Theta Phase Synchronization between the Human
Hippocampus and Prefrontal Cortex Increases
during Encoding of Unexpected Information:
Un estudio de caso
Matthias J. Gruber1,2, Liang-Tien Hsieh2,3, Bernhard P. Staresina4, Christian E. Elger5,
Juergen Fell5, Nikolai Axmacher6, and Charan Ranganath2
Abstracto
■ Events that violate predictions are thought to not only modu-
late activity within the hippocampus and PFC but also enhance
communication between the two regions. Scalp and intracranial
EEG studies have shown that oscillations in the theta frequency
band are enhanced during processing of contextually unexpected
información. Some theories suggest that the hippocampus and
PFC interact during processing of unexpected events, and it is
possible that theta oscillations may mediate these interactions.
Aquí, we had the rare opportunity to conduct simultaneous elec-
trophysiological recordings from the human hippocampus and
PFC from two patients undergoing presurgical evaluation for
pharmacoresistant epilepsy. Recordings were conducted during
a task that involved encoding of contextually expected and un-
expected visual stimuli. Across both patients, hippocampal–
prefrontal theta phase synchronization was significantly higher
during encoding of contextually unexpected study items, rel-
ative to contextually expected study items. Además, el
hippocampal–prefrontal theta phase synchronization was
larger for contextually unexpected items that were later remem-
bered compared with later forgotten items. Además, we did not
find increased theta synchronization between the PFC and rhinal
corteza, suggesting that the observed effects were specific to
prefrontal–hippocampal interactions. Our findings are consistent
with the idea that theta oscillations orchestrate communication
between the hippocampus and PFC in support of enhanced en-
coding of contextually deviant information. ■
INTRODUCCIÓN
Unexpected events that violate internal predictions are
more likely to be successfully encoded to memory
(p.ej., Elhalal, Davelaar, & Ujier, 2014; Murty & Adcock,
2014; Schomaker et al., 2014; Axmacher et al., 2010). Él
has been proposed (Lisman & Grace, 2005; Ranganath
& Rainer, 2003) that the hippocampus and PFC play a
critical role in the detection and formation of memories
of contextually unexpected events (p.ej., rare events of a
specific category that are randomly encountered within
most events of a different category; Von Restorff, 1933).
Consistent with this idea, fMRI studies in humans have in-
dicated that processing of contextually unexpected in-
formation is associated with increased activation in the
hippocampus and PFC (and other cortical/subcortical re-
gions; Murty & Adcock, 2014; Murty, Ballard, Macduffie,
krebs, & Adcock, 2013; Yassa & Rígido, 2008; Bunzeck &
Düzel, 2006; Strange & Dolan, 2001). Curiosamente, func-
1Universidad de Cardiff, 2Universidad de California, davis, 3Universidad
of California, berkeley, 4University of Birmingham, 5Universidad
of Bonn, 6Ruhr University Bochum
© 2018 Instituto de Tecnología de Massachusetts
tional connectivity between the hippocampus and PFC is
enhanced during successful memory encoding and re-
trieval (p.ej., Nee & En Jonia, 2008; Ranganath, infierno,
cohen, Brozinsky, & Rissman, 2005; grady, McIntosh, &
Craik, 2003). Por lo tanto, a currently unresolved question
is whether or how these regions interact during encoding
or processing of contextually unexpected events.
Several EEG studies have suggested that neural oscilla-
tions in the theta band are enhanced after contextually
unexpected events. Studies using intracranial EEG have
shown that hippocampal theta power is increased during
encoding of contextually unexpected information (Chen
et al., 2013; Axmacher et al., 2010). Además, contex-
tually unexpected events elicit increases in scalp-recorded
frontal theta power (p.ej., Cavanagh & Franco, 2014;
Cavanagh, Figueroa, cohen, & Franco, 2012; Walsh &
anderson, 2012), and recent EEG studies have demon-
strated increases in theta phase synchrony between
frontal and temporal scalp sites during contextually un-
expected stimuli (Harper, Malone, & Iacono, 2017; Sotavento,
Sotavento, kim, & Jung, 2014). In light of this evidence, es
possible that theta oscillations facilitate communication
between PFC and the hippocampus during encoding of
contextually unexpected events.
Revista de neurociencia cognitiva 30:11, páginas. 1646–1656
doi:10.1162/jocn_a_01302
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
1
1
6
4
6
1
7
8
7
8
6
9
/
/
j
oh
C
norte
_
a
_
0
1
3
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Results from other paradigms have indicated that inter-
actions between PFC and the hippocampus could be
mediated by theta coupling. Por ejemplo, intracranial
EEG studies in humans have reported increased theta
phase synchronization between PFC and medial temporal
lobe cortical regions during virtual navigation and memory
retrieval (Watrous, Tandon, Conner, Pieters, & Ekstrom,
2013; anderson, Rajagovindan, Ghacibeh, Meador, &
Ding, 2010; Kahana, Sekuler, Caplan, Kirschen, & Madsen,
1999; but see Raghavachari et al., 2006), but these studies
did not report changes in phase synchrony specifically
with the hippocampus. Recent studies on memory re-
trieval in humans using source localization on magneto-
encephalography data or combined EEG–fMRI data also
suggest that theta oscillations correlate with hippocampal–
PFC connectivity (Kaplan et al., 2017; Herweg et al., 2016;
Fuentemilla, Barnes, Duzel, & Levin, 2014).
Consistent with the idea that theta oscillations might
facilitate communication between PFC and the hippo-
campus, local field potential recordings in rodents have
shown that salient events (p.ej., those occurring at choice
points in a maze learning task) increase oscillatory power
in the theta band (4–8 Hz) within the hippocampus and
PFC (p.ej., Donnelly et al., 2014; O’Neill, gordon, &
Sigurdsson, 2013; Totah, Jackson, & Moghaddam, 2013;
Hasselmo, Bodelón, & Wyble, 2002; Winson, 1978). Fur-
thermore, recordings in rodents and nonhuman primates
have also shown that theta oscillations synchronize be-
tween the two areas (Brincat & Molinero, 2015; Fujisawa &
Buzsáki, 2011; Benchenane et al., 2010; Hyman, Zilli,
Paley, & Hasselmo, 2005; jones & wilson, 2005). por ejemplo-
amplio, enhanced theta phase synchrony between the
hippocampus and PFC has been shown during perfor-
mance of a spatial T-maze task (Benchenane et al.,
2010) and during retrieval of object–context associations
(Place, Farovik, Brockmann, & Eichenbaum, 2016). Estos
findings in the rodent brain are consistent with the idea
that phase synchronization in the theta frequency band is
relevant for spike-timing-dependent plasticity (Fell &
Axmacher, 2011). Sin embargo, little is known about the ex-
tent to which the findings of frontal–hippocampal synchro-
nization in rodents correspond to activity in the human
cerebro.
en este estudio, we used intracranial EEG to determine
(i) whether human hippocampal–PFC theta phase syn-
chrony is enhanced during processing of contextually
unexpected events and (ii) whether hippocampal–PFC
theta phase synchrony predicts later memory perfor-
mance. We used a Von Restorff paradigm ( Von Restorff,
1933) in which patients encoded trial-unique images
from two different categories (for exemplary trials, ver
Figura 2A). En tono rimbombante, one type of stimuli comprised
most encoding stimuli in a given encoding-test block
(contextually expected items; p.ej., grayscale faces on a
red background), and the other type of stimuli only
comprised a small percentage (es decir., 14%) of the encod-
ing stimuli in a given encoding-test block (contextually
unexpected items; p.ej., grayscale houses on a green
fondo). During the encoding phase, we recorded
intracranial EEG simultaneously from the hippocampus
and PFC in two patients with pharmacoresistant epilepsy.
