La semántica de la sintaxis: La base de lo transitivo
and Intransitive Constructions
Wessel O. van Dam and Rutvik H. Desai
Abstracto
■ Embodied theories of language maintain that brain areas as-
sociated with perception and action are also involved in the
processing and representation of word meaning. A number of
studies have shown that sentences with action verbs elicit ac-
tivation within sensory–motor brain regions, arguing that
sentence-induced mental simulations provide a means for
grounding their lexical-semantic meaning. Constructionist the-
ories argue, sin embargo, that form–meaning correspondence is
present not only at the lexical level but also at the level of
constructions. We investigated whether sentence-induced mo-
tor resonance is present for syntactic constructions. We mea-
sured the BOLD signal while participants read sentences with
(di)transitive (caused motion) or intransitive constructions that
contained either action or abstract verbs. The results showed a
distinct neuronal signature for caused motion and intransitive
syntactic frames. Caused motion frames activated regions asso-
ciated with reaching and grasping actions, including the left
anterior intraparietal sulcus and the parietal reach region. En-
transitive frames activated lateral temporal regions commonly
associated with abstract word processing. The left pars orbitalis
showed an interaction between the syntactic frame and verb
class. These findings show that sensory–motor activation elic-
ited by sentences entails both motor resonance evoked by sin-
gle words as well as at the level of syntactic constructions. ■
INTRODUCCIÓN
Traditionally, perceptual and motor brain systems were
seen as categorically distinct from our representational
systems subserving cognition. In this symbolic view, estafa-
ceptual representations were taken to be of a symbolic
and amodal nature and lexical-semantic meaning inde-
pendent of sensory–motor systems of the brain (Kintsch,
2008; Pylyshyn, 1984; Fodor, 1983; katz & Fodor, 1963).
Opponents of such a symbolic account have raised the
problem of how symbols can be mapped to their refer-
ents in the real world (Harnad, 1990) and its failure to
provide an adequate description of the transduction pro-
cess that maps perceptual states to amodal conceptual
symbols (Barsalou, 1999). In contrast to such a symbolic
cuenta, embodied theories argue that experiential
traces stored in sensory–motor brain areas can pro-
vide the means for grounding lexical-semantic meaning
(Barsalou, 2008; Pulvermüller, 1999; Glenberg, 1997). En
la última década, a plethora of studies using various ex-
perimental techniques have directly contrasted predic-
tions from symbolic and embodied accounts. Findings
from behavioral experiments suggest that processes that
play a role in perception and action are also involved in
comprehending language about action (see Fisher &
Zwaan, 2008; Zwaan & Kaschak, 2008, for reviews). en un
similar vein, fMRI studies have shown that comprehen-
sion of action verbs, action sentences, and words denot-
University of South Carolina
© 2016 Instituto de Tecnología de Massachusetts
ing manipulable objects frequently activate sensory–
motor brain areas (see Kiefer & Pulvermüller, 2012;
Meteyard, Cuadrado, Bahrami, & Vigliocco, 2012; Binder
& Desai, 2011, for reviews).
This work has focused almost exclusively on meanings
of nouns and verbs, either isolated or embedded in sen-
tenencias. Several studies that use sentences compared
action-related sentences with abstract sentences that have
an identical or similar sentence structure (p.ej., Desai,
Conant, Binder, Parque, & Seidenberg, 2013; Desai, Binder,
Conant, Mano, & Seidenberg, 2011; Desai, Binder,
Conant, & Seidenberg, 2009; Raposo, Moss, Stamatakis,
& tyler, 2009; Aziz-Zadeh, wilson, Rizzolatti, & Iacoboni,
2006; Tettamanti et al., 2005). Por ejemplo, Tettamanti
et al. (2005) showed that listening to sentences that de-
scribe actions (p.ej., “I bite an apple”) as compared with
similar transitive sentences with abstract content (p.ej.,
“I appreciate sincerity”) activated parts of the motor system.
These findings have been taken to show that, to convey
the meaning of a sentence as a whole, we rely on ex-
periential traces of the referent action stored in sensory–
motor brain areas. Además, recent studies have
suggested that conceptual features contribute to word
and sentence level meaning in a flexible and context-
dependent manner (Kemmerer, 2015; De la presa, Van Dijk,
Bekkering, & Rueschemeyer, 2012; Hoenig, Sim, Bochev,
Herrnberger, & Kiefer, 2008). Sin embargo, in most (if not all)
of the studies in the conceptual representation literature,
the effects of sentence structure are subtracted out by
diseño.
Revista de neurociencia cognitiva 28:5, páginas. 693–709
doi:10.1162/jocn_a_00926
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
oh
4
C
7
norte
0
_
7
a
/
_
j
0
oh
0
C
9
norte
2
6
_
a
pag
_
d
0
0
b
9
y
2
gramo
6
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
i
2
3
mi
s
/
j
.
/
t
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Traditionally, it is assumed that argument structures con-
vey their meaning by means of the main verb. Por ejemplo,
in “Pat gave Chris a cake,” the meaning of transfer apparent
in the overall expression is specified by the three-argument
verb “give.” That is, the three-argument verb “give” is ex-
pected to appear with phrases corresponding to the three
characters required in the act of giving: a giver, a recipient,
and something that is given. Por lo tanto, it can be argued
that the interpretation of this sentence pattern as a
whole can be determined on the basis of its main verb.
Sin embargo, in many instances, the main verb does not re-
liably determine the interpretation of a sentence. por ejemplo-
amplio, “Liza bought a book for Zach” can mean that Liza
bought a book for a third person because Zach did not
have time to buy it himself. Sin embargo, “Liza bought Zack
a book” can only mean that Liza intended to give the
book to Zach (Goldberg, 2003). Eso es, although both
sentences involve the same verb “to buy,” the interpreta-
tion of these sentences is not reliably determined by its
independent specification. The verb-centric view deals
with such cases by proposing that verbs have multiple
distinct senses because of the different linking patterns
between syntax and semantics (Levin, 1995; Pinker,
1989). The main criticism of this view is that it leads to
a proliferation of polysemous lexical entries (Goldberg,
1995).
An alternative is provided by a family of theories
known as “contructionist” approaches (p.ej., Bergen &
Chang, 2003; Goldberg, 1995, 2003; Croft, 2001; Langacker,
1999; Fillmore, kay, & O’Connor, 1988; Lakoff, 1987). Estafa-
structions are stored pairings of form and function, incluir-
ing words, idioms, or linguistic patterns that may be fully or
partially filled. A central idea in these approaches is that
constructions themselves have meaning. Eso es, form–
meaning correspondences do not only exist at the level
of verbs or single words but also at the level of construc-
ciones. Constructions are pairings of form and meaning,
and therefore, a difference in syntactic form always spells
a difference in meaning (Langacker, 1999; Wierzbicka,
1988; Haiman, 1985; bolinger, 1968). This idea that argu-
ment structure constructions themselves carry meaning is
often referred to as the “principle of no synonymy of gram-
matical forms” (bolinger, 1968).
