A Differentiation Account of Recognition Memory:
Evidence from fMRI
Amy H. Criss1*, 马克E. Wheeler2*, 和詹姆斯·L. McClelland3
抽象的
■ Differentiation models of recognition memory predict a
strength-based mirror effect in the distributions of subjective
memory strength. Subjective memory strength should increase
for targets and simultaneously decrease for foils following a
strongly encoded list compared with a weakly encoded list. 一个
alternative explanation for the strength-based mirror effect is that
participants adopt a stricter criterion following a strong list than
a weak list. Behavioral experiments support the differentiation
帐户. The purpose of this study was to identify the neural
bases for these differences. Encoding strength was manipulated
(强的, weak) in a rapid event-related fMRI paradigm. To inves-
tigate the effect of retrieval context on foils, foils were presented
in test blocks containing strong or weak targets. Imaging analy-
ses identified regions in which activity increased faster for foils
tested after a strong list than a weak list. The results are inter-
preted in support of a differentiation account of memory and
are suggestive that the angular gyrus plays a role in evaluating evi-
dence related to the memory decision, even for new items. ■
介绍
Episodic memory is the ability to mentally time travel to a
past experience. One method for testing episodic memory
is a recognition task where participants are asked to en-
dorse targets that were studied and reject foils that were
不是. Memory has been extensively studied with imaging
and computational modeling techniques. 然而,
research connecting the two fields is slim (比照, Norman &
奥莱利, 2003). Our goal is to initiate a framework for
combining behavioral analysis, 成像, and modeling to
understand the role of strength in recognition.
Models of memory have successfully accounted for
many details of performance (see Malmberg, 2008). 一
exception is the role of list strength. When strength is
manipulated between lists, the result is a strength-based
mirror effect (SBME; Stretch & Wixted, 1998). Hit rates
(人力资源) are higher and false alarm rates (FAR) are lower for
a strongly encoded list compared with a weakly encoded
list and typically the HR differences are larger than the
FAR differences. Higher HRs for a strongly encoded list
are predicted by all models of memory. A challenge to
many models, 然而, is posed by the finding that the
FAR differs between the strong and weak lists. Foils are
drawn from the same set of items and are randomly placed
into a test list following weak or strong encoding. 有
no objective difference between foils tested after a weakly
versus a strongly encoded list other than the encoding
conditions of the target items. The criterion shift and dif-
1Syracuse University, 2University of Pittsburgh, 3斯坦福大学
*A. H. C. 和M. 乙. 瓦. contributed equally to this article.
ferentiation assumptions have been offered as competing
accounts for the SBME.
The Criterion Shift Assumption
One assumption is meta-cognitive; participants become
aware that accuracy for a strong list is high, during encod-
ing or the initial test trials, and consequently adopt a
strict criterion. The reduction in FAR for a strong relative
to a weak study list is accounted for by a change in the
criterion. This assumption is prominent in the class of
models that assumes the subjective memory strength of
unrelated foils is not affected by encoding strength, 作为
如图所示 1 (顶部). Dual process models assume
two different sources on which to base the memory deci-
锡安 (例如, recollecting the specific details of the event or
an overall feeling of familiarity absent the details) 然而
single process models assume just one basis for memory
决定. The criterion shift hypothesis has been used
in both types of models, with the change in criterion af-
fecting familiarity-based decisions (Cary & Reder, 2003;
Stretch & Wixted, 1998). Critically, these models assume
that the memorial evidence that a foil is from the study
列表 (例如, subjective memory strength) does not differ for
strong-list and weak-list foils.
Differentiation Models
In differentiation models, foils following a weakly and a
strongly encoded list differ in their distributions of sub-
jective memory strength. Such models need not assume
a change in the criterion to account for the SBME (看
© 2013 麻省理工学院
认知神经科学杂志 25:3, PP. 421–435
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
哦
8
C
6
n
5
_
7
A
/
_
j
0
哦
0
C
2
n
9
2
_
A
p
_
d
0
0
乙
2
y
9
G
2
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
F
t
.
/
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
Whether strengthening items on a list results in a change
in the memory distribution for foils and targets or a change
in criterion placement is an important theoretical ques-
的. Discriminating between a differentiation account
and criterion shift account of the SBME with only HRs
and FARs is impossible because the signal detection
parameters are saturated (four data points and four critical
参数). Evidence for differentiation models is accu-
mulating with studies using more informative dependent
措施. 例如, direct ratings of subjective mem-
ory strength support a priori predictions of differentiation
型号. 那是, participant-generated distributions of
memory strength following a strong and a weak list differ
as predicted by differentiation models. Critically, 这些
participant-generated distributions of memory strength
do not change with differences in target probability, A
classic response bias manipulation (Criss, 2009).
Analysis of RT distributions within the diffusion model
框架 (例如, 拉特克利夫, 1978) support differentiation
型号 (比照, Starns, 白色的, & 拉特克利夫, 2010). The diffusion
model describes how information is accumulated to reach
a decision. The better the quality of evidence provided by
the stimulus or the decision-maker, the faster the rate of
evidence accumulation, called the drift rate. In recognition
记忆, drift rate maps onto subjective memory strength
(拉特克利夫, 1978), thus differences in drift rate were pre-
dicted for strong-list foils and weak-list foils. Empirical dis-
tributions of RT were best fit by differences in the rate of
evidence accumulation (例如, drift rate) for strong and weak
targets and strong-list and weak-list foils, with larger mag-
nitudes of drift rates for targets and foils from the strong
列表 (Criss, 2010). The magnitude of the strength differences
and corresponding changes in drift rate need not be and
was not identical for targets and foils. 相比之下, manipu-
lating criterion by changing the percentage of targets at test
resulted in a different pattern of RT distributions that was
best accounted for by changes in the starting point of the
evidence accumulation process, not drift rate (Criss, 2010).
Despite a considerable body of data, 如上所述,
supporting predictions of differentiation models, 可能是
possible to account for the same findings within a criterion
shift account. 为此原因, we explored the possibility that
evidence from functional imaging studies could provide ad-
ditional evidence about the two accounts. The critical ques-
tion in the debate between criterion shift and differentiation
accounts of list strength is whether a change in response
bias or memory strength is necessary for the changes in
the FAR. We attempt to answer that question by comparing
strong versus weak conditions within brain regions where
activation is related to memory strength or response bias.
Neural Correlates of Memory Strength and
Response Bias
The neural correlates of episodic memory have been well
documented (for reviews, see Wagner, Shannon, Kahn, &
巴克纳, 2005; 巴克纳 & Wheeler, 2001). A collection
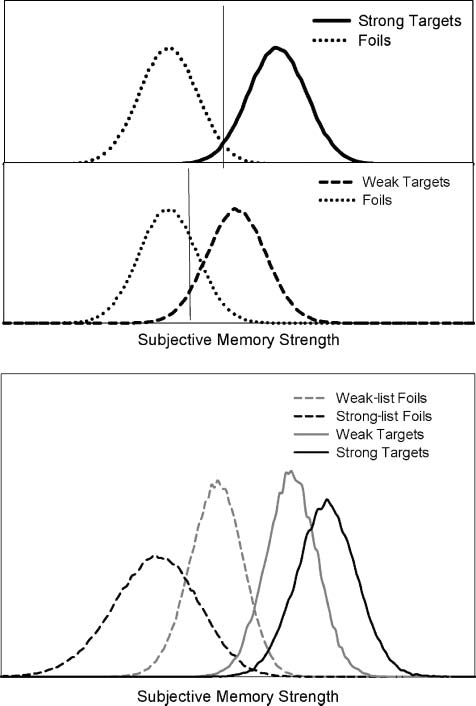
数字 1. Distributions of memory strength illustrating a criterion
shift account of the SBME (顶部). The vertical line represents the
criterion placement. Distributions generated from a differentiation
model showing that the memory strength of targets and foils differs
for strong and weak lists (底部).
数字 1, 底部; Criss & 麦克莱兰, 2006; 麦克莱兰 &
Chappell, 1998; Shiffrin & Steyvers, 1997). Differentiation
models assume that better encoding of target items re-
sults in more accurate memory traces. The more accurate
a given memory trace, the less likely it is that it will match
a foil decreasing the FAR. 换句话说, the more that
is known about an item, the less confusable that item is
with other items. This assumption causes the distribution
of subjective memory strength to increase for targets and
simultaneously decrease for foils tested after a strong
list compared with a weak list (see Criss, 2006, 2009,
2010). Like all memory models, differentiation models
do include a criterion that can be used strategically. 为了
例子, the criterion is changed in response to the pro-
portion of targets on the test list (例如, Criss, 2009). 在
this sense the differentiation hypothesis is more robust
whereas the criterion placement hypothesis can be re-
futed either by no evidence of a criterion shift or by evi-
dence for differentiation. For the present purposes, 这
critical point made by differentiation models is that the
criterion does not play a causal role in the SBME.
422
认知神经科学杂志
体积 25, 数字 3
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
哦
8
C
6
n
5
_
7
A
/
_
j
0
哦
0
C
2
n
9
2
_
A
p
_
d
0
0
乙
2
y
9
G
2
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
.
t
/
F
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
of retrieval success areas (RSA) consistently show differ-
ential activation when hits are contrasted with correct re-
jections (CRs) in recognition memory tasks. RSA include
inferior and superior parietal cortex, 前额皮质, pre-
cuneus, and cingulate gyrus (Simons et al., 2008; Wheeler &
巴克纳, 2004; Konishi, Wheeler, Donaldson, & 巴克纳,
2000; 麦克德莫特, 琼斯, 彼得森, Lageman, & Roediger,
2000; Henson, Rugg, Shallice, Josephs, & Dolan, 1999).
Very few studies of RSA have explicitly differentiated
between the contribution of memory strength and re-
sponse bias to memory performance and the resulting
neural activation in these areas. Many studies have shown
that some RSA track factors related to memory strength.
