Assessing the Effect of Early Visual Cortex Transcranial
Magnetic Stimulation on Working
Memory Consolidation
Amanda E. van Lamsweerde and Jeffrey S. 约翰逊
抽象的
■ Maintaining visual working memory ( VWM) 陈述
recruits a network of brain regions, including the frontal, 后部
顶叶, and occipital cortices; 然而, it is unclear to what ex-
tent the occipital cortex is engaged in VWM after sensory encoding
is completed. Noninvasive brain stimulation data show that stim-
ulation of this region can affect working memory (WM) during the
early consolidation time period, but it remains unclear whether it
does so by influencing the number of items that are stored or their
precision. In this study, we investigated whether single-pulse
transcranial magnetic stimulation (spTMS) to the occipital cortex
during VWM consolidation affects the quantity or quality of VWM
陈述. In three experiments, we disrupted VWM consol-
idation with either a visual mask or spTMS to retinotopic early vi-
sual cortex. We found robust masking effects on the quantity of
VWM representations up to 200 msec poststimulus offset and
较小, more variable effects on WM quality. 相似地, spTMS
decreased the quantity of VWM representations, but only when
it was applied immediately following stimulus offset. Like visual
masks, spTMS also produced small and variable effects on WM
precision. The disruptive effects of both masks and TMS were
greatly reduced or entirely absent within 200 msec of stimulus
抵消. 然而, there was a reduction in swap rate across all time
intervals, which may indicate a sustained role of the early visual
cortex in maintaining spatial information. ■
介绍
Research examining the neural bases of working memory
( WM) has suggested that, whereas early sensory cortex is
primarily engaged during stimulus encoding, 形式-
tion and maintenance of WM representations are most
likely mediated by sustained activation in “higher-level”
areas such as the parietal and frontal cortex (Postle,
2006; 徐 & Chun, 2006; Todd & Marois, 2004; 考特尼,
Ungerleider, Keil, & Haxby, 1997; Goldman-Rakic, 1995).
More recent evidence, 然而, has demonstrated that
stimulus attributes, such as the color or direction of mo-
tion of a stimulus, can be decoded from delay period ac-
tivity in sensory cortex using pattern classifiers (Emrich,
Riggall, LaRocque, & Postle, 2013; Ester, 安德森, &
Serences, 2013; 哈里森 & Tong, 2009; Serences, Ester,
沃格尔, & Awh, 2009), and the accuracy of these classifiers
has been correlated with the precision of stored informa-
的 (Emrich et al., 2013). These data suggest that early
sensory areas may play an important role in the forma-
tion and short-term retention of WM representations, 在
addition to their well-established role in perceptual en-
编码. The correlational nature of the majority of these
学习, 然而, has made it difficult to draw strong in-
ferences regarding the causal role of early sensory areas
North Dakota State University
© 2017 麻省理工学院
in WM functions. To address this, several studies have
used noninvasive brain stimulation methods to demon-
strate a causal link between early sensory areas and WM
(Makovski & Lavidor, 2014; van de Ven & Sack, 2013; van
de Ven, Jacobs, & Sack, 2012; 卡塔内奥, Vecchi, Pascual-
莱昂内, & Silvanto, 2009).
例如, van de Ven et al. (2012) used single-pulse
transcranial magnetic stimulation (spTMS) to examine
the contribution of the early visual cortex to visual work-
ing memory ( VWM). Results showed that spTMS of the
visual cortex produced a retinotopically specific disrup-
tion of performance on a shape change detection task
when applied 200 msec poststimulus offset (350 毫秒
poststimulus onset), but not when applied at 100 或者
400 毫秒. 此外, this disruption was only observed
in a high-load condition in which three shapes were
remembered; no effects of spTMS were observed when
only a single item was remembered. 至关重要的是, the TMS-
related decrease in performance occurred at the same time
period as visual mask-related decreases in performance
observed in a second experiment. The authors concluded
from these results that occipital-cortex-mediated VWM con-
solidation occurs early during the retention interval (看
also Cattaneo et al., 2009) and is both topographically orga-
nized and highly capacity limited. Underlying this conclu-
sion is the idea that spTMS and visual masks disrupt
performance in an all-or-none fashion by interfering with
认知神经科学杂志 29:7, PP. 1226–1238
土井:10.1162/jocn_a_01113
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
9
/
7
2
9
1
/
2
7
2
/
6
1
1
2
9
2
5
6
2
/
7
1
5
7
7
8
哦
6
C
4
n
0
_
3
A
/
_
j
0
哦
1
C
1
n
1
3
_
A
p
_
d
0
1
乙
1
y
1
G
3
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
F
t
.
/
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
visual cortex activity that is critical for the formation of
stimulus representations in WM. 那是, TMS prevents
one or more items from being successfully consolidated
and maintained in WM. This possibility appears consistent
with evidence suggesting that the functional effect of TMS
is to interrupt ongoing neural activity, rather than to add
random noise to the signal (哈里斯, Clifford, & Miniussi,
2008; but see Abrahamyan, Clifford, Arabzadeh, & 哈里斯,
2011; 施华蔻, Silvanto, & Rees, 2011). 其他
possibility, 然而, is that both forms of disruption
produce their effects by reducing mnemonic precision
(IE。, the quality of information in VWM), 而不是
preventing the consolidation or maintenance of some
项目 (IE。, the quantity of information in VWM). This could
happen if the disruption caused by TMS (or visual masks)
prevents the further accumulation of high-resolution
stimulus information for a subset of the remembered
项目, rather than having an all-or-none effect on
consolidation. Previous TMS studies, which have relied
exclusively on variants of the change detection paradigm,
are not well suited to addressing this possibility, 因为
they do not allow the quantity and quality of WM represen-
tations to be separately estimated.
In this study, we sought to clarify the functional role of
early visual cortex in WM by applying single pulses of
TMS to retinotopic visual cortex while participants per-
formed a cued recall WM task. This task required partic-
ipants to remember three colors presented in either the
lower left or lower right visual field, randomly deter-
mined by a spatial cue presented at the beginning of each
审判 ( 沃格尔 & Machizawa, 2004), and to estimate a given
remembered color at test by selecting its value from a
continuous representation of the color space (张 &
Luck, 2008; Wilken & Ma, 2004). Critically, 虽然
visual field location of memory stimuli varied from trial to
审判, the stimulated hemisphere was held constant. 阿迪-
理论上, the timing of TMS relative to the offset of the
memory display was varied randomly across trials. 这些
design elements allowed us to assess the topographic
and temporal specificity of TMS effects on performance
(as in van de Ven et al., 2012). To assess the functional
contribution of early visual cortex activity to WM, 我们
adopted a mixture modeling approach (Bays, Catalao, &
Husain, 2009; 张 & Luck, 2008) that attributes recall
response errors to three different underlying sources: (1)
response variability, measured as the standard deviation
(标清) of a circular Gaussian (von Mises) distribution cen-
tered on the target color, a proxy for memory quality; (2)
the probability of uniform responding (IE。, guessing,
denoted as g), proposed to reflect the proportion of trials
on which no information about the cued item is present
in WM, a proxy for WM capacity; 和 (3) nontarget
responses, in which participants mistakenly report the
color of one of the un-cued items at test.
Our initial hypothesis was that TMS would produce
retinotopically specific effects on estimates of the SD
(rather than g). This was based on previous findings,
suggesting that the ability to decode stimulus identity
from patterns of activation in early visual cortex is predic-
tive of SD (Emrich et al., 2013), and on theories, 建议-
ing that the representation of high-precision featural
information in WM involves the recruitment by attention
of early visual areas involved in the initial processing of that
信息 (D’Esposito, 2007; Postle, 2006; Pasternak &
Greenlee, 2005). 然而, a previous study examining
the effects of visual masks on recall performance (张
& Luck, 2008) revealed selective effects on random
responding, rather than SD. If TMS influences WM through
a similar mechanism, it could also be expected to affect the
rate of random responding. 最后, if retinotopic early
visual cortex plays a functional role in both the initial en-
coding as well as maintenance of information in VWM,
we expected TMS pulses to continue to exert an influence
on behavior across each of the time points tested (0, 100,
或者 200 msec poststimulus offset).
Results showed that, contrary to our initial prediction,
TMS-related declines in performance were most promi-
nently reflected in a decrease in g, although smaller ef-
fects were also observed on SD. 此外, TMS
produced retinotopically specific improvements in per-
formance; the likelihood of making a swap error was
reduced for targets contralateral to the stimulated
hemisphere. In a corresponding experiment, we found
a similar pattern of disruptive effects when visual masks
were used to interfere with performance; visual masks
produced a significant increase in g and smaller more var-
iable effects on SD. Visual masks had no effect on swap
速度. Although the disruptive effects of visual masks were
larger and extended further into the delay than the TMS-
induced effects, in both cases the effects were either ab-
sent or considerably reduced by 200 msec poststimulus
(350 msec after stimulus onset).
EXPERIMENT 1: EFFECTS OF VISUAL MASKS
ON CHANGE DETECTION
Before examining the effects of masks and TMS on recall
表现, we conducted an initial experiment to de-
termine the timing of masking effects in the context of
change detection, the VWM task utilized by van de Ven
等人. (2012). In a change detection task, 参与者
are asked to remember a small set of simple objects
(例如, colored squares, oriented bars, abstract shapes,
ETC。) across a short retention interval (∼800 msec to
3 秒), by asking them to judge whether the items in a
test display are the same as or different than the items
they saw originally (Luck & 沃格尔, 1997). In a shape
change detection task, van de Ven et al. (2012) 成立
masking effects at 200 msec poststimulus offset (350 毫秒
post onset), but not at 100 或者 400 msec poststimulus off-
放. 相比之下, 沃格尔, Woodman, and Luck (2006) 成立
that visual masks disrupted performance on a color
change detection task until approximately 183 msec after
the onset of the memory display (33 msec poststimulus
van Lamsweerde and Johnson
1227
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
9
/
7
2
9
1
/
2
7
2
/
6
1
1
2
9
2
5
6
2
/
7
1
5
7
7
8
哦
6
C
4
n
0
_
3
A
/
_
j
0
哦
1
C
1
n
1
3
_
A
p
_
d
0
1
乙
1
y
1
G
3
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
t
.