Given that embodiment theories reject the idea that
language can convey meaning without it somehow being
grounded in real-world sensory–motor experiences, este
would suggest that semantics of constructions should be
grounded in a similar fashion. In line with this idea, recent
theorists have argued that sensory–motor activation elic-
ited by sentences might be a complex phenomenon that
entails both motor resonance evoked by single words
and word combinations, but also more general motor res-
onance at the level of constructions (Kemmerer, 2006).
Bergen and Chang (2003), in their Embodied Construction
Grammar approach, argue that syntactic constructions can
drive specific perceptual and motor simulations of a lan-
guage user. Por ejemplo, the directed motion construction
may lead to the mental simulation of an event involving an
animate mover moving along a path, even if the main verb
in the construction does not explicitly specify motion (as in
“The cat meowed down the street”). Experimental evi-
dence for the principle of no synonymy of grammatical
forms has been provided by, Por ejemplo, work showing
that comprehenders access different meanings for sen-
tences using transitive versus ditransitive constructions
(Kaschak & Glenberg, 2000). A ditransitive construction
is a construction consisting of a verb, an agent argument,
a recipient-like argument, and a theme argument (p.ej., “He
threw the ball to John”). It can be argued that, Por ejemplo,
the ditransitive (double-object) construction activates
a transfer-of-possession schema (X causes Y to have
z), whereas a prepositional dative may activate a caused-
motion schema (Goldberg, 1995).
These findings mesh with evidence from action observa-
ción, which suggests that the transitivity of an action is a
crucial factor in determining the type of simulations. El
finding that some mirror neurons of the macaque monkey
were only responsive to object-directed actions suggests
that activation of the parietal-frontal network depends on
whether an action is goal directed (Tkach, Reimer, &
Hatsopoulos, 2007; Gallese, Fadiga, Fogassi, & Rizzolatti,
1996; Rizzolatti, Fadiga, Gallese, & Fogassi, 1996).
A number of fMRI studies also suggest a neural disso-
ciation between these two types of actions. The inferior
frontal gyrus (IFG), lateral precentral gyrus, inferior pari-
etal lobule (IPL), posterior middle temporal gyrus (MTG),
and the posterior part of the superior parietal lobule have
been shown to play a role in the representation of tran-
sitive actions (Caspers, Zilles, Laird, & Eickhoff, 2010;
Sakreida, Schubotz, Wolfensteller, & por cramon, 2005;
Buccino et al., 2001). Intransitive actions, en el otro
mano, seem to rely more on posterior regions in the pa-
rietal cortex, the angular gyrus (AG), and STS regions
(Grosbras, Beaton, & Eickhoff, 2012; Lui et al., 2008). Re-
cruitment of partially different brain regions might reflect
the fact that transitive actions are object related and tend
to be more complex (p.ej., “throwing a ball”), whereas in-
transitive actions are not related to an object (p.ej., “waving
goodbye”). Additional evidence for a neural dissociation
between transitive and intransitive actions comes from
patient studies. Por ejemplo, watson, Fleet, Rothi, y
Heilman (1986) observed an isolated disturbance of tran-
sitive but not intransitive movements in bilateral apraxia
(see also Mozaz, Rothi, anderson, Crucian, & Heilman,
2002; Foundas et al., 1999).
We hypothesized that mental simulations in service of
language comprehension will be modulated by whether a
construction (or syntactic frame) is of a transitive (caused
movimiento) or intransitive nature. En el estudio actual, nosotros
investigated whether distinct patterns of sensory–motor ac-
tivation can be observed for different syntactic construc-
ciones, independent of the main verb. We also investigated
if language-induced mental simulations differ as a function
of the class of main verb used within the construction and
694
Revista de neurociencia cognitiva
Volumen 28, Número 5
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
oh
4
C
7
norte
0
_
7
a
/
_
j
0
oh
0
C
9
norte
2
6
_
a
pag
_
d
0
0
b
9
y
2
gramo
6
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
i
2
3
mi
s
/
j
F
t
/
.
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
whether the factors of construction and verb class interact
with each other.
Para tal fin, we measured changes in the hemodynamic
response of participants while they read transitive and in-
transitive sentences that either contained a concrete or ab-
stract main verb. We used a specific type of transitive
oraciones; transitives were defined here as a construction
with the syntactic realization (NP V NP PP). Such a transitive
construction is characterized by a transitive verb, an agent
argumento, a recipient-like argument, and a theme argu-
mento (Malchukov, Haspelmath, & Comrie, 2010). El
schematic constructional meaning of this phrase is that
of the causer argument directly causing the theme argu-
ment to move toward a recipient-like argument (X causes
Y to move to Z), when used with a concrete verb. Tenga en cuenta que
the schematic constructional meaning is closely related to
the transfer-of-possession schema (X causes Y to have Z),
with the difference that the sentence “You threw the ball to
her” does not actually entail that the recipient-like argu-
ment receives the ball (es decir., a change of possession), Alabama-
though that may typically be the case. Throughout the
manuscript, we refer to our main constructions of interest
as transitive (caused motion) and intransitive construc-
ciones. It needs to be noted that these terms are very broad
in nature and actually embrace a whole family of construc-
tions with more specific meanings. Por ejemplo, a more
specific label for the construction used in our transitive
sentences would be the “caused motion construction.”
Por lo tanto, to be more specific, we will adopt the term
“caused motion construction” from here on.
We hypothesized a main effect of verb class, with sen-
tences containing a concrete/action-related main verb eli-
citing stronger activation in the anterior IPL, primary
motor cortex, premotor cortex, posterior MTG and infe-
rior temporal gyrus (ITG), superior frontal gyrus (SFG),
and IFG. Por otro lado, for sentences containing an
abstract main verb, we expect stronger activation within
the anterior and middle STS (Desai et al., 2009, 2011,
2013). Además, given that transitive sentences are
strongly associated with a caused motion (X causes Y to
move Z) schema (and take a direct object), we expect
these sentences to elicit stronger activation in regions
within the posterior middle/inferior temporal cortex that
are involved in the processing of motion (Chen, Widick,
& Chatterjee, 2008) and within the anterior IPL, a brain
region that is involved in representing complex hand–
object interactions and tool use. Stronger activation for
intransitive sentences is hypothesized in the anterior and
middle lateral temporal lobes, given that these regions
are associated with more abstract semantics.
