Uso de electroencefalografía de alta densidad para explorar
Spatiotemporal Representations of Object Categories
in Visual Cortex
Gennadiy Gurariy1, Ryan E. B. Mruczek2, Jacqueline C. Snow3, and Gideon P. Caplovitz3
Abstracto
■ Visual object perception involves neural processes that
unfold over time and recruit multiple regions of the brain. Aquí,
we use high-density EEG to investigate the spatiotemporal rep-
resentations of object categories across the dorsal and ventral
pathways. In Experiment 1, human participants were presented
with images from two animate object categories (birds and
insects) and two inanimate categories (tools and graspable
objects). In Experiment 2, participants viewed images of tools
and graspable objects from a different stimulus set, one in which
a shape confound that often exists between these categories
(elongation) was controlled for. To explore the temporal dynam-
ics of object representations, we employed time-resolved multi-
variate pattern analysis on the EEG time series data. This was
performed at the electrode level as well as in source space of
two regions of interest: one encompassing the ventral pathway
and another encompassing the dorsal pathway. Nuestros resultados
demonstrate shape, exemplar, and category information can
be decoded from the EEG signal. Multivariate pattern analysis
within source space revealed that both dorsal and ventral path-
ways contain information pertaining to shape, inanimate object
categories, and animate object categories. Of particular interest,
we note striking similarities obtained in both ventral stream and
dorsal stream regions of interest. These findings provide insight
into the spatio-temporal dynamics of object representation and
contribute to a growing literature that has begun to redefine the
traditional role of the dorsal pathway. ■
INTRODUCCIÓN
In this article, we describe the results of two experiments
in which we applied high-density EEG (HD-EEG) a
explore the spatio-temporal representations of visual
objects in the human brain. The successful perception of
visual objects poses serious computational challenges
given that the signals traversing the human retina are
ambiguous, noisy, and in constant flux. Despite these
obstacles, visual perception is a rapid and efficient pro-
impuesto. Por ejemplo, discernment of different object catego-
ries can happen in as little as 120 mseg (Kirchner &
Thorpe, 2006), whereas neural activity corresponding to
some object categories can be detected after only 80 mseg
(Crouzet, Kirchner, & Thorpe, 2010). The neural activity
associated with object perception is not static but rather
evolves over time (Contini, Wardle, & Carlson, 2017) y
has been studied in terms of various event-related
componentes (Bentin et al., 1996) that can be measured
with magnetoencephalography/electroencephalography
(M/EEG) técnicas.
The cascade of neural activity evoked by object percep-
tion is thought to progress across multiple visual regions
throughout the brain. Proceeding from V1 to the inferotem-
poral cortex, neuronal populations display increasingly
1Medical College of Wisconsin, 2College of the Holy Cross, 3Uni-
versity of Nevada, Reno
© 2022 Instituto de Tecnología de Massachusetts
larger receptive fields and increasingly complex tuning
propiedades, culminating in regions that appear to be
category selective (Booth & Rolls, 1998; Desimone,
Albright, Bruto, & bruce, 1984; Bruto, Rocha-Miranda, &
Bender, 1972). fMRI studies have also uncovered object-
selective responses in the human ventral temporal cortex
(Malach et al., 1995) as well as selectivity toward specific
object categories including faces (Ishai, Ungerleider, Martín,
Schouten, & Haxby, 1999; Kanwisher, McDermott, &
Chun, 1997), lugares (Epstein, harris, Stanley, & Kanwisher,
1999; Epstein & Kanwisher, 1998), bodies (Downing, Jiang,
Shuman, & Kanwisher, 2001), and words (cohen &
Dehaene, 2004). Neural regions displaying selectivity
toward broader, superordinate visual categories, como
animacy (Grill-Spector & Weiner, 2014; Martín, 2007), tener
also been reported. Además, many of these areas exhibit
invariance to low-level manipulations such as size, visita
perspectiva, and contour definition (Sawamura, Georgieva,
Vogels, Vanduffel, & Orban, 2005; Vuilleumier, Henson,
Conductor, & Dolan, 2002; Kourtzi & Kanwisher, 2000; Grill-
Spector et al., 1998).
Classically, studies of visual object perception have
focused on the ventral pathway, a network of neural
regions thought to generate invariant representations of
the world for the purposes of recognition. Sin embargo, a
growing body of empirical results suggests that the dorsal
pathway (thought to be involved in spatial knowledge and
Revista de neurociencia cognitiva 34:6, páginas. 967–987
https://doi.org/10.1162/jocn_a_01845
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
visuomotor interaction) may also contribute to this pro-
impuesto. This conclusion is corroborated by findings demon-
strating that the dorsal pathway can process shape as well
as certain object categories (collins, Freud, Kainerstorfer,
Cao, & behrman, 2019; Chen, Snow, Culham, & Goodale,
2018; Erlikhman, Gurariy, Mruczek, & Caplovitz, 2016;
Zachariou, Klatzky, & behrman, 2014; Mruczek, von
Loga, & Kastner, 2013; Almeida, Mahon, & Caramazza,
2010; Almeida, Mahon, Nakayama, & Caramazza, 2008;
Konen & Kastner, 2008; Valyear, Cavina-Pratesi, Stiglick,
& Culham, 2007; Weisberg, Van Turennout, & Martín,
2007; Fang & Él, 2005; chao & Martín, 2000). Graspable
objects constitute one category of stimuli that has been
shown to drive activation in the dorsal pathway. Tools
are a special subset of graspable objects characterized
by a specific associated motor plan learned through
experiencia (Frey, 2007) and have been shown to activate
regions in the ventral as well as the dorsal pathway
(Garcea & Mahon, 2014; Mruczek et al., 2013; Hermsdörfer,
Terlinden, Mühlau, Goldenberg, & Wohlschläger, 2007;
chao & Martín, 2000; Chao et al., 1999).
Some have questioned whether the aforementioned
findings truly demonstrate neural tuning toward abstract
categories, arguing that studies on object perception are
often confounded by features that covary within a particu-
lar object class. Por ejemplo, category members often
share similarities across a number of dimensions such as
real-world size (Konkle & Oliva, 2012), manipulability
(Mahon et al., 2007), the potential for self-initiated behav-
ior (Martín & Weisberg, 2003), and numerous low-level
image properties (Andrews, watson, Rice, & Hartley,
2015; Rice, watson, Hartley, & Andrews, 2014; watson,
Hartley, & Andrews, 2014; Baldassi et al., 2013; O’Toole,
Jiang, Abdi, & Haxby, 2005). Multivariate analyses of
neural data offer greater sensitivity to subtle differences
in patterns of activation (Kriegeskorte, 2011; Kriegeskorte,
Goebel, & Bandettini, 2006; Lange et al., 1999) and have
further challenged the notion of clearly delineated,
object-selective regions. Por ejemplo, Haxby et al.
(2001) showed that category representations may be
overlapping and distributed throughout ventral temporal
corteza, as opposed to being localized within functionally
homogenous neural modules. Similarmente, some have
questioned studies purporting to show object selectivity
in the dorsal pathway, suggesting that shape rather than
categorical membership might explain these results. Para
ejemplo, tools are more likely to be elongated along the
principal axis relative to members of other nontool catego-
ries. Además, many studies that have found dorsal
activation to tools have failed to control for elongation
(Chen et al., 2018; Sakuraba, Sakai, Yamanaka, Yokosawa,
& Hirayama, 2012), leading to the possibility that shape,
rather than category membership, best describes tool-
evoked activity in the dorsal pathway (Almeida et al.,
2014; Sakuraba et al., 2012; Sakata et al., 1998). Juntos,
such findings complicate theoretical interpretations of
previous research and underscore the importance of
adequate controls and consilience across different
methodologies.
en este estudio, we explored the dynamics of object per-
ception using HD-EEG with a particular focus on the neu-
ral representation of visual objects in the dorsal and ventral
pathways as well as the contribution of shape and category
to these representations. We define representations as
informational neural states that may subserve numerous
processes such as recognition, visuomotor interactions,
Etcétera. En tono rimbombante, we extended our analysis across
both space and time by conducting multivariate pattern
análisis (MVPA) on source-localized dipoles within
broadly defined dorsal and ventral ROIs. In the first exper-
mento, participants viewed images of objects from differ-
ent categories. Two superordinate categories (animate
and inanimate) were further subdivided into four catego-
ries (bird, insect, tool, and graspable object). These object
categories were chosen based on their relationship to the
known computational properties of the dorsal and ventral
pathways. Específicamente, regions along the ventral pathway
have been shown to process object identity and category
membership for both animate and inanimate object cate-
gories (macdonald & Culham, 2015; Garcea & Mahon,
2014; Kravitz, Saleem, Panadero, Ungerleider, & Mishkin,
2013; Mahon et al., 2007). Mientras tanto, regions along the
dorsal pathway exhibit sensitivity to particular object cate-
gories based on the presence of affordances and/or spe-
cific motor plans in the case of tools (Luis, 2006; Fang
& Él, 2005; chao & Martín, 2000). It is important to note
that the inanimate stimuli (tools and graspable objects)
used in Experiment 1 contained the shape confound
(elongation along the principal axis for tools) commonly
found in previous studies. De este modo, a follow-up experiment
(Experimento 2) was conducted in which participants
viewed images of tools and graspable objects that were
matched for global shape (elongated vs. stubby), permitiendo
shape-related signals to be disambiguated from category-
related responses.
Juntos, these experiments allowed us to explore the
extent to which spatio-temporal responses from neural
populations in dorsal and ventral cortex vary according
to global shape characteristics versus higher-level object
categoría. On the basis of the preponderance of previous
investigación, we expected that activity within the ventral path-
way should contain information pertaining to both shape
and category membership. With respect to the dorsal
pathway, we expected above-chance classification for
tools versus graspable objects using stimuli from Experi-
mento 1. Además, if the computations along the dorsal
pathway extract category information in a manner analo-
gous to the canonical properties of the ventral pathway,
then we would also expect above-chance classification
for birds versus insects. Además, the temporal reso-
lution afforded by EEG allows for additional inferences
regarding the nature of the computations along the two
pathways. Both the onset and peak of time-resolved sig-
nals provide information regarding the nature of neural
968
Revista de neurociencia cognitiva
Volumen 34, Número 6
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Procesando. Por ejemplo, comparing these metrics across
conditions has been used to make inferences regarding
the hierarchy of object recognition (es decir., the order of
visual processing stages; Contini et al., 2017; Carlson,
Tovar, Alink, & Kriegeskorte, 2013) and to test hypotheses
regarding feedback and feedforward modulation between
regiones del cerebro (Martin et al., 2019). In the context of the
estudio actual, comparing the temporal dynamics between
the dorsal and ventral pathways can yield insights regard-
ing the relationship and computational dependencies
between these networks. Por ejemplo, category informa-
tion within the dorsal pathway could either be computed
independently or arise as a product of feedforward
input from the ventral pathway (Takemura et al., 2016;
Cloutman, 2013). De este modo, observing a latency offset whereby
significant classification of object categories in the ventral
pathway precedes significant classification in the dorsal
pathway would be consistent with a model in which dorsal
object representations are partially dependent on feedfor-
ward ventral projections. En cambio, similar temporal
dynamics across dorsal and ventral ROIs would be consis-
tent with a model in which object representations are com-
puted independently across the two pathways (Freud,
Plaut, & behrman, 2016).
Aquí, we leverage HD-EEG, source localization, y
MVPA to explore the spatio-temporal dynamics of object
processing across the dorsal and ventral pathways. Specif-
icamente, this study addresses the following questions: (a)
when does category-specific information arise within the
EEG signal, (b) how are object categories represented
across dorsal and ventral neural pathways, y (C) to what
extent does category, as opposed to shape, contribute to
the above representations?
EXPERIMENT 1
The purpose of Experiment 1 was to explore the spatio-
temporal dynamics of object processing using stimuli
belonging to animate (bird and insect) and inanimate (tool
and graspable object) categories. Participants viewed
images from both object categories as electrophysiological
data were recorded from 256 electrodes. MVPA was per-
formed on the EEG time-series data to ascertain when
category-specific information emerges in the brain. Given
the spatial limitations of EEG, source localization, en
conjunction with MVPA, was used to examine the spatio-
temporal dynamics of object perception in dorsal and
ventral regions of the brain.
Métodos
Participantes
Twenty, right-handed, neurotypical adults with normal or
corrected-to-normal visual acuity participated in the study
(12 hombres, ages 18–38 years). Each participant provided
informed written consent. All protocols received approval
by the institutional review board at the University of
Nevada, Reno. We based our sample size on previous
EEG studies that we have successfully carried out in the
pasado (Killebrew, Gurariy, Peacock, Berryhill, & Caplovitz,
2018; Gurariy, Killebrew, Berryhill, & Caplovitz, 2016;
Peterson et al., 2014). Además, other studies in the
empirical literature that have adopted similar methodo-
logical approaches contain samples sizes that are similar
o, en algunos casos, smaller (Grootswagers, Wardle, &
Carlson, 2017; Carlson et al., 2016; Cichy, Pantazis, & Oliva,
2014; van de Nieuwenhuijzen et al., 2013). To further
improve signal-to-noise ratio (SNR), our study contained
grande, centrally presented images in addition to a very large
number of trials (1680 total; 420 per condition). Even after
data cleaning and trial exclusion, the average number of
total trials per participant was 1365. Given precedent set
by previously published studies and the measures taken
to improve SNR, we believe that the sample size used in
this study is sufficient to address the empirical questions
posed here.
Apparatus
Stimuli were displayed on a Mitsubishi Diamond Pro270
CRT monitor (20 in., 1024 × 768) with a 120-Hz refresh
tasa, running via a 2.6-Mhz Mac Mini (Apple, Cª) y
presented using the PsychToolbox (Kleiner et al., 2007;
Brainard, 1997; Pelli, 1997) for MATLAB (MathWorks
Cª, 2007). Viewing distance was 57 cm.
EEG Data Acquisition
The EEG signal was continuously recorded using a 256-
channel HydroCel Geodesic Sensor Net via an EGI Net
Amps Bio 300 amplifier (Electrical Geodesics Inc.)
sampling at 1000 Hz. The digital data were recorded using
Netstation 5.0(1) software. Impendence values were kept
at or below 100Ω. A photodiode was used to validate
frame-accurate timing of stimulus presentation.