F
/
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
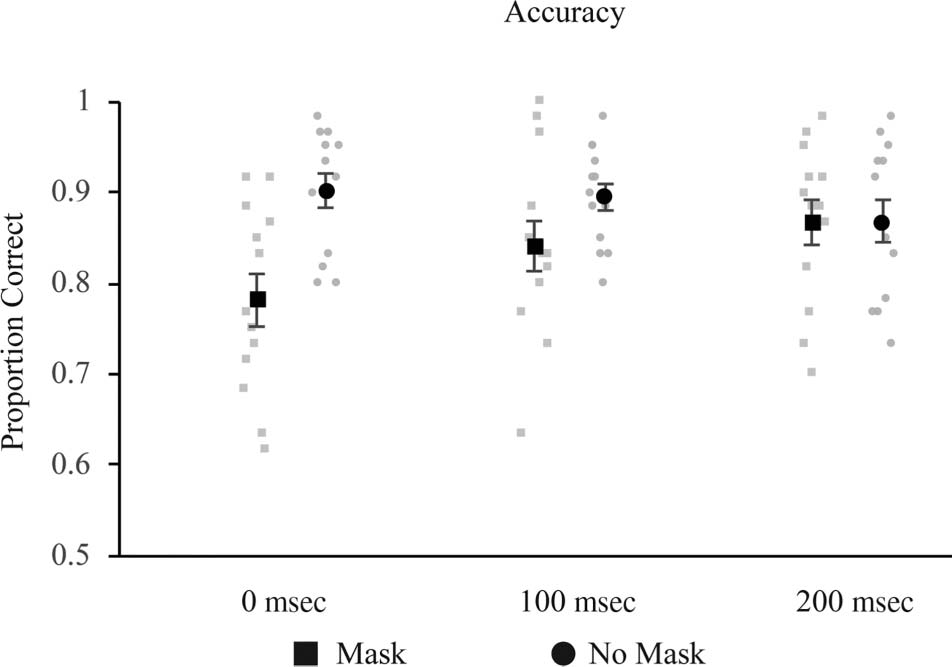
抵消). 所以, in Experiment 1, we utilized a change
detection task with masks at three time points: 0, 100, 或者
200 msec following the offset of the stimulus display. 这
方式, we could determine whether mask timings differ for
change detection versus recall tasks before administering
spTMS in a cued recall task (实验 3).
方法
参加者
Thirteen undergraduate students (10 女性, average
age = 24.5 年) participated in this experiment for
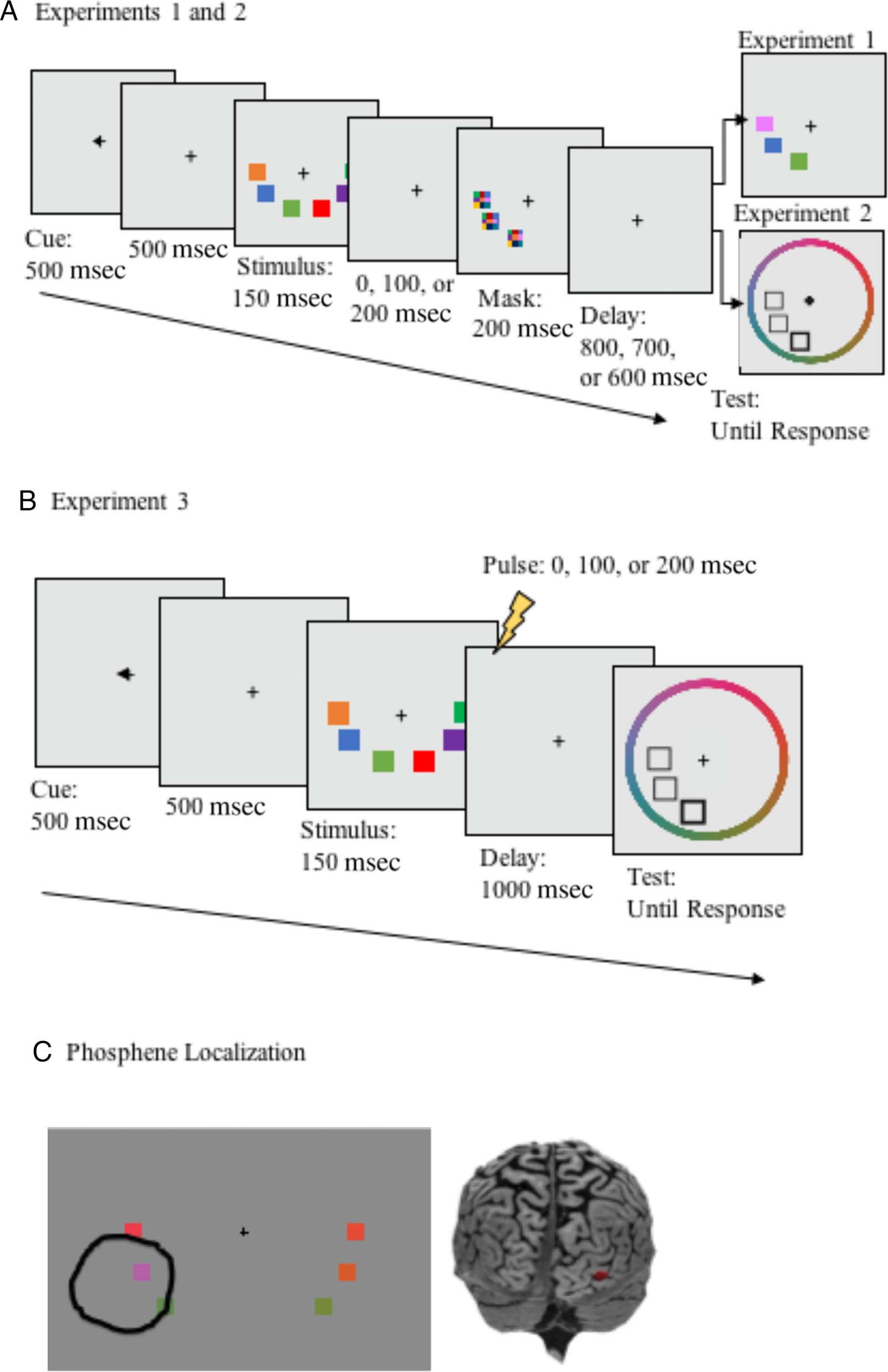
monetary compensation ($10/小时). All participants had normal or corrected-to-normal visual acuity and normal color vision and provided informed consent before participation. Study protocols for this and all subsequent experiments were approved by the North Dakota State University institutional review board. Stimuli Stimulus presentation and response recording was con- trolled by a PC running Matlab (The Mathworks, 公司, Natick, 嘛) with Psychophysics Toolbox extensions (Brainard, 1997; Pelli, 1997). Stimulus displays contained three colored squares subtending 1.23° × 1.23° of visual angle on either side of fixation at a viewing distance of 70 厘米 (见图 1). Individual colors were selected at random from a set of 180 colors equally distributed in CIELAB (1976) color space (centered at CIE L*a*b* coor- dinates: L = 70, A = 28, B = 12). All objects were pre- sented in the lower left and right visual hemifields, equally spaced on an invisible circle centered at fixation with a radius of 6.31° of visual angle and squares within each hemifield were spaced 3.3° from each other, center to center. Procedure The task is shown in Figure 1A. Each trial began with a precue (500 毫秒), directing participants to attend to either the left or right visual hemifield, followed by a 500-msec fixation screen, then the stimulus display (150 毫秒), and then a 1000-msec delay. During the de- 躺着, a pattern mask (composed of randomly selected colors from the possible colorspace) flashed for 200 msec over the spatial location occupied by each object on the left side of the screen. 所以, when cued to attend to the left visual hemifield, the memory representations were masked, but when cued to attend to the right visual hemifield, memory representations were unmasked (the left side alone was masked to correspond with the TMS experiment, in which stimulation was always applied to the right hemisphere). Masks appeared at delays of 0, 100, 或者 200 msec after the offset of the stimulus display (randomly intermixed). Because the duration of the stim- ulus display was 150 毫秒, these timings correspond to SOAs between the onset of the stimulus display and the onset of the mask of 150, 250, 和 350 毫秒, 重新指定- 主动地. These timings were selected on the basis of previ- ous findings related to the time course of consolidation of color stimuli in VWM, which revealed that visual masks continue to disrupt memory performance at SOAs up to183 msec (see Vogel et al., 2006, 实验 2). 福尔- lowing the delay, the test display was presented, which was either identical to the memory display or contained a color change in one of the objects. When a color changed, the new color was selected at random from the total set of possible colors, with the constraint that it must be different from the sample stimulus by at least 20° in color space. Participants indicated whether the test display was the same as or different than the stimulus display by pressing one of two keys on their keyboards. Participants completed 60 experimental trials in each condition (30 改变, 30 no change; 360 trials total) divided into 10 blocks with an even number of each trial type, plus a block of 36 practice trials. Results A mask-related decrease in performance was found at 0 和 100 msec poststimulus offset, 但不是 200 毫秒 (见图 2). This was supported by a 2 (Mask: mask, no mask) × 3 (Timing: 0, 100, 200 毫秒) repeated-measures ANOVA. Where the assumption of sphericity was violated, a Greenhouse–Geisser correction was applied. In this and all subsequent experiments, post hoc tests were cor- rected with the Holm–Bonferroni correction for family- wise error. With this correction, comparisons are ranked from smallest to largest p value and are compared against a critical p value, calculated separately for each hypothesis using the formula P(k) > α/m + 1 − k, where P refers to the obtained p value, α represents the selected criterion for rejecting the null hypothesis (.05 in the present case), m is an index representing the order of p values for each hypothesis tested, from lowest to highest, and k is the minimal index for which the obtained p value does not exceed the significance criterion (α/m + 1 − k; Holm, 1979). In all cases, three comparisons were made. 因此, the threshold for significance for the smallest p value is p < .0167, the second largest p < .025, and the largest p value p < .05. The ANOVA revealed a significant effect of Mask, F(1, 12) = 12.83, p = .004, ηp 2 = 0.52, no main effect of Tim- ing, F(1.33, 15.924) = 2.33, p = .14, ηp 2 = 0.16, and a sig- nificant Mask × Timing interaction, F(2, 24) = 20.62, p < .001, ηp 2 = 0.63. Post hoc t tests revealed a significant dif- ference between the mask and no-mask conditions at 0 msec, t(12) = −7.39, p < .001 (critical p = .0167), a marginally significant effect at 100 msec, t(12) = −2.53, p = .03 (critical p = .025), and no effect at 200 msec, t(12) = .01, p = .99 (critical p = .05). 1228 Journal of Cognitive Neuroscience Volume 29, Number 7 D o w n l o a d e d f r o m l l / / / / j t t f / i t . : / / h t t p : / D / o m w i n t o p a r d c e . d s f i r o l m v e h r c p h a d i i r r e . c c t . o m m / j e d o u c n o / c a n r a t r i t i c c l e e - p - d p d 2 f 9 / 7 2 9 1 / 2 7 2 / 6 1 1 2 9 2 5 6 2 / 7 1 5 7 7 8 o 6 c 4 n 0 _ 3 a / _ j 0 o 1 c 1 n 1 3 _ a p _ d 0 1 b 1 y 1 g 3 u . e p s t d o f n b 0 y 7 S M e I p T e m L i b b e r r a 2 r 0 2 i 3 e s / j t . f / u s e r o n 1 7 M a y 2 0 2 1 D o w n l o a d e d f r o m l l / / / / j f / t t i t . : / / h t t p : / D / o m w i n t o p a r d c e . d s f i r o l m v e h r c p h a d i i r r e . c c t . o m m / j e d o u c n o / c a n r a t r i t i c c l e e - p - d p d 2 f 9 / 7 2 9 1 / 2 7 2 / 6 1 1 2 9 2 5 6 2 / 7 1 5 7 7 8 o 6 c 4 n 0 _ 3 a / _ j 0 o 1 c 1 n 1 3 _ a p _ d 0 1 b 1 y 1 g 3 u . e p s t d o f n b 0 y 7 S M e I p T e m L i b b e r r a 2 r 0 2 i 3 e s / j f t / . u s e r o n 1 7 M a y 2 0 2 1 Figure 1. Tasks used in Experiments 1–3. In Experiment 1 (A), participants detected changes in a single color between stimulus and test displays; visual masks were displayed 0, 100, or 200 msec into the 1000-msec delay period. In Experiment 2, the task was identical to Experiment 1, except that a recall task was used, rather than change detection. In Experiment 3 (B), the task was identical to Experiment 2, except that instead of a visual mask, spTMS was applied at 0, 100, or 200 msec into the delay period. Brain regions targeted for stimulation (C) were selected before the experiment with a phosphene localization procedure. This figure shows the targeted brain region for one participant, based on the phosphene area circled by that participant. Phosphene localization response is shown overlayed on top of a sample stimulus, for illustrative purposes only. The stimulus display was not present during phosphene localization. van Lamsweerde and Johnson 1229 change detection. The task was identical to the change detection task up until the presentation of the test dis- play. In this task, the participant is asked to recall a par- ticular cued item by selecting its value from a continuous representation of the feature space (Zhang & Luck, 2008; Wilken & Ma, 2004). We then examined the impact of visual masks on both total error (the absolute difference between the recalled and actual color across trials) and on different putative sources of error using a mixture modeling approach (Bays et al., 2009; Zhang & Luck, 2008), which allows both the quantity and quality of VWM representations to be separately estimated. Methods Participants Twenty-two undergraduate and graduate students (age M = 22.09 years, 19 women) participated in this experi- ment for either course credit or monetary compensation ($10/ hr). All participants had normal or corrected-to-