The locations of the implanted prefrontal electrodes also
allowed us to explore whether theta phase synchroniza-
tion with the hippocampus might be evident with specific
subregions of PFC. Además, we also investigated
phase synchronization between PFC and sites in the
rhinal cortex.
MÉTODOS
We recorded intracranial EEG from two pharmacorefrac-
tory epileptic patients at the Department of Epileptology
at the University of Bonn, Alemania. Both patients (uno
woman; 46 y 48 years old) were implanted with bilat-
eral depth electrodes in the hippocampus and the adja-
cent rhinal cortex as well as with bilateral subdural
electrodes covering parts of PFC (es decir., one frontopolar
and one frontolateral electrode strip bilaterally covering
rostral/anterior and lateral PFC regions, respectivamente; ver
Cifra 1). From the larger sample of patients reported in
Axmacher et al. (2010), the two patients were the only
patients who had both implanted hippocampal and PFC
electrodes. Details about the patients and analyses of
ERPs and oscillatory power from hippocampal sites in
these two patients are presented in Axmacher et al.
(2010). Because epileptic seizures were focused on left
hippocampal and surrounding medial temporal lobe
areas in one patient and left medial temporal lobe areas
and left temporolateral areas in the other patient, nosotros sólo
considered data from the hippocampal, rhinal, and PFC
electrodes on the right hemisphere. The local ethics
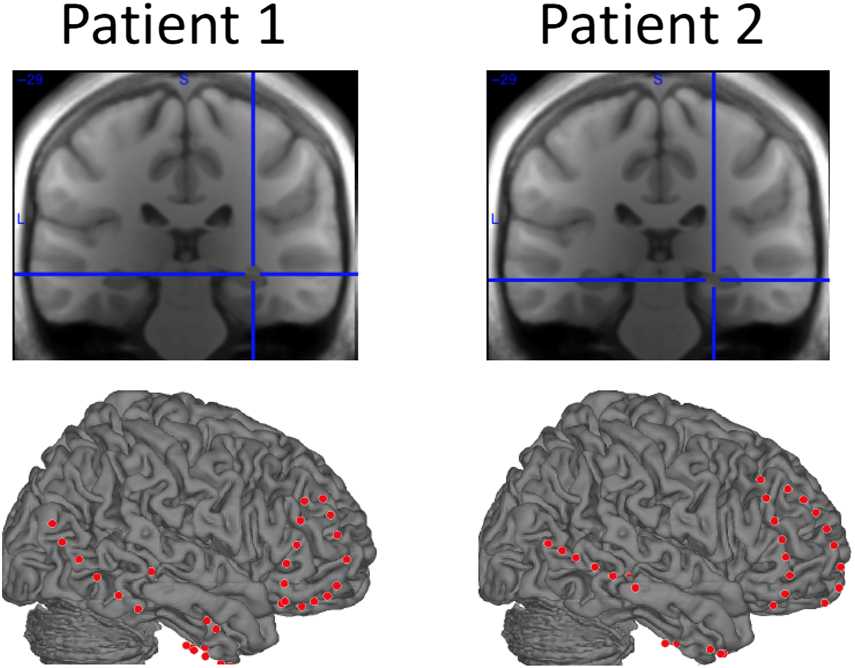
Cifra 1. Locations of hippocampal and prefrontal electrodes. Sobre el
arriba, the location of the selected hippocampal electrode is depicted for
each patient (Patient 1: MNI = 32, −29, −7; Patient 2: MNI = 26, −29,
−10). On the bottom, all implanted subdural strip electrodes covering the
right hemisphere are depicted for each patient. Only the frontopolar and
frontolateral strips were analyzed for each patient.
Gruber et al.
1647
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
1
1
6
4
6
1
7
8
7
8
6
9
/
/
j
oh
C
norte
_
a
_
0
1
3
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
Primero, we restricted our iEEG analyses to contextually un-
esperado (Patient 1: 32 ensayos; Patient 2: 15 ensayos) y estafa-
textually expected items (Patient 1: 68 ensayos; Patient 2: 45
ensayos) that were later correctly recognized in the recogni-
tion memory test (es decir., collapsed across correct confident
old and unconfident old responses), to examine effects of
contextual unexpectedness. This approach gave us a suf-
ficient number of trials and did not confound effects
driven by contextual unexpectedness with memory en-
codificación. Segundo, we asked whether any potential theta
phase synchronization effects further predict later mem-
ory. Para tal fin, we compared the later remembered
elementos (from the previous analysis) with the later forgot-
ten items separately in the contextually unexpected
condición (forgotten items: Patient 1, 17 ensayos; Patient
2, 25 ensayos) and the contextually expected condition (para-
gotten items: Patient 1, 18 ensayos; Patient 2, 29 ensayos). Para-
gotten trials included items with incorrect confident new
and unconfident new responses as well as items for
which the patients did not give any response during
the recognition test.
Because electrode placement varied across patients
because of the clinical needs of each patient, we focused
our analyses on hippocampal contacts that were most
consistently localized across the two patients. Eso es,
we first selected one hippocampal electrode per patient
that had maximal anatomical overlap between the two
patients. The selected hippocampal electrode pair (uno
electrode from each patient) had the smallest Euclidean
distance between the two patients (7-mm distance; Patient 1:
Instituto Neurológico de Montreal [MNI] = 32, −29, −7;
Patient 2: MNI = 26, −29, −10; ver figura 1). Nosotros entonces
used the EEGLAB toolbox (Delorme & Makeig, 2004) a
segment the iEEG data into epochs from −2 to +3 segundo
relative to the onset of all items. To preprocess these
datos, primero, we used an automated artifact detection pro-
cedure implemented in EEGLAB, in which EEG activity
that exceeded more than three 3 SDs from the mean
on that electrode or 5 SDs across all electrodes was ex-
cluded from the analyses (Gruber, Watrous, Ekstrom,
Ranganath, & Otten, 2013). Segundo, in line with our orig-
inal data set (Axmacher et al., 2010), we then visually
inspected the hippocampal and prefrontal raw data and
further manually discarded trials containing EEG artifacts
and epileptiform activity from any further analyses (es decir.,
trials discarded because of artifacts or epileptic signals
detected in a given channel were also excluded from
the analysis for all other electrodes).
