. (2001). The magical number 4 in short-term
memory: A reconsideration of mental storage capacity.
Behavioral and Brain Sciences, 24, 87–114; discussion
114–185.
Cubells, J. F., van Kammen, D. P., Kelley, M. E., Anderson,
G. M., O’Connor, D. T., Price, L. H., et al. (1998).
Dopamine beta-hydroxylase: Two polymorphisms in
linkage disequilibrium at the structural gene DBH
associate with biochemical phenotypic variation. Human
Genetics, 102, 533–540.
Curtis, C. E., & D’Esposito, M. (2003). Persistent activity in the
prefrontal cortex during working memory. Trends in
Cognitive Sciences, 7, 415–423.
Dani, J. A., & Bertrand, D. (2007). Nicotinic acetylcholine
receptors and nicotinic cholinergic mechanisms of the
central nervous system. Annual Review of Pharmacology
and Toxicology, 47, 699–729.
Devauges, V., & Sara, S. J. (1990). Activation of the
noradrenergic system facilitates an attentional shift in
the rat. Behavioural Brain Research, 39, 19–28.
Dyon-Laurent, C., Herve, A., & Sara, S. J. (1994).
Noradrenergic hyperactivity in hippocampus after
partial denervation: Pharmacological, behavioral, and
electrophysiological studies. Experimental Brain
Research, 99, 259–266.
Engle, R. W., Kane, M. J., & Tuholski, S. W. (Eds.) (1999).
Individual differences in working memory capacity and
what they tell us about controlled attention, general fluid
intelligence, and functions of the prefrontal cortex.
Cambridge: Cambridge University Press.
Eriksen, C. W., & St James, J. D. (1986). Visual attention within
and around the field of focal attention: A zoom lens model.
Perception & Psychophysics, 40, 225–240.
Espeseth, T., Endestad, T., Rootwelt, H., & Reinvang, I. (2007).
Nicotine receptor gene CHRNA4 modulates early
event-related potentials in auditory and visual oddball
target detection tasks. Neuroscience, 147, 974–985.
Espeseth, T., Greenwood, P. M., Reinvang, I., Fjell, A. M.,
Walhovd, K. B., Westlye, L. T., et al. (2006). Interactive effects
of APOE and CHRNA4 on attention and white matter volume
in healthy middle-aged and older adults. Cognitive, Affective,
& Behavioral Neuroscience, 6, 31–43.
Everitt, B. J., & Robbins, T. W. (1997). Central cholinergic
systems and cognition. Annual Review of Psychology, 48,
649–684.
Feng, Y., Niu, T., Xing, H., Xu, X., Chen, C., Peng, S., et al.
(2004). A common haplotype of the nicotine acetylcholine
receptor alpha 4 subunit gene is associated with vulnerability
to nicotine addiction in men. American Journal of Human
Genetics, 75, 112–121.
Flores, C. M., DeCamp, R. M., Kilo, S., Rogers, S. W., &
Hargreaves, K. M. (1996). Neuronal nicotinic receptor
expression in sensory neurons of the rat trigeminal ganglion:
Demonstration of alpha3beta4, a novel subtype in the
mammalian nervous system. Journal of Neuroscience, 16,
7892–7901.
Flores, C. M., Rogers, S. W., Pabreza, L. A., Wolfe, B. B., &
Kellar, K. J. (1992). A subtype of nicotinic cholinergic
receptor in rat brain is composed of alpha 4 and beta 2
subunits and is up-regulated by chronic nicotine treatment.
Molecular Pharmacology, 41, 31–37.
Folstein, M. F., Folstein, S. E., & McHugh, P. R. (1975).
‘‘Mini-mental state’’—A practical method for grading the
cognitive state of patients for the clinician. Journal of
Current Psychiatric Research, 12, 189–198.
Fossella, J., Sommer, T., Fan, J., Wu, Y., Swanson, J. M., Pfaff,
D. W., et al. (2002). Assessing the molecular genetics of
attention networks. BMC Neuroscience, 3, 14.
Fossella, J. A., & Casey, B. J. (2006). Genes, brain, and behavior:
Bridging disciplines. Cognitive, Affective & Behavioral
Neuroscience, 6, 1–8.
Foxe, J. J., & Simpson, G. V. (2002). Flow of activation from V1
to frontal cortex in humans. A framework for defining
‘‘early’’ visual processing. Experimental Brain Research,
142, 139–150.
