Adjustments to Proactive Motor Inhibition without
Effector-Specific Foreknowledge Are Reflected in a
Bilateral Upregulation of Sensorimotor β-Burst Rates
Cheol Soh1, Megan Hynd1, Benjamin O. Rangel2, and Jan R. Wessel1,2
抽象的
■ Classic work using the stop-signal task has shown that humans
can use inhibitory control to cancel already initiated movements.
Subsequent work revealed that inhibitory control can be proac-
tively recruited in anticipation of a potential stop-signal, 从而
increasing the likelihood of successful movement cancellation.
然而, the exact neurophysiological effects of proactive inhib-
itory control on the motor system are still unclear. On the basis of
classic views of sensorimotor β-band activity, as well as recent
findings demonstrating the burst-like nature of this signal, 我们
recently proposed that proactive inhibitory control is imple-
mented by influencing the rate of sensorimotor β-bursts during
movement initiation. 这里, we directly tested this hypothesis
using scalp EEG recordings of β-band activity in 41 healthy human
adults during a bimanual RT task. By comparing motor responses
made in two different contexts—during blocks with or without
stop-signals—we found that premovement β-burst rates over both
contralateral and ipsilateral sensorimotor areas were increased in
stop-signal blocks compared to pure-go blocks. 而且, 这
degree of this burst rate difference indexed the behavioral imple-
mentation of proactive inhibition (IE。, the degree of anticipatory
response slowing in the stop-signal blocks). 最后, exploratory
analyses showed that these condition differences were explained
by a significant increase in β bursting that was already present
during baseline period before the movement initiation signal.
一起, this suggests that the strategic deployment of proactive
inhibitory motor control is implemented by upregulating the tonic
inhibition of the motor system, signified by increased sensorimotor
β-bursting both before and after signals to initiate a movement. ■
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
哦
C
n
_
A
_
0
1
6
8
2
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
介绍
Inhibitory control is a fundamental cognitive control ability
that supports the implementation of many flexible and
adaptive behaviors. In the motoric domain, inhibitory
control allows humans to stop a movement outright, 甚至
if that movement has already been initiated. In the real
世界, this ability can help humans adapt their behavior
to a rapidly changing environment—be it by stopping to
walk into the street when suddenly noticing a previously
overlooked car or by stopping to pick up a berry when
realizing that a snake is lurking in the underwood. 在里面
laboratory, the ability to implement inhibitory control is
most often studied in the stop-signal task (SST; Verbruggen
等人。, 2019; Logan, 考恩, & 戴维斯, 1984). In this task,
participants first initiate an action to a go-signal that is pre-
sented on every trial. On some trials, a second signal—the
stop-signal—is then presented with some delay after the
initial go-signal, and participants have to attempt to cancel
their response to the go-signal.
Subsequent work using this experimental paradigm has
shown that successful action stopping in the SST depends
on a mixture of two types of inhibitory control processes:
1University of Iowa, 2University of Iowa Hospital and Clinics
© 2021 麻省理工学院
reactive and proactive (Braver, 2012). Reactive inhibition is
triggered by the stop-signal itself, whereas proactive pro-
cesses are recruited before the stop-signal, in anticipation
of potentially having to cancel the movement ( 韦塞尔,
2018; Elchlepp, Lavric, Chambers, & Verbruggen, 2016;
Aron, 2011; Verbruggen & Logan, 2009). In real-world
scenarios, proactive inhibitory control is evident by the in-
crease in motor caution that is exerted in some environ-
mental contexts. 例如, when approaching a busy
intersection, humans may slow their walk in anticipation
of an approaching vehicle. 相似地, when foraging in
dense foliage, they may more cautiously grasp for foods
in anticipation of potentially lurking carnivorous reptiles.
To emulate such proactive control behaviors in the labora-
保守党, researchers typically compare responses made in
experimental blocks in which participants are instructed
to expect no stop-signals (IE。, a pure-go RT task) to blocks
in which stop-signals are expected (IE。, the classic SST
设计). Using this approach, studies have consistently
found that go-responses made in stop-signal blocks are
made more slowly compared to go-responses made in
pure-go blocks, reflecting the heightened motor caution
that humans exert when they anticipate potentially having
to stop in the real world (Swann et al., 2012; Chikazoe et al.,
2009; Jahfari, Stinear, Claffey, Verbruggen, & Aron, 2009;
认知神经科学杂志 33:5, PP. 784–798
https://doi.org/10.1162/jocn_a_01682
Verbruggen & Logan, 2009). 至关重要的是, such studies also
consistently found that the engagement of proactive inhib-
itory control directly benefits the implementation of reac-
tive inhibitory control: 具体来说, participants who show
greater degrees of anticipatory motor slowing in stop-
signal blocks also show faster stop-signal RTs (SSRTs).
This also suggests that humans can strategically balance
the degree to which they rely on either proactive or reac-
tive control mechanisms (Braver, 2012), 取决于,
例如, on how strongly they prioritize successful
stopping compared to fast responding (Greenhouse &
韦塞尔, 2013) or on how high they deem the relative
likelihood of a stop-signal to be on a given trial ( Vink,
Kaldewaij, Zandbelt, Pas, & du Plessis, 2015).
With regard to the neural basis of inhibitory control, cogni-
tive neuroscience has contributed much to our knowledge
of how both reactive and proactive inhibitory control are
implemented in the brain. The study of the basic stop-
signal paradigm has elucidated a fronto-BG network of
brain regions that is activated by stop-signals and imple-
ments reactive inhibitory control (for reviews, see Schmidt
& Berke, 2017; 韦塞尔 & Aron, 2017; Jahanshahi, Obeso,
Rothwell, & Obeso, 2015; Aron, Robbins, & Poldrack,
2014; 里德林克霍夫, Forstmann, Wylie, Burle, & van den
Wildenberg, 2011; Robbins, 2007). The neural underpin-
nings of proactive inhibitory control, 然而, are com-
paratively less clearly defined (迈耶 & Bucci, 2016;
凯尼曼斯, 2015; Aron, 2011). Broadly speaking, 学习
either report that proactive inhibitory control is imple-
mented via the anticipatory recruitment of the same
control circuitry that is underlying reactive inhibitory
控制 (例如, Cunillera, Brignani, Cucurell, Fuentemilla,
& Miniussi, 2016; Cai et al., 2015; Cunillera, Fuentemilla,
Brignani, Cucurell, & Miniussi, 2014; Swann et al., 2012;
Jahfari et al., 2009), via the attentional tuning of sensory
processes that help detect potential stop-signals (例如,
Elchlepp et al., 2016; 凯尼曼斯, 2015), or via entirely sep-
arate neural circuitry (例如, Cai, 陈, Ide, 李, & Menon,
2016; Vink et al., 2015; Majid, Cai, Corey-Bloom, & Aron,
2013; Zandbelt & Vink, 2010; Jaffard et al., 2008)—with
many studies finding a mixture of these patterns. The most
informative insights into the implementation of proactive in-
hibitory control come from studies that relate its purported
neural signatures to its behavioral expression—that is, 他们
show a systematic relationship between the relative slowing
of motor responses in the stop-signal context (from here
forward referred to as “proactive RT slowing”) and the pur-
ported neural signature of proactive inhibition. In one of
the earliest studies of this type, Chikazoe et al. (2009) 成立
那, across participants, the degree of neural activity within
the reactive control network after stop-signals was inversely
related to the degree of their proactive RT slowing. 这
provided one of the first pieces of neuroscientific support
for the notion that the behavioral recruitment of proactive
inhibition reduces the demand on the neural reactive inhi-
bition system when movement cancellation is needed.
Using a similar approach, Jahfari et al. (2009) have found
that proactive RT slowing was correlated with increased ac-
tivity in a broad and distributed array of control-related
frontoparietal brain regions activated on go-trials—most
notably prefrontal brain regions that are also active during
reactive control. This finding supports the view that pro-
active control can be implemented by an anticipatory
recruitment of reactive control circuitry (but likely also
involves additional structures and processes).
Further studies have lent additional evidence to the view
that proactive inhibitory control is not purely attributable
to the tuning of attentional processes that support stop-
signal detection, but can instead directly affect the activity
of the motor system. In that vein, studies using TMS have
found that the recruitment of proactive inhibitory control
can override the broad, nonselective suppression of motor
excitability that is typically found during the reactive period
after the stop-signal, thereby helping participants to more
selectively target specific parts of the motor system when
executing a stop (Duque, Greenhouse, Labruna, & Ivry,
2017; Greenhouse, Oldenkamp, & Aron, 2011). 然而,
尤其, the vast majority of studies that measure such
changes in corticospinal excitability during the proactive
control period use “foreknowledge” paradigms—that is,
tasks in which participants know ahead of their motor ini-
tiation which specific motor effector they ultimately may
have to stop (Cai, 乔治, Verbruggen, Chambers, &
Aron, 2012; Greenhouse et al., 2011; Majid, Cai, 乔治,
Verbruggen, & Aron, 2011; Claffey, Sheldon, Stinear,
Verbruggen, & Aron, 2010). By utilizing this additional
信息, participants are able to selectively inhibit the
critical effector without affecting the rest of the motor sys-
TEM (for reviews, see Duque et al., 2017; Aron, 2011). 在
contrast to this well-established literature on proactive,
selective inhibition under conditions of foreknowledge,
much less is known about whether (以及如何) proactive
control affects the motor system when the to-be-inhibited
motor effector is not known ahead of time—that is, 什么时候
responses have to be made under a “global” state of cau-
tion and inhibition and cannot be targeted at a previously
已知的, specific motor representation. 而且, the neu-
rophysiological changes that are underlying these ob-
served changes in motor system excitability because of
proactive control are still largely unknown.
In the latter respect, the neurophysiological activity of
the sensorimotor cortex can be noninvasively investigated
using scalp EEG. A substantial literature of existing research
has consistently shown that the sensorimotor cortices
display prominent activity in the β-frequency band (∼15–
29 赫兹). This β-band activity is suppressed leading up to and
during movement (Tzagarakis, 西方, & Pellizzer, 2015;
Kilavik, Zaepffel, Brovelli, MacKay, & Riehle, 2013;
Pogosyan, Gaynor, Eusebio, & 棕色的, 2009; 锄,
Oostenveld, & 薯条, 2008; 张, 陈, Bressler, &
Ding, 2008; 贝克, 2007; Neuper, Wörtz, & Pfurtscheller,
2006; Pfurtscheller, Graimann, Huggins, 莱文, & Schuh,
2003; McFarland, Miner, Vaughan, & Wolpaw, 2000) 和
is directly related to corticospinal excitability (Schulz,
Soh et al.
785
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
哦
C
n
_
A
_
0
1
6
8
2
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
Übelacker, Keil, 穆勒, & 魏兹, 2013). There are several
proposed neurophysiological and functional interpretations
of β-band activity during movement (例如, Kilavik et al., 2013;
恩格尔 & 薯条, 2010). One of the earliest and most enduring
ideas is the proposal that sensorimotor β-band activity
reflects a net-inhibited state of the motor system (Schulz
等人。, 2013; Pfurtscheller, Neuper, Flotzinger, & Pregenzer,
1997). Concomitantly, its reduction during movement initi-
ation purportedly reflects the net disinhibition of the
motor system in preparation for movement.
Building on that theory, two recent stop-signal studies
have proposed that proactive inhibitory control may be im-
plemented via a preparatory upregulation of sensorimotor
β-band activity (韦塞尔, 2020; Muralidharan, 于, 科恩, &
Aron, 2019). Muralidharan et al. (2019) 表明, 什么时候
foreknowledge is present, the selective, effector-specific
proactive inhibitory control that is typically found in such
paradigms is indexed by increased β-band activity over the
contralateral sensorimotor site. 反过来, the Wessel
(2020) study used a standard SST without foreknowledge,
where inhibition cannot be targeted at a specific motor
effector ahead of time. In that study, we first showed that
trial-to-trial β-band activity in the sensorimotor cortex
occurs in brief, transient bursts, rather than in the form
of steady oscillations (which is in line with recent findings
from the animal literature; 比照. Sherman et al., 2016;
Feingold, 吉布森, DePasquale, & Graybiel, 2015). We then
showed that the rate of these β bursts steadily declines in
the lead-up to the movement, mirroring previous find-
ings from the classic, power-based β literature (reviewed
多于). 至关重要的是, we then found that the amount of senso-
rimotor β-bursts in the early premovement period positively
predicted go-trial RT (more sensorimotor β-bursts =
longer RTs) and negatively predicted SSRT (more bursts =
faster SSRT). 所以, we tentatively proposed that early
sensorimotor β-bursting on go-trials could reflect the de-
gree to which participants engaged general, nonspecific
proactive inhibition of all potential responses.