We also excluded data from the first electrode of each
PFC electrode strip (es decir., most inferior electrode) for both
patients because of a very low signal-to-noise ratio as
compared with all other remaining PFC electrodes (es decir.,
no visible event-related evoked responses across contex-
tually unexpected and expected trials). Artifact-free iEEG
data were then imported into the Fieldtrip toolbox
(Oostenveld, Fries, Maris, & Schoffelen, 2011) for further
análisis. Primero, standard time–frequency decomposition
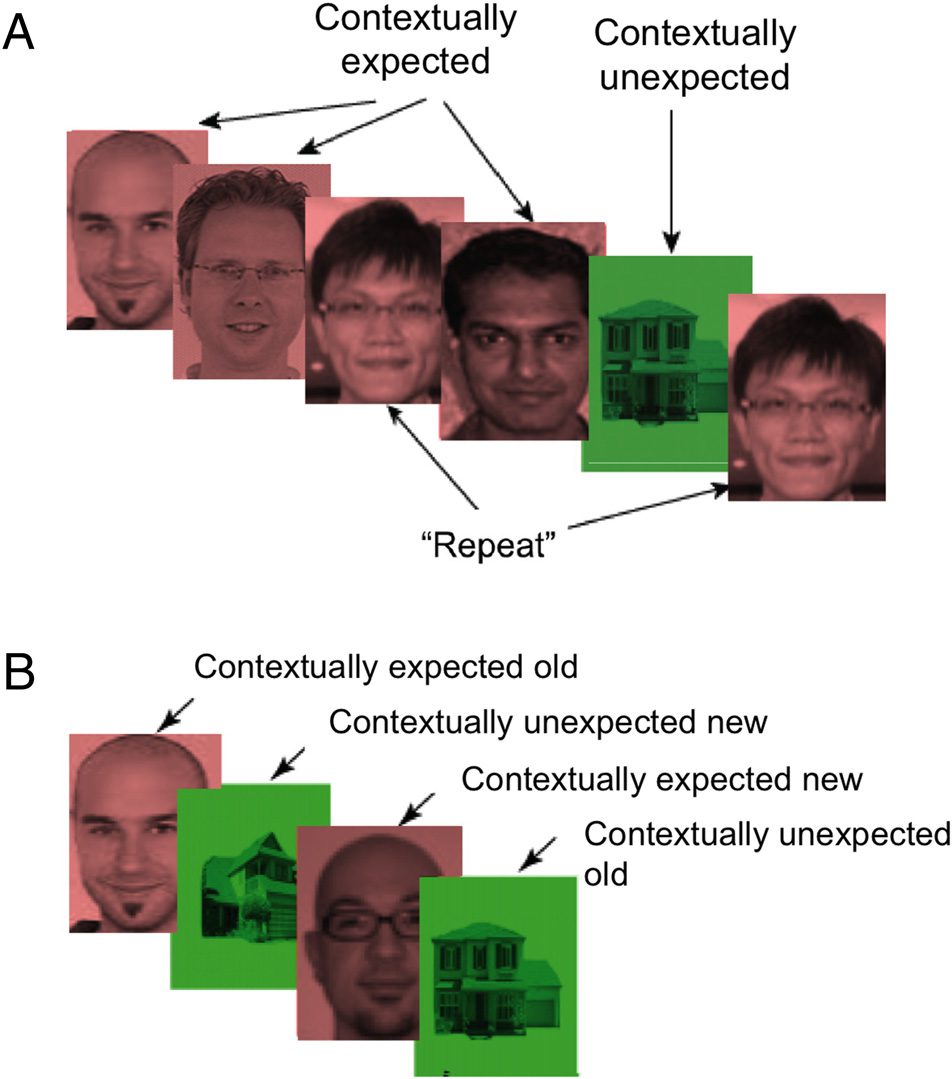
Cifra 2. Experimental procedure. (A) During the encoding phase for
which iEEG results are reported here, patients encoded images of
stimuli that comprised most encoding stimuli (contextually expected
elementos), and the other type of stimuli only comprised a small
porcentaje (contextually unexpected items). Categories (es decir., faces,
houses) and colors (es decir., rojo, verde) of contextually expected and
unexpected stimuli were counterbalanced across blocks in each
patient. (B) After an encoding block, patients completed a recognition
memory test.
committee approved the study, and both patients gave
written informed consent.
Both patients took part in a variant of a Von Restorff par-
adigm (Von Restorff, 1933; for details of the experimental
procedimiento, see Axmacher et al., 2010). During the en-
coding phase for which iEEG results are reported here,
patients encoded trial-unique images from two different
categories (for exemplary trials, see Figure 2A). Impor-
tantly, one type of stimuli comprised most encoding stim-
uli in a given encoding-test block (contextually expected
elementos; p.ej., grayscale faces on a red background as shown
in Figure 2A), and the other type of stimuli only comprised
a small percentage (es decir., 14%) of the encoding stimuli in a
given encoding-test block (contextually unexpected items;
p.ej., grayscale houses on a green background as shown in
Figura 2A). Categories and colors of contextually expect-
ed and unexpected stimuli were counterbalanced across
blocks in each patient. After the encoding phase, patients
completed a recognition memory test for these images
(Figura 2B). Memory accuracy (es decir., hits − false alarms
collapsed across confident old and unconfident old re-
sponses) was higher for contextually unexpected compared
with expected events in Patient 1 (40% vs. 35%) but not in
Patient 2 (44% vs. 53%).
1648
Revista de neurociencia cognitiva
Volumen 30, Número 11
yo
D
oh
w
norte
oh
a
d
mi
d
F
r
oh
metro
h
t
t
pag
:
/
/
d
i
r
mi
C
t
.
metro
i
t
.
mi
d
tu
/
j
/
oh
C
norte
a
r
t
i
C
mi
–
pag
d
yo
F
/
/
/
3
0
1
1
1
6
4
6
1
7
8
7
8
6
9
/
/
j
oh
C
norte
_
a
_
0
1
3
0
2
pag
d
.
F
b
y
gramo
tu
mi
s
t
t
oh
norte
0
8
S
mi
pag
mi
metro
b
mi
r
2
0
2
3
was performed on artifact-free raw EEG data to obtain
power and phase information. We used a Morlet wavelet
decomposition method with a width of 5 cycles in indi-
vidual frequencies. Decomposition was conducted within
the epoch period of −0.2 to 1.2 segundo (t1 = onset of event)
in steps of 0.02 sec and in the frequency range of 2–20 Hz.
Segundo, to address the role of theta phase synchrony
between the hippocampus and PFC, we calculated phase
synchrony indices between the previously selected hip-
pocampal electrode and each of the artifact-free frontal
electrodes, Resultando en 14 hippocampal–PFC electrode
pairs for each patient. Phase synchrony was separately
quantified for all four conditions (contextually unex-
pected remembered, contextually unexpected forgotten,
contextually expected remembered, and contextually ex-
pected forgotten) using the debiased estimator of the
squared weighted phase lag index (d-WPLI) implemented
in Fieldtrip. The d-WPLI has the advantage that it allevi-
ates problems related to volume conduction and other
noise-related issues (Vinck, Oostenveld, van Wingerden,
battaglia, & Pennartz, 2011).
To statistically determine whether contextually unex-
pected compared with expected items show a significant
theta phase synchrony increase, we used a nonparametric
statistical approach that randomly permutes condition
labels to correct for multiple comparisons across electrode
pares. Analyses were conducted separately in each patient
for all data points within a selected time–frequency range
(time range = −0.2 to 1.2 segundo, frequency range = 2–20 Hz).
This analysis approach had the strength to reveal signifi-
cant time–frequency clusters without prior selection of a
specific time–frequency bin of interest. The steps are as
follows: (1) We computed the d-WPLI values within the
selected time–frequency range for each condition (a
use an identical approach as for the surrogate data, nosotros
randomly selected equal trial numbers from two condi-
tions of interest based on the minimum number of trials
in one condition). We then computed the difference of
the d-WPLI values between the conditions of interest
(es decir., first analysis: contextually unexpected vs. esperado
elementos; second and third analyses: remembered vs. forgot-
ten items in the contextually unexpected and contextually
expected conditions, separately). Thereby, we obtained
the empirical difference in theta phase synchrony (es decir.,
d-WPLI) between two conditions. (2) We shuffled trial
labels by randomly selecting equal trial numbers from
the two conditions based on the minimum number of tri-
als in one condition, calculated surrogate phase synchrony
values for all 14 electrode pairs, took the difference be-
tween the surrogate conditions for all 14 electrode pairs,
and saved the maximum surrogate phase synchrony differ-
ence across all 14 electrode pairs (es decir., electrode-pairmax).
(3) Step 2 was repeated 500 veces. On the basis of the
500 permutations, we created a null distribution of all
electrode-pairmax difference values and determined the
alpha cutoff point ( pag < .05, one-sided; i.e., 475th data point
in surrogate difference distribution) to test the statistical
significance of the empirical theta phase synchrony values
for all electrode pairs. This stringent approach allowed us
to correct for multiple comparisons across electrodes.