Estímulos
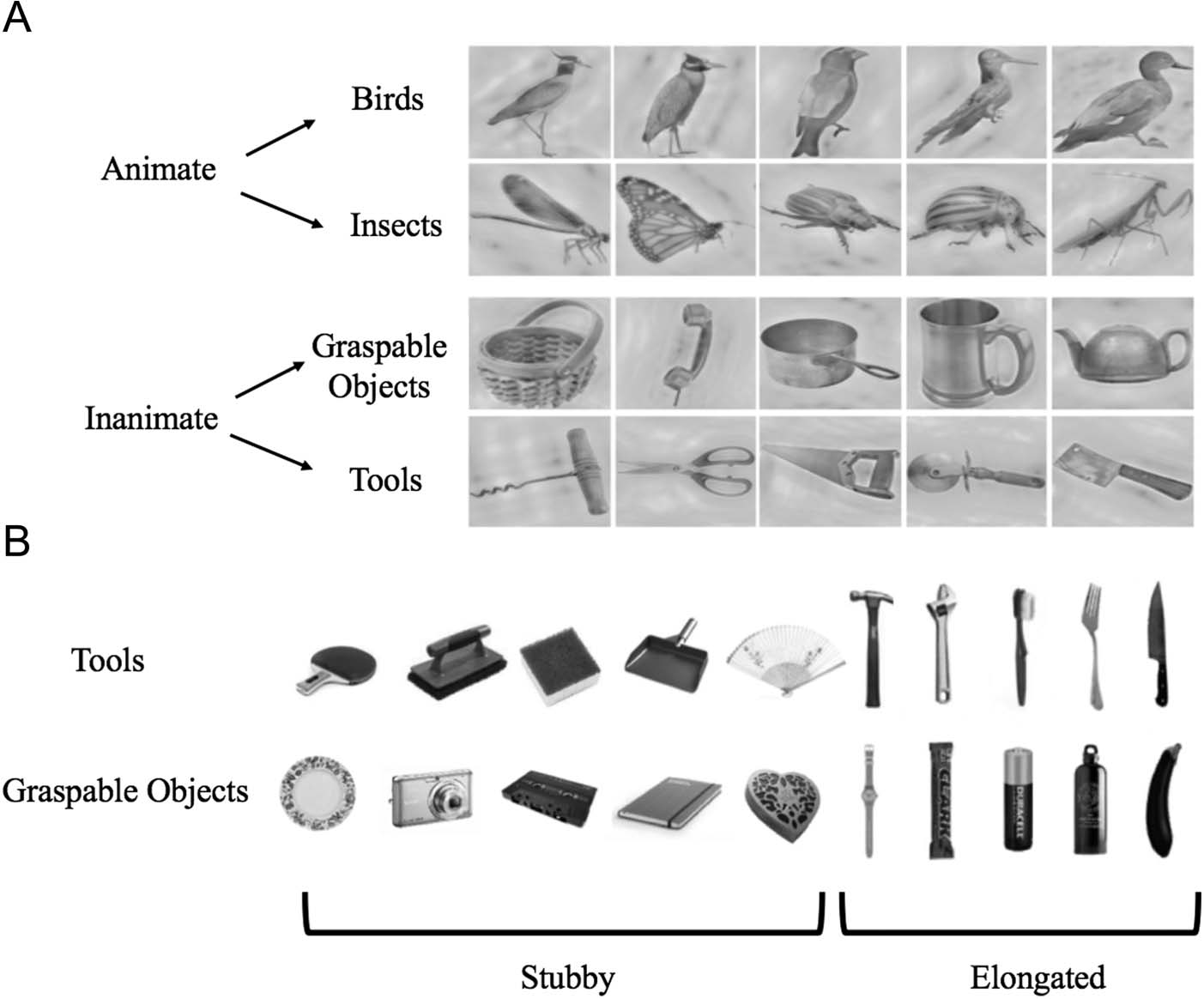
The stimuli used in Experiment 1 (Figura 1A) were chosen
from two superordinate categories of animate and inani-
mate objects. Each superordinate category was in turn
composed of two basic categories, each consisting of five
exemplars (Resultando en 20 unique, monochrome images).
In the case of animate objects, the two categories were
bird and insect, whereas in the case of inanimate objects,
the two categories were tools and graspable (nontool)
objects. Although both tools and graspable objects evoke
inferred action affordances (Gibson, 2014), pictures of
tools are associated with a stereotypical motor plan (p.ej.,
the stereotypical twisting motion associated with a screw-
conductor) more so than are graspable objects (Frey, 2007).
All stimuli were processed using the SHINE toolbox
( Willenbockel et al., 2010) to control for low-level
Gurariy et al.
969
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 1. The stimuli used in
Experimento 1 (A) came from
two superordinate categories:
animate and inanimate. Cada
superordinate category was
composed of two basic
categories (animate: bird and
insect; inanimate: tools and
graspable objects). Images
were processed using the
SHINE toolbox to match
lower level features, incluido
luminance and spatial
frequency. The stimulus
set used for Experiment 2
(B) consisted of images
from two categories: herramientas
and graspable objects. Del
10 exemplars that comprised
each category, five were
“stubby” (es decir., foreshortened)
and five were elongated.
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
differences in luminance and spatial frequency. In brief,
the Fourier power spectrum of the images without opti-
mization of the structural similarity were matched,
followed by an equating of luminance histograms over
the entire image.
Experimental Procedure
For each trial, an image was presented at the center of the
pantalla (15° × 15°) para 300 msec followed by an ISI lasting
entre 800 y 1200 mseg. During each trial, participar-
pants were instructed to maintain central fixation upon a
black fixation square in the center of the screen. Cada uno de
el 20 exemplars that made up the four categories was
presentado 84 times resulting in 420 trials per condition
y 1680 trials in total. The order of presentation was ran-
domized. To compel attentive viewing, participants were
instructed to press the space bar if an image appeared at a
reduced luminance (50%), which occurred on 5% del
ensayos. The data from these reduced luminance (es decir., objetivo)
trials were removed from further analysis. Although this
task was orthogonal to the neural processes explored in
our main analysis, object perception (especially in the
context of well-known object categories) is thought to
be an efficient, rapid, and fairly automatic process
(Hung, Kreiman, Pogio, & DiCarlo, 2005; Dell’acqua &
Job, 1998; Thorpe, Fize, & Marlot, 1996; Potter & Exacción,
1969). Además, the stimuli were large and visible for
300 mseg, leaving ample time for participants to appre-
hend each image at the level of identity. Although slower
presentation times are not required for successful object
identification (Thorpe et al., 1996; Potter & Exacción, 1969),
they are conducive to activation of higher-level visual
areas and more abstract levels of visual processing
(Grootswagers, robinson, & Carlson, 2019; robinson,
Grootswagers, & Carlson, 2019).
Multiple published studies examining the neural
dynamics of visual object processing have successfully uti-
lized behavioral tasks that were orthogonal to perceptual
categorization (Cichy, Pantazis, & Oliva, 2016; Kaneshiro,
Perreau Guimaraes, kim, Norcia, & Suppes, 2015; Cichy
et al., 2014; Carlson et al., 2013; van de Nieuwenhuijzen
et al., 2013). Además, object-related activity in the
dorsal pathway has been observed in the absence of action
planificación (Kourtzi & Kanwisher, 2000; Faillenot, Decety, &
Jeannerod, 1999; Grill-Spector et al., 1999; Sereno &
Maunsell, 1998), and evidence exists that the elicitation
of affordances and motor plans can occur automatically
(Grèzes, Tucker, Armony, Ellis, & Passingham, 2003;
Gentilucci, 2002) and in response to 2-D images (Craighero,
Bello, Fadiga, & Rizzolatti, 2002; Craighero, Fadiga,
Rizzolatti, & Umilta, 1998; Craighero, Fadiga, Umiltà, &
Rizzolatti, 1996). De este modo, although the behavioral task did
not require identification of or attention to image cate-
gory, our experimental design was sufficient to elicit
neural activity corresponding to categorical identity, en
addition to low-level features. Además, using a task
that involves category judgments would elicit additional
cognitive operations that could introduce confounds into
the results of our classification analysis.
970
Revista de neurociencia cognitiva
Volumen 34, Número 6
EEG Preprocessing
Analysis of EEG data was carried out using the FieldTrip
Toolbox (Oostenveld, Fries, Maris, & Schoffelen, 2011)
along with custom scripts written in MATLAB. A bandpass
filter (0.5–40 Hz) was applied to remove slow drift and
electrical noise. Próximo, the data were rereferenced from
Cz to an average reference. The filtered time series was
then segmented into 550-msec epochs (a 50-msec base-
line followed by 500 msec of electrophysiological data
after stimulus onset). Segmentation was performed using
trigger markers that were sent to the acquisition computer
at the onset of each trial. The temporal offset that existed
between the physical presentation of the stimulus and
the registration of the stimulus marker in the acquisition
computer was measured using a photodiode and cor-
rected for during trial segmentation. Próximo, ocular artifacts
(blinks and saccades) were identified using FieldTrip
funciones (ft_artifact_eog and ft_artifact_zvalue).
En breve, these functions detect ocular artifacts by thresh-
olding the z-transformed value of the preprocessed raw
datos. Any trial contaminated by the presence of either
artifact was removed from further analysis. Trials in
which participants performed the contrast discrimination
task were also discarded. After data cleaning and trial
removal, all participants had a minimum of 1224 ensayos,
whereas the average number of trials across all partici-
pants was 1365. Using a combination of custom MATLAB
scripts and visual inspection, bad EEG channels were iden-
tified for each trial. Data from these faulty channels were
replaced with a weighted average of all neighboring
electrodes using spherical spline interpolation (Perrin,
Pernier, Bertrand, & Echallier, 1989). Para cada participante,
the EEG epochs described above were grouped into four
condiciones: bird, insect, tool, and graspable object.
Sin embargo, trial rejection resulted in an unequal number
of trials across these conditions. To remedy this issue,
the condition with the least number of usable trials was
identified for each participant and a random subset of tri-
als equaling this number was selected for the remaining
three conditions. This resulted in each condition having
an equal number of trials for each participant with no
condition having fewer than 295 ensayos.
Multivariate Analysis
To determine the degree of object information present in
the electrophysiological data, we employed MVPA with the
aid of the CoSMoMVPA MATLAB toolbox (Oosterhof,
Connolly, & Haxby, 2016). Given the relative abundance
of trials for each condition, we were able to perform sub-
averages to increase the SNR of the data (Grootswagers
et al., 2017). To perform these averages, trials within each
condition were grouped by exemplars, split into subsets of
five trials (all of the same exemplar), and averaged
together. Por ejemplo, if a condition initially contained
420 ensayos (84 trials of each exemplar), then performing
the subaverage reduced the total number of trials within
that condition to 84 (each subaveraged trial being an aver-
age of five trials from that condition). Próximo, MVPA classifi-
cation was conducted independently for each participant
on the subaveraged trials discussed above. A naive Bayes
classifier was trained to discriminate patterns of neural
activity evoked by trials across the conditions of interest.
Classifier performance was evaluated using the leave-one-
out cross-validation method. For each millisecond, el
EEG data were organized into a matrix consisting of
samples by features, where samples refer to amplitude
values for each subaveraged trial and features refer to
EEG electrodes (256 en total). Próximo, the data were split
en 10 chunks and organized into test and training sets
such that nine of the chunks were randomly placed into
the training set and one chunk was held out as the test
colocar. This was done 10 times so that, on each iteration, a
different chunk was held out as the test data. Classifier
performance was averaged across the 10 folds at each
time point, followed by a grand average across all partici-
pants. The contrasts of interest on which MVPA was
performed were as follows: animate (bird vs. insect),
inanimate (tool vs. graspable object), animacy (animate
vs. inanimate), and all categories (bird vs. insect vs. tool
vs. graspable objects).
To identify statistically significant time points and deal
with the problem of multiple comparisons, we employed
a Monte Carlo simulation technique modeled on the
methods of Bae and Luck (2018). Primero, for each time point,
the classifier performance was submitted to a one-tailed,
one-sample t test against chance performance (1/number
of conditions). Próximo, we located “temporal clusters”
(temporally contiguous time points that were statistically
significant at an alpha level of .05) and summed the
t scores across these data points to generate a single t score
per cluster. Próximo, we reran the entire time-resolved multi-
variate analysis 1000 times using randomly reshuffled
labels for the training data. The summed t score corre-
sponding to the largest temporal cluster was stored on
each iteration. This procedure allowed us to generate a
Monte Carlo null distribution of summed t scores and
derive a threshold corresponding to the 95th percentile
( pag = .05). Finalmente, only clusters from the main analysis
whose summed t score exceeded this critical value were
treated as statistically significant. Statistical significance
for comparisons between conditions was also evaluated
using the same Monte Carlo simulation method described
arriba.
To explore the nature of the underlying neural repre-
sentation, the time series EEG data were subjected to a
multivariate time generalization analysis (Rey & Dehaene,
2014). Unlike standard time-resolved MVPA, in a time gen-
eralization analysis, a classifier trained at a given time point
is tested on all other time points of the experimental
epoch. The results can be represented as a temporal
cross-decoding matrix with each cell depicting classifier
performance for a particular combination of training
Gurariy et al.
971
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
and testing times. Standard decoding of time-resolved
M/EEG data allows for limited inferences regarding the
nature of mental representations being decoded. Cómo-
alguna vez, the time generalization method offers further
insight into these representations in the context of
spatio-temporal dynamics. Específicamente, examination of
the temporal cross-decoding matrix can reveal informa-
tion regarding the structure of underlying representa-
ciones, when those representations are activated and
how they change over time. We performed this analysis
using the same methodological approach and parameters
that were used for regular time-resolved MVPA, pero el
classifier trained on each time point was subsequently
tested on every other time point. These results were eval-
uated via comparison to chance using a one-tailed, uno-
sample t test and an adjusted p value corresponding to
a false discovery rate corrected threshold of q = .05
(Benjamini & Hochbery, 1995).
Source Localization
Source localization was performed using Brainstorm soft-
mercancía (Tadel, Baillet, Mosher, Pantazis, & Leahy, 2011),
which is documented and freely available for download
online under the GNU general public license (neuroimage
.usc.edu/brainstorm). This was done for all 20 Participantes.
Sin embargo, En el caso de 10 Participantes, the source local-
ization was further constrained by anatomical T2 MRI
scans (3-T Skyra MRI, 64-channel phased-array head coil;
repetition time = 3 segundo, echo time = 304 mseg, flip angle =
7°, 640 × 640 matrix, resolution = 0.375 × 0.375 ×
0.8 mm) collected from these individuals on a previous
occasion. For the remaining participants, the default
Colin27 MNI brain (Holmes et al., 1998) was used to con-
strain the results of the source analysis. These anatomical
images were transformed into a unique cortical space for
each participant consisting of 15,000 hypothetical sources
(7500 per hemisphere) oriented orthogonal to the cortical
sheath. Próximo, the boundary element method (Gramfort,
Papadopoulo, Olivi, & Clerc, 2010) was used to model
Cifra 2. Dorsal (A) and ventral
(B) pathway ROIs, shown in the
izquierda (l) y correcto (R) hemispheres
on inflated cortical surfaces.
each participant’s inner skull, outer skull, and head sur-
rostro. Using the EGI GPS Solver software, physical locations
of all electrodes during the experiment were triangulated,
imported into Brainstorm, and coregistered with existing
head surfaces. In the case of participants for whom only
the default MRI anatomy was available, their surfaces
(source space, inner skull, outer skull, and head surface)
were warped to match the head shape and size generated
by the electrode location data (Leahy, Mosher, Spencer,
Huang, & Lewine, 1998). Próximo, a forward model was com-
puted using the OpenMEEG software (Kybic et al., 2005)
followed by a noise covariance matrix from all individual
trials for a single participant. Finalmente, the inverse model
was calculated using minimum norm estimation, con un
current density map as the measure and constrained
diploe orientation (normal to cortex).