Franowicz, J. S., & Arnsten, A. F. (2002). Actions of alpha-2
noradrenergic agonists on spatial working memory and
blood pressure in rhesus monkeys appear to be mediated by
Greenwood et al.
2151
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
1
-
p
2
d
1
f
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
o
3
c
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
1
n
1
6
0
4
8
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
f
/
.
.
.
t
o
n
1
8
M
a
y
2
0
2
1
the same receptor subtype. Psychopharmacology (Berlin),
162, 304–312.
Furey, M. L., Pietrini, P., & Haxby, J. V. (2000). Cholinergic
enhancement and increased selectivity of perceptual
processing during working memory. Science, 290,
2315–2319.
Furey, M. L., Pietrini, P., Haxby, J. V., & Drevets, W. C.
(2008). Selective effects of cholinergic modulation
on task performance during selective attention.
Neuropsychopharmacology, 33, 913–923.
Goldberg, T. E., & Weinberger, D. R. (2004). Genes and the
parsing of cognitive processes. Trends in Cognitive Sciences,
8, 325–335.
Greenwood, P., Kumar, R., Sundararajan, R., Fryxell, K. J.,
Lin, M., & Parasuraman, R. (2006). Effects of attention
on working memory are modulated by a nicotinic
SNP (CHRNA4) but not a noradrenergic SNP (DBH).
Paper presented at the Society for Neuroscience, Atlanta,
Georgia.
Greenwood, P. M., Fossella, J. A., & Parasuraman, R. (2005).
Specificity of the effect of a nicotinic receptor polymorphism
on individual differences in visuospatial attention. Journal of
Cognitive Neuroscience, 17, 1611–1620.
Greenwood, P. M., Lambert, C., Sunderland, T., &
Parasuraman, R. (2005). Effects of apolipoprotein E
genotype on spatial attention, working memory, and their
interaction in healthy, middle-aged adults: Results from the
National Institute of Mental Health’s BIOCARD study.
Neuropsychology, 19, 199–211.
Greenwood, P. M., & Parasuraman, R. (1999). Scale of
attentional focus in visual search. Perception &
Psychophysics, 61, 837–859.
Greenwood, P. M., & Parasuraman, R. (2003). Normal genetic
variation, cognition, and aging. Behavioral and Cognitive
Neuroscience Reviews, 2, 278–306.
Greenwood, P. M., & Parasuraman, R. (2004). The scaling of
spatial attention in visual search and its modification in
healthy aging. Perception & Psychophysics, 66, 3–22.
Greenwood, P. M., Sunderland, T., Putnam, K., Levy, J., &
Parasuraman, R. (2005). Scaling of visuospatial attention
undergoes differential longitudinal change as a function of
APOE genotype prior to old age: Results from the NIMH
BIOCARD study. Neuropsychology, 19, 830–840.
Griffin, I. C., & Nobre, A. C. (2003). Orienting attention to
locations in internal representations. Journal of Cognitive
Neuroscience, 15, 1176–1194.
Gu, Q. (2002). Neuromodulatory transmitter systems in the
cortex and their role in cortical plasticity. Neuroscience, 111,
815–835.
Hare, M. P., & Palumbi, S. R. (1999). The accuracy of
heterozygous base calling from diploid sequence and
resolution of haplotypes using allele-specific sequencing.
Molecular Ecology, 8, 1750–1752.
characterization, pharmacological specification, and
anatomic localization. Psychopharmacology (Berlin), 184,
523–539.
Levin, E. D., & Simon, B. B. (1998). Nicotinic acetylcholine
involvement in cognitive function in animals.
Psychopharmacology (Berlin), 138, 217–230.
Mao, Z. M., Arnsten, A. F., & Li, B. M. (1999). Local infusion of
an alpha-1 adrenergic agonist into the prefrontal cortex
impairs spatial working memory performance in monkeys.
Biological Psychiatry, 46, 1259–1265.
Matsukura, M., Luck, S. J., & Vecera, S. P. (2007). Attention
effects during visual short-term memory maintenance:
Protection or prioritization? Perception & Psychophysics, 69,
1422–1434.
Noesselt, T., Hillyard, S. A., Woldorff, M. G., Schoenfeld, A.,
Hagner, T., Jancke, L., et al. (2002). Delayed striate
cortical activation during spatial attention. Neuron, 35,
575–587.