MÉTODOS
Participantes
Fourteen individuals participated in the study, all of whom
were right-handed and between 19 y 22 years old (m =
20.64 years old, DE = 0.93 years old; six men). All partic-
ipants had normal or corrected-to-normal vision and no
history of neurological disorders. Before the experiment,
participants were informed about the experimental pro-
cedures, signed informed consent forms, and were given
practice trials according to a protocol sanctioned by the
institutional review board of the University of South
Carolina. Participants were paid for their participation.
Every participant (excepto por uno) underwent two scans
on two different days within a 1-week period.
Estímulos
The four main experimental conditions contained 35 sen-
tences each. They were (1) concrete caused motion
(CCM; p.ej., “You threw the ball to her”; “He gave the
pizza to you”), (2) concrete intransitive (CI; p.ej., "Tú
waved at her whole family”; “He clapped after your big
concert”), (3) abstract caused motion (ACM; p.ej., "Tú
delegated the task to her”; “He communicated the news
to you”), y (4) abstract intransitive (AI; p.ej., "Tú
thought about her feelings”; “He cared about your new
program”). The caused motion sentences used a ditran-
sitive verb with a preposition. Además, 30 filler sen-
tences were also included (p.ej., “You deduced the
truth”; “He met the person”). The concrete conditions
used an action-related verb, whereas the abstract condi-
tions used verbs that did not have a strong association
with actions. Verb concreteness ratings showed no differ-
ence between the CCM and CI conditions and the ACM
and AI conditions (all ps > .5). Concreteness ratings for
the main verb used in the concrete sentences (m = 3.92)
were significantly higher than for the main verb used in
the abstract sentences (m = 2.48; pag < .001). There were
also pseudoword conditions in the experiment, but we
do not discuss them here and focus on the interpretable
conditions.
The sentences of all experimental conditions were
matched on a number of psycholinguistic variables (see
Table 1). Two-sample t tests ensured that all sentences
were matched on number of words (all ps > .10), total
number of phonemes (all ps > .20), total number of syl-
lables (all ps > .07), total number of letters (all ps > .10),
and the number of persons mentioned in a sentence (ex-
act match). Además, 11 participants performed a be-
havioral version of our experiment in which we collected
RT data for the different sentences in a meaningfulness
judgment task. Paired-sample t tests showed that our
stimuli were not perfectly matched on RTs. Participantes
were faster to respond to ACM (m = 1356 mseg) than
to AI (m = 1506 mseg) oraciones ( pag < .01). Likewise, par-
ticipants were faster to respond to CCM (M = 1404 msec)
than to CI (M = 1532 msec) sentences ( p = .001). In ad-
dition, participants were faster to respond to ACM (M =
1356 msec) than to CI (M = 1532 msec) sentences
( p < .001). A regressor representing the mean-centered
RT for each sentence from the behavioral experiment
van Dam and Desai
695
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
o
4
c
7
n
0
_
7
a
/
_
j
0
o
0
c
9
n
2
6
_
a
p
_
d
0
0
b
9
y
2
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 1. Psycholinguistic Variables
No. of words
No. of phonemes
No. of syllables
Length
Verb concreteness
Noun concreteness
Relative freq verb/tran
Relative freq verb/intran
RT
Log HAL freq
ACM
6.03 (0.17)
21.26 (2.77)
8.60 (1.09)
25.74 (2.60)
2.60 (0.52)
3.28 (0.83)
0.57 (0.20)
0.23 (0.15)
1356 (362)
9.84 (0.82)
AI
5.91 (0.95)
22.23 (3.39)
8.71 (1.56)
27.49 (4.40)
2.38 (0.65)
3.11 (0.95)
0.31 (0.22)
0.49 (0.25)
1506 (411)
9.78 (1.50)
CCM
6.26 (0.82)
20.57 (3.39)
8.03 (1.46)
26.14 (4.53)
3.82 (0.43)
4.70 (0.37)
0.46 (0.16)
0.31 (0.19)
1404 (354)
8.86 (1.10)
CI
6.17 (1.18)
21.37 (3.95)
7.97 (1.93)
27.23 (5.34)
4.01 (0.48)
4.52 (0.56)
0.43 (0.16)
0.35 (0.17)
1532 (382)
9.38 (1.14)
was used as an additional item-wise regressor to account
for variance due to time-on-task and difficulty.
In addition, we calculated the mean concreteness rat-
ing of the nouns of each of the four experimental condi-
tions using the Brysbaert, Warriner, and Kuperman
(2013) database. In the case in which the database did
not have an entry for a compound word (e.g., “dining
table”), we used the concreteness rating for the head
noun (i.e., “table”) of the compound word. Two-sample
t tests revealed an overall difference in the mean noun
concreteness ratings between the concrete sentences
(M = 4.61) and the abstract sentences (M = 3.20; p <
.001). No difference was observed between transitive
(caused motion) and intransitive frames, neither for con-
structions with a concrete verb or an abstract main verb
(Table 1).
A number of studies show, using single-verb stimuli,
that brain activations can be affected by other character-
istics of verb classes, such as whether they allow causative
alternation or number of obligatory arguments. The rele-
vance of these results is not clear in the present case be-
cause we used sentences instead. Single verbs were used
in these experiments as stimuli to examine the dominant
or default response to the verb (e.g., those that require
two arguments vs. three or those with and without caus-
ative alternation). When the argument structure is real-
ized, the salience of these defaults (beyond general
difficulty effects because of low familiarity of certain us-
age that is allowed but is uncommon) can be expected
to reduce. We are not aware of any experiments that
show similar differences between verb classes while using
them in sentences. Nonetheless, we examined three such
variables as a cautionary measure.
First, verbs with causative alternation (those that can
be used both transitively and intransitively) and those
without can potentially differ. A recent study by Meltzer-
Asscher, Schuchard, den Ouden, and Thompson (2012)
suggests that the pattern of neural activity elicited by a
verb is different for verbs, which can alternate between
transitive and intransitive frames and those that cannot.
We used the VALEX database to obtain information on
the relative frequency with which the main verb appears
in a transitive versus intransitive construction (Korhonen,
Krymolowski, & Briscoe, 2006; see Table 1). Two-sample
t tests indicated that caused motion and intransitive sen-
tences differed in the relative frequency with which its
main verb was used in a caused motion versus intransitive
construction ( p < .001). That is, the main verbs in the
caused motion condition occur relatively more often in
caused motion constructions, whereas the main verbs
in the intransitive condition occur relatively more often
in intransitive constructions. Importantly, the concrete
and abstract sentences did not differ in the relative fre-
quency with which its main verb was used transitively
versus intransitively ( p > .50).
