El seguimiento de las secuencias de pulsaciones de teclas a nivel cortical revela
the Dynamics of Serial Order Production
Svetlana Pinet1,2, Gary S. Dell3, and F.-Xavier Alario1
Abstracto
■ Response selection is often studied by examining single re-
patas, although most actions are performed within an over-
arching sequence. Understanding processes that order and
execute items in a sequence is thus essential to give a complete
picture of response selection. en este estudio, we investigate re-
sponse selection by comparing single responses and response
sequences as well as unimanual and bimanual sequences. Nosotros
recorded EEG while participants were typing one- or two-
keystroke sequences. Irrespective of stimulus modality (visual
or auditory), response-locked analysis revealed distinct contra-
lateral and ipsilateral components previously associated with
activation and inhibition of alternative responses. Unimanual
sequences exhibited a similar activation/inhibition pattern as
single responses, but with the activation component of the
pattern expressed more strongly, reflecting the fact that the
hand will be used for two strokes. A diferencia de, bimanual se-
quences were associated with successive activation of each of
the corresponding motor cortices controlling each keystroke
and no traceable inhibitory component. En breve, the activation
component of the two-keystroke sequence EEG pattern can be
understood from the addition of activation components of
single-stroke sequences; the inhibition of the hand not being
used is only evidenced when that hand is not planned for the
next stroke. ■
INTRODUCCIÓN
Producing a behavioral sequence involves retrieving and
activating each sequence unit in an ordered and timely
moda, a requirement often referred to as the problem
of serial order (Lashley, 1951). An important theory of se-
rial order is expressed by competitive queuing (CQ)
modelos. These models have been successfully applied
in several cognitive domains, such as memory (p.ej., serial
recordar; Houghton, 1990) and language (p.ej., typing;
Rumelhart & Norman, 1982). CQ models propose that
all elements of a sequence are activated, such that there
is a gradient of activation consistent with the order of
items in the sequence. Por ejemplo, the gradient may
be set up by each item inhibiting all subsequent items,
and the sequence may be executed by selecting the cur-
rently most activated item, and then inhibiting it, de este modo
leading to the next item in the queue possessing the
most activation. Entonces, a balance of activation and inhi-
bition of items leads to the correct production of the
planned sequence.
The CQ model’s activation and inhibition processes
constitute an algorithmic (Marr, 1982) cuenta, one that
makes no claims about physiological implementation. A
significant step toward such an implementation was taken
by Averbeck, Chafee, Crowe, and Georgopoulos (2002),
who showed that pFC of behaving monkeys displayed a
gradient of activation of future items, with their degree of
activation following their order in the sequence. Estos
data show that at least some assumptions from cognitive
models of sequence planning can be linked to physiolog-
ical activities and to their neural implementation.
In the case of humans, when they give manual re-
sponses with either hand, it is possible to follow the ac-
tivation of a response and the inhibition of its alternative
with EEG recordings (p.ej., Servidor, Blanco, Montagnini, &
Burle, 2016). Readiness potentials and their lateralized
cálculo (lateralized readiness potential [LRP]) tener
been widely used to describe such dynamics (Coles,
1989; de Jong, Wierda, mulder, & mulder, 1988;
graton, Coles, Sirevaag, Eriksen, & Donchin, 1988).
Sin embargo, a more detailed approach consists of using a
spatial filter (Laplacian transform) that allows a more
direct assessment of the activity of each motor cortex
( Vidal et al., 2015; Perrin, Pernier, Bertrand, & Echallier,
1989). Using this approach, the component recorded
over the contralateral motor cortex of a response effector
has been directly linked to the activation of that cortical
área, and the analogous ipsilateral component has been
linked to the inhibition of the corresponding ipsilateral
área (Selen, Shadlen, & Wolpert, 2012; Paralí, Siegel,
Fries, & ángel, 2009; Burle, Vidal, Tandonnet, &
Hasbroucq, 2004; Vidal, Grapperon, & Bonnet, 2003).
1Aix-Marseille University, CNRS, LPC, 2Universidad Johns Hopkins,
3University of Illinois, Urbana-Champaign
What this research on single responses does not
address is whether and how these components and
© 2019 Instituto de Tecnología de Massachusetts
Revista de neurociencia cognitiva 31:7, páginas. 1030–1043
doi:10.1162/jocn_a_01401
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
1
7
1
0
3
0
1
7
8
8
6
3
1
/
/
j
oh
C
norte
_
a
_
0
1
4
0
1
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
processes would be involved in sequence production,
minimally a sequence of two strokes on a keyboard.