normal vision and normal color vision.
Stimuli and Procedure
The stimuli and procedure were identical to Experiment 1,
with the exception that a cued recall rather than change
detection test was used (Figure 1B). The test display
contained a filled white square at the location of a ran-
domly selected test item, with empty white box place-
holders in the locations of the nontargets in the attended
hemifield only. These boxes were surrounded by a color
wheel centered at fixation with a radius of 8.13° of visual
angle, which contained all possible colors equally distribut-
ed in steps of 2° and was randomly rotated on each trial so
that participants could not generate anticipatory responses
before the onset of the test display. Participants were in-
structed to report the color of the cued item by clicking
on the color wheel using the computer mouse (see
Figure 1). As participants moved the mouse around the
color wheel, the cued square was filled with the selected
color. Once participants made a response, the target square
was filled with the response color, and a border of the
correct color was added so that participants could compare
their response to the correct one. Additional feedback was
provided in the form of a black bar that appeared on the
outside the color wheel, marking the correct color. Partici-
pants completed 100 trials in each condition (600 trials
total), evenly distributed across 10 experimental blocks,
plus one practice block of 60 trials.
Modeling Recall Response Distributions
Participants’ data were analyzed using the MemToolbox
(Suchow, Brady, Fougnie, & Alvarez, 2013). To analyze
performance in the recall task, we made use of analytic
techniques proposed by Zhang and Luck (2008) and Bays
Figure 2. Results from Experiment 1. Mask-related decreases in
performance were found at 0 and 100 msec, but not 200 msec.
Discussion
Visual masks produced a temporally graded pattern of
disruption of color change detection, producing a large
effect when the mask appeared immediately after stimu-
lus offset, a marginal effect 100 msec later, and no effect
at all by 200 msec (SOA between onset of stimulus and
onset of mask = 350 msec), similar to the timing effects
found by Vogel et al. (2006). This is in contrast to van de
Ven et al. (2012), who found masking-related decreases
in performance only at 200 msec (sample-mask onset
SOA = 350 msec), but not at either earlier (100 msec)
or later (400 msec) time points. The reasons for this dis-
crepancy are unclear. van de Ven et al. suggest that the
timing of the interference effect observed in their exper-
iment could be due to interference with a later-occurring
sweep of feedback input from higher-level areas to the
visual cortex, which, they propose, may be critical for suc-
cessful consolidation. Differences in the timing of mask-
ing effects between our studies could potentially be
explained if we assume that the timing and/or necessity
of such feedback interactions for consolidation differ de-
pending on the specific stimuli used (e.g., complex
shapes vs. colors). Determining whether this is in fact
the case is beyond the scope of this study and will re-
quire further research, ideally using neuroimaging
methods that make it possible to carefully track the pat-
terns of activity involved in task performance. Of greater
relevance, the results of Experiment 1 provide a range of
sample-mask SOAs that were utilized in Experiment 2 in
the context of a recall WM task. Use of recall, rather than
change detection, allowed us to determine whether visu-
al masks influence g (as a proxy for capacity) or SD (as a
proxy for mnemonic precision).
EXPERIMENT 2: EFFECTS OF VISUAL MASKS
ON RECALL PERFORMANCE
Experiment 2 was identical to Experiment 1, with the ex-
ception that a cued recall test was used, rather than
1230
Journal of Cognitive Neuroscience
Volume 29, Number 7
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
7
2
9
1
/
2
7
2
/
6
1
1
2
9
2
5
6
2
/
7
1
5
7
7
8
o
6
c
4
n
0
_
3
a
/
_
j
0
o
1
c
1
n
1
3
_
a
p
_
d
0
1
b
1
y
1
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
et al. (2009), in which recall response distributions are
assumed to reflect a mixture of response types drawn
from different distributions. According to the logic of
this approach, when the cued item is successfully
remembered, recall responses are drawn from a circular
Gaussian (i.e., Von Mises) distribution, in which the
mean indicates how close recall responses were on
average to the actual target value across trials, and the
standard deviation reflects the precision (quality) of the
recalled items. If the item was not successfully stored,
recall responses are assumed to be drawn from a
uniform distribution in which individual colors are
selected equiprobably from the color wheel; this “guess
rate” can then be used to estimate the quantity of items
stored in VWM. Finally, accurate performance on this task
necessitates keeping track of which colors appeared in
each location in the original memory display. A failure
to do so can result in a third type of recall response in
which one of the noncued memory items is recalled
instead of the cued item, known as a “swap error”
(Bays et al., 2009). Therefore, this method makes it
possible to separately estimate the quantity and quality
of stored information, as well as the likelihood of
mistaking the cued item for one of the other items in
WM. For each participant, we compared goodness of fit
between the three-component variant of the mixture
model and the two-component model proposed by
Zhang and Luck (2008), which does not include swap
errors, using the log likelihood and the corrected
Akaike information criteria (cAIC). For all model
comparisons, the three-component model provided the
superior fit for the majority of participants (100% and
86% for the log likelihood and cAIC, respectively). For
both the log likelihood and cAIC, we compared the
computed scores for each model with a paired sample
t test and report the mean difference between the scores
as the two component score minus the three component
score. The three component model provided a signifi-
cantly superior fit for both the cAIC, t(21) = −.30, p <
.001 (mean difference = 13.53, SD = 14.76), and log
likelihood, t(21) = −5.49, p < .001 (mean difference =
−8.07, SD = 6.89). Therefore, the three-component
model was used for all analyses.