例如, regions in or near the left intraparietal sul-
cus are modulated by the subjective memory decision
(例如, differential activity to false alarms vs. misses; Kahn,
Davachi, & 瓦格纳, 2004; Wheeler & 巴克纳, 2003) 和
left lateral parietal regions near the angular gyrus (AG) 是
modulated by whether the response is recollection or fa-
miliarity based (Vilberg & Rugg, 2008; Wheeler & 巴克纳,
2004; Henson et al., 1999). Using subjective ratings of
strength, other studies have found a set of regions in which
activity correlates positively with memory strength, 在-
cluding left lateral parietal and inferior frontal cortex, 左边
thalamus, and bilateral medial parietal cortex (Montaldi,
Spencer, 罗伯茨, & Mayes, 2006; Yonelinas, Otten, Shaw,
& Rugg, 2005). Relatively few studies have evaluated the
role of RSA in response bias manipulations. Herron, Henson,
and Rugg (2004) manipulated the percentage of test items
that were targets and found that, as the ratio of old to new
items decreased, the difference in Hit and CR activation
increased in left superior parietal, left inferior frontal, 和
bilateral anterior frontal regions.
OʼConnor, Han, and Dobbins (2010) found correlations
between signal detection measures of memory strength
0) and response bias (C) with the contrast of Hit–CR
(d
across many frontal and parietal regions including several
RSA. Like OʼConnor et al., we differentiate between those
RSA that are modulated by response bias (RSAc) 和
those RSA modulated by accuracy (RSAd0).
预测
The goal of this study was to evaluate whether a crite-
rion shift or differentiation causes list strength effects for
FARs. Responses to targets as a function of list strength are
less informative because both accounts predict an increase
in HRs with list strength. The theories differ in their pre-
dictions for the memory strength of foils: Differentiation
models predict that the memory strength of foils decreases
as strength of targets increase and criterion shift models do
不是. We therefore focus our analyses on differences in ac-
tivations produced by foils at different levels of list strength.
We first identify retrieval success regions and determine
0, 因此
whether those areas are correlated with d
considered candidate memory strength areas or are corre-
lated with c, and therefore considered candidate response
bias areas. Such areas are designated RSAd0 and RSAc, 重新指定-
主动地. We then compare strong-list foils and weak-list foils
within the RSAc and RSAd0 to evaluate whether criterion shift
or differentiation accounts best describe the processes un-
derlying the SBME. If a criterion shift underlies list strength
effects, then we should see greater activity for strong list foil
trials than weak-list foil trials in RSAc (reflecting a list-wide
shift in the decision process).
If differentiation underlies list strength effects, we should
see differences between strong-list and weak-list foils in
regions sensitive to memory strength. 然而, 有
two alternative ways of thinking of how these differences will
be manifest. One possibility is that RSA represent the
strength of memory activation. 在这种情况下, illustrated in
数字 1, strong-list foils should produce even less activa-
tion than weak-list foils in these areas. Another possibility
and the one that is our primary focus here is that these
areas should be thought of as associated with accumulation
of evidence toward either of the two possible responses.
Strong-list foils lead to faster accumulation of evidence
toward the “new’ response than weak-list foils. 因此, 如果
RSA contain accumulators of evidence, we would expect to
see a faster change of activation in these areas to strong-list
foils than to weak-list foils (consistent with the Criss, 2010,
application of the diffusion model to the SBME), 尤其
in the most confident responses that represent the extreme
edges of the memory strength distributions.
The neural correlates of information accumulation have
been well documented in nonhuman primates and include
FEFs, lateral intraparietal area, superior colliculus, 和
dorsolateral pFC (金子 & 沙德伦, 2007; 沙尔, 2001).
Relatively less research has evaluated the neural substrates
associated with evidence accumulation in humans. Ho,
棕色的, and Serences (2009) identified a subregion of the
right insula whose activity is consistent with a modality-
independent evidence accumulator. In another case, Ploran
等人. (2007) controlled the amount of information pro-
vided by the stimulus as a means of manipulating informa-
tion accumulation. Information accumulation regions were
defined as those that became active immediately following
onset of the stimulus (例如, early in the time course) 和
whose subsequent rate of increase in activity was related
to the time taken to identify the stimulus. The rise in activ-
ity was steep for easy stimuli (identified early) and gradually
decreased with stimulus difficulty. We are not aware of any
studies that have evaluated the neural correlates of infor-
mation accumulation in memory. Our focus builds on the
rationale of Ploran et al. (2007), leading us to compare
the rate of information accumulation in strong-list versus
weak-list foils; 然而, we also consider alternatives.
方法
参加者
Twenty-nine right-handed native English-speaking volun-
teers with normal or corrected-to-normal vision participated
Criss, Wheeler, and McClelland
423
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
哦
8
C
6
n
5
_
7
A
/
_
j
0
哦
0
C
2
n
9
2
_
A
p
_
d
0
0
乙
2
y
9
G
2
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
t
/
F
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
in the study. Data from six participants were discarded:
three for chance level performance, two for insufficient
numbers of observations in some conditions, and one for
excessive head motion. Of the remaining 23 参与者,
15 were women. Participants ranged in age from 21 到
29 (米= 21.9). Participants gave informed consent as re-
quired by the institutional review board of the University
of Pittsburgh and were paid $25/hr for participation.
Stimuli
The word pool consisted of 700 words between 4 和
14 letters in length with a log frequency range between
8 和 14 (米= 10.483) among approximately 131 百万
words in the corpus (Balota et al., 2007).
Behavioral Paradigm
Studies focused on the cause of the SBME typically manip-
ulate strength via repetition, and we follow that tradition
这里 (though deep vs. shallow encoding may also produce
an SBME). Critically, the mechanisms underlying recog-
nition models operate in the same way whether informa-
tion is strengthened via encoding task or repetition (例如,
Shiffrin, 拉特克利夫, & 克拉克, 1990).
At study, participants received a weak and a strong
block, order counterbalanced. Each study list consisted of
100 unique words. For the weak lists, study words were
presented a single time for 1.5 sec with a 500-msec ISI.
For the strong lists, study words were presented for five
such presentations and the entire set of 100 words was
randomly ordered anew and presented before any word
repeated (equating study test lag for the most recent
presentation of weak and strong targets). The test list
began immediately after the study list with 100 targets
和 100 foils each presented for 750 msec followed by
A 2250 msec ISI. Participants responded to the question
“was this word on the list you just studied?” on a 4-point
规模 (sure yes, maybe yes, maybe no, sure no). The test
trials were intermixed with 100 fixation trials, 还 3 秒
in duration. Order of the test trials was generated using
the Buracas and Boynton (2002) 方法.
Image Acquisition
Images were acquired on a 3-T Siemens (Malvern, PA)
MAGNETOM Allegra system at the University of Pittsburghʼs
Brain Imaging Research Center. Before functional imaging,
a T1-weighted high-resolution magnetization prepared
rapid gradient-echo image (192 parasagittal slices, 1 mm3
voxels, repetition time = 1540 毫秒, echo time = 3.04 毫秒,
flip angle = 8°, inversion time = 800 毫秒) was acquired.
Functional images were collected during task perfor-
mance using a T2*-weighted echo-planar pulse sequence
sensitive to BOLD contrast (Kwong et al., 1992; Ogawa
等人。, 1992; repetition time = 1500 毫秒, echo time =
25 毫秒, flip angle = 79°, in-plane resolution = 3.2 ×
3.2 毫米, slice thickness = 3.5 毫米, 35 slices, interleaved
acquisition). The first five image acquisitions per run
were discarded to allow net magnetization to reach a
steady state.
程序
The entire experiment was conducted inside the scanner;
然而, functional images were only collected during
the test blocks. Participants were fully informed about
the experimental design before entering the scanner. A
brief practice block preceded the experimental blocks.
Responses were collected with a glove on each hand. 在-
dex fingers corresponded to “sure” and middle fingers
corresponded to “maybe.” The hand used to respond
yes or no was counterbalanced across subjects. Partici-
pants were asked to respond as quickly as possible with-
out sacrificing accuracy. The experiment was conducted
using E-prime (Psychology Tools, 公司, Pittsburgh, PA)
and stimuli were projected from the rear of the scanner
to a mirror positioned above the participantsʼ eyes. Anal-
ysis of ROI-based data was conducted using JMP software
( Version 8, SAS Institute, 公司, Cary, NC).
Functional MRI Data Analysis
Functional data were corrected for slice timing differences
using sinc interpolation, realigning all slices to the first
slice. Head motion was corrected within and across runs
using a rigid body algorithm with three translational and
three rotational parameters (斯奈德, 1996). Whole-brain
adjustment normalized the modal voxel value for all par-
ticipants to a value of 1000 to facilitate comparison be-
tween data sets (Ojemann et al., 1997).
After preprocessing, functional data from each partici-
pant were analyzed on a voxel-by-voxel basis using a gen-
eral linear model (GLM) 方法 (Ollinger, 舒尔曼, &
科尔贝塔, 2001; 米津, Maccotta, Ollinger, 彼得森, &
巴克纳, 2000; 弗里斯顿, Jezzard, & 车工, 1994). 这
BOLD data in each voxel were modeled as the sum of
coded effects at each time point, produced by modeled
events and by unexplained variance. Event regressors
were coded into each model at trial onset according to
list strength (强的, weak), item type (目标, foil), 和
准确性 (正确的, 不正确). Events were modeled
超过 12 time points beginning at trial onset, 生产
a time course of BOLD activity for each event spanning
18 秒. Linear trend and constant regressors were in-
cluded for each run. A series of delta functions described
event-related effects as estimates of the percent signal
change from the baseline term. This approach makes
no assumptions about the shape of the BOLD response.
Image processing and analyses were conducted using soft-
ware developed at Washington University (Ollinger et al.,
2001).
To identify RSA, an ANOVA was conducted using GLM
parameter estimates from each participant, with Subject
424
认知神经科学杂志
体积 25, 数字 3
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
哦
8
C
6
n
5
_
7
A
/
_
j
0
哦
0
C
2
n
9
2
_
A
p
_
d
0
0
乙
2
y
9
G
2
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
F
/
.
t
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
treated as a random factor and Time (each whole-brain
体积) as a repeated measure. A voxelwise mixed effects
repeated-measures ANOVA, with fixed factors of Strength
(强的, weak), Item Type (目标, foil) and Time (12 时间
点), was computed on correct trials, collapsing over
confidence response. This analysis produced a set of
main effect and interaction images, one for each term
in the ANOVA model. The interaction image of Item
Type (correct target vs. correct foil) × Time was used to
define retrieval success ROIs. This image displays the
degree to which activity on correct target trials differs from
activity on correct foil trials across the 12 modeled time
点.