RESULTS
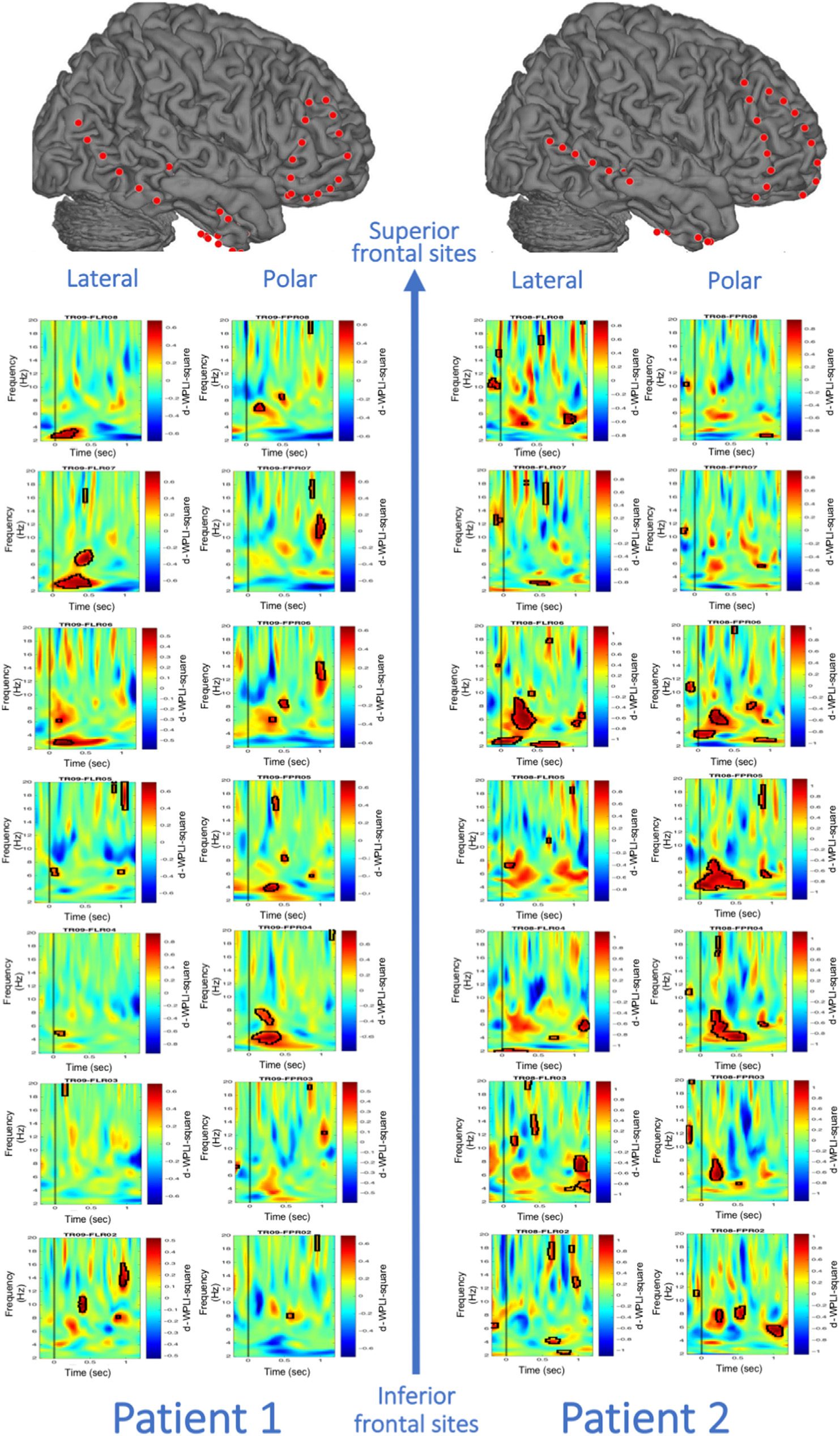
As shown in Figure A1, in both patients, permutation
tests that corrected for multiple comparisons revealed
that frontopolar (within Brodmann’s area 10) and dorso-
lateral (within Brodmann’s area 46) prefrontal electrode
sites showed significantly increased theta phase syn-
chrony with the hippocampus during encoding of contex-
tually unexpected compared with expected items.
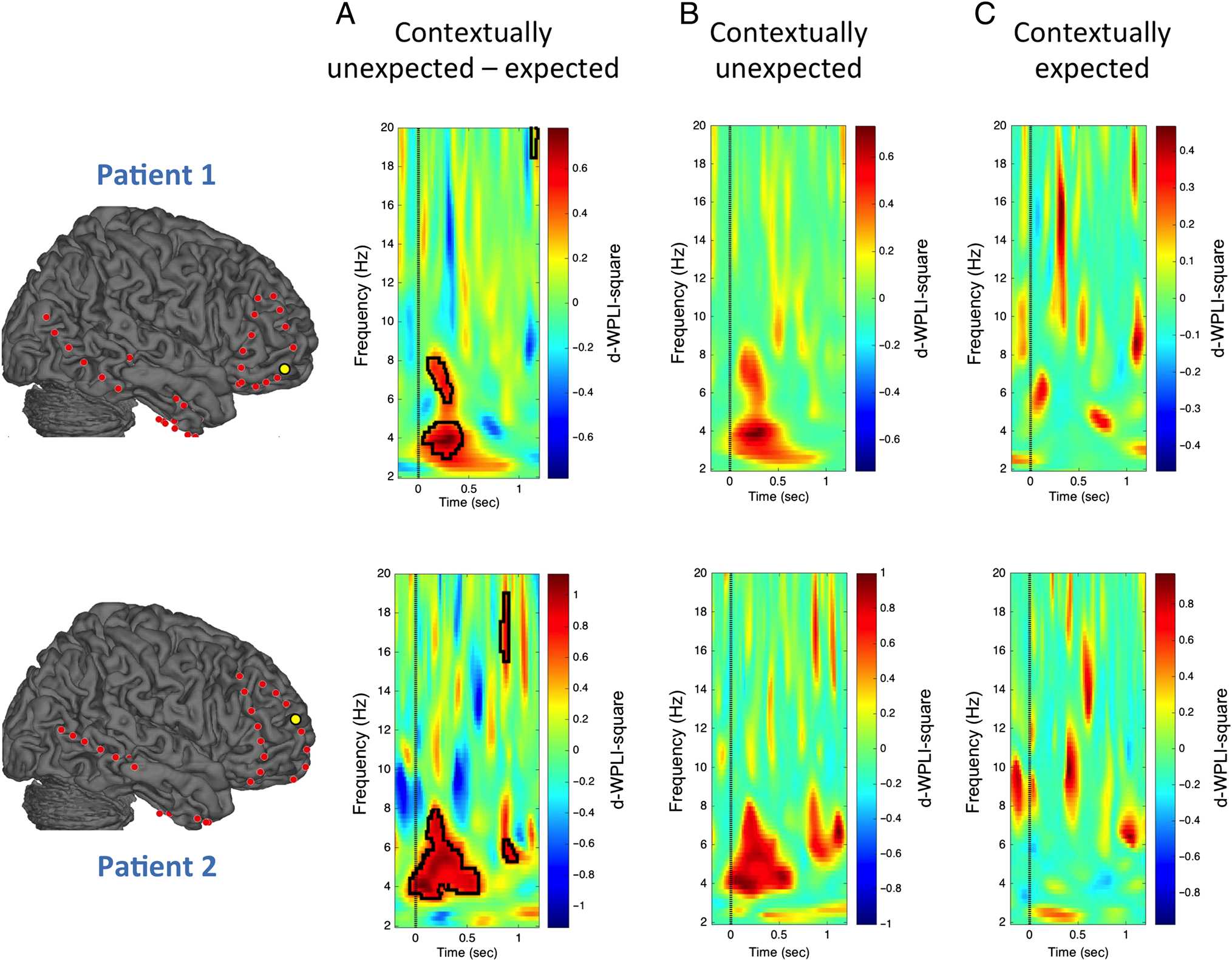
Figure 3 depicts one selected hippocampal–frontopolar
electrode pair per patient showing phase synchronization
increases for contextually unexpected compared with ex-
pected items in the theta frequency range (∼3–8 Hz;
black contours show the permutation-based significant
difference clusters in Figure 3A). We found that the theta
phase synchronization increase is specific to contextually
unexpected items (Figure 3B) and absent in contextually
expected items (Figure 3C).