Results
Analysis of Absolute Error
The present experiment manipulated the timing of visual
masks and whether the attended visual field location was
masked or unmasked. To examine the effects of mask
timing and presence/absence on recall performance, we
first performed a two-way (2 Mask × 3 Timing) repeated-
measures ANOVA on the absolute error (absolute differ-
ence between the recalled and the actual target color).
This revealed a significant main effect of Mask, F(1, 21) =
45.78, p < .0001, ηp
2 = 0.69, a significant main effect of
Timing, F(2, 42) = 27.99, p < .0001, ηp
2 = 0.57, and a
significant Mask × Timing interaction, F(2, 42) = 14.145,
p < .0001, ηp
2 = 0.40. Post hoc t tests revealed significant
mask-related elevations in absolute error at each sample-
mask timing (all ps < .001), although the effect grew
substantially smaller at longer sample-mask delays (see
Figure 3A).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
7
2
9
1
/
2
7
2
/
6
1
1
2
9
2
5
6
2
/
7
1
5
7
7
8
o
6
c
4
n
0
_
3
a
/
_
j
0
o
1
c
1
n
1
3
_
a
p
_
d
0
1
b
1
y
1
g
3
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
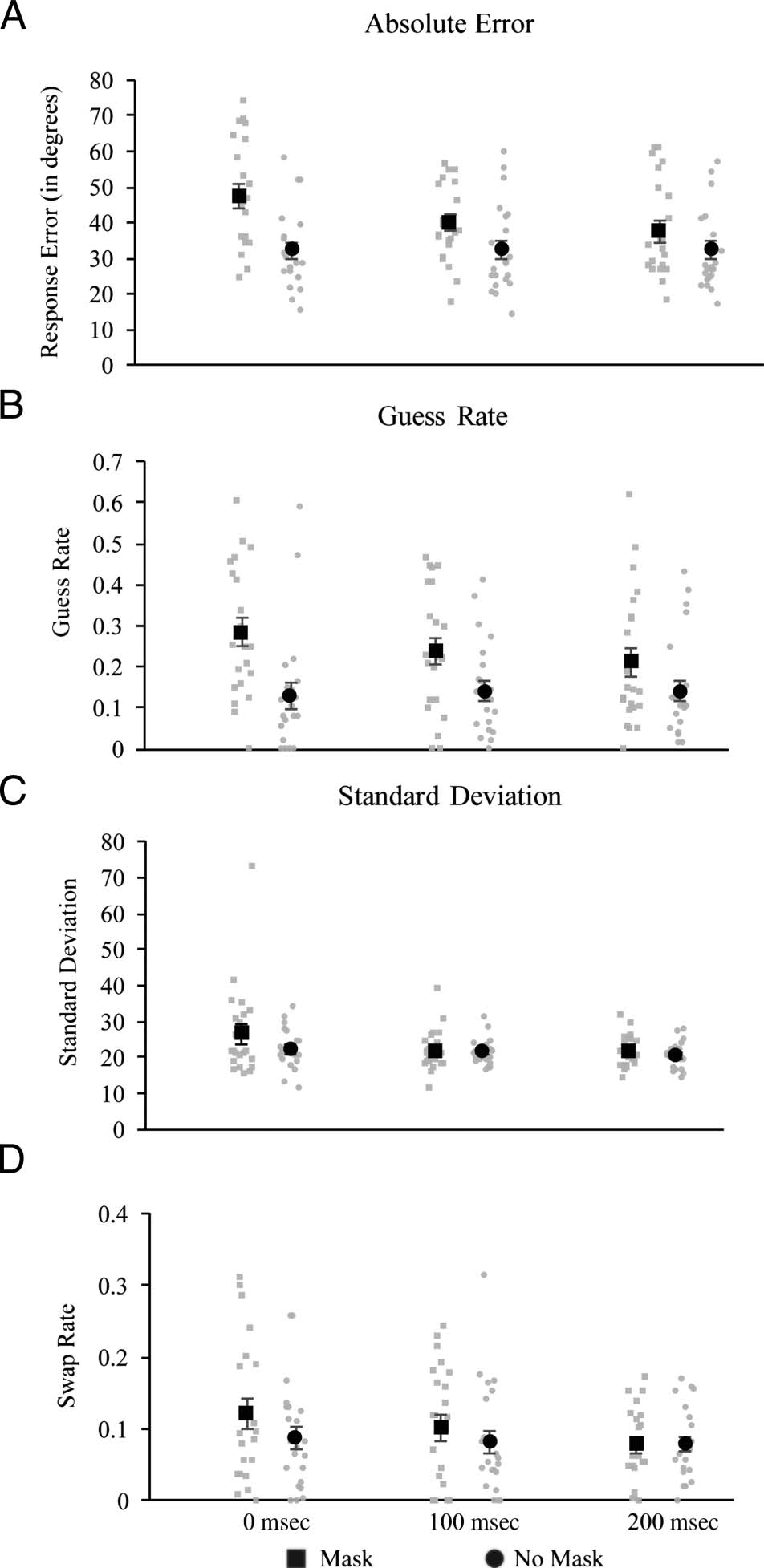
Figure 3. Results from Experiment 2. Mask-related decreases in (A)
absolute error were found at all time periods. In the mixture model
parameters, mask-related decreases were found at all time points for
guess rate (B), with a decreasing effect of mask with increased time.
Standard deviation was overall greater with a mask (C) and no effects
were found on swap rate (D).
van Lamsweerde and Johnson
1231
Analysis of Mixture Model Fits
Results of the mixture model analysis can be seen in
Figure 3B–D. Differences between conditions were
assessed with separate 2 (Mask: mask, no mask) × 3
(Timing: 0, 100, 200) repeated-measures ANOVAs con-
ducted for each parameter (g, SD, swap rate). When the
assumption of sphericity was violated, a Greenhouse–
Geisser correction was applied.
Guess rate. The results indicate a masking-related in-
crease in guess rate at all three time points, although
effects tended to decrease over time. Confirming this pat-
tern, ANOVA revealed a main effect of Mask, F(1, 21) =
19.29, p < .001, ηp
2 = 0.48, no effect of Timing, F(2, 42) =
1.14, p = .33, ηp
2 = 0.05, and a significant Mask × Timing
interaction, F(2, 42) = 3.72, p = .03, ηp
2 = 0.15. Post hoc
t tests revealed a significant increase in guess rate as a result
of the mask at 0 msec, t(21) = 4.63, p < .001 (critical p =
.0167), and 100 msec, t(21) = 3.56, p = .002 (critical
p = .025), and a smaller effect at 200 msec, t(21) =
2.09, p = .049 (critical p = .05).
Standard deviation. The ANOVA only revealed a main
effect of Mask, F(1, 21) = 4.39, p = .049, ηp
2 = 0.17, with
a marginal effect of Timing, F(1.55, 32.49) = 3.20, p =
.065, ηp
2 = 0.13, and no significant interaction, F(1.51,
31.77) = 1.58, p = .22, ηp
2 = 0.07.
Swap rate. No masking effects on swap rate were found.
The ANOVA revealed no effects of Mask, F(1, 21) = 2.73,
p = .11, ηp
2 = 0.12, Timing, F(2, 42) = 2.43, p = .10,
ηp
2 = 0.10, or a Mask × Timing interaction, F(2, 42) =
1.00, p = .38, ηp
2 = 0.05.
Discussion
Using a recall test, Experiment 2 revealed the presence of
masking effects at all tested intervals, although the mag-
nitude of the effect decreased over time. Although the
greater number of participants in Experiment 2 makes
it difficult to directly compare these results to Experi-
ment 1, this result suggests that the recall task may have
greater sensitivity to reveal masking effects than the
change detection task. The time required to fully consol-
idate VWM representations may therefore be somewhat
longer than suggested by studies examining this issue
using the change detection task ( Vogel et al., 2006). In
keeping with the findings of Zhang and Luck (2008),
the masks in this experiment primarily influenced the
likelihood of generating a guess-like response. However,
we also observed a small, mask-related increase in SD
that did not vary based on mask timing. Thus, these find-
ings are broadly consistent with the proposal that encod-
ing in VWM is an all-or-none process, as opposed to a
process characterized by a gradual accumulation of fea-
tural information over time (Zhang & Luck, 2008).
EXPERIMENT 3: EFFECTS OF spTMS OF
RETINOTOPIC VISUAL CORTEX ON
RECALL PERFORMANCE
The goal of Experiment 3 was to determine whether TMS
affects the quantity and quality of information in VWM in a
manner similar to the visual masks used in Experiment 2.
The same procedure was used as in Experiment 2, ex-
cept that instead of visual masks, spTMS was applied
to retinotopic visual cortex at variable delays relative to
stimulus offset. It was expected that disruptions in per-
formance would occur when participants encoded infor-
mation presented in the visual field contralateral to the
stimulated hemisphere (as in van de Ven et al., 2012);
contralateral stimulation trials therefore served the same
function as the “mask” trials in Experiments 1 and 2. The
trials in which participants attended to the hemifield ip-
silateral to stimulation (i.e., right hemifield during right-
occipital stimulation) served as a hemispheric control.
Although it is common to also include a sham control
condition in TMS experiments, the use of a control
hemisphere has the advantage that (1) trials probing
the control and target hemispheres are randomly inter-
mixed throughout the session, ensuring that the state of
the participant is roughly equivalent across conditions,
and (2) the tactile sensation produced by the coil’s dis-
charge is identical across conditions (acoustic sensations
were largely eliminated by the use of masking noise dur-
ing the session). For these reasons, a sham control was
not included.