Parasuraman, R., & Greenwood, P. M. (2004). Molecular
genetics of visuospatial attention and working memory.
In M. I. Posner (Ed.), Cognitive neuroscience of attention
(pp. 245–259). New York: Guilford.
Parasuraman, R., Greenwood, P. M., Haxby, J. V., & Grady, C. L.
(1992). Visuospatial attention in dementia of the Alzheimer
type. Brain, 115, 711–733.
Parasuraman, R., Greenwood, P. M., Kumar, R., & Fossella, J.
(2005). Beyond heritability: Neurotransmitter genes
differentially modulate visuospatial attention and working
memory. Psychological Science, 16, 200–207.
Phillips, J. M., McAlonan, K., Robb, W. G., & Brown, V. J. (2000).
Cholinergic neurotransmission influences covert orientation
of visuospatial attention in the rat. Psychopharmacology
(Berlin), 150, 112–116.
Posner, M. I., Rothbart, M. K., & Sheese, B. E. (2007). Attention
genes. Developmental Science, 10, 24–29.
Postle, B. R. (2006). Working memory as an emergent
property of the mind and brain. Neuroscience, 139,
23–38.
Ranganath, C. (2006). Working memory for visual objects:
Complementary roles of inferior temporal, medial
temporal, and prefrontal cortex. Neuroscience, 139,
277–289.
Rezvani, A. H., Bushnell, P. J., & Levin, E. D. (2002). Effects of
nicotine and mecamylamine on choice accuracy in an
operant visual signal detection task in female rats.
Psychopharmacology (Berlin), 164, 369–375.
Robbins, T. W., Everitt, B. J., Marston, H. M., Wilkinson, J.,
Jones, G. H., & Page, K. J. (1989). Comparative effects
of ibotenic acid- and quisqualic acid-induced lesions
of the substantia innominata on attentional function
in the rat: Further implications for the role of the
cholinergic neurons of the nucleus basalis in cognitive
processes. Behavioural Brain Research, 35, 221–240.
Hasselmo, M. E., Linster, C., Patil, M., Ma, D., & Cekic, M.
Robbins, T. W., Semple, J., Kumar, R., Truman, M. I., Shorter, J.,
(1997). Noradrenergic suppression of synaptic transmission
may influence cortical signal-to-noise ratio. Journal of
Neurophysiology, 77, 3326–3339.
Kent, L., Middle, F., Hawi, Z., Fitzgerald, M., Gill, M.,
Feehan, C., et al. (2001). Nicotinic acetylcholine
receptor alpha4 subunit gene polymorphism and
attention deficit hyperactivity disorder. Psychiatric Genetics,
11, 37–40.
Levin, E. D., Kaplan, S., & Boardman, A. (1997). Acute nicotine
interactions with nicotinic and muscarinic antagonists:
Working and reference memory effects in the 16-arm
radial maze. Behavioural Pharmacology, 8, 236–242.
Levin, E. D., McClernon, F. J., & Rezvani, A. H. (2006).
Nicotinic effects on cognitive function: Behavioral
Ferraro, A., et al. (1997). Effects of scopolamine on
delayed-matching-to-sample and paired associates tests
of visual memory and learning in human subjects:
Comparison with diazepam and implications of dementia.
Psychopharmacology (Berlin), 134, 95–106.
Sahakian, B., Jones, G., Levy, R., Gray, J., & Warburton, D.
(1989). The effects of nicotine on attention, information
processing, and short-term memory in patients with
dementia of the Alzheimer type. British Journal of
Psychiatry, 154, 797–800.
Sara, S. J. (1989). Noradrenergic–cholinergic interaction: Its
possible role in memory dysfunction associated with senile
dementia. Archives of Gerontology and Geriatrics:
Supplement, 1, 99–108.
2152
Journal of Cognitive Neuroscience
Volume 21, Number 11
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
1
-
p
2
d
1
f
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
o
3
c
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
1
n
1
6
0
4
8
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
/
f
.
t
.
.
o
n
1
8
M
a
y
2
0
2
1
Sarter, M., & Bruno, J. P. (2004). Developmental origins of the
age-related decline in cortical cholinergic function and
associated cognitive abilities. Neurobiology of Aging, 25,
1127–1139.