During group analyses, BOLD data were resampled into
2-mm isotropic voxels and transformed into stereotaxic atlas
space by aligning an individual participantʼs T1-weighted
image to a Talairach atlas-transformed T1-weighted tem-
plate using a series of affine transformations (狐狸, 斯奈德,
Barch, Gusnard, & Raichle, 2005; Michelon, 斯奈德,
巴克纳, McAvoy, & Zacks, 2003; Lancaster et al., 1995;
Talairach & Tournoux, 1988). The ANOVA produced an
F-to-z-transformed statistical image, smoothed using
a 6-mm FWHM Gaussian kernel, for each term in the
ANOVA. These uncorrected images were then corrected
for multiple comparisons and sphericity. Criteria for multi-
ple comparison corrections were based on Monte Carlo
simulations (McAvoy, Ollinger, & 巴克纳, 2001), 与一个
cluster-size Type I error rate of p < .05 at a 70 voxel extent.
Although all reported data are from voxels meeting correc-
tion criteria, both corrected and uncorrected images were
retained for use in the ROI definition procedure (described
in the next section). Some activations and ROIs are dis-
played on cortical surface representations using Caret soft-
ware (Van Essen et al., 2001) using the population-average,
landmark, and surface-based atlas (Van Essen, 2005).
Region Definition Procedures
To define ROIs, uncorrected smoothed group z statistical
images were resmoothed using a 4-mm hard sphere
kernel to reduce the number of peaks in the volume. An
algorithm searched for the location of signal change peaks
exceeding p < .001 significance, and ROI volumes were
grown up to a maximum 10 mm radius of contiguous
voxels around the peak coordinates, including only voxels
passing threshold. Peaks separated by <10 mm were con-
solidated by averaging their xyz coordinates. Voxels failing
to pass the sphericity and multiple comparisons correc-
tions described in the previous section were then excluded
from the ROIs. Using this approach, only corrected voxels
were retained in the ROIs.
Correlational Analyses of fMRI and
Behavioral Measures
The relationship between memory performance and re-
trieval success activity was assessed by determining the
0 and c statistics cor-
degree to which a participantʼs d
related with an estimate of the retrieval success effect
(RSE), the difference in peak activity on Hit and CR trials
(OʼConnor et al., 2010). To compute peak activity in each
ROI, the magnitude of the BOLD response on correct
target and foil trials was averaged over Time Points 4,
5, and 6 (4.5, 6.0, and 7.5 sec from trial onset). This range
of time points was selected because it includes the peak
time points in most regions. We note that retrieval suc-
cess has been characterized both by an interaction of
Item Type and Time (in Functional MRI Data Analysis
section) and by differences in peak activity between
correct target and foil trials (this section). The two ap-
proaches should have high agreement because most
of the time the interaction of item type with time was
significant due to differences at peak. This was verified
by evaluating Hit–CR differences at the peak of the
time course (defined as Time Points 4, 5, and 6) in each
ROI (listed in Table 3). We conducted a 2 × 2 ANOVA
with factors of Item Type (target, foil) and Strength
(strong, weak). The main effect of Item Type was sig-
nificant ( p < .05) in all ROIs except ROI 8, left thalamus,
F(1, 22) = 2.89, p = .09, no corrections for multiple
comparisons.
The ROI analysis was followed by an exploratory voxel-
wise analysis in which correct target and foil time series
were convolved with a gamma function. This approach
was chosen because it provided increased power, helpful
in voxelwise analyses, relative to the time-course-based
analysis used in the ROI analysis. The scale parameter
0 and c in two separate anal-
(β) was regressed against d
yses. Images were smoothed using a 4-mm FWHM Gaussian
smoothing kernel. The resulting statistical maps were
corrected for multiple comparisons as described in
the previous section, with a cluster size threshold of
100 voxels and Type I error rate of p < .05 (McAvoy
et al., 2001).
RESULTS
Behavioral Results
As shown in Table 1, we find an SBME as expected. One-
tailed paired t tests showed that HRs were higher for
strong than weak targets, t(22) = 8.74, p < .001. FARs were
numerically smaller for strong-list than weak-list foils, but
the magnitude failed to reach significance, t(22) = 1.200,
p = .122. The power to detect significant changes in be-
havior is relatively low given the small number of partici-
pants. We collected pilot data in the behavioral paradigm
in which 18 volunteers participated in the exact behavioral
paradigm used in the scanner including fixation trials. The
combined data (excluding 1 pilot participant whose per-
formance was at chance) show an SBME: Strong HRs
are higher than weak HRs, t(39) = 9.39, p < .001, and
strong-list FARs are lower than weak-list FARs, t(39) =
2.47, p = .009. For comparison with neural activation
Criss, Wheeler, and McClelland
425
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
o
8
c
6
n
5
_
7
a
/
_
j
0
o
0
c
2
n
9
2
_
a
p
_
d
0
0
b
2
y
9
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 1. The Probability of Calling a Test Item Old as a Function of Type of Test Item and List Strength
fMRI Subjects
Strong
Weak
Pilot and fMRI Participants
Strong
Weak
Targets
Foils
.775 (.026)
.350 (.036)
.609 (.031)
.390 (.031)
Targets
Foils
.795 (.019)
.310 (.032)
.603 (.026)
.377 (.027)
times and for archival purposes, median RT is included in
Table 2.
Imaging Results
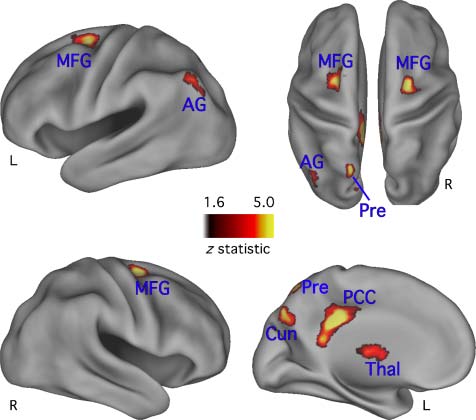
Retrieval Success
The retrieval success analysis revealed reliably different
Hit versus CR activity in or near bilateral middle frontal
gyrus (MFG) and the left posterior cingulate gyrus, AG,
thalamus (thal), and precuneus (Table 3 and Figure 2).
These locations are consistent with commonly reported
RSA (Simons et al., 2008; Vilberg & Rugg, 2008; Wagner
et al., 2005; Buckner & Wheeler, 2001). To investigate
the nature of the RSEs, ROIs were defined from the
corrected retrieval success statistical map (see Methods)
and activity on strong-list and weak-list target and foil
trials, averaging over levels of confidence, was evaluated
in each ROI. The time course of the BOLD response from
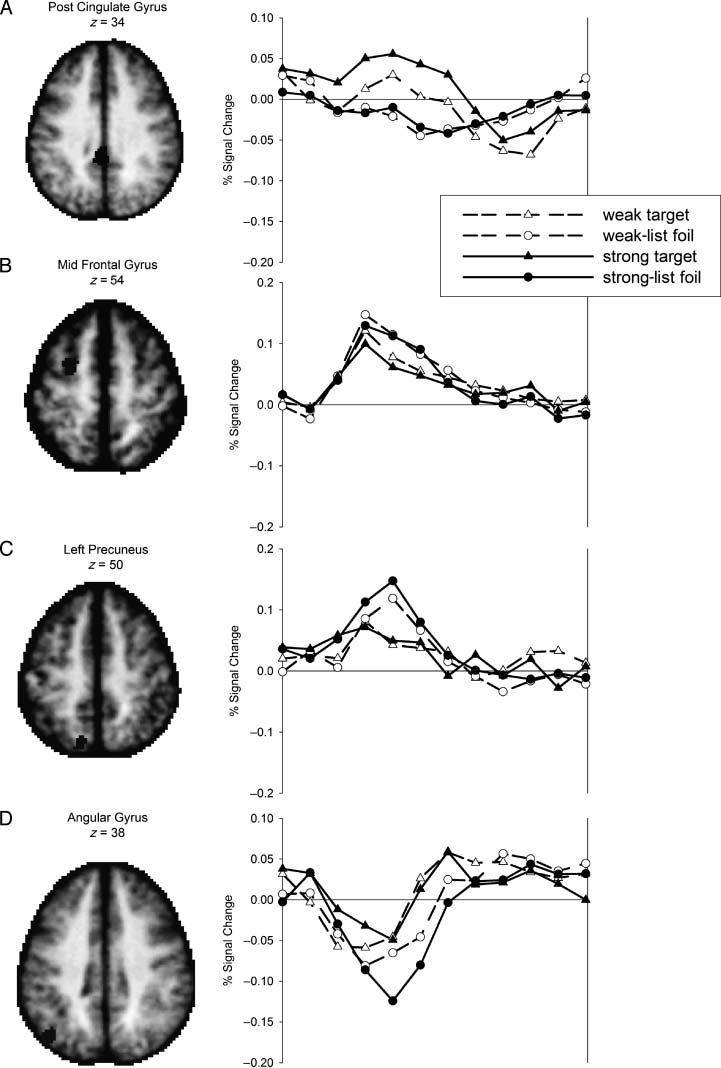
four regions are displayed in Figure 3. Consistent with prior
reports (Nelson et al., 2010; Wheeler & Buckner, 2004),
activity in the AG ROI (Figure 3D) displayed a decrease
relative to the GLM baseline term, with a greater mag-
nitude of decrease on foil than target trials (Figure 3D).
Whereas activity in retrieval success ROIs is often greater
on target than on foil trials, activity in four of the retrieval
success ROIs showed the reverse pattern. These regions
were located in or near bilateral MFG (Table 3, Nos. 3, 5,
and 6) and left precuneus (Table 3, No. 7). This pattern of
activation may reflect facilitation in processing related to
prior exposure of the targets (e.g., Grill-Spector, Henson,
& Martin, 2006).