Methods
Participants
Twenty-two participants recruited from the North Dakota
State University undergraduate and graduate population
completed the experiment for paid compensation
($20/hr). All participants were between the ages of 18
and 35 years, (age M = 22 years, 15 women), were right-
handed, reported normal or corrected-to-normal visual
acuity, and had normal color vision (as established with
the Ishihara color vision test). Participants gave informed
consent and were screened for the presence of neurolog-
ical and psychiatric conditions and other risk factors
related to the application of both MRI and TMS before par-
ticipation (based on guidelines for TMS safety set forth in
Rossi, Hallett, Rossini, & Pascual-Leone, 2009). One partic-
ipant was excluded because of very low performance
(guess rate > 2 Sds below the mean across all conditions),
bringing the final N to 21. The results do not change when
this participant is included.
TMS Targeting and Stimulation
TMS was delivered with a Magstim Super Rapid 2 mag-
netic stimulator fit with a focal bipulse, figure of eight
70-mm stimulating coil (Magstim, Whitland, 英国). TMS
1232
认知神经科学杂志
体积 29, 数字 7
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
9
/
7
2
9
1
/
2
7
2
/
6
1
1
2
9
2
5
6
2
/
7
1
5
7
7
8
哦
6
C
4
n
0
_
3
A
/
_
j
0
哦
1
C
1
n
1
3
_
A
p
_
d
0
1
乙
1
y
1
G
3
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
t
/
F
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
targeting and online guidance was achieved using a
Visor2 neuronavigation system (Advanced Neuro Tech-
科学, Enschede, 荷兰人) that uses infrared-
based frameless stereotaxy to map the position of the coil
and the participant’s head within the reference space of
the individual’s high-resolution anatomical MRI. Whole-
brain T1-weighted anatomical MRI scans were acquired
with a GE Signa HD 1.5-T MRI scanner for each partici-
pant before participation (206 axial slices, with a resolu-
tion of 1 毫米). Throughout the experiment, 参与者
listened to constant masking noise played through a pair
of inserted earplugs. The volume of the masking noise,
which never exceeded 90 分贝, was adjusted immediately
before the experimental session for each participant until
the “click” produced by the discharge of the TMS coil
could no longer be heard ( 约翰逊, Kundu, Casali, &
Postle, 2012; Esser et al., 2006).
Phosphene Localization and Thresholding
Phosphene localization procedures and all subsequent
stimulus presentation and response recording were con-
trolled by a PC running Matlab with Psychophysics Tool-
box extensions (Brainard, 1997; Pelli, 1997). The right
primary visual cortex was first identified and targeted
on the basis of individual anatomy, and target coordi-
nates were further refined by determining the coil posi-
tion and orientation that elicited visual phosphenes in a
particular region of visual space (van de Ven et al., 2012).
要做到这一点, participants fixated a central white cross pre-
sented against a black background while single pulses of
TMS were administered to visual cortex at 70% of stimu-
lator output. Coil position and orientation were then ad-
justed until reproducible phosphenes were visible in the
hemifield contralateral to stimulation. Once phosphenes
were induced, participants used a computer mouse to
draw a circle around the region in the visual field where
the phosphenes appeared. Responses were recorded as
both an image file and as the x–y coordinates of all
mouse locations. The coil position was then adjusted un-
til reliable phosphenes were produced overlapping or ad-
jacent to the location of the sample display items (为了
stimulus positions, see Experiment 1 方法). Phos-
phenes were considered to be elicited when the follow-
ing criteria were met: Phosphenes could be localized in
both left and right hemispheres, phosphenes were elicit-
ed with eyes shut, and phosphenes moved with fixation
(Kammer, 1998). Following phosphene localization,
phosphene thresholds were established. Stimulator in-
tensity was reduced by increments of 5%, 和 10 pulses
were delivered at each intensity until phosphene thresh-
老的, defined as the minimum intensity required to elicit
phosphenes 50% 当时的, was established. During the
实验, pulses were administered to the right hemi-
sphere at 110% of phosphene threshold. Average stimu-
lation intensity was at 67% of stimulator output (range =
57–82%). Postexperiment debriefing confirmed that TMS
at this intensity did not give rise to visible phosphenes
during performance of the color recall task, 其中
stimuli were presented against a light gray background
and attention was focused on the task.
Stimuli and Procedure
Stimuli and procedure were identical to Experiment 2,
except that TMS pulses, rather than visual masks, 是
applied at varying intervals relative to stimulus onset
(see Figure 1C). Viewing distance was held constant at
70 厘米, and head position was stabilized using a chinrest.
spTMS was administered either 0, 100, 或者 200 msec post-
stimulus offset.
结果
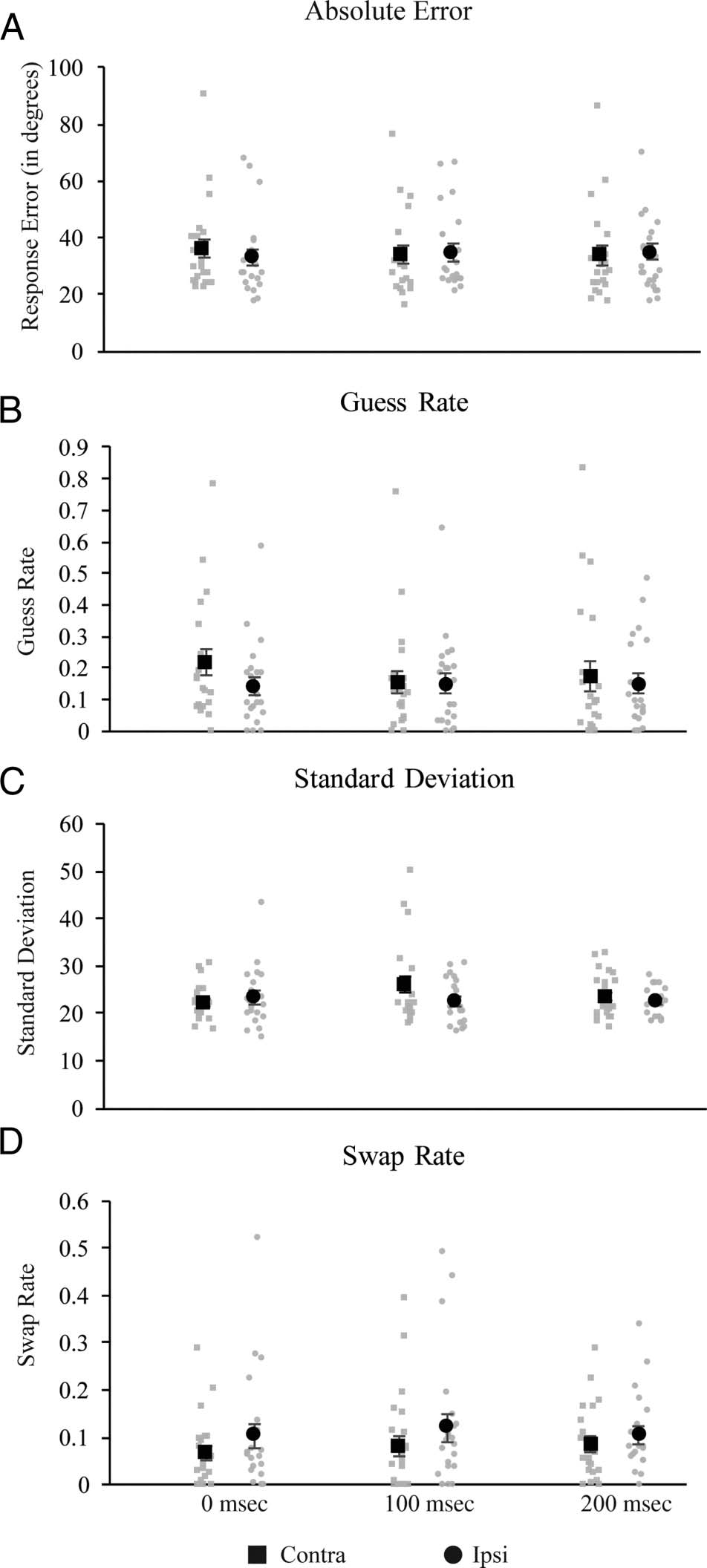
Analysis of Absolute Error
Absolute error for each combination of TMS side and tim-
ing can be seen in Figure 4A. As in Experiment 2, differ-
ences in absolute error across conditions were assessed
by a two-way (2 Mask × 3 Timing) repeated-measures
ANOVA. Contrary to the masking data, neither main
effect reached significance ( ps = .20 和 .23 for side
and timing, 分别). 然而, there was a trend to-
ward a significant Side × Timing interaction, F(2, 40) =
2.608, p = .086, ηp
2 = 0.12, with somewhat elevated
absolute error for targets contra versus ipsi to the stimu-
lated hemisphere when TMS was applied 0-msec post-
stimulus offset.
Analysis of Mixture Model Fits
As with Experiment 2, estimates of response errors were
derived from the three-component mixture model pro-
posed by Bays et al. (2009). Comparison of model fits
using the log likelihood and cAIC suggested that the
three-component model performed better than the
two-component model for the majority of participants
(100% 相对 82% for log likelihood and cAIC, 重新指定-
主动地). Average estimated model parameters are de-
picted in Figure 4B. As in Experiment 2, three separate
2 (Visual hemifield: contra vs. ipsi to stimulated hemi-
sphere) × 3 (TMS timing: 0, 100, 或者 200 msec after stim-
ulus display offset) repeated-measures ANOVAs were
实施, one each for g, 标清, and swap errors. For both
the log likelihood and cAIC, we compared the computed
scores for each model with a paired sample t test. 这
three component model provided a significantly superior
fit for both the cAIC, t(20) = 2.34, p = .03 (mean differ-
ence = 26.41, 标准差= 51.70), and log likelihood, t(20) =
−2.52, p = .02 (mean difference = −14.22, 标准差= 25.85).
Guess rate. A TMS-related increase in g was found when
stimulation was applied coincident with stimulus offset.