Source-Localized MVPA
To avoid some of the potential limitations and impreci-
sions associated with EEG source localization, MVPA was
performed within two broadly defined ROIs, each cover-
ing a substantial swathe of the cortex. This rather conser-
vative approach allowed us to disassociate computations
between the dorsal and ventral pathways without making
overly precise claims regarding the neural loci of this
actividad. The two ROIs included a ventral and dorsal region
(Cifra 2). Each ROI was collated using multiple regions
extracted from the Desikan–Killiany Atlas (Desikan et al.,
2006). The bilateral dorsal ROIs were composed of the
superior parietal, inferior parietal, and supramarginal
regiones, whereas the bilateral ventral ROIs consisted of
the fusiform, inferior temporal, parahippocampal, y
entorhinal regions.
Accurate source localization of EEG data depends on
several parameters, Por ejemplo, realistic modeling of
the cortex, inner skull, outer skull, and head surface; acumular-
rate coregistration of electrode locations, Etcétera.
Given the numerous challenges involved in source local-
ización, dorsal and ventral ROIs were defined very broadly.
972
Revista de neurociencia cognitiva
Volumen 34, Número 6
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Sin embargo, further measures were taken to ensure that the
activity extracted from the two ROIs can be reasonably
expected to represent actual signal from those regions.
Simulated EEG waveforms were generated by projecting
activity from an ROI to the surface electrodes by multiply-
ing the source time series with the forward model. Este
was done separately in dorsal and ventral ROIs while activ-
ity in all other regions was set to zero. Próximo, these simu-
lated waveforms were localized back into dipole space
using the source localization process described above.
Examining the localization results for a known source
was used to examine the accuracy of the forward and
inverse models.
To perform MVPA on source-localized data, individual
preprocessed trials from each participant (described in
the EEG Data Preprocessing section) were averaged
together to create subaverages, each composed of five
ensayos. The trials that composed each subaverage were of
the same exemplar image, and the total number of newly
created subaveraged trials was held constant across all
condiciones. Próximo, source localization (see previous section
for details) was performed on each subaveraged trial, y
individual sources from each ROI were extracted. Este
resulted in a distribution of cortical sources for each
millisecond of the experimental epoch for each ROI.
Given the substantial number of individual sources within
each ROI and the fact that this number differed between
the ROIs, principal component analysis was performed on
los datos. The first 40 components were retained to be
used as feature inputs for the MVPA. The classification pro-
cedure used for these data recapitulated those used in the
EEG analysis at the sensor level. At each millisecond, el
data were divided into 10 chunks, nine of which would
serve as the training set and one as the test set. Próximo, a
leave-one-out method was used where, on each fold, a dif-
ferent chunk was held out as the test set. The performance
of a naive Bayes classifier was averaged across the 10 folds
for each time point, averaged across all participants, y
statistically evaluated using the Monte Carlo simulation
described in the Multivariate Analysis section.
Resultados
Electrode Time Course MVPA
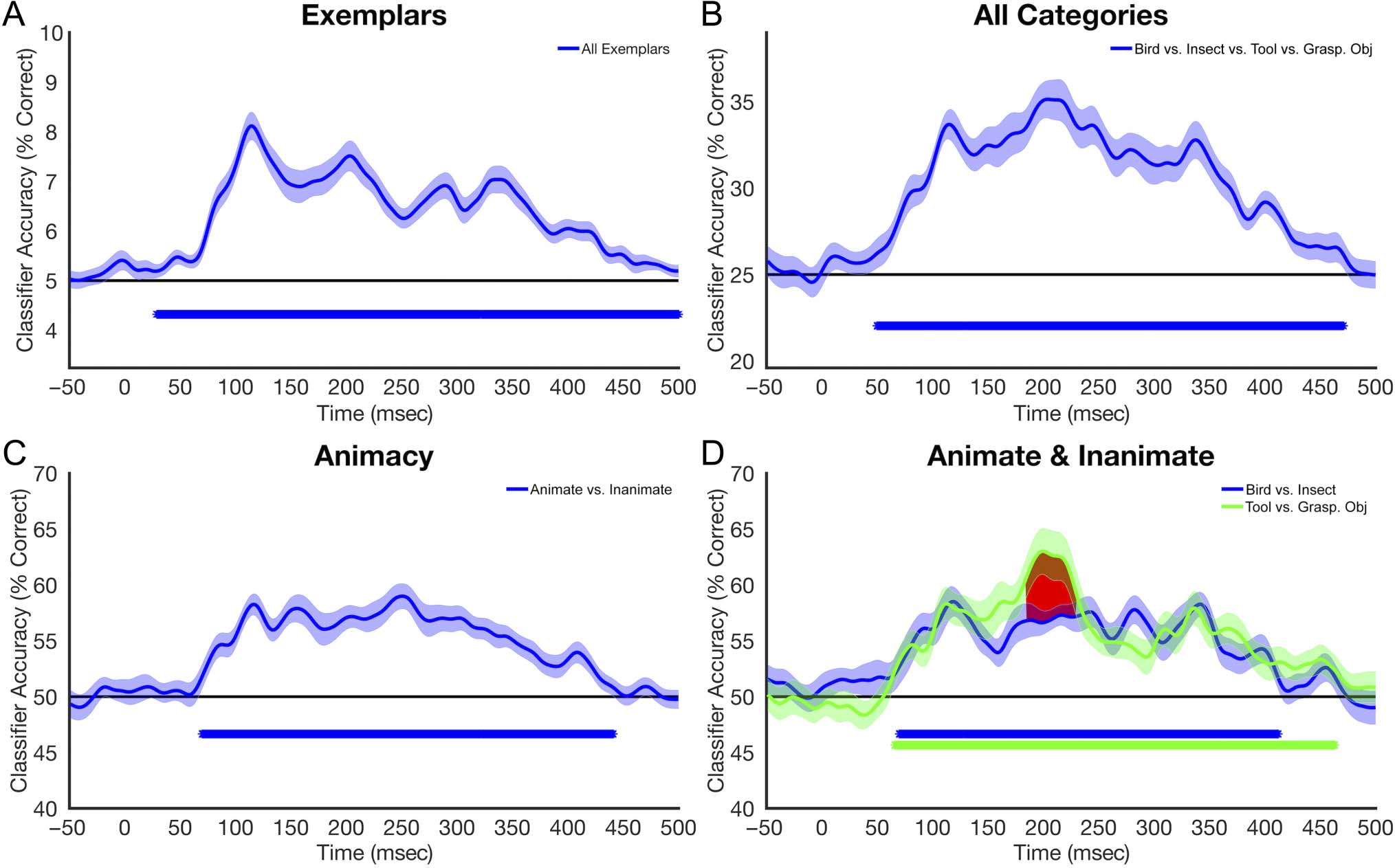
Cifra 3 depicts the results of the MVPA performed on the
time course EEG data. Above-chance classification was
observed for all examined comparisons (exemplars; todo
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 3. Decoding performance averaged across participants, plotted for each millisecond of the experimental epoch. Shaded regions around the
curve represent SE. The black horizontal line represents chance performance. Asterisks below the line of chance represent time points at which
classification was statistically significant after correcting for multiple comparisons. (A) Decoding of individual exemplars. (B) Decoding between the
four basic categories (insect vs. bird vs. tool vs. graspable object). (C) Decoding of animacy (animate vs. inanimate). (D) Decoding within animate
(bird vs. insect) and within inanimate (tool vs. graspable objects) object categories. Red shading represents time points at which the two curves were
significantly different from one another after multiple comparisons correction. Grasp. Obj = graspable object.
Gurariy et al.
973
categories, animacy; bird vs. insect; tool vs. graspable
object). The onset for significant classification occurred
at around 60 msec after stimulus onset in all comparisons
and slightly earlier (alrededor 40 mseg) for the individual
exemplar comparison. In comparing the classification per-
formance between the animate and inanimate conditions,
the above-chance classification onset occurred at the same
estado latente (∼70 msec). Sin embargo, in the tool versus graspable
object comparison, we observed a pronounced peak from
aproximadamente 180 a 230 msec that was not evident for the
animate conditions. Peak latencies for the exemplar, basic,
and superordinate contrasts (Figure 3A–C) occurred at
114, 207, y 249 mseg, respectivamente.
Although classification accuracy suggests the presence of
category-specific information in the neural data, este
measure says relatively little regarding the nature of the
underlying representation. This question was explored
using the time generalization method in which a classifier
was trained at each time point and subsequently tested
on all other time points. The resulting temporal cross-
decoding matrix illustrates whether the neural representa-
tion at any single time point generalizes to other latencies,
hence providing a window into the neural dynamics of the
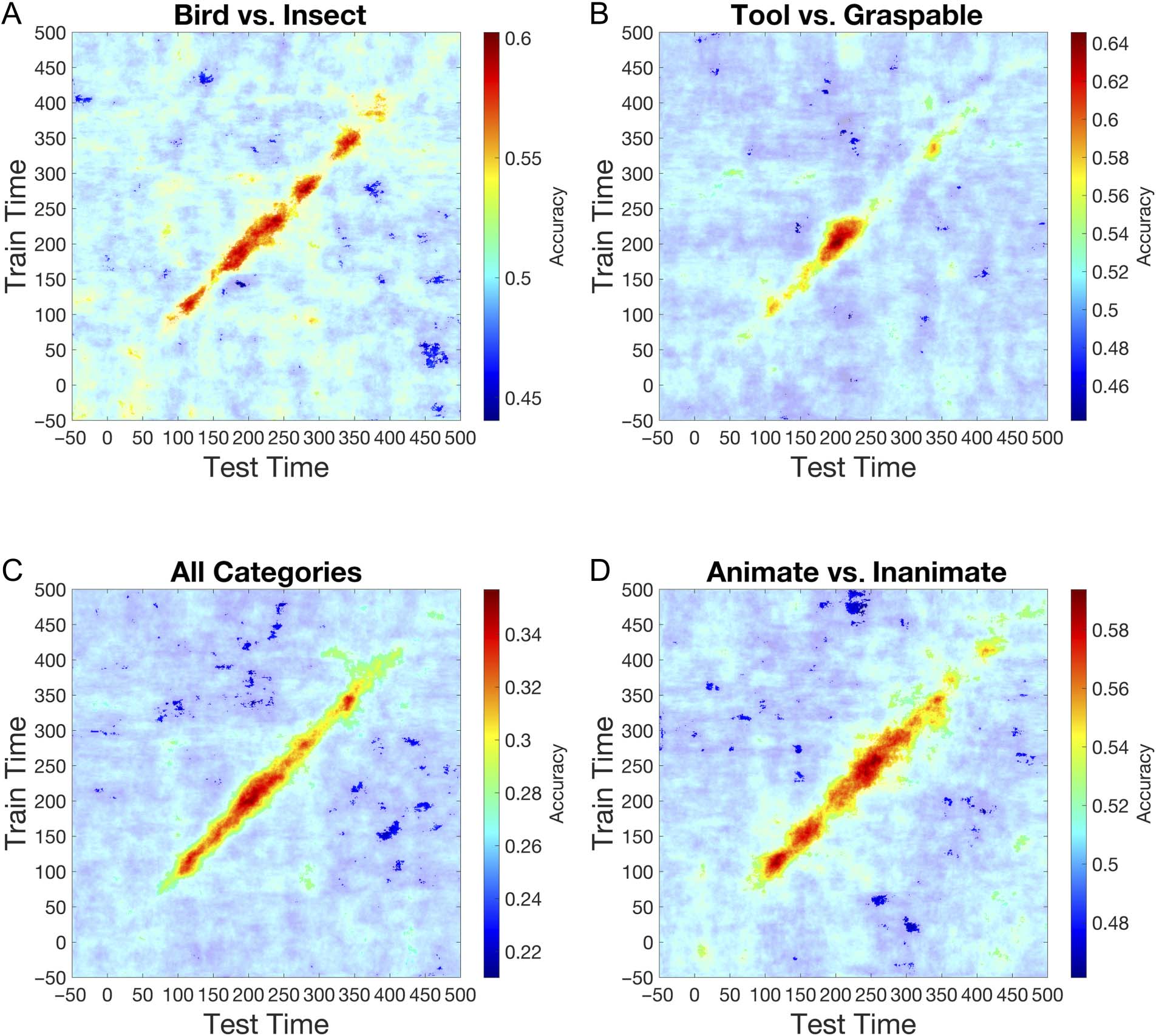
underlying representation. The results (Cifra 4) depict a
similar pattern for the four MVPA conditions explored in
this analysis (all categories; bird vs. insect, animate vs.
inanimate, tool vs. graspable object). Específicamente, cells
with above-chance classification are confined within a
relatively narrow region of the matrix. The temporal
window within which the classifier could generalize was
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 4. Temporal cross-decoding matrices averaged across participants. Classifiers trained on each time point of the experimental epoch ( y axis)
were then tested on every other time point (x axis). Values plotted in the matrix represent classifier accuracy at each combination of points.
Highlighted regions signify time points that were statistically significant at a false discovery rate corrected p value. (A) Temporal cross-decoding
matrix for basic object categories (insect vs. bird vs. tool vs. graspable objects). (B) Temporal cross-decoding matrix for animate (bird vs. insect)
object categories. (C) Temporal cross-decoding matrix for animacy (animate vs. inanimate). (D) Temporal cross-decoding matrix for inanimate (tool
vs. graspable object) object categories.
974
Revista de neurociencia cognitiva
Volumen 34, Número 6
approximately 30–40 msec, suggesting a highly dynamic
and evolving neural representation.
Source-Localized MVPA
The presence of a known source allows for the evaluation
of the forward and inverse model in terms of spatial accu-
racy. We therefore conducted an analysis in which simu-
lated EEG waveforms were generated by multiplying the
source time series by the forward model and then localiz-
ing this activity back into dipole space. The results of the
model evaluation are shown in Figure 5 for a representa-
tive participant. A qualitative inspection of the data sug-
gests that activity originating from the dorsal or ventral
pathway can be correctly localized. En tono rimbombante, actividad
localized to dorsal ROIs did not appear to contaminate
activity in ventral ROIs, y viceversa.
