Adaptive Top–Down Suppression of Hippocampal
Activity and the Purging of Intrusive Memories
from Consciousness
Roland G. Benoit1, Justin C. Hulbert2, Ean Huddleston3,
and Michael C. Anderson3
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
抽象的
■ When reminded of unwanted memories, 人们经常
attempt to suppress these experiences from awareness. 事先的
work indicates that control processes mediated by the dorso-
lateral prefrontal cortex (DLPFC) modulate hippocampal activity
during such retrieval suppression. It remains unknown whether
this modulation plays a role in purging an intrusive memory from
consciousness. 这里, we combined fMRI and effective connectiv-
ity analyses with phenomenological reports to scrutinize a role
for adaptive top–down suppression of hippocampal retrieval
processes in terminating mnemonic awareness of intrusive mem-
ories. Participants either suppressed or recalled memories of pic-
tures depicting faces or places. After each trial, they reported their
success at regulating awareness of the memory. DLPFC activation
was greatest when unwanted memories intruded into conscious-
ness and needed to be purged, and this increased engagement
predicted superior control of intrusive memories over time.
然而, hippocampal activity was decreased during the sup-
pression of place memories only. 重要的, the inhibitory
influence of the DLPFC on the hippocampus was linked to the
ensuing reduction in intrusions of the suppressed memories.
Individuals who exhibited negative top–down coupling during
early suppression attempts experienced fewer involuntary mem-
ory intrusions later on. Over repeated suppressions, the DLPFC–
hippocampus connectivity grew less negative with the degree
that they no longer had to purge unwanted memories from
意识. These findings support a role of DLPFC in counter-
manding the unfolding recollection of an unwanted memory
via the suppression of hippocampal processing, a mechanism
that may contribute to adaptation in the aftermath of traumatic
经历. ■
介绍
Not everything stored in memory is something that we
wish was there. The unpleasantness that sometimes
accompanies human experience is all too often remem-
bered, and when we are reminded of these unwelcome
事件, we often strive to exclude them from awareness.
Recent years have witnessed accumulating evidence that
targeted efforts to suppress an experience from aware-
ness can weaken the underlying memory representations
of the excluded trace and eventually cause forgetting
(Küpper, Benoit, Dalgleish, & 安德森, 2014; Noreen &
MacLeod, 2013; 安德森 & Huddleston, 2011; 安德森
& 绿色的, 2001). 在本文中, we examine the neural
mechanisms supporting the purging of an unwanted mem-
ory at moments when that memory involuntarily enters
意识. We seek, 尤其, to examine the mecha-
nisms that tie the regulation of awareness to reductions in
a memoryʼs later propensity to intrude into consciousness.
In so doing, we hope to provide a foundation for under-
standing how people adapt memory by attenuating in-
1哈佛大学, 剑桥, 嘛, 2普林斯顿大学, 新泽西州,
3MRC Cognition and Brain Sciences Unit, 剑桥, 英国
voluntary recollections that often pervade their experience
in the aftermath of unpleasant life events.
Much is already known about suppressing unwanted
回忆. 例如, attempts to suppress retrieval
are associated with increased activation in right dorsolat-
eral prefrontal cortex (DLPFC) and diminished activation
of the hippocampus (HC; Benoit & 安德森, 2012; 看
also Gagnepain, Henson, & 安德森, 2014; Paz-Alonso,
Bunge, 安德森, & Ghetti, 2013; Depue, 柯兰, & Banich,
2007; Anderson et al., 2004). Given the fundamental role
of the latter region in recollection (例如, Eichenbaum,
Yonelinas, & Ranganath, 2007; Eldridge, Knowlton,
Furmanski, Bookheimer, & 恩格尔, 2000), these data sug-
gest that the DLPFC exerts inhibitory control over critical
retrieval processes supported by the HC. Effective connectiv-
ity analyses provided evidence for this direct suppression
机制 (Benoit & 安德森, 2012): The DLPFC in-
fluenced hippocampal activation, and a more negative cou-
pling between these structures was associated with greater
forgetting of unwanted memories (see also Gagnepain
等人。, 2014). 重要的, this negative modulation of
HC activation did not arise for participants asked to avoid
the unwanted memory by generating a distracting thought;
it only arose for those told to directly suppress the retrieval
© 2014 麻省理工学院. Published under a
Creative Commons Attribution 3.0 Unported (CC-BY) 执照
认知神经科学杂志 27:1, PP. 96–111
土井:10.1162/jocn_a_00696
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
e
j
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
哦
2
C
0
n
3
_
7
A
/
_
j
0
哦
0
C
6
n
9
_
6
A
p
_
d
0
0
乙
6
y
9
G
6
你
.
e
p
s
d
t
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
t
F
/
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
过程 (for suppression vs. distraction, see also Bergström,
de Fockert, & Richardson-Klavehn, 2009; see Anderson &
Hanslmayr, 2014, for a review). Because negative modu-
lation of HC activity was associated with instructions to
terminate episodic retrieval, the inhibitory control mecha-
nisms supporting this process may underpin the subjective
experience of intentionally terminating awareness of an
unwelcome reminding.
The current study examines the linkage between the
engagement of top–down fronto-hippocampal modu-
latory processes and the regulation of awareness. Spe-
cifically, we test the possibility that this mechanism gets
adaptively upregulated when intruding memories need
to be purged from consciousness. Although existing data
on retrieval suppression are consistent with this possibil-
性, the key inference for this hypothesis is indirect. 在
Benoit and Anderson (2012), 例如, whether memo-
ries intruded during suppression attempts is unknown
and even if this was known, it remains possible that a
different mechanism purged the memory from aware-
内斯, in parallel to the modulation of HC activation by
DLPFC. HC modulation might instead serve a proactive
function of preventing recollection. To establish that DLPFC
is involved in purging momentary awareness (a form of
adaptive control), requires that we link the occurrence of
intrusions and their control to the DLPFC–HC suppression
机制.
Suggestive evidence already exists for an adaptive con-
trol process that reactively purges memories from aware-
ness via hippocampal downregulation. A recent study
observed reduced HC activation when people suppressed
unwanted memories but only when those memories in-
truded into awareness, as indexed by trial-by-trial intrusion
判断 (征收 & 安德森, 2012). 很遗憾, 这
control process that reduced HC activation during intru-
sions was not strongly established in that study, leaving it
uncertain whether the DLPFC mechanism was involved in
purging memories from awareness.
In this study, we seek to identify two key features of the
DLPFC–HC mechanism that would provide evidence for
adaptive control. 第一的, the top–down influence exerted
by the DLPFC should be dynamically upregulated during
intrusive memories, and such reactive engagement should
contribute to reduced involuntary awareness of the intrud-
ing memory over time. 第二, if this mechanism renders
the memories less intrusive, there would be less need for
inhibitory top–down control after repeated suppression
attempts for a given memory. 例如, in research
on retrieval-induced forgetting, PFC regions are recruited
most during the initial retrieval of a target memory when
competition from an interfering memory needs to be
overcome (Wimber et al., 2011; Kuhl, Dudukovic, Kahn,
& 瓦格纳, 2007); 批判地, activation in those regions is
attenuated during later retrieval attempts, when the com-
peting memory had presumably been weakened and thus
had become less interfering. On the basis of these find-
英格斯, Kuhl et al. (2007) argued that the diminished need
to engage cognitive control over repetitions was an adap-
tive benefit of forgetting competing memories. 如果是这样, 一个
analogous benefit of successful retrieval suppression may
be observed here, with diminished engagement of control
over repeated suppression attempts, as intrusions of un-
wanted memories are downregulated. 因此, the DLPFC–
HC suppression mechanism could be considered an adap-
tive control process if it gets dynamically upregulated to
contend with involuntary intrusions, and it becomes pro-
gressively disengaged over trials with diminishing need to
purge unwanted memories from awareness.
Participants first extensively encoded cue–picture asso-
ciations via a procedure designed to induce strong and
intrusive memories. Half of the pictures depicted faces;
the other half, 地方. Using two types of material allowed
us to examine whether the prefrontal inhibition mecha-
nism is engaged irrespective of the mnemonic content
(Benoit, 吉尔伯特, Frith, & 伯吉斯, 2012; Burgess et al.,
2006). After training, participants performed a think/no-
think task (安德森 & 绿色的, 2001), as they were scanned
with fMRI. They were presented with the cues from studied
pairs and repeatedly recalled some associated pictures (关于-
call condition) and attempted to suppress others (suppress
状况). Critically, after each trial, participants indi-
cated whether the picture had entered awareness (征收
& 安德森, 2012). This method of introspection, intro-
duced in attention research (Corallo, 萨克尔, 德阿内, &
Sigman, 2008; 军士 & 德阿内, 2004), enables us to
identify activation associated with intrusions (IE。, picture
involuntarily came to mind) versus nonintrusions (IE。, pic-
ture was kept out of awareness). It provides our central
behavioral dependent measure, as it may be used to quan-
tify the degree to which participants progressively gained
control over intrusive memories.
If DLPFC supports an adaptive control mechanism that is
instrumental to purging the contents of mnemonic aware-
内斯, we expected several core findings, both on individ-
ual trials as a function of intrusions and over the course
of the think/no-think phase as a function of a diminishing
intrusion frequency. 第一的, DLPFC should be more robustly
engaged when people need to countermand intruding
memories than on trials during which they do not experi-
ence intrusions. If this mechanism is instrumental in purg-
ing memories from awareness, stronger engagement of
DLPFC during intrusions may be associated with a greater
decline in the memoriesʼ intrusiveness over time. 第二,
effective connectivity analyses should reveal a modulatory
influence of DLPFC on HC during suppression. If a negative
top–down coupling is implemented to purge unwanted
memories from awareness, its engagement may be tied
to the degree that such memories intrude into conscious-
内斯. It should be especially pronounced when individuals
efficiently cope well with unwanted memories, as indi-
cated by a greater subsequent reduction in intrusions. 作为
a consequence of such efficient coping, there would sub-
sequently be less demand for the purging mechanism, 和
we accordingly expect a successive attenuation of negative
Benoit et al.