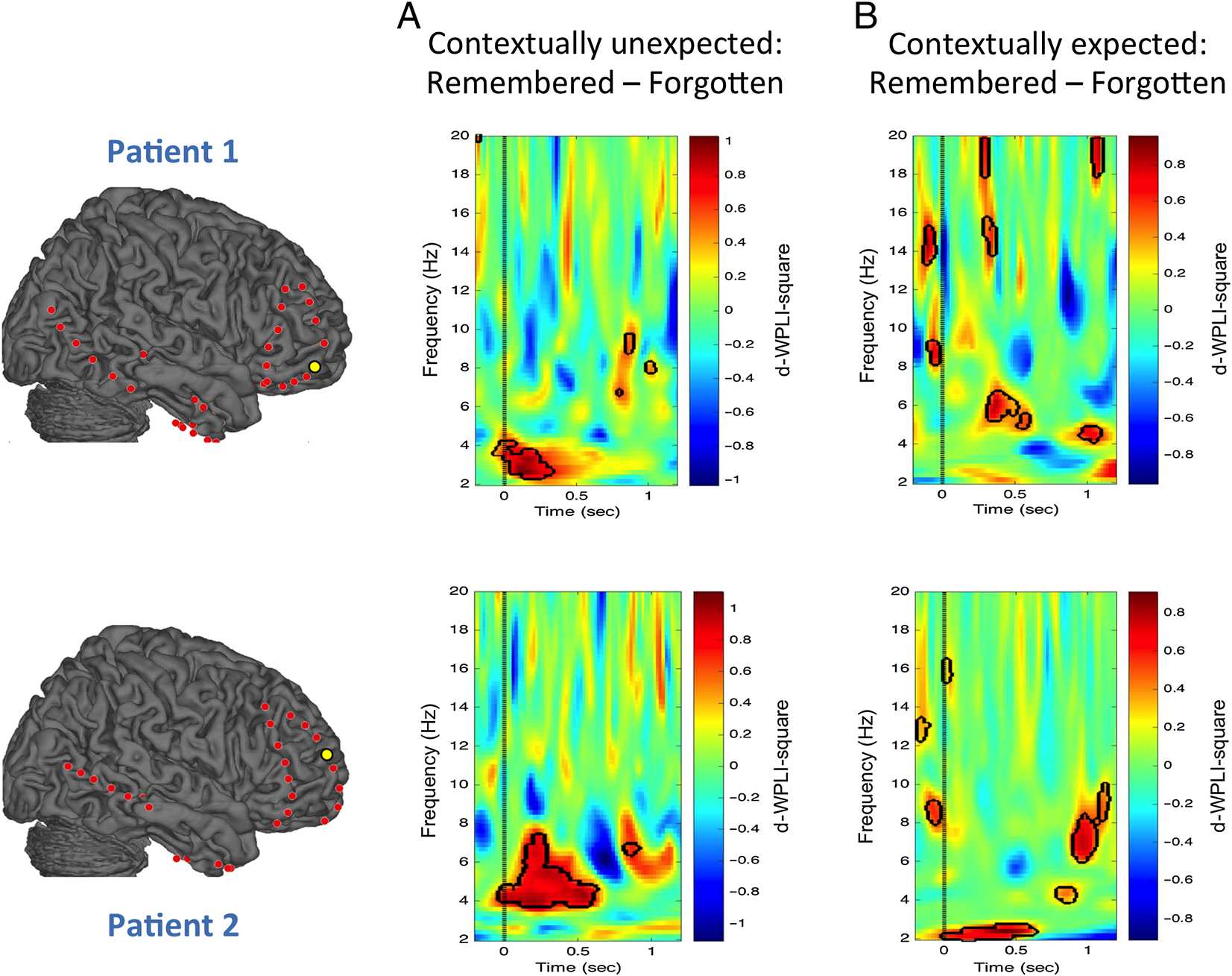
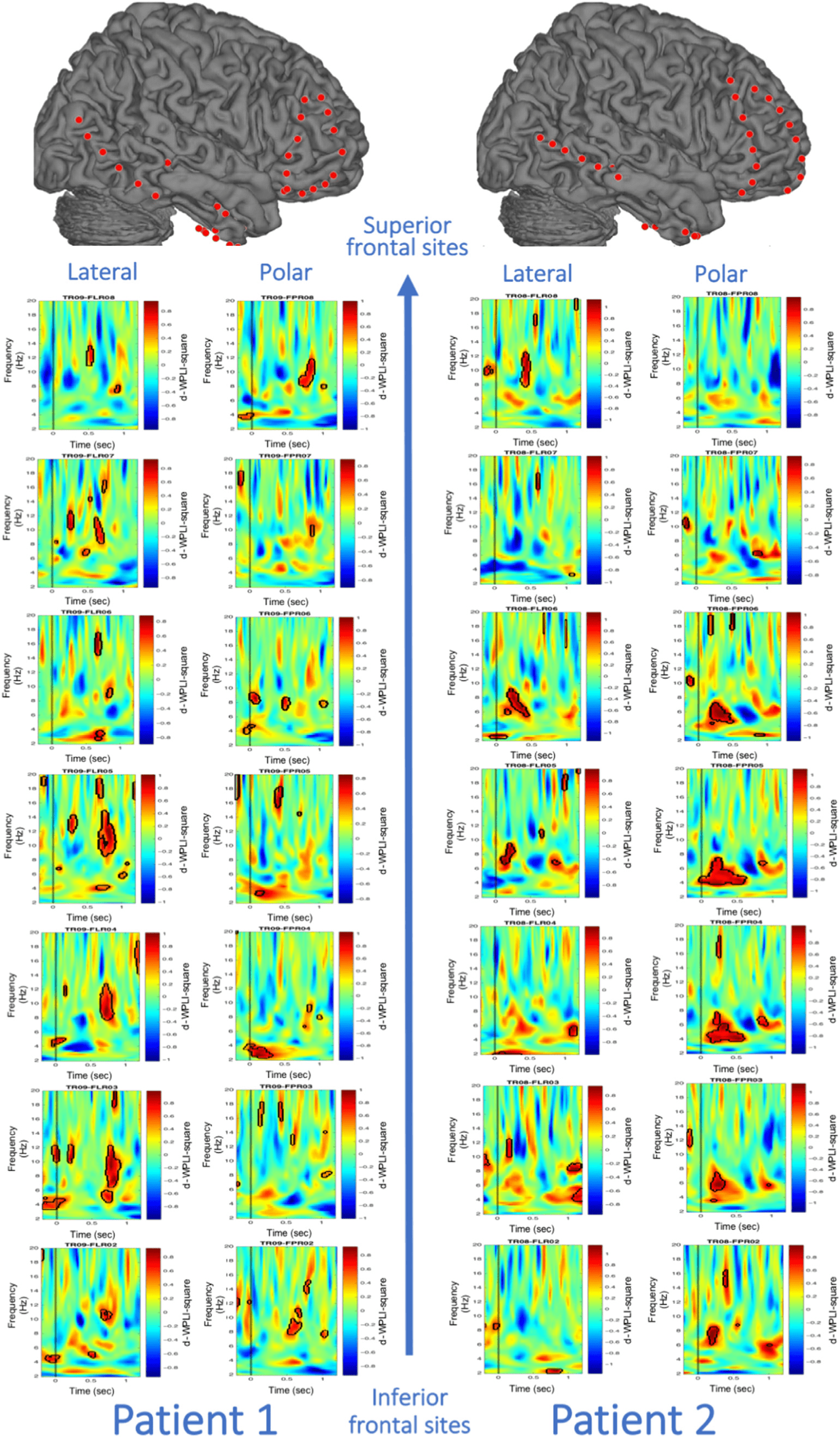
In a second set of analyses, we investigated whether
the increased theta phase synchronization related to con-
textual unexpectedness predicted later memory perfor-
mance. Importantly, in the “contextually unexpected”
condition, across both patients, permutations tests re-
vealed a significant increase in hippocampal–frontopolar
theta phase synchronization for later remembered com-
pared with later forgotten unexpected information
(Figure 4A). As such, the significant cluster of the contex-
tually unexpected subsequent memory effect overlapped
with the significant time–frequency cluster of the con-
textually unexpected–expected contrast (see Figure 4
for the same hippocampal–frontopolar electrode pairs
shown in Figure 3). In contrast, the subsequent memory
analysis for the “contextually expected” condition only
showed smaller significant clusters that did not overlap
in the time–frequency domain with the original clusters
from the contextually unexpected–expected contrast
(Figure 4B). For completeness, Figure A2 shows all 14
electrode pairs for the encoding-related phase synchro-
nization in the contextually unexpected condition.