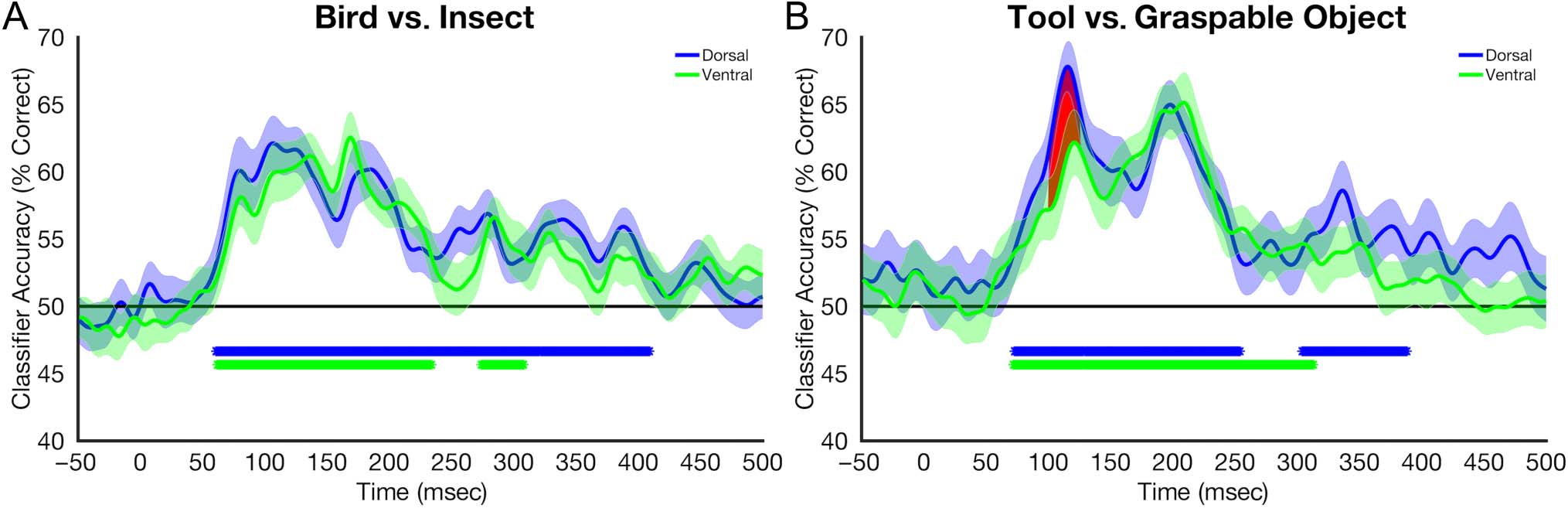
Given previously reported differences in object selec-
tivity in the dorsal and ventral pathways, we analyzed
the time course classification performance based on
source-localized data extracted from dorsal and ventral
ROI. For both the animate (bird vs. insect) and inanimate
(tool vs. graspable object) contrasts, above-chance
performance was observed in both ventral and dorsal
ROI (Cifra 6). Onset latencies for above-chance classi-
fication were similar across both ROIs, starting at approx-
imately 75 msec after stimulus onset. Regions shaded in
red represent time windows during which classification
differed significantly between the two pathways. Of note,
significantly higher performance was observed in the dor-
sal ROI from 100 a 130 msec for the comparison of tools
versus graspable objects. For the decoding of the animate
object categories (birds vs. insects), the classification time
courses were similar across both ROIs with no time points
at which statistically significant differences we observed.
Above-chance classification of object categories within
the ventral pathway has been observed before and is
consistent with the known properties of ventral neural
poblaciones. Sin embargo, above-chance classification of birds
versus insects in the dorsal pathway was an unexpected
finding—one that may call for a reevaluation of dorsal
pathway computations and its role in visual perception.
Successful decoding was observed although the stimuli
used in this experiment were processed to control for
low-level differences (luminance and spatial frequency).
Sin embargo, global shape differences between birds and
insects (although not as pronounced as those between
tools and graspable objects) may still exist and account
for some of the results reported in this section. Interest-
ingly, the onset of above-chance classification for the
birds–insects contrast did not differ across the two path-
maneras. This suggests that dorsal object representations
reported here are not likely to be a consequence of feed-
forward signals from ventral regions. Bastante, these results
suggest that the dorsal pathway is capable of forming
independent object representations in the absence of
visuomotor behaviors for animate categories lacking
motor plans.
Cifra 5. EEG recordings were simulated by projecting dipole activity in dorsal and ventral ROIs (separately) while activity in all other regions was
set to zero. Próximo, source localization was performed on the resulting, simulated waveforms using the same parameters and surfaces as described in
the main experiment. The resulting source-localized diploe activity is plotted at different time points in dorsal (teal) and ventral (pink) ROI. (A)
Simulations of ventral activity. (B) Simulations of dorsal activity.
Gurariy et al.
975
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 6. Decoding performance averaged across participants, plotted across time, and quantified as percent correct. Shaded regions around the
curve represent SE. The black horizontal line represents chance performance. Asterisks below the line of chance represent time points at which
classification was statistically significant after correcting for multiple comparisons. (A) Decoding of bird versus insects. (B) Decoding of tools versus
graspable objects.
EXPERIMENT 2
The stimuli used in Experiment 1 contain a potential con-
found that is especially evident for the inanimate condi-
ciones. Específicamente, there are systematic differences in
shape that can be seen between exemplars that make up
the tools and graspable object categories. Tools tend to be
elongated along the principal axis, whereas graspable
objects are typically foreshortened, or “stubby,” along
the central axis. Previous research suggests that the
selectivity toward tools observed within the dorsal path-
way may be, at least in part, explained by this systematic
elongation that tends to co-occur in objects that comprise
most tool stimulus sets (Chen et al., 2018; Sakuraba et al.,
2012). De este modo, it is not clear whether the classification
performance observed for tools versus graspable objects
in Experiment 1 was driven by shape rather than category
membership. To address this issue, Experimento 2
replicates the analyses from Experiment 1 on tool and
graspable object images chosen from a new stimulus
set—one designed to control for shape differences. Este
new stimulus set contains novel exemplars of tools and
graspable objects, with each category including both
elongated and stubby exemplars. Además, it allows
for the disambiguation of shape from toolness and an
examination of how these parameters contribute to the
neural representation of objects.
Métodos
Participantes
Twenty, right-handed, neurotypical adults with normal or
corrected-to-normal visual acuity participated (11 hombres,
ages 18–43 years) and provided informed written consent.
De estos 20 individuals, 10 also participated in Experiment 1.
The reason for running some participants in both studies
had to do with the availability of anatomical MRI scans
(which help improve source localization accuracy). El
institutional review board at the University of Nevada,
Reno approved all protocols. The corresponding section
of Experiment 1 provides a discussion and justification
of the sample size used in the current study.
Apparatus and EEG Data Acquisition
The stimulus display and EEG recording methods were
identical to those used for Experiment 1.
Estímulos
The stimuli used in Experiment 2 (Figura 1B) consistió en
20 unique, monochrome images that varied along two
dimensions: shape and toolness. Along the toolness dimen-
sión, each image could be classified as either a tool or a
graspable object (see the Stimuli subsection of Experiment 1
Methods for the specific distinction between these catego-
ries). Along the shape dimension, the profile of each image
was either stubby or elongated. De este modo, the stimuli could be
organized into the following nonoverlapping categories:
stubby tool, stubby graspable object, elongated tool, y
elongated graspable object. Unlike the stimulus set used
for Experiment 1, these stimuli were not processed with
the SHINE toolbox given the importance of texture and
shading cues for the dorsal pathway (Freud et al., 2016).
Experimental Procedure
The experimental design used in Experiment 2 mimicked
that of Experiment 1. Please see the Experimental Proce-
dure section of Experiment 1 for details.
EEG Preprocessing
The preprocessing pipeline was identical to the one used
for Experiment 1. See the EEG Preprocessing section in
Experimento 1 for specific details. In brief, data were
976
Revista de neurociencia cognitiva
Volumen 34, Número 6
filtered, referenced, and segmented into individual trials.
Data from defective EEG channels were interpolated via
averaging, and trials that were contaminated by the pres-
ence of ocular artifacts were removed from further analy-
hermana. After data cleaning and trial removal, all participants
had a minimum of 1048 ensayos, whereas the average num-
ber of trials across all participants was 1294. Additional tri-
als were randomly removed to ensure that all conditions
within a participant had an equal number of trials. All par-
ticipants had a minimum of 257 trials per condition.
Multivariate Analysis
The steps involved in the multivariate analysis were
identical to those used in Experiment 1. See the Multivar-
iate Analysis section of Experiment 1 for specific details.
The contrasts of interest on which MVPA was performed
were as follows: forma (all elongated vs. all stubby objects),
toolness (all tools vs. all graspable objects), elongated tools
versus elongated graspable objects, and stubby tools ver-
sus stubby graspable objects. A time generalization analysis
using the methods described in Experiment 1 was also per-
formed for the toolness and shape contrasts.
Source Localization and Source-Localized MVPA
The steps involved in source localization and the corre-
sponding MVPA were identical to those used in Experiment
1. See the corresponding sections of Experiment 1 para
details. At the completion of this process, source-localized
activity was extracted from dorsal and ventral ROIs and
used as input into the MVPA classifier. The classifier was
trained to discriminate between the following conditions
in both dorsal and ventral ROIs: forma (all elongated vs.
all stubby objects), toolness (all tools vs. all graspable
objects), elongated tools versus elongated graspable
objects, and stubby tools versus stubby graspable objects.
Resultados
Time Course MVPA
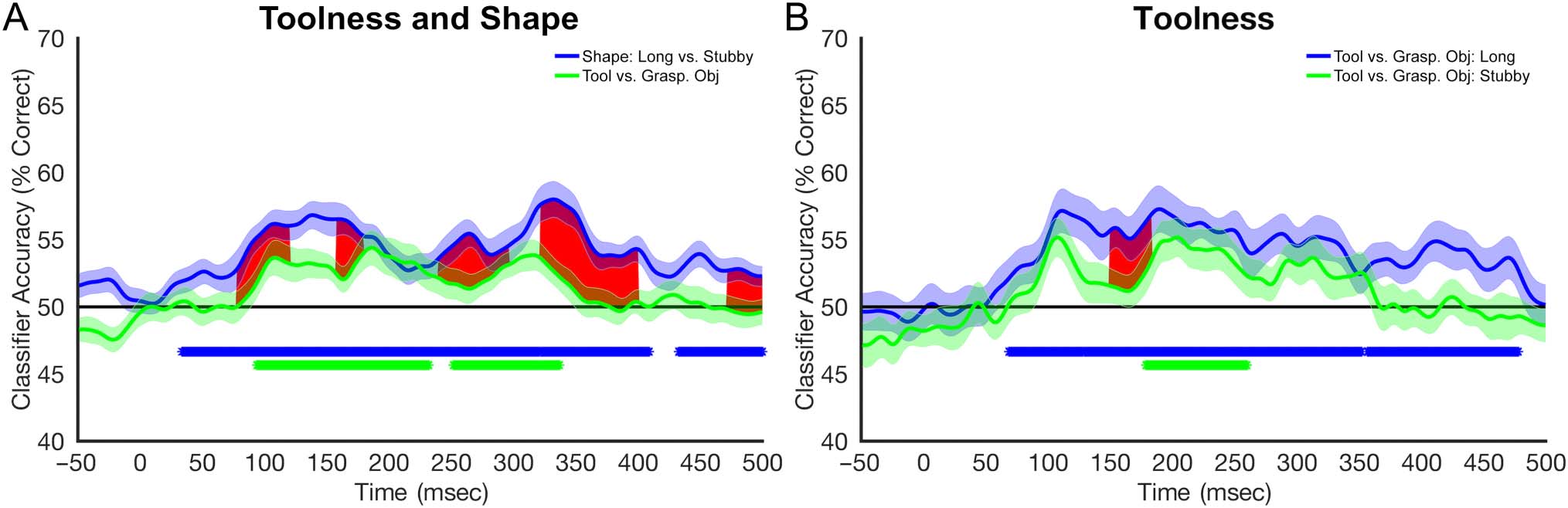
Cifra 7 depicts the results of the MVPA performed on
the time course EEG data. Above-chance classification per-
formance was observed for all of the examined contrasts.
Figure 7A compares classification of shape (all long vs. todo
stubby) and toolness (all tools vs. all graspable objects).
Classification of object shape reached significance at
∼60 msec, whereas classification of toolness reached sig-
nificance approximately 35 msec later (∼95 msec). At mul-
tiple time windows, performance was statistically higher
for shape as compared to the toolness condition. Specifi-
cally, this occurred at 80–120, 158–180, 240–297, 320–400,
and 470–550 msec.
The stimuli used in classifying all tools versus all
graspable objects included both stubby and elongated
exemplars within each category. Figure 7B displays the
classification between tools and graspable objects when
shape was held constant. The results suggest that, incluso
when shape is controlled for, the EEG signal contains
information distinguishing tools from graspable objects.
En general, performance was better when the classifier was
trained on elongated exemplars of tools and graspable
objects as opposed to stubby exemplars from these cate-
gories. The onset of significant classification for elongated
objects began at approximately 70 mseg. Classification of
stubby objects was weaker and less consistent, alcanzando
significance starting at approximately 184 mseg. Statisti-
cally significant differences between the two conditions
were observed within the time window of 150–182 msec,
with higher performance for the elongated condition. Como
in Experiment 1, the time generalization analysis shows
that above-chance classification was constrained to a rela-
tively narrow time window of approximately 30–40 msec,
suggesting a dynamic and evolving neural representation
(Cifra 8).
Cifra 7. Decoding performance averaged across participants, plotted across time, and quantified as percent correct. Shaded regions around the
curve represent SE. The black horizontal line represents chance performance. Red shading highlights time points at which the two curves were
significantly different from one another after multiple comparisons correction. Asterisks below the line of chance represent time points at which
classification was statistically significant after correcting for multiple comparisons. (A) Decoding of toolness and shape. (B) Decoding tool versus
graspable object (long or stubby).
Gurariy et al.
977
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 8. Temporal cross-decoding matrices averaged across participants. Classifiers trained on each time point of the experimental epoch ( y axis)
were then tested on every other time point (x axis). Values plotted in the matrix represent classifier accuracy at each combination of points.
Highlighted regions signify time points that were statistically significant at a false discovery rate corrected p value. (A) Temporal cross-decoding
matrix for toolness (tool vs. graspable object). (B) Temporal cross-decoding matrix for object shape (elongated vs. stubby).
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Cifra 9. Classification performance averaged across participants and plotted across time. Shaded regions around the curve represent SE. Regions
shaded in red represent time windows during which accuracy differed significantly between the two curves following corrections for multiple
comparisons. The black horizontal line represents chance performance. Asterisks below the line of chance represent time points at which classification
was statistically significant after correcting for multiple comparisons. (A) Decoding object shape. (B) Decoding toolness without controlling for
elongation. (CD) Decoding toolness while controlling for elongation.