A second factor is whether verbs used in the transitive
(caused motion) conditions obligatorily take two or three
argumentos. Some studies have indicated inferior parietal
and posterior superior temporal regions to be involved in
processing argument structure complexity, mostrando que
verbs with more obligatory arguments show a stronger
response than verbs with fewer obligatory arguments
(Thompson, Bonakdarpour, & Fix, 2010; Thompson
et al., 2007; Ben-Shachar, Hendler, Kahn, Ben-Bashat, &
Grodzinsky, 2003). We conducted an analysis on the
number of arguments a verb obligatorily takes by using
VerbNet (Kipper, Dang, & Palmer, 2000). This analysis in-
dicated that there was no difference in the number of ar-
guments a verb obligatorily takes between the CCM (m =
2.11) and ACM (m = 2.17) condiciones ( pag = .50).
Correspondingly, intransitive verbs have been subcate-
gorized as unergatives and unaccusatives. Both verb
types have the same number of arguments (es decir., a single
argumento) but differ syntactically. Unergatives and unac-
cusatives are often delineated in terms of whether the
verb takes an internal versus external argument. Such a
696
Revista de neurociencia cognitiva
Volumen 28, Número 5
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
oh
4
C
7
norte
0
_
7
a
/
_
j
0
oh
0
C
9
norte
2
6
_
a
pag
_
d
0
0
b
9
y
2
gramo
6
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
i
2
3
mi
s
/
j
.
F
t
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
distinction is, sin embargo, associated with a syntactic move-
ment analysis, which is only one of many approaches
used in linguistic studies of unaccusativity (see Roehm,
Sorace, & Bornkessel-Schlesewsky, 2012). Different ap-
proaches in this branch of linguistics tend to agree that,
in an unaccusative construction, the participant is likely
to be a patient or a nonvolitional causer of the event,
whereas in the unergative construction, some property
inherent in the argument of the verb is responsible for
bringing about the event. Some studies with healthy par-
ticipants and brain-damaged patients suggest that the
neural structures that underlie the processing of these
subtypes of intransitive verbs are dissociable (Agnew,
van de Koot, McGettigan, & Scott, 2014; Shetreet &
Friedmann, 2012; McAllister, Bachrach, Waters, Michaud,
& Caplan, 2009; Luzzatti et al., 2002). Aquí, there was no
difference in the number of unergative verbs between
the CI (norte = 5) and AI (norte = 3) condiciones ( pag = .46). Sev-
eral recent authors, sin embargo, have argued that the use of
the terms “unergative” and “unaccusative” might be
problematic, given that this distinction in intransitive
verbs cannot account for the acceptability of the use of
certain syntactic constructions (for an extensive discus-
sión, see Kuno & Takami, 2004). Además, Ha sido
argued that the labels “unergative” and “unaccusative”
are used to describe a wide variety of phenomena in dif-
ferent languages and therefore render them vague and
imprecise (dixon, 2010).
Procedimiento
The order of stimulus presentation was randomized indi-
vidually for each participant. A single trial lasted 7.4 segundo
and constituted of the presentation of a single sentence.
A variable jitter time of 0, 500, 1000, o 1500 msec was
included at the beginning of each trial to enhance the
temporal resolution of the acquired signal. After this, a
fixation cross was presented in the center of the screen.
A 400-msec blank screen followed the fixation cross; allá-
después, the sentence appeared on screen for 3000 mseg. El
time between offset of the sentence and onset of the next
trial was filled with a blank screen, to ensure that each trial
lasted exactly 7.4 segundo. Participants were instructed to read
all sentences carefully to be able to answer a surprise yes/no
question after some of the sentences (responses were
made by performing a right-hand button click with their
thumb). Thirty comprehension questions (p.ej., “Did he
clap after your big concert?") were used. This catch trial
design was used to encourage participants to read all sen-
tences for comprehension while avoiding motor execution
after every trial.
fMRI Data Acquisition
Functional images were acquired on a Siemens TRIO
3.0-T MRI system (Siemens, Erlangen, Alemania) equipado
with a 12-channel head coil. BOLD-sensitive functional
images were acquired using a single-shot gradient EPI se-
quence (echo time/repetition time = 34/1850 mseg, 34 ax-
ial slices in ascending order, slice gap = 0.60 mm, field of
view = 208 mm, flip angle = 90°, voxel size = 3.25 ×
3.25 × 3.60 mm3). High-resolution anatomical images were
acquired using a magnetization prepared rapid gradient-
echo sequence (echo time = 4.15 mseg, voxel size =
1 × 1 × 1 mm3, 192 sagittal slices, campo de visión = 256 mm).
fMRI Data Analysis
Functional data were preprocessed and analyzed with the
AFNI software package (Cox, 1996). A standardized pre-
processing pipeline involved registration of functional im-
ages to the anatomy (Saad et al., 2009). Después,
functional images were co-registered (Cox & Jesmanowicz,
1999) and projected into standard stereotaxic space
(Talairach & Tournoux, 1988). The normalized images
were smoothed with an isotropic 5-mm FWHM Gaussian
kernel, and the run mean of each voxel was scaled to
100. The ensuing preprocessed fMRI time series were ana-
lyzed on a participant-by-participant basis using an event-
related approach in the context of voxelwise multiple linear
regression with regressors for each condition (ACM, AI,
CCM, CI,) as well as the filler items and question trials con-
volved with a canonical hemodynamic response function.
Six motion parameters and the signal extracted from the
ventricles were included as noise covariates of no interest.
General linear tests were conducted to obtain the main ef-
fect of Transitivity and Concreteness and the Transitivity ×
Condition interactions.
In a random effects analysis, group maps were created
by comparing activations against a constant value of 0.
The group maps were thresholded at voxelwise p < .01
and corrected for multiple comparisons by removing
clusters smaller than 800 μl to achieve a map-wise cor-
rected two-tailed p < .05.1 Using the 3dClustSim pro-
gram with 10,000 iterations, the cluster threshold was
determined through Monte Carlo simulations that esti-
mate the chance probability of spatially contiguous voxels
exceeding the voxelwise p threshold. The analysis was re-
stricted to a mask that excluded areas outside the brain as
well as deep white matter areas and the ventricles. This
mask is based on the probabilistic Desikan–Killiany atlas
that contains 35 cortical areas in each hemisphere (Desikan
et al., 2006) and the subcortical parcellation provided by
FreeSurfer. In addition to this whole-brain analysis, two
other (bilateral) ROIs were defined for a more sensitive
analysis. One used the primary motor and sensory cortex
(M1 and S1) as defined by the HMAT atlas (Mayka, Corcos,
Leurgans, & Vaillancourt, 2006). Given its association with
transitive actions and action sentence processing, we also
defined a bilateral supramarginal gyrus (SMG) ROI. This
ROI was defined on the basis of the maximum probability
map provided with AFNI, which is based on the Destrieux
atlas (Destrieux, Fischl, Dale, & Halgren, 2010). Small
volume correction was applied in these ROIs to achieve a
van Dam and Desai
697
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
o
4
c
7
n
0
_
7
a
/
_
j
0
o
0
c
9
n
2
6
_
a
p
_
d
0
0
b
9
y
2
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
corrected p < .05, determined in the same manner as
above.