然而, although this study employed a large sample
size of over 200 participants and therefore revealed reliable
brain–behavior associations, the standard SST used therein
did not include a pure-go condition to provide a control
comparison without proactive inhibition. 因此, 它是
impossible to definitively conclude that early sensorimotor
β-bursting truly reflects an active, strategic deployment of
general proactive control. 的确, a plausible alternative
explanation is that some participants may innately show
higher sensorimotor β bursting (IE。, a more strongly inhib-
ited motor system overall), which would lead them to show
longer RTs in general (IE。, a slower go-process), 不只是
within the context of an SST. In the stop-signal context,
然后, participants with such a slowed go-process would
have a systematic bias in the “race” between the stop-
process and the go-process, resulting in more efficient
reactive stopping (indexed by shorter SSRT). In that sce-
成员, the observed direct relationship between β-bursts
and both go- and stop-signal RTs could be explained
without any relationship between proactive inhibitory
control and β-band activity.
所以, in the current study, we set out to directly test
whether increases in sensorimotor β-bursts actually reflect
a genuine strategic deployment of proactive inhibitory
控制. As in previous studies of proactive control in the
absence of effector-specific foreknowledge ( 韦塞尔,
Conner, Aron, & Tandon, 2013; Swann et al., 2012), 参与-
爱普茨 (n = 41) alternated between performing blocks of
the SST and a pure-go task. We used scalp EEG to measure
sensorimotor β-burst rates during go-trials performed in
both contexts and aimed to directly test whether sensori-
motor β-burst rates contralateral and/or ipsilateral to the
response hand were increased in the stop-signal context
compared to the pure-go context. We also aimed to relate
these changes in sensorimotor β-bursting to proactive
RT slowing. 最后, we aimed to explore whether the
purported increase in β-bursting during the stop-signal
blocks was specific to the period immediately after the
go-signal (suggesting a strategic, transient deployment of
proactive control specific to the movement initiation
时期) or whether such increases were found even before
the go-signal was presented (suggesting a more broad
control state that is sustained throughout the task).
方法
Data Availability
All data, analysis scripts, and task codes can be down-
loaded from the Open Science Framework at https://osf
.io/49hts/.
参加者
Forty-one healthy adult participants (27 女性) partici-
pated in this study (mean age = 21.6 年, 标准差= 3.85, range =
18–30 years) in exchange for either class credit or an hourly
compensation of $15. All procedures were approved by
the University of Iowa’s institutional review board (IRB
#201511709). The same data were used to investigate a
hypothesis about the fronto-central P3 event-related poten-
tial in the context of reactive inhibitory control (Hynd, Soh,
Rangel, & 韦塞尔, 2021).
Stimulus Presentation
All stimuli were presented on a 19-in. Dell flat-screen mon-
itor connected to an IBM-compatible PC running Fedora
Linux and MATLAB 2015b (The MathWorks, 公司). Stimuli
were presented using Psychtoolbox 3 (Brainard, 1997).
Responses were made using a standard QWERTY USB
keyboard.
Experimental Task
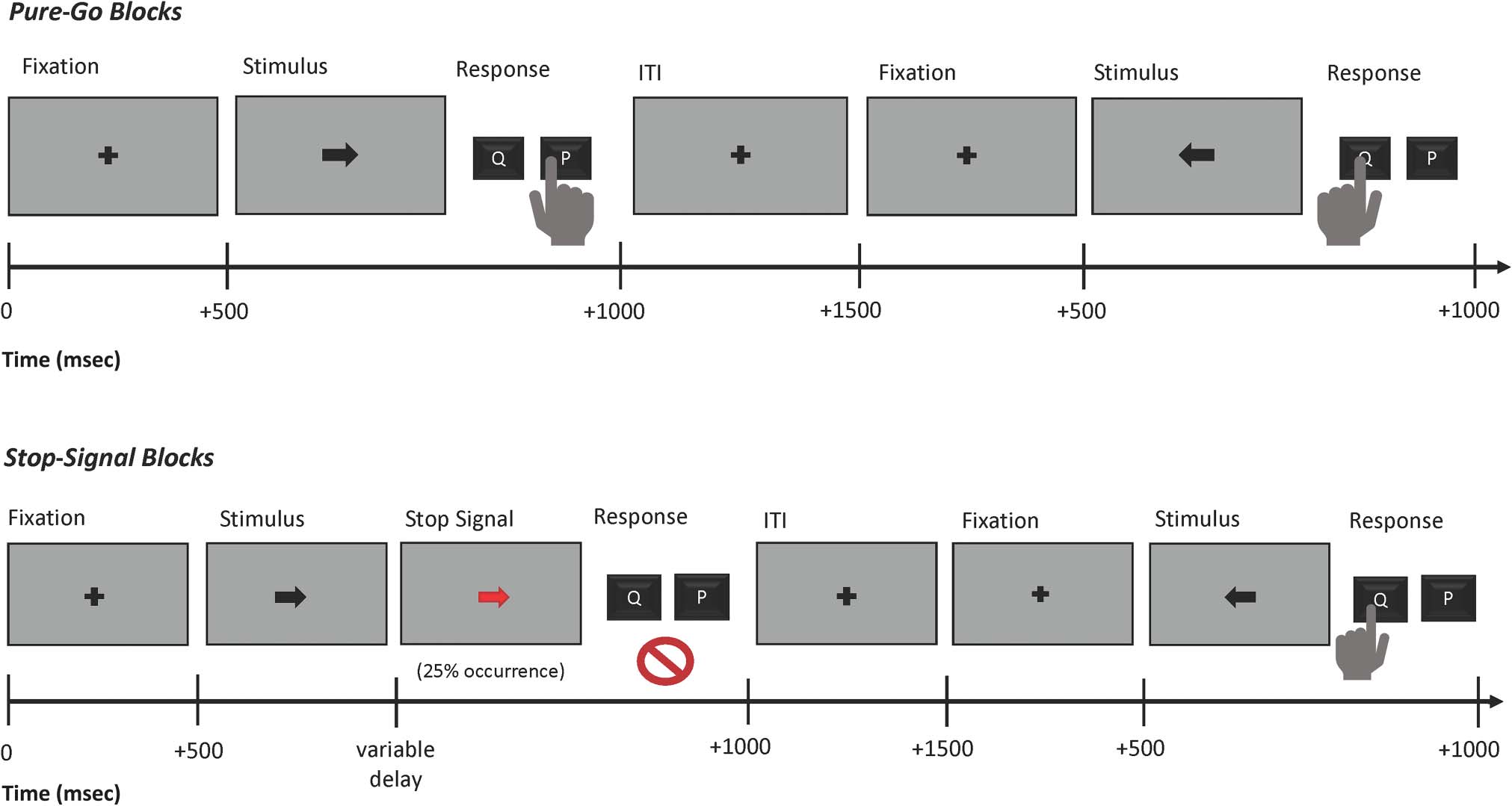
Stimuli were presented on a gray background (数字 1).
Each trial began with a black fixation cross (500 毫秒),
786
认知神经科学杂志
体积 33, 数字 5
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
哦
C
n
_
A
_
0
1
6
8
2
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
我
D
哦
w
n
哦
A
d
e
d
F
r
哦
米
H
t
t
p
:
/
/
d
我
r
e
C
t
.
米
我
t
.
e
d
你
/
j
/
哦
C
n
A
r
t
我
C
e
–
p
d
我
F
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
哦
C
n
_
A
_
0
1
6
8
2
p
d
.
F
乙
y
G
你
e
s
t
t
哦
n
0
8
S
e
p
e
米
乙
e
r
2
0
2
3
数字 1. Task diagram.
followed by a black arrow (go-signal) pointing either left or
正确的, displayed for 1000 毫秒. Participants were instructed
to press the “q” key on the keyboard with their left index
finger in case of a left-facing arrow and “p” with their right
index finger in case of a right-facing arrow. Responses were
to be made within the 1000-msec window during which the
stimulus was presented on the screen. Each trial was
followed by a 1500-msec intertrial interval. If no response
was made during the response window, the first 1000 毫秒
of the intertrial interval included a red “Too Slow!” message
presented on the screen. In stop-signal blocks, a stop-
signal (IE。, the black go-signal arrow changing to red color)
was presented after the go-signal on 25% 的考验. The stop-
signal delay was initially set to 200 msec and was then sub-
sequently adjusted in steps of 50 毫秒 (which were added
to the stop-signal delay after successful stop-trials and sub-
tracted after failed stop-trials), with the goal of achieving an
overall p(停止) of approximately .5. Participants completed
two blocks of practice with the SST and then performed 10
total blocks, alternating between stop-signal blocks
(“There will be stop-signals. Responding quickly on go-
trials and stopping successful on stop-trials are equally
important.”) and pure-go blocks (“There will be no stop-
signals. Respond as fast as possible.”). We altered the type
of the first block after each participant to counterbalance
the order. To achieve a balanced number of go-trials from
each task context, each block contained 48 go-trials. 在
添加, the stop-signal blocks contained 16 stop-trials.
总共, this resulted in 240 go-trials from pure-go blocks,
240 go-trials from stop-signal blocks, 和 80 stop-trials per
参与者.
Behavioral Analysis
Variables of interest were go-trial RT from both task contexts
and SSRT, failed stop RT, 和 p(停止) from the stop-signal
blocks. Within the stop-blocks, 去- and failed stop-trial RTs
were compared to ensure that the requirements of the
horse race model were met (failed stop RT < go-trial RT;
Logan et al., 1984). p(Stop) was investigated to ensure that
the stop-signal delay staircase algorithm was effective
in achieving an approximate stopping success rate of .5.
SSRT was calculated using the integration method with
replacement of go-trial omission errors (Verbruggen et al.,
2019).
The primary behavioral variable of interest to the main
hypothesis was the relative slowing of go-trial RT that was
expected in stop-signal blocks compared to pure-go blocks
(proactive RT slowing). First, these go-trial RTs from both
task contexts were compared with a paired-samples t test.
For the purposes of the brain–behavior correlation, a nor-
malized proactive RT slowing value was then computed for
each participant. We used a percent-change measure that
normalizes the RT condition difference by the pure-go-
trial RT, to account for baseline differences in RT. The
resulting formula:
Proactive RT slowing
¼ 100 * ðstop (cid:2) signal block go (cid:2) RT
(cid:2) pure (cid:2) go block go (cid:2) RTÞ
= pure (cid:2) go block go (cid:2) RT
Soh et al.
787
expresses proactive RT slowing as the percentage of
slowing found in stop-signal blocks relative to pure-go
blocks.
EEG Recording and Preprocessing
These steps were performed exactly as described in our
recent paper ( Wessel, 2020). The description is adapted
from therein. In brief, we will discuss them below.
Scalp EEG was recorded using a 64-channel active EEG
system (BrainProducts actiChamp). The ground was
placed at electrode Fz, and the reference was placed at
electrode Pz. Sampling rate was 500 Hz, with hardware
filters set to time-constant high-pass 10 sec and low-pass
1000 Hz.
Data were preprocessed using custom routines in
MATLAB, incorporating functions from EEGLAB (Delorme
& Makeig, 2004). The data were imported into MATLAB and
then filtered using symmetric two-way least-squares finite
impulse response filters (high-pass cutoff: 0.3 Hz, low-pass
cutoff: 30 Hz). Nonstereotyped artifacts were automatically
removed from further analysis using segment statistics
applied to each second-long segment of data joint proba-
bility and joint kurtosis, with both cutoffs set to 5 SDs (cf.
