Biological Control
Variously Materialized:
Modeling,
Experimentation
and Exploration in
Multiple Media
Tarja Knuuttila
维也纳大学
Andrea Loettgers
维也纳大学
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
This paper examines two parallel discussions of scientific modeling which have in-
voked experimentation in addressing the role of models in scientific inquiry. One side
discusses the experimental character of models, whereas the other focuses on their ex-
ploratory uses. Although both relate modeling to experimentation, they do so differ-
ently. The former has considered the similarities and differences between models and
实验, addressing, 尤其, the epistemic value of materiality. By con-
特拉斯特, the focus on exploratory modeling has highlighted the various kinds of explor-
atory functions of models in the early stages of inquiry. These two perspectives on
modeling are discussed through a case study in the field of synthetic biology. 这
research practice in question explores biological control by making use of an ensemble
of different epistemic means: mathematical models and simulations, synthetic genetic
circuits and intracellular measuring devices, and finally electronic circuits. We argue
that the study of exploratory modeling should trace the ways different epistemic
方法, in different materialities, are being combined over time. 最后, the epistemic
status of such novel investigative objects as synthetic genetic circuits is evaluated, 和
the conclusion that they can function as both experiments and models.
The authors wish to thank Michael Elowitz, Jordi García-Ojalvo, Axel Gelfert and an anon-
ymous referee for this special issue. This project received funding from the European Re-
search Council (ERC) under the European Union’s Horizon 2020 research and innovation
programme (grant agreement No 818772).
科学观点 2021, 卷. 29, 不. 4
© 2021 由麻省理工学院.
根据知识共享署名发布 4.0 国际的 (抄送 4.0) 执照
https://doi.org/10.1162/posc_a_00379
468
科学观点
469
介绍
1.
The philosophical discussion of scientific models is undergoing a distin-
guishable turn towards the practices of modeling, and away from issues rep-
resentation. Two recent discussions, 尤其, have attempted to flesh
out the idea that models are objects with which we do something in our
epistemic activities. While the one discussion has considered the experimen-
tal character of models (例如, Cartwright 1999; Guala 2002; Mäki 2005;
摩根 2003, 2005), the other one has addressed their exploratory functions
(例如, Gelfert 2016, 2018; Fisher 2017; Massimi 2018). Both discussions
relate modeling to experimentation, but they do so differently. The discus-
sion of the experimental character of models likens them more directly to
实验, highlighting those aspects of modeling and simulation
that come close to experimentation. 因此, a new set of questions has
出现了, concerning the ways modeling and experimentation resemble, 或者
differ from, 彼此, and whether those differences are due to the infer-
ences they license, or the different role materiality plays in experimentation
vis-à-vis modeling.
In contrast to viewing models as a kind of experiment, the interest in explor-
atory modeling does not directly build on the analogy between modeling and
实验. It rather takes the discussion of exploratory experimentation as
its starting point in focusing on the exploratory modes of reasoning character-
istic of early stages of inquiry in which “an established prior body of knowledge”
“cannot be assumed, or is itself at issue” (Gelfert 2018). Experimentation in such
situations takes up the role of conceptual and theoretical exploration (Steinle
1997), as well that of stabilization of phenomena (Burian 1997). 然而,
when applied to modeling, the idea of exploration gets new dimensions due
to the fact that much of modeling is of a theoretical nature at the outset, 和
in many fields the theoretical activity consists predominantly of modeling.
Given these two parallel discussions of modeling, both inspired by the
philosophical literature on experimentation, it seems interesting to discuss
them side-by-side, in order to better understand the experimentable and
exploratory nature of modeling. The two discussions seem to place emphasis
on different dimensions of modeling. The discussion of the experimental
character of models concentrates on models themselves as workable, exper-
imental objects, addressing in particular the epistemic value of materiality.
The focus on exploratory modeling, 反过来, highlights the different uses of
models in explorative activities. In order to better understand such exploratory
流程, we analyze (parts of) a research trajectory of one leading research
group within the field of synthetic biology. Such a processual approach
allows us to more fully understand exploratory modeling. Another impor-
tant aspect of exploratory modeling that we wish to highlight is the use of
various kinds of epistemic means—different kinds of models, 实验
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
470
Biological Control Materialized
and measurements—that any genuinely exploratory process typically
involves. We submit that is crucial not just to study single models but also
to focus on multiple models, in multiple materialities.
The exploratory process we will examine focuses on biological control,
and on how scientists combined different models of various materialities with
彼此, and with other epistemic means. The ensemble of different epistemic
means used to explore the role of noise in biological organization consisted of
mathematical models and simulations, synthetic genetic circuits and measuring
设备, and finally electronic circuits. The status of synthetic and electronic cir-
cuits in this fabric is especially interesting. While they can be considered as ex-
periments due to their material make-up, they can also be conceptualized as
theoretical explorations. Especially synthetic genetic circuits lie squarely be-
tween modeling and experimentation as they are constructed of the “same stuff”
as the studied biological systems, yet are carefully designed making use of math-
ematical models as blueprints. 或者, the role of electronic circuits in this
exploratory process seems equally intriguing. Why did scientists, already armed
with mathematical models, simulations, and synthetic genetic circuits built
from genes and proteins, start to construct electronic circuits to study biological
控制?
Our case on biological control is based on a laboratory study of the Elowitz Lab
at the California Institute of Technology, including its collaborations, 尤其
with physicist Jordi García-Ojalvo at Universitat Pompeu Fabra and his co-
工人. Michael Elowitz is the co-author of the Repressilator, which is one
of the first synthetic genetic circuits to have been published, as well as probably
the most famous one, due to its pioneering nature (Elowitz and Leibler 2000).
There is already ample philosophical literature on the Repressilator (例如,
Knuuttila and Loettgers 2011, 2013A, 2013乙), but the exploratory process it
gave impetus to, has neither been reported, nor studied. We will show how
the process of exploratory modeling has established more conclusive evidence,
now at the molecular level, for the role of stochastic fluctuations in biology. 什么
is more, this evidence amounted to a redefinition of the target system: 尽管
human-made control systems are built to minimize noise, the scientists were able
to make a convincing case that noise is an essential part of biological control.
We will begin with an overview on the philosophical discussion of the
experimental character of models, turning then to exploratory modeling.
After that, we will consider the exploratory research program on biological
control and noise in which the Repressilator model has played a crucial
role in its many incarnations.
2. Models as Experiments?
Once models are couched as experimentable objects, it has proven difficult,
at least on a general philosophical level, to clearly distinguish between modeling
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
科学观点
471
and experimentation. Although it seems intuitive to think that there are cru-
cial differences between modeling and experimentation in terms of their re-
spective targets, epistemic results, or materiality, several philosophers have
presented counterarguments that largely bring modeling/simulation and ex-
perimentation together. The recent philosophical discussion has pointed out
two ways in which they resemble each other. 第一的, modeling and experimen-
tation have been viewed as largely analogous operations aiming to isolate some
core causal factors and their effects. The argument is that both in modeling
and in experimentation one aims to seal off the influence of other causal factors
in order to study how a single causal factor operates on its own. Whereas in
experimentation this sealing off happens through experimental controls, 模组-
elers use various techniques, such as abstraction, idealization, and omission as
vehicles of isolation (例如, Cartwright 1999; Mäki 2005).
One central problem of the isolationist view is due to the fact that idealizing
and simplifying assumptions made in modeling are often driven by the require-
ments of tractability and mathematical convenience rather than those of isola-
的 (例如, Cartwright 1999). This feature of mathematical models is further
enhanced by their use of general, cross-disciplinary computational templates
那是, in the modeling process, adjusted to fit the field of application (例如,
Humphreys 2004; Knuuttila and Loettgers 2011, 2016). Such templates are
often transferred from other disciplines, as in the case of synthetic biology, 在哪里
many models, formal methods, and related concepts originate from physics and
工程 (例如, the concepts of oscillator, feedback mechanism, and noise—
见下文).
第二, when it comes to simulation, numerous philosophers and scientists
have pointed out their experimental nature as kinds of “numerical experiments”
(Winsberg 2003).1 Instead of isolation, the stress here is on intervention: both in
modeling and experimentation one seeks to intervene on a system in the light of
the results of this intervention. 最后, simulations can be thought of as
experiments performed on mathematical models, but the question is how deep
this resemblance cuts. Two issues, 尤其, have sparked discussion: 这
supposed target systems of simulations versus experiments, and the role of
materiality they incorporate.