具体来说, the ANOVA revealed no main effects of
van Lamsweerde and Johnson
1233
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
9
/
7
2
9
1
/
2
7
2
/
6
1
1
2
9
2
5
6
2
/
7
1
5
7
7
8
哦
6
C
4
n
0
_
3
A
/
_
j
0
哦
1
C
1
n
1
3
_
A
p
_
d
0
1
乙
1
y
1
G
3
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
/
t
.
F
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
p = .02, which was just above the corrected threshold
value for significance (critical p = .0167). No effects of
TMS on guess rate were found at either 100 毫秒, t(20) =
.48, p = .63, 或者 200 毫秒, t(20) = .99, p = .33.
Standard deviation. The ANOVA revealed a main effect
of Stimulation side, F(1, 20) = 5.05, p = .04, ηp
2 = 0.20, 但
no main effect of Timing, F(2, 40) = 1.45, p = .25, ηp
2 =
0.07, and a marginal interaction, F(2, 40) = 2.80, p = .07,
ηp
2 = 0.12. 然而, post hoc t tests revealed no significant
differences in standard deviation between the contralateral
and ipsilateral hemifield when TMS was applied at 0, t(20) =
−1.40, p = .18, 100, t(20) = 2.14, p = .045 (critical p =
.0167), 或者 200 毫秒, t(20) = .83, p = .42.
有趣的是, analysis of swap rate revealed
Swap rate.
an overall decrease in swap errors for targets in the con-
tralateral hemifield, but this was not specific to a particu-
lar TMS timing. The ANOVA revealed a main effect of
Stimulation side, F(1, 20) = 5.44, p = .03, ηp
2 = 0.21,
but no effects of timing, F(2, 40) = .44, p = .65, ηp
2 =
0.02, and no interaction, F(2, 40) = 1.14, p = .33, ηp
2 =
0.05.
讨论
These data demonstrate that TMS of the early visual cor-
tex produced temporally and topographically specific ef-
fects on g, an index of the number of items that were
successfully stored in VWM. 具体来说, when applied
coincident with the offset of the stimulus display, spTMS
increased the likelihood of making a guess response for
targets appearing in the visual hemifield contralateral to
the stimulated cortex (IE。, the left visual field following
right visual cortex stimulation). Contrary to our predic-
系统蒸发散, TMS only produced a small effect on SD. 虽然
the effect on SD appeared to be specific to the 100-msec
状况, the TMS Timing × Visual Hemifield interac-
tion of the ANOVA was not significant, nor was the
post hoc t test looking at differences in SD at this interval.
此外, contrary to our predictions, 的影响
spTMS on both g and SD were entirely absent by
200 msec after stimulus offset (IE。, 350 msec poststimu-
lus onset). Contrasting with these disruptive effects,
spTMS also produced a temporally nonspecific reduction
in the likelihood of making swap errors for targets in the
contralateral hemifield (IE。, swap errors were less likely
when estimating the color of targets contra vs. ipsi to
the stimulated hemisphere). The effects of TMS on g
and SD are qualitatively similar to the findings of van
de Ven et al. (2012), who found that spTMS was no lon-
ger effective at later time points, when consolidation is
presumably already completed. 然而, these results
go beyond their findings by demonstrating temporally
and topographically specific effects on the parameters
of the mixture model, which may map on to qualitatively
distinct sources of error in recall WM tasks.
数字 4. Results from Experiment 3. A TMS-related increase in guess
rate was found in (A) absolute error and (乙) guess rate at 0 毫秒
poststimulus offset and an increase in standard deviation (C) was found
在 100 msec post offset. A TMS-related increase in swap rate was found
across all time points (D).
either stimulation side, F(1, 20) = 2.41, p = .14, ηp
2 =
0.11, or TMS timing, F(2, 40) = 1.33, p = .28, ηp
2 =
0.06, but there was a significant interaction between Tim-
ing and Stimulation side, F(2, 40) = 3.69, p = .03, ηp
2 =
0.16. Follow-up post hoc t tests revealed an increase in
guess rate for targets in the contralateral versus ipsilateral
hemifield when TMS was applied at 0 毫秒, t(20) = 2.50,
1234
认知神经科学杂志
体积 29, 数字 7
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
9
/
7
2
9
1
/
2
7
2
/
6
1
1
2
9
2
5
6
2
/
7
1
5
7
7
8
哦
6
C
4
n
0
_
3
A
/
_
j
0
哦
1
C
1
n
1
3
_
A
p
_
d
0
1
乙
1
y
1
G
3
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
/
.
t
F
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
GENERAL DISCUSSION
This study sought to clarify the functional relevance of
early visual cortex contributions to VWM. 要做到这一点,
we presented visual pattern masks or administered single
pulses of TMS to retinotopic visual cortex at different
time points relative to the offset of a stimulus display.
To determine the nature of the influence of masks and
TMS on the formation of VWM representations, parti-
cipants’ memory for the sample display colors was
assessed using either delayed recognition (change detec-
的) or cued recall, which made it possible to separately
estimate the number of items stored in WM and the pre-
cision of the stored information. Critically, 虽然
memory stimuli could be encoded from either the lower
left or lower right visual fields, as determined by a spatial
cue presented at the beginning of each trial, both masks
and TMS selectively targeted the lower left visual field.
因此, performance for stimuli encoded from the lower
right visual field served as a within-subject control (作为
in van de Ven et al., 2012). Across experiments, we ob-
served similar effects of masks and TMS on memory for
items appearing in the affected visual field, 虽然
effects of masks were generally more pronounced. Spe-
cifically, visual masks impaired change detection perfor-
曼斯 (实验 1) and increased the likelihood of
making a guess-like response (实验 2) for targets
appearing in the lower-left visual field, and spTMS in-
duced a similar increase in guessing for stimuli in the vi-
sual field contralateral to the targeted hemisphere
(实验 3). Both masks and TMS produced the larg-
est effects when applied coincident with stimulus offset,
with the disruption either entirely dissipating (Experi-
评论 1 和 3) or growing progressively smaller (Exper-
iment 2) the further into the retention interval it was
applied. Visual masks and TMS also produced small and
variable effects on the standard deviation of responses.
最后, TMS produced an unexpected decrease in swap
errors for targets appearing in the affected hemifield.
These findings have several implications for our under-
standing of the time course and functional relevance of
early visual cortex to VWM.
Regarding the question of whether masks (and TMS)
affect the number of items stored or their quality, 结果
of the mixture model analysis in Experiment 2 confirmed
the findings of Zhang and Luck (2008), who found that
masks primarily affect performance by reducing the prob-
ability that an item is encoded into VWM, rather than in-
creasing the variability of recall responses (标清). 相似地,
实验 3 revealed that TMS applied to early visual
cortex coincident with stimulus offset produced an in-
crease in guess rate for targets presented in the contra-
lateral visual field. Although there were also effects on
标清, these effects were more variable, and post hoc tests
suggested that they were not reliable at any of the tested
latencies. 因此, 放在一起, these results support the
proposal of van de Ven et al. (2012) that interfering with
activity in early visual cortex likely disrupts an ongoing
process of memory consolidation, suggesting a functional
role for early sensory areas in the initial formation of
VWM representations.
Although the mechanisms underlying the functional
effects of spTMS are not well understood, research by
Harris and colleagues (2008) suggests that the pattern
of disruption observed here and in the study of van de
Ven et al. (2012) may have been caused by a TMS-related
decrease in signal intensity, rather than an increase in ran-
dom image noise (Abrahamyan et al., 2011; 施华蔻
等人。, 2011). This would explain the finding of more robust
and reliable effects on guessing, rather than standard devi-
化. Although the precise mechanism by which TMS in-
duces these effects is unclear, a recent optical imaging
study examining the effects of TMS on cat visual cortex ac-
tivity suggests that the reduction in signal intensity may be
caused by TMS-induced local cortical inhibition (Kozyrev,
Eysel, & Jancke, 2014). In their study, single pulses of
TMS applied to the visual cortex were found to induce a
localized pattern of inhibition that lasted approximately
300 msec before returning to baseline. Assuming similar
mechanisms are at work in human visual cortex, this brief
period of inhibition may be adequate to disrupt the initial
formation of WM representations, without affecting later
activity once consolidation is complete. Whether this is cor-
rect will require further work examining the neural effects
of TMS. 尤其, this question could be profitably ad-
dressed by the adoption of a computational neurostimula-
tion approach, in which the effects of simulated TMS
pulses are examined in the context of realistic neural
models of the consolidation and maintenance of informa-
tion in WM (for discussion of this approach, see Bestmann
& Feredoes, 2013).
Another notable aspect of the data that speaks to the
functional role of the early visual cortex in VWM is the
time course of the effects of masks and TMS on perfor-
曼斯. If activity in the early visual cortex is critical for
both the initial consolidation and later maintenance of
information in VWM, as has been proposed by sensory
recruitment models of WM (D’Esposito & Postle, 2015),
we expected TMS to disrupt performance at all latencies
经测试. Contrary to this possibility, in each experiment,
the effects of TMS and of masks were either substantially
reduced or entirely absent when stimulation was applied
at later time points, when consolidation was nearing
completion and short-term maintenance had presumably
begun. These findings match those of van de Ven et al.
(2012), who observed a significant disruption of perfor-
mance when spTMS was applied 200 msec after memory
display offset, 但不是 200 msec later. 相似地, Beckers
and Hömberg (1991) reported no effect on performance
when spTMS was applied to the visual cortex during the
retention interval of a delayed match to sample task re-
quiring memory for faces, although performance was dis-
rupted by TMS applied during retrieval.