97
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
e
j
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
哦
2
C
0
n
3
_
7
A
/
_
j
0
哦
0
C
6
n
9
_
6
A
p
_
d
0
0
乙
6
y
9
G
6
你
.
e
p
s
d
t
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
/
t
.
F
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
top–down control over trials (比照. Wimber et al., 2011; Kuhl
等人。, 2007).
方法
参加者
全部 18 volunteers were right-handed and not color blind,
reported no history of neurological or psychiatric disorder,
and gave written informed consent as approved by the
local ethics research committee. Two participants were
excluded: one because of poor recall of faces and one
for noncompliance with the instructions as assessed by
a postexperimental questionnaire (derived from Hertel &
Calcaterra, 2005). 因此, 数据来自 16 participants re-
mained for further analyses (8 男人, mean age = 22 年,
range = 18–30 years).
程序
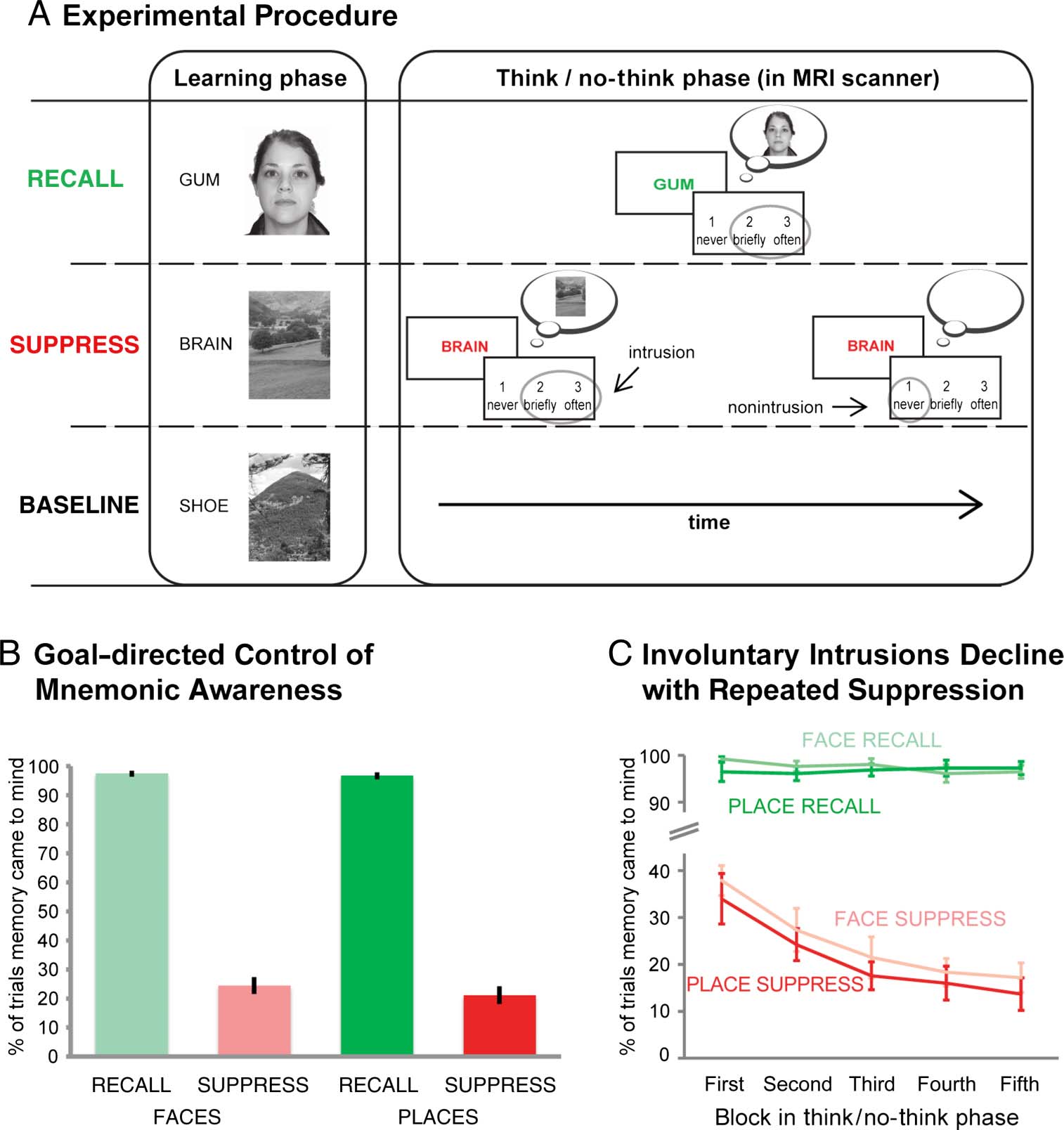
The procedure was based on the think/no-think task
(安德森 & 绿色的, 2001) and was divided into three
阶段 (Figure 1A): (A) an elaborate study phase, 期间
which participants encoded cue–memory associations;
(乙) the critical think/no-think phase, during which we
assessed the key behavioral measure, 那是, the experi-
ence of involuntary memory intrusions; 和 (C) a final rec-
ognition phase.
During study, participants encoded 24 critical cue–
picture pairings. The cues were short nouns, and all pic-
tures were presented in grayscale. Half of the images
depicted unfamiliar faces (IE。, white female and male
faces); the other half, 地方 (水, 树, mountain, 和
prairie scenes). The pictures were assigned to the sup-
press, 记起, and baseline conditions, counterbalanced
across participants. Participants also encoded 12 filler pair-
ings for practice. The study phase was composed of two
stages designed to create strong memories that would
likely intrude into awareness involuntarily. 第一的, each cue–
picture pairing was presented for 6 秒. 第二, we en-
sured successful learning by a category recall task (IE。,
presenting each cue word and asking participants to clas-
sify the retrieved scenes as a face or place) followed by a
four-choice recognition task. The foils were sampled from
数字 1. (A) Participants were
trained on reminder–picture
pairings, before they suppressed
memories cued with red
reminders and recalled those
cued by green reminders. 后
每次试验, they indicated whether
the memory did come to mind
(either briefly or often) 或者
did not enter awareness at
全部. These critical trial-by-trial
reports allowed us to identify
brain activation associated with
intrusions and to determine the
degree to which participants
gained better control over
involuntary retrievals with
repeated suppression attempts.
(乙) Whereas participants
reported awareness of the
reminded memory on most trials
of the recall condition, 他们
did experience intrusions of
to-be-suppressed memories
on a smaller number of trials.
(C) The intrusion frequency
of these items decreased over
blocks of the experiment.
98
认知神经科学杂志
体积 27, 数字 1
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
e
j
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
哦
2
C
0
n
3
_
7
A
/
_
j
0
哦
0
C
6
n
9
_
6
A
p
_
d
0
0
乙
6
y
9
G
6
你
.
e
p
s
d
t
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
F
.
/
t
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
the pairings and were of the same type as the critical picture
(例如, male faces). By this, participants had to remember
exact perceptual features. They received feedback after
each test and were shown the cues along with the correct
pictures for 2 sec at the end. A cue–picture pairing was re-
moved from the training set, only if it had been recognized
and its category had been recalled on the same trial. 我们
thus ensured accurate memory for all pictures and further
strengthened their representations by feedback.
After a practice session, participants performed the
think/no-think phase in the MRI scanner. We presented
cues in either red or green for 3 秒. Participants always
read and comprehended the cue first. For green cues,
they then covertly recalled the associated pictures (记起
状况). 相比之下, for red cues, they avoided thinking
of the associated picture at all (suppress condition),
in whatever way they saw fit (as in Anderson et al., 2004;
安德森 & 绿色的, 2001). Participants were thus free to
adopt their own strategy, as long as they did not shift their
eyes or attention away from the cue. After cue offset, 他们
reported the extent to which they had thought about its
associated picture by pressing one of three buttons: never,
briefly, 并且经常. This was followed by a jittered ISI (M ±
标准差= 3500 ± 1588 毫秒), optimized to increase the effi-
ciency of the event-related design using optseq2 (surfer.
nmr.mgh.harvard.edu/optseq/). There were five blocks,
during each of which all recall and suppress cues were
randomly presented twice, although any cue could only
be repeated once all the others had been shown. 因此, 在
全部的, each suppress and recall memory was cued 10 次.
We did not present cues for baseline memories during this
phase.
Outside the scanner, after-effects of suppression on
recognition memory were assessed. Given the extensive
training of the memories and based on pilot data, 我们
expected nearly perfect recognition accuracy. We there-
fore examined whether repeated purging from awareness
slowed subsequent recognition. A trial started with a cue
presented in white and the prompt to recall the category
of the associated picture within 5 秒. The response was
immediately followed by a blank screen (500 毫秒), 前
we presented the correct picture and a single foil. Partici-
pants had up to 5 sec to respond on this forced-choice
recognition task. Analyses focused on correct recognition
时间, conditioned on items for which participants had
also correctly recalled the category (比照. Detre, Natarajan,
格什曼, & Norman, 2013).
fMRI Acquisition and Preprocessing
A 3-T Siemens TIM Trio MRI scanner was used for acquisi-
tion of T2*-weighted EPIs (64 × 64; 3 × 3 mm pixels;
3-mm thick, oriented to the AC–PC plane; repetition
time = 2 秒; echo time = 30 毫秒; flip angle = 78°). Five
runs were acquired, with the number of volumes varying
从 193 到 226 because of the self-paced intrusion ratings.
此外, magnetization prepared rapid gradient echo
structural images were acquired (256 × 240 × 192; 1-mm3
isotropic voxels; repetition time = 2,250 毫秒; echo
time = 2.99 毫秒; flip angle = 9°).
Preprocessing and univariate analyses were performed
with SPM8 (www.fil.ion.ucl.ac.uk/spm/software/spm8).