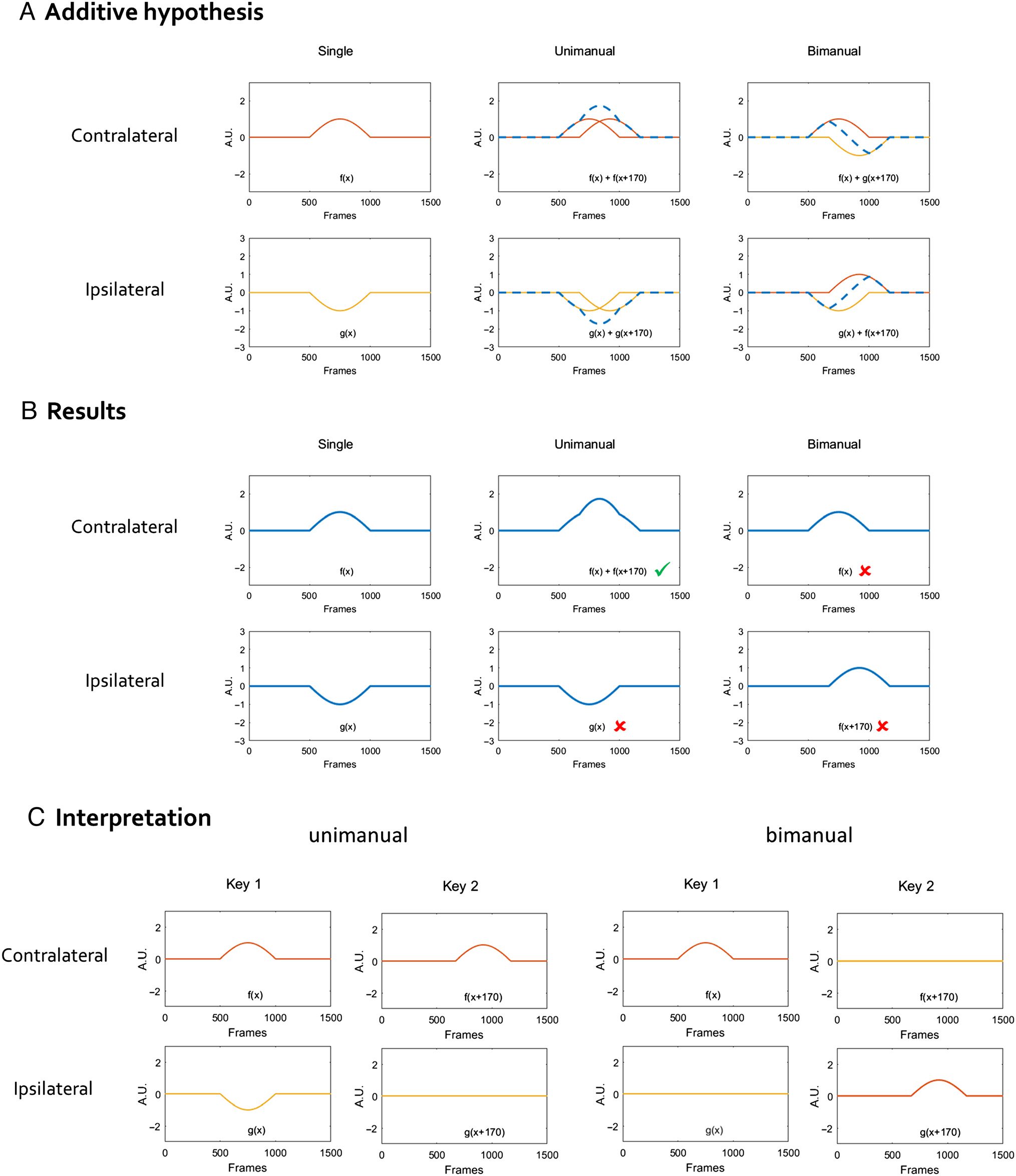
The simplest additive hypothesis would state that the
production of two responses can be understood from
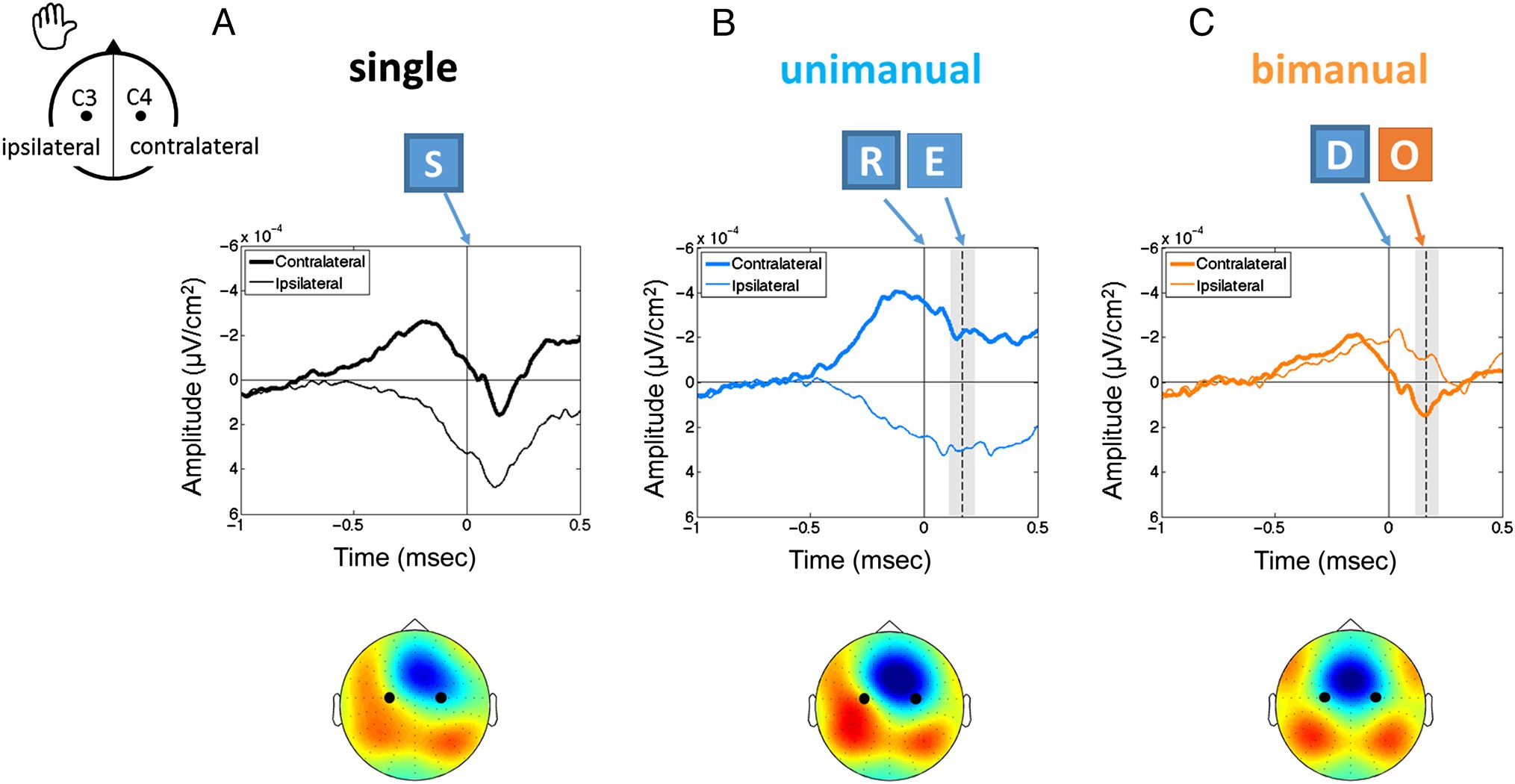
the addition of each single response. Cifra 1 regalos
the predictions of such a hypothesis on the associated
motor components, separately for unimanual (both re-
sponses use the same hand) and bimanual (each re-
sponse uses a different hand) secuencias, based on the
components known to be associated with single re-
patas. The additive hypothesis was schematized by
considering that a two-stroke sequence consists of the
simple addition of each component from the single
ataque, adjusted for the side of each keystroke, y con
a 170-msec delay between keystrokes (estimated from
the recorded average interkeystroke interval). For unim-
anual sequences, the activation and inhibition compo-
nents associated with each keystroke are added, y
each hemisphere would then exhibit something like
double the amplitude compared with the components
associated with single responses. Bimanual sequences
require the inversion of the role of each hemisphere in
terms of activation and inhibition from the first to second
keystroke. The addition of the resulting components
yields specific patterns for each hemisphere, cada uno de
which is quite different from the expectation for uniman-
ual sequences. Any deviation from the expected patterns
displayed in Figure 1 would indicate the existence of
specific sequence mechanisms.