978
Revista de neurociencia cognitiva
Volumen 34, Número 6
Source-Localized Time Course MVPA
As in Experiment 1, we analyzed the time course classifica-
tion performance based on source-localized data extracted
from dorsal and ventral ROIs (Cifra 9). As demonstrated
in Figure 9A, shape could be successfully decoded in both
ROIs beginning at 55 msec in the ventral pathway and
66 msec in the dorsal pathway. Classification accuracy
for both ROIs remained fairly similar until approximately
337 mseg, after which significantly better performance was
observed in the dorsal ROI. Figure 9B displays classifica-
tion performance between elongated tools and stubby
graspable objects. This contrast was chosen as it most
closely approximates the stimulus characteristics used in
Experimento 1. As is evidenced by the data, failure to con-
trol for shape between object categories resulted in higher
exactitud (relative to Figure 9C and D, in which shape was
controlled for). Above-chance classification of long tools
and stubby graspable objects occurred in the ventral path-
way at 43 msec after stimulus onset, whereas dorsal classi-
fication reached statistical significance at 63 mseg. Finalmente,
Figure 9C and D suggests that category information
regarding toolness was present in the data even when
the shape confound between tools and graspable objects
was controlled for. Específicamente, for both dorsal and ventral
ROI, significant decoding of tool versus graspable objects
was observed when limiting the comparison to only long
or only stubby objects. Juntos, these results suggest
that both shape and object identity are represented within
neural activity across both the dorsal and ventral visual
pathways. Además, given similar onset times for suc-
cessful decoding, these data are consistent with a model in
which neural representations of objects are computed
independently within the two pathways.
DISCUSIÓN
The purpose of this study was to explore the spatio-
temporal dynamics of visual object processing in the
human cortex. To achieve this goal, we collected evoked
responses to different object categories using HD-EEG.
MVPA was used to detect the presence of category-specific
information within the EEG signal and to explore the tem-
poral dynamics of category classification. Source localiza-
tion combined with MVPA was employed to explore the
neural origins of the EEG signal, focusing on the temporal
dynamics within ventral and dorsal neural pathways. Nosotros
report successful classification of object categories at both
the electrode level and the source-localized dipoles
extracted from these two neural pathways.
In Experiment 1, participants viewed images from two
superordinate categories, each in turn consisting of two
basic categories (animate: bird and insect; inanimate: tool
and graspable object). Electrode-level MVPA results from
Experimento 1 showed that objects could be successfully
classified across different hierarchical levels of categoriza-
ción. Above-chance classification was observed when
testing and training sets were organized into superordi-
nate level categories (animate vs. inanimate) and basic
level categories (bird vs. insect vs. tool vs. graspable
object) as well as individual exemplars. Onset latencies
were similar across these conditions, whereas the peak
latencies revealed a distinct temporal trend; exemplar
decoding showed the earliest peak, followed by the basic
and superordinate categories, respectivamente. These tempo-
ral patterns have been observed before (Contini et al.,
2017; Cichay et al., 2014; Carlson et al., 2013) and have
implications regarding the neural bases of object percep-
ción. En particular, these data suggest that object represen-
tations in the brain evolve from lower to increasingly
higher levels of abstraction (exemplar → basic → superor-
dinate). This is consistent with an object perception
framework in which categorization can be understood as
evidence accumulation over time (Mack & Palmeri, 2011;
Philiastides & Sajda, 2006). This conclusion is further
corroborated by a time generalization analysis showing
that the neural representations underlying the object
categories were not stable but rather evolved dynamically
over the course of the epoch.
Whereas some low-level features (es decir., luminance) eran
controlled in Experiment 1, other potentially confounding
variables were not. Específicamente, the stimulus set contained
shape confounds that were especially pronounced
between tools and graspable objects (elongation along
the principal axis). To remedy this issue, a follow-up
Experimento 2 was performed in which tools and graspable
objects were composed of both elongated and stubby
exemplars. This change allowed us to examine the contri-
bution of shape to the neural representation of object cat-
egories. Data from Experiment 2 suggest that information
pertaining to both category and shape are present in the
EEG signal. This is evidenced by successful classification
for tools versus graspable objects even when restricting
the analysis to only elongated or only stubby exemplars.
We used source localization to explore the spatio-
temporal dynamics of visual object processing within
two broadly defined neural regions: dorsal and ventral
corteza. In Experiment 1, without controlling for elonga-
ción, we observed above-chance classification for tools
versus graspable objects in both dorsal and ventral
stream ROIs. The time course of classifier performance
was similar across the two pathways, with the exception
of a dorsal stream advantage (higher classification accu-
racy) from approximately 100 a 130 mseg. A similar pat-
tern was observed in Experiment 2 when the classifier
was trained to discriminate between elongated tools and
stubby graspable objects (a replication of the conditions
in Experiment 1). Restricting the analysis to all elongated
versus all stubby objects (irrespective of toolness) de nuevo
produced a qualitatively similar time course in both dorsal
and ventral ROIs. Además, performance for shape
(elongated vs. stubby) was overall more robust compared
to category (tool vs. graspable object), partially validating
concerns regarding shape as a potential confound. Allá
Gurariy et al.
979
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
has been a long-standing debate in the literature about
whether the organizing principles of category-selective
neural regions are best described by shape and low-level
propiedades (Nasr, Echavarria, & Tootell, 2014; Rice et al.,
2014; Watson et al., 2014; yue, Pourladian, Tootell, &
Ungerleider, 2014; Baldassi et al., 2013; Rajimehr, Devaney,
Bilenko, Joven, & Tootell, 2011) or category membership
(Kriegeskorte et al., 2008; Kiani, Esteky, Mirpour, & Tanaka,
2007). Bracci and Op de Beeck (2016) addressed this
issue by generating a two-factorial stimulus set in which
the contribution of category and shape could be dissoci-
ated in humans. Their results suggest that shape and cat-
egory are coded independently but nevertheless interact
in important ways throughout the visual hierarchy. Nuestro
findings are in line with those of Bracci and Op de
Beeck (2016), showing that successful decoding of tools
versus graspable objects can be achieved at the electrode
nivel, as well as in the source-localized data, even when
shape has been controlled for. Grootswagers, robinson,
Shatek, and Carlson (2019) reached a similar conclusion
upon measuring EEG responses to intact objects from
different conceptual categories as well as “textform” ver-
sions of each image that were rendered unrecognizable
while preserving numerous midlevel features. Su
results demonstrate that the statistical regularities main-
tained in the scrambled images were indeed sufficient for
above-chance decoding of animacy. Sin embargo, classifier
performance for intact images was significantly more
robusto, suggesting that featural confounds contribute to
but cannot fully account for the classification of conceptual
categories (see Long, Yu, & Konkle, 2018, for similar
findings using fMRI).
We also observed that classification performance was
generally better when the tool category consisted of elon-
gated (rather than stubby) images. This may reflect the fact
that tools tend be elongated in real life, and hence these
images may constitute better exemplars of the tool cate-
gory. Previous studies have reported the existence of
regions in the dorsal pathway that show selectivity for
elongated objects, irrespective of their semantic category
(Sakata et al., 1998). The existence of such regions may
partially explain why classification performance was ele-
vated for elongated versus stubby exemplars, especially
in the dorsal pathway. Chen et al. (2018) also investigated
the relationship between elongation and toolness using
resonancia magnética funcional. They reported that both toolness and elongation
are processed by both separate and common regions
and that elongated tools (but not stubby tools) facilitate
reciprocal connectivity between the ventral and dorsal
regiones.
One unexpected finding revealed by our analyses was
the striking similarities between the dorsal and ventral
pathways pertaining to the classifier’s performance in
the decoding of different object categories as well as the
categories that could be successfully decoded. The results
of Experiment 1 suggest that information in the dorsal
pathway is not restricted to inanimate objects with
affordances (tools and graspable objects) but also extends
to animate objects categories (birds and insects). Este
result supports and extends previous findings of object
selectivity in the dorsal pathway (Freud et al., 2017; jeong
& Xu, 2016; Zachariou et al., 2014; Konen & Kastner, 2008;
Valyear et al., 2007; Fang & Él, 2005; chao & Martín, 2000;
Sereno & Maunsell, 1998). Además, in addition to evi-
dence of the dorsal pathway exhibiting properties similar
to the ventral pathway (Konen & Kastner, 2008), Tiene
been shown that the ventral pathway contains spatial infor-
formación (hong, Yamins, Majaj, & Dicarlo, 2016; Zoccolan,
Kouh, Pogio, & DiCarlo, 2007) and can be modulated
by motor attributes (Gallivan, Chapman, Mclean, Flanagan,
& Culham, 2013; Astafiev, Stanley, Shulman, & Corbetta,
2004). In light of such findings, the utility of the dorsal–
ventral dichotomy has been questioned, with some sug-
gesting the existence of additional pathways (Haak &
beckman, 2018) and others arguing for a “patchwork”
model defined by multiple interacting neural regions
(de Haan & Cowey, 2011). At the very least, such findings
urge a reassessment of the two-pathway hypothesis
y, as it relates to this study, the role of the dorsal
pathway in visual object perception (Freud, behrman,
& Snow, 2020; Erlikhman, Caplovitz, Gurariy, Medina, &
Snow, 2018; Freud et al., 2016).
Interesting possibilities regarding dorsal involvement in
visual object perception are bolstered by the anatomical
conectividad (Takemura et al., 2016; Cloutman, 2013)
and cross-talk (hutchison & Gallivan, 2018; Janssen,
Verhoef, & Premereur, 2018; de Haan & Cowey, 2011;
Schenk & McIntosh, 2010) between the two pathways as
well as the finding that neural signals propagate faster
through the dorsal relative to the ventral pathway (Sim,
Helbig, Graf, & Kiefer, 2015; Srivastava, Orban, De
Mazière, & Janssen, 2009; Norman, 2002). Juntos, estos
functional and anatomical properties give rise to the pos-
sibility that object representations computed first in the
dorsal pathway may be capable of priming or otherwise
modulating object-related computations in the ventral
pathway via feedback. Por ejemplo, Sim et al. (2015) found
that priming the dorsal pathway with movie clips of tool
use modulated ventral activity related to the recognition
of tools. Additional evidence of dorsal pathway involve-
ment in object perception comes from neuropsychology.
Several studies have documented that the behavioral
deficits associated with dorsal lesions are not always
restricted to visuomotor interactions but can also perturb
certain perceptual abilities such as global shape percep-
tion and 3-D processing ( Van Dromme, Premereur,
Verhoef, Vanduffel, & Janssen, 2016; Gillebert et al.,
2015; Lestou, Justicia, Humphreys, Kourtzi, & Humphreys,
2014). De hecho, under some circumstances, agnosic
patients with a spared dorsal pathway can outperform
somebody with the opposite pattern of damage. Para
ejemplo, agnosic patients with ventral damage (but intact
dorsal pathways) were able to identify the presence of an
object defined by disparity, whereas patients with dorsal
980
Revista de neurociencia cognitiva
Volumen 34, Número 6
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
damage were unable to detect such objects at all (Vaina,
1989). The involvement of the dorsal pathway in 3-D shape
perception has also been documented using neuroimaging
in humans (Freud et al., 2017; Katsuyama, Usui, Nose, &
Taira, 2011; Georgieva, Peeters, Kolster, Todd, & Orban,
2009; Vanduffel et al., 2002) in addition to monkey
physiology (Alizadeh, van Dromme, Verhoef, & Janssen,
2018; Janssen, Srivastava, Ombelet, & Orban, 2008).
Sin embargo, several studies (including this one) tener
found that computations in the dorsal pathway may
extend beyond the processing of 3-D shape. En efecto, evi-
dence exists that dorsal activity can be elicited from 2-D
images, even during passive viewing (Zachariou et al.,
2014; Konen & Kastner, 2008; chao & Martín, 2000;
Sereno & Maunsell, 1998). An influential article by Konen
and Kastner (2008) showed that, in addition to selectivity
for 2-D objects, the dorsal pathway shows evidence of size,
retinal position, and viewpoint invariance—properties
typically associated with ventral stream computations
(Sawamura et al., 2005; Vuilleumier et al., 2002; Kourtzi
& Kanwisher, 2000; Grill-Spector et al., 1998). Otro
findings suggest that the dorsal pathway might facilitate
perceptual tasks that require configural processing.
Zachariou, Nikas, Safiullah, Gotts, and Ungerleider
(2017) used fMRI to show that configural versus featural
face differences elicited greater activity in dorsal regions
and correlated with behavioral performance on configural
tasks whereas the deactivation of the posterior parietal
cortex using TMS adversely affected behavior on such
tareas. Other findings also suggest that the dorsal pathway
may be involved in aspects of perceptual organization
occurring over space (Xu & Chun, 2007) as well as time
(Erlikhman & Caplovitz, 2017; Erlikhman et al., 2016;
McCarthy, Kohler, Tse, & Caplovitz, 2015).
An important caveat to the above discussion is that the
dorsal pathway is not a single, functionally homogenous
neural region but rather is composed of multiple neural
regiones, each defined by its own functional profile.
Research suggests that the functional–anatomical proper-
ties of the dorsal pathway are best captured by a posterior-
to-anterior gradient. Along this gradient, neural signals
appear to be transformed from perceptual representations
to those in service of motoric interactions. In the posterior
region of the dorsal pathway, neural populations show
selectivity toward object shape, 3-D cues, and global form
(Frued et al., 2017; Erlikhman et al., 2016; Tsutsui, Jiang,
Yara, Sakata, & Taira, 2001) as well as ventral-like proper-
corbatas, such as invariance to low-level transformations
(Konen & Kastner, 2008). En cambio, anterior regions
feed into motor cortex and exhibit corresponding sensitiv-
ity to hand position, motor plans, and other computations
geared toward visuomotor interactions (Rígido & Zohary,
2008; Shmuelof & Zohary, 2005). This gradient can help
explain the range of disparate findings in the literature as
they relate to the functional role of the dorsal pathway.
localization methods applied to M/EEG data constitute
an indirect estimation of the underlying neural signal.
These methods, although generally considered reliable,
lack the spatial precision of fMRI and should be inter-
preted with some caution. To mitigate some of the uncer-
tainties pertaining to source localization, we opted for a
conservative approach in choosing ROIs. Our dorsal and
ventral ROIs covered a substantial swathe of cortex.
Although this approach minimizes the possibility of false
claims regarding activity in specific neural loci, it also pre-
cludes a more refined analysis regarding the functional
properties of different regions that make up the dorsal
and ventral pathways.
Another issue relates to the stimulus sets used for both
experimentos. The stimuli used in Experiment 1 eran
manipulated to control luminance and spatial frequency
diferencias; sin embargo, there were still shape confounds
that existed across object category. This was especially
true for the inanimate object categories where systematic
differences in elongation can be seen between tools and
graspable objects; de este modo, interpretation of results should be
undertaken with these confounds in mind. Experimento 2
addressed this issue by subdividing each basic category
into an equal number of elongated and stubby exemplars,
thereby controlling for global shape differences; sin embargo,
this experiment used regular gray-scale images and did not
control for other low-level confounds. This decision was
made in light of evidence suggesting that the dorsal path-
way may be sensitive to texture and shading cues (Freud
et al., 2016). Sin embargo, this makes it harder to rule out
other low-level confounds in the data. Finalmente, objects were
not matched for familiarity. This control may be especially
germane to the selection of tool exemplars given that
membership into this category is defined by a stereotypic
motor plan that is instantiated through experience with
tool-like objects. Future experiments may do a better job
controlling for the myriad confounds such as low-level fea-
turas, real-world size, and familiarity that lurk between
object categories. Sin embargo, controlling for every con-
found in a single study is often unrealistic, and all studies
must face trade-offs between controls and image integrity.