To examine whether the theta phase synchrony effects
were specific to hippocampal–PFC interactions, we per-
formed control analyses in which we quantified theta
phase synchrony between rhinal and PFC electrodes.
We selected an electrode contact for each patient from
the rhinal cortex (perirhinal/entorhinal cortex) based on
the smallest Euclidean distance between rhinal contacts
in both patients resulting in a 9-mm distance between
both patients (distance between rhinal and hippocampal
contact: 41 and 36 mm for Patients 1 and 2, respectively).
Importantly, permutation tests that corrected for multi-
ple comparisons across electrode pairs revealed that
the frontal electrodes that showed increases in theta
phase synchrony with the hippocampus did not show
Gruber et al.
1649
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
1
1
6
4
6
1
7
8
7
8
6
9
/
/
j
o
c
n
_
a
_
0
1
3
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
1
1
6
4
6
1
7
8
7
8
6
9
/
/
j
o
c
n
_
a
_
0
1
3
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Increases in hippocampal–prefrontal theta phase synchrony for contextually unexpected compared with expected information for one selected
frontopolar electrode per patient (highlighted in yellow). In both patients, frontopolar and dorsolateral prefrontal electrode sites showed significantly
increased theta phase synchrony with the hippocampus during encoding of contextually unexpected compared with expected items (see Figure A1 for all
14 hippocampal-PFC electrode pairs). Phase synchrony was measured via the debiased WPLI-square estimator (d-WPLI) ( Vinck et al., 2011). Top row
depicts findings for Patient 1 and bottom row depicts findings for Patient 2. (A) Significant clusters revealed via permutation tests are depicted with black
contours ( p < .05 family-wise error corrected). (B) Theta phase synchronization was evident for contextually unexpected events and (C) absent in
contextually expected events.
enhanced theta phase synchrony with the rhinal cortex
for contextually unexpected compared with contextually
expected trials.
DISCUSSION
Our study demonstrates that theta phase synchrony
between the hippocampus and PFC is enhanced during
unexpected, contextually deviant events. Moreover, par-
ticularly at sites in the frontopolar cortex, results from both
participants converged in revealing that hippocampal–
frontopolar synchronization predicted later memory
performance. These findings are consistent with the idea
that theta oscillations facilitate communication between
PFC and the hippocampus in support of successful mem-
ory encoding.
Although electrophysiological recording studies in
rodents and nonhuman primates have provided evidence
for task-evoked changes in theta synchronization between
the hippocampus and PFC (Place et al., 2016; Brincat &
Miller, 2015; Fujisawa & Buzsáki, 2011; Benchenane
et al., 2010; Hyman et al., 2005), it is worth noting that
nonhuman and human electrophysiological studies typi-
cally assess synchrony in different ways. Studies in rodents
often measure synchrony via single-unit spiking activity that
is phase-locked to theta oscillations or via amplitude-based
coherence of local field potentials between two regions
(e.g., Benchenane et al., 2010; Jones & Wilson, 2005).
Human studies, in contrast, commonly measure synchrony
via phase alignment of theta oscillations between distant
brain regions (e.g., Kaplan et al., 2017; Backus, Schoffelen,
Szebényi, Hanslmayr, & Doeller, 2016; Watrous et al.,
1650
Journal of Cognitive Neuroscience
Volume 30, Number 11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
1
1
6
4
6
1
7
8
7
8
6
9
/
/
j
o
c
n
_
a
_
0
1
3
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Subsequent memory analyses for the ‘contextually unexpected’ and ‘contextually expected’ condition. Across both patients, permutation
tests revealed strong encoding-related hippocampal-frontopolar theta phase synchronization (same hippocampal-PFC electrode pairs as shown
in Figure 3) in the ‘contextually unexpected’ condition that overlapped with the time-frequency cluster of the previously observed theta phase
synchronization increase for contextually unexpected events (see Figure A2 for all 14 hippocampal–PFC electrode pairs). In contrast, encoding-
related theta phase synchronization in the “contextually expected” condition was limited to small clusters that did not overlap with the previously
reported phase synchronization for unexpected events.
2013). Despite these methodological differences in the
measurement of synchrony, our findings in humans con-
verge with findings in rodents in that they support the
idea that theta synchrony facilitates interactions between
the hippocampus and PFC and thereby facilitates memory
formation.
Our findings are consistent with recent findings in rodents
(Place et al., 2016) that have shown that hippocampal–PFC
phase synchronization represents long-range communica-
tion. On the basis of the findings by Place et al. (2016) that
the mnemonic operation determines the direction of
information flow between the two regions, we speculate
that information flow from the hippocampus to PFC might
underlie the encoding of unexpected events into memory.
However, our analyses do not allow making any claim
about the directionality, and more advanced analyses
would be needed to address this question.
It could be argued that theta synchronization might be
a ubiquitous phenomenon during encoding, but at least
two aspects of our findings are not consistent with this
idea. First, theta synchrony between the two regions
was larger for contextually unexpected compared with
expected events, and second, this synchrony increase
was specific between PFC and the hippocampus but
did not extend to a cortical medial temporal lobe region
(i.e., no evidence for rhinal–PFC theta synchrony). There-
fore, our findings suggest that increased theta synchrony
might be specific to a brain network (involving PFC and
hippocampus) that detects the salience of information
rather than being a ubiquitous property during encoding.
We found an increase in theta phase synchrony dur-
ing an early period during the presentation of a contex-
tually unexpected event. Further control analyses of
time–frequency power for the hippocampus and PFC
contacts did not reveal consistent early theta power in-
creases for contextually unexpected events in the two
patients. It is therefore unlikely that theta power effects
in the two regions drove the phase synchronization find-
ings. However, the early theta hippocampal–PFC syn-
chrony coincides with our previously shown early ERP
finding in the human hippocampus (Axmacher et al.,
2010). Therefore, the increase in theta synchrony be-
tween PFC and the hippocampus, together with this early
hippocampal ERP, might suggest an early detection
Gruber et al.