Previous studies about bimanual sequence production
are not incompatible with the above predictions.
Focusing on the preparation period before an execution
cue, the LRP can index the amount of preparation of
the following sequence. The LRP displays a higher ampli-
tude when two movements are being prepared rather
than one (Gladwin, No es Hart, & de Jong, 2008; de Jong,
Gladwin, & No es Hart, 2006). Sin embargo, results also show
that bimanual movements cannot be easily understood
as the sum of two unimanual movements as evidenced
in particular on the motor potentials, suggesting that
specific processes might be in place during sequence
preparation (Cual & Deecke, 1999). En particular, el
effectors involved in a movement sequence influence
how the sequence is being prepared, initiated, and exe-
lindo. Sin embargo, without separating activity coming from
each hemisphere, it is not possible to formulate precise
predicciones, especially those regarding the inhibition
(ipsilateral) componente, and thus linking neurophysio-
logical results with the hypothesis of cognitive models.
In contrast with evoked potentials, time–frequency
estimates reveal the selection of motor goals or the fore-
knowledge of a sequence (Parque, kim, & Chung, 2013;
Gladwin et al., 2008). Both alpha and beta bands show
desynchronization at the start of a movement sequence,
sustained for the whole duration of the movement
(Alegre et al., 2004). Because of the slower time frame
of processes indexed by time–frequency, sin embargo, después-
tentials seem more appropriate to test the predictions
laid out by the additive model, as they reflect processes
closer to response initiation, which is of particular inter-
est here (Gladwin et al., 2008).
The activation and inhibition components described
above for single responses have been shown to be
present in the context of word typing (Scaltritti, Pinet,
Longcamp, & Alario, 2017; Pinet, Dubarry, & Alario,
2016; Pinet, Hamamé, Longcamp, Vidal, & Alario, 2015;
Cifra 1. Schematic illustration of the additive hypothesis. Electrodes contralateral and ipsilateral to the first keystroke are plotted separately.
An orange line represents the contralateral component f(X), and a yellow line represents the ipsilateral component g(X). A dashed blue line
corresponds to the addition of the components for each electrode and represents the components predicted by the additive hypothesis (see main
text for details). Equations on each graph specify the addition of components that were computed. The second keystroke is modeled to be
delayed 170 msec relative to the first keystroke.
Pinet, Dell, and Alario
1031
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
1
7
1
0
3
0
1
7
8
8
6
3
1
/
/
j
oh
C
norte
_
a
_
0
1
4
0
1
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
see also Logan, Molinero, & Strayer, 2011), a quintessential
serial order task that could provide a simple and natural
domain for addressing sequence production. Most re-
cently, Scaltritti, Alario, and Longcamp (2018) explored
the scope of planning in typing. They showed that the
neurophysiological signature associated with response
selection is modulated by item position within the up-
coming sequence (close vs. distant). Consecuentemente,
Behmer et al. (2018) used TMS to show a gradient of ac-
tivation of motor cortex that corresponded to the graded
position of future keystrokes.