Still, some promising approaches exist that can help
researchers mitigate the impact of low-level confounds.
Por ejemplo, Weisberg et al. (2007) presented participants
with novel, artificially created objects. Over multiple ses-
siones, participants used these objects to perform specific
funciones, thereby facilitating a transition from object to
tool via the acquisition of a specific motor plan. Compar-
ing neural activity before and after training serves as a
control for low-level confounds and ensures that the
demarcation between tool and graspable object maps on
to the participant’s experience.
Conclusión
A number of limitations that may affect the interpre-
tation of our data should be discussed. Primero, source
En resumen, our study used MVPA on HD-EEG time series
data in conjunction with source localization to explore the
Gurariy et al.
981
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
spatio-temporal dynamics of object processing in the
cerebro humano. Our results suggest the following: (1) Suc-
cessful classification of object categories was observed
across different levels of abstraction, including exemplar,
basic (bird, insect, tool, graspable object), and superordi-
nate (animacy); (2) shape and category information are
both represented throughout the ventral and dorsal path-
maneras; (3) successful classification of object categories in
the dorsal pathway was not restricted to inanimate catego-
ries (tools and graspable nontools) but also included
animate categories (birds and insects); y (4) en general,
prominent similarities were observed between the two
pathways, both with regard to temporal dynamics as well
as the neural representations decoded by the MVPA
classifier.
Understanding the neural basis of object processing
remains an important challenge. The methodological
approach used in this study offers a promising avenue
for better understanding the neural dynamics of object
perception via the combination of spatial and temporal
analiza. Además, the successful decoding of ani-
mate object categories in the dorsal pathway as well as
the general similarities observed between the two path-
ways raises intriguing questions regarding the role that
the dorsal stream plays in visual perception. A pesar de
these findings do not undermine the substantial body of
work that shows that, broadly speaking, each pathway is
specialized toward differing behavioral goals, the notion
of functional independence must be reevaluated. Future
work is needed to further elucidate the computational
processes of the dorsal pathway and how these processes
modulate visual object perception.
Reprint requests should be sent to Gennadiy Gurariy, Biomed-
ical Engineering, Medical College of Wisconsin, o por correo electrónico:
ggurariy@mcw.edu.
Contribuciones de autor
Gennadiy Gurariy: Conceptualización; Curación de datos;
Análisis formal; Metodología; Visualización; Writing—
Original draft; Writing—Review & edición. Ryan E. B.
Mruczek: Conceptualización; Metodología; Writing—
Revisar & edición. Jacqueline C. Snow: Conceptualización;
Metodología; Writing—Review & edición. Gideon P.
Caplovitz: Conceptualización; Adquisición de financiación;
Metodología; Administración de proyecto; Recursos; Supervi-
sión; Writing—Review & edición.
Información de financiación
Fundación Nacional de Ciencia (NSF) (https://dx.doi.org/10
.13039/100000001), conceder número: EPSCoR Research
Infrastructure Awards 1632849 (GRAMO. PAG. C. & j. S.) y
1632738 (GRAMO. PAG. C.); National Institute of General Medical
Sciences of the National Institutes of Health (NIH), grant
number: P20 GM103650; National Eye Institute of Health,
award number: R01EY026701 ( j. S.).The content is solely
the responsibility of the authors and does not necessarily
represent the official views of the NIH or the NSF.
Diversidad en las prácticas de citas
Retrospective analysis of the citations in every article
published in this journal from 2010 a 2021 reveals a
persistent pattern of gender imbalance: Aunque el
proportions of authorship teams (categorized by esti-
mated gender identification of first author/last author)
publishing in the Journal of Cognitive Neuroscience ( JoCN)
during this period were M(un)/m = .407, W.(Omán)/m =
.32, M/M = .115, and W/ W = .159, the comparable pro-
portions for the articles that these authorship teams cited
were M/M = .549, W/M = .257, M/M = .109, and W/ W =
.085 (Postle and Fulvio, JoCN, 34:1, páginas. 1–3). Consecuencia-
frecuentemente, JoCN encourages all authors to consider gender
balance explicitly when selecting which articles to cite
and gives them the opportunity to report their article’s
gender citation balance.
REFERENCIAS
Alizadeh, A. METRO., van Dromme, I., Verhoef, B. MI., & Janssen, PAG.
(2018). Caudal intraparietal sulcus and three-dimensional
visión: A combined functional magnetic resonance imaging
and single-cell study. Neuroimagen, 166, 46–59. https://doi
.org/10.1016/j.neuroimage.2017.10.045, PubMed: 29080712
Almeida, J., Mahon, B. Z., & Caramazza, A. (2010). El rol de
the dorsal visual processing stream in tool identification.
ciencia psicológica, 21, 772–778. https://doi.org/10.1177
/0956797610371343, PubMed: 20483820
Almeida, J., Mahon, B. Z., Nakayama, K., & Caramazza, A. (2008).
Unconscious processing dissociates along categorical lines.
procedimientos de la Academia Nacional de Ciencias, EE.UU.,
105, 15214–15218. https://doi.org/10.1073/pnas.0805867105,
PubMed: 18809923
Andrews, t. J., watson, D. METRO., Rice, GRAMO. MI., & Hartley, t. (2015).
Low-level properties of natural images predict topographic
patterns of neural response in the ventral visual pathway.
Revista de visión, 15, 3. https://doi.org/10.1167/15.7.3,
PubMed: 26024512
Astafiev, S. v., Stanley, C. METRO., Shulman, GRAMO. l., & Corbetta, METRO.
(2004). Extrastriate body area in human occipital cortex
responds to the performance of motor actions. Naturaleza
Neurociencia, 7, 542–548. https://doi.org/10.1038/nn1241,
PubMed: 15107859
Bae, G.-Y., & Luck, S. j. (2018). Dissociable decoding of spatial
attention and working memory from EEG oscillations and
sustained potentials. Revista de neurociencia, 38, 409–422.
https://doi.org/10.1523/JNEUROSCI.2860-17.2017, PubMed:
29167407
Baldassi, C., Alemi-Neissi, A., Pagan, METRO., DiCarlo, j. J., Zecchina, r.,
& Zoccolan, D. (2013). Shape similarity, better than semantic
membership, accounts for the structure of visual object
representations in a population of monkey inferotemporal
neuronas. Biología Computacional PLoS, 9, e1003167. https://
doi.org/10.1371/journal.pcbi.1003167, PubMed: 23950700
Booth, METRO. C. A., & Rolls, mi. t. (1998). View-invariant
representations of familiar objects by neurons in the inferior
temporal visual cortex. Corteza cerebral, 8, 510–523. https://
doi.org/10.1093/cercor/8.6.510, PubMed: 9758214
982
Revista de neurociencia cognitiva
Volumen 34, Número 6
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Bracci, S., & Op de Beeck, h. (2016). Dissociations and
associations between shape and category representations
in the two visual pathways. Revista de neurociencia, 36,
432–444. https://doi.org/10.1523/JNEUROSCI.2314-15.2016,
PubMed: 26758835
Brainard, D. h. (1997). The Psychophysics toolbox. Visión espacial,
10, 433–436. https://doi.org/10.1163/156856897X00357,
PubMed: 9176952
Carlson, T., Tovar, D. A., Alink, A., & Kriegeskorte, norte. (2013).
Representational dynamics of object vision: The first
1000 EM. Revista de visión, 13, 1–19. https://doi.org/10.1167
/13.10.1, PubMed: 23908380
chao, l. l., & Martín, A. (2000). Representation of manipulable
man-made objects in the dorsal stream. Neuroimagen, 12,
478–484. https://doi.org/10.1006/nimg.2000.0635, PubMed:
10988041
Chen, J., Snow, j. C., Culham, j. C., & Goodale, METRO. A. (2018).
What role does “elongation” play in “tool-specific” activation
and connectivity in the dorsal and ventral visual streams?
Corteza cerebral, 28, 1117–1131. https://doi.org/10.1093
/cercor/bhx017, PubMed: 28334063
Cichy, R. METRO., Pantazis, D., & Oliva, A. (2014). Resolving human
object recognition in space and time. Neurociencia de la naturaleza,
17, 455–462. https://doi.org/10.1038/nn.3635, PubMed:
24464044
Cichy, R. METRO., Pantazis, D., & Oliva, A. (2016). Similarity-based
fusion of MEG and fmri reveals spatio-temporal dynamics
in human cortex during visual object recognition. Cerebral
Corteza, 26, 3563–3579. https://doi.org/10.1093/cercor
/bhw135, PubMed: 27235099
Cloutman, l. l. (2013). Interaction between dorsal and ventral
processing streams: Where, when and how? Brain and
Idioma, 127, 251–263. https://doi.org/10.1016/j.bandl.2012
.08.003, PubMed: 22968092
cohen, l., & Dehaene, S. (2004). Specialization within the
ventral stream: The case for the visual word form area.
Neuroimagen, 22, 466–476. https://doi.org/10.1016/j
.neuroimage.2003.12.049, PubMed: 15110040
collins, MI., Freud, MI., Kainerstorfer, j. METRO., Cao, J., &
behrman, METRO. (2019). Temporal dynamics of shape
processing differentiate contributions of dorsal and ventral
visual pathways. Revista de neurociencia cognitiva, 31,
821–836. https://doi.org/10.1162/jocn_a_01391, PubMed:
30883289
Contini, mi. w., Wardle, S. GRAMO., & Carlson, t. A. (2017). Decoding
the time-course of object recognition in the human brain:
From visual features to categorical decisions. Neuropsicología,
105, 165–176. https://doi.org/10.1016/j.neuropsychologia
.2017.02.013, PubMed: 28215698
Craighero, l., Bello, A., Fadiga, l., & Rizzolatti, GRAMO. (2002). Hand
action preparation influences the responses to hand pictures.
Neuropsicología, 40, 492–502. https://doi.org/10.1016
/S0028-3932(01)00134-8, PubMed: 11749979
Craighero, l., Fadiga, l., Umiltà, C. A., & Rizzolatti, GRAMO. (1996).
Evidence for visuomotor priming effect. NeuroReport, 8,
347–349. https://doi.org/10.1097/00001756-199612200-00068,
PubMed: 9051808
Craighero, l., Fadiga, l., Rizzolatti, GRAMO., & Umilta, C. (1998).
Visuomotor priming. Visual Cognition, 5, 109–125. https://
doi.org/10.1080/713756780
Crouzet, S. METRO., Kirchner, h., & Thorpe, S. j. (2010). Fast
saccades toward faces: Face detection in just 100 EM. Diario
of Vision, 10, 16.1–16.17. https://doi.org/10.1167/10.4.16,
PubMed: 20465335
de Haan, mi. h. F., & Cowey, A. (2011). On the usefulness of
‘what’ and ‘where’ pathways in vision. Tendencias en Cognitivo
Ciencias, 15, 460–466. https://doi.org/10.1016/j.tics.2011.08
.005, PubMed: 21906989
Dell’acqua, r., & Job, R. (1998). Is object recognition automatic?
Boletín psiconómico & Revisar, 5, 496–503. https://doi.org
/10.3758/BF03208828
Desikan, R. S., Ségonne, F., pescado, B., quinn, B. T., Dickerson,
B. C., Blacker, D., et al. (2006). An automated labeling system
for subdividing the human cerebral cortex on MRI scans into
gyral based regions of interest. Neuroimagen, 31, 968–980.
https://doi.org/10.1016/j.neuroimage.2006.01.021, PubMed:
16530430
Desimone, r., Albright, t. D., Bruto, C. GRAMO., & bruce, C. (1984).
Stimulus-selective properties of inferior temporal neurons
in the macaque. Revista de neurociencia, 357, 219–240.
https://doi.org/10.1523/JNEUROSCI.04-08-02051.1984,
PubMed: 6470767
Downing, PAG. MI., Jiang, y., Shuman, METRO., & Kanwisher, norte. (2001).
A cortical area selective for visual processing of the human
body. Ciencia, 293, 2470–2473. https://doi.org/10.1126
/science.1063414, PubMed: 11577239
Epstein, r., harris, A., Stanley, D., & Kanwisher, norte. (1999). El
parahippocampal place area: Reconocimiento, navigation, o
encoding? Neurona, 23, 115–125. https://doi.org/10.1016
/S0896-6273(00)80758-8, PubMed: 10402198
Epstein, r., & Kanwisher, norte. (1998). A cortical representation
the local visual environment. Naturaleza, 392, 598–601. https://
doi.org/10.1038/33402, PubMed: 9560155
Erlikhman, GRAMO., & Caplovitz, GRAMO. PAG. (2017). Decoding information
about dynamically occluded objects in visual cortex.
Neuroimagen, 146, 778–788. https://doi.org/10.1016/j
.neuroimage.2016.09.024, PubMed: 27663987
Erlikhman, GRAMO., Caplovitz, GRAMO. PAG., Gurariy, GRAMO., Medina, J., & Snow,
j. C. (2018). Towards a unified perspective of object
shape and motion processing in human dorsal cortex.
Consciousness and Cognition, 64, 106–120. https://doi.org
/10.1016/j.concog.2018.04.016, PubMed: 29779844
Erlikhman, GRAMO., Gurariy, GRAMO., Mruczek, R. mi. B., & Caplovitz, GRAMO. PAG.
(2016). The neural representation of objects formed through
the spatiotemporal integration of visual transients. Neuroimagen,
142, 67–78. https://doi.org/10.1016/j.neuroimage.2016.03.044,
PubMed: 27033688
Faillenot, I., Decety, J., & Jeannerod, METRO. (1999). Human brain
activity related to the perception of spatial features of objects.
Neuroimagen, 10, 114–124. https://doi.org/10.1006/nimg.1999
.0449, PubMed: 10498441
Fang, F., & Él, S. (2005). Cortical responses to invisible objects in
the human dorsal and ventral pathways. Neurociencia de la naturaleza,
8, 1380–1385. https://doi.org/10.1038/nn1537, PubMed:
16136038
Freud, MI., behrman, METRO., & Snow, j. C. (2020). What does
dorsal cortex contribute to perception? Mente abierta, 4, 40–56.
https://doi.org/10.1162/opmi_a_00033, PubMed: 33225195
Freud, MI., Ganel, T., Shelef, I., Hammer, METRO. D., Avaro, GRAMO., &
behrman, METRO. (2017). Three-dimensional representations of
objects in dorsal cortex are dissociable from those in ventral
corteza. Corteza cerebral, 27, 422–434. https://doi.org/10.1093
/cercor/bhv229, PubMed: 26483400
Freud, MI., Plaut, D. C., & behrman, METRO. (2016). ‘What’ is
happening in the dorsal visual pathway. Tendencias en Cognitivo
Ciencias, 20, 773–784. https://doi.org/10.1016/j.tics.2016.08
.003, PubMed: 27615805
Frey, S. h. (2007). What puts the how in where? Tool use and
the divided visual streams hypothesis. Corteza, 43, 368–375.
https://doi.org/10.1016/S0010-9452(08)70462-3, PubMed:
17533760
Gallivan, j. PAG., Chapman, C. S., Mclean, D. A., Flanagan, j. r., &
Culham, j. C. (2013). Activity patterns in the category-selective
occipitotemporal cortex predict upcoming motor actions.