Delorme, Sejnowski, & Makeig, 2007). Trials that included
a rejected data segment were excluded from further anal-
ysis. After removal of nonstereotypic artifacts, the data
were rereferenced to common average and subjected to
a temporal infomax independent component analysis
(Bell & Sejnowski, 1995), with extension to sub-Gaussian
sources (Lee, Girolami, & Sejnowski, 1999). Components
representing eye-movement and electrode artifacts were
identified using outlier statistics and were removed from
the data, alongside nondipolar components’ residual vari-
ance cutoff at 15% (Delorme, Palmer, Onton, Oostenveld,
& Makeig, 2012). The remaining components were back-
projected into channel space and subjected to further
analyses. The data were then transformed to a reference-
free montage using the current source density (CSD)
method (Tenke & Kayser, 2005; Perrin, Pernier, Bertrand,
& Echallier, 1989).
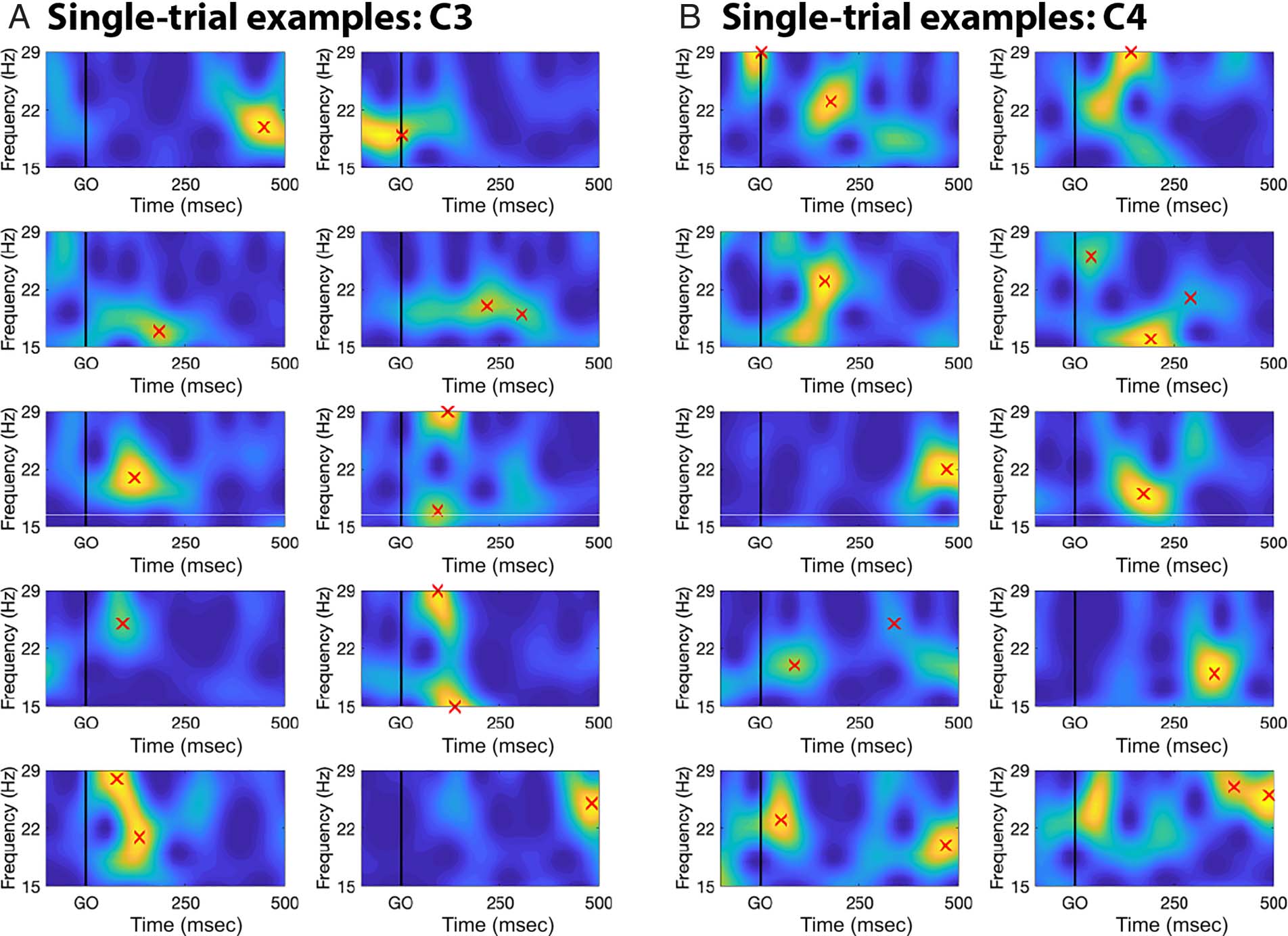
β-Burst Detection
Sensorimotor β-bursts from go-trials in both contexts were
quantified at electrode sites C3 and C4 from the CSD-
converted montage. β-burst detection was performed as
described in Shin, Law, Tsutsui, Moore, and Jones (2017)
and implemented in our previous paper for human scalp
EEG ( Wessel, 2020). The description is adapted from
therein.
First, each electrode’s data were convolved with a com-
plex Morlet wavelet of the form:
Þ ¼ A exp − t2
2σ2
t
w t; fð
(cid:1)
(cid:3)
ð
exp 2iπft
Þ
p
ffiffiffiffiffi
2π
; A ¼ 1
σt
with σ ¼ m
, and m = 7 (cycles) for each of 15
2πf
evenly spaced frequencies spanning the β-band (15–29 Hz).
Time–frequency power estimates were extracted by calcu-
lating the squared magnitude of the complex wavelet-
convolved data. These power estimates were then epoched
relative to the events in question (ranging from −500 to
+1000 msec with respect to go-signals). Individual β-bursts
were defined as local maxima in the trial-by-trial β-band
time–frequency power matrix for which the power ex-
ceeded a set cutoff of 6× the median power of the entire
time–frequency epoch power matrix for that electrode
(i.e., the median thresholding was performed across all
epochs). Local maxima within each 1500-msec epoch were
identified using the MATLAB function imregional().
Example trials from one participant are depicted in
Figure 2. Although the median thresholding procedure
provides a rather coarse and simplistic threshold that
is invariant across participants, this procedure matches
work in both magnetoencephalography (Shin et al.,
2017) and EEG ( Wessel, 2020). Hence, this approach is
consistent with existing literature—although further
method development in this regard is necessary to iden-
tify more sophisticated and flexible β-burst detection
procedures.
Statistical Analyses
As in our previous study (Wessel, 2020), the rate of β-bursts
at both electrode sites was quantified in bins, time-locked
to the onset of the go-signal. This binning procedure was
performed after the initial burst detection (described
above)—that is, each burst was uniquely assigned to one
bin according to its timing. Each of the five bins was 100 msec
in length, together spanning the first 500 msec after the
go-signal. This bin size was chosen because it represented
an ideal trade-off between the dynamics of the sensorimotor
β-burst counts and the amount of statistical tests to be per-
formed. Within each bin, we compared the amount of
sensorimotor β-bursts using a 2 × 2 repeated-measures
ANOVA with the two factors Context (stop-signal vs. pure-go
blocks) and Side (contralateral vs. ipsilateral to the cued
response). Because this resulted in five separate ANOVAs
(one for each bin), the resulting p values were corrected
for multiple comparisons using the false discovery rate
procedure (FDR; Benjamini, Krieger, & Yekutieli, 2006).
Moreover, to test whether the purported increase in sen-
sorimotor β-bursts was sustained throughout the entire
stop-signal block or whether sensorimotor β-bursts were
specifically upregulated during movement initiation, we also
compared the amount of β-bursts in the baseline fixation
period (500 msec before the go-signal) between both
conditions. This was done using a paired-samples t test
(because this measurement was taken before the go-
signal, Laterality was not a factor in this analysis). To enable
a comparison of this measurement to the burst rates during
the 100-msec post-go-signal bins, this measurement was
788
Journal of Cognitive Neuroscience
Volume 33, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
o
c
n
_
a
_
0
1
6
8
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
o
c
n
_
a
_
0
1
6
8
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Demonstration of the burst detection procedure using a random selection of example trials from one random participant, focused on the
β-band. Red crosses mark detected bursts.
converted into “β-bursts per 100 msec” by dividing the
number of β-bursts in the 500-msec fixation period by a
factor of 5.
We tested two types of brain–behavior relationships
using Pearson’s product–moment correlation coefficient.
First, to confirm that sensorimotor β-bursts are meaning-
fully related to movement initiation in both task contexts,
we aimed to replicate our previous finding of a positive
cross-participant correlation between the amount of early
contralateral β-bursts on go-trials and go-trial RT (Wessel,
2020). We tested this correlation for each of the five bins
and the two task contexts separately, correcting the re-
sulting p values for multiple comparisons using the FDR
method. We expected a positive correlation in the early
bins in both task contexts.
Second, to test the main brain–behavior hypothesis re-
garding the relationship between the hypothesized general
β-burst upregulation in the stop-signal blocks and the be-
havioral expression of proactive control, we identified the
bins in which the main 2 × 2 ANOVA revealed a main effect
of Context and no influence of Laterality. The mean β-burst
counts from these bins were then converted to the same
percent-increase measure that was used for the behavioral
go-RT data. The following formula was used to quantify
this measurement for each participant:
β(cid:2)burst increase ¼ 100
ð
* stop (cid:2) signal block β(cid:2)bursts
(cid:2) pure (cid:2) go block β(cid:2)burstsÞ
= pure (cid:2) go block β(cid:2)bursts:
We hypothesized that there would be a significantly
positive correlation between the percentage of increase
in sensorimotor β-bursting found on stop-signal versus
pure-go blocks and proactive RT slowing (quantified as
explained in the Behavioral Analysis section).
RESULTS
Behavior
In line with the predictions from the horse race model of
the SST, all participants showed longer go-trial RTs (mean =
553 msec, SD = 117) compared to failed stop-trial RTs
(mean = 488 msec, SD = 114) in the stop-signal blocks.
Soh et al.