Identifying RSAc and RSAd0
To identify memory strength and response bias compo-
nents, the peak activity on Hit minus peak activity on CR
trials was computed for each participant. For each par-
ticipant, this value was correlated with two measures of
Table 2. Median RT in Milliseconds for Each Condition and
Each Response Type
“Old”
“New”
Targets
Foils
Strong
527
903.5
Weak
705
828.5
Strong
Weak
975
797.5
837
796
behavioral performance, bias (c) to identify RSAc and sen-
0) to identify RSAd0. Strong and weak trials were
sitivity (d
analyzed separately. R values for each ROI for strong and
weak conditions are listed in Table 3.
This analysis identified correlations of the RSE with c in
both left MFG ROIs in the strong condition only (R = .47,
p = .02 and R = .42, p = .04). RSE did not correlate with
0 for the strong condition in either of these regions (R =
d
−.32, p = .13 and R = −.35, p = .11). However, a correla-
0 was found in the left AG ROI in the strong
tion with d
condition (R = .53, p = .009). The RSE in AG did not,
however, correlate with c (R = .26, p = .23). The only
0 was the left
other retrieval success ROI correlating with d
caudate nucleus (R = .46, p = .03), but only in the weak
condition. Correlations for the strong condition in the AG
ROI are plotted in Figure 4A. On the basis of these analyses,
the left AG is a RSAd0 and candidate memory strength re-
gion, and the left and right MFG are RSAc and candidate
response bias regions.
Analysis of Foils in RSAd0
If differentiation underlies list strength effects, we should
see faster onset of information accumulation for strong-list
than weak-list foil trials in RSAd0. As described earlier, in
an information accumulation framework (e.g., diffusion
model), stimuli with high-quality evidence accumulate
activity toward a decision boundary quicker than stimuli
with lower quality evidence. According to differentiation
models, strong-list foils have higher-quality evidence (more
extreme memory evidence) than weak-list foils, and there-
fore, strength should affect the rate of increase in activity
from trial onset. CRs (not FARs) provide critical evidence
of the difference in rate of accumulation because they
end at the correct decision boundary, whereas FARs ter-
minate at the wrong decision boundary (and may not fol-
low the expected pattern based on the magnitude of the
drift rates).
To evaluate this prediction, we used the Ploran et al.
(2007) method, focusing analysis on activity early in the time
series, just after trial onset (averaged across Time Points 2
and 3). Ploran et al. (2007) found early differences in the
time to reach peak activity (rather than the magnitude of
peak activation) for stimuli that were easier to identify in a
perceptual task (e.g., provided better evidence). Following
their logic, we looked for differences in early activation
for strong-list foils that are easier to reject (e.g., provided
better evidence). A differentiation account would be
supported by strength-based differences in the magnitude
426
Journal of Cognitive Neuroscience
Volume 25, Number 3
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
o
8
c
6
n
5
_
7
a
/
_
j
0
o
0
c
2
n
9
2
_
a
p
_
d
0
0
b
2
y
9
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 3. ROI Atlas Coordinates and Approximate Anatomic Locations of Retrieval Success Regions and Correlations with d
0 and c
No.
Hem
Anatomic Location
x
1
2
3
4
5
6
7
8
9
10
11
L
L
R
L
L
L
L
L
L
L
L
Post. Cingulate G.
Cuneus
Mid. Frontal G.
Thalamus
Mid. Frontal G.
Mid. Frontal G.
Precuneus
Thalamus
Post. Cingulate G.
Angular G.
Caudate
−01
−04
+22
−10
−24
−17
−13
−03
−03
−42
−10
y
−39
−79
−07
−05
−03
+04
−68
−17
−47
−70
+08
z
+29
+34
+55
+14
+55
+57
+52
+11
+17
+37
+08
∼BA
#Vox
c-str
c-wk
0
d
-str
0
-wk
d
31
19
6
NA
6
6
7
NA
30
39
NA
306
289
191
141
171
104
74
93
106
74
80
.18
.03
.27
−.05
.47*
.42*
.37
.15
.22
.26
−.27
.17
.02
−.03
.09
.34
−.19
.37
−.02
.05
.24
.08
.32
.36
−.39
−.13
−.32
−.35
.28
.15
.37
.53**
−.21
.21
.23
−.24
−.09
−.15
.12
−.22
−.28
.10
.35
.46*
Hem = hemisphere; L = left; R = right; G = gyrus; Inf = inferior; Mid = middle; Post = posterior; BA = Brodmannʼs area; NA = not applicable;
#vox = number of voxels in ROI; all locations and Brodmannʼs areas are approximate. The rightmost four columns are Pearson r values for the para-
meter (c or d
0) and RSE for the strong (str) and weak (wk) conditions.
*p < .05.
**p ≤ .01.
of activity at Time Points 2 and 3 (strong-list foil CRs > weak-
list foil CRs, indicating a faster approach to peak activation
for strong-list foils) in the left AG, the candidate memory
strength region. From a diffusion model perspective, 峰值
magnitudes need not differ; however the rate of accumula-
tion should be faster for strong-list foils. 然而, t tests
数字 2. Corrected retrieval success map, projected onto inflated
cortical surfaces. Lateral view of the left and right hemispheres are in
the first and second row, 分别. Top row right side shows a dorsal
view of both hemispheres. Bottom row right side shows a medial view
of the left hemisphere. The reliability of activation is indicated by the
scale bar, in z score units. L = left; Cun = cuneus; PCC = posterior
cingulate cortex; Pre = precuneus; Thal = thalamus.
(one-tailed) revealed no strong > weak differences in the
left AG or any other retrieval success ROI (all p ≥ .23).
Because the most robust memory strength effects are
observed in the most confident responses (例如, see Criss,
2009, 2010), we performed the same analyses using only
“sure” trials. Including confidence in the analysis required
us to eliminate five participants who lacked a sufficient
number of trials in each condition. This analysis revealed
a significant early strength effect (strong > weak) on “sure”
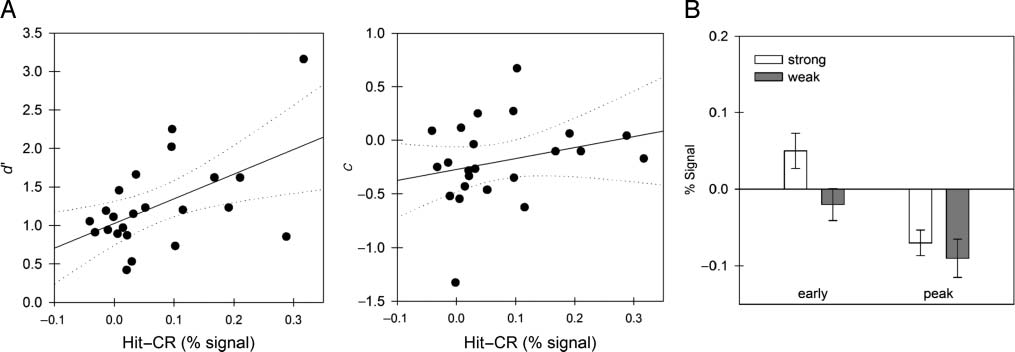
responses to foils in the left AG ROI, t(17) = 2.01, p = .03,
shown in the left half of Figure 4B (“early”). None of the
other retrieval success ROIs approached significance. Peak
activity in AG did not differ between conditions, as shown
in Figure 4B (“peak”). It should be noted that the “peak”
activations in this case are actually well below the baseline.
This is true both for Hits and CR (see Figure 3D). 这是
consistent with our account under the assumption that
there is an overall drop in activation in this region during
memory task performance; the evidence accumulation
process may occur as a positive-going activation on top
of this overall drop in activation, or it may occur as a de-
crease in activity. Although we have stressed the possibil-
ity that left AG might profitably be viewed as an evidence
accumulator region, with activation increasing as evidence
is accumulated, one may also consider the possibility that
AG activation might correspond to the total memory activa-
tion produced by the stimulus. In this way of thinking, 我们
would expect the BOLD signal strength to be greatest for
strong targets, then to decrease for weak targets and
weak-list foils, with the lowest signal strength for strong-list
foils (as seen in Figure 1, bottom panel). The expected dif-
ference between weak-list and strong-list foils is visually
Criss, Wheeler, and McClelland
427
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
哦
8
C
6
n
5
_
7
A
/
_
j
0
哦
0
C
2
n
9
2
_
A
p
_
d
0
0
乙
2
y
9
G
2
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
.
/
F
t
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
数字 3. Time courses of the
BOLD response for a subset of
the RSA. Percent signal change
from baseline is plotted over
12 time points from stimulus
onset for Post Cingulate Gyrus
(A), Mid Frontal Gyrus (乙), 左边
Precuneus (C), and Angular
Gyrus (D).
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
哦
8
C
6
n
5
_
7
A
/
_
j
0
哦
0
C
2
n
9
2
_
A
p
_
d
0
0
乙
2
y
9
G
2
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
t
/
F
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
suggestive in left AG (see Figure 3D). 然而, 不同之处-
ence in the peak the BOLD response is not significant.
Analysis of RSAc
If a criterion shift underlies list strength effects, 然后我们
should see different activity for strong-list compared with
weak-list foil trials in RSAc (left and right MFG). Data from
each retrieval success ROI were entered into separate
mixed effects repeated-measures ANOVA models, 和
Item Type (correct target, correct foil), Strength (强的,
weak), and Time (12 time points) as factors (描述的
in Retrieval Success section). A list strength effect was
evaluated using the Strength × Time and Item Type ×
Strength × Time interaction terms. No interactions of
these types were observed in the retrieval success ROIs.
Thus RSAc failed to respond to list strength.
In Analysis of Foils in RSAd0 section, we report that five
participants were excluded from analyses of “sure” re-
sponses to foils because they lacked a sufficient number
of such responses. To ensure that these participants did
not obscure the analysis just reported, we reran the analysis
428
认知神经科学杂志
体积 25, 数字 3
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
哦
8
C
6
n
5
_
7
A
/
_
j
0
哦
0
C
2
n
9
2
_
A
p
_
d
0
0
乙
2
y
9
G
2
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
/
F
t
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
0; 左边) 和
数字 4. The two panels in Panel A show the correlation between activity in left AG and measures of performance. Discrimination (d
bias (C; 中间) are plotted as a function of RSE (Hit–CR percent signal change) for each individual participant. The RSE is correlated with d0 but
not c. Panel B shows the magnitude of activation early (averaged over Time Points 2 和 3) and at peak (averaged over Time Points 4, 5, 和 6) in AG.