The goal of our work was to identify the physiological
implementation of some assumptions made by cognitive
modelos, such as the CQ model, and particularly the as-
sumptions regarding the successive activation of each
item and the inhibition of upcoming items. en este estudio,
we tracked the cortical dynamics driving response program-
ming in single keystrokes and two-keystroke sequences
that were either unimanual or bimanual. Because pre-
vious studies allowed us to formulate precise predictions
on activation/inhibition components (Cifra 1), eran-
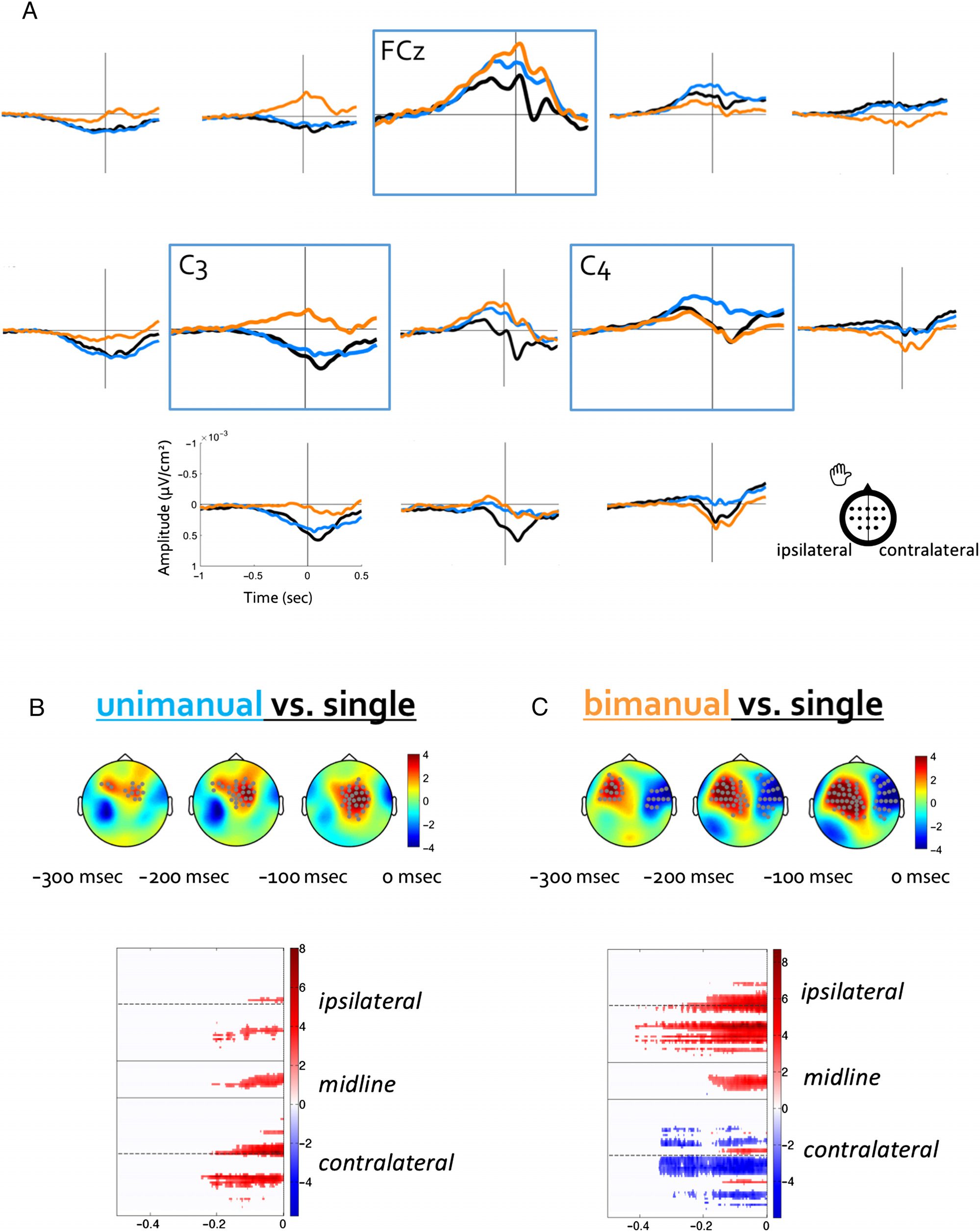
stricted our analysis to the time domain. Comparación
single keystrokes with two-keystroke sequences informs
us about whether and when activation and inhibition
processes identified for single responses are modified
by the requirement to type an additional keystroke in a
sequence context. Each contrast of sequence type (unim-
anual or bimanual) versus single strokes informs us
further about the interplay between the hemispheres.
Específicamente, the unimanual versus single contrast keeps
the recruited effectors constant but increases the number
of strokes in the unimanual case, perhaps requiring more
processing from the contralateral hemisphere. El
bimanual versus single contrast tests the role of both ef-
fectors, particularly the possible influence of inhibition of
the hemisphere not involved in the immediate keystroke.
MÉTODOS
Participantes
Eighteen right-handed participants were selected for
their ability to touch-type, eso es, typing with all (o
almost all) 10 fingers without the need to look at their
manos. Four participants were excluded because of prob-
lematic eye movements around the moment of response
or poor signal-to-noise ratio. The final sample (five men,
nine women) era 24.9 years old on average (rango: 20–
33 años). Participants’ typing ability was assessed via
a typing test (p.ej., Pinet et al., 2016). Participants had a
mean accuracy of 84% (rango: 78–91%) and a mean
typing speed of 48 palabras por minuto (wpm; five-character
wpm; rango: 34.6–57.5 wpm).