789
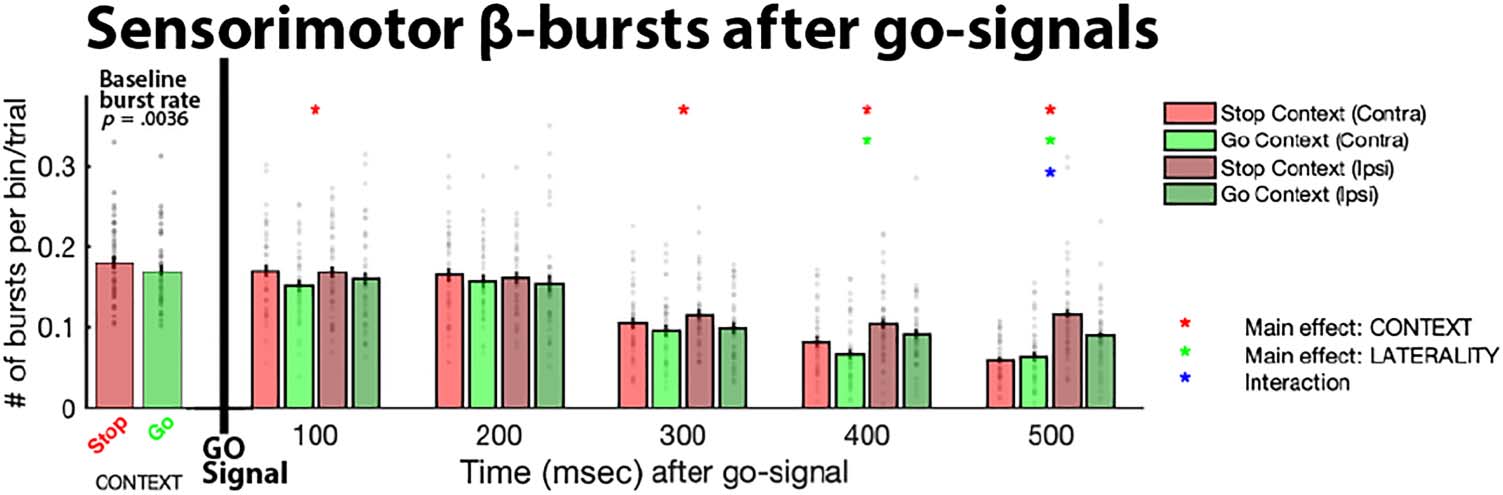
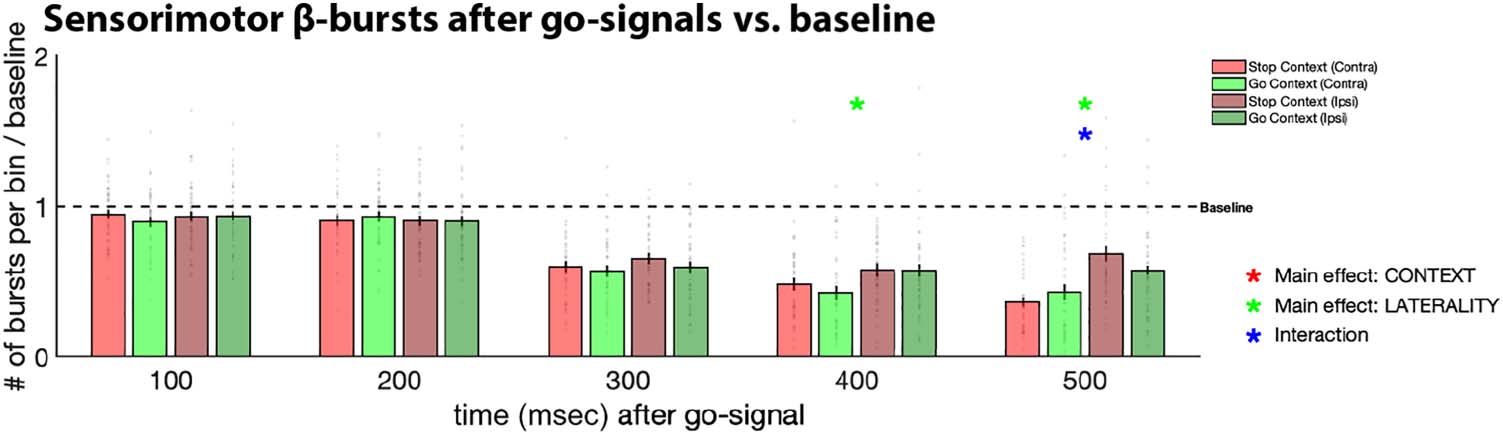
Figure 3. Sensorimotor
β-bursts in five consecutive time
bins after go-signals in both
task contexts, separately for
contralateral (Contra) and
ipsilateral (Ipsi) electrode sites,
as well as during the pre-go
baseline period (where the
bars express β-burst rate per
100 msec to match the scaling
of the post-go period). Gray
dots represent individual
participant means; black bars
denote the standard error of the
group mean. Asterisks denote significance of the respective main effect or interaction within the specified bin, corrected for multiple comparisons
using FDR to a family-wise p < .05. For the baseline period, the exact p value (.0036) is listed above the bar plot.
p(stop) was .53 (SD = .05), indicating the effectiveness of
the staircase procedure. Mean SSRT was 237 msec (SD = 30).
In line with our hypothesis and prior studies, go-trial RT
was strongly affected by task context (stop-signal blocks:
553 msec, SD = 117; pure-go blocks: 393 msec, SD = 50;
t(40) = 10.16, p = 1.21*10−12, d = 1.81), indicating an
average percent increase of 41.23% (SD = 27%) that ranged
from 2.26% to 108% in individual participants.
Post-go Sensorimotor β-Burst Condition Averages
In line with our main hypothesis, sensorimotor β-bursts
were increased in stop-signal blocks compared to pure-
go blocks (Figure 3), moreover at both contralateral and
ipsilateral electrode sites. After FDR correction, there was
a significant main effect of Context in Bins 1 and 3–5.
Moreover, in line with our previous report (Wessel, 2020),
the contralateral electrode site showed lower β-burst
rates in the bins toward the end of the preresponse
period, resulting in a significant main effect of Laterality in
Bins 4 and 5 and a significant Context × Laterality interac-
tion in Bin 5. Notably, no effect of Laterality was found
during the early bins, showing that β-bursts were increased
in the stop-signal context over both the ipsilateral and con-
tralateral electrode sites. Test statistics and p values for all
three factors across the five bins can be found in Table 1.
In response to a reviewer suggestion, we also explored
other burst parameters (burst duration, frequency range,
and power), although these variables have not been found
to relate to behavior to the same extent as burst rate (Shin
et al., 2017). Indeed, neither burst duration nor burst
power showed any FDR-corrected significant task context
effects in either of the windows. Burst frequency range did
show a significant increase in the stop-signal context in the
200- to 300-msec window ( p = .004, η2 = .19). Although we
had no a priori hypothesis regarding the interpretation of
the burst frequency range parameter, one possible expla-
nation for this result could be that the increase in burst rate
in this window could lead to instances in which two bursts
occur in neighboring frequency bands, with only one ex-
ceeding the median power threshold for detection (cf.
top right trial example in Figure 2). However, at this point,
this is conjectural.
Brain–Behavior Correlations: β-Burst Rates vs.
Basic RT
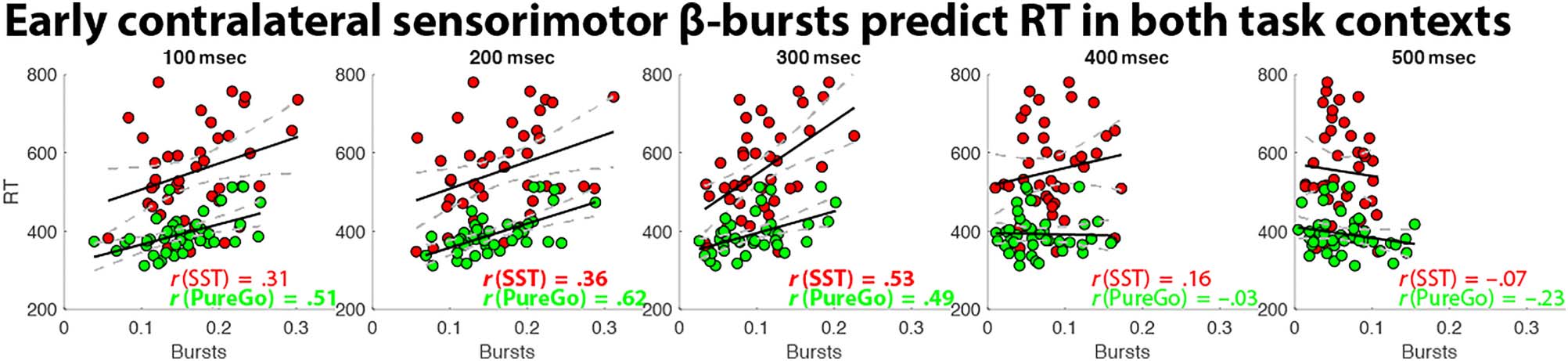
Replicating our previous investigation (Wessel, 2020), early
contralateral β-bursts after the go-signal predicted RT
across participants (Figure 4). Importantly, extending
those earlier findings, this was true for both task contexts,
showing that there is a general relationship between
Table 1. Test Statistics for Bin-Wise ANOVAs of Sensorimotor β-Bursts After Go-Signals
Context
p
.04
.22
.02
.02
.03
F
4.29
1.54
6.37
6.42
4.95
η 2
.10
.04
.14
.14
.11
Variable
Laterality
p
.51
.54
.10
.00
.00
F
0.43
0.39
2.86
16.68
31.56
η 2
.01
.01
.07
.29
.44
Interaction
F
0.97
0.01
0.93
0.07
12.65
p
.33
.91
.34
.79
.00
η 2
.02
.00
.02
.00
.24
Bin
0–100
100–200
200–300
300–400
400–500
p Values are uncorrected; bold print denotes significant tests after FDR correction.
790
Journal of Cognitive Neuroscience
Volume 33, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
o
c
n
_
a
_
0
1
6
8
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
o
c
n
_
a
_
0
1
6
8
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Brain–behavior correlations between go-trial RT and contralateral sensorimotor β-bursts in both task contexts. Across both tasks, early
levels of β-bursting positively predicted RT, with increased bursting leading to slower RTs. Values in bold print were significant after FDR correction
to a family-wise p = .05 (see Table 2 for exact p values).
sensorimotor β-bursts and basic RT that is not specific to
whether proactive inhibitory control is present. After FDR
correction, positive correlations were observable between
β-bursts and go-trial RT in Bins 2 and 3 for stop-signal
blocks and in Bins 1–3 for pure-go blocks (Figure 4 and
Table 2).
Brain–Behavior Correlations: Context-dependent
β-Burst Increase vs. Proactive RT Slowing
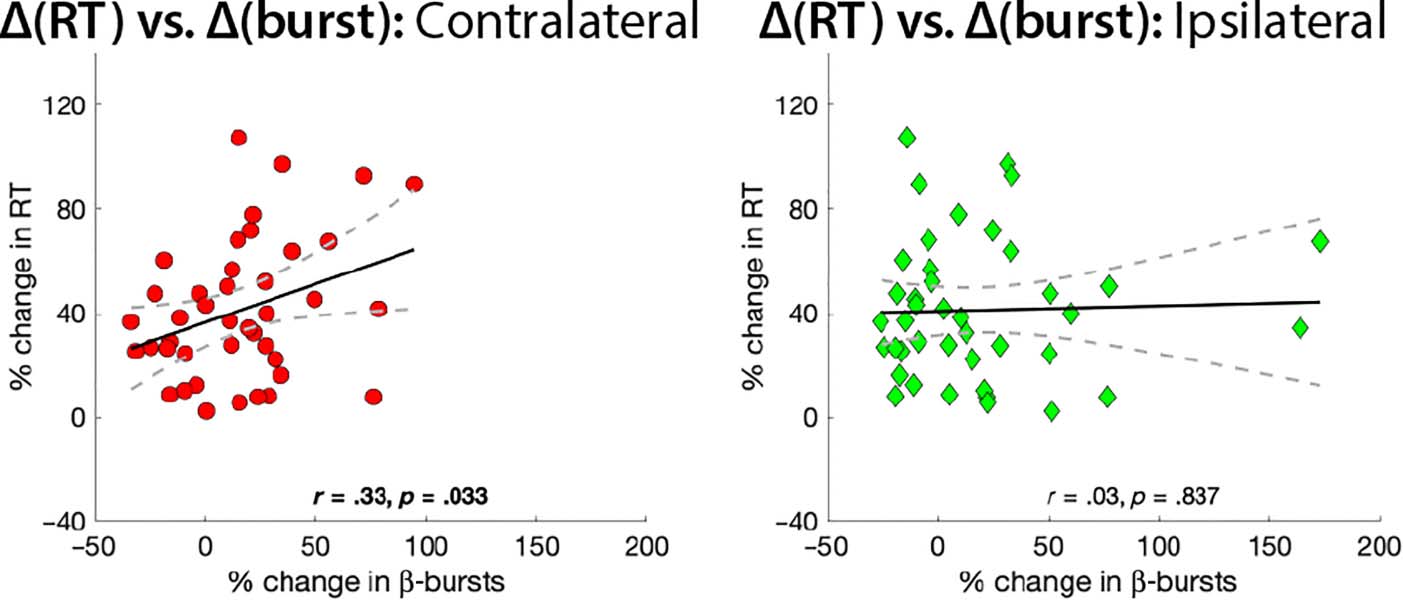
In line with the main hypothesis of the current study, there
was also a significantly positive relationship between the
degree of increase in sensorimotor β-burst rate each partic-
ipant showed in the stop-signal compared to the pure-go
context and the degree to which they slowed their
responses in the stop-signal context (Figure 5). Notably,
this was only true for the contralateral β-bursts (r = .33,
p < .05), whereas the ipsilateral β-bursts did not show
the same relationship (r = .03, p = .84).