Differences between strong-list and weak-list foils early in the time course indicate differences in the rate of evidence accumulation, consistent with
differentiation models.
excluding those participants. No interactions were observed.
此外, we conducted the ANOVAs separately for
“sure” and “maybe” responses and found no interactions.
Despite multiple attempts, we found no evidence support-
ing the hypothesis that RSAc respond to list strength.
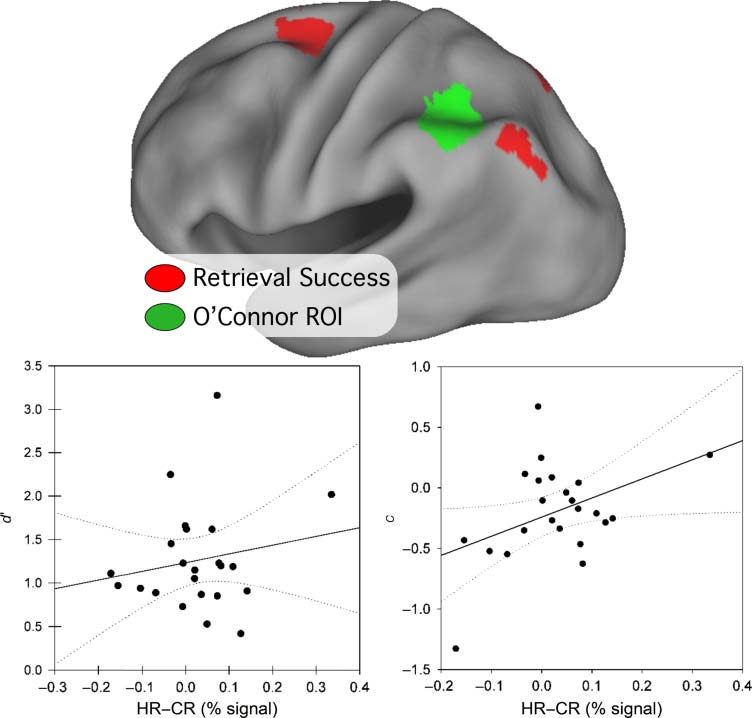
Comparison with OʼConnor et al. (2010)
OʼConnor et al. (2010) identified a region of left parietal
lobe that correlated significantly with bias (C), 但不是
0). Together with a voxel-based analysis, 他们的
灵敏度 (d
findings suggest that the function of the parietal RSA are
related more to overcoming response bias than success
at retrieval or memory accuracy. To compare our findings
with those of OʼConnor and colleagues (2010), a 10-mm
diameter sphere was created around their peak AG co-
纵坐标 (Talairach x, y, z = −42, −50, 41), after using the
nonlinear transformation method developed by Matthew
Brett (imaging.mrc-cbu.cam.ac.uk/imaging/MniTalairach)
to convert their Montreal Neurological Institute (MNI) 共-
ordinates to Talairach atlas space. The resulting ROI, 迪斯-
played in Figure 5 along with our retrieval success ROIs, 是
located anterior to our AG RSAc. Time courses were ex-
tracted for Hits and CRs, and correlation analyses were per-
形成的. Consistent with the findings of OʼConnor et al., 我们
found that the RSE in their ROI correlated positively and
reliably with c, but only in the strong condition (R = 0.42,
p = .04; weak: R = 0.30, p = .17), 如图 5.
0 for strong (R = .18,
There were no correlations with d
p = 42) or weak (R = .07, p = .76) 状况.
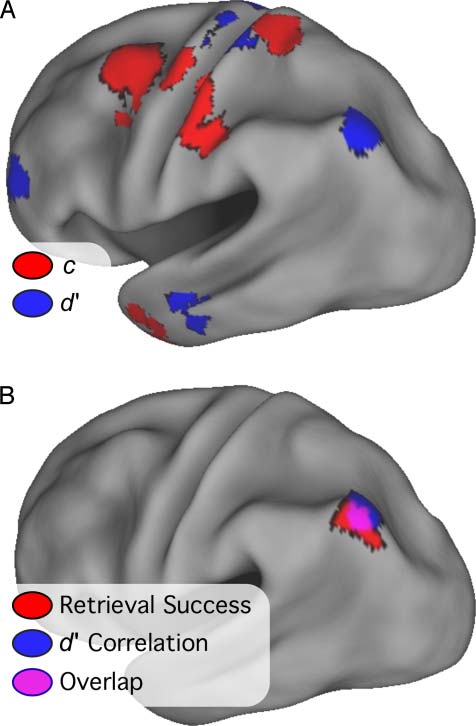
voxel-by-voxel analyses, with Subject as a random factor.
The z-transformed and multiple-comparison corrected
images are shown in Figure 6A, overlaid onto an inflated
left hemisphere cortical surface. Regions in which activity
0 were found in bilateral MFG
significantly correlated with d
near BA 6 and BA 10, left postcentral gyrus, left AG (−36,
−70, 39), and bilateral cerebellum (桌子 4).
Regions in which activity correlated positively with c
included bilateral precentral and postcentral gyrus, bilateral
MFG, bilateral cerebellum, right inferior parietal lobe, 和
right fusiform gyrus (Figure 6A; 桌子 5). Many of the re-
gions in this map were located in or near structures typi-
cally involved in motor planning or execution, 包括
premotor cortex, primary somatosensory/motor cortex,
cerebellum, and BG. All regions in the left parietal lobe
were located anterior or superior to the lateral parietal
0. There were very few voxels
regions that correlated with d
in which retrieval success activity correlated with both
0 and c (Figure 6B, overlap would appear in violet).
d
None of these voxels formed an isolated region, 但反而
0
existed at the borders of regions correlating either with d
or c. This observation indicates a sharp distinction between
regions involved in signaling successful retrieval and those
involved in attentional or strategic demands.
The AG region identified in the exploratory voxelwise
correlation analysis was located near the AG retrieval success
ROI reported in Table 3. The spatial relationship between
these two regions was explored by overlaying them onto
an inflated cortical surface. As shown in Figure 6B, 那里
was a high degree of overlap between the ROIs.
Exploratory Voxelwise Analysis of the Relationship
0
between the RSE and d
and c
讨论
Differentiation in Episodic Memory
To explore the distribution of voxels sensitive to d
RSE was correlated with d
0 and c,
0 and c in separate mixed-effects
The goal of this article was to use fMRI to evaluate whether
differentiation or criterion shifts underlie the SBME in
Criss, Wheeler, and McClelland
429
0 (左边) and c
数字 5. The ROI derived
from OʼConnor and colleagues
(2010) is shown in green,
overlaid with the retrieval
success ROIs (红色的) onto an
inflated cortical surface. 这
bottom row plot d
(正确的) as a function of RSE
(HR–CR percent signal change)
from the OʼConnor ROI. 每个
data point represents values
from a single subject. c but not
0 is correlated with RSE in this
d
ROI, replicating the OʼConnor
等人. finding.
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
哦
8
C
6
n
5
_
7
A
/
_
j
0
哦
0
C
2
n
9
2
_
A
p
_
d
0
0
乙
2
y
9
G
2
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
F
.
t
/
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
behavioral data. It is of course possible that both differen-
tiation and a criterion shift contribute to the magnitude of
the SBME. 然而, several imaging analyses converged
to indicate accumulator-like behavior in or near the AG,
supporting a differentiation account. AG showed a Hit–
CR difference that grew in magnitude across subjects as
0 增加. This region also displayed early differential
d
strength-based effects in the foil conditions, with activity
increasing more during the early time points for strong-
list foils than weak-list foils on high confidence trials (如图-
ure 4B, “early”). A voxelwise analysis revealed a region
0 and that overlapped
in left AG that correlated with d
with the left AG retrieval success ROI (Figure 6B). 最后,
AG activity showed no relation to response bias. One inter-
pretation of this set of converging findings is that the left
AG serves as one source of information accumulation rel-
evant for a memory decision. 然而, a conclusive state-
ment about the role of the AG in memory awaits further
证据.
A variety of criterion shift models can mimic predictions
from differentiation models (例如, see Stretch & Wixted,
1998). 然而, this reasoning fails when faced with the
full set of relevant data. Investigations of differentiation
with purely behavioral data included an important com-
parison condition that is not included here, 即, 与-
dition manipulating the percentage of test trials that are
targets. This manipulation is widely regarded as a response
bias manipulation. If both paradigms (% target manipula-
tion and SBME) are driven by changes in criterion location,
then a similar pattern of behavior should emerge. 在一个
sophisticated analysis of RT distributions (Criss, 2010)
and of participant-generated memory strength distribu-
系统蒸发散 (Criss, 2009), qualitatively different patterns of
结果, requiring different model parameters, 出现了.
Two different processes appear to underlie behavioral per-
formance in the SBME and % target paradigms. Our data
cannot evaluate the neural correlates of manipulating
target probability. 幸运的是, there are two reports mea-
suring fMRI in a % target paradigm discussed in the
Response Bias in Anterior and Superior Parietal Lobe sec-
的. As a preview, both experiments are consistent with
the conclusion we have reached based on the data re-
ported here: Regions sensitive to response bias are not
the same as regions sensitive to memory accuracy.
Characterizing neural activation as reflecting memory
strength and response bias and especially describing the
role of the left AG as reflecting differentiation is a slightly
different nomenclature than is typical in the literature.
430
认知神经科学杂志
体积 25, 数字 3
of parietal regions and evaluated RSEs from six studies
across regions. Their analyses also categorized the left AG
(−45, −67, 36 in MNI-to-Talairach coordinates) with other
default mode regions, including the right AG, 后部
cingulate, medial and superior frontal, and anterior tem-
poral cortex. Vincent and colleagues used hippocampal
ROIs as seed points and found correlated voxels in bilateral
AG (Kahn, Andrews-Hanna, Vincent, 斯奈德, & 巴克纳,
2008; Vincent et al., 2006).