A common intuition seems to be that, whereas in experimentation one in-
tervenes on the real target system of interest, in modeling one merely interacts
with a model system (例如, Barberousse et al. 2009). 然而, a closer examination has
assured several philosophers that these intuitions may be deceptive. Winsberg
argues that both “experiments and simulations have objects on the one hand
1. Another similarity between modeling and experimentation is related to data. 两个都
activities produce data, and deal with data analysis and error management (see Winsberg
2003, Barberousse et al. 2009). We will not discuss this third commonality.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
472
Biological Control Materialized
and targets on the other, 然后, in each case, one has to argue that the object is
suitable for studying the target” (2009, p. 579; see also Guala 2002). 因此,
both experimentation and modeling/simulation seem to display features
of surrogate reasoning (Swoyer 1991), which is visible, 例如, 在里面
experimentation on model organisms instead of the actual organisms of
兴趣. 最后, the closeness of a relationship of a model or experiment
to its respective target need not distinguish the two activities from each other.
Even though modeling and experimentation appear to come close to each
其他, at the level of scientific practice we often do not have any difficulties in
distinguishing model systems from experimental systems, although borderline
cases do exist. Models and simulations are considered kinds of representations
and are typically are expressed in other media than what their targets are made
的, whereas experimental objects are supposed to share at least partly the same
material make-up as the systems of interest. 的确, the right kind of mate-
riality has been claimed to be the distinguishing mark of experiments and even
the reason for their epistemic superiority to simulations. Guala (2002) 和
摩根 (2003) have argued that the relationship between a simulation and its
target is nevertheless abstract, while the relationship between an experimental
system and its target is grounded in the same material being governed by the
same kinds of causes. 最后, while in simulation one experiments with a
(formal) representation of the target system, in experimentation the experimental
and target systems are made of the “same stuff.” This difference also explains,
according to Morgan and Guala, why experiments have more epistemic leverage
than simulations. 例如, anomalous experimental findings are more likely
to incur change in our theoretical commitments than unexpected results from
simulations (摩根 2005).
Despite the intuitive appeal of the importance of the “same” materiality, 它
has been contested on different grounds. Morrison (2009) points out that even in
experimental contexts the causal connection with the physical systems of inter-
est is often established via models. 最后, materiality is not able to de-
liver an unequivocal epistemic standard that distinguishes simulation outputs
from experimental results. 派克 (2009) questions the alleged significance of
the “same stuff.” She interprets the “same stuff” to mean for instance the same
fluid, and points out that in traditional laboratory experiments on fluid phe-
nomena, and many other things such as the depth of the fluid and the size,
shape, roughness and the movement of any container holding it may matter.
This leads her to suggest that it is the “relevant similarities” that matter for
the justified inferences about the phenomena. Our case study on synthetic
modeling shows that the “same stuff” was crucial in the study of genetic circuits.
然而, due to the complexity of intracellular mechanisms, and our scant
knowledge of them, other kinds of models were needed that were triangulated
with synthetic models in an exploratory fashion.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
科学观点
473
Exploratory Modeling
3.
The discussion of exploratory modeling builds on the already established lit-
erature on experimental exploration (例如, Steinle 1997, 1998; Burian 1997).
Friedrich Steinle introduced the notion of exploratory experimentation in his
study of Ampère’s experiments on Oerstedt’s discovery concerning the influ-
ence of a nearby electric current on a compass needle. This discovery suggested
that there is a connection between electricity and magnetism. Steinle’s studies
of Ampère’s lab books lay out the methodological care with which Ampère
proceeded. His endeavor was not based on trial and error, although no theo-
retical framework that would have explained the connection between electric-
ity and magnetism was available. 例如, Ampère realized right in the
beginning that if he wanted to investigate whether there is a connection be-
tween electricity and magnetism, he had to isolate terrestrial magnetism from
magnetism caused by electricity.2 In order to separate the two effects, he de-
veloped a specific experimental setup. The case study shows that experiments
do not have to be guided by an encompassing theory, since there are experi-
mental guidelines such as systematic variation of parameters, formulation of
stable empirical rules, and the exploration of which of the experimental con-
ditions were necessary for the effect, and which were not. 而且, such ex-
ploratory experimentation can lead to conceptual developments especially in
the early stages of inquiry.
Contemporaneously to Steinle, and in line with his findings, Burian (1997,
p. 15) used the notion of exploratory experimentation to characterize the “elab-
orate series of interconnected experiments” that were used to (关于)identify and
localize nucleic acids in the early phases of the study of these still ephemeral
entities that are nowadays called mRNA and tRNA. The notion of exploratory
experimentation provided Burian an answer to the puzzle of how the work of the
experimentalists in different groups and even in different disciplines could con-
verge and stabilize around some shared ‘theoretical’ entities. The various proce-
dures for experimental localization of entities provided exploratory means for
scientists that did not depend “wholly on the specific disciplinary or theoretical
background of the experimenters, who initiated the work on those objects”
(Burian 1997, p. 17).
Instead of trying to highlight the experimental features of modeling, 这
philosophers interested in exploratory modeling have rather focused on some
explorative features that are specific to modeling (although they may be found
to apply to other epistemic practices as well). What the two discussions of
2.
Such separation between the two kinds of magnetism has a parallel in the case
discussed in this article, where the scientists investigating noise proceeded to construct
an experimental set-up that would allow them to distinguish between internal and external
noise (见下文).
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
474
Biological Control Materialized
exploration share, 然后, is the focus on the early stages of inquiry. 然而,
Gelfert (2018) is careful to point out that the question is not just about the
chronicle of events. Exploration seeks to get “a grasp of a phenomenon or sci-
entific problem in the absence of a well-understood and workable theory of the
domain in question” (Gelfert 2018, p. 4). He argues that this “early” character
of exploration distinguishes it from heuristics or abduction. While heuristics is
utilized in all stages of research, abduction, in Gelfert’s construal, applies to
later stages of inquiry, in which there already is a stable target phenomenon
requiring explanation. But such a well-delineated target phenomenon cannot
be taken for granted in the early stages of research. 的确, one of the four
exploratory uses of models that Gelfert puts forth is precisely that of “reasses-
sing the suitability of the target.”
The three other exploratory functions of models that Gelfert introduces are
the uses of models as starting points of inquiry, as proofs of principle, 并作为
sources of potential explanation. These uses are overlapping. Often proofs of
principle provide sources of potential explanation, and as such, starting points
of inquiry—the last one of these uses being so generally characterized that it
does not carry too much analytical potential of its own. The functioning of
models as proofs of principle and potential explanations has been commonly
recognized in the literature of modeling—covering e.g., how-possibly, fictional
and toy models—though the existing discussion has not addressed them
collectively as various modes of exploration. Apart from providing the unify-
ing notion of exploration for many so far separately examined activities of
造型, Gelfert’s discussion of reassessing the suitability of a target system
appears to us to break new ground. He studies such a process through Turing
图案, which present a case of a general mathematical model looking for
suitable targets. Turing’s model (1952) of the chemical basis of morphogenesis
seeks to explain pattern formation in an otherwise homogeneous system
through oscillations in concentrations resulting from the interaction between
two chemical substances. The system consists of an “activator” that can make
more of itself, and a quickly diffusing “inhibitor” that slows down the activa-
托尔. This simple reaction-diffusion system is able to produce very complex pat-
terns and might explain, 除其他事项外, spots and stripes in animals.
With respect to model-based exploration, especially three observations by
Gelfert concerning the history of the Turing patterns seem particularly relevant.
第一的, the biographies of scientific models are anything but linear, “models can
go out of fashion and subsequently rebound” (Gelfert 2018, p. 266). It was the
better simulation techniques that sparked a new interest in Turing patterns, 喜欢
in the case of many other simple mathematical models examining complex phe-
nomena. 第二, Turing patterns offered only a potential explanation and con-
关注的, in Turing’s own understanding, “imaginary biological systems.” Such
potential explanations are modal in nature; they target principles that might
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
科学观点
475
apply to a very wide range of different kinds of systems. 第三, Gelfert men-
tions that apart from the availability of advanced simulation models, also their
interplay with experimental manipulation was crucial for the revival of the
Turing patterns.
We suggest that these two latter points are substantially intertwined.
即, insofar as mathematical models study abstractly the dynamics of
some general phenomena, and in doing so only provide potential explanations
for particular real-world regularities or processes, the experimental grounding
of these models in the real phenomena becomes crucial. Such grounding does
not amount to testing of the very general theory, as pointed out by the liter-
ature on exploratory experimentation. 相当, it situates the general model
into the context of some real-world problem, and the existing knowledge con-
cerning it, through an exploratory interplay of modeling and experimentation.