The volumes were realigned, corrected for different slice
acquisition times, and coregistered with the structural
图像. This was spatially normalized, and the resulting
parameters served to normalize the functional images
进入 3 × 3 × 3 mm3 voxels by fourth degree B-spine
interpolation (using the Montreal Neurological Institute
reference brain). The images were then smoothed by
an isotropic 8-mm FWHM Gaussian kernel.
fMRI Analyses
The variance in BOLD signal was decomposed in a general
linear model (Friston et al., 1995) on the concatenated
data from all runs to optimize estimation of intrusion
事件. Six regressors coded the 3 sec of each trial, sepa-
rately for recall, nonintrusion (IE。, suppress trials with a
“never” rating), and intrusion (IE。, other suppress trials)
events of the faces and places trials. These regressors were
convolved with the canonical hemodynamic response func-
的. The full model was also composed of regressors for
each run representing residual movement artifacts. We mod-
eled trends over time by including, for each run, a linear-
trend predictor, a six-predictor Fourier basis for nonlinear
趋势 (sines and cosines of up to three cycles per run),
and a confound-mean predictor (Kriegeskorte, 穆尔, &
Bandettini, 2008). Parameters were estimated from the least
mean-squares fit of the model to the data. For second-level
分析, contrast estimates for recall, nonintrusion, 并在-
trusion events (for both faces and places) were entered into
a repeated-measures ANOVA using nonsphericity correction,
with Individuals as between-subject factor.
We modeled effective connectivity with DCM12 as im-
plemented in SPM12b, following the procedure employed
by Benoit and Anderson (2012). DCM explains regional ef-
fects in terms of changing patterns of connectivity during
experimentally induced contextual modulations (Stephan
等人。, 2010). It allows for inferences about the presence
and directions of causal connections, such as whether
activity in the HC is influenced by activity in the DLPFC
during suppression attempts (Benoit & 安德森, 2012).
These inferences are based on evidence from Bayesian
model selection (BMS; 见下文), which indicates the
probability that a given model is more likely to have gen-
erated the data than the other models (例如, a model that
entails a modulation of the connectivity from DLPFC to HC
与. a model with a modulation of the reversed connectiv-
ity from HC to DLPFC; Penny et al., 2010). DCM requires
univariate effects of interest in the respective brain areas
(Stephan et al., 2010). Given that we did not observe
hippocampal modulation during the suppression of face
pictures (see Results), our analyses of effective connectivity
are thus restricted to place data.
Benoit et al.
99
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
e
j
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
哦
2
C
0
n
3
_
7
A
/
_
j
0
哦
0
C
6
n
9
_
6
A
p
_
d
0
0
乙
6
y
9
G
6
你
.
e
p
s
d
t
哦
F
n
乙
0
y
7
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
.
/
t
F
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
All models were variations of a standard model that is
composed of the two regions (DLPFC and HC) as nodes,
within-region inhibitory auto-connections, and bidirec-
tional intrinsic connections (representing the connectiv-
ity between the regions across all conditions). Any cue
to recall or to suppress could elicit responses in this net-
工作. This driving input was modeled as a series of delta
functions at the respective cue onsets. It could enter the
network either via the HC, the DLPFC, or both nodes,
thus constituting three basic models (Figure 3A). Criti-
卡莉, we wanted to assess whether models could account
better for the data that feature a modulation of the top–
down connectivity from DLPFC to HC during attempts
to suppress an unwanted memory. We thus created four
model families, each of which was composed of modified
versions of all three basic models. Across the model fam-
ilies, we varied the connection that could be modulated
during suppression (Figure 3A). The modulatory compo-
nent reflects the change in coupling from the average
connectivity that arises specifically during suppression
attempts. Family 1 did not have any such modulatory
成分, whereas Family 2 allowed for modulation of
the connection from HC to DLPFC. 因此, these two fami-
lies do not feature a modulation of the top–down con-
nection, and they are incongruent with the hypothesized
suppression mechanism. 相比之下, Family 3 entailed
modulation of the connection from DLPFC to HC, 和
Family 4 allowed both the DLPFC-to-HC and HC-to-DLPFC
connections to be modulated during suppress events. 因此,
these latter two families are congruent with the hypothe-
sized suppression mechanism. The modulatory input was
defined as changes in connectivity induced during the first
second after the onset of a suppress event.
The models were estimated separately for each partici-
pant and for each of the five functional runs. By this, 我们
were able to assess changes in top–down coupling across
the think/no-think phase. We first identified the group
peaks for the suppress-versus-recall contrast within a
sphere (r = 10 毫米) centered on the DLPFC peak previ-
ously associated with direct memory suppression (X, y,
z = 36, 38, 31; Benoit & 安德森, 2012; suppress > recall)
and within an anatomical mask of the right HC (Malian,
Laurienti, Kraft, & Burdette, 2003; suppress < recall). Both
peaks survived small-volume family-wise error correction
(DLPFC: 39, 29, 34, zmax = 3.99; HC: 33, −25, −11,
zmax = 4). We then identified the subject-specific peaks
within a sphere (r = 10 mm) centered on the observed
group peaks (Stephan et al., 2010). For the HC, the indi-
vidual peaks also had to be within the anatomical mask.
The individual peaks then served as centers for subject-
specific, spherical ROIs (r = 5 mm). The first eigenvariate
from an ROI (i.e., the first principal component of the
time series of the voxels), adjusted for the effects of inter-
est, constituted the regional activation. Model fitting was
based on these data and was achieved by adjusting the
parameters to maximize the free-energy estimate of the
model evidence (Friston, Harrison, & Penny, 2003).
On the estimated models, we ran BMS in a random-
effects approach, which does not assume that the optimal
model is identical across all participants and which is also
less susceptible to outliers than a fixed-effects approach
(Stephan et al., 2010). This procedure reports the exceed-
ance probability (EP), that is, the probability to which a
given model is more likely than the others to have gener-
ated the data from a randomly selected participant (Penny
et al., 2010). The EPs of all included models sum to 1.
Note that BMS penalizes for model complexity. We then
analyzed the effective connectivity (i.e., the sum of the
intrinsic and modulatory parameters) of the winning
model family, to examine the relationship between top–
down coupling during suppression and control of involun-
tary intrusions. Specifically, we performed Bayesian model
averaging (BMA; Penny et al., 2010), which computes
weighted averages of each model parameter, where the
weighting is determined by the posterior probability of
each model. To examine changes in connectivity across
the think/no-think phase, we conducted BMA separately
for each of the five blocks. We also performed BMA on
models from the first and second blocks (approximating
the first half of the think/no-think phase) to assess the
relationship between negative coupling during this period
and subsequent reductions in involuntary intrusions.
RESULTS
Behavioral Results
Repeated Suppression Attempts Decrease the Intrusion
of Unwanted Memories
We first analyzed the frequency with which people reported
awareness of the associated memory on both recall and
suppress trials. As might be expected based on the exten-
sive training of pairs, on recall trials, participants reported
they were nearly always successful at bringing the asso-
ciated picture into awareness during the think/no-think
phase (M ± SD: faces = 98 ± 4%; places = 97 ± 5%). More-
over, the frequency of reported retrieval did not change
reliably across the session (i.e., from the first block to the
last block) as indicated by an ANOVA with the factors
Block and Material (Block: F(1, 15) = 1.44, p = .25).
On suppress trials, participants reported that pictures
came into awareness far less frequently (faces: 24 ± 12%;
places: 21 ± 12%) than during recall trials (Figure 1B).
This dramatic difference in reported mnemonic awareness
was corroborated by an ANOVA with the factors Retrieval
goal (recall, suppress) and Material (faces, places) that
yielded a significant effect of Retrieval Goal only (F(1,
15) = 699.31, p < .001). Thus, participants exhibited a
highly robust ability to control awareness of the pictures
(a nearly 75% difference in awareness across conditions).
Nevertheless, the images intruded frequently during sup-
pression trials, although the intrusion frequency declined
from the first block to the last block (cf. Levy & Anderson,
2012): An ANOVA with the factors Block and Material
100
Journal of Cognitive Neuroscience
Volume 27, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
o
2
c
0
n
3
_
7
a
/
_
j
0
o
0
c
6
n
9
_
6
a
p
_
d
0
0
b
6
y
9
g
6
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
yielded a substantial effect of Block (F(1, 15) = 60.3, p <
.001), reflecting diminished intrusion frequencies for both
faces and places with repeated suppression attempts (Fig-
ure 1B). Neither the Material effect nor the interaction was
significant (all Fs < 1.09, all ps > .3). 因此, our partici-
pants showed evidence of substantial improvements in
coping with intrusive memories, irrespective of mnemonic
内容. This pattern may reflect a progressive weakening
of the memories because of repeated suppression. 我们的
central objective is to understand the mechanisms under-
lying this highly robust reduction in intrusive memories
over time and whether an adaptive upregulation of the
modulatory DLPFC influence on HC contributes to it.
Suppressing Unwanted Memories Slows
Later Recognition
We expected nearly perfect performance on our final test
because of our extensive training procedure. 的确,
peopleʼs performance at classifying the category of the
picture in response to the cue word (例如, as either a face
or a place) was very high (M ± SEM: 0.92 ± 0.02 为了
地方, 0.94 ± 0.02 for faces), and an ANOVA with the
factors Retrieval goal (suppress, 记起, 基线) and Mate-
rials (faces, 地方) yielded no reliable differences or inter-
行动 (all Fs < 1.3, all ps > .28). 相似地, as expected
based on the extensive training and piloting results, 识别-
nition performance was virtually perfect across all condi-
系统蒸发散 (桌子 1; all Fs = 1.06, all ps = .319).
On measures of recognition time, 然而, an ANOVA
with the factors Retrieval goal and Materials revealed
a significant effect of Retrieval goal on response times
(F(2, 30) = 7.42, p < .003; Table 1), and a follow-up
ANOVA indicated that responses for suppress items were
slower than those for baseline items (F(1, 15) = 8.78,
p < .02). Thus, people were slower to recognize sup-
pressed pictures. Surprisingly, however, we also found
that recall items were recognized more slowly than base-
Table 1. Recognition Performance
Recognition
Recognition
Times (msec)
Mean
SEM
Mean
SEM
Faces
Baseline
Recall
Suppress
Places
Baseline
Recall
Suppress
0.97
0.95
0.95
0.96
0.95
0.93
0.01
0.02
0.03
0.03
0.02
0.03
1219
1384
1341
1452
1652
1610
90
93
103
81
110
111
line items (F(1, 15) = 16.31, p < .002), which could
not have been produced by a suppression process. The
main effects of Material were also significant, reflecting
faster responses for faces (all Fs > 8.96, all ps < .01).