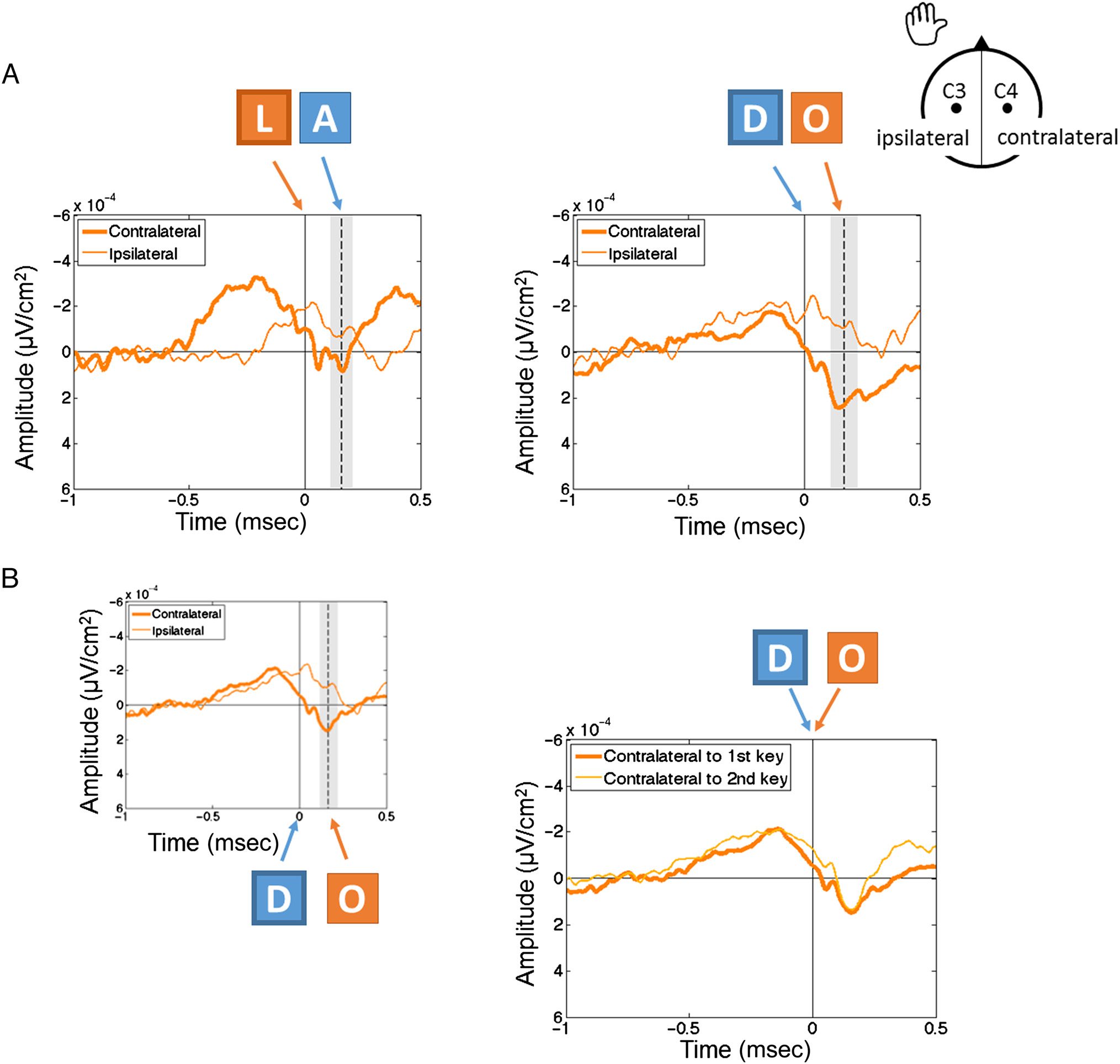
mano (“Z,” “E,” “S,” “D”); and the other half, with the right
mano ("I,” “O,” “K,” “L”) in the AZERTY keyboard. El
words selected were the French names of musical notes,
to ensure homogeneity of meaning and because they
present interesting sequence properties. Four musical
note names started with the left hand (“DO,” “RE,"
“FA,” and “SI”), and two started with the right hand
(“MI” and “LA”); half were typed with one hand (uniman-
ual: “RE,” “FA,” “MI”), and the other half were typed with
both hands (bimanual: “DO,” “SI,” “LA”). Por eso, estímulos
were divided into three conditions: single (one letter),
unimanual (two letters, one hand), and bimanual (dos
letters, two hands). The only three-letter musical note
(“SOL”) was also included for completeness, but data
from SOL trials were not analyzed. Stimulus presentation
was visual (printed letters) and auditory (spoken letter or
music note names). Stimulus modality (visual vs. audi-
conservador) and stimulus type (letters vs. musical notes) eran
presented in four independent blocks. Each participant
saw all conditions in a randomized order.