Exploratory Analysis: Pre-go Baseline
In addition to the β-burst rates that occurred in the move-
ment initiation period after the go-signal (the investigation
Table 2. Test Statistics for Bin-Wise Correlations between
Contralateral Sensorimotor β-Bursts and Go-Trial RT Across
Both Task Contexts
Context
Stop-Signal
Pure-go
r
.310
.364
.532
.161
−.071
p
.045
.019
.000
.314
.659
r
.505
.620
.494
−.032
−.226
p
.001
.000
.001
.844
.155
Bin
0–100
100–200
200–300
300–400
400–500
Bold print signifies significant tests after FDR correction to a family-wise
p < .05.
of which was motivated by our previous study; Wessel,
2020), we also explored whether potential differences in
β-burst rates between the stop- and pure-go contexts were
already present before the go-signal. This would indicate
that proactive control is tonically deployed throughout
the task, whereas a null finding would indicate that proac-
tive control is deployed in a more transient, phasic fashion
during movement initiation itself. In line with the former
interpretation, prestimulus sensorimotor β-burst rate in
stop-blocks was 0.181 bursts per 100 msec (SD = 0.049),
whereas the same period in pure-go blocks showed a
rate of 0.171 bursts per 100 msec (SD = 0.046, t(40) =
3.1, p = .0036, d = 0.22). This shows that sensorimotor
β-bursting was increased even before the to-be-initiated
movement was known or cued. However, unlike burst
rates in the movement initiation period (Figure 5), this
pre-go baseline increase in sensorimotor β-bursts was
not related to behavior in the form of proactive RT slowing
(r = .06, p = .7). This suggests that, although β is increased
even before the movement is known, this increase only
becomes effective in influencing behavior once an actual
movement has been cued and is initiated.
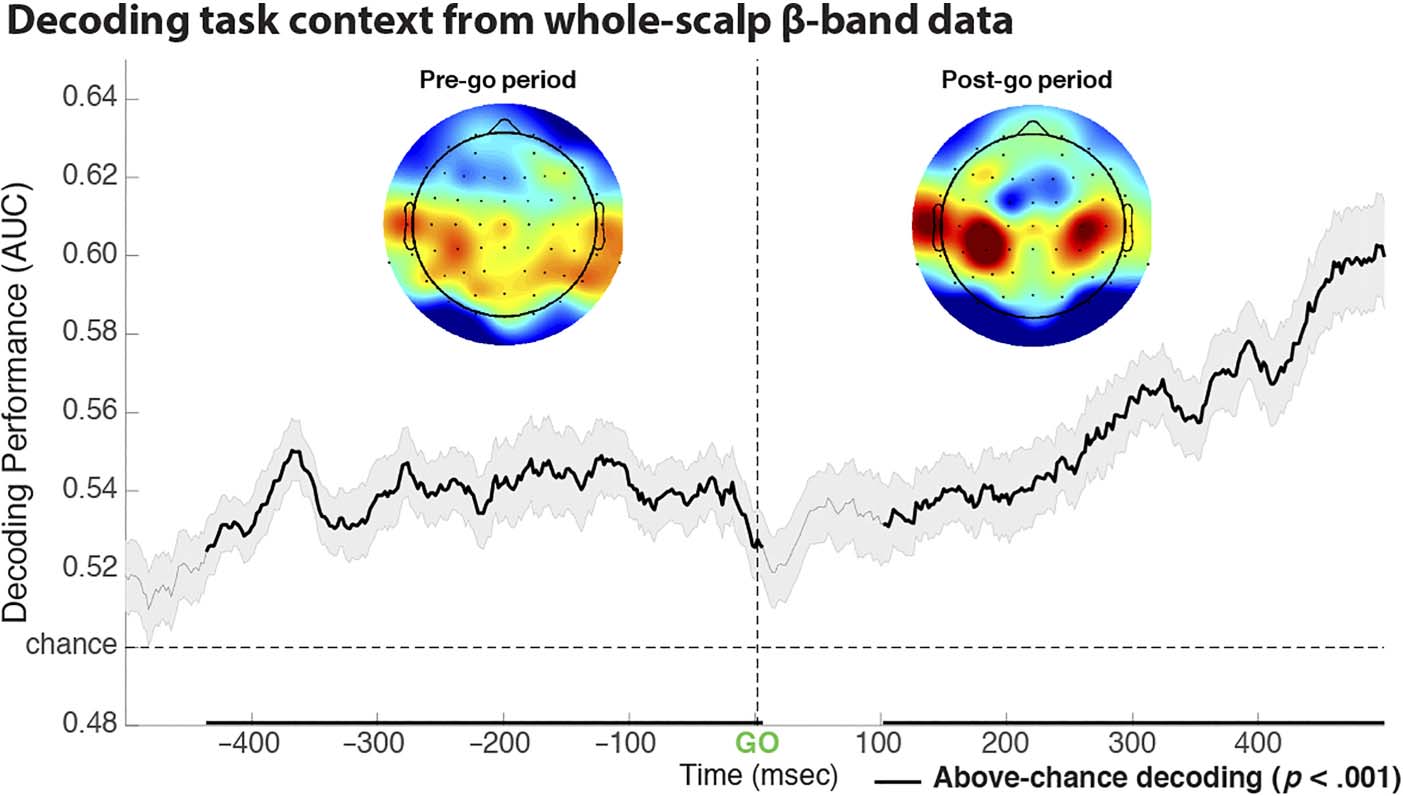
Exploratory Analysis: Multivariate Pattern Analysis
of the Pre-go Baseline Period
To confirm the abovementioned exploratory finding of in-
creased sensorimotor β-bursting during the prestimulus
baseline period in stop-signal blocks, we attempted to rep-
licate the finding using a complete different analysis
approach. Specifically, we explored if the task context (stop
or pure-go) could be decoded from the whole-scalp
β-band activity even before go-stimulus onset (i.e., during
the baseline). These analyses were performed using the
multivariate pattern analysis (MVPA) method implemented
in the ADAM toolbox (Fahrenfort, Van Driel, Van Gaal, &
Olivers, 2018).
To this end, we used the same data as for the abovemen-
tioned β-burst analysis (i.e., the CSD-transformed and
cleaned data on which the burst detection was performed)
and epoched the β-band amplitude signal for each trial
(15–29 Hz, extracted using the seven-cycle Morlet wavelet
Soh et al.
791
Figure 5. Correlation between
the change in early β-bursting
between both task contexts
(during the bins that showed a
significant increase in average
burst rate after FDR correction;
cf. Figure 3) and proactive RT
slowing across participants,
separately for contralateral and
ipsilateral electrode sites.
described above) from −500 to 500 msec around the go-
stimulus. To avoid biasing the classifier, trial counts were
equated across both conditions by removing a random se-
lection of trials from the condition with more exemplars.
To train and test the classifier, we used linear discriminant
analysis (Grootswagers, Wardle, & Carlson, 2017) on the
single-trial data for each participant, with the quality of
the classification evaluated with leave-one-out cross-
validation. We then quantified the area under the resultant
receiver operating characteristic curve (i.e., the cumulative
probability of correct classification plotted against the cu-
mulative probability of false-positive classification) for this
validation procedure. Each participant’s resulting vector of
area under the resultant receiver operating characteristic
curve data at each time point was then submitted to sample
point-wise paired-samples t test against chance (.5), cor-
rected for multiple comparisons using cluster-based per-
mutation testing (Maris & Oostenveld, 2007; 10,000
iterations, cluster p value = .05, alpha = .001).
Confirming the findings from the β-burst analysis, the
MVPA showed that the task condition could indeed be
successfully decoded from β activity. In the post-go period,
decoding accuracy notably increased in the later periods,
which likely reflects the earlier occurrence of postmove-
ment β rebound ( Jurkiewicz, Gaetz, Bostan, & Cheyne,
2006; Pfurtscheller, Neuper, Brunner, & Da Silva, 2005)
in the pure-go condition (which contained faster responses
on average). Notably, however, in line with the exploratory
β-burst analysis, classification was already above chance
during the pre-go baseline period (Figure 6). Moreover,
the topography of the forward-reconstructed weight
matrix reveals that most of the contribution to decoding
performance came from sensorimotor sites. This confirms
the analysis from the exploratory burst analysis.
Furthermore, identical analyses of theta (5–8 Hz) and
alpha (9–12 Hz) activity showed that neither frequency
band showed above-chance decoding in the pre-go base-
line period, underscoring that β-band activity in the base-
line period is uniquely related to proactive control.
Exploratory Analysis: Baseline-Normalized Post-go
β-Burst Changes
Following up on the finding that differences in sensorimo-
tor β-burst rates were already present in the prestimulus
baseline period, we also reinvestigated the post-go data.
Figure 6. Using whole-scalp
β-band (15–29 Hz) data to
decode task conditions using
MVPA. Confirming the results
from both the hypothesis-
driven analysis of the post-go
β-band burst data and the
exploratory analysis of the
pre-go baseline period, task
context (stop vs. pure-go)
could be successfully decoded
from β-band data in both
periods. Moreover, forward-
reconstructed decoding weight
matrices at the significant
clusters in both time ranges
show that most explanatory
power was provided by activity
over bilateral sensorimotor sites.
792
Journal of Cognitive Neuroscience
Volume 33, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
o
c
n
_
a
_
0
1
6
8
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 3. Test Statistics for Bin-Wise ANOVAs of Sensorimotor β-Bursts After Go-Signals, Normalized by Baseline (cf. Figure 5)
Context
p
.52
.74
.12
.35
.52
F
0.42
0.11
2.59
0.91
0.41
η 2
.01
.00
.06
.02
.01
Variable
Laterality
p
.68
.62
.13
.00
.00
F
0.17
0.25
2.33
15.49
26.28
η 2
.00
.01
.06
.28
.40
Interaction
F
0.69
0.24
0.40
0.69
9.81
p
.41
.63
.53
.41
.00
η 2
.02
.01
.01
.02
.20
Bin
0–100
100–200
200–300
300–400
400–500
p Values are uncorrected; bold print denotes significant tests after FDR correction.
In particular, instead of investigating raw burst counts, we
reran the same analyses performed in the main hypothesis
test, but normalizing each bin and condition by the appro-
priate baseline burst rate (i.e., contralateral post-go activity
for a left-hand response trial in the stopping condition was
normalized by the mean baseline burst rate in the stopping
condition at electrode C4, and so forth). This revealed that,
although the late-period effects of laterality (in the 300- to
400-msec and 400- to 500-msec windows) and the interac-
tion in the final bin (400–500 msec) remained significant,
none of the five post-go bins retained any significant burst
rate differences between the two task contexts (i.e., be-
tween stop and pure-go blocks). The exact statistics for
each bin can be found in Table 3, and the data are visualized
in Figure 7.
What this shows is that the post-go differences in raw
burst rates (Figure 1) can be, at least to a substantial de-
gree, attributed to condition differences that are already
present in the baseline burst rates. In other words, it
appears that broad, nonselective inhibitory control in the
absence of foreknowledge is implemented by a global in-
crease of β-bursting both before and after a signal to initiate
a movement. Notably, the main brain–behavior analysis in
the hypothesis-testing part of this study showed that the
contralateral β-burst rate difference in the post-go period
did predict proactive RT slowing—that is, the behavioral
expression of proactive inhibition (this association
remained significant when the data were corrected for
baseline differences as well; r = .35, p = .027 ).
Importantly, as noted above, this was not the case for the
burst-rate condition differences in the pre-go baseline
period.
DISCUSSION
β-Band activity is a fundamental neurophysiological signa-
ture of the motor system. Our current study shows that,
when human participants strategically implement proac-
tive inhibitory motor control in anticipation of potentially
having to stop a movement, β-burst rates over bilateral sen-
sorimotor cortex were increased. Further analyses showed
that this relative increase was already present in the base-
line period—that is, before the movement in question was
known or cued. Indeed, this was further underscored by
MVPAs that demonstrated that the task condition (and
the associated engagement of proactive control) could
already be successfully decoded from β-band activity that
occurred before any signal to initiate a response (i.e., before
the go-signal). Brain–behavior correlation analyses then re-
vealed that this tonic increase in sensorimotor β-bursting
throughout the task period affected behavior specifically
during the movement initiation period: Participants with
greater relative increases in sensorimotor β-bursting
Figure 7. Baseline-normalized
version of Figure 3. Sensorimotor
β-bursts in five consecutive time
bins after go-signals in both
task contexts, separately for
contralateral (Contra) and
ipsilateral (Ipsi) electrode sites.
Gray dots represent individual
participant means; black bars
denote the standard error of the
group mean. Asterisks denote
significance of the respective
main effect or interaction within
the specified bin, corrected for
multiple comparisons using FDR
to a family-wise p < .05.
Soh et al.
793
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
o
c
n
_
a
_
0
1
6
8
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
between the two task contexts also showed greater proac-
tive RT slowing.