Slowed RTs for suppress and recall items (compared with
baseline items) did not correlate (faces: r(14) = .098,
p = .719; places: r(14) = −.003, p = .99), suggesting
that the slowed recognition for these two item types
may reflect different mechanisms (see Discussion). How-
ever, exploratory analyses did not yield any significant
relationships between suppression-induced slowing and
any of the brain measures linked to the efficient coping
with unwanted memory intrusions reported in the next
sections.
fMRI Results
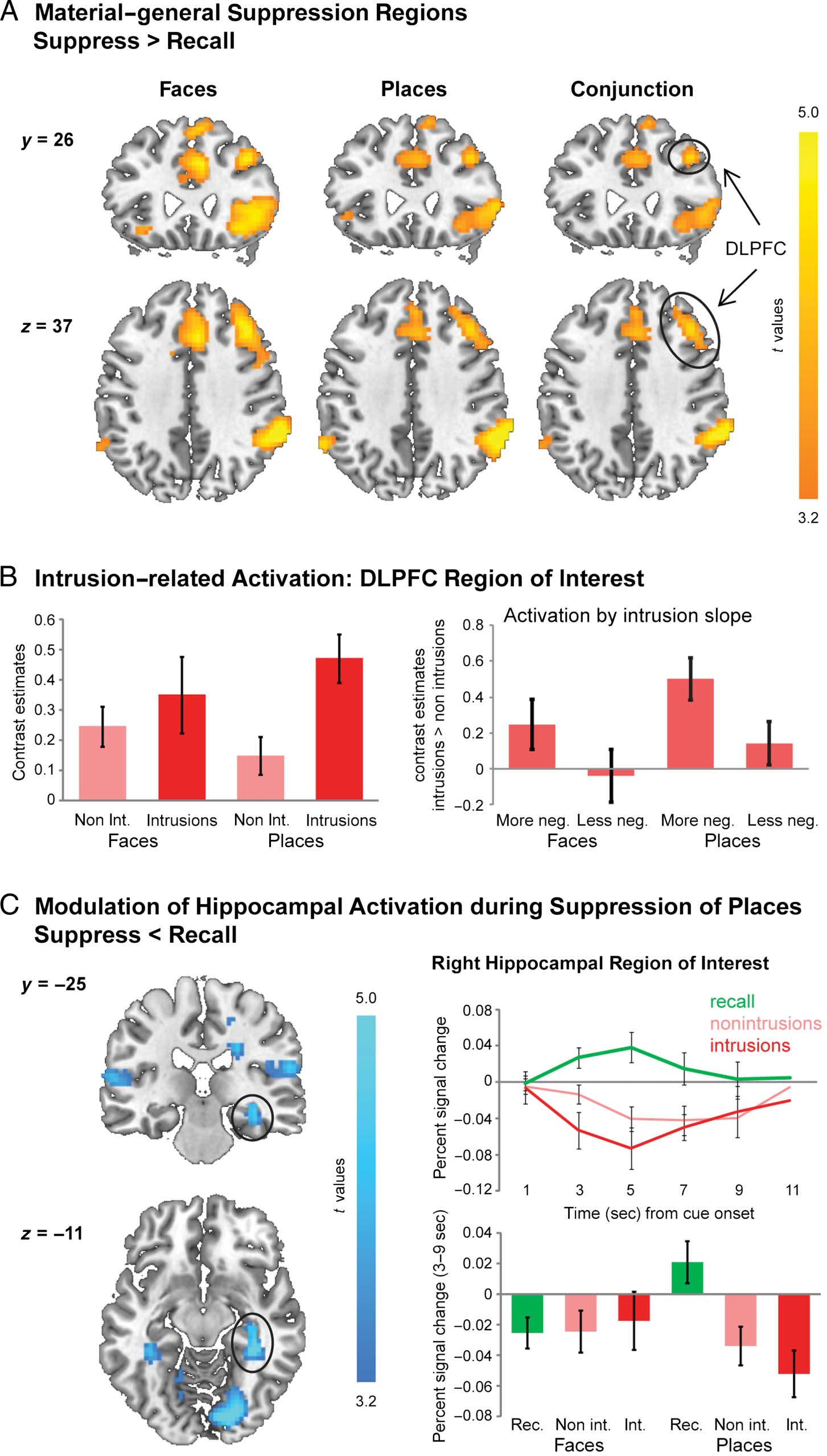
DLPFC Activation Is Greater during Memory Intrusions
To establish the involvement of a content-general suppres-
sion mechanism in DLPFC, we performed a conjunction
analysis of the suppress-versus-recall contrasts. This analy-
sis averaged estimates of intrusion and nonintrusion trials
and thus reveals those regions that are engaged across
both types of suppression trials. We then performed a
conjunction analysis of the contrasts for the faces and
places items to identify regions showing this effect for
both material types. Critically, an extensive right DLPFC
area centered on the middle frontal gyrus survived whole-
brain cluster correction (peak: x, y, z = 42, 26, 37; zmax =
4.23; 160 voxels; Figure 2A). This area included those
voxels previously implicated in direct memory suppression
(i.e., around x, y, z = 36, 38, 31; Benoit & Anderson, 2012;
for complete whole-brain analyses, see Tables 2–5). Thus,
a common DLPFC region is robustly engaged during the
suppression of images of faces and places and words
(as previously shown).
Having identified this control region, we next examined
whether it functioned to proactively gate retrieval or,
instead, could be adaptively upregulated to purge intrud-
ing mental content from awareness. The previous analysis
collapsed across contrast estimates for all suppression
trials, that is, irrespective of the occurrence of intrusions.
The obtained DLPFC cluster is thus derived from a contrast
vector that is orthogonal to the comparison of intrusion
versus nonintrusion trials. Hence, we extracted contrast
estimates for intrusions and nonintrusions from this cluster
and conducted an ANOVA with the factors Awareness
(intrusion, nonintrusion) and Material (faces, places). This
analysis revealed greater DLPFC activation when partici-
pants experienced intrusions than when they did not (F(1,
15) = 7.68, p < .02; Figure 2B). Thus, although DLPFC
was generally engaged when participants suppressed
memories, it was more strongly recruited in situations
that required the countermanding of intruding memories.
However, there was a trend for the interaction between
this effect and material type (F(1, 15) = 3.53, p = .08).
Benoit et al.
101
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
o
2
c
0
n
3
_
7
a
/
_
j
0
o
0
c
6
n
9
_
6
a
p
_
d
0
0
b
6
y
9
g
6
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. (A) Collapsing across
estimates for intrusion and
nonintrusion trials, DLPFC
showed stronger activation
during attempts to suppress
(vs. recall) memories of both
faces and places (conjunction
analysis). (B) Consistent
with a reactive account
of memory suppression,
activation in the DLPFC
cluster was greater during
intrusions than nonintrusions
(left), and the activation
difference between intrusions
and nonintrusions was greater
for individuals who were
more successful at decreasing
intrusion frequency over
time (i.e., with a more
negative intrusion slope; right).
(C) The right HC yielded the
expected reduced activation
during suppression of place
memories only. For display
purposes, statistical maps
are thresholded at p < .001,
uncorrected.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
o
2
c
0
n
3
_
7
a
/
_
j
0
o
0
c
6
n
9
_
6
a
p
_
d
0
0
b
6
y
9
g
6
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
102
Journal of Cognitive Neuroscience
Volume 27, Number 1
Table 2. Regions in Which BOLD Signal Was Significantly Greater during Suppression than Recall of Place Memories
MNI (Peak)
Region
SFG/MiFG
MiFG/SFG
ACC
MiFG/SFG
IFG
STG
SFG/MeFG
IFG
PreG/PosG
IPC
SMG/IPC
∼BA
10/11
10
32
9
45/47
13
6/8
47
4/3
40
40
Hemi.
l
r
r/l
r
r
r
r
l
l
r
l
x
−24
27
3
3
42
30
51
57
33
42
12
18
6
−45
−33
−51
66
−60
y
59
56
35
23
26
38
14
26
23
17
23
11
11
23
23
−13
−43
−52
z
−8
22
31
37
37
37
40
4
−14
−11
61
70
58
−5
−14
46
40
37
Voxels
Zmax
14
75
172
Same cluster
195
Same cluster
Same cluster
273
Same cluster
Same cluster
120
Same cluster
Same cluster
48
Same cluster
31
268
34
3.69
4.09
4.21
3.91
4.23
3.65
3.64
4.22
4.05
4.01
4.19
3.45
3.45
3.76
3.52
3.98
5.31
4.24
Threshold at p < .001, uncorrected, and at least 10 contiguous voxels. ACC = anterior cingulate cortex; IFG = inferior frontal gyrus; IPC = inferior
parietal cortex; MeFG = medial frontal gyrus; MiFG = middle frontal gyrus; PosG = postcentral gyrus; PreG = precentral gyrus; SFG = superior
frontal gyrus; SMG = supramarginal gyrus; STG = superior temporal gyrus; Hemi = hemisphere; r = right; l = left.
Stronger DLPFC Upregulation Predicts Better Coping
with Intrusive Memories
Purging a memory from awareness may be achieved by
inhibitory processes that reduce its tendency to intrude
again on later trials. If so, participants who recruit the
DLPFC region more during intrusions than during non-
intrusions may exhibit a steeper decline in intrusion fre-
quency over repeated suppression attempts. To test this
prediction, we performed a median split of our sample
based on the baseline-corrected intrusion slope (i.e., the
decrease in intrusion frequency over repetitions of sup-
pression, divided by the initial frequency of intrusions,
as reported by Levy & Anderson, 2012). By correcting
for individual differences in the initial intrusion frequency,
this measure allows for the definition of groups who were
high or low in their ability to cope with intrusive memories
over time. Given the absence of interactions between
material type and the decline in intrusion frequency (see
above), this split was performed on intrusion slopes
averaged across places and faces data. The individuals
exhibiting a more negative slope trended to experience
intrusions on fewer trials overall (M ± SEM = 18 ± 3%
vs. 28 ± 4%; t(14) = –1.97, p = .069), consistent with a
superior control ability.