In the following we will study exploratory modeling of genetic circuitry that
made use of modeling and experimentation in various modes and materiali-
领带: mathematical models and their simulations, synthetic genetic circuits
and measuring devices, and electronic circuits. Different models were con-
structed in different stages of the exploratory process that led, much to the
scientists’ surprise, to the redefinition of the original target, biological control.
Exploration in Biological Control
4.
Control in biological systems has been a central topic of biological sciences
for a long time. Biological control is related to the high degree of organi-
zation in biological systems, from the molecular and cellular levels up to
the organismic level, including properties that are considered to be biology
specific such as their ability to maintain themselves, 发展, 并重现-
duce. Biological control became a subject of systematic study in the begin-
ning of the twentieth century. Françoise Jacob and Jacque Monod’s (1961)
discovery that gene regulation takes place via transcription factors was a
milestone in experimental molecular biology. These findings were accom-
panied by theoretical studies making use of engineering principles, math-
ematical modeling, and general systems theory (Von Bertalanffy 1969).
Early work in the 1960s on cybernetics (Wiener 1948) and information
理论 (Shannon 1948) proceeded along the same lines. These different
developments influenced the modeling of genetic and metabolic regulation
in terms of feedback loops, 在哪里, for example genes are controlled at the
level of transcription by the products of other genes. With the introduction
of synthetic biology at the turn of the twentieth century, it finally became
possible to study such feedback systems within living cells. 中的一个
pioneers of this approach is the Elowitz lab (Caltech), whose study of gene
regulation extended into the examination of the role of noise in biological
组织.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
476
Biological Control Materialized
4.1. Mathematical and Synthetic Modeling in the Exploration of Noise
The research on noise in synthetic biology emerged from the question of how
biological control in biological systems, such as in the circadian clock, 可以
have been implemented (Loettgers 2009). Experiments on the circadian clock
in molecular biology, as well as mathematical models by mathematical biolo-
gists like Brian Goodwin (Goodwin 1963) and Arthur Winfree (Winfree 1990)
suggested that this kind of control is based on oscillations in protein levels.
Michael Elowitz together with Stanislas Leibler constructed a synthetic ge-
netic circuit, the Repressilator (Elowitz and Leibler 2000), that aimed to
furnish a proof of principle that such oscillations could be produced by var-
ious kinds of molecular feedback systems.
The Repressilator consists of three genes that repress the protein production
of each of its neighbor gene in the fashion of the rock/paper/scissors game. 这
mathematical model underlying the Repressilator is a system of non-linear
coupled differential equations of the following form:
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
dmi
dt
dpi
dt
¼ −mi þ
(西德:1)
A
1 þ pn
j
(西德:3) þ α0
¼ −β pi − mi
ð
Þ
(西德:4)
i ¼ lacI; tetR; cl
j ¼ cl; lacl; tetR
(西德:5)
和:
In this set of equations pi stands for the concentrations of the proteins sup-
pressing the function of the neighbor genes and mi (where i stands for lacI, tetR,
or λcl) are the corresponding concentrations of mRNA. There are six molecule
物种 (3 proteins functioning as repressors and 3 基因), each of them taking
part in transcription, 翻译, and degradation reactions. 一般来说, 有
no analytical solutions for such non-linear coupled differential equations, 所以
Elowitz and Leibler performed computer simulations on the basis of this math-
ematical model. The main purpose of these simulations was the identification of
relevant experimental parameters as well as the different possible states that
could be exhibited by the system. There are two such states: a steady state, 和
a state in which the system performs limit-cycle-oscillations. Being interested in
biological control, Elowitz and Leibler aimed for limit-cycle oscillations. 为了
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
科学观点
477
attaining these oscillations, the experimental parameters were of critical impor-
坦斯, and the simulations showed that such oscillations require, 例如,
strong promoters and tight transcriptional repression. This information was put
to use in the construction of the actual genetic network, the Repressilator, 为了
which the mathematical model functioned as a blueprint.
Although the computer simulations provided some important experi-
mental parameters, most of the biochemical parameters remained unknown.
The Repressilator did not seek to represent any naturally existing genetic
circuit, it was a very simple construct made of well-characterized molecular
components adopted from different contexts of research in view of obtaining
robust oscillations. The same applies to the mathematical model that under-
lies it. Both models, the mathematical and the synthetic, can better be un-
derstood from the exploratory than the traditional representational
看法; they were purposefully constructed epistemic artifacts for prob-
ing possible architectures and dynamics of genetic circuits (参见部分 5 是-
low for further discussion).
Another important exploratory dimension of the Repressilator was its
implementation within a living bacterial cell that allowed the model to
be explored in its natural biochemical environment. The actual genetic network
was constructed by making use of a plasmid that was introduced into a cell.
Plasmids are extra-chromosomal DNA molecules that are self-replicating.
数字 1 shows the architecture of the Repressilator and the interaction be-
tween the three genes.
The genes of the Repressilator are connected via a negative feedback loop
through which they repress each other’s protein production. Green fluorescent
蛋白质 (GFP) is used as a reporter and it is fused to the tetR gene. This construct
makes the oscillations in the protein level of the gene visible through fluores-
cence microscopy. The entire construct, consisting of the Repressilator and the
记者, is integrated into E. coli bacteria. Its being constructed from biological
成分, and integrated into the bacteria, made the system more biology
喜欢. Although the biochemical interactions in the cell are largely unknown,
this embedment, as Waters (2012) has lucidly spelled out, “avoids having to
数字 1. The diagrams show sketches of the Repressilator and the reporter
(Elowitz and Leibler 2000).
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
478
Biological Control Materialized
understand the details of the complexity, not by assuming that complexity is
irrelevant but by incorporating the complexity in the models.” The Repressila-
tor was able to produce oscillations, but they turned out noisy (相比之下
to what the underlying mathematical model predicted). 数字 2 节目
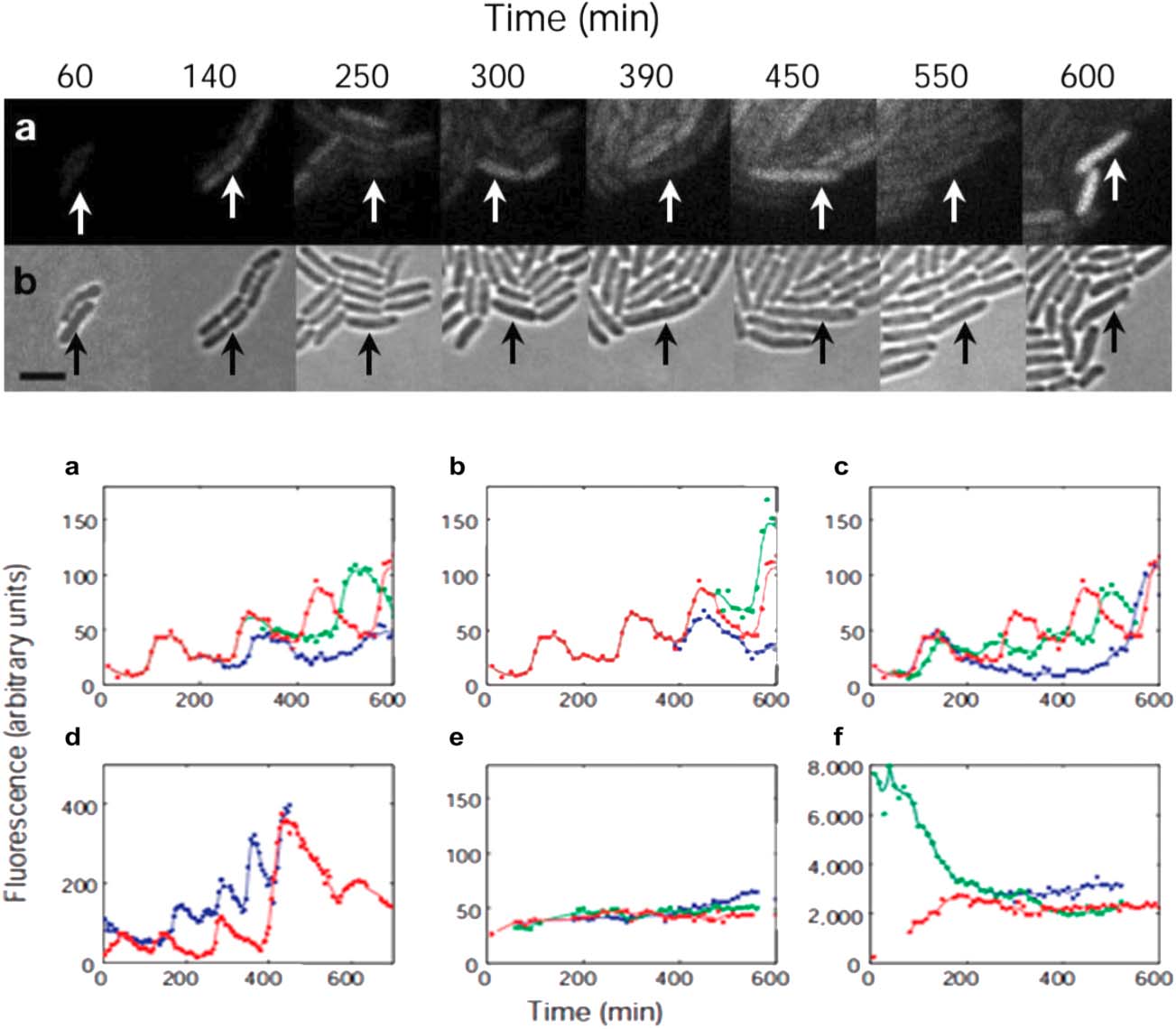
the oscillations of the Repressilator, both in the growing bacteria colony,
and in the diagrams depicting the measurements of oscillations in indi-
vidual Repressilators.