The current data have several notable features. First, in-
dependent of whether proactive control was engaged or
not, sensorimotor β-burst rates during movement initia-
tion predicted RTs in both task contexts (Figure 4). This
confirms that β-bursts may reflect a general signature of a
net-inhibited motor system, which has to be overcome to
initiate a movement (Pfurtscheller et al., 1997). Moreover,
it speaks toward recent conceptualizations of motor inhibi-
tion as a generic, universal mechanism that is involved in
movement planning, with the degree of its deployment de-
pending on—among other things—proactive control set-
tings (Raud et al., 2020; Greenhouse, Sias, Labruna, &
Ivry, 2015). Second, adjustments to proactive inhibitory
control resulted in an increase of sensorimotor burst rates
both before and after a signal to initiate a movement. This
shows that humans adjust the overall, general inhibition of
their corticomotor system when they anticipate that they
may have to stop an action after its initiation, even when
they have no foreknowledge of the specific movement that
they may have to stop. Although this bilateral increase in
net inhibition of the motor system is already observable
before the go-signal even appeared, it does appear to be
specifically implemented in service of inhibiting behavior
during the movement initiation period (which is when it
becomes predictive of condition-related increases in pro-
active RT slowing). Moreover, whereas the overall increase
in β activity was observed bilaterally, the predictive rela-
tionship between β and RT was specific to contralateral β
increases. Hence, humans appear to nonselectively deploy
proactive inhibitory control as a broad task setting—both
during and before movement initiation, and both at contra-
lateral and ipsilateral motor sites—but with the goal of
achieving a site-specific inhibition of the to-be-performed
movement specifically during its initiation. Third, the pre-
go baseline increase in sensorimotor β-burst rates suggests
that, in the absence of foreknowledge, humans implement
proactive inhibitory control via a sustained increase of sen-
sorimotor β-bursting, instead of a transient engagement of
β-bursting that is specific to the movement initiation period.
A highly notable implication of this finding is that studies
that seek to identify the cognitive control signatures that
produce the ultimate effect of increased sensorimotor
β-bursting observed here should avoid baseline-corrected
contrasts of post-go activity. Instead, potential condition
differences in the upstream processes should be investigated
as sustained differences that are likely present throughout
most (or all) of the respective task periods, not just during
movement initiation.
The current study shows that proactive inhibitory con-
trol in the context of the SST is implemented at the level
of the motor system by sustaining the inhibited state of
the sensorimotor cortex past the signal to initiate an action.
In other words, if a movement is executed under an in-
creased amount of caution, the corticomotor system re-
mains inhibited for an extended period. This is in line
with existing literature that used TMS to probe the net ex-
citability of entire corticospinal tracts during stop-signal-
type tasks (Duque et al., 2017; Cowie, MacDonald, Cirillo,
& Byblow, 2016; Greenhouse et al., 2011; Claffey et al.,
2010). Specifically, these studies show that the implemen-
tation of proactive motor inhibition directly affects the mo-
tor system and is not solely attributable to attentional
processes that aid stop-signal detection. Whereas most of
these studies have shown that this is the case when partic-
ipants have specific foreknowledge of the to-be-inhibited
response, the current results show that this is also true
when the to-be-inhibited response is unknown ahead of
time. More generally, these findings support the view that
sensorimotor β-band activity reflects an inhibited state of
the motor system (Schulz et al., 2013; Zhang et al., 2008;
Pfurtscheller et al., 1997).
Notably, motor caution is an executive function that is
implemented in many controlled behaviors that occur even
outside SSTs. Indeed, increases in motor threshold are pur-
ported to underlie more cautious, controlled responding in
a slew of cognitive control scenarios that are subject to sci-
entific study, including error commission (Fischer, Nigbur,
Klein, Danielmeier, & Ullsperger, 2018; Ullsperger &
Danielmeier, 2016; Ridderinkhof, 2002; Ridderinkhof et al.,
2002), response conflict (Wessel, Waller, & Greenlee, 2019;
Klein, Petitjean, Olivier, & Duque, 2014; Burle, Possamaï,
Vidal, Bonnet, & Hasbroucq, 2002; Botvinick, Braver,
Barch, Carter, & Cohen, 2001), task switching (Mansfield,
Karayanidis, Jamadar, Heathcote, & Forstmann, 2011), and
decision conflict (Cavanagh et al., 2011; Frank, 2006). It is
tempting to assume that a global, bilateral increase of
sensorimotor β activity could reflect an increase in overall
response threshold, implemented via a broad recruit-
ment of proactive inhibitory control. Indeed, along those
lines, some studies have recently connected sensorimotor
β-band activity to adjustments in response thresholds in
situations that demand cognitive control, even outside
explicit stop-signal scenarios (e.g., Fischer et al., 2018;
Tan, Wade, & Brown, 2016). The potential universality of
this signature outside the SST and its direct relationship to
the inhibition of the motor system therefore yields a
promising avenue for future study.
On the methodological side, the current study provides
another demonstration of the recent development toward
investigating sensorimotor β-band activity as rate-modulated
burst-like activity, rather than as amplitude-modulated
oscillations (e.g., Wessel, 2020; Shin et al., 2017; Sherman
et al., 2016; Feingold et al., 2015), a notion for which there
is now converging evidence from both the rodent and pri-
mate literature (both in the human and nonhuman realms).
In our previous investigation of the SST, we have found that
burst quantifications can reveal brain–behavior correlations
that are absent when quantifying the signal in a classic
amplitude-modulated fashion (Wessel, 2020). Indeed, the
clarity of the brain–behavior relationships in the current
study (both in terms of simple RT and even in terms of stra-
tegic changes in RT across two different task contexts)
794
Journal of Cognitive Neuroscience
Volume 33, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
o
c
n
_
a
_
0
1
6
8
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
suggests that even noninvasive scalp recordings can, with-
out any additional preprocessing or specific spatial filters,
provide sufficient power to investigate movement-related
brain activity, if β is investigated using a burst-based quanti-
fication. Moreover, there is a growing body of recent human
clinical literature suggesting that even subcortical β activity
(e.g., in the BG) may be better characterized by short, tran-
sient bursts, whose features index specific motor-related
symptoms in populations with movement disorders (Little,
Bonaiuto, Barnes, & Bestmann, 2019; Anidi et al., 2018;
Tinkhauser et al., 2017). Given that both proactive and reac-
tive control deficits are well-known symptoms of such disor-
ders (Benis et al., 2014; Obeso et al., 2013, 2014; van den
Wildenberg et al., 2006), burst-based quantifications of
β-band activity from both invasive and noninvasive record-
ings yield a tremendous potential for future clinical investi-
gations of abnormalities in inhibitory control and their
neurophysiological basis.
In summary, the current study provides strong evidence
for the fact that sensorimotor β-bursting is a general signa-
ture of an inhibited motor system, which can be globally
upregulated to implement inhibitory motor control, even
when the specific motor effector to be inhibited is not
known ahead of time. The results also provide further ev-
idence for the view that proactive inhibitory control is
achieved via direct effects on the motor system and against
the view that proactive control is solely achieved by atten-
tional tuning.
Reprint requests should be sent to Jan R. Wessel or Cheol Soh,
Department of Psychological and Brain Sciences, University of
Iowa, 444 MRC, Iowa City, IA 52242, or via e-mail: Jan-Wessel
@uiowa.edu, Cheol-Soh@uiowa.edu.
Author Contributions
Cheol Soh: Conceptualization; Formal analysis; Investigation;
Visualization; Writing—Review & editing. Megan Hynd:
Investigation; Writing—Review & editing. Benjamin Rangel:
Investigation; Writing—Review & editing. Jan Wessel:
Conceptualization; Funding acquisition; Investigation;
Project administration; Supervision; Visualization; Writing—
Original draft; Writing—Review & editing.
Funding Information
National Institute of Neurological Disorders and Stroke
(http://dx.doi.org/10.13039/100000065), grant number: R01
NS117753. National Science Foundation (http://dx.doi
.org/10.13039/100000001), grant number: CAREER 1752355.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed a
persistent pattern of gender imbalance: Although the
proportions of authorship teams (categorized by estimated
gender identification of first author/last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/W = .108, and W/W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/W = .102, and W/W = .076
(Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently, JoCN
encourages all authors to consider gender balance explicitly
when selecting which articles to cite and gives them the
opportunity to report their article’s gender citation balance.
REFERENCES
Anidi, C., O’Day, J. J., Anderson, R. W., Afzal, M. F., Syrkin-Nikolau,
J., Velisar, A., et al. (2018). Neuromodulation targets pathological
not physiological β bursts during gait in Parkinson’s disease.
Neurobiology of Disease, 120, 107–117. DOI: https://doi.org
/10.1016/j.nbd.2018.09.004, PMID: 30196050, PMCID:
PMC6422345
Aron, A. R. (2011). From reactive to proactive and selective
control: Developing a richer model for stopping inappropriate
responses. Biological Psychiatry, 69, e55–e68. DOI: https://
doi.org/10.1016/j.biopsych.2010.07.024, PMID: 20932513,
PMCID: PMC3039712
Aron, A. R., Robbins, T. W., & Poldrack, R. A. (2014). Inhibition
and the right inferior frontal cortex: One decade on. Trends
in Cognitive Sciences, 18, 177–185. DOI: https://doi.org/10
.1016/j.tics.2013.12.003, PMID: 24440116
Baker, S. N. (2007). Oscillatory interactions between
sensorimotor cortex and the periphery. Current Opinion in
Neurobiology, 17, 649–655. DOI: https://doi.org/10.1016
/j.conb.2008.01.007, PMID: 18339546, PMCID: PMC2428102
Bell, A. J., & Sejnowski, T. J. (1995). An information-maximization
approach to blind separation and blind deconvolution. Neural
Computation, 7, 1129–1159. DOI: https://doi.org/10.1162/neco
.1995.7.6.1129, PMID: 7584893
Benis, D., David, O., Lachaux, J.-P., Seigneuret, E., Krack, P.,
Fraix, V., et al. (2014). Subthalamic nucleus activity dissociates
proactive and reactive inhibition in patients with Parkinson’s
disease. Neuroimage, 91, 273–281. DOI: https://doi.org/10
.1016/j.neuroimage.2013.10.070, PMID: 24368260
Benjamini, Y., Krieger, A. M., & Yekutieli, D. (2006). Adaptive
linear step-up procedures that control the false discovery rate.
Biometrika, 93, 491–507. DOI: https://doi.org/10.1093/biomet
/93.3.491
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S., &
Cohen, J. D. (2001). Conflict monitoring and cognitive
control. Psychological Review, 108, 624–652. DOI: https://
doi.org/10.1037/0033-295X.108.3.624, PMID: 11488380
Brainard, D. H. (1997). The psychophysics toolbox. Spatial Vision,
10, 433–436. DOI: https://doi.org/10.1163/156856897X00357
Braver, T. S. (2012). The variable nature of cognitive control: A
dual mechanisms framework. Trends in Cognitive Sciences,
16, 106–113. DOI: https://doi.org/10.1016/j.tics.2011.12.010,
PMID: 22245618, PMCID: PMC3289517
Burle, B., Possamaï, C.-A., Vidal, F., Bonnet, M., & Hasbroucq, T.
(2002). Executive control in the Simon effect: An
electromyographic and distributional analysis. Psychological
Research, 66, 324–336. DOI: https://doi.org/10.1007/s00426
-002-0105-6, PMID: 12466929
Cai, W., Chen, T., Ide, J. S., Li, C.-S. R., & Menon, V. (2016).
Dissociable fronto-operculum–insula control signals for
anticipation and detection of inhibitory sensory cue. Cerebral
Soh et al.