We then performed an ANOVA on the measure of DLPFC
upregulation (i.e., the contrast estimates for intrusions >
nonintrusions) with the between-subject factor Intrusion
slope (more negative, less negative) and the within-subject
factor Material (地方, faces). Consistent with the predic-
的, participants who more effectively coped with intru-
西翁, reducing their frequency over time (IE。, 一个更多
negative slope), showed a significantly greater intrusion-
related upregulation of DLPFC activation (F(1, 15) = 5.83,
p < .05; Figure 2C). This effect did not interact with Material
type (F(1, 15) = 0.1, p = .76). Thus, a greater increase in
DLPFC activation when memories intruded into awareness
was associated with a greater decline in intrusion frequency,
irrespective of the nature of the unwanted memories.
Hippocampal Activation Is Reduced during Attempts
to Suppress Unwanted Place Memories
The hypothesized purging mechanism is thought to sup-
press awareness of intruding memories via the modulation
Benoit et al.
103
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
o
2
c
0
n
3
_
7
a
/
_
j
0
o
0
c
6
n
9
_
6
a
p
_
d
0
0
b
6
y
9
g
6
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
of HC activation by DLPFC. Because DLPFC has been
found to modulate right HC during direct suppression
(Benoit & Anderson, 2012), we extracted contrast
estimates for recall, nonintrusion, and intrusion trials from
an anatomical mask of this region (Malian et al., 2003). In
the current data, hippocampal activation seemed to be
reduced during attempts to suppress place memories only
(Figure 2C). This impression was corroborated by an
ANOVA with the factors Retrieval status (recall, nonintru-
sions, intrusions) and Material (faces, places) that yielded
a significant interaction (F(2, 30) = 4.15, p < .03). For
places, activation was lower during attempts to suppress
(compared with recall) unwanted memories both in the
absence (t(15) = 4.24, p < .001, one-tailed) and presence
Table 3. Regions in Which BOLD Signal Was Significantly Greater during Recall than Suppression of Place Memories
MNI (Peak)
Region
ACC
ACC
∼BA
24
23/32
Hemi.
l
r/l
STG/PosG
22/42/40
PHC
Striatum
Striatum
PosG
Insula
STG
CG
Insula
PHC
HC
PHC
Striatum
HC
CG
Cerebellum
MTG/SOG
MTG/SOG
Cuneus
PCC
40
13
41
31
13
37
36/37
31
39/19
39/19
17/18
31
l
r
l
r
r
r
r
l
l
r
l
r
l
r
r
x
−18
6
−6
−48
−63
−57
39
30
−21
−21
24
60
48
57
6
−27
−36
−27
36
30
36
30
−12
−24
42
−39
−33
18
15
12
y
32
23
20
−10
−25
−22
−13
−13
−19
−7
−25
−28
−7
−19
−34
−34
−40
−37
−34
−34
−43
−31
−37
−37
−76
−82
−76
−91
−76
−67
z
1
−2
−2
7
13
7
−26
−17
31
28
31
19
13
10
34
16
−8
4
−14
10
4
−2
46
−47
22
22
31
4
16
13
Voxels
Zmax
11
29
Same cluster
184
Same cluster
Same cluster
18
Same cluster
10
Same cluster
41
312
Same cluster
Same cluster
10
119
Same cluster
Same cluster
237
Same cluster
Same cluster
Same cluster
43
25
45
67
Same cluster
3074
Same cluster
Same cluster
3.63
3.73
3.49
3.94
3.92
3.63
3.6
3.25
3.34
3.32
4.01
4.65
4.1
4.03
3.67
4.3
4.24
3.26
5.19
4.51
4.03
3.48
3.82
3.63
4.03
4.2
3.62
6.84
5.43
5.29
Threshold at p < .001, uncorrected, and at least 10 contiguous voxels. CG = cingulate gyrus; PCC = posterior cingulate cortex; PHC = parahippocampal
cortex; SOG = superior occipital gyrus. See Table 2 for additional acronyms.
104
Journal of Cognitive Neuroscience
Volume 27, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
o
2
c
0
n
3
_
7
a
/
_
j
0
o
0
c
6
n
9
_
6
a
p
_
d
0
0
b
6
y
9
g
6
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 4. Regions in Which BOLD Signal Was Significantly Greater during Suppression than Recall of Face Memories
MNI (Peak)
∼BA
Hemi.
Region
MiFG/SFG
ACC
IFG/insula
10
6
32
47/13
IFG/insula
45/47/13
Striatum
ITG
PreG/PosG
IPC/SMG
Cerebellum
SMG
IOG/MOG/ITG
MTG/ITG
IOG
MOG
20
4/3
40
40
19/18
37
18/19
19
r
r/l
l
r
r
r
L
r
l
l
l
l
l
l
x
33
15
6
−30
−36
48
33
45
15
48
−48
60
60
51
−33
−66
−45
−45
−36
−36
y
53
14
32
20
14
17
20
23
8
−1
−16
−46
−43
−49
−49
−52
−79
−67
−85
−94
z
22
61
31
−11
1
4
−11
−5
7
−35
46
25
34
34
−32
31
−5
1
−8
13
Voxels
1345
Same cluster
Same cluster
101
Same cluster
533
Same cluster
Same cluster
22
13
15
391
Same cluster
Same cluster
37
18
55
Same cluster
Same cluster
38
Zmax
5.36
5.08
4.81
4.48
3.47
5.34
4.99
4.49
3.54
3.6
3.67
5.21
5.05
4.68
3.84
3.74
3.59
3.43
3.33
3.89
Threshold at p < .001, uncorrected, and at least 10 contiguous voxels. IOG = inferior occipital gyrus; ITG = inferior temporal gyrus; MOG = middle
occipital gyrus. See previous tables for additional acronyms.
of intrusions (t(15) = 2.04, p < .03, one-tailed), although
inspection of the time course data (as in Levy & Anderson,
2012) indicates that intrusion trials were associated with
numerically, yet not significantly, lower activation. In
contrast, for faces, activation was reduced neither during
nonintrusions (t(15) = 0.91, p = .19, one-tailed) nor intru-
sions (t(15) = 0.03, p = .49, one-tailed). Thus, whereas
the right DLPFC was recruited during attempts to sup-
press both faces and places, we observed evidence for
the complementary reduction in HC activation only during
the suppression of places.
The Top–Down Connectivity from DLPFC to HC Is
Modulated during Memory Suppression
During the suppression of place memories, we observed
the activation pattern previously associated with direct
Table 5. Regions in Which BOLD Signal Was Significantly Greater during Recall than Suppression of Face Memories
Region
ACC
SPC
Cuneus/MOG/LG
∼BA
25/24/32
7
17/18
Hemi.
l/r
l
r
MNI (Peak)
y
20
17
−76
−94
x
−6
6
−39
18
z
−2
−5
46
4
Voxels
18
24
132
Zmax
3.73
3.24
4.05
4.6
Threshold at p < .001, uncorrected, and at least 10 contiguous voxels. LG = lingual gyrus; SPC = superior parietal cortex. See previous tables for additional
acronyms.
Benoit et al.
105
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
o
2
c
0
n
3
_
7
a
/
_
j
0
o
0
c
6
n
9
_
6
a
p
_
d
0
0
b
6
y
9
g
6
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
memory suppression: increased DLPFC and reduced HC
activation. We hypothesized that the DLPFC would cause
this reduction to effectively purge unwanted memories
from awareness and that such suppression would be asso-
ciated with evidence of peopleʼs ability to efficiently cope
with memory intrusions over time. Efficient coping would
be reflected in two ways. First, we predicted a relationship
between the effective top–down connectivity and changes
in the frequency of involuntary intrusions. Those individ-
uals who exhibit an inhibitory (i.e., negative) influence
during early suppression trials should be more likely to
disrupt intruding traces and, as a result, experience fewer
involuntary intrusions on later suppression trials. Second,
to the extent that memories are rendered less intrusive
over time by early and efficient control, the need to engage
top–down control may be reduced. Hence, we used DCM
to examine the effective connectivity between the DLPFC
and the HC during the suppression of place memories
and how this changed over blocks. It was not possible to
model DLPFC–HC interactions during face suppression
because DCM requires all univariate effects of interest in
the respective ROIs (Stephan et al., 2010).
DCM explains regional effects in terms of dynamically
changing patterns of connectivity during experimentally
induced contextual changes. Thereby, it allows inferences
about the direction of causal connections, that is, whether
suppress events are more likely to modulate the top–down
connection from DLPFC to HC or the reverse, bottom–up
connection. We first tested whether models could account
best for the fMRI data that include a modulation of the
top–down connectivity from DLPFC to HC during mem-
ory suppression, similar to what has been observed in
prior work (Gagnepain et al., 2014; Benoit & Anderson,
2012). We then examined the consequences of this puta-
tive top–down influence. Specifically, we examined whether
it could be linked to the reduction in involuntary intrusions
with repeated suppression attempts.