The closer look at the pictures of the blinking bacteria colonies reveals that
the blinking of the bacteria, IE。, the oscillations made visible by the reporter, 是
not synchronized. This non-synchronization is manifested even more obviously
in the lower diagrams (a–c), showing the fluorescence of different sibling cells.
Here the red line is a reference line representing the oscillations of the whole
bacteria colony and the blue and green lines belong to oscillations of single
sibling cells. The diagrams show that the amplitudes of the oscillations of
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
数字 2. The upper picture shows the growing population of E. coli bacteria
carrying the Repressilator. The lower picture shows the oscillations of single E. 大肠杆菌
bacteria over time (Elowitz and Leibler 2000, p. 336).
科学观点
479
the sibling cells change over time, meaning that there is a difference in the
amount of proteins produced over time by the reporter gene. 第二, 这
phase of the oscillations in the two bacteria shift over time. 换句话说,
the sibling cells show some individual behavior (phase shift) but there is also
some variability in this individual behavior (changes in amplitude). (这
图形 (d) presents oscillations obtained in different experiments, 和 (e–f )
are the result of negative control experiments.)
The single cell measurements the Elowitz lab performed were laborious.
One source of difficulties was due to choosing the “right” parameter values.
Elowitz explained this in the following way:
[氧]ne thing we’ve seen is that, the first generation of synthetic
circuits were often overexpressing proteins a lot, and consumed a lot
of resources and [那] often makes the cells grow a little bit slower
than other cells that don’t have the synthetic circuit [……] it just
means that maybe they’re not optimized yet to be sufficiently
独立的 [……] the other side of it is that the environment inside
the cell is not the environment of our model where there [是], 你
知道, continuous variables and continuous trajectories. (Elowitz in
interview conducted by Andrea Loettgers)
Another source has been the large apparatus consisting of microscopes,
image processing utilities and computer programs that were brought
into play and adjusted for the analysis of the dynamics of the synthetic
系统. The complexity of this experimental set up contrasts with the
apparent simplicity of the synthetic genetic circuit. Only when one
focuses on the noisy character of the observed fluctuations, is one able to
get a glimpse of the complexity of even such simple systems as the
Repressilator.
The observed individual behavior of cells, shown in the phase shifts and fluc-
tuations in Figure 1, provided a first clue that the fluctuations could be of a
stochastic nature. Most probably, the researchers assumed, they were caused
by the limited number of molecules in cells. In order to explore the noisy be-
havior exhibited by the Repressilator, the researchers performed computer sim-
ulations of a stochastic version of the initial mathematical model that seemed to
confirm the stochastic nature of the observed fluctuations. Two related questions
appeared: 第一的, how are regular oscillations possible at all in the stochastic en-
vironment of a cell, 和, 第二, how are stochastic fluctuations related to other
sources of noise that occur independently from the observed stochastic fluctua-
系统蒸发散. Both questions were explored by further models and synthetic constructs.
Of a particular interest is the synthetic intracellular measuring device that the
Elowitz group developed to explore stochastic fluctuations by making them
“visible” on a single cell level.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
480
Biological Control Materialized
4.2. Measuring Stochastic Fluctuations in Biological Systems
A question, which was unable to be explored by the Repressilator, 是
total amount of noise in a biological system. The total amount of noise is
given by the sum of extrinsic and intrinsic noise. The Repressilator only pro-
vides insight into stochastic noise, which is part of intrinsic noise that has at
least two sources:
(西德:129) Individual messenger RNA does not get translated only one time during
the transcription process, but can be translated many times, 导致
the production of many proteins instead of one.
(西德:129) Promoters that regulate the transcription process by turning the genes
‘off’ and ‘on,’ can switch between long-lived ‘off’ and ‘on’ states.
As further experiments on stochastic fluctuations showed, both processes re-
sult in bursts of mRNA and, related to it, bursts of proteins reflected by the
observed fluctuations in the protein level (Ingram et al. 2008, 卷. 4). Intrinsic
noise is closely linked to extrinsic noise. While intrinsic noise are fluctuations
generating bursts of proteins, extrinsic noise are the propagated bursts of pro-
teins affecting the expression and protein production of other genes in the sys-
TEM. There are various other extrinsic sources of noise that arise independently of
the gene, yet act on it, IE。, the stage of the cell cycle, the mRNA degradation
machinery and the cell environment fluctuations. Although extrinsic fluctua-
tions are part of the dynamic and behavior of the Repressilator, they cannot
be identified in its oscillations.
The basic motivation for constructing the synthetic intracellular measur-
ing device was to distinguish between extrinsic and intrinsic fluctuations by
separating them from each other (Elowitz et al. 2002; Swain et al. 2002). 在
constructing the measuring device Elowitz and colleagues made again use
of E. coli bacteria. This time they integrated into the chromosome of the
bacteria cyan cfp and yellow yfp alleles of green fluorescent proteins, 哪个
were then put under the control of identical promoters. In this set-up the
intrinsic noise affects each of the promoters of the two reporter genes sepa-
率地. The two genes are uncorrelated, meaning that the proteins produced
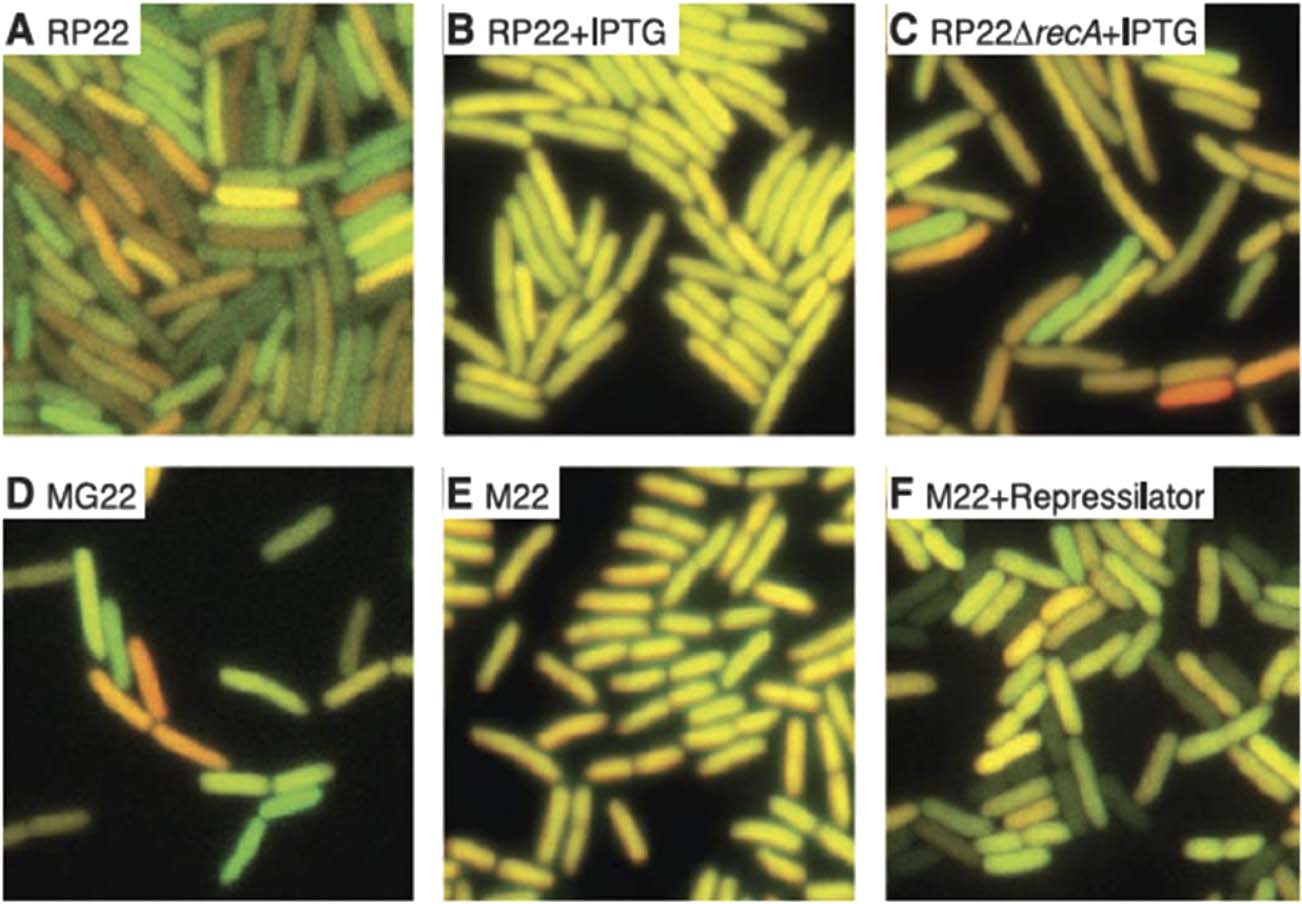
by the genes fluctuate in an uncorrelated fashion (see the lower part B of
数字 3). This gives rise to a population of cells, in which some cells express
more of one fluorescent protein than the others. 作为结果, the cells in
the populations appear in different colors such as yellow, orange, 红色的, 和
绿色的.