Procedimiento
Visual stimuli were displayed on a 17-in. CRT computer
screen placed at about 70 cm from the participant.
Auditory stimuli were presented binaurally through
earphones. Responses were collected from a DirectIN
High Speed Keyboard PCB v2010 (Empirisoft) to obtain
keystroke timing data with at least 1-msec accuracy.
Stimuli presentation and response acquisition were con-
trolled using the Presentation software (NeuroBehavioral
Sistemas).
Participants sat in a dimly lighted, electrically shielded
habitación. Their sitting position was adjusted so that they
were comfortable typing. The task was to type the letter
or musical note name on the keyboard as fast and accu-
rately as possible after presentation. Letter blocks com-
prised 200 ensayos (25 repetitions of each of the eight
estímulos). Music blocks comprised 350 ensayos (50 reps-
etitions of each of the seven note names). Each block
included a short break.
A trial consisted of a fixation cross presented during a
random duration from 400 a 600 mseg, followed by pre-
sentation of the stimulus (maximal duration = 2000 mseg).
After the response, the stimulus stayed on for 500 mseg,
and then feedback (correct or incorrect, indicated by a
green check mark or a red cross or by high- and low-pitch
beeps) was displayed for 500 msec in the same modality
as the stimulus (visual or auditory). The screen then
remained black for 500 mseg. Every two to three trials,
there was a 2-sec interval to allow participants to blink
and avoid contamination of the signal.
Stimuli and Design
Behavioral Data Analysis
Eight single letters and six two-letter words were used as
estímulos. Half of the single letters were typed with the left
RTs (time of the first keystroke) and accuracy rates were
analyzed with linear mixed-effect models, as is commonly
1032
Revista de neurociencia cognitiva
Volumen 31, Número 7
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
1
7
1
0
3
0
1
7
8
8
6
3
1
/
/
j
oh
C
norte
_
a
_
0
1
4
0
1
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
done in cognitive studies of language (Baayen, Davidson,
& Batio, 2008; Jaeger, 2008). RTs were log-transformed,
following the result of a Box Cox test. RTs below 300 mseg
or incorrect responses were removed from the RT
análisis. RTs and accuracy rates were analyzed according
to the following predictors: modality (auditory or visual),
first keystroke hand (left or right), and second keystroke
(single, unimanual, bimanual). To account for the three
levels of the second keystroke predictor, the predictors
were Helmert-coded to contrast sequence length (uno
key vs. two keys) and second keystroke side (same/
diferente). By-participant random slopes were added for
all predictors of interest. Random intercepts by partic-
ipant and by item were also added to the model. Trial
number was added as a control variable. Accuracy rate
analysis included a random slope for modality only, como
the model would not converge otherwise.
EEG Recordings and Preprocessing
EEG was acquired from 128 scalp locations using Ag/AgCl
active electrodes (BioSemi Active Two system), referirse-
enced to the CMS-DRL ground. The sampling rate was
512 Hz (filters: DC to 104 Hz, 3-dB/octave slope).
Vertical and horizontal EOGs were recorded with three
surface electrodes (Ag/AgCl) placed below the left eye
and next to the two outer canthi.
Offline analyses were performed using the MATLAB
toolboxes Brainstorm (Tadel, Baillet, Mosher, Pantazis,
& Leahy, 2011) and Fieldtrip (Oostenveld, Fries, Maris,
& Schoffelen, 2011), as well as custom routines. Datos
were rereferenced to the average of both mastoids.
Continuous data were filtered (Butterworth Filter Order
4: 0.1–100 Hz). Bad electrodes were removed and inter-
polated. A first mild artifact rejection was performed
before computing independent component analysis (Alabama-
gorithm: Infomax from EEGLAB toolbox; Delorme &
Makeig, 2004). Component(s) corresponding to eye
movements was (eran) removed. A second stricter arti-
fact rejection was then performed to reject remaining
noisy segments. A −200- to 0-msec prestimulus baseline
was applied by subtraction. Response-locked epochs
were then extracted (−1500 to +800 msec around the
first or second keystroke).