Shirtcliff, E. A., & Marrocco, R. T. (2003). Salivary cotinine levels
in human tobacco smokers predict the attentional validity
effect size during smoking abstinence. Psychopharmacology
(Berlin), 166, 11–18.
Steinlein, O. K., Deckert, J., Nothen, M. M., Franke, P., Maier,
W., Beckmann, H., et al. (1997). Neuronal nicotinic
acetylcholine receptor alpha 4 subunit (CHRNA4) and panic
disorder: An association study. American Journal of
Medical Genetics, 74, 199–201.
Super, H., Spekreijse, H., & Lamme, V. A. (2001). A neural
correlate of working memory in the monkey primary visual
cortex. Science, 293, 120–124.
Thiel, C. M., Zilles, K., & Fink, G. R. (2005). Nicotine modulates
reorienting of visuospatial attention and neural activity in
human parietal cortex. Neuropsychopharmacology, 30,
810–820.
Vogel, E. K., & Machizawa, M. G. (2004). Neural activity
predicts individual differences in visual working memory
capacity. Nature, 428, 748–751.
Vogel, E. K., McCollough, A. W., & Machizawa, M. G.
(2005). Neural measures reveal individual differences in
controlling access to working memory. Nature, 438, 500–503.
Voytko, M. L. (1996). Cognitive functions of the basal forebrain
cholinergic system in monkeys: Memory or attention?
Behavioural Brain Research, 75, 13–25.
single-tube PCR with Tm-shift primers. BioTechniques, 39,
885–893.
Wechsler, D. (1981). Wechsler Adult Intelligence
Scale—revised: Manual. New York: Psychological
Corporation.
Wechsler, D. (1987). Wechsler Memory Scale—revised:
Manual. New York: Psychological Corporation.
White, H. K., & Levin, E. D. (2004). Chronic transdermal
nicotine patch treatment effects on cognitive performance in
age-associated memory impairment. Psychopharmacology
(Berlin), 171, 465–471.
Winterer, G., Musso, F., Konrad, A., Vucurevic, G., Stoeter, P.,
Sander, T., et al. (2007). Association of attentional network
function with exon 5 variations of the CHRNA4 gene.
Human Molecular Genetics, 16, 2165–2174.
Witte, E. A., Davidson, M. C., & Marrocco, R. T. (1997).
Effects of altering brain cholinergic activity on covert
orienting of attention: Comparison of monkey and
human performance. Psychopharmacology (Berlin), 132,
324–334.
Witte, E. A., & Marrocco, R. T. (1997). Alteration of brain
noradrenergic activity in rhesus monkeys affects the alerting
component of covert orienting. Psychopharmacology
(Berlin), 132, 315–323.
Wood, J. G., Joyce, P. R., Miller, A. L., Mulder, R. T., &
Kennedy, M. A. (2002). A polymorphism in the dopamine
beta-hydroxylase gene is associated with ‘‘paranoid
ideation’’ in patients with major depression. Biological
Psychiatry, 51, 365–369.
Voytko, M. L., Olton, D. S., Richardson, R. T., Gorman, L. K.,
Yago, E., Duarte, A., Wong, T., Barcelo, F., & Knight, R. T.
Tobin, J. R., & Price, D. L. (1994). Basal forebrain lesions in
monkeys disrupt attention but not learning and memory.
Journal of Neuroscience, 14, 167–186.
(2004). Temporal kinetics of prefrontal modulation of the
extrastriate cortex during visual attention. Cognitive,
Affective & Behavioral Neuroscience, 4, 609–617.
Wang, J., Chuang, K., Ahluwalia, M., Patel, S., Umblas, N.,
Yu, A. J., & Dayan, P. (2005). Uncertainty, neuromodulation,
Mirel, D., et al. (2005). High-throughput SNP genotyping by
and attention. Neuron, 46, 681–692.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
t
f
r
p
o
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
h
.
m
a
i
r
e
.
d
c
u
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
e
1
1
-
p
2
d
1
f
3
/
9
2
1
1
9
/
3
1
7
1
5
/
5
2
4
1
o
3
c
9
n
/
2
1
0
7
0
5
8
9
8
2
4
1
1
6
6
/
4
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
1
n
1
6
0
4
8
.
S
p
e
d
p
f
e
m
b
y
b
e
r
g
u
2
0
e
2
s
3
t
/
j
.
/
.
.
t
f
o
n
1
8
M
a
y
2
0
2
1
Greenwood et al.
2153