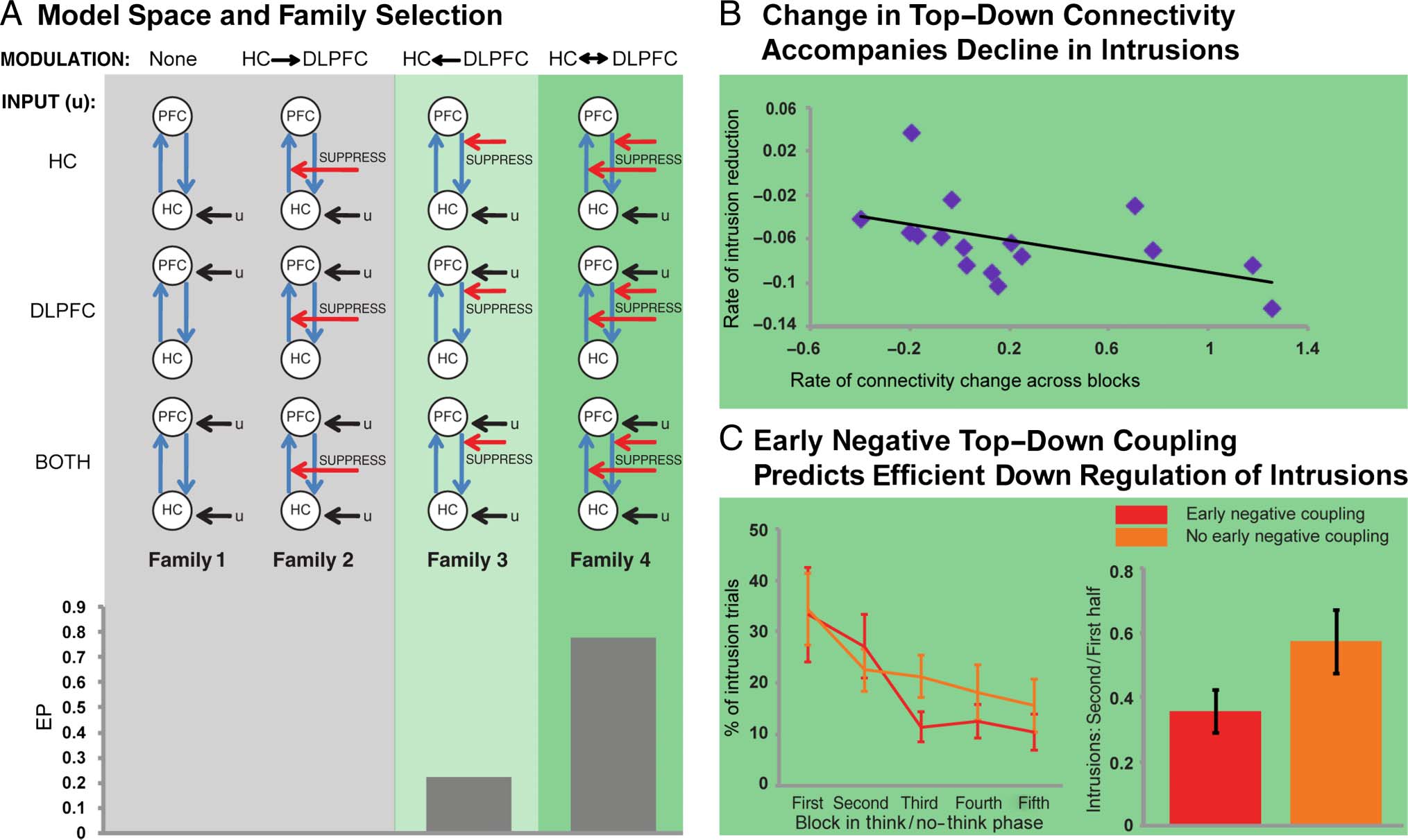
As detailed in Methods, we created four model families
that varied the connection that could be modulated during
suppression (Figure 3A). Family 1 did not have any modu-
latory component, whereas Family 2 entailed a modulation
of the connectivity from HC to DLPFC. Thus, these two
families are incongruent with the hypothesized suppres-
sion mechanism because they do not feature a modulation
of the top–down connection. By contrast, the remain-
ing two families are congruent with the hypothesized
increased top–down influence: Family 3 entailed a modu-
lation of the connection from DLPFC to HC, and Family 4
allowed both the DLPFC-to-HC and HC-to-DLPFC con-
nections to be modulated.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
o
2
c
0
n
3
_
7
a
/
_
j
0
o
0
c
6
n
9
_
6
a
p
_
d
0
0
b
6
y
9
g
6
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 3. (A) BMS indicated that Family 4 could account best for the places data, consistent with the hypothesized increased impact of DLPFC on
activation changes in the HC during suppression. (B) Changes in effective connectivity across the think/no-think phase were stronger for those individuals
who exhibited a steeper decline in the number of involuntary intrusions, suggesting that a reduced requirement to purge unwanted memories leads
to a disengagement of top–down inhibitory control. (C) Individuals exhibiting negative DLPFC–HC coupling early on (i.e., across the first two blocks)
subsequently experienced a greater reduction of involuntary intrusions.
106
Journal of Cognitive Neuroscience
Volume 27, Number 1
On the estimated models, we ran BMS, which returns
EPs, that is, the probability for each model family to be
more likely than the other families to have generated the
data (Penny et al., 2010). The selection procedure followed
the approach adopted by Benoit and Anderson (2012).
First, we compared the four families, and BMS indicated
that Family 4 was superior with an EP of 0.78 (Figure 3A).
Thus, replicating Benoit and Anderson (2012), those
models accounted best for the data that featured a modula-
tion of the critical connection from DLPFC to HC during
suppression as well as of the reverse, bottom–up connec-
tion. We then compared the three models within the win-
ning Family 4, that is, the models that varied in the location
of the driving input. Within this set, there was a clear pref-
erence for the model that received its driving input both
via the HC and the DLPFC (EP > 0.99). 总结一下,
consistent with our hypothesis, the winning model
featured a modulation of the connectivity from DLPFC
to HC, suggesting that DLPFC influences HC activation
specifically during memory suppression.
In the following, we further tested the adaptive control
account by examining the nature of the effective top–
down connectivity. The effective connectivity is the sum
of the intrinsic (IE。, average connectivity) and modulatory
connectivity parameters (IE。, change in connectivity spe-
cifically during suppress trials) and thus indicates the
implementation of top–down control that is specific to
memory suppression. BMA of the members of family IV
indicated that, across the entire course of the think/no-
think phase, the effective connectivity from the DLPFC to
the HC was not reliably different from 0 (米= 0.9, SEM =
1.15; t(15) = 0.78, p = .45). 然而, as described next,
this overall result masks a critical dynamic adjustment in
top–down suppression over blocks predicted by the adap-
tive control hypothesis.
Top–Down Suppression Changes with Diminishing
Need to Purge Involuntary Intrusions
People who cope efficiently with intrusions should show a
rapid decline in intrusions across the think/no-think phase,
reflecting the accumulating effects of control on the sup-
pressed memory representations. As a result of mounting
effects of control, the need to further engage top–down
inhibitory control should decline as well, reflecting the
comparative rarity of intrusion events that require action.
The adaptive control account posits that this reduction
in the need for control should be reflected in dynamic
adaptation of DLPFC–HC coupling over blocks (比照. Wimber
等人。, 2011; Kuhl et al., 2007). Those individuals who
experience a greater reduction in involuntary intrusions
should progressively disengage this mechanism, as there
is diminishing need to purge unwanted memories from
意识. 因此, changes in DLPFC–HC coupling
over the course of the suppression phase should be tied to
the rate that the intrusion frequency declines across the
think/no-think phase.
We thus performed BMA of the members of Family 4
separately for each of the five runs and then calculated
the slope of the top–down effective connectivity across
blocks. This slope measure is positive for individuals who
show a weaker negative coupling over blocks. 尤其,
some participants exhibited negative slopes, reflecting that
they only developed a negative top–down connectivity
with repeated suppression attempts.
下一个, we examined the critical hypothesis that individ-
uals who cope better with intrusions, and who therefore
show a steep decline in intrusions over blocks (IE。, 一个更多
negative baseline-corrected behavioral intrusion slope),
will show a decreasing need to implement top–down in-
hibitory control as blocks progress. The coupling between
DLPFC and HC for efficient individuals should initially be
negative and becomes less negative over blocks; 因此,
their change in coupling should be more positive. Taken
一起, the adaptive control hypothesis therefore pre-
dicts a negative correlation between behavioral intrusion
slopes (which are more negative for highly efficient indi-
个人) and coupling parameter slopes (which should be
positive for efficient individuals who bring intrusions under
better control).
We observed this predicted negative correlation be-
tween the decline in intrusions and change in connectivity
(Figure 3B; r(16) = −0.49, p < .03, one-tailed). This sug-
gests that participants who showed the most precipitous
reduction in involuntary intrusions also showed a stronger
disengagement of the suppression mechanism over blocks.
Conversely, those who were least successful in reducing
memory intrusions may have modulated HC less overall.
Large Reductions in Intrusions Are Predicted
by Negative DLPFC–HC Coupling during
Early Suppressions
The relationship between change in connectivity strength
and decline in intrusion frequency suggests that an initially
negative coupling leads to a reduction in the number of
experienced intrusions on subsequent trials. Accordingly,
individuals who already exhibit a negative top–down
connectivity during earlier suppression attempts should
experience fewer involuntary intrusions of the suppressed
memories later on. Thus, the nature of the top–down con-
nectivity during the first half of the think/no-think phase
should predict the reduction in intrusions that participants
experience during the second half. To test this predic-
tion, we calculated the ratio of involuntary memory intru-
sions in the second versus first half, separately for each
participant. This value is smaller for those who experience
relatively fewer intrusions over time. We then split our
sample based on the effective top–down connectivity
parameter estimates to compare the intrusion reduction
for those individuals who exhibited negative coupling
parameters across the first two blocks of the think/no-think
phase (i.e., approximating the first half of the session) and
those who did not show evidence yet of implementing
Benoit et al.
107
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
o
2
c
0
n
3
_
7
a
/
_
j
0
o
0
c
6
n
9
_
6
a
p
_
d
0
0
b
6
y
9
g
6
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
an inhibitory suppression mechanism during that same
period. Consistent with a role of inhibitory modulation
during earlier trials in reducing involuntary intrusions on
later trials, early negative couplers experienced a greater
reduction in intrusions than individuals who did not exhibit
early negative coupling (Figure 3C; t(14) = −1.79, p < .05,
one-tailed).
Taken together, the data provide evidence for the adap-
tive deployment of the inhibitory top–down mechanism in
the purging of unwanted memories from awareness. Acti-
vation in DLPFC was strongest when unwanted memories
intruded into consciousness, and a greater upregulation
of this activation was associated with a steeper decline of
involuntary retrievals. Moreover, for place memories, a
negative DLPFC-to-HC coupling during earlier suppression
attempts predicted a greater reduction of involuntary in-
trusions on later trials. The coupling became weaker as
there was diminishing need for a reactive control mecha-
nism, further indicating that it was implemented adaptively
when there was a greater demand to counteract involun-
tary retrievals.