In the absence of intrinsic noise at the two reporter genes, the two genes,
which are located in the same cell, are only exposed to extrinsic noise that is
the same for each of the genes. 最后, the cells with the same amount
of each protein appear yellow, as shown by the upper part A of Figure 3.
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
科学观点
481
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
数字 3. The fluctuations due to the extrinsic (A) and intrinsic (乙) noise and
the corresponding variations in fluorescence (Elowitz et al. 2002, p. 1185).
In an interview Michael Elowitz reflected on his expectations concerning
the experiment:
[瓦]hen I first was doing that experiment I really didn’t know [……].
There had been a lot of work theoretically, simulations, on how noise
should be significant. But depending on the effective biochemical
parameters inside the cell, which really were not known, 和
depending on how big the extrinsic noise is, it was hard to say
whether the extrinsic or intrinsic effects would be stronger. 我认为
what was interesting for me was, going into it, I really had no idea
whether noise would just be a small perturbation on top of these big
extrinsic fluctuations, or whether actually everything would be
dominated by intrinsic noise. 所以, I think that was what was kind of
fun about it [……] (interview conducted by Andrea Loettgers)
In exploring the relation of intrinsic and extrinsic noise under different con-
版本, experiments were performed by introducing the reporter genes into
different strains of E. 大肠杆菌. The differences between these strains depended on
how strongly the genes of the regulator sequences, to which the reporter genes
were fused, were transcribed. By performing these experiments, Elowitz and
co-workers were able to explore how the transcription of a gene is related to
intrinsic and extrinsic noise. The results were differently colored colonies of
cells depending on the level of intrinsic and extrinsic noise (见图 4).
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
482
Biological Control Materialized
而且, to study the interplay between the regulatory dynamics and noise,
researchers introduced the Repressilator into one of the strains. From the re-
苏丹, they concluded that changes in the regulatory dynamics may cause sub-
stantial changes in noise levels.
These measurements were constrained by how much the parameters of
the synthetic measuring device could be changed in a controlled fashion.
Even this comparatively simple system turned out to be very complex and
experimentally difficult to access. In addition to the Repressilator, 和
measuring device for distinguishing between extrinsic and intrinsic noise,
a third model was designed and used to explore noise in biological systems.
The third system was not synthetic, instead it was an electronic version of
the Repressilator. What could be learned from an electronic model about
noise in biological systems?
The Electronic Repressilator
4.3.
Even though Elowitz and co-workers had been able to distinguish between
extrinsic and intrinsic noise, and to explore them under different conditions,
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
数字 4. The pictures show the outcome of the experiments in which noise was
explored under different experimental conditions by, 例如, changing the
reporter genes. (Elowitz et al. 2002, p. 1186). The last picture (F) shows the
influence of the periodic dynamic of the Repressilator on the level of intrinsic and
extrinsic noise in the cells (IE。, the contrast between pictures E and F).
科学观点
483
the question of how regular oscillations are possible at all under these cir-
cumstances still needed to be answered. To answer this question an elec-
tronic Repressilator was constructed (Buldú et al. (2007). The electronic
Repressilator was inspired partly by the work of Mason et al. (2004), WHO
constructed an electronic circuit based on ordinary differential equations
modeling a genetic network. Such an electronic circuit provides a good
model for the study of robust oscillations, 自从, as the researchers put it
“this system is subject to electronic noise and time delays associated with
its operation, and since its parameters depend on the actual values of
capacitances and resistors [……].” (Mason et al. 2004, p. 709).
While Mason et al. (2004) constructed a hybrid digital-analog circuit,
the electronic Repressilator of Buldú et al. (2007) is purely analog. Among
the advantages of this approach is the experimental flexibility of electronic
circuits as well as the possibility of constructing integrated circuits with a
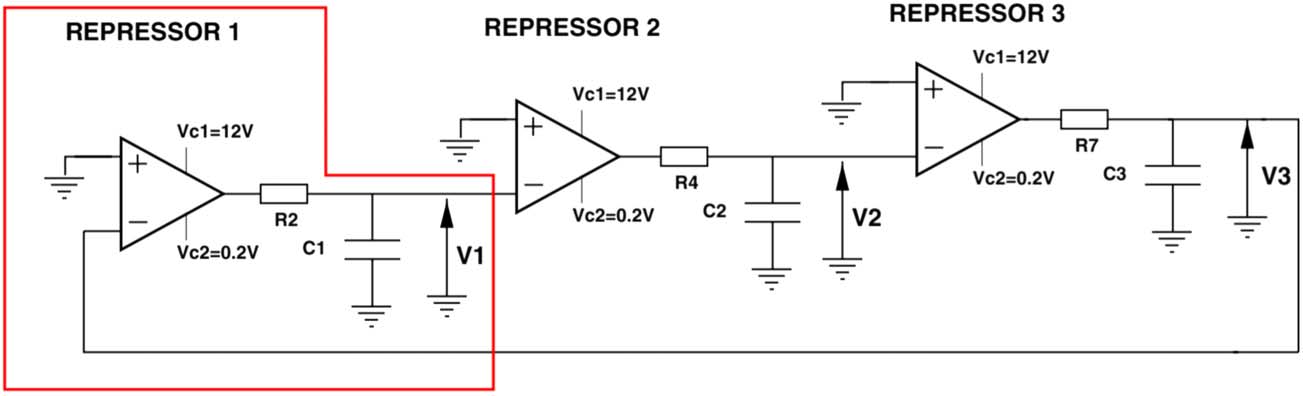
big number of elements. 数字 5 shows the layout of this electronic cir-
崔特. It is based on the same network design as the biological Repressilator
consisting of “three dynamical elements coupled in chain with an inhibi-
tory interaction” (Buldú et al. 2007, p. 3508).