RESULTS
Concrete–Abstract
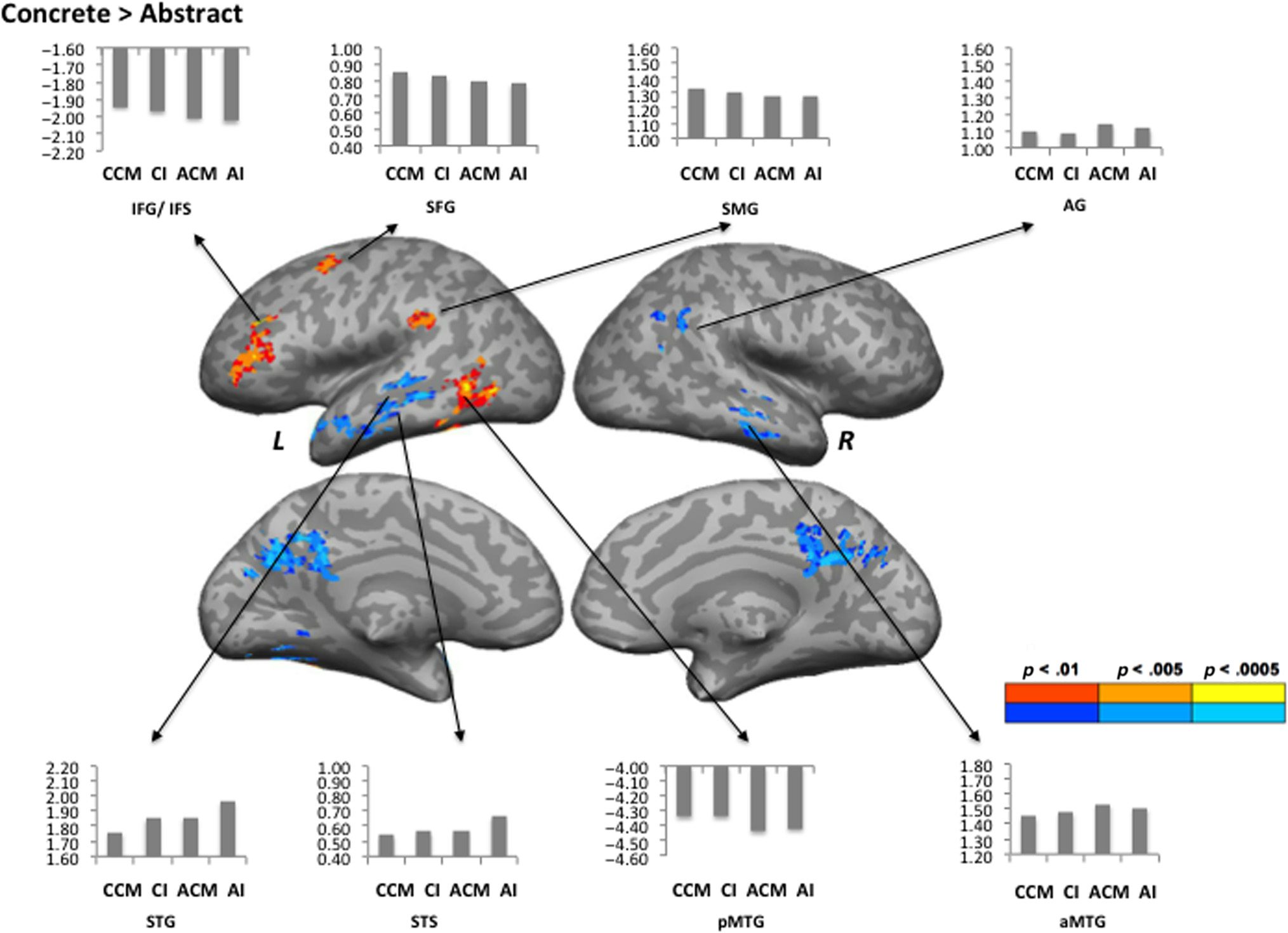
Areas activated to a greater extent by the Concrete con-
dition relative to the Abstract condition included the left
MTG and ITG, superior frontal sulcus, inferior frontal sul-
cus (IFS), and IFG (pars triangularis). The ROI analyses
revealed an additional cluster in the left SMG. The Ab-
stract condition relative to the Concrete condition led
to stronger activation in the left superior temporal gyrus
(STG), STS, fusiform gyrus, right precuneus, MTG, and
AG (Figure 1, Table 2).
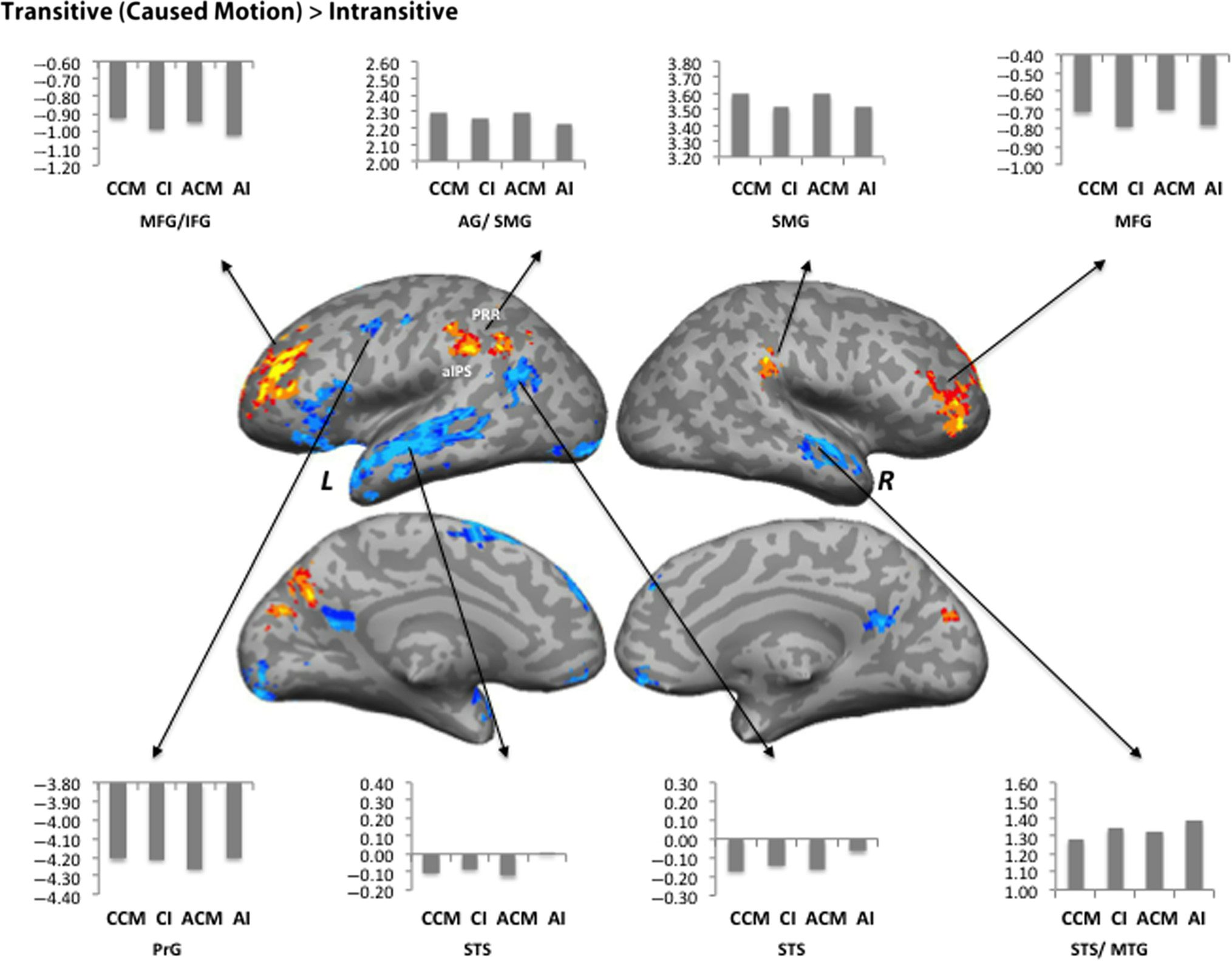
Caused Motion (Transitive)–Intransitive
Caused Motion sentences elicited greater levels of activa-
tion than Intransitive sentences within the bilateral mid-
dle frontal gyrus (MFG), SMG, and left AG as well as
precuneus. Structures more strongly activated by the In-
transitive condition relative to the Caused Motion condi-
tion included bilateral STS and rectal gyrus, left occipital
pole, SFG, and precentral gyrus as well as the right MTG