1651
process that is elicited when expectations are violated
and that the ongoing encoding processes need to be flex-
ibly adapted toward the contextually unexpected infor-
mation (cf. Axmacher et al., 2010). As pointed out in
our earlier study (Axmacher et al., 2010), we cannot rule
out that a third source might have driven the observed
effect between the hippocampus and PFC. For example,
as unexpected information depends on activity within a
cortico-mesolimbic circuit, it would be interesting to test
how other regions within the circuit might affect hippo-
campal–PFC synchrony (e.g., Fujisawa & Buzsáki, 2011;
Benchenane et al., 2010). Because of the sparse implan-
tation scheme of intracranial EEG, this method is not
ideally suited to investigate this question.
Although the understanding of the direct anatomical
connections between the hippocampus and frontopolar
cortex is complicated by the fact that frontopolar cortex
may be differentially organized in humans, as compared
with nonhuman primates or rodents (Semendeferi, Lu,
Schenker, & Damasio, 2002), one possible route could
be via the nucleus reuniens of the ventral midline thala-
mus (Bokor, Csáki, Kocsis, & Kiss, 2002; Herkenham,
1978), which has been shown to support long-term mem-
ory formation (Barker & Warburton, 2018). Alternative
routes could be via the entorhinal cortex and parahippo-
campal cortex/retrosplenial cortex (Ranganath & Ritchey,
2012).
One limitation of this study is that only two patients
had electrodes placed in both the hippocampus and
PFC. It would be beneficial for future studies to investi-
gate this question with a larger sample and a sufficient
number of trials to test the reproducibility of the data.
In addition, future research would need to address how
the observed theta phase synchronization for contex-
tually unexpected information that predicts later memory
generalizes to different forms of salient stimuli (e.g.,
novel or rewarded information).
In conclusion, we have shown that contextually un-
expected information elicits increased theta phase syn-
chrony between the hippocampus and frontopolar cortex,
and this increase in theta phase synchrony is associated
with successful memory formation. Consistent with the
literature on the relationship between theta activity
and memory (for reviews, see Hsieh & Ranganath, 2014;
Düzel, Penny, & Burgess, 2010), we suggest that theta
synchrony between the hippocampus and PFC may be
an important neural mechanism that helps to facilitate
memory formation of novel, unexpected information.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
1
1
6
4
6
1
7
8
7
8
6
9
/
/
j
o
c
n
_
a
_
0
1
3
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
1652
Journal of Cognitive Neuroscience
Volume 30, Number 11
APPENDIX
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
1
1
6
4
6
1
7
8
7
8
6
9
/
/
j
o
c
n
_
a
_
0
1
3
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure A1. Increase in theta phase synchronization for contextually unexpected compared with contextually expected events between the
hippocampus and frontopolar and dorsolateral PFC electrode sites. All selected 14 hippocampal–PFC electrode pairs are shown. Significant clusters
revealed via permutation tests are depicted with black contours.
Gruber et al.
1653
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
1
1
6
4
6
1
7
8
7
8
6
9
/
/
j
o
c
n
_
a
_
0
1
3
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure A2. Encoding-related increase in theta phase synchronization in the “contextually unexpected” condition between the hippocampus and
frontopolar electrode sites across both patients. All selected 14 hippocampal–PFC electrode pairs are shown. Significant clusters revealed via
permutation tests are depicted with black contours.
1654
Journal of Cognitive Neuroscience
Volume 30, Number 11
Acknowledgments
Work on this paper was supported by a German Research
Foundation (DFG) Postdoctoral Fellowship (2014–2016) and a
COFUND Early Career Fellowship from the European Commis-
sion and the Welsh Government (2016–curent) for M. J. G., a
NARSAD Young Investigator grant from the Brain & Behavior
Research Foundation for L.-T. H., a Wellcome Trust/Royal Soci-
ety Sir Henry Dale Fellowship for B. P. S. (107672/Z/15/Z), and
funding via SFB874, SFB1280, and DFG project AX82/3 for N. A.
Reprint requests should be sent to Matthias J. Gruber, School of
Psychology, Cardiff University, Park Place, Cardiff CF10 3AT,
United Kingdom, or via e-mail: GruberM@cardiff.ac.uk.
REFERENCES
Anderson, K. L., Rajagovindan, R., Ghacibeh, G. A., Meador, K. J.,
& Ding, M. (2010). Theta oscillations mediate interaction
between prefrontal cortex and medial temporal lobe in
human memory. Cerebral Cortex, 20, 1604–1612.
Axmacher, N., Cohen, M. X., Fell, J., Haupt, S., Dümpelmann,
M., Elger, C. E., et al. (2010). Intracranial EEG correlates
of expectancy and memory formation in the human
hippocampus and nucleus accumbens. Neuron, 65, 541–549.
Backus, A. R., Schoffelen, J.-M., Szebényi, S., Hanslmayr, S., &
Doeller, C. F. (2016). Hippocampal–prefrontal theta oscillations
support memory integration. Current Biology, 26, 450–457.
Barker, G. R. I., & Warburton, E. C. (2018). A critical role for the
nucleus reuniens in long-term, but not short-term associative
recognition memory formation. Journal of Neuroscience,
38, 3208–3217.
Benchenane, K., Peyrache, A., Khamassi, M., Tierney, P. L.,
Gioanni, Y., Battaglia, F. P., et al. (2010). Coherent theta
oscillations and reorganization of spike timing in the
hippocampal-prefrontal network upon learning. Neuron,
66, 921–936.
Bokor, H., Csáki, A., Kocsis, K., & Kiss, J. (2002). Cellular
architecture of the nucleus reuniens thalami and its putative
aspartatergic/glutamatergic projection to the hippocampus
and medial septum in the rat. European Journal of
Neuroscience, 16, 1227–1239.
Brincat, S. L., & Miller, E. K. (2015). Frequency-specific
hippocampal–prefrontal interactions during associative
learning. Nature Neuroscience, 18, 576–581.
Bunzeck, N., & Düzel, E. (2006). Absolute coding of stimulus
novelty in the human substantia nigra/ VTA. Neuron, 51,
369–379.
Cavanagh, J. F., Figueroa, C. M., Cohen, M. X., & Frank, M. J.
(2012). Frontal theta reflects uncertainty and unexpectedness
during exploration and exploitation. Cerebral Cortex, 22,
2575–2586.
Cavanagh, J. F., & Frank, M. J. (2014). Frontal theta as a
mechanism for cognitive control. Trends in Cognitive
Sciences, 18, 414–421.
Chen, J., Dastjerdi, M., Foster, B. L., LaRocque, K. F., Rauschecker,
A. M., Parvizi, J., et al. (2013). Human hippocampal increases
in low-frequency power during associative prediction violations.
Neuropsychologia, 51, 2344–2351.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21.
Donnelly, N. A., Holtzman, T., Rich, P. D., Nevado-Holgado,
A. J., Fernando, A. B. P., Van Dijck, G., et al. (2014). Oscillatory
activity in the medial prefrontal cortex and nucleus accumbens
correlates with impulsivity and reward outcome. PLoS One,
9, e111300.
Düzel, E., Penny, W. D., & Burgess, N. (2010). Brain
oscillations and memory. Current Opinion in Neurobiology,
20, 143–149.
Elhalal, A., Davelaar, E. J., & Usher, M. (2014). The role of the
frontal cortex in memory: An investigation of the Von Restorff
effect. Frontiers in Human Neuroscience, 8, 410.
Fell, J., & Axmacher, N. (2011). The role of phase
synchronization in memory processes. Nature Reviews
Neuroscience, 12, 105–118.
Fuentemilla, L., Barnes, G. R., Duzel, E., & Levine, B. (2014).