These findings suggest that early visual cortex in-
volvement in VWM may be restricted to an early consol-
idation time window during which fragile sensory
van Lamsweerde and Johnson
1235
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
9
/
7
2
9
1
/
2
7
2
/
6
1
1
2
9
2
5
6
2
/
7
1
5
7
7
8
哦
6
C
4
n
0
_
3
A
/
_
j
0
哦
1
C
1
n
1
3
_
A
p
_
d
0
1
乙
1
y
1
G
3
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
/
F
t
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
representations are being transformed into more dura-
ble VWM representations.
This conclusion is consistent with several lines of evi-
dence suggesting that, although initial perceptual process-
ing depends on activity in early visual areas, maintenance in
VWM likely depends on activity in higher-order cortical
地区, such as the parietal or frontal cortex (Bettencourt
& 徐, 2016; Mendoza-Halliday, Torres, & Martinez-Trujillo,
2014; 徐, 2010; 徐 & Chun, 2006; Todd & Marois, 2004,
2005). 例如, using fMRI and multivoxel pattern
classification, Bettencourt and Xu (2016) reported that
the ability to decode the orientation of a remembered
stimulus from patterns of activity in specific regions of
the occipital and parietal cortex differed depending on
whether new, task-irrelevant stimuli were presented
during the delay. Although it was possible to decode item
orientation from both areas during an unfilled delay,
when task-irrelevant distractors were presented, decoding
from early visual areas was no longer possible, 甚至
though task performance was unaffected by the new in-
放. 相比之下, decoding from the superior intraparietal
sulcus, a region that has been implicated in setting capac-
ity limits in WM (Todd & Marois, 2004, 2005), 留下来
intact and closely tracked WM task performance.
相似地, a recent neurophysiological recording study
in macaques (Mendoza-Halliday et al., 2014) found that,
although the direction of motion of a stored stimulus
could be decoded from local field potential activity
recorded in the motion-selective middle temporal area
(公吨), stimulus-specific spiking activity was only ob-
served in higher-order, multimodal areas. 而且, 低的-
frequency neural oscillations in MT were phase-coherent
with spiking activity in the frontal cortex, suggesting that
the local field potential activity supporting decoding in
MT was likely driven by feedback inputs from the frontal
or parietal cortex, rather than reflecting memory-related
activity arising in early visual area MT itself. Findings such
as these suggest that, although it may be possible to
decode stimulus identity from the early visual cortex in
一些案例, this activity may not be causally necessary
for storage in WM.
In a reply to the study of Bettencourt and Xu (2016),
Ester, Rademaker, and Sprague (2016) pointed out that a
failure to decode from a given area does not constitute
strong evidence against this area’s involvement in the
process in question. 例如, it remains possible that
the visual cortex was engaged in storage in the distractor
condition of Bettencourt and Xu (2016), but at a level
that was inaccessible to multivoxel decoding analyses
(see Dubois, de Berker, & Tsao, 2015, for evidence dem-
onstrating such a failure of decoding in the macaque face
patch system). Similar arguments have been forwarded
to explain several notable failures to decode item identity
from delay period activity in both frontal and parietal re-
祇翁 (Emrich et al., 2013; 林登, Oosterhof, 克莱因, &
唐宁, 2012; Riggall & Postle, 2012). 例如, 在
a recent review paper, Riley and Constantinidis (2016)
argue that the spatial resolution of fMRI may be insuffi-
cient to support successful decoding of item identity
from the frontal cortex, which is characterized by non-
topographic organization and the representation of in-
formation at spatial scales that can be an order of
magnitude finer than the resolution of fMRI. 鉴于这种,
failures to decode the contents of WM from the frontal
cortex would be expected, even if this information is
robustly represented in this area.
This discussion highlights potential limitations in the
kinds of inferences that can be drawn from pattern clas-
sification analyses alone. 一方面, as noted by
Ester et al. (2016) and Riley and Constantinidis (2016),
a failure to decode does not necessarily constitute strong
evidence against a particular area’s involvement in a
given process. 另一方面, the results of
Bettencourt and Xu (2016) and Mendoza-Halliday et al.
(2014) suggest that the ability to decode from a particular
area is no guarantee that this area is causally necessary
for storage. One way of addressing these inferential lim-
itations is to adopt causal methods such as TMS, 和我们一样
have done here. 至关重要的是, in both our study and that of
van de Ven et al. (2012), TMS only disrupted perfor-
mance when applied relatively early in the delay period
(see also Beckers & Hömberg, 1991). These findings
strongly suggest that, although it is sometimes possible
to decode item identity from this area, the early visual
cortex is likely not causally necessary to represent item
information beyond the time period of initial memory
consolidation.
最后, in addition to the temporally specific disruptive
effects of TMS on guess rate, we also observed a signifi-
cant main effect of stimulation side on swap rate (IE。, 在
the likelihood of confusing the cued item with one of the
other items in memory). Contrary to what we would have
预期的, the swap rate was significantly reduced for tar-
gets appearing in the affected hemifield across all inter-
vals tested. 那是, TMS appears to have had a reliable
enhancing effect for contralateral targets, 增加
likelihood of correctly binding individual colors to specif-
ic locations. According to Bays et al. (2009), swap errors
likely arise as a result of coarse coding of the spatial po-
sitions of remembered features. This can lead to confu-
sion about which item to report at test, particularly when
either a large number of items need to be remembered
(Bays et al., 2009) or when stimuli are presented very
close together in space (Emrich & Ferber, 2012). 一
means by which TMS could reduce this type of error,
所以, is if it serves to sharpen the spatial tuning of
neurons in the stimulated area. Although we are unaware
of any direct evidence supporting this possibility, the re-
sults of Kozyrev et al. (2014) discussed above are sugges-
主动的. In their study, single pulses of TMS were found to
produce a short-lived, localized increase in inhibition,
which they attribute to a selective effect of TMS on inhib-
itory interneurons. Increased inhibition has, 反过来, 到过
shown to produce a sharpening of neural tuning in
1236
认知神经科学杂志
体积 29, 数字 7
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
9
/
7
2
9
1
/
2
7
2
/
6
1
1
2
9
2
5
6
2
/
7
1
5
7
7
8
哦
6
C
4
n
0
_
3
A
/
_
j
0
哦
1
C
1
n
1
3
_
A
p
_
d
0
1
乙
1
y
1
G
3
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
F
.
/
t
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
various sensory cortical areas (Isaacson & Scanziani,
2011). If spTMS of the visual cortex in our study in-
creased inhibition and this increase sharpened the tuning
of cells coding for the spatial position of remembered
项目, this could explain the beneficial effect of TMS on
color–location binding. This possibility is frankly specula-
主动的, but it could be tested by assessing the impact of
spTMS of early visual cortex on recall of the spatial posi-
的, rather than color, of a remembered item. If this were
the case, we would expect TMS to produce an improve-
ment in the resolution of spatial recall (IE。, reduced SD).
TMS as applied here is a powerful method of determin-
ing the functional relevance of a particular brain area.
Future work could benefit by stimulating a wider range
of areas and/or localizing TMS targets using functional
criteria. 例如, current models and data suggest
that WM functions are likely supported by distributed ac-
tivity within a network of partly functionally specialized
brain regions, rather than being mediated by a single area
(Ester et al., 2016; D’Esposito & Postle, 2015; Ester,
Sprague, & Serences, 2015). Future research could ben-
efit by targeting other regions within this network, 这样的
as the parietal or pFC, to determine their specific contri-
butions to WM. 此外, although the current study
suggests that early visual cortex is not causally necessary
for WM storage, different results may be achieved by
targeting specific subregions of the visual cortex on the
basis of functional criteria, rather than phosphene
perception alone. 例如, TMS targeting could be
guided by fMRI scans aimed at localizing specific areas
exhibiting particular functional characteristics (例如, 颜色
selectivity or load sensitivity) by selecting regions on the
basis of accurate classification of stimulus features or via
TMS-based localization of feature-specific regions. 作为一个
example of the latter, Banissy, Walsh, and Muggleton
(2012) have shown that continuous theta-burst stimula-
tion applied to area V4 disrupted color priming effects
in a subsequent task. A procedure such as this one could
be used to localize color-selective areas of visual cortex
before the application of TMS during a color WM task.
It is possible that localizing stimulus-selective regions in
this way could reveal a role of visual cortex in WM main-
租赁. This is still to be determined however, and ad-
ditional research will be required to determine the
functional roles of each region in the network of brain
areas associated with WM functioning.
综上所述, results of this study suggest that inter-
fering with visual cortex activity by either presenting
visual masks or directly stimulating the cortex with TMS
disrupts performance primarily by influencing the num-
ber of items that are successfully stored, with smaller,
more variable disruptions of the quality of VWM repre-
sentation. 在每种情况下, observed disruptive effects were
largest when interference was applied very early during
the retention interval, growing smaller or disappearing
altogether at later intervals. 此外, TMS also pro-
duced longer-lasting, location-specific improvements in
color–location binding. 合在一起, this pattern of
results echoes recent findings suggesting that early visual
cortex involvement in VWM for object features may be
restricted to the period of time during which fragile
sensory representations are being transformed into more
durable VWM representations, with its role dissipating
the further one goes into the retention interval.
Reprint requests should be sent to Amanda E. van Lamsweerde,
North Dakota State University, Dept. 2765, P.O. Box 6050,
Fargo, ND 58108, or via e-mail: amanda.vanlamsweerde@
ndsu.edu.
参考
Abrahamyan, A。, Clifford, C. 瓦. G。, Arabzadeh, E., & 哈里斯, J. A.
(2011). Improving visual sensitivity with subthreshold
transcranial magnetic stimulation. 神经科学杂志,
31, 3290–3294.
Banissy, 中号. J。, Walsh, 五、, & Muggleton, 氮. G. (2012). A
disruption of colour priming following continuous theta
burst transcranial magnetic stimulation. Cortex, 48,
1359–1361.
Bays, 磷. M。, Catalao, 右. F. G。, & Husain, 中号. (2009). The precision
of visual working memory is set by allocation of a shared
resource. Journal of Vision, 9, 1–11.