and precuneus (Figure 2, Table 2).
Overlap
Some similarities as well as differences were found be-
tween the Concrete > Abstract and the Caused Motion >
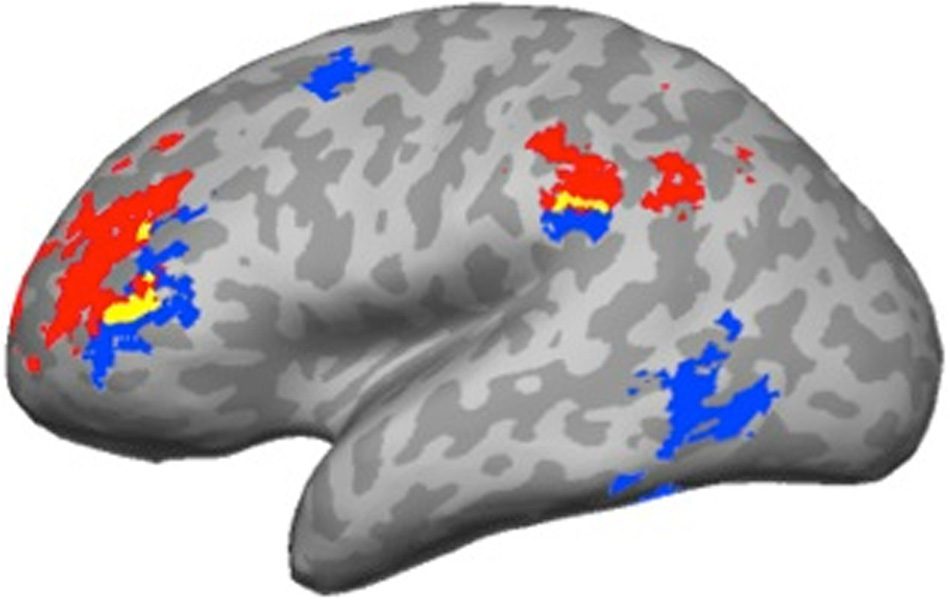
Intransitive contrasts (Cifra 3). Both contrasts activated
the left IFG and IFS (pars triangularis), with the former
extending ventrally and the latter extending dorsally
into MFG. A similar dorsal–ventral pattern was found
in the left IPL. Both contrasts activated the SMG, con
the Concrete > Abstract contrast extending ventrally
into the parietal operculum, whereas the Caused Motion >
Intransitive contrast extending dorsally into the anterior
intraparietal sulcus (IPS).
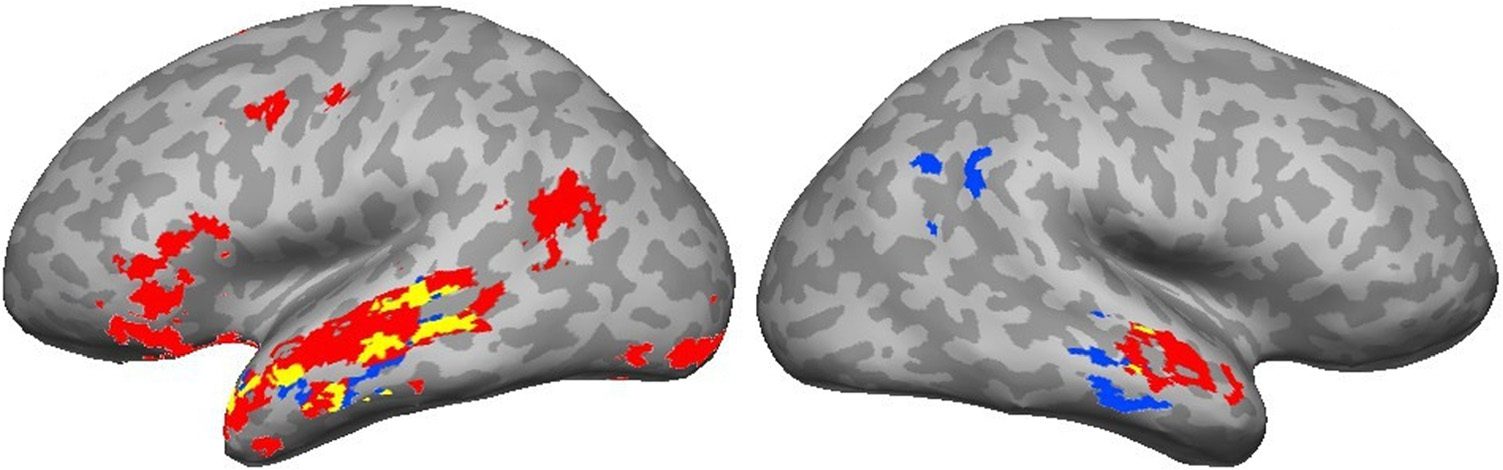
The Abstract > Concrete and Intransitive > Caused Mo-
tion comparisons also showed some similarities (Cifra 4),
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
oh
4
C
7
norte
0
_
7
a
/
_
j
0
oh
0
C
9
norte
2
6
_
a
pag
_
d
0
0
b
9
y
2
gramo
6
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
i
2
3
mi
s
/
j
/
t
.