795
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
o
c
n
_
a
_
0
1
6
8
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Cortex, 27, 4073–4082. DOI: https://doi.org/10.1093/cercor
/bhw219, PMID: 27473319, PMCID: PMC6059112
Cai, W., George, J. S., Verbruggen, F., Chambers, C. D., & Aron,
A. R. (2012). The role of the right presupplementary motor
area in stopping action: Two studies with event-related
transcranial magnetic stimulation. Journal of Neurophysiology,
108, 380–389. DOI: https://doi.org/10.1152/jn.00132.2012,
PMID: 22514296, PMCID: PMC3404792
Cai, Y., Li, S., Liu, J., Li, D., Feng, Z., Wang, Q., et al. (2015). The
role of the frontal and parietal cortex in proactive and reactive
inhibitory control: A transcranial direct current stimulation
study. Journal of Cognitive Neuroscience, 28, 177–186. DOI:
https://doi.org/10.1162/jocn_a_00888, PMID: 26439269
Cavanagh, J. F., Wiecki, T. V., Cohen, M. X., Figueroa, C. M.,
Samanta, J., Sherman, S. J., et al. (2011). Subthalamic nucleus
stimulation reverses mediofrontal influence over decision
threshold. Nature Neuroscience, 14, 1462–1467. DOI:
https://doi.org/10.1038/nn.2925, PMID: 21946325, PMCID:
PMC3394226
Chikazoe, J., Jimura K, Hirose S, Yamashita, K.-I., Miyashita, Y.,
& Konishi, S. (2009). Preparation to inhibit a response
complements response inhibition during performance of a
stop-signal task. Journal of Neuroscience, 29, 15870–15877.
DOI: https://doi.org/10.1523/JNEUROSCI.3645-09.2009,
PMID: 20016103, PMCID: PMC6666181
Claffey, M. P., Sheldon, S., Stinear, C. M., Verbruggen, F., &
Aron, A. R. (2010). Having a goal to stop action is associated
with advance control of specific motor representations.
Neuropsychologia, 48, 541–548. DOI: https://doi.org/10
.1016/j.neuropsychologia.2009.10.015, PMID: 19879283,
PMCID: PMC2813913
Cowie, M. J., MacDonald, H. J., Cirillo, J., & Byblow, W. D.
(2016). Proactive modulation of long-interval intracortical
inhibition during response inhibition. Journal of
Neurophysiology, 116, 859–867. DOI: https://doi.org/10.1152
/jn.00144.2016, PMID: 27281744, PMCID: PMC4995280
Cunillera, T., Brignani, D., Cucurell, D., Fuentemilla, L., &
Miniussi, C. (2016). The right inferior frontal cortex in
response inhibition: A tDCS–ERP co-registration study.
Neuroimage, 140, 66–75. DOI: https://doi.org/10.1016
/j.neuroimage.2015.11.044, PMID: 26619787
Cunillera, T., Fuentemilla, L., Brignani, D., Cucurell, D., &
Miniussi, C. (2014). A simultaneous modulation of reactive
and proactive inhibition processes by anodal tDCS on the
right inferior frontal cortex. PLoS One, 9, e113537. DOI:
https://doi.org/10.1371/journal.pone.0113537, PMID:
25426713, PMCID: PMC4245139
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21. DOI: https://doi.org/10.1016/j.jneumeth
.2003.10.009, PMID: 15102499
Delorme, A., Palmer, J., Onton, J., Oostenveld, R., & Makeig, S.
(2012). Independent EEG sources are dipolar. PLoS One, 7,
e30135. DOI: https://doi.org/10.1371/journal.pone.0030135,
PMID: 22355308, PMCID: PMC3280242
Delorme, A., Sejnowski, T., & Makeig, S. (2007). Enhanced
detection of artifacts in EEG data using higher-order statistics
and independent component analysis. Neuroimage, 34,
1443–1449. DOI: https://doi.org/10.1016/j.neuroimage.2006
.11.004, PMID: 17188898, PMCID: PMC2895624
Duque, J., Greenhouse, I., Labruna, L., & Ivry, R. B. (2017).
Physiological markers of motor inhibition during human
behavior. Trends in Neurosciences, 40, 219–236. DOI:
https://doi.org/10.1016/j.tins.2017.02.006, PMID: 28341235,
PMCID: PMC5389740
Elchlepp, H., Lavric, A., Chambers, C. D., & Verbruggen, F. (2016).
Proactive inhibitory control: A general biasing account. Cognitive
Psychology, 86, 27–61. DOI: https://doi.org/10.1016/j.cogpsych
.2016.01.004, PMID: 26859519, PMCID: PMC4825542
Engel, A. K., & Fries, P. (2010). β-band oscillations—Signalling the
status quo? Current Opinion in Neurobiology, 20, 156–165. DOI:
https://doi.org/10.1016/j.conb.2010.02.015, PMID: 20359884
Fahrenfort, J. J., Van Driel, J., Van Gaal, S., & Olivers, C. N.
(2018). From ERPs to MVPA using the Amsterdam decoding
and modeling toolbox (ADAM). Frontiers in Neuroscience,
12, 368. DOI: https://doi.org/10.3389/fnins.2018.00368,
PMID: 30018529, PMCID: PMC6038716
Feingold, J., Gibson, D. J., DePasquale, B., & Graybiel, A. M.
(2015). Bursts of β oscillation differentiate postperformance
activity in the striatum and motor cortex of monkeys
performing movement tasks. Proceedings of the National
Academy of Sciences, U.S.A., 112, 13687–13692. DOI: https://
doi.org/10.1073/pnas.1517629112, PMID: 26460033, PMCID:
PMC4640760
Fischer, A. G., Nigbur, R., Klein, T. A., Danielmeier, C., &
Ullsperger, M. (2018). Cortical β power reflects decision
dynamics and uncovers multiple facets of post-error adaptation.
Nature Communications, 9, 5038. DOI: https://doi.org/10.1038
/s41467-018-07456-8, PMID: 30487572, PMCID: PMC6261941
Frank, M. J. (2006). Hold your horses: A dynamic computational
role for the subthalamic nucleus in decision making. Neural
Networks, 19, 1120–1136. DOI: https://doi.org/10.1016/j.neunet
.2006.03.006, PMID: 16945502
Greenhouse, I., Oldenkamp, C. L., & Aron, A. R. (2011). Stopping a
response has global or nonglobal effects on the motor system
depending on preparation. Journal of Neurophysiology, 107,
384–392. DOI: https://doi.org/10.1152/jn.00704.2011, PMID:
22013239, PMCID: PMC3349702
Greenhouse, I., Sias, A., Labruna, L., & Ivry, R. B. (2015).
Nonspecific inhibition of the motor system during response
preparation. Journal of Neuroscience, 35, 10675–10684.
DOI: https://doi.org/10.1523/JNEUROSCI.1436-15.2015,
PMID: 26224853, PMCID: PMC4518047
Greenhouse, I., & Wessel, J. R. (2013). EEG signatures associated
with stopping are sensitive to preparation. Psychophysiology,
50, 900–908. DOI: https://doi.org/10.1111/psyp.12070, PMID:
23763667, PMCID: PMC3745550
Grootswagers, T., Wardle, S. G., & Carlson, T. A. (2017). Decoding
dynamic brain patterns from evoked responses: A tutorial on
multivariate pattern analysis applied to time series neuroimaging
data. Journal of Cognitive Neuroscience, 29, 677–697. DOI:
https://doi.org/10.1162/jocn_a_01068, PMID: 27779910
Hynd, M., Soh, C., Rangel, B. O., & Wessel, J. R. (2021). Paired-
pulse TMS and scalp EEG reveal systematic relationship
between inhibitory GABAa signaling in M1 and fronto-central
cortical activity during action stopping. Journal of Neural
Physiology, 125, 648–660. DOI: https://doi.org/10.1152/jn
.00571.2020
Jaffard, M., Longcamp, M., Velay, J.-L., Anton, J.-L., Roth, M.,
Nazarian, B., et al. (2008). Proactive inhibitory control of
movement assessed by event-related fMRI. Neuroimage, 42,
1196–1206. DOI: https://doi.org/10.1016/j.neuroimage.2008
.05.041, PMID: 18588986
Jahanshahi, M., Obeso, I., Rothwell, J. C., & Obeso, J. A. (2015).
A fronto–striato–subthalamic–pallidal network for goal-directed
and habitual inhibition. Nature Reviews Neuroscience, 16,
719–732. DOI: https://doi.org/10.1038/nrn4038, PMID:
26530468
Jahfari, S., Stinear, C. M., Claffey, M., Verbruggen, F., & Aron, A. R.
(2009). Responding with restraint: What are the neurocognitive
mechanisms? Journal of Cognitive Neuroscience, 22,
1479–1492. DOI: https://doi.org/10.1162/jocn.2009.21307,
PMID: 19583473, PMCID: PMC2952035
Jurkiewicz, M. T., Gaetz, W. C., Bostan, A. C., & Cheyne, D.
(2006). Post-movement beta rebound is generated in motor
796
Journal of Cognitive Neuroscience
Volume 33, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
o
c
n
_
a
_
0
1
6
8
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
cortex: Evidence from neuromagnetic recordings. Neuroimage,
32, 1281–1289. DOI: https://doi.org/10.1016/j.neuroimage
.2006.06.005, PMID: 16863693
Kenemans, J. L. (2015). Specific proactive and generic reactive
inhibition. Neuroscience & Biobehavioral Reviews, 56, 115–126.
DOI: https://doi.org/10.1016/j.neubiorev.2015.06.011, PMID:
26116545
Kilavik, B. E., Zaepffel, M., Brovelli, A., MacKay, W. A., & Riehle, A.
(2013). The ups and downs of β oscillations in sensorimotor
cortex. Experimental Neurology, 245, 15–26. DOI: https://doi
.org/10.1016/j.expneurol.2012.09.014, PMID: 23022918
Klein, P.-A., Petitjean, C., Olivier, E., & Duque, J. (2014). Top–
down suppression of incompatible motor activations during
response selection under conflict. Neuroimage, 86, 138–149.
DOI: https://doi.org/10.1016/j.neuroimage.2013.08.005,
PMID: 23939021
Lee, T. W., Girolami, M., & Sejnowski, T. J. (1999). Independent
component analysis using an extended infomax algorithm
for mixed subgaussian and supergaussian sources. Neural
Computation, 11, 417–441. DOI: https://doi.org/10.1162
/089976699300016719, PMID: 9950738
Little, S., Bonaiuto, J., Barnes, G., & Bestmann, S. (2019).
Human motor cortical β bursts relate to movement planning
and response errors. PLoS Biology, 17, e3000479. DOI: https://
doi.org/10.1371/journal.pbio.3000479, PMID: 31584933,
PMCID: PMC6795457
Logan, G. D., Cowan, W. B., & Davis, K. A. (1984). On the ability
to inhibit simple and choice reaction time responses: A
model and a method. Journal of Experimental Psychology:
Human Perception and Performance, 10, 276–291. DOI:
https://doi.org/10.1037/0096-1523.10.2.276
Majid, D. S. A., Cai, W., Corey-Bloom, J., & Aron, A. R. (2013).
Proactive selective response suppression is implemented via
the basal ganglia. Journal of Neuroscience, 33, 13259–13269.
DOI: https://doi.org/10.1523/JNEUROSCI.5651-12.2013,
PMID: 23946385, PMCID: PMC3742918
Majid, D. S. A., Cai, W., George, J. S., Verbruggen, F., & Aron, A. R.
(2011). Transcranial magnetic stimulation reveals dissociable
mechanisms for global versus selective corticomotor
suppression underlying the stopping of action. Cerebral
Cortex, 22, 363–371. DOI: https://doi.org/10.1093/cercor
/bhr112, PMID: 21666129, PMCID: PMC3256406
Mansfield, E. L., Karayanidis, F., Jamadar, S., Heathcote, A., &
Forstmann, B. U. (2011). Adjustments of response threshold
during task switching: A model-based functional magnetic
resonance imaging study. Journal of Neuroscience, 31,
14688–14692. DOI: https://doi.org/10.1523/JNEUROSCI
.2390-11.2011, PMID: 21994385, PMCID: PMC6703389
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Journal of Neuroscience
Methods, 164, 177–190. DOI: https://doi.org/10.1016
/j.jneumeth.2007.03.024, PMID: 17517438
McFarland, D. J., Miner, L. A., Vaughan, T. M., & Wolpaw, J. R.
(2000). Mu and beta rhythm topographies during motor
imagery and actual movements. Brain Topography, 12,
177–186. DOI: https://doi.org/10.1023/A:1023437823106,
PMID: 10791681
Meyer, H. C., & Bucci, D. J. (2016). Neural and behavioral
mechanisms of proactive and reactive inhibition. Learning
& Memory, 23, 504–514. DOI: https://doi.org/10.1101/lm
.040501.115, PMID: 27634142, PMCID: PMC5026209
Muralidharan, V., Yu, X., Cohen, M. X., & Aron, A. R. (2019).