The AG may temporarily maintain information retrieved
from memory as indicated by several studies showing a
relationship between the magnitude of signal change and
the degree to which retrieval involves recollection. 为了
例子, studies using source memory or remember/
知道 (RK) paradigms have been associated with less of a
decrease in activity in lateral parietal ROIs near the AG
when retrieval involves recollection than when it involves
familiarity (Wheeler & 巴克纳, 2004; Dobbins, Foley,
Schacter, & 瓦格纳, 2002; Henson et al., 1999). 两个螺柱-
ies using a graded memory strength measure (Montaldi
等人。, 2006; Yonelinas et al., 2005) found nearby parietal
地区 ( Yonelinas: −33, −56, 36; Montaldi: −39, −68,
39; MNI-to-Talairach coordinates) in which there was less
of a decrease in activity as subjective memory strength
增加. Vilberg and Rugg (2007) had subjects study pairs
of picture stimuli and used a variant of the RK paradigm
with two levels of remember response based on amount
of recollected information (IE。, 和 [R2] or without [R1]
the studied associate). They found a parietal region near
the AG (−39, −77, 40 in MNI-to-Talairach coordinates) 那
was selective for remember responses and increased ac-
tivity with amount recollected (R2 > R1). 作者
posited that the region operates as an “episodic buffer”
桌子 4. List of ROIs with Positive Correlations between
0
RSE and d
# Hem Anatomic Location
X
y
z
∼BA #Vox
1
2
3
4
5
6
7
8
9
10
11
L Med. Frontal G.
−07 −24
72
6
R Med. Frontal G.
03
50
07 10
L
L
右
Postcentral G.
Postcentral G.
小脑
−29 −28
−06 −41
67
72
3
5
47 −63 −32 NA
L Mid. Frontal G.
−38
44
13 10
L Mid. Temporal G.
−54 −18 −20 21
196
152
169
141
222
109
106
L
右
L
L
Angular G./Precuneus −36 −70
39 39/19
102
小脑
小脑
小脑
05 −25 −34 NA
−15 −42 −48 NA
−37 −44 −49 NA
108
133
126
Hem = hemisphere; R = right; L = left; Med = medial; Mid = middle;
G = gyrus; BA = Brodmannʼs area; #vox = number of voxels in ROI; 全部
locations and Brodmannʼs areas are approximate.
Criss, Wheeler, and McClelland
431
数字 6. Significant and corrected voxelwise correlations with c (红色的)
and d0 (蓝色的) are overlaid onto an inflated cortical surface of the left
hemisphere (顶部). The overlap between activations in or near the AG
0 voxelwise correlation (蓝色的), 和
from the retrieval success (红色的), d
analyses is shown in the bottom. Areas of overlap are in violet.
所以, the following discussion is aimed at placing our
results in the context of prior findings and considering the
possibility of reinterpreting prior results.
Memory Retrieval and Left AG
The left parietal cortex, including AG, is frequently re-
ported in fMRI studies of retrieval success (Simons et al.,
2008). Activity in some parts of AG tends to decrease be-
low baseline during performance of goal-directed tasks
suggesting it is part of the “default mode” network. 使用
functional connectivity density mapping, Tomasi and
Volkow (2011) found that bilateral AG belonged to a de-
fault mode network that included the parahippocampal
gyrus and medial parietal and frontal cortex.
Analyses of spontaneous low-frequency fluctuations
in BOLD fMRI data demonstrate a link between AG and
medial-temporal and parietal structures. Nelson and col-
联赛 (2010) used resting state functional connectiv-
ity analyses and graph theoretical tools to define a grid
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
哦
8
C
6
n
5
_
7
A
/
_
j
0
哦
0
C
2
n
9
2
_
A
p
_
d
0
0
乙
2
y
9
G
2
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
t
F
.
/
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
桌子 5. List of ROIs with Positive Correlations between
RSE and C
不. Hem Anatomic Location
X
y
z
∼BA #Vox
1
2
3
4
5
6
7
8
9
10
11
12
13
14
15
16
17
18
19
20
21
22
23
L
右
L
L
右
右
L
L
L
L
L
右
右
L
右
右
L
L
L
L
右
L
右
中. Frontal G.
Inf. Parietal Lobe
Postcentral G.
Postcentral G.
中. Frontal G.
中. Frontal G.
Postcentral G.
Precentral G.
小脑
−34 −00
48 −62
−47 −26
−52 −22
48
46
23
11
−31 −21
−40 −09
51
43
38
51
31
37
37
55
6
40
2
2
9
8
1
6
−17 −47 −37 NA
Postcentral G.
−21 −39
62
5
小脑
−03 −42 −35 NA
Inf. Temporal G.
40 −05 −40
20
小脑
小脑
小脑
33 −55 −34 NA
−33 −41 −42 NA
26 −41 −32 NA
中. Frontal G.
20 −02
52
6
Red Nucleus
−08 −22 −04 NA
Precentral G.
−35 −05
37
6
Pulvinar
Precuneus
小脑
−13 −27
06 NA
−12 −62
25
31
32 −38 −43 NA
中. Temporal G. −47
01 −30
Fusiform G.
26 −91 −13
21
18
287
143
162
159
101
135
116
118
141
204
129
113
107
114
146
115
101
106
155
119
108
157
158
Hem = hemisphere; L = left; R = right; Inf = inferior; G = gyrus;
Mid = middle; BA = Brodmannʼs area; #vox = number of voxels in
ROI; all locations and Brodmannʼs areas are approximate.
for on-line maintenance of retrieved information. 相似地,
Guerin and Miller (2011) found a more ventral left parietal/
middle temporal region (−44, −64, 22 in MNI-to-Talairach
coordinates) that decreased activity less as more informa-
tion was retrieved. Although unilateral lesions to the lateral
parietal do not affect source recollection of word labels and
faces (Simons et al., 2008), there is evidence that bilateral
lesions are associated with reduced perceptual detail in
recollection (Berryhill, Phuong, Picasso, Cabeza, & 奥尔森,
2007). This inconsistency between lesion and fMRI studies
deserves further attention. Collectively these studies have
been used to conclude that AG is associated with recol-
lection, and the association with the amount of content
precluded a role in memory decisions. 然而, 在下面
an accumulator hypothesis, the amount of content may
very well play an important role: The better the evidence
for the memory decision, the faster the information ac-
cumulates toward a decision and the faster AG activity
变化. 换句话说, the accumulator hypothesis
provides an alternative explanation attributing the role of
AG in recollection tasks to high evidence situations.
It has also been proposed that parietal regions play
a role in attention to memory. Proposals include the
Attention-to-Memory (AtoM; Cabeza, Ciaramelli, 奥尔森, &
Moscovitch, 2008; Ciaramelli, Grady, & Moscovitch, 2008)
and the Dual-Attentional Processes (Cabeza, 2008) hy-
potheses. In these accounts, ventral parietal regions
mediate reflexive attentional capture of relevant retrieved
information whereas dorsal parietal regions are involved
in top–down control. In the AtoM framework, the AG is
included in the reflexive ventral system and should be
more active during target and foil decisions made with
high than low confidence because the former items are
more salient (Cabeza, 2008). To test whether our findings
in AG were consistent with AtoM, we conducted a 2 ×
2 ANOVA on the peak estimates from “sure” and “maybe”
targets and foils (collapsed over list strength) 在
18 participants with sufficient data. This analysis identified
a main effect of Confidence (sure > maybe; F(1, 17) =
13.87, p < .001) and, as expected due to the region
definition procedure, a main effect of Item Type (target >
foil) with no interaction ( p = .31). 因此, consistent with
AtoM predictions, activity was greater on “sure” than
“maybe” trials for both targets and foils. 然而, 这
is also consistent with accumulator activity as described
下一个.
AG Demonstrates Accumulator-like Activity
during Episodic Memory
如上所述, the AG region displayed differential
strength-based effects early in the time course for foils,
suggesting that the momentary signal in this region may
be related to the impending memory decision. This find-
ing is similar to the patterns of accumulating activity in a
perceptual identification task where participants identify
an item that is masked and slowly revealed (Ploran,
Tremel, 纳尔逊, & Wheeler, 2011; Wheeler, 彼得森,
纳尔逊, Ploran, & Velanova, 2008; Ploran et al., 2007). 在
those studies, the onset of activity in left parietal cortex
ROIs occurred early in the trial and increased at a rate
that correlated with the time of identification: Activity
increased faster when identification occurred earlier in
the trial. The observed patterns of data may reflect an
integration-to-bound mechanism (金子 & 沙德伦, 2007;
Hanes & 沙尔, 1996) because the information processed
was relevant to the decision. That parietal damage does
not cause major disruption to memory suggests that the
AG is not the sole source of evidence. We note that Ploran
等人. (2007) also reported “accumulator-like” regions that
decreased rather than increased activity relative to base-
线 (Figure 3B from that manuscript). 因此, the absolute
direction of activity (negative in the AG region reported
432
认知神经科学杂志
体积 25, 数字 3
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
F
/
t
t
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
哦
8
C
6
n
5
_
7
A
/
_
j
0
哦
0
C
2
n
9
2
_
A
p
_
d
0
0
乙
2
y
9
G
2
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
/
t
F
.
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
这里) may be less important than the pattern of accumula-
的. We also note that their peak voxels (Ploran et al.,
2007: −26 −68 38; Ploran et al., 2011: −24, −57, 45
and −24, −71, 34) were ∼15 mm closer to the midline
of the cerebral hemispheres than the peak voxel in our
AG (AG: −42, −70, 37).
在我们的数据中, left AG is activated more for correct target
than foil trials and more for “sure” than “maybe” trials. 这
U-shaped function is consistent with findings from sev-
eral other studies (IE。, Daselaar, Fleck, & Cabeza, 2006;
Yonelinas et al., 2005). This is the expected pattern of data
obtained when considering SBME data with the diffusion
model framework and is consistent with some models of
evidence accumulation in macaque LIP (例如, Mazurek,
Roitman, Ditterich, & 沙德伦, 2003): Items receive a
high-confidence response because they provide high-
quality evidence, which has a faster rate of accumulation.