DISCUSSION
When unwanted memories intrude into consciousness,
people often seek to cope with these experiences by lim-
iting the memoriesʼ duration in awareness. What mecha-
nism achieves this purging, and how is it implemented in
the brain? Past work provided evidence for a direct sup-
pression mechanism that stops episodic retrieval, via neg-
ative coupling from DLPFC to HC (Gagnepain et al., 2014;
Benoit & Anderson, 2012). Here, we showed that the im-
pact of DLPFC on hippocampal activity can be upregulated
adaptively to purge unwanted memories from awareness
and that such counterintentional retrievals may actually
foster the engagement of processes that minimize invol-
untary memory intrusions over time.
To isolate the role of the DLPFC in purging memories
from awareness, participants reported memory intrusions
on a trial-by-trial basis. Whereas all suppression attempts
engaged the DLPFC, intrusions did so more robustly than
did nonintrusions. This indicates that processes supported
by DLPFC can be upregulated to countermand an unfold-
ing recollection. This elevated intrusion response occurred
independently of the mnemonic content (faces or places),
suggesting that DLPFC supports a central process during
memory suppression (cf. Burgess et al., 2006). Critically,
the DLPFC engagement during intrusions appeared effec-
tive in reducing the later intrusiveness of the suppressed
memories, predicting the decline in intrusions over sup-
pression repetitions. This pattern may indicate that the
reactivation of an intruding memory renders it more vul-
nerable to disruption (Detre et al., 2013; Levy & Anderson,
2012; Norman, Newman, & Detre, 2007), echoing research
on reconsolidation (Dudai, 2004; Nader, Schafe, & Le
Doux, 2000). In this reactivated state, inhibitory processes
supported by DLPFC may be particularly effective in dis-
rupting the memoryʼs trace, for example, by distorting its
hippocampal replay. This, in turn, would make it less
likely for the suppressed memory to be reactivated invol-
untarily on later trials, as indexed by a decrease in memory
intrusions.
The role of the DLPFC in reactive control received fur-
ther support from connectivity analyses. The models that
could account best for the place data were those that fea-
tured a modulation of the top–down connection from
DLPFC to HC, consistent with prior work on direct sup-
pression (Gagnepain et al., 2014; Benoit & Anderson,
2012). The effective connectivity between these regions
changed with the rate that participants had gained better
control over intruding memories. Those who had already
exhibited a negative DLPFC–HC coupling during initial
suppression attempts experienced less involuntary intru-
sions later on, and this top–down influence became progres-
sively weaker as the demand to purge involuntary intrusions
diminished. This pattern thus provides direct evidence for
the central assertion of the adaptive control hypothesis
(Kuhl et al., 2007), that is, that a benefit of weakening un-
wanted memories is the subsequent reduced demand on
mnemonic control. Whereas previous support for this
account was based on the assumption that successive re-
ductions in prefrontal engagement were a consequence of
successive weakening of the memories ( Wimber et al.,
2011; Kuhl et al., 2007), our online assessment of intrusions
allowed us to directly link the rate of decreasing control
with the rate of reduction in the memoriesʼ intrusiveness.
Together, these findings indicate that, for places at least,
the DLPFC adaptively modulates HC activity particularly
during intrusions and that a negative coupling contributes
to diminished intrusiveness of suppressed memories. This
modulation need not be direct, however, and may be
achieved via relay nodes such as other medial temporal
lobe structures or the retrosplenial cortex (Morris, Pandya,
& Petrides, 1999; Goldman-Rakic, Selemon, & Schwartz,
1984), especially given the lack of evidence for mono-
synaptic connections from DLPFC to HC.
The best models in the current experiment also fea-
tured a modulation of the connectivity from the HC to
the DLPFC during suppression, consistent with previous
work (Benoit & Anderson, 2012). Given that activation in
the HC may signal the involuntary retrieval of an unwanted
memory (cf. Levy & Anderson, 2012), the HC-to-DLPFC
connection could constitute a feedback loop that transmits
the requirement for an upregulation of the inhibitory pro-
cesses supported by the DLPFC.
Despite similar DLPFC activation for places and faces,
we did not observe reliable reductions in HC activation
when participants suppressed faces. Because of the ab-
sence of this effect, we could not use DCM for this class
of stimuli. A recent study reported a similar pattern of
material-specific differences during memory suppression
(Detre et al., 2013): whereas a classifier of fMRI data yielded
evidence for involuntary reactivation of place informa-
tion, this was not the case for faces. More specifically, the
108
Journal of Cognitive Neuroscience
Volume 27, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
o
2
c
0
n
3
_
7
a
/
_
j
0
o
0
c
6
n
9
_
6
a
p
_
d
0
0
b
6
y
9
g
6
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
present lack of a difference in HC activation may reflect
less dependence on HC processes during the retrieval of
faces, as suggested by striking material-specific effects in
amnesia showing that memory for unfamiliar faces may
be supported by structures outside the HC (Aly, Knight,
& Yonelinas, 2010; Bird & Burgess, 2008). The present lack
of HC activation for faces on recall trials, despite partici-
pants reporting face retrieval on 97% of the trials, may be
another instance of this material effect. As the neural basis
of covert episodic recall of faces is not widely studied, this
possibility must remain speculative. Our DCM evidence
for HC modulation during the suppression of places con-
verges, however, with similar modulations observed during
the suppression of verbal materials (Benoit & Anderson,
2012) and visual objects (Gagnepain et al., 2014), indi-
cating the generalization of this suppression mechanism
across other material types.
The progressive weakening of inhibited memories, as
indexed by a reduction in involuntary memory intrusions,
may eventually also lead to impaired voluntary access. In
the current study, we employed a training regimen that
was designed to create strong memories overall; yet, on
the final test, participants were slower to recognize pictures
that they had previously suppressed compared with ini-
tially equally strong baseline memories. This finding adds
to recent evidence that suppressing a memory can im-
pede subsequent recognition (Kim, Oh, Kim, Sim, & Lee,
2013; Hart & Schooler, 2012; Waldhauser, Lindgren, &
Johansson, 2012), thus extending previous evidence
for suppression-induced deficits in recall (Anderson &
Huddleston, 2011). Surprisingly, however, participants
were also slower to recognize pictures that they had
repeatedly recalled during the think/no-think phase,
although they had reported recalling those memories on
nearly all trials. Importantly, the slowed recognition of re-
call and suppress items did not correlate across subjects,
suggesting a different underlying mechanism. Our exten-
sive training of the cue–picture pairs may have contributed
to this effect, because further attempts to retrieve Recall
images during the Think/No-Think phase would have led
to minimal further strengthening. Instead, additional re-
trieval may have contaminated the memory representa-
tions of the items by incorporating additional incorrect
features (Schacter, Guerin, & St Jacques, 2011). Thus, on
the final test, a mismatch between the probe image and
its memory representation may have slowed recognition.
Future research is required to understand the cause of
this intriguing impairment.
Although our data indicate a role of DLPFC in purging
a memory from awareness, DLPFC may also serve a pro-
active gating function that suppresses retrieval before
an item reaches awareness. Several observations are consis-
tent with this possibility. First, the frequency of intrusions
on suppress trials was far lower than that of successful re-
calls on recall trials, even on the first repetition, suggesting
a considerable ability to prevent retrieval. Second, although
DLPFC was significantly more engaged by intrusions, it was
also recruited in the absence of intrusions (compared with
the recall condition), suggesting that it may not require
awareness of the unwanted memory to engage this control
process. Thus, whereas involuntary retrievals may trigger
elevated control, control may be present in all cases.
This raises the possibility that a memory intrusion may
simply be a proxy for a high level of reactivation of the
unwanted memory. If so, enhanced direct suppression
may not be triggered by an intrusion into awareness but
rather may be engaged to the degree to which a mem-
ory gets reactivated, irrespective of awareness (Levy &
Anderson, 2012). If suppression depends on a reactivation
level, this may explain why HC activation was also reduced
for nonintrusions, whereas it was not in prior work (Levy
& Anderson, 2012). The extensive training procedure
employed here created strong memory traces, as evident
by the overall high recognition rate on the final test. There-
fore, even those memories that did not intrude are likely
to have reached a moderate activation level in response
to their cues, necessitating some engagement of control.
However, when memories surpassed the activation thresh-
old for recollection, the suppression mechanism might
have had to be dynamically upregulated to purge the
intruding memory. Whether or not awareness is necessary,
our findings show that, when people attempt to exclude
an intrusive memory from awareness, they appear to
dynamically adjust a top–down mechanism that suppresses
retrieval processes supported by the HC.
Efforts to suppress awareness of unwanted memories
are ubiquitous in the aftermath of trauma, and involuntary
memory intrusions constitute an important symptom pres-
ent in many psychological disorders, including depression
and post-traumatic stress disorder (Ehlers et al., 2002). To
the extent that it disrupts traces, suppressing memories
may reduce intrusions over time. Protracted engagement
of suppression as a coping mechanism may, moreover,
alter the potential for top–down modulation of HC in gen-
eral. Indeed, in the year after a trauma, people undergo
significant cortical plasticity, confined largely to DLPFC re-
gions (Lyoo et al., 2011). Notably, individual variations in
the increase in cortical thickness predicted the long-term
recovery from post-traumatic stress disorder intrusion symp-
toms, and over time, cortical thickness returned to baseline
with the degree that the patients experienced an alleviation
of their symptoms. Although the mechanistic origins of this
DLPFC plasticity and its function remain speculative, the
present linkage between DLPFC engagement and the de-
mand to terminate awareness provides an important candi-
date process. By identifying foundational neurocognitive
mechanisms underlying peopleʼs coping response, the pres-
ent findings thus introduce an important hypothesis about
what alters peopleʼs mnemonic landscape and what permits
adjustment after trauma. This discovery has translational
potential for ameliorating the suffering of individuals for
whom these mechanisms break down. More broadly,
these findings point to a fundamental process by which we
regulate the contents of our awareness.