Preparing to stop action increases β band power in
contralateral sensorimotor cortex. Journal of Cognitive
Neuroscience, 31, 657–668. DOI: https://doi.org/10.1162
/jocn_a_01373, PMID: 30633601
Neuper, C., Wörtz, M., & Pfurtscheller, G. (2006). ERD/ERS
patterns reflecting sensorimotor activation and deactivation.
In C. Neuper & W. Klimesch (Eds.), Progress in brain
research (pp. 211–222). Elsevier. DOI: https://doi.org/10
.1016/S0079-6123(06)59014-4
Obeso, I., Cho, S. S., Antonelli, F., Houle, S., Jahanshahi, M., Ko,
J. H., et al. (2013). Stimulation of the pre-SMA influences
cerebral blood flow in frontal areas involved with inhibitory
control of action. Brain Stimulation, 6, 769–776. DOI:
https://doi.org/10.1016/j.brs.2013.02.002, PMID: 23545472
Obeso, I., Wilkinson, L., Casabona, E., Speekenbrink, M., Luisa
Bringas, M., Álvarez, M., et al. (2014). The subthalamic
nucleus and inhibitory control: Impact of subthalamotomy in
Parkinson’s disease. Brain, 137, 1470–1480. DOI: https://doi
.org/10.1093/brain/awu058, PMID: 24657985
Perrin, F., Pernier, J., Bertrand, O., & Echallier, J. F. (1989).
Spherical splines for scalp potential and current density mapping.
Electroencephalography and Clinical Neurophysiology, 72,
184–187. DOI: https://doi.org/10.1016/0013-4694(89)90180-6
Pfurtscheller, G., Graimann, B., Huggins, J. E., Levine, S. P.,
& Schuh, L. A. (2003). Spatiotemporal patterns of β
desynchronization and gamma synchronization in
corticographic data during self-paced movement. Clinical
Neurophysiology, 114, 1226–1236. DOI: https://doi.org
/10.1016/S1388-2457(03)00067-1
Pfurtscheller, G., Neuper, C., Brunner, C., & Da Silva, F. L.
(2005). Beta rebound after different types of motor imagery
in man. Neuroscience Letters, 378, 156–159. DOI: https://doi
.org/10.1016/j.neulet.2004.12.034, PMID: 15781150
Pfurtscheller, G., Neuper, C., Flotzinger, D., & Pregenzer, M.
(1997). EEG-based discrimination between imagination of
right and left hand movement. Electroencephalography and
Clinical Neurophysiology, 103, 642–651. DOI: https://doi
.org/10.1016/S0013-4694(97)00080-1
Pogosyan, A., Gaynor, L. D., Eusebio, A., & Brown, P. (2009).
Boosting cortical activity at β-band frequencies slows
movement in humans. Current Biology, 19, 1637–1641. DOI:
https://doi.org/10.1016/j.cub.2009.07.074, PMID: 19800236,
PMCID: PMC2791174
Raud, L., Huster, R., Ivry, R. B., Labruna, L., Messel, M. S., &
Greenhouse, I. (2020). A single mechanism for global and
selective response inhibition under the influence of motor
preparation. Journal of Neuroscience, 40, 7921–7935. DOI:
https://doi.org/10.1523/JNEUROSCI.0607-20.2020, PMID:
32928884, PMCID: PMC7548697
Ridderinkhof, R. K. (2002). Micro- and macro-adjustments
of task set: Activation and suppression in conflict tasks.
Psychological Research, 66, 312–323. DOI: https://doi.org
/10.1007/s00426-002-0104-7, PMID: 12466928
Ridderinkhof, K. R., de Vlugt, Y., Bramlage, A., Spaan, M., Elton,
M., Snel, J., et al. (2002). Alcohol consumption impairs
detection of performance errors in mediofrontal cortex.
Science, 298, 2209–2211. DOI: https://doi.org/10.1126
/science.1076929, PMID: 12424384
Ridderinkhof, K., Forstmann, B. U., Wylie, S. A., Burle, B., &
van den Wildenberg, W. P. M. (2011). Neurocognitive
mechanisms of action control: Resisting the call of the
Sirens. Wiley Interdisciplinary Reviews: Cognitive Science,
2, 174–192. DOI: https://doi.org/10.1002/wcs.99, PMID:
26302009
Robbins, T. W. (2007). Shifting and stopping: Fronto-striatal
substrates, neurochemical modulation and clinical implications.
Philosophical Transactions of the Royal Society of London,
Series B, Biological Sciences, 362, 917–932. DOI: https://doi
.org/10.1098/rstb.2007.2097, PMID: 17412678, PMCID:
PMC2430006
Schmidt, R., & Berke, J. D. (2017). A pause-then-cancel model
of stopping: Evidence from basal ganglia neurophysiology.
Philosophical Transactions of the Royal Society of London,
Series B, Biological Sciences 372, 20160202. DOI: https://
Soh et al.
797
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
o
c
n
_
a
_
0
1
6
8
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
doi.org/10.1098/rstb.2016.0202, PMID: 28242736, PMCID:
PMC5332861
Schoffelen, J.-M., Oostenveld, R., & Fries, P. (2008). Imaging
the human motor system’s β-band synchronization during
isometric contraction. Neuroimage, 41, 437–447. DOI: https://
doi.org/10.1016/j.neuroimage.2008.01.045, PMID: 18396062
Schulz, H., Übelacker, T., Keil, J., Müller, N., & Weisz, N. (2013).
Now I am ready—Now I am not: The influence of pre-TMS
oscillations and corticomuscular coherence on motor-evoked
potentials. Cerebral Cortex, 24, 1708–1719. DOI: https://doi
.org/10.1093/cercor/bht024, PMID: 23395847
Sherman, M. A., Lee, S., Law, R., Haegens, S., Thorn, C. A.,
Hämäläinen, M. S., et al. (2016). Neural mechanisms of
transient neocortical β rhythms: Converging evidence from
humans, computational modeling, monkeys, and mice.
Proceedings of the National Academy of Sciences, U.S.A.,
113, E4885–E4894. DOI: https://doi.org/10.1073/pnas
.1604135113, PMID: 27469163, PMCID: PMC4995995
Shin, H., Law, R., Tsutsui, S., Moore, C. I., & Jones, S. R. (2017). The
rate of transient β frequency events predicts behavior across
tasks and species. eLife, 6, e29086. DOI: https://doi.org/10.7554
/eLife.29086, PMID: 29106374, PMCID: PMC5683757
Swann, N. C., Cai, W., Conner, C. R., Pieters, T. A., Claffey, M. P.,
George, J. S., et al. (2012). Roles for the pre-supplementary
motor area and the right inferior frontal gyrus in stopping
action: Electrophysiological responses and functional and
structural connectivity. Neuroimage, 59, 2860–2870. DOI:
https://doi.org/10.1016/j.neuroimage.2011.09.049, PMID:
21979383, PMCID: PMC3322194
Tan, H., Wade, C., & Brown, P. (2016). Post-movement β
Activity in sensorimotor cortex indexes confidence in the
estimations from internal models. Journal of Neuroscience,
36, 1516–1528. DOI: https://doi.org/10.1523/JNEUROSCI
.3204-15.2016, PMID: 26843635, PMCID: PMC4737767
Tenke, C. E., & Kayser, J. (2005). Reference-free quantification
of EEG spectra: Combining current source density (CSD) and
frequency principal components analysis (fPCA). Clinical
Neurophysiology, 116, 2826–2846. DOI: https://doi.org
/10.1016/j.clinph.2005.08.007, PMID: 16257577
Tinkhauser, G., Pogosyan, A., Little, S., Beudel, M., Herz, D. M.,
Tan, H., et al. (2017). The modulatory effect of adaptive deep
brain stimulation on β bursts in Parkinson’s disease. Brain,
140, 1053–1067. DOI: https://doi.org/10.1093/brain/awx010,
PMID: 28334851, PMCID: PMC5382944
Tzagarakis, C., West, S., & Pellizzer, G. (2015). Brain oscillatory
activity during motor preparation: Effect of directional
uncertainty on β, but not alpha, frequency band. Frontiers in
Neuroscience, 9, 246. DOI: https://doi.org/10.3389/fnins
.2015.00246, PMID: 26257597, PMCID: PMC4508519
Ullsperger, M., & Danielmeier, C. (2016). Reducing speed and
sight: How adaptive is post-error slowing? Neuron, 89,
430–432. DOI: https://doi.org/10.1016/j.neuron.2016.01.035,
PMID: 26844827
Verbruggen, F., Aron, A. R., Band, G. P., Beste, C., Bissett, P. G.,
Brockett, A. T., et al. (2019). A consensus guide to capturing
the ability to inhibit actions and impulsive behaviors in the stop-
signal task. eLife, 8, e46323. DOI: https://doi.org/10.7554/eLife
.46323, PMID: 31033438, PMCID: PMC6533084
Verbruggen, F., & Logan, G. D. (2009). Proactive adjustments
of response strategies in the stop-signal paradigm. Journal
of Experimental Psychology: Human Perception and
Performance, 35, 835–854. DOI: https://doi.org/10.1037
/a0012726, PMID: 19485695, PMCID: PMC2690716
Vink, M., Kaldewaij, R., Zandbelt, B. B., Pas, P., & du Plessis, S.
(2015). The role of stop-signal probability and expectation in
proactive inhibition. European Journal of Neuroscience, 41,
1086–1094. DOI: https://doi.org/10.1111/ejn.12879, PMID:
25832122
Wessel, J. R. (2018). Surprise: A more realistic framework for
studying action stopping? Trends in Cognitive Sciences, 22,
741–744. DOI: https://doi.org/10.1016/j.tics.2018.06.005,
PMID: 30122169, PMCID: PMC7714639
Wessel, J. R. (2020). β-bursts reveal the trial-to-trial dynamics of
movement initiation and cancellation. Journal of Neuroscience,
40, 411–423. DOI: https://doi.org/10.1523/JNEUROSCI.1887
-19.2019, PMID: 31748375, PMCID: PMC6948942
Wessel, J. R., & Aron, A. R. (2017). On the globality of motor
suppression: Unexpected events and their influence on
behavior and cognition. Neuron, 93, 259–280. DOI: https://
doi.org/10.1016/j.neuron.2016.12.013, PMID: 28103476,
PMCID: PMC5260803
Wessel, J. R., Conner, C. R., Aron, A. R., & Tandon, N. (2013).
Chronometric electrical stimulation of right inferior frontal
cortex increases motor braking. Journal of Neuroscience, 33,
19611–19619. DOI: https://doi.org/10.1523/JNEUROSCI
.3468-13.2013, PMID: 24336725, PMCID: PMC3858630
Wessel, J. R., Waller, D. A., & Greenlee, J. D. W. (2019). Non-
selective inhibition of inappropriate motor-tendencies during
response-conflict by a fronto-subthalamic mechanism. eLife,
8, e42959. DOI: https://doi.org/10.7554/eLife.42959, PMID:
31063130, PMCID: PMC6533064
van den Wildenberg, W. P. M., van Boxtel, G. J. M., van der Molen,
M. W., Bosch, D. A., Speelman, J. D., & Brunia, C. H. M. (2006).
Stimulation of the subthalamic region facilitates the selection
and inhibition of motor responses in Parkinson’s disease.
Journal of Cognitive Neuroscience, 18, 626–636. DOI: https://
doi.org/10.1162/jocn.2006.18.4.626, PMID: 16768365
Zandbelt, B. B., & Vink, M. (2010). On the role of the striatum in
response inhibition. PLoS One, 5, e13848. DOI: https://
doi.org/10.1371/journal.pone.0013848, PMID: 21079814,
PMCID: PMC2973972
Zhang, Y., Chen, Y., Bressler, S. L., & Ding, M. (2008). Response
preparation and inhibition: The role of the cortical
sensorimotor β rhythm. Neuroscience, 156, 238–246. DOI:
https://doi.org/10.1016/j.neuroscience.2008.06.061, PMID:
18674598, PMCID: PMC2684699
798
Journal of Cognitive Neuroscience
Volume 33, Number 5
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
3
5
7
8
4
1
9
0
0
8
1
0
/
j
o
c
n
_
a
_
0
1
6
8
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3