European Journal of Neuroscience, 38, 2408–2424. https://
doi.org/10.1111/ejn.12215, PubMed: 23581683
Gurariy et al.
983
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Garcea, F. MI., & Mahon, B. z. (2014). Parcellation of left
parietal tool representations by functional connectivity.
Neuropsicología, 60, 131–143. https://doi.org/10.1016/j
.neuropsychologia.2014.05.018, PubMed: 24892224
Gentilucci, METRO. (2002). Object motor representation and
reaching–grasping control. Neuropsicología, 40, 1139–1153.
https://doi.org/10.1016/S0028-3932(01)00233-0, PubMed:
11931918
Georgieva, S., Peeters, r., Kolster, h., Todd, j. T., & Orban, GRAMO. A.
(2009). The processing of three-dimensional shape from
disparity in the human brain. Revista de neurociencia, 29,
727–742. https://doi.org/10.1523/JNEUROSCI.4753-08.2009,
PubMed: 19158299
Gibson, j. j. (2014). The ecological approach to visual
percepción (1st ed., pag. 346). Prensa de Psicología. https://doi.org
/10.4324/9781315740218
Gillebert, C. r., Schaeverbeke, J., Bastin, C., Neyens, v., Bruffaerts,
r., De Weer, A.-S., et al. (2015). 3D shape perception in
posterior cortical atrophy: A visual neuroscience
perspectiva. Revista de neurociencia, 35, 12673–12692.
https://doi.org/10.1523/JNEUROSCI.3651-14.2015, PubMed:
26377458
encoding. Más uno, 11, e0167022. https://doi.org/10.1371
/diario.pone.0167022, PubMed: 27902738
Haak, k. v., & beckman, C. F. (2018). Objective analysis of
the topological organization of the human cortical visual
connectome suggests three visual pathways. Corteza, 98, 73–83.
https://doi.org/10.1016/j.cortex.2017.03.020, PubMed: 28457575
Haxby, j. v., Gobbini, METRO. I., Furey, METRO. l., Ishai, A., Schouten, j. l., &
Pietrini, PAG. (2001). Distributed and overlapping representations
of faces and objects in ventral temporal cortex. Ciencia, 293,
2425–2430. https://doi.org/10.1126/science.1063736, PubMed:
11577229
Hermsdörfer, J., Terlinden, GRAMO., Mühlau, METRO., Goldenberg, GRAMO., &
Wohlschläger, A. METRO. (2007). Neural representations of
pantomimed and actual tool use: Evidence from an
event-related fMRI study. Neuroimagen, 36, T109–T118.
https://doi.org/10.1016/j.neuroimage.2007.03.037, PubMed:
17499158
holmes, C. J., Hoge, r., collins, l., Woods, r., Toga, A. w., &
evans, A. C. (1998). Enhancement of MR images using
registration for signal averaging. Journal of Computer
Assisted Tomography, 22, 324–333. https://doi.org/10.1097
/00004728-199803000-00032, PubMed: 9530404
Gramfort, A., Papadopoulo, T., Olivi, MI., & Clerc, METRO. (2010).
hong, h., Yamins, D. l. K., Majaj, norte. J., & Dicarlo, j. j. (2016).
OpenMEEG: Opensource software for quasistatic
bioelectromagnetics. Biomedical Engineering Online, 45.
https://doi.org/10.1186/1475-925X-9-45, PubMed: 20819204
Grèzes, J., Tucker, METRO., Armony, J., Ellis, r., & Passingham, R. mi.
(2003). Objects automatically potentiate action: An fMRI
study of implicit processing. European Journal of
Neurociencia, 17, 2735–2740. https://doi.org/10.1046/j.1460
-9568.2003.02695.X, PubMed: 12823480
Grill-Spector, K., Kushnir, T., Edelman, S., Avaro, GRAMO., Itzchak,
y., & Malach, R. (1999). Differential processing of objects
under various viewing conditions in the human lateral
occipital complex. Neurona, 24, 187–203. https://doi.org/10
.1016/S0896-6273(00)80832-6, PubMed: 10677037
Grill-Spector, K., Kushnir, T., Hendler, T., Edelman, S., Itzchak,
y., & Malach, R. (1998). A sequence of object-processing
stages revealed by fMRI in the human occipital lobe. Humano
Mapeo cerebral, 6, 316–328. https://doi.org/10.1002/(CIENCIA)
1097-0193(1998)6:4<316::AID-HBM9>3.0.CO;2-6, PubMed:
9704268
Grill-Spector, K., & Weiner, k. S. (2014). The functional
architecture of the ventral temporal cortex and its role in
categorization. Naturaleza Reseñas Neurociencia, 15, 536–548.
https://doi.org/10.1038/nrn3747, PubMed: 24962370
Grootswagers, T., robinson, A. K., & Carlson, t. A. (2019).
The representational dynamics of visual objects in rapid
serial visual processing streams. Neuroimagen, 188, 668–679.
https://doi.org/10.1016/j.neuroimage.2018.12.046, PubMed:
30593903
Grootswagers, T., robinson, A. K., Shatek, S. METRO., & Carlson,
t. A. (2019). Untangling featural and conceptual object
representaciones. Neuroimagen, 202, 116083. https://doi.org/10
.1016/j.neuroimage.2019.116083, PubMed: 30593903
Grootswagers, T., Wardle, S. GRAMO., & Carlson, t. A. (2017).
Decoding dynamic brain patterns from evoked responses: A
tutorial on multivariate pattern analysis applied to time series
neuroimaging data. Revista de neurociencia cognitiva, 29,
677–697. https://doi.org/10.1162/jocn_a_01068, PubMed:
27779910
Bruto, C. GRAMO., Rocha-Miranda, C. MI., & Bender, D. B. (1972).
Visual properties of neurons in inferotemporal cortex of the
macaque. Revista de neurofisiología, 35, 96–111. https://
doi.org/10.1152/jn.1972.35.1.96, PubMed: 4621506
Gurariy, GRAMO., Killebrew, k. w., Berryhill, METRO. MI., & Caplovitz, GRAMO. PAG.
(2016). Induced and evoked human electrophysiological
correlates of visual working memory set-size effects at
Explicit information for category-orthogonal object
properties increases along the ventral stream. Naturaleza
Neurociencia, 19, 613–622. https://doi.org/10.1038/nn.4247,
PubMed: 26900926
Hung, C. PAG., Kreiman, GRAMO., Pogio, T., & DiCarlo, j. j. (2005). Fast
readout of object identity from macaque inferior temporal
corteza. Ciencia, 310, 863–866. https://doi.org/10.1126/science
.1117593, PubMed: 16272124
hutchison, R. METRO., & Gallivan, j. PAG. (2018). Functional coupling
between frontoparietal and occipitotemporal pathways
during action and perception. Corteza, 98, 8–27. https://doi
.org/10.1016/j.cortex.2016.10.020, PubMed: 27890325
Ishai, A., Ungerleider, l. GRAMO., Martín, A., Schouten, j. l., & Haxby,
j. V. (1999). Distributed representation of objects in the
human ventral visual pathway. Actas del Nacional
Academia de Ciencias, EE.UU., 96, 9379–9384. https://doi.org
/10.1073/pnas.96.16.9379, PubMed: 10430951
Janssen, PAG., Srivastava, S., Ombelet, S., & Orban, GRAMO. A. (2008).
Coding of shape and position in macaque lateral
intraparietal area. Revista de neurociencia, 28, 6679–6690.
https://doi.org/10.1523/JNEUROSCI.0499-08.2008, PubMed:
18579742
Janssen, PAG., Verhoef, B. MI., & Premereur, mi. (2018). Funcional
interactions between the macaque dorsal and ventral visual
pathways during three-dimensional object vision. Corteza,
98, 218–227. https://doi.org/10.1016/j.cortex.2017.01.021,
PubMed: 28258716
jeong, S. K., & Xu, Y. (2016). Behaviorally relevant abstract
object identity representation in the human parietal cortex.
Revista de neurociencia, 36, 1607–1619. https://doi.org/10
.1523/JNEUROSCI.1016-15.2016, PubMed: 26843642
Kaneshiro, B., Perreau Guimaraes, METRO., kim, H.-S., Norcia, A. METRO.,
& Suppes, PAG. (2015). A representational similarity analysis of
the dynamics of object processing using single-trial EEG
clasificación. Más uno, 10, e0135697. https://doi.org/10.1371
/diario.pone.0135697, PubMed: 26295970
Kanwisher, NORTE., McDermott, J., & Chun, METRO. METRO. (1997). El
área de la cara fusiforme: A module in human extrastriate cortex
specialized for face perception. Revista de neurociencia, 17,
4302–4311. https://doi.org/10.1523/JNEUROSCI.17-11-04302
.1997, PubMed: 9151747
Katsuyama, NORTE., Usui, NORTE., Nose, I., & Taira, METRO. (2011). Percepción
of object motion in three-dimensional space induced by cast
shadows. Neuroimagen, 54, 485–494. https://doi.org/10.1016/j
.neuroimage.2010.07.075, PubMed: 20692350
984
Revista de neurociencia cognitiva
Volumen 34, Número 6
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Kiani, r., Esteky, h., Mirpour, K., & Tanaka, k. (2007). Object
Lestou, v., Justicia, j. METRO. l., Humphreys, K., Kourtzi, Z., &
category structure in response patterns of neuronal
population in monkey inferior temporal cortex. Diario de
Neurofisiología, 97, 4296–4309. https://doi.org/10.1152/jn
.00024.2007, PubMed: 17428910
Killebrew, k. w., Gurariy, GRAMO., Peacock, C. MI., Berryhill, METRO. MI., &
Caplovitz, GRAMO. PAG. (2018). Electrophysiological correlates of
encoding processes in a full-report visual working memory
paradigma. Cognitivo, Afectivo & Neurociencia conductual,
18, 353–365. https://doi.org/10.3758/s13415-018-0574-8,
PubMed: 29446044
Rey, j. r., & Dehaene, S. (2014). Characterizing the
dynamics of mental representations: The temporal
generalization method. Tendencias en Ciencias Cognitivas, 18,
203–210. https://doi.org/10.1016/j.tics.2014.01.002, PubMed:
24593982
Kirchner, h., & Thorpe, S. j. (2006). Ultra-rapid object detection
with saccadic eye movements: Visual processing speed
revisited. Vision Research, 46, 1762–1776. https://doi.org/10
.1016/j.visres.2005.10.002, PubMed: 16289663
Kleiner, METRO., Brainard, D. h., Pelli, D. GRAMO., Broussard, C., Lobo, T.,
& Niehorster, D. (2007). What’s new in Psychtoolbox-3?
Percepción, 36, 1-dieciséis.
Konen, C. S., & Kastner, S. (2008). Two hierarchically organized
neural systems for object information in human visual cortex.
Neurociencia de la naturaleza, 11, 224–231. https://doi.org/10.1038
/nn2036, PubMed: 18193041
Konkle, T., & Oliva, A. (2012). A familiar-size Stroop effect:
Real-world size is an automatic property of object
representación. Revista de Psicología Experimental:
Percepción y desempeño humanos, 38, 561–569. https://
doi.org/10.1037/a0028294, PubMed: 22545601
Kourtzi, Z., & Kanwisher, norte. (2000). Cortical regions involved in
perceiving object shape. Revista de neurociencia, 20,
3310–3318. https://doi.org/10.1523/JNEUROSCI.20-09-03310
.2000, PubMed: 10777794
Kravitz, D. J., Saleem, k. S., Panadero, C. I., Ungerleider, l. GRAMO., &
Mishkin, METRO. (2013). The ventral visual pathway: An expanded
neural framework for the processing of object quality. Trends
in Cognitive Sciences, 17, 26–49. https://doi.org/10.1016/j.tics
.2012.10.011, PubMed: 23265839
Kriegeskorte, norte. (2011). Pattern-information analysis: De
stimulus decoding to computational-model testing. Neuroimagen,
56, 411–421. https://doi.org/10.1016/j.neuroimage.2011.01
.061, PubMed: 21281719
Kriegeskorte, NORTE., Goebel, r., & Bandettini, PAG. (2006).
Information-based functional brain mapping. Actas
of the National Academy of Sciences, EE.UU., 103, 3863–3868.
https://doi.org/10.1073/pnas.0600244103, PubMed: 16537458
Kriegeskorte, NORTE., Mur, METRO., Fallar, D. A., Kiani, r., Bodurka, J.,
Esteky, h., et al. (2008). Matching categorical object
representations in inferior temporal cortex of man and
monkey. Neurona, 60, 1126–1141. https://doi.org/10.1016/j
.neuron.2008.10.043, PubMed: 19109916
Kybic, J., Clerc, METRO., Abboud, T., Faugeras, o., Keriven, r., &
Papadopoulo, t. (2005). A common formalism for the
integral formulations of the forward EEG problem. IEEE
Transactions on Medical Imaging, 24, 12–28. https://doi.org
/10.1109/TMI.2004.837363, PubMed: 15638183
Lange, NORTE., Strother, S. C., anderson, j. r., Nielsen, F. Å., holmes,
A. PAG., Kolenda, T., et al. (1999). Plurality and resemblance in
fMRI data analysis. Neuroimagen, 10, 282–303. https://doi
.org/10.1006/nimg.1999.0472, PubMed: 10458943
Humphreys, GRAMO. W.. (2014). A dorsal visual route necessary for
global form perception: Evidence from neuropsychological
resonancia magnética funcional. Revista de neurociencia cognitiva, 26, 621–634.
https://doi.org/10.1162/jocn_a_00489, PubMed: 24047377
Luis, j. W.. (2006). Cortical networks related to human use of
herramientas. neurocientífico, 12, 211–231. https://doi.org/10.1177
/1073858406288327, PubMed: 16684967
Largo, B., Yu, C.-P., & Konkle, t. (2018). Mid-level visual features
underlie the high-level categorical organization of the ventral
stream. procedimientos de la Academia Nacional de Ciencias,
115, E9015–E9024. https://doi.org/10.1073/pnas.1719616115,
PubMed: 30171168
macdonald, S. NORTE., & Culham, j. C. (2015). Do human brain areas
involved in visuomotor actions show a preference for real
tools over visually similar non-tools? Neuropsicología, 77,
35–41. https://doi.org/10.1016/j.neuropsychologia.2015.08
.004, PubMed: 26253009
Mack, METRO. l., & Palmeri, t. j. (2011). The timing of visual object
categorization. Fronteras en psicología, 2, 165. https://doi
.org/10.3389/fpsyg.2011.00165, PubMed: 21811480
Mahon, B. Z., Milleville, S. C., Negri, GRAMO. A. l., Rumiati, R. I.,
Caramazza, A., & Martín, A. (2007). Action-related properties
shape object representations in the ventral stream. Neurona,
55, 507–520. https://doi.org/10.1016/j.neuron.2007.07.011,
PubMed: 17678861
Malach, r., Reppas, j. B., Benson, R. r., Kwong, k. K., Jiang, h.,
Kennedy, W.. A., et al. (1995). Object-related activity revealed
by functional magnetic resonance imaging in human occipital
corteza. procedimientos de la Academia Nacional de Ciencias,
EE.UU., 92, 8135–8139. https://doi.org/10.1073/pnas.92.18
.8135, PubMed: 7667258
Martín, A. (2007). The representation of object concepts in
the brain. Annual Review of Psychology, 58, 25–45. https://
doi.org/10.1146/annurev.psych.57.102904.190143, PubMed:
16968210
Martín, A., & Weisberg, j. (2003). Neural foundations for
understanding social and mechanical concepts. Cognitivo
Neuropsychology, 20, 575–587. https://doi.org/10.1080
/02643290342000005, PubMed: 16648880
Martín, A. B., Cual, X., Saalmann, Y. B., Wang, l., Shestyuk, A.,
lin, j. J., et al. (2019). Temporal dynamics and response
modulation across the human visual system in a spatial
attention task: An ECoG study. Revista de neurociencia, 39,
333–352. https://doi.org/10.1523/JNEUROSCI.1889-18.2018,
PubMed: 30459219
MathWorks Inc. (2007). MATLAB data analysis. MATLAB.