Benoit et al.
109
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
o
2
c
0
n
3
_
7
a
/
_
j
0
o
0
c
6
n
9
_
6
a
p
_
d
0
0
b
6
y
9
g
6
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Acknowledgments
We thank I. Charest and P. Gagnepain for assistance in data analy-
sis and A. Gazzaley and J. Rissman for providing the pictures.
This work was supported by the UK Medical Research Council
(MC-A060-5PR00).
Reprint requests should be sent to Roland G. Benoit, Department
of Psychology, Harvard University, Room 860, William James
Hall, 33 Kirkland Street, Cambridge, MA 02138, or via e-mail:
rbenoit@fas.harvard.edu.
REFERENCES
Aly, M., Knight, R. T., & Yonelinas, A. P. (2010). Faces are
special but not too special: Spared face recognition in
amnesia is based on familiarity. Neuropsychologia, 48,
3941–3948.
Anderson, M. C., & Green, C. (2001). Suppressing unwanted
memories by executive control. Nature, 410, 366–369.
Anderson, M. C., & Hanslmayr, S. (2014). Neural mechanisms
of motivated forgetting. Trends in Cognitive Sciences, 18,
279–292.
Anderson, M. C., & Huddleston, E. (2011). Towards a cognitive
and neurobiological model of motivated forgetting. In
R. F. Belli (Ed.), True and false recovered memories:
Toward a reconciliation of the debate, Vol. 58, Nebraska
symposium on motivation (pp. 53–120). New York: Springer.
Anderson, M. C., Ochsner, K. N., Kuhl, B., Cooper, J.,
Robertson, E., Gabrieli, S. W., et al. (2004). Neural systems
underlying the suppression of unwanted memories. Science,
303, 232–235.
Benoit, R. G., & Anderson, M. C. (2012). Opposing mechanisms
support the voluntary forgetting of unwanted memories.
Neuron, 670, 450–460.
Benoit, R. G., Gilbert, S. J., Frith, C. D., & Burgess, P. W.
(2012). Rostral prefrontal cortex and the focus of attention
in prospective memory. Cerebral Cortex, 22, 1876–1886.
Bergström, Z. M., de Fockert, J. W., & Richardson-Klavehn, A.
(2009). ERP and behavioural evidence for direct suppression
of unwanted memories. Neuroimage, 48, 726–737.
Bird, C. M., & Burgess, N. (2008). The hippocampus supports
recognition memory for familiar words but not unfamiliar
faces. Current Biology, 18, 1932–1936.
Burgess, P. W., Alderman, N., Forbes, C., Costello, A.,
Coates, L. M., Dawson, D. R., et al. (2006). The case for
the development and use of “ecologically valid” measures
of executive function in experimental and clinical
neuropsychology. Journal of the International
Neuropsychological Society, 12, 194–209.
Corallo, G., Sackur, J., Dehaene, S., & Sigman, M. (2008).
Limits on introspection: Distorted subjective time during
the dual-task bottleneck. Psychological Science, 19,
1110–1117.
Depue, B. E., Curran, T., & Banich, M. T. (2007). Prefrontal
regions orchestrate suppression of emotional memories
via a two-phase process. Science, 317, 215–219.
Detre, G. J., Natarajan, A., Gershman, S. J., & Norman, K. A.
(2013). Moderate levels of activation lead to forgetting in the
think/no-think paradigm. Neuropsychologia, 51, 2371–2388.
Dudai, Y. (2004). The neurobiology of consolidations, or,
how stable is the engram? Annual Review of Psychology,
55, 51–86.
Ehlers, A., Hackmann, A., Steil, R., Clohessy, S., Wenninger, K.,
& Winter, H. (2002). The nature of intrusive memories
after trauma: The warning signal hypothesis. Behaviour,
Research & Therapy, 40, 995–1002.
Eichenbaum, H., Yonelinas, A. P., & Ranganath, C. (2007).
The medial temporal lobe and recognition memory.
Annual Review of Neuroscience, 30, 123–152.
Eldridge, L. L., Knowlton, B. J., Furmanski, C. S., Bookheimer,
S. Y., & Engel, S. A. (2000). Remembering episodes:
A selective role for the hippocampus during retrieval.
Nature Neuroscience, 3, 1149–1152.
Friston, K. J., Harrison, L., & Penny, W. D. (2003). Dynamic
causal modelling. Neuroimage, 19, 1273–1302.
Friston, K. J., Holmes, A. P., Worsley, K. J., Poline, J.-P., Frith,
C. D., & Frackowiack, R. S. J. (1995). Statistical parametric
maps in functional imaging: A general linear approach.
Human Brain Mapping, 2, 189–210.
Gagnepain, P., Henson, R., & Anderson, M. C. (2014).
Suppressing unwanted memories reduces their unconscious
influence via targeted cortical inhibition. Proceedings of
the National Academy of Sciences, U.S.A., 111, 1310–1319.
Goldman-Rakic, P. S., Selemon, L. D., & Schwartz, M. L. (1984).
Dual pathways connecting the dorsolateral prefrontal cortex
with the hippocampal formation and parahippocampal
cortex in the rhesus monkey. Neuroscience, 12, 719–743.
Hart, R. E., & Schooler, J. W. (2012). Suppression of novel
stimuli: Changes in the accessibility of suppressed,
nonverbalizable shapes. Consciousness and Cognition,
21, 1541–1546.
Hertel, P. T., & Calcaterra, G. (2005). Intentional forgetting
benefits from thought substitution. Psychonomic Bulletin
& Review, 12, 484–489.
Kim, D., Oh, D., Kim, S. H., Sim, K., & Lee, J. (2013). Effects
of intentional suppression of recall of unwanted images
in repressors and non-repressors. Social Behavior and
Personality, 41, 319–326.
Kriegeskorte, N., Mur, M., & Bandettini, P. (2008).
Representational similarity analysis—Connecting the
branches of systems neuroscience. Frontiers in Systems
Neuroscience, 2, 4.
Kuhl, B. A., Dudukovic, N. M., Kahn, I., & Wagner, A. D. (2007).
Decreased demands on cognitive control reveal the neural
processing benefits of forgetting. Nature Neuroscience,
10, 908–914.
Küpper, C. S., Benoit, R. G., Dalgleish, T., & Anderson, M. C.
(2014). Direct suppression as a mechanism of controlling
unpleasant memories in daily life. Journal of Experimental
Psychology: General, 143, 1443–1449.
Levy, B. J., & Anderson, M. C. (2012). Purging of memories
from conscious awareness tracked in the human brain.
The Journal of Neuroscience, 32, 16785–16794.
Lyoo, I. K., Kim, J. E., Yoon, S. J., Hwang, J., Bae, S., & Kim,
D. J. (2011). The neurobiological role of the dorsolateral
prefrontal cortex in recovery from trauma. Longitudinal brain
imaging study among survivors of the South Korean subway
disaster. Archives of General Psychiatry, 68, 701–713.
Malian, J. A., Laurienti, P. J., Kraft, R. A., & Burdette, J. H.
(2003). An automated method for neuroanatomic and
cytoarchitectonic atlas-based interrogation of fMRI data
sets. Neuroimage, 19, 1233–1239.
Morris, R., Pandya, D. N., & Petrides, M. (1999). Fiber
system linking the mid-dorsolateral frontal cortex with the
retrosplenial/presubicular region in the rhesus monkey.
Journal of Comparative Neurology, 407, 183–192.
Nader, K., Schafe, G. E., & Le Doux, J. E. (2000). Fear memories
require protein synthesis in the amygdala for reconsolidation
after retrieval. Nature, 406, 722–726.
Noreen, S., & MacLeod, M. D. (2013). Itʼs all in the detail:
Intentional forgetting of autobiographical memories
using the autobiographical think/no-think task. Journal
of Experimental Psychology: Learning, Memory, and
Cognition, 39, 375–393.
110
Journal of Cognitive Neuroscience
Volume 27, Number 1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
o
2
c
0
n
3
_
7
a
/
_
j
0
o
0
c
6
n
9
_
6
a
p
_
d
0
0
b
6
y
9
g
6
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Norman, K. A., Newman, E. L., & Detre, G. (2007). A neural
network model of retrieval-induced forgetting. Psychological
Review, 114, 887–953.
Paz-Alonso, P. M., Bunge, S. A., Anderson, M. C., & Ghetti, S.
(2013). Strength of coupling within a mnemonic control
network differentiates those who can and cannot suppress
memory retrieval. The Journal of Neuroscience, 33,
5017–5026.
Penny, W. D., Stephan, K. E., Daunizeau, J., Rosa, M. J., Friston,
K. J., Schofield, T. M., et al. (2010). Comparing families of
dynamic causal models. PLoS Computational Biology, 6,
e1000709.
Schacter, D. L., Guerin, S. A., & St Jacques, P. L. (2011). Memory
distortion: An adaptive perspective. Trends in Cognitive
Sciences, 5, 467–474.
Sergent, C., & Dehaene, S. (2004). Neural processes underlying
conscious perception: Experimental findings and a global
neuronal workspace framework. The Journal of Physiology,
98, 374–384.
Stephan, K. E., Penny, W. D., Moran, R. J., den Ouden, H. E.,
Daunizeau, J., & Friston, K. J. (2010). Ten simple rules for
dynamic causal modeling. Neuroimage, 49, 3099–3109.
Waldhauser, G., Lindgren, M., & Johansson, M. (2012).
Intentional suppression can lead to a reduction of memory
strength: Behavioral and electrophysiological findings.
Frontiers in Psychology, 3, 401.
Wimber, M., Schott, B. H., Wendler, F., Seidenbecher, C. I.,
Behnisch, G., Macharadze, T., et al. (2011). Prefrontal
dopamine and the dynamic control of human long-term
memory. Translational Psychiatry, 1, e15.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
7
9
/
6
1
1
/
9
9
4
6
7
/
9
1
2
7
4
8
o
2
c
0
n
3
_
7
a
/
_
j
0
o
0
c
6
n
9
_
6
a
p
_
d
0
0
b
6
y
9
g
6
u
.
e
p
s
d
t
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Benoit et al.
111