As an alternative to the notion that left AG should be
viewed as an evidence accumulator region, we also con-
sidered the possibility that left AG activation might cor-
respond to the total memory strength produced by the
刺激. This is consistent with a number of features
数据的, including that this is a RSAd0. This possibility
is also consistent with the finding of Rissman, Greely,
and Wagner (2010) that activity in left AG (除其他外
地区) is predictive of subjective memory for items. 和-
der this view, we would expect less activation in AG from
strong-list foil CRs than from weak-list foil CRs (as illus-
trated in Figure 1, bottom panel), and such a trend is
visually apparent after onset of the memory probe (时间
Points 5 和 6, Figure 3D). 然而, this finding was not
statistically reliable at our estimated peak time of Time
Points 4, 5 和 6, and more confident CRs were asso-
ciated with greater, not less activation that less confident
CRs, a finding more consistent with the evidence accu-
mulator hypothesis. The picture presented by the full
pattern is tantalizing in suggesting possible roles for both
evidence accumulation and total memory activation in
the same brain area. We look forward to further investi-
gations that may help clarify the possibly complex role of
left AG in recognition memory.
Response Bias in Anterior and Superior
Parietal Lobe
Our response bias analyses found parietal regions that
were located anterior and superior to the AG regions track-
ing retrieval success. These included a region (数字 5)
taken from a recent study (OʼConnor et al., 2010) reporting
0) in parietal cortex
a significant correlation with c (but not d
and regions defined in a voxelwise analysis correlating c
with RSE on a subject-by-subject basis (Figure 6A). 一些-
what strikingly, the correlation between c and RSE in our
estimate of the OʼConnor ROI matched very well with the
correlation in their ROI, indicating that the finding is reli-
able across independent studies. Plotting peak coordinates
from those studies revealed a section of left lateral parietal
cortex near the AG and supramarginal gyrus that is fre-
quently associated with memory operations (Hit > CR)
but rarely reported in studies of attention. Findings from
the current study are thus consistent with the claim by
OʼConnor et al. (2010) that anterior and superior regions
of the left parietal cortex are related to strategic attention,
but inconsistent in that lateral regions were found to be
associated with operations pertaining to the memory de-
切除术. OʼConnor et al. reported no ROIs where the RSE
0 may be
correlated with d
related to differences in task or region selection (例如, 这
0 and c correlations were conducted on an unpublished
d
数据集, not the experiment manipulating cue validity
reported in their article).
0. The discrepant findings for d
Our findings are consistent with data reported by
Aminoff et al. (2011) and Herron et al. (2004), WHO
manipulated response bias by varying the percent of tar-
gets in the test block. In both of these studies and in our
数据, there is considerable overlap with RSA and regions
modulated by response bias. As suggested by OʼConnor
等人。, the term “retrieval success” is perhaps a misnomer
as these regions reflect the contribution of both memory
retrieval and response bias. Another common finding
across these studies is a lack of response bias-dependent
activation for left AG. Herron et al. report an RSE but no
interaction with response bias for a region near left AG
(MNI coordinates −33, −72, 30) and Aminoff et al. 做
not report a contribution of AG in a regression analysis
accounting for their data. 最后, all three studies report
many regions of parietal cortex that are modulated by
response bias.
Summary
We found evidence suggesting a differentiation account
of the SBME, specifically the AG may be accumulating
or maintaining accumulated evidence. 此外, 我们
found regions whose retrieval success activity correlates
0 and with response bias with few such areas that
with d
重叠. 然而, theories of parietal function in memory
are diverse (OʼConnor et al., 2010; Cabeza, 2008; Ciaramelli
等人。, 2008; Vilberg & Rugg, 2008; Wagner et al., 2005;
Wheeler & 巴克纳, 2003), and a definitive statement of
the role of the AG in memory awaits further research.
致谢
Modeling and other data analysis was supported by the National
Science Foundation (0951612 to A. H. C。). This research was
also supported by the National Institute of Mental Health
(R01-MH086492 to M. 乙. 瓦. and P50-MH64445 to J. L. M。).
We thank Sarah Woo for assistance with data collection and
加工.
Reprint requests should be sent to Amy H. Criss, Department of
心理学, Syracuse University, 477 Huntington Hall, 锡拉丘兹,
纽约 13244, or via e-mail: acriss@syr.edu.
Criss, Wheeler, and McClelland
433
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
哦
8
C
6
n
5
_
7
A
/
_
j
0
哦
0
C
2
n
9
2
_
A
p
_
d
0
0
乙
2
y
9
G
2
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
t
/
.
F
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
参考
Aminoff, E., 弗里曼, S。, Clewett, D ., Tipper, C。, Frithsen, A。,
约翰逊, A。, 等人. (2011). Neural correlates of criterion
shifting in memory. Poster presented at the Cognitive
Neuroscience Meeting, 旧金山, CA.
Balota, D. A。, Yap, 中号. J。, Cortese, 中号. J。, Hutchinson, K. A。,
Kessler, B., Neely, J. H。, 等人. (2007). The English lexicon
项目. Behavior Research Methods, 39, 445–449.
Berryhill, 中号. E., Phuong, L。, Picasso, L。, Cabeza, R。, & 奥尔森,
我. 右. (2007). Parietal lobe and episodic memory: Bilateral
damage causes impaired free recall of autobiographical
回忆. 神经科学杂志, 27, 14415–14423.
巴克纳, 右. L。, & Wheeler, 中号. 乙. (2001). The cognitive
neuroscience of remembering. 自然评论
神经科学, 2, 624–634.
Buracas, G. T。, & Boynton, G. 中号. (2002). Efficient design
of event-related fMRI experiments using M-sequences.
Neuroimage, 16, 801–813.
Cabeza, 右. (2008). Role of parietal regions in episodic
memory retrieval: The dual attentional processes
假设. Neuropsychologia, 46, 1813–1827.
Cabeza, R。, Ciaramelli, E., 奥尔森, 我. R。, & Moscovitch, 中号.
(2008). The parietal cortex and episodic memory: 一个
attentional account. 自然评论神经科学, 9,
613–625.
Cary, M。, & Reder, L. 中号. (2003). A dual-process account of the
list-length and strength-based mirror effects in recognition.
记忆与语言杂志, 49, 231–248.
Ciaramelli, E., Grady, C. L。, & Moscovitch, 中号. (2008). Top–down
and bottom–up attention to memory: A hypothesis (AtoM)
on the role of the posterior parietal cortex in memory
恢复. Neuropsychologia, 46, 1828–1851.
Guerin, S. A。, & 磨坊主, 中号. 乙. (2011). Parietal cortex tracks
the amount of information retrieved even when it is
not the basis of a memory decision. Neuroimage, 55,
801–807.
Hanes, D. P。, & 沙尔, J. D. (1996). Neural control of voluntary
movement initiation. 科学, 274, 427–430.
Henson, 右. N。, Rugg, 中号. D ., Shallice, T。, Josephs, 奥。, &
Dolan, 右. J. (1999). Recollection and familiarity in recognition
记忆: An event-related functional magnetic resonance
imaging study. 神经科学杂志, 19, 3962–3972.
Herron, J. E., Henson, 右. N。, & Rugg, 中号. D. (2004). 可能性
effects on the neural correlates of retrieval success:
An fMRI study. Neuroimage, 21, 302–310.
Ho, 时间. C。, 棕色的, S。, & Serences, J. 时间. (2009). Domain general
mechanisms of perceptual decision making in human
cortex. 神经科学杂志, 29, 8675–8687.
Kahn, 我。, Andrews-Hanna, J. R。, Vincent, J. L。, 斯奈德, A. Z。,
& 巴克纳, 右. L. (2008). Distinct cortical anatomy linked
to subregions of the medial temporal lobe revealed
by intrinsic functional connectivity. 杂志
Neurophysiology, 100, 129–139.
Kahn, 我。, Davachi, L。, & 瓦格纳, A. D. (2004). Functional-
neuroanatomic correlates of recollection: Implications for
models of recognition memory. 神经科学杂志,
24, 4172–4180.
Konishi, S。, Wheeler, 中号. E., Donaldson, D. 我。, & 巴克纳,
右. L. (2000). Neural correlates of episodic retrieval success.
Neuroimage, 12, 276–286.
Kwong, K. K., 贝利沃, J. W., Chesler, D. A。, Goldberg, 我. E.,
Weisskoff, 右. M。, Poncelet, 乙. P。, 等人. (1992). 动态的
magnetic resonance imaging of human brain activity during
primary sensory stimulation. 国家会议录
Academy of Sciences, 美国。, 89, 5675–5679.
Criss, A. H. (2006). The consequences of differentiation in
兰卡斯特, J. L。, Glass, 时间. G。, Lankipalli, 乙. R。, Downs, H。,
episodic memory: Similarity and the strength based mirror
影响. 记忆与语言杂志, 55, 461–478.
Criss, A. H. (2009). The distribution of subjective memory
strength: List strength and response bias. 认知的
心理学, 59, 297–319.
Criss, A. H. (2010). Differentiation and response bias in
episodic memory: Evidence from reaction time distributions.
实验心理学杂志: 学习, 记忆,
和认知, 36, 484–499.
Criss, A. H。, & 麦克莱兰, J. L. (2006). Differentiating the
differentiation models: A comparison of the retrieving
effectively from memory model (REM) and the subjective
likelihood model (SLiM). Journal of Memory and
语言, 55, 447–460.
Daselaar, S. M。, Fleck, 中号. S。, & Cabeza, 右. (2006). Triple
dissociation in the medial temporal lobes: Recollection,
familiarity, and novelty. 神经生理学杂志, 96,
1902–1911.
Dobbins, 我. G。, Foley, H。, Schacter, D. L。, & 瓦格纳, A. D.
(2002). Executive control during episodic retrieval: 多种的
prefrontal processes subserve source memory. 神经元,
35, 989–996.
狐狸, 中号. D ., 斯奈德, A. Z。, Barch, D. M。, Gusnard, D. A。,
& Raichle, 中号. 乙. (2005). Transient BOLD responses at
block transitions. Neuroimage, 28, 956–966.
弗里斯顿, K., Jezzard, P。, & 车工, 右. (1994). Analysis of
functional MRI time-series. 人脑图谱,
1, 153–171.
金子, J. 我。, & 沙德伦, 中号. 氮. (2007). The neural basis of
decision making. Annual Review of Neuroscience, 30,
535–574.
Mayberg, H。, & 狐狸, 磷. 时间. (1995). A modality-independent
approach to spatial normalization of tomographic
images of the human brain. 人脑图谱, 3,
209–223.