Beckers, G。, & Hömberg, V. (1991). Impairment of visual
perception and visual short term memory scanning by
transcranial magnetic stimulation of occipital cortex.
Experimental Brain Research, 87, 421–432.
Bestmann, S。, & Feredoes, 乙. (2013). 组合
neurostimulation and neuroimaging in cognitive
神经科学: 过去的, 展示, and future. Annals of the
New York Academy of Sciences, 1296, 11–30.
Bettencourt, K. C。, & 徐, 是. (2016). Decoding the content of
visual short-term memory under distraction in occipital and
parietal areas. 自然神经科学, 19, 150–157.
Brainard, D. H. (1997). The psychophysics toolbox. 空间
Vision, 10, 433–436.
卡塔内奥, Z。, Vecchi, T。, Pascual-Leone, A。, & Silvanto, J. (2009).
Contrasting early visual cortical activation states causally
involved in visual imagery and short-term memory.
欧洲神经科学杂志, 30, 1393–1400.
考特尼, S. M。, Ungerleider, L. G。, Keil, K., & Haxby, J. V.
(1997). Transient and sustained activity in a distributed
neural system for human working memory. 自然,
386, 608–611.
D’Esposito, 中号. (2007). From cognitive to neural models of
工作记忆. Philosophical Transactions of the Royal
Society of London, Series B, Biological Sciences, 362,
761–772.
D’Esposito, M。, & Postle, 乙. 右. (2015). The cognitive
neuroscience of working memory. Annual Review of
心理学, 66, 115–142.
Dubois, J。, de Berker, A. 奥。, & Tsao, D. 是. (2015). Single-unit
recordings in the macaque face patch system reveal
limitations of fMRI MVPA. 神经科学杂志, 35,
2791–2802.
Emrich, S. M。, & Ferber, S. (2012). Competition increases
binding errors in visual working memory. Journal of Vision,
1212, 1–16.
Emrich, S. M。, Riggall, A. C。, LaRocque, J. J。, & Postle, 乙. 右.
(2013). Distributed patterns of activity in sensory cortex
reflect the precision of multiple items maintained in visual
short-term memory. 神经科学杂志, 33, 6516–6523.
van Lamsweerde and Johnson
1237
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
9
/
7
2
9
1
/
2
7
2
/
6
1
1
2
9
2
5
6
2
/
7
1
5
7
7
8
哦
6
C
4
n
0
_
3
A
/
_
j
0
哦
1
C
1
n
1
3
_
A
p
_
d
0
1
乙
1
y
1
G
3
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
F
t
/
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
Esser, S. K., Huber, R。, Massimini, M。, 彼得森, 中号. J。, Ferrarelli,
F。, & 托诺尼, G. (2006). A direct demonstration of cortical
LTP in humans: A combined TMS/EEG study. 脑研究
Bulletin, 69, 86–94.
Ester, 乙. F。, 安德森, D. E., & Serences, J. 时间. (2013). A neural
measure of precision in visual working memory. 杂志
Cognitive Neuroscience, 25, 754–761.
Ester, 乙. F。, Rademaker, 右. L。, & Sprague, 时间. C. (2016). 怎么办
visual and parietal cortex contribute to visual short-term
记忆? eNeuro, 3, ENEURO.0041.
Ester, 乙. F。, Sprague, 时间. C。, & Serences, J. 时间. (2015). Parietal and
frontal cortex encode stimulus-specific mnemonic
representations during visual working memory parietal and
frontal cortex encode stimulus-specific mnemonic
representations during visual working memory. 神经元,
87, 893–905.
Goldman-Rakic, 磷. S. (1995). Architecture of the prefrontal
cortex and the central executive. Annals of the New York
Academy of Sciences, 769, 71–83.
哈里斯, J. A。, Clifford, C. 瓦. G。, & Miniussi, C. (2008). 这
functional effect of transcranial magnetic stimulation: Signal
suppression or neural noise generation? 认知杂志
神经科学, 20, 734–740.
哈里森, S. A。, & Tong, F. (2009). Decoding reveals the
contents of visual working memory in early visual areas.
自然, 458, 632–635.
Holm, S. (1979). A simple sequentially rejective multiple test
procedure. Scandanavial Journal of Statistics, 6, 65–70.
Isaacson, J. S。, & Scanziani, 中号. (2011). How inhibition shapes
cortical activity. 神经元, 72, 231–243.
约翰逊, J. S。, Kundu, B., Casali, A. G。, & Postle, 乙. 右. (2012).
Task-dependent changes in cortical excitability and effective
连接性: A combined TMS-EEG study. 杂志
Neurophysiology, 107, 2383–2392.
Kammer, 时间. (1998). Phosphenes and transient scotomas
induced by magnetic stimulation of the occipital lobe: 他们的
topographic relationship. Neuropsychologia, 37, 191–198.
Kozyrev, 五、, Eysel, U. T。, & Jancke, D. (2014). Voltage-sensitive
dye imaging of transcranial magnetic stimulation-induced
intracortical dynamics. 国家科学院院刊
科学系, 美国。, 111, 13553–13558.
林登, D. 乙. J。, Oosterhof, 氮. N。, 克莱因, C。, & 唐宁, 磷. 乙.
(2012). Mapping brain activation and information during
category-specific visual working memory. 杂志
Neurophysiology, 107, 628–639.
Luck, S. J。, & 沃格尔, 乙. K. (1997). The capacity of visual working
memory for features and conjunctions. 自然, 390,
281–297.
Makovski, T。, & Lavidor, 中号. (2014). Stimulating occipital cortex
enhances visual working memory consolidation. Behavioural
脑研究, 275, 84–87.
Mendoza-Halliday, D ., Torres, S。, & Martinez-Trujillo, J. C.
(2014). Sharp emergence of feature-selective sustained
activity along the dorsal visual pathway. 自然
神经科学, 17, 1255–1262.
Pasternak, T。, & Greenlee, 中号. 瓦. (2005). Working memory in
primate sensory systems. 自然评论神经科学,
6, 97–107.
Pelli, D. G. (1997). The VideoToolbox software for visual
psychophysics: Transforming numbers into movies. 空间
Vision, 10, 437–442.
Postle, 乙. 右. (2006). Working memory as an emergent property
of the mind and brain. 神经科学, 139, 23–38.
Riggall, A. C。, & Postle, 乙. 右. (2012). The relationship between
working memory storage and elevated activity as measured
with functional magnetic resonance imaging. 杂志
神经科学, 32, 12990–12998.
Riley, 中号. R。, & Constantinidis, C. (2016). Role of prefrontal
persistent activity in working memory. Frontiers in Systems
神经科学, 9, 1–14.
Rossi, S。, Hallett, M。, Rossini, 磷. M。, & Pascual-Leone, A. (2009).
Safety, ethical considerations, and application guidelines for
the use of transcranial magnetic stimulation in clinical practice
and research. Clinical Neurophysiology, 120, 2008–2039.
施华蔻, D. S。, Silvanto, J。, & Rees, G. (2011). Stochastic
resonance effects reveal the neural mechanisms of
transcranial magnetic stimulation. 神经科学杂志,
31, 3143–3147.
Serences, J。, Ester, E., 沃格尔, 乙. K., & Awh, 乙. (2009). Stimulus-
specific delay activity in human primary visual cortex.
心理科学, 20, 207–214.
Suchow, J. W., 布雷迪, 时间. F。, Fougnie, D ., & 阿尔瓦雷斯, G. A. (2013).
Modeling visual working memory with the MemToolbox.
Journal of Vision, 13, 1–8.
Todd, J. J。, & Marois, 右. (2004). Capacity limit of visual short-
term memory in human posterior parietal cortex. 自然,
428, 751–754.
Todd, J. J。, & Marois, 右. (2005). Posterior parietal cortex activity
predicts individual differences in visual short-term memory
容量. 认知的, Affective, & Behavioral Neuroscience,
5, 144–155.
van de Ven, 五、, Jacobs, C。, & Sack, A. 时间. (2012). Topographic
contribution of early visual cortex to short-term memory
consolidation: A transcranial magnetic stimulation study.
神经科学杂志, 32, 4–11.
van de Ven, 五、, & Sack, A. 时间. (2013). Transcranial magnetic
stimulation of visual cortex in memory: Cortical state,
interference and reactivation of visual content in memory.
Behavioural Brain Research, 236, 67–77.
沃格尔, 乙. K., & Machizawa, 中号. G. (2004). Neural activity predicts
individual differences in visual working memory capacity.
自然, 428, 748–751.
沃格尔, 乙. K., Woodman, G. F。, & Luck, S. J. (2006). The time
course of consolidation in visual working memory. 杂志
实验心理学: Human Perception and
Performance, 32, 1436–1451.
Wilken, P。, & Ma, 瓦. J. (2004). A detection theory account of
change detection. Journal of Vision, 4, 1120–1135.
徐, 是. (2010). The neural fate of task-irrelevant features in object-
based processing. 神经科学杂志, 30, 14020–14028.
徐, Y。, & Chun, 中号. 中号. (2006). Dissociable neural mechanisms
supporting visual short-term memory for objects. 自然,
440, 91–95.
张, W., & Luck, S. J. (2008). Discrete fixed-resolution
representations in visual working memory. 自然, 453,
233–236.
1238
认知神经科学杂志
体积 29, 数字 7
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
9
/
7
2
9
1
/
2
7
2
/
6
1
1
2
9
2
5
6
2
/
7
1
5
7
7
8
哦
6
C
4
n
0
_
3
A
/
_
j
0
哦
1
C
1
n
1
3
_
A
p
_
d
0
1
乙
1
y
1
G
3
你
.
e
p
s
t
d
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
2
我
3
e
s
/
j
F
/
t
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1