Current source density transformation was applied
(number of spherical splines: 4, maximal degree of
Legendre polynomial: 20, smoothing parameter: 10-5)
to enhance spatial resolution of the EEG signal (Perrin
et al., 1989).
EEG Statistical Analysis
Unless stated otherwise, activities on the contralateral
hemisphere from left- and right-hand responses were av-
eraged together. Similarmente, activities on the ipsilateral
hemisphere from left- and right-hand responses were
averaged together. The signal is presented relative to
left-hand responses (es decir., ipsilateral hemisphere corre-
sponds to the left hemisphere).
Two main contrasts (unimanual vs. single and bimanual
vs. single) were tested from −500 to 0 msec including
todo 128 electrodes, using cluster-based nonparametric sta-
tistics (Maris & Oostenveld, 2007). This procedure allows
us to deal with the multiple-comparison problem by clus-
tering significant neighboring channels and time points
and building the statistical distribution directly from the
observed data by successive permutations. More specifi-
cally, two conditions are compared with a paired t test at
each time point. Clusters are composed of the time
points presenting t values above a specific threshold
( pag < .05) based on spatial and temporal adjacency. The
sum of t values within each cluster gives the cluster-
level statistic, and the significance of each cluster is
computed using permutations. However, one should
refrain from overinterpreting the spatial or temporal exten-
sion of any cluster. Because the significance is only tested
between conditions, one should not conclude strongly
about the boundaries of any cluster (any point is not tested
against another). Nevertheless, we depict the significant
clusters as Channel × Time matrices for illustrative pur-
poses. Although all electrodes within a cluster show a
significant effect, each electrode can display a slightly
different pattern from other electrodes of the same clus-
ter. For that reason, we present an array of electrodes
rather than the averaged signal over cluster electrodes.
To compare our results with previous literature, we also
display specific electrodes over the motor cortex, close to
C3/C4 (i.e., D18 and B21 in our 128-electrode system),
based on Pinet et al. (2016).
The two main contrasts were run for data collapsed
over modality and hand (main analysis reported). They
were also run for each modality and hand separately
(eight additional contrasts).
RESULTS
Behavioral Results
The RT analysis (see Table 1) revealed a main effect of
Modality, with auditory stimuli eliciting slower responses
than visual stimuli (ß = 0.14, t = 4.5, 95% CI [0.076,
0.20], Mvisual = 697.7 ± 231.4 msec, Mauditory = 792.4 ±
255.3 msec). RTs also exhibited a practice effect, strongly
decreasing with trial number (ß = −8.43E-5, t = −13.2,
95% CI [−9.7E-5, −7.2E-5]). None of the other effects
was significant.
Mean accuracy was 93.5%. Accuracy rate analysis (see
Table 2) revealed that unimanual sequences were pro-
duced more accurately than bimanual sequences (ß =
1.1, z = 3.4, 95% CI [0.45, 1.7], Munimanual = 94.1 ±
2.8%, Mbimanual = 93.1 ± 5.0%). None of the other
effects was significant.
Pinet, Dell, and Alario
1033
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
1
7
1
0
3
0
1
7
8
8
6
3
1
/
/
j
o
c
n
_
a
_
0
1
4
0
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Coefficient
SE
t
95% CI
Table 1. Mixed Model Regression Results for RT
Fixed Effects
Intercept
Modality (visual/auditory)
First key (left/right)
Second key (one/two keys)
Second key (same/different hand)
Trial
Random Effects
Participant intercept
Modality | participant
First key | participant
Second key (one/two keys) | participant
Second key (same/different hand) | participant
Item intercept
Residual
0.046
0.031
0.029
0.035
0.043
0.000
6.568
0.136
−0.051
0.036
−0.012
−8.43E-05
Variance
0.027
0.013
0.006
0.011
0.015
0.001
0.046
143.115
4.46
−1.78
1.04
−0.28
6.478
0.076
−0.108
−0.032
−0.097
6.658
0.196
0.005
0.104
0.073
p
<2e-16
6.49E-04
.090
.312
.782
−13.22
−9.68E-05
−7.18E-05
<2e-16
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
>