McCarthy, j. D., Kohler, PAG. J., Tse, PAG. Ud., & Caplovitz, GRAMO. PAG.
(2015). Extrastriate visual areas integrate form features over
space and time to construct representations of stationary and
rigidly rotating objects. Revista de neurociencia cognitiva,
27, 2158–2173. https://doi.org/10.1162/jocn_a_00850,
PubMed: 26226075
Mruczek, R. mi. B., von Loga, I. S., & Kastner, S. (2013). El
representation of tool and non-tool object information in the
human intraparietal sulcus. Revista de neurofisiología, 109,
2883–2896. https://doi.org/10.1152/jn.00658.2012, PubMed:
23536716
Nasr, S., Echavarria, C. MI., & Tootell, R. B. h. (2014). Thinking
outside the box: Rectilinear shapes selectively activate
scene-selective cortex. Revista de neurociencia, 34,
6721–6735. https://doi.org/10.1523/JNEUROSCI.4802-13
.2014, PubMed: 24828628
Leahy, R. METRO., Mosher, j. C., Spencer, METRO. MI., Huang, METRO. X., &
Norman, j. (2002). Two visual systems and two theories of
Lewine, j. D. (1998). A study of dipole localization accuracy
for MEG and EEG using a human skull phantom.
Neuroimagen, 7, S676. https://doi.org/10.1016/S1053-8119
(18)31509-X
percepción: An attempt to reconcile the constructivist and
ecological approaches. Ciencias del comportamiento y del cerebro,
25, 73–96. https://doi.org/10.1017/S0140525X0200002X,
PubMed: 12625088
Gurariy et al.
985
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Oostenveld, r., Fries, PAG., Maris, MI., & Schoffelen, j. METRO. (2011).
FieldTrip: Open source software for advanced analysis
of MEG, EEG, and invasive electrophysiological data.
Computational Intelligence and Neuroscience, 2011, 156869.
https://doi.org/10.1155/2011/156869, PubMed: 21253357
Oosterhof, norte. NORTE., Connolly, A. C., & Haxby, j. V. (2016).
CoSMoMVPA: Multi-modal multivariate pattern analysis of
neuroimaging data in MATLAB/GNU octave. Fronteras en
Neuroinformatics. https://doi.org/10.3389/fninf.2016.00027,
PubMed: 27499741
O’Toole, A. J., Jiang, F., Abdi, h., & Haxby, j. V. (2005). Partially
distributed representations of objects and faces in ventral
temporal cortex. Revista de neurociencia cognitiva, 17,
580–590. https://doi.org/10.1162/0898929053467550,
PubMed: 15829079
Pelli, D. GRAMO. (1997). The VideoToolbox software for visual
psychophysics: Transforming numbers into movies. Espacial
Vision, 10, 437–442. https://doi.org/10.1163/156856897X00366,
PubMed: 9176953
Perrin, F., Pernier, J., Bertrand, o., & Echallier, j. F. (1989).
Spherical splines for scalp potential and current density
mapping. Electroencephalography and Clinical
Neurofisiología, 72, 184–187. https://doi.org/10.1016/0013
-4694(89)90180-6
Peterson, D. J., Gurariy, GRAMO., Dimotsantos, GRAMO. GRAMO., Arciniega, h.,
Berryhill, METRO. MI., & Caplovitz, GRAMO. PAG. (2014). The steady-state
visual evoked potential reveals neural correlates of the items
encoded into visual working memory. Neuropsicología, 63,
145–153. https://doi.org/10.1016/j.neuropsychologia.2014.08
.020, PubMed: 25173712
Philiastides, METRO. GRAMO., & Sajda, PAG. (2006). Temporal characterization
of the neural correlates of perceptual decision making in the
cerebro humano. Corteza cerebral, 16, 509–518. https://doi.org/10
.1093/cercor/bhi130, PubMed: 16014865
Potter, METRO. C., & Exacción, mi. I. (1969). Recognition memory for
a rapid sequence of pictures. Revista de experimentación
Psicología, 81, 10-15. https://doi.org/10.1037/h0027470,
PubMed: 5812164
Rajimehr, r., Devaney, k. J., Bilenko, norte. y., Joven, j. C., &
Tootell, R. B. h. (2011). The “parahippocampal place area”
responds preferentially to high spatial frequencies in humans
and monkeys. Biología PLoS, 9, e1000608. https://doi.org/10
.1371/journal.pbio.1000608, PubMed: 21483719
Rice, GRAMO. MI., watson, D. METRO., Hartley, T., & Andrews, t. j. (2014).
Low-level image properties of visual objects predict patterns
of neural response across category-selective regions of the
ventral visual pathway. Revista de neurociencia, 34,
8837–8844. https://doi.org/10.1523/JNEUROSCI.5265-13
.2014, PubMed: 24966383
robinson, A. K., Grootswagers, T., & Carlson, t. A. (2019). El
influence of image masking on object representations during
rapid serial visual presentation. Neuroimagen, 197, 224–231.
https://doi.org/10.1016/j.neuroimage.2019.04.050, PubMed:
31009746
Sakata, h., Taira, METRO., Kusunoki, METRO., Murata, A., Tanaka, y., &
Tsutsui, k. I. (1998). Neural coding of 3D features of objects
for hand action in the parietal cortex of the monkey.
Philosophical Transactions of the Royal Society of London,
Serie B, Ciencias Biologicas, 353, 1363–1373. https://doi.org
/10.1098/rstb.1998.0290, PubMed: 9770229
Sakuraba, S., Sakai, S., Yamanaka, METRO., Yokosawa, K., &
Hirayama, k. (2012). Does the human dorsal stream really
process a category for tools? Revista de neurociencia, 32,
3949–3953. https://doi.org/10.1523/JNEUROSCI.3973-11
.2012, PubMed: 22423115
Sawamura, h., Georgieva, S., Vogels, r., Vanduffel, w., & Orban,
GRAMO. A. (2005). Using functional magnetic resonance imaging to
assess adaptation and size invariance of shape processing
by humans and monkeys. Revista de neurociencia, 25,
4294–4306. https://doi.org/10.1523/JNEUROSCI.0377-05
.2005, PubMed: 15858056
Schenk, T., & McIntosh, R. D. (2010). Do we have independent
visual streams for perception and action? Cognitivo
Neurociencia, 1, 52–62. https://doi.org/10.1080
/17588920903388950, PubMed: 24168245
Sereno, A. B., & Maunsell, j. h. R. (1998). Shape selectivity in
primate lateral intraparietal cortex. Naturaleza, 395, 500–503.
https://doi.org/10.1038/26752, PubMed: 9774105
Shmuelof, l., & Zohary, mi. (2005). Dissociation between ventral
and dorsal fMRI activation during object and action
recognition. Neurona, 47, 457–470. https://doi.org/10.1016/j
.neuron.2005.06.034, PubMed: 16055068
Sim, mi. J., Helbig, h. B., Graf, METRO., & Kiefer, METRO. (2015). Cuando
action observation facilitates visual perception: Activation in
visuo-motor areas contributes to object recognition. Cerebral
Corteza, 25, 2907–2918. https://doi.org/10.1093/cercor
/bhu087, PubMed: 24794918
Srivastava, S., Orban, GRAMO. A., De Mazière, PAG. A., & Janssen, PAG. (2009).
A distinct representation of three-dimensional shape in
macaque anterior intraparietal area: Fast, métrico, and coarse.
Revista de neurociencia, 29, 10613–10626. https://doi.org
/10.1523/JNEUROSCI.6016-08.2009, PubMed: 19710314
Rígido, A., & Zohary, mi. (2008). Parietal mapping of visuomotor
transformations during human tool grasping. Cerebral
Corteza, 18, 2358–2368. https://doi.org/10.1093/cercor
/bhm260, PubMed: 18252741
Tadel, F., Baillet, S., Mosher, j. C., Pantazis, D., & Leahy, R. METRO.
(2011). Brainstorm: A user-friendly application for MEG/EEG
análisis. Computational Intelligence and Neuroscience,
2011, 879716. https://doi.org/10.1155/2011/879716, PubMed:
21584256
Takemura, h., Rokem, A., Winawer, J., Yeatman, j. D., Wandell,
B. A., & Pestilli, F. (2016). A major human white matter
pathway between dorsal and ventral visual cortex. Cerebral
Corteza, 26, 2205–2214. https://doi.org/10.1093/cercor/bhv064,
PubMed: 25828567
Thorpe, S., Fize, D., & Marlot, C. (1996). Speed of processing in
the human visual system. Naturaleza, 381, 520–522. https://doi
.org/10.1038/381520a0, PubMed: 8632824
Tsutsui, k. I., Jiang, METRO., Yara, K., Sakata, h., & Taira, METRO. (2001).
Integration of perspective and disparity cues in surface-
orientation-selective neurons of area CIP. Diario de
Neurofisiología, 86, 2856–2867. https://doi.org/10.1152/jn
.2001.86.6.2856, PubMed: 11731542
Vaina, l. METRO. (1989). Selective impairment of visual motion
interpretation following lesions of the right occipito-parietal
area in humans. Cibernética biológica, 61, 347–359. https://
doi.org/10.1007/BF00200800, PubMed: 2790066
Valyear, k. F., Cavina-Pratesi, C., Stiglick, A. J., & Culham, j. C.
(2007). Does tool-related fMRI activity within the intraparietal
sulcus reflect the plan to grasp? Neuroimagen, 36(Suplemento. 2),
T94–T108. https://doi.org/10.1016/j.neuroimage.2007.03.031,
PubMed: 17499175
van de Nieuwenhuijzen, METRO. MI., Backus, A. r., Bahramisharif, A.,
Doeller, C. F., Jensen, o., & van Gerven, METRO. A. j. (2013).
MEG-based decoding of the spatiotemporal dynamics of
visual category perception. Neuroimagen, 83, 1063–1073.
https://doi.org/10.1016/j.neuroimage.2013.07.075, PubMed:
23927900
Van Dromme, I. C., Premereur, MI., Verhoef, B. MI., Vanduffel, w.,
& Janssen, PAG. (2016). Posterior parietal cortex drives
inferotemporal activations during three-dimensional object
visión. Biología PLoS, 14, e1002445. https://doi.org/10.1371
/journal.pbio.1002445, PubMed: 27082854
Vanduffel, w., Fize, D., Peuskens, h., Denys, K., Sunaert, S.,
Todd, j. T., et al. (2002). Extracting 3D from motion:
986
Revista de neurociencia cognitiva
Volumen 34, Número 6
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Differences in human and monkey intraparietal cortex.
Ciencia, 298, 413–415. https://doi.org/10.1126/science
.1073574, PubMed: 12376701
Vuilleumier, PAG., Henson, R. NORTE., Conductor, J., & Dolan, R. j. (2002).
Multiple levels of visual object constancy revealed by
event-related fMRI of repetition priming. Naturaleza
Neurociencia, 5, 491–499. https://doi.org/10.1038/nn839,
PubMed: 11967545
watson, D. METRO., Hartley, T., & Andrews, t. j. (2014). Patrones
of response to visual scenes are linked to the low-level
properties of the image. Neuroimagen, 99, 402–410. https://doi
.org/10.1016/j.neuroimage.2014.05.045, PubMed: 24862072
Weisberg, J., Van Turennout, METRO., & Martín, A. (2007). A neural
system for learning about object function. Corteza cerebral,
17, 513–521. https://doi.org/10.1093/cercor/bhj176, PubMed:
16581980
Willenbockel, v., Sadr, J., Fiset, D., Horne, GRAMO. o., Gosselin, F., &
Tanaka, j. W.. (2010). Controlling low-level image properties:
The SHINE toolbox. Behavior Research Methods, 42, 671–684.
https://doi.org/10.3758/BRM.42.3.671, PubMed: 20805589
Xu, y., & Chun, METRO. METRO. (2007). Visual grouping in human parietal
corteza. procedimientos de la Academia Nacional de Ciencias,
EE.UU., 104, 18766–18771. https://doi.org/10.1073/pnas
.0705618104, PubMed: 17998539
yue, X., Pourladian, I. S., Tootell, R. B. h., & Ungerleider, l. GRAMO.
(2014). Curvature-processing network in macaque visual
corteza. procedimientos de la Academia Nacional de Ciencias,
EE.UU., 13, E3467–E3475. https://doi.org/10.1073/pnas
.1412616111, PubMed: 25092328
Zachariou, v., Klatzky, r., & behrman, METRO. (2014). Ventral
and dorsal visual stream contributions to the perception of
object shape and object location. Revista de Cognitivo
Neurociencia, 26, 189–209. https://doi.org/10.1162/jocn_a
_00475, PubMed: 24001005
Zachariou, v., Nikas, C. v., Safiullah, z. NORTE., Gotts, S. J., &
Ungerleider, l. GRAMO. (2017). Spatial mechanisms within the
dorsal visual pathway contribute to the configural processing
of faces. Corteza cerebral, 27, 4124–4138. https://doi.org/10
.1093/cercor/bhw224, PubMed: 27522076
Zoccolan, D., Kouh, METRO., Pogio, T., & DiCarlo, j. j. (2007).
Trade-off between object selectivity and tolerance in monkey
inferotemporal cortex. Revista de neurociencia, 27,
12292–12307. https://doi.org/10.1523/JNEUROSCI.1897-07
.2007, PubMed: 17989294
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
/
3
4
6
9
6
7
2
0
1
9
2
8
2
/
j
oh
C
norte
_
a
_
0
1
8
4
5
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Gurariy et al.
987