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Cifra 1. Areas activated by the Concrete–Abstract contrast. Red-orange colors show greater activation for the Concrete condition; blue-cyan colors
show greater activation for the Abstract condition. Mean percent signal change relative to rest is shown for the CCM, CI, ACM, and AI conditions, en un
sphere of 5-mm radius around the peak voxel. L = left hemisphere; R = right hemisphere.
698
Revista de neurociencia cognitiva
Volumen 28, Número 5
Mesa 2. Activations in the Main Contrasts of Interest
Volume Max
X
y
z
Estructura
where the left middle and anterior STS were activated by
both contrasts.
Concrete > Abstract
3078
2268
918
675
3.61 −52 −58
0 L MTG
3.79 −40
3.15 −22
34
1
14 L IFG, IFS (pars triangularis)
44 L sup frontal s
2.95 −49 −34
32 L SMG
Abstract > Concrete
5832
2538
1377
1053
1026
810
−4.20
4 −52
26 R precuneus
−3.95 −52 −37
0 L STS
−4.09 −25 −67 −12 L occ temp g
−3.67
−3.29
61 −19 −9 R MTG
55 −55
20 R AG
−4.35 −49
13 −15 L STG
Caused Motion > Intransitive
4.53 −40
5913
37
26 L MFG
3294
2781
2268
1188
810
3.84 −40 −52
44 L AG, L SMG
3.58
43
46
11 R MFG
3.57 −7 −73
29 L precuneus
3.65
3.70
19
52
17 R MFG
61 −31
32 R SMG
Intransitive > Caused Motion
13986 −4.86 −52 −40
2 L STS
−3.06 −52
22
8 L IFG (pars triangularis)
−3.78 −34
25 −3 L IFG (pars orbitalis)
3915
1890
1674
1647
1620
1323
1188
1134
−3.93 −16 −88 −6 L occipital pole
−3.98 −43 −55
23 L STS
−4.34
52 −1 −12 R STS; R MTG
−4.62 −4
−3.61 −4
4
52
62 L SFG
32 L SFG
−3.50
−3.58
4 −52
11 R precuneus
1
52 −9 L rectus; R rectus
−3.99 −49 −10
44 L precentral g
The volume of the cluster (μl), peak z score, Talairach coordinates,
and the anatomical structures that the clusters overlap are shown. L =
left hemisphere; R = right hemisphere; g = gyrus; s = sulcus; sup =
superior.
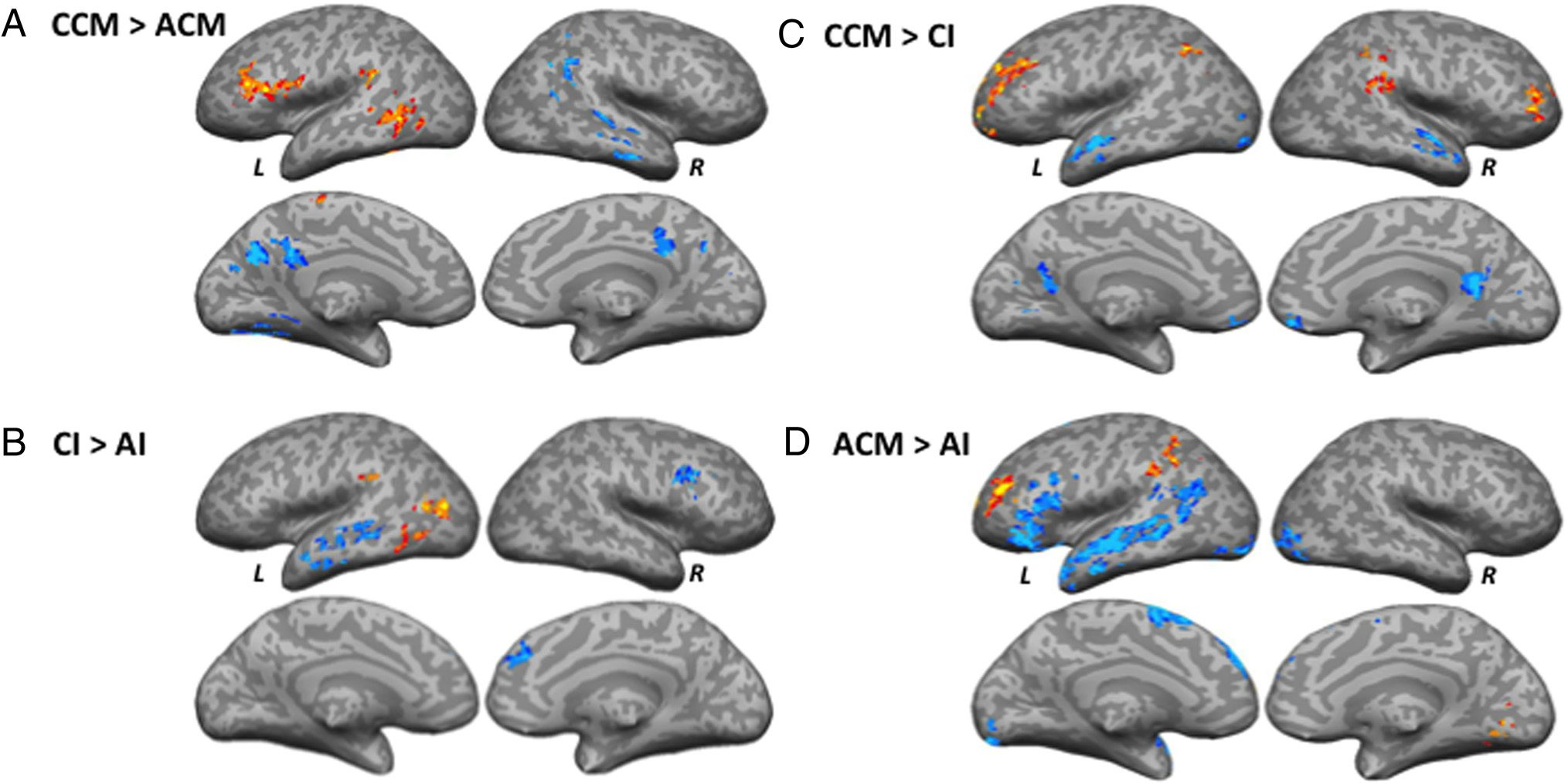
CCM–ACM
The CCM condition relative to the ACM condition led to
stronger activation in the left IFG (pars opercularis),
MTG, SMG, and paracentral gyrus. Areas activated to a
greater extent by the ACM condition relative to the
CCM condition included the left fusiform gyrus, precu-
neus, posterior dorsal cingulum, right AG, and MTG
(Figure 5A, Mesa 3).