The electronic version of the Repressilator is, as is the corresponding syn-
thetic model, based on a mathematical model. It consists of three elements,
each of them modeling a gene, which becomes repressed by the proteins pro-
duced by its neighbor gene. The three elements themselves consist of opera-
tional amplifiers and RC (R=Resistance C=Capacitor) circuits. 这
operational amplifier, designated as a triangle in the diagram, functions as a
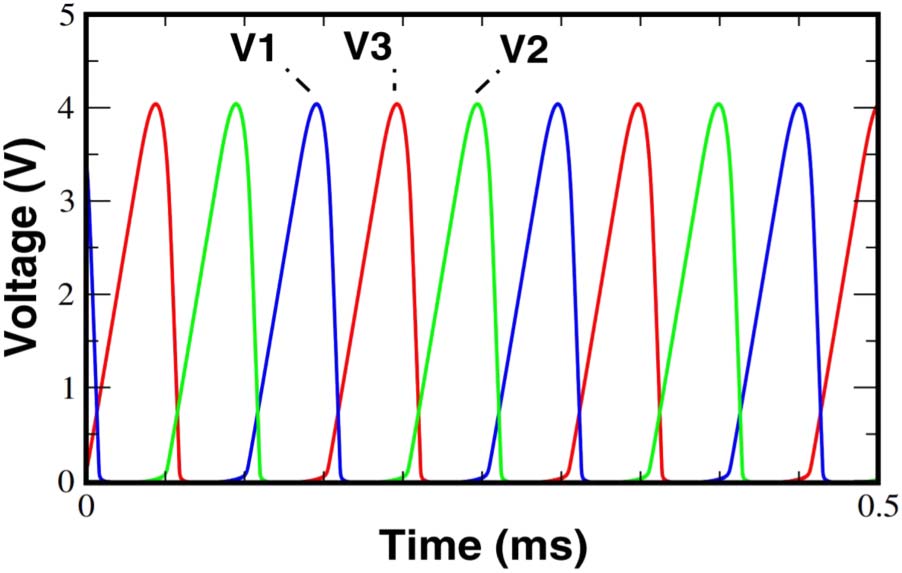
comparator and the RC circuit as filter. The voltages V1, V2, and V3 are the
analogues to the protein concentrations in the synthetic model. The dynamic
of the electronic circuit can be described in the following way: in case the volt-
age V2 increases, it induces a reduction of the following output voltage V3,
which again lead to an increase of V1. The model results in regular oscil-
lations in the three output voltages of the electronic circuit and is known
数字 5. The diagram is a sketch of the electronic version of the Repressilator
(Buldú et al. 2007, p. 3508).
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
484
Biological Control Materialized
in electrical engineering as a ring oscillator (displaying the engineering
origin of the Repressilator) (数字 6).
The electronic Repressilator shows that robust oscillations are possible
despite of the presence of noise. But how could this result be transferred into the
context of biology? The problem becomes one of relating the parameters of the
electronic circuit to the genetic circuit, and it has so far remained unsolved.
因此, it seems that there are limitations to the widespread practice in
synthetic biology of drawing analogies between electronic and genetic circuits.
The electronic Repressilator turned out to be a valuable model in the exploration
of possible network designs, 然而, due to its combination of experimental
flexibility and implementation of noise in the electronic components. An exam-
ple of this line of work is the interdisciplinary collaboration between Michael
Elowitz, his long-time collaborator physicist Jordi García-Ojalvo, and mathe-
matician Steven Strogatz. They constructed a mathematical model of a popula-
tion of Repressilators, coupled by quorum sensing (García-Ojalvo et al. 2004).
Quorum sensing is the common way of how bacteria communicate with each
other by exchanging molecules. Following Strogatz’s interest in synchronization
of oscillatory systems such as the blinking of fireflies, García-Ojalvo et al. 用过的
the mathematical model together with a computer simulation to explore con-
ditions under which the coupled Repressilators would synchronize. 甚至
though the model predicted that “a diverse and noisy community of such
genetic oscillators interacting through quorum-sensing mechanism should
self-synchronize in a robust way” (García-Ojalvo et al. 2004, p. 10955), 这
attempt of constructing a synthetic system of coupled Repressilators failed.
Buldú et al. (2007) succeeded, 然而, in constructing an electronic version
of the population of Repressilators that was able to exhibit synchronized
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
数字 6. The diagram shows the oscillations in voltages in the electronic
circuit (Buldú et al. 2007, p. 3508).
科学观点
485
oscillations. 尽管如此, given the problem of relating the electronic and
biochemical parameters, they could not answer the question of why the
synthetic system failed to synchronize.
总结, the Repressilator showed that it was possible to build, 根据
to engineering and physical principles, a biological circuit able to oscillate. 如何-
曾经, as these oscillations turned out to be noisy, the Elowitz lab went on to study
the sources of noise, 一方面, and the possibility of robust oscillations and
the synchronization of oscillating bacteria, 在另一. While the former studies
were performed with synthetic systems, the latter were carried out with electronic
circuits and mathematical modeling and simulation. 还, elsewhere in synthetic
biology a lot of effort was put into designing robust genetic circuits. Here espe-
cially the work of the Hasty lab at the University of California San Diego needs to
be mentioned. Hasty and his co-workers were able to construct a robust oscilla-
tory genetic circuit by paying close attention to the biochemical parameters, ba-
sically involving an unintended interaction between the synthetic genetic circuit
and the host cell (Cookson et al. 2009; Stricker et al. 2008).
最后, Elowitz and his co-workers also addressed the role of noise in biolog-
ical organization. In several studies, the members of the lab showed that noise is
not a nuisance in biology but has a functional meaning (Süel et al. 2007; Eldar
and Elowitz 2010). Noise, in the form of stochastic fluctuations, triggers as well
as controls processes in cells. In addressing the functional meaning of noise, 这
Elowitz lab drew inspiration from the study of neurodynamics and lasers, 男人-
tioning also the classic experiments by Spudich and Koshland (1976) on non-
genetic variability of the bacterial chemotaxis. Spudich and Koshland had
concluded that “nongenetic variability would be a preferred mechanism for ac-
commodation to random fluctuations in the environment and genetic variability
the preferred mechanism for accommodation to long lasting environmental
changes” (1976, p. 470). With the benefit of new technologies, the Elowitz
lab was able to study such nongenetic variability in the form of stochastic fluc-
tuations within living cells. 重要的, their exploration on noise led to a de-
velopment of an experimental-cum-conceptual framework in which noise and
control became intertwined and redefined.
5. Discussion and Conclusions
The exploratory research program on biological control and noise by the
Elowitz lab and its collaborators portrays an intricate fabric of mathematical
modeling and simulation, intracellular measurement, and the construction of
synthetic genetic circuits and electronic circuits. We deem multiple modeling
and the combination of different epistemic means a central scientific practice
that merits more research from the exploratory perspective. That contemporary
modeling practices typically employ multiple models and triangulate them
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
486
Biological Control Materialized
with other epistemic activities is something that has been recognized and ana-
lyzed by the philosophical discussion surrounding robustness. 然而, robust-
ness analysis has been focused on convergence; on either the use of multiple
related models to causally isolate a core mechanism producing certain phenom-
ena (Knuuttila and Loettgers 2011), or on the use of independent epistemic
方法 (型号, 实验, and observations) to “triangulate the existence
and character of a common phenomenon, object or result” (Wimsatt 2007,
p. 43). Although both of these aspects of robustness analysis are present in
our case study, the notion of exploratory modeling better captures how the
Elowitz lab and its collaborators studied biological control, and noise. In line
with robustness analysis, the scientists were using multiple, materially distinct
and thus partially independent epistemic means, in order to study whether the
simple network design could create robust oscillations at the molecular level.
然而, this triangulation process is better conceived of as providing an initial
proof of principle, in the form of the Repressilator model that then led into
further exploration into the sources and role of noise in biological control. 在
this explorative process, materially different models were used rather in a dia-
logue with each other than only as an attempt to isolate a core causal mechanism
common to all of them. 相比之下, the models studied were different material
realizations (digital, synthetic, and electronic) of a particular circuit design.3
Massimi (2018) discusses multiple models as means of exploration, and casts,
而且, exploratory modeling in a modal idiom. She presents a case in Beyond
Standard Model (BMS) 物理, where different models perform the exploratory
function in charting and carving out the space of possibilities (Massimi 2018,
p. 349). The Elowitz lab’s research on biological control was also importantly
modal in that it was motivated by the question of whether the kind of feedback
systems already familiar from physics and engineering, and theorized since the
early 1960s (例如, Jacob and Monod 1961) could be realizable in biological or-
ganisms. Synthetic biology finally gave means to scientists to study such possi-
ble general design principles of biological organization within living cells. 这
underlying additive character of Massimi’s “perspectival modeling” does not
easily accommodate our example, 然而. In her construal, perspectival models
provide each a partial account of the phenomenon of interest, where the “explor-
atory function is performed jointly” (Massimi 2018, p. 350). The multiple
modeling process of the Elowitz lab had more of an unfolding character. 他们的
models and measurements functioned as kinds of target systems/research ob-
jects for each other in the investigation of the entangled, and largely unknown
phenomena of noise and control. In this process the phenomenon of interest,
3. The epistemic importance of the different materialities and formats of models has
also been studied by e.g., Chandrasekharan and Nersessian (2011); Vorms (2012).