Theta oscillations orchestrate medial temporal lobe and
neocortex in remembering autobiographical memories.
Neuroimage, 85, 730–737.
Fujisawa, S., & Buzsáki, G. (2011). A 4 Hz oscillation adaptively
synchronizes prefrontal, VTA, and hippocampal activities.
Neuron, 72, 153–165.
Grady, C. L., McIntosh, A. R., & Craik, F. I. M. (2003). Age-related
differences in the functional connectivity of the hippocampus
during memory encoding. Hippocampus, 13, 572–586.
Gruber, M. J., Watrous, A. J., Ekstrom, A. D., Ranganath, C.,
& Otten, L. J. (2013). Expected reward modulates
encoding-related theta activity before an event. Neuroimage,
64, 68–74.
Harper, J., Malone, S. M., & Iacono, W. G. (2017). Theta- and
delta-band EEG network dynamics during a novelty oddball
task. Psychophysiology, 54, 1590–1605.
Hasselmo, M. E., Bodelón, C., & Wyble, B. P. (2002). A proposed
function for hippocampal theta rhythm: Separate phases of
encoding and retrieval enhance reversal of prior learning.
Neural Computation, 14, 793–817.
Herkenham, M. (1978). The connections of the nucleus reuniens
thalami: Evidence for a direct thalamo-hippocampal pathway
in the rat. Journal of Comparative Neurology, 177, 589–610.
Herweg, N. A., Apitz, T., Leicht, G., Mulert, C., Fuentemilla, L.,
& Bunzeck, N. (2016). Theta–alpha oscillations bind the
hippocampus, prefrontal cortex, and striatum during
recollection: Evidence from simultaneous EEG–fMRI.
Journal of Neuroscience, 36, 3579–3587.
Hsieh, L.-T., & Ranganath, C. (2014). Frontal midline theta
oscillations during working memory maintenance and
episodic encoding and retrieval. Neuroimage, 85, 721–729.
Hyman, J. M., Zilli, E. A., Paley, A. M., & Hasselmo, M. E. (2005).
Medial prefrontal cortex cells show dynamic modulation
with the hippocampal theta rhythm dependent on behavior.
Hippocampus, 15, 739–749.
Jones, M. W., & Wilson, M. A. (2005). Theta rhythms coordinate
hippocampal–prefrontal interactions in a spatial memory
task. PLoS Biology, 3, e402.
Kahana, M. J., Sekuler, R., Caplan, J. B., Kirschen, M., & Madsen,
J. R. (1999). Human theta oscillations exhibit task dependence
during virtual maze navigation. Nature, 399, 781–784.
Kaplan, R., Bush, D., Bisby, J. A., Horner, A. J., Meyer, S. S., &
Burgess, N. (2017). Medial prefrontal–medial temporal theta
phase coupling in dynamic spatial imagery. Journal of
Cognitive Neuroscience, 29, 507–519.
Lee, G.-T., Lee, C., Kim, K. H., & Jung, K.-Y. (2014). Regional
and inter-regional theta oscillation during episodic novelty
processing. Brain and Cognition, 90, 70–75.
Lisman, J. E., & Grace, A. A. (2005). The hippocampal–VTA
loop: Controlling the entry of information into long-term
memory. Neuron, 46, 703–713.
Murty, V. P., & Adcock, R. A. (2014). Enriched encoding: Reward
motivation organizes cortical networks for hippocampal
detection of unexpected events. Cerebral Cortex, 24,
2160–2168.
Murty, V. P., Ballard, I. C., Macduffie, K. E., Krebs, R. M., &
Adcock, R. A. (2013). Hippocampal networks habituate as
novelty accumulates. Learning and Memory, 20, 229–235.
Gruber et al.
1655
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
1
1
6
4
6
1
7
8
7
8
6
9
/
/
j
o
c
n
_
a
_
0
1
3
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Nee, D. E., & Jonides, J. (2008). Neural correlates of access to
short-term memory. Proceedings of the National Academy
of Sciences, U.S.A., 105, 14228–14233.
O’Neill, P. K., Gordon, J. A., & Sigurdsson, T. (2013). Theta
oscillations in the medial prefrontal cortex are modulated
by spatial working memory and synchronize with the
hippocampus through its ventral subregion. Journal of
Neuroscience, 33, 14211–14224.
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M. (2011).
FieldTrip: Open source software for advanced analysis
of MEG, EEG, and invasive electrophysiological data.
Computational Intelligence and Neuroscience, 2011, 156869.
Place, R., Farovik, A., Brockmann, M., & Eichenbaum, H. (2016).
Bidirectional prefrontal-hippocampal interactions support
context-guided memory. Nature Neuroscience, 19, 992–994.
Raghavachari, S., Lisman, J. E., Tully, M., Madsen, J. R., Bromfield,
E. B., & Kahana, M. J. (2006). Theta oscillations in human
cortex during a working-memory task: Evidence for local
generators. Journal of Neurophysiology, 95, 1630–1638.
Ranganath, C., Heller, A., Cohen, M. X., Brozinsky, C. J., &
Rissman, J. (2005). Functional connectivity with the
hippocampus during successful memory formation.
Hippocampus, 15, 997–1005.
Ranganath, C., & Rainer, G. (2003). Neural mechanisms for
detecting and remembering novel events. Nature Reviews
Neuroscience, 4, 193–202.
Ranganath, C., & Ritchey, M. (2012). Two cortical systems for
memory-guided behaviour. Nature Reviews Neuroscience,
13, 713–726.
Schomaker, J., Berendse, H. W., Foncke, E. M. J., van der Werf,
Y. D., van den Heuvel, O. A., Theeuwes, J., et al. (2014).
Novelty processing and memory formation in Parkinson’s
disease. Neuropsychologia, 62, 124–136.
Semendeferi, K., Lu, A., Schenker, N., & Damasio, H. (2002).
Humans and great apes share a large frontal cortex. Nature
Neuroscience, 5, 272–276.
Strange, B. A., & Dolan, R. J. (2001). Adaptive anterior hippocampal
responses to oddball stimuli. Hippocampus, 11, 690–698.
Totah, N. K. B., Jackson, M. E., & Moghaddam, B. (2013).
Preparatory attention relies on dynamic interactions between
prelimbic cortex and anterior cingulate cortex. Cerebral
Cortex, 23, 729–738.
Vinck, M., Oostenveld, R., van Wingerden, M., Battaglia, F.,
& Pennartz, C. M. A. (2011). An improved index of
phase-synchronization for electrophysiological data in
the presence of volume-conduction, noise and sample-size
bias. Neuroimage, 55, 1548–1565.
Von Restorff, H. (1933). Über die wirkung von bereichsbildungen
im spurenfeld. Psychologische Forschung, 18, 299–342.
Walsh, M. M., & Anderson, J. R. (2012). Learning from
experience: Event-related potential correlates of reward
processing, neural adaptation, and behavioral choice.
Neuroscience & Biobehavioral Reviews, 36, 1870–1884.
Watrous, A. J., Tandon, N., Conner, C. R., Pieters, T., & Ekstrom,
A. D. (2013). Frequency-specific network connectivity
increases underlie accurate spatiotemporal memory retrieval.
Nature Neuroscience, 16, 349–356.
Winson, J. (1978). Loss of hippocampal theta rhythm results in
spatial memory deficit in the rat. Science, 201, 160–163.
Yassa, M. A., & Stark, C. E. L. (2008). Multiple signals of
recognition memory in the medial temporal lobe.
Hippocampus, 18, 945–954.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
1
1
1
6
4
6
1
7
8
7
8
6
9
/
/
j
o
c
n
_
a
_
0
1
3
0
2
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
1656
Journal of Cognitive Neuroscience
Volume 30, Number 11