Malmberg, K. J. (2008). Recognition memory: A review of the
critical findings and an integrated theory for relating them.
认知心理学, 57, 335–384.
Mazurek, 中号. E., Roitman, J. D ., Ditterich, J。, & 沙德伦, 中号. 氮.
(2003). A role for neural integrators in perceptual decision
制作. 大脑皮层, 13, 1257–1269.
McAvoy, 中号. P。, Ollinger, J. M。, & 巴克纳, 右. L. (2001). Cluster
size thresholds for assessment of significant activation in
功能磁共振成像. Neuroimage, 13, S198.
麦克莱兰, J. L。, & Chappell, 中号. (1998). Familiarity breeds
differentiation: A subjective-likelihood approach to the
effects of experience in recognition memory. Psychological
审查, 105, 724–760.
麦克德莫特, K. B., 琼斯, 时间. C。, 彼得森, S. E., Lageman, S. K.,
& Roediger, H. L。, 三、 (2000). Retrieval success is accompanied
by enhanced activation in anterior prefrontal cortex during
recognition memory: An event-related fMRI study. 杂志
of Cognitive Neuroscience, 12, 965–976.
Michelon, P。, 斯奈德, A. Z。, 巴克纳, 右. L。, McAvoy, M。, &
Zacks, J. 中号. (2003). Neural correlates of incongruous visual
信息. An event-related fMRI study. Neuroimage,
19, 1612–1626.
米津, F. M。, Maccotta, L。, Ollinger, J. M。, 彼得森, S. E., &
巴克纳, 右. L. (2000). Characterizing the hemodynamic
response: Effects of presentation rate, sampling procedure,
and the possibility of ordering brain activity based on
relative timing. Neuroimage, 11, 735–759.
烧烤-斯佩克特, K., Henson, R。, & 马丁, A. (2006). Repetition
and the brain: Neural models of stimulus-specific effects.
认知科学的趋势, 10, 14–23.
Montaldi, D ., Spencer, 时间. J。, 罗伯茨, N。, & Mayes, A. 右.
(2006). The neural system that mediates familiarity
记忆. Hippocampus, 16, 504–520.
434
认知神经科学杂志
体积 25, 数字 3
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
哦
8
C
6
n
5
_
7
A
/
_
j
0
哦
0
C
2
n
9
2
_
A
p
_
d
0
0
乙
2
y
9
G
2
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
F
/
.
t
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1
纳尔逊, S. M。, 科恩, A. L。, 力量, J. D ., 假发, G. S。, 米津,
F. M。, Wheeler, 中号. E., 等人. (2010). A parcellation scheme
for human left lateral parietal cortex. 神经元, 67,
156–170.
Norman, K. A。, & 奥莱利, 右. C. (2003). Modeling hippocampal
and neocortical contributions to recognition memory:
A complementary learning systems approach.
心理评论, 110, 611–646.
OʼConnor, A. R。, Han, S。, & Dobbins, 我. G. (2010). The inferior
parietal lobule and recognition memory: Expectancy violation
or successful retrieval? 神经科学杂志, 30,
2924–2934.
Ogawa, S。, Tank, D. W., Menon, R。, Ellerman, J. M。, Kim,
S. G。, Merkle, H。, 等人. (1992). Intrinsic signal changes
accompanying sensory stimulation: Functional brain mapping
with magnetic resonance imaging. 诉讼程序
美国国家科学院, 美国。, 89, 5951–5955.
Ojemann, J. G。, Akbudak, E., 斯奈德, A. Z。, McKinstry, 右. C。,
Raichle, 中号. E., & Conturo, 时间. 乙. (1997). Anatomic localization
and quantitative analysis of gradient refocused echo-planar
fMRI susceptibility artifacts. Neuroimage, 6, 156–167.
Ollinger, J. M。, 舒尔曼, G. L。, & 科尔贝塔, 中号. (2001).
Separating processes within a trial in event-related
functional MRI I. 方法. Neuroimage, 13, 210–217.
Ploran, 乙. J。, 纳尔逊, S. M。, Velanova, K., Donaldson, D. 我。,
彼得森, S. E., & Wheeler, 中号. 乙. (2007). 证据
accumulation and the moment of recognition: Dissociating
perceptual recognition processes using fMRI. 杂志
神经科学, 27, 11912–11924.
Ploran, 乙. J。, Tremel, J. J。, 纳尔逊, S. M。, & Wheeler, 中号. 乙.
(2011). High quality but limited quantity perceptual evidence
produces neural accumulation in frontal and parietal
cortex. 大脑皮层, 21, 2650–2662.
拉特克利夫, 右. (1978). A theory of memory retrieval. Psychological
审查, 85, 59–108.
Rissman, J。, Greely, H. T。, & 瓦格纳, A. D. (2010). Detecting
individual memories through the neural decoding of memory
states and past experience. 国家会议录
Academy of Sciences, 美国。, 107, 9849–9854.
沙尔, J. D. (2001). 决策的神经基础, choosing and
acting. 自然评论神经科学, 2, 33–42.
Shiffrin, 右. M。, 拉特克利夫, R。, & 克拉克, S. 乙. (1990). List-strength
影响: 二. Theoretical mechanisms. 实验杂志
心理学: 学习, 记忆, 和认知, 16, 179–195.
Shiffrin, 右. M。, & Steyvers, 中号. (1997). A model for recognition
记忆: REM-retrieving effectively from memory.
Psychonomic Bulletin & 审查, 4, 145–166.
Simons, J. S。, Peers, 磷. 五、, Hwang, D. Y。, Ally, 乙. A。, 弗莱彻, 磷. C。,
& Budson, A. 乙. (2008). Is the parietal lobe necessary for
recollection in humans? Neuropsychologia, 46, 1185–1191.
斯奈德, A. Z. (1996). Difference image versus ratio image
error function forms in PET-PET realignment. 在D中. 贝利
& 时间. 琼斯 (编辑。), Quantification of brain function using
PET (PP. 131–137). 圣地亚哥, CA: 学术出版社.
Starns, J. J。, 白色的, C. N。, & 拉特克利夫, 右. (2010). A direct test of
the differentiation mechanism: REM, BDCMEM, 和
strength-based mirror effect in recognition memory.
记忆与语言杂志, 63, 18–34.
Stretch, 五、, & Wixted, J. 时间. (1998). On the difference between
strength-based and frequency-based mirror effects in
recognition memory. 实验心理学杂志:
学习, 记忆, 和认知, 24, 1379–1396.
Talairach, J。, & Tournoux, 磷. (1988). Co-planar stereotaxic atlas
of the human brain. Thieme, 纽约.
Tomasi, D ., & Volkow, 氮. D. (2011). Association between
functional connectivity hubs and brain networks. Cerebral
Cortex, 21, 2003–2013.
Van Essen, D. C. (2005). A population-average, landmark-
and surface-based (PALS) atlas of human cerebral cortex.
Neuroimage, 28, 635–662.
Van Essen, D. C。, Dickson, J。, Harwell, J。, Hanlon, D ., 安德森,
C. H。, & Drury, H. A. (2001). An integrated software suite
for surface-based analyses of cerebral cortex. 杂志
the American Medical Informatics Association, 41,
1359–1378. See also brainmap.wustl.edu/caret.
Vilberg, K. L。, & Rugg, 中号. D. (2007). Dissociation of the neural
correlates of recognition memory according to familiarity,
recollection, and amount of recollected information.
Neuropsychologia, 45, 2216–2225.
Vilberg, K. L。, & Rugg, 中号. D. (2008). Memory retrieval and the
parietal cortex: A review of evidence from a dual-process
看法. Neuropsychologia, 46, 1787–1799.
Vincent, J. L。, 斯奈德, A. Z。, 狐狸, 中号. D ., Shannon, 乙. J。, Andrews,
J. R。, Raichle, 中号. E., 等人. (2006). Coherent spontaneous
activity identifies a hippocampal-parietal memory network.
神经生理学杂志, 96, 3517–3531.
瓦格纳, A。, Shannon, B., Kahn, 我。, & 巴克纳, 右. L. (2005).
Parietal lobe contributions to episodic memory retrieval.
认知科学的趋势, 9, 445–453.
Wheeler, 中号. E., & 巴克纳, 右. L. (2003). Functional dissociation
among components of remembering: 控制, perceived
oldness, and content. 神经科学杂志, 23,
3869–3880.
Wheeler, 中号. E., & 巴克纳, 右. L. (2004). Functional-anatomic
correlates of remembering and knowing. Neuroimage, 21,
1337–1349.
Wheeler, 中号. E., 彼得森, S. E., 纳尔逊, S. M。, Ploran, 乙. J。,
& Velanova, K. (2008). Dissociating early and late error
signals in perceptual recognition. 认知杂志
神经科学, 12, 2211–2225.
Yonelinas, A. P。, Otten, L. J。, Shaw, K. N。, & Rugg, 中号. D. (2005).
Separating the brain regions involved in recollection and
familiarity in recognition memory. 神经科学杂志,
25, 3002–3008.
Criss, Wheeler, and McClelland
435
D
哦
w
n
我
哦
A
d
e
d
F
r
哦
米
我
我
/
/
/
/
j
t
t
F
/
我
t
.
:
/
/
H
t
t
p
:
/
D
/
哦
米
w
我
n
t
哦
p
A
r
d
C
e
.
d
s
F
我
r
哦
我
米
v
e
H
r
C
p
H
A
d
我
我
r
r
e
.
C
C
t
.
哦
米
米
/
j
e
d
哦
你
C
n
哦
/
C
A
n
r
A
t
r
我
t
我
C
C
我
e
e
–
p
–
d
p
d
2
F
5
/
3
2
5
4
/
2
3
1
/
1
4
9
2
4
1
4
/
9
1
4
7
9
7
哦
8
C
6
n
5
_
7
A
/
_
j
0
哦
0
C
2
n
9
2
_
A
p
_
d
0
0
乙
2
y
9
G
2
你
.
e
p
s
t
d
哦
F
n
乙
0
y
8
S
中号
e
我
p
时间
e
米
L
我
乙
乙
e
r
r
A
2
r
0
我
2
3
e
s
/
j
.
t
/
F
你
s
e
r
哦
n
1
7
中号
A
y
2
0
2
1