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
科学观点
487
biological control, became reassessed, as researchers realized that noise might
play a crucial role in it.
最后, the synthetic biology program of the Elowitz lab provides in-
sight into the discussion of models as experiments. 的建设
synthetic constructs—synthetic genetic circuits and synthetic intracellular
measuring devices—each functioning within individual, and multiplying,
bacterial cells, provides the particular novelty of the research practice of the
Elowitz lab. For considering the experimental character of models, syn-
thetic genetic circuits seem especially exciting, as they seem difficult to
place either in the category of modeling or that of experimentation. 这
comparison of synthetic genetic circuits to the electronic circuits on the
一只手, and to the synthetic intracellular measuring devices on the other,
helps to identify the model-like and experimental features of synthetic ge-
netic circuits.
第一的, while both electronic and synthetic genetic circuits are material model
systems used experimentally to study biological control, we seem more inclined
to treat such electronic circuits as models than synthetic circuits. 为什么? 这
answer appears to be that because electronic circuits are implemented in another
medium than the biological systems of interest, they are more readily called
型号. 例如, Rheinberger (2015) distinguishes what he calls “prepara-
tions” from models on this basis: while preparations “participate in the materi-
ality of the object of knowledge in question,” models do not (p. 325). 我们不
consider this a very substantial point. Both electronic circuits and synthetic cir-
cuits can be considered models in that they are tightly constrained and (到一个
程度) self-contained constructions designed to explore certain pertinent theo-
retical and empirical questions (Knuuttila 2011, 2017). Neither of them aims
to represent some particular naturally occurring target systems, they rather pres-
ent and study very rudimentary and highly hypothetical design principles, ten-
tatively applying to actual and possible non-actual biological systems alike.
Such a program is in line with the grander vision of synthetic biology: “the ex-
pansion of biology from a discipline that focuses on natural organisms to one
that includes potential organisms” (Elowitz and Lim 2010, p. 889). This modal
character of synthetic biology is precisely one of the most important reasons for
approaching it from the exploratory perspective (Knuuttila forthcoming;
Knuuttila and Koskinen 2020; Gelfert 2016, 2018).
The comparison of the Repressilator to the synthetic measuring device, 这
noise sensor, further underlines its model-like character. In contrast to the Re-
pressilator, the noise sensor was not constructed to fulfill some specific biological
function and, 最后, to exhibit at least partially independent behavior
from the rest of the cell. In designing the Repressilator, 相比之下, the scientists
aimed for a synthetic module that would, by being able to create new behavior,
provide a tool for the exploration of possible design principles of biological
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
488
Biological Control Materialized
有机体 (see above). The noise sensor as opposed to the Repressilator did not
have a dynamic of its own. The independent functioning of the Repressilator
was crucial for its role as an exploratory tool, whereas the noise sensor was
implemented as an integral part of the cell and was supposed to be responsive
to various conditions taking place in it. 因此, although the Repressilator and
the noise sensor were both parts of the cell, they were considered different
kinds of things with only the former having a proper model-like character.
然而, 同时, the material make-up of the Repressilator, 它是
being of the same stuff as the naturally occurring genetic circuits, and fur-
thermore implemented in the natural cell environment, gave it a highly
experimental character. 像这样, it was exposed to the (largely unknown)
constraints of naturally evolved biological systems which explains how the
researchers reacted to the unexpected results. The irregular oscillations
were taken more seriously than any particular results derived from math-
ematical models would have been, spawning a new research program on
noise. The fusion of the model-like and experimental features of the Re-
pressilator speaks in favor of considering it from a broad exploratory per-
观望的. There is no need to dress such human epistemic artifacts as
synthetic genetic circuits in either the experimental or modeling guise,
as they can function in both modes, even simultaneously.
参考
Barberousse, Anouk, Sara Franceschelli, and Cyrille Imbert. 2009. “康-
puter Simulations as Experiments.” Synthese 169: 557–574. https://土井
.org/10.1007/s11229-008-9430-7
Buldú, Javier M., Jordi García-Ojalvo, Alexandre Wagemakers, and Miguel A. F.
桑胡安. 2007. “Electronic Design of Synthetic Genetic Networks.” Interna-
tional Journal of Bifurcation and Chaos 17(10): 3507–3511. https://doi.org/10
.1142/S0218127407019275
Burian, 理查德. 1997. “Exploratory Experimentation and the Role of Histo-
chemical Techniques in the Work of Jean Brachet, 1938–1952.” History and
Philosophy of the Life Sciences 19(1): 27–45. 考研: 9284641
Cartwright, 南希. 1999. “The Vanity of Rigour in Economics: Theoretical
Models and Galilean Experiments.” Discussion Paper Series 43/99, Centre
for Philosophy of Natural and Social Sciences, London School of Economics.
Chandrasekharan, Sanjay, and Nancy Nersessian. 2011. “Building Cognition:
The Construction of External Representations for Discovery.” Proceedings of
认知科学学会 33: 267–272.
Cookson, Natalia A., Lev S. Tsimring, and Jeff Hasty. 2009. “The Pedestrian
Watchmaker: Genetic Clocks from Engineered Oscillators.” FEBS Letters
583: 3931–3937. https://doi.org/10.1016/j.febslet.2009.10.089, 考研:
19903483
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
科学观点
489
Eldar, Avigdor, and Michael B. Elowitz. 2010. “Functional Roles for Noise in
Genetic Circuits.” Nature 467: 167–173. https://doi.org/10.1038
/nature09326, 考研: 20829787
Elowitz, Michael B., and Stanislas Leibler. 2000. “A Synthetic Oscillatory
Network of Transcriptional Regulators.” Nature 403: 335–338. https://
doi.org/10.1038/35002125, 考研: 10659856
Elowitz, Michael B., Arnold J. 莱文, 埃里克·D. Siggia, and Peter S. Swain. 2002.
“Stochastic Gene Expression in a Single Cell.” Science 297(5584):
1183–1186. https://doi.org/10.1126/science.1070919, 考研: 12183631
Elowitz, Michael B., and Wendell A. Lim. 2010. “Build Life to Under-
stand It.” Nature 468: 889–890.
Fisher, 授予. 2017. “Diagnostics and the ‘Deconstruction’ of Models.”
Phil-Sci Archive. philsci-archive.pitt.edu/12573
García-Ojalvo, Jordi, Michael B. Elowitz, and Steven H. Strogatz. 2004.
“Modeling a Synthetic Multicellular Clock: Repressilators coupled by
Quorum Sensing.” Proceedings of the National Academy of Science 101:
10955–10960. https://doi.org/10.1073/pnas.0307095101, 考研:
15256602
Gelfert, Axel. 2016. How to Do Science With Models: A Philosophical Primer.
占婆: 施普林格. https://doi.org/10.1007/978-3-319-27954-1
Gelfert, Axel. 2018. “Models in Search of Targets: Exploratory Modelling
and the Case of Turing Patterns.” Pp. 245–271 in Philosophy of Science: 是-
tween the Natural Sciences, the Social Sciences, and the Humanities. Edited by
Alexander Christian, David Hommen, Nina Retzlaff, and Gerhard Schurz.
纽约: 施普林格. https://doi.org/10.1007/978-3-319-72577-2_14
Goodwin, Brian C. 1963. “Temporal Organization in Cells: A Dynamic Theory of
Cellular Control Processes.” London and New York: 学术出版社. https://
doi.org/10.5962/bhl.title.6268
Guala, Francesco. 2002. “Models, Simulations, and Experiments.” Pp. 59–74
in Model-Based Reasoning: 科学, 技术, Values. Edited by Lorenzo
Magnani and Nancy J. Nersessian. 纽约: Kluwer. https://doi.org
/10.1007/978-1-4615-0605-8_4
Humphreys, 保罗. 2004. “Extending Ourselves: Computational Science, Empiricism,
and Scientific Method.” Oxford: 牛津大学出版社. https://doi.org/10
.1093/0195158709.001.0001
Ingram, Piers J., 迈克尔·P. Stumpf, and Jaroslav Stark. 2008. “Nonidentifia-
bility of the Source of Intrinsic Noise in Gene Expression from Single-Burst
Data.” PLoS Computational Biology 4(10): 1–10. https://doi.org/10.1371
/journal.pcbi.1000192, 考研: 18846201
雅各布, François, and Jacques Monod. 1961. “Genetic Regulatory Mechanisms
in the Synthesis of Proteins.” Journal of Molecular Biology 3: 318–356. https://
doi.org/10.1016/S0022-2836(61)80072-7
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
490
Biological Control Materialized
Knuuttila, Tarja. 2011. “Modeling and Representing: An Artefactual Ap-
proach.” Studies in History and Philosophy of Science 42: 262–271. https://土井
.org/10.1016/j.shpsa.2010.11.034
Knuuttila, Tarja. 2017. “Imagination Extended and Embedded: Artifactual
Versus Fictional Accounts of Models.” Synthese. https://doi.org/10.1007
/s11229-017-1545-2 (Accessed 22 七月 2021)
Knuuttila, Tarja. 2021. “Epistemic Artifacts and the Modal Dimension of
Modeling.” European Journal for Philosophy of Science 11(65). https://doi.org
/10.1007/s13194-021-00374-5 (Accessed 23 七月 2021)
Knuuttila, Tarja, and Rami Koskinen. 2020. “Synthetic Fictions: Turning
Imagined Biological Systems into Concrete Ones.” Synthese. https://土井
.org/10.1007/s11229-020-02567-6
Knuuttila, Tarja, and Andrea Loettgers. 2011. “Causal Isolation Robustness
分析: The Combinatorial Strategy in Synthetic Biology.” Biology and
Philosophy 26: 773–791. https://doi.org/10.1007/s10539-011-9279-x
Knuuttila, Tarja, and Andrea Loettgers. 2013A. “The Productive Tension:
Mechanisms vs. Templates in Modeling the Phenomena.” Pp. 3–24 in
Representations, 楷模, and Simulations. Edited by Pauly Humphreys and
Cyrille Imbert. 纽约: 劳特利奇.
Knuuttila, Tarja, and Andrea Loettgers. 2013乙. “Synthetic Modeling and
the Mechanistic Account: Material Recombination and Beyond.” Philosophy
of Science 80: 874–885. https://doi.org/10.1086/673965
Knuuttila, Tarja, and Andrea Loettgers. 2016. “Model Templates Within
and Between Disciplines; From Magnets to Gases—and Socio-Economic
Systems.” European Journal for Philosophy of Science 6(3): 377–400. https://
doi.org/10.1007/s13194-016-0145-1
Loettgers, 安德里亚. (2009). “Synthetic Biology and the Emergence of a Dual
Meaning of Noise.” Biological Theory 4: 340–356. https://doi.org/10.1162
/BIOT_a_00009
Mäki, Uskali. 2005. “Models Are Experiments, Experiments Are Models.”
Journal of Economic Methodology 12: 303–315. https://doi.org/10.1080
/13501780500086255
石匠, 乔纳森, Paul S. Linsay, 詹姆斯·J. 柯林斯, and Leon Glass. 2004.
“Evolving Complex Dynamics in Electronic Models of Genetic Networks.”
Chaos 14: 707–715. https://doi.org/10.1063/1.1786683, 考研: 15446982
Massimi, Michela. 2018. “Perspectival Modeling.” Philosophy of Science 85:
335–359. https://doi.org/10.1086/697745
摩根, Mary S. 2003. “Experiments without Material Intervention:
Model Experiments, Virtual Experiments and Virtually Experiments.”
Pp. 216–235 in The Philosophy of Scientific Experimentation. Edited by Hans
Radder. Pittsburgh: University of Pittsburgh Press. https://doi.org/10
.2307/j.ctt5hjsnf.15
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
科学观点
491
摩根, Mary S. 2005. “Experiments versus Models: New Phenomena, Infer-
ence and Surprise.” Journal of Economic Methodology 12: 317–329. https://土井
.org/10.1080/13501780500086313
Morrison, 玛格丽特. 2009. “Models, Measurement and Computer Simula-
系统蒸发散: The Changing Face of Experimentation.” Philosophical Studies 143:
33–57. https://doi.org/10.1007/s11098-008-9317-y
派克, Wendy. 2009. “Does Matter Really Matter? Computer Simulations,
Experiments and Materiality.” Synthese 169: 483–496. https://doi.org/10
.1007/s11229-008-9434-3
Rheinberger, Hans-Jörg. 2015. “Preparations, 楷模, and Simulations.”
History and Philosophy of the Life Sciences 36(3): 321–334. https://doi.org
/10.1007/s40656-014-0049-3, 考研: 26013191
Shannon, Claude. 1948. “A Mathematical Theory of Communication.”
Bell System Technical Journal. Short Hills, 新泽西州. 27: 379–423, 623–656.
https://doi.org/10.1002/j.1538-7305.1948.tb01338.x
Spudich, John L., and Daniel E. Koshland. 1976. “Non-genetic Individ-
uality: Chance in the Single Cell.” Nature 262: 467–471. https://doi.org
/10.1038/262467a0, 考研: 958399
Steinle, 弗里德里希. 1997. “Entering New Fields: Exploratory Uses of Exper-
imentation.” Philosophy of Science (会议记录) 64: 65–74. https://doi.org
/10.1086/392587
Steinle, 弗里德里希. 1998. “Exploratives vs. Theoriebestimmtes Experimen-
tieren: Ampères frühe Arbeiten zum Elektromagnetismus.” Experimental
随笔 – Versuche zum Experiment. Edited by Michael Heidelberger and
Friedrich Steinle. Baden-Baden: Nomos Verlagsgesellschaft.
Stricker, Jesse, Scott Cookson, Matthew R. Bennet, William H. Mather,
Lev S. Tsimring, and Jeff Hasty. 2008. “A Fast, Robust and Tunable
Synthetic Gene Oscillator.” Nature 456: 516–519. https://doi.org/10
.1038/nature07389, 考研: 18971928
Süel, Gürol M., Rajan P. Kulkarin, Jonathan Dworkin, Jordi Garcia-Ojalvo,
and Michael B. Elowitz. 2007. “Tunability and Noise Dependence in Dif-
ferentiation Dynamics.” Science 315: 1716–1719. https://doi.org/10.1126
/science.1137455, 考研: 17379809
Swain, Peter S., Michael B. Elowitz, and Eric D. Siggia. 2002. “Intrinsic and
Extrinsic Contributions to Stochasticity in Gene Expression.” Proceedings of
the National Academy of Science 99(20): 12795–12800. https://doi.org/10
.1073/pnas.162041399, 考研: 12237400
Swoyer, Chris. 1991. “Structural Representation and Surrogative Reason-
ing.” Synthese 87(3): 449–508. https://doi.org/10.1007/BF00499820
图灵, Alan M. 1952. “The Chemical Basis of Morphogenesis.” Philosophical
Transactions of the Royal Society of London B 237 (641): 37–72. https://doi.org
/10.1098/rstb.1952.0012
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3
492
Biological Control Materialized
Von Bertalanffy, 路德维希. 1969. General Systems Theory. Foundations, Develop-
蒙特, Application. 纽约: George Braziller.
Vorms, Marion. 2012. “Formats of Representation in Scientific Theoris-
ing.” Pp. 250–273 in Models, Simulations, and and Representations. Edited
by Paul Humphreys and Cyrille Imbert. 纽约: 劳特利奇.
沃特斯, C. Kenneth. 2012. “Experimental Modeling as a Form of Theoret-
ical Modeling.” Paper presented at the Philosophy of Science Association
23rd Meeting, 15–17 November, 圣地亚哥.
Wiener, Norbert. 1948. Cybernetics or Control and Communication in the Animal
and Machine. 巴黎: Hermann & Cie/Cambridge Mass.: 与新闻界.
Winfree, Arthur T. 1990. The Geometry of Biological Time. Berlin and Heidelberg:
施普林格.
Winsberg, Eric. 2003. “Simulated Experiments: Methodology for a Virtual
World.” Philosophy of Science 70: 105–125. https://doi.org/10.1086/367872
Winsberg, Eric. 2009. “A Tale of Two Methods.” Synthese 169: 575–592.
https://doi.org/10.1007/s11229-008-9437-0
Wimsatt, William C. 2007. Re-Engineering Philosophy for Limited Beings: Approx-
imations to Reality. 剑桥: 哈佛大学出版社. https://doi.org/10
.2307/j.ctv1pncnrh
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
/
e
d
你
p
哦
s
C
/
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
2
9
4
4
6
8
2
0
3
2
7
4
3
p
哦
s
C
_
A
_
0
0
3
7
9
p
d
.
/
F
乙
y
G
你
e
s
t
t
哦
n
0
7
S
e
p
e
米
乙
e
r
2
0
2
3