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
oh
4
C
7
norte
0
_
7
a
/
_
j
0
oh
0
C
9
norte
2
6
_
a
pag
_
d
0
0
b
9
y
2
gramo
6
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
i
2
3
mi
s
/
j
F
.
t
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
CI–AI
Areas activated to a greater extent by the CI condition rel-
ative to the AI condition included the left middle occipital
gyrus (MOG), MTG, and SMG. Stronger activation was
observed for the AI condition relative to the CI condition
in the left MTG, STS, right MFG, and SFG (Figure 5B,
Mesa 3).
CCM–CI
Stronger activation was observed for the CCM condition
relative to the CI condition in the bilateral MFG, left IPS,
frontomarginal gyrus and sulcus, and right SMG. Areas ac-
tivated to a greater extent by the CI condition relative to
the CCM condition included the bilateral MTG, left MOG,
right rectus, and posterior ventral cingulum (Figure 5C,
Mesa 3).
ACM–AI
The ACM condition elicited stronger activation relative to
the AI condition in the left MFG, AG, and right occipito-
temporal lingual gyrus. The ROI analyses revealed an addi-
tional cluster in the left SMG. Areas activated to a greater
extent by the AI condition relative to ACM condition in-
cluded the left IFG (pars opercularis), STS, MTG, SFG,
occipital pole, and right MOG (Figure 5D, Mesa 3).
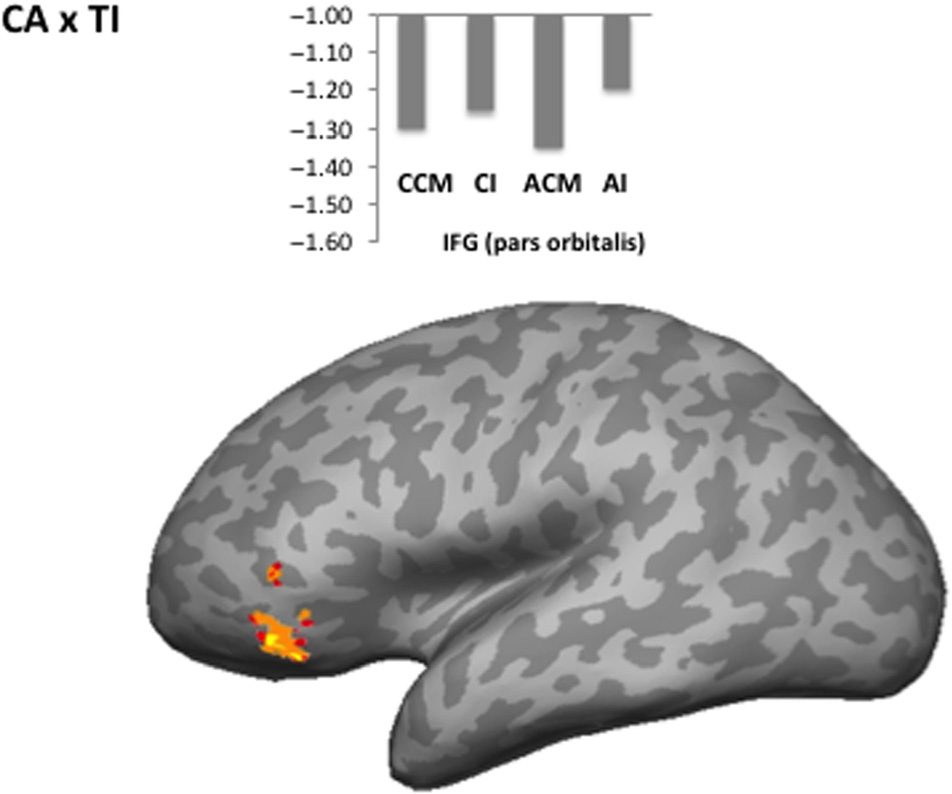
CA × TI Interaction
The left IFG (mostly in pars orbitalis, approximately BA
47) showed an interaction between Concreteness (Estafa-
crete vs. Abstracto) and Transitivity (Transitive [Caused
Motion] vs. Intransitive; Cifra 6, Mesa 3). This interac-
tion arose from a small difference between CCM and CI
conditions and a bigger difference between the abstract
condiciones (AI > ACM).
DISCUSIÓN
The current experiment explored if distinct patterns of
sensory–motor activation can be observed for different
van Dam and Desai
699
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
oh
4
C
7
norte
0
_
7
a
/
_
j
0
oh
0
C
9
norte
2
6
_
a
pag
_
d
0
0
b
9
y
2
gramo
6
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
i
2
3
mi
s
/
j
.
/
t
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Cifra 2. Areas activated by the Caused Motion–Intransitive contrast. Red-orange colors show greater activation for the Transitive/Caused Motion
condición; blue-cyan colors show greater activation for the Intransitive condition. Mean percent signal change relative to rest is shown for the CCM,
CI, ACM, and AI conditions, in a sphere of 5-mm radius around the peak voxel. L = left hemisphere; R = right hemisphere.
syntactic constructions and how these patterns relate to
those induced by verbs.
Main Effect of Verb Concreteness
En el estudio actual, constructions that contained a con-
crete verb elicited greater levels of activation in the ante-
rior part of the left IPL (aIPL) and the left inferior MTG
(ITG) and inferior temporal sulcus. The IPL has shown
to be critical for the representation of action plans and
goals and tool use (Luis, 2006; Rumiati et al., 2004)
and the performance of complex hand–object interac-
ciones (Ramayya, vidrio, & Rilling, 2010; hamilton &
Grafton, 2006). It has also been shown to be part of a tool
use network encompassing the middle/inferior temporal
and inferior frontal regions (Ramayya et al., 2010). Dam-
age to the aIPL/IPL is associated with ideomotor apraxia,
which results in impairments in skilled performance of
motor acts, difficulty imitating gestures, impairments in
Cifra 3. Areas activated by the Transitive/Caused Motion >
Intransitive (rojo) and Concrete > Abstract (azul) contrasts. Yellow
indicates the overlap between the two.
700
Revista de neurociencia cognitiva
Volumen 28, Número 5
Cifra 4. Areas activated by the
Intransitive > Transitive/Caused
Motion (rojo) and Abstract >
Concrete (azul) contrasts.
Yellow indicates the overlap
between the two.
performing appropriate actions in response to a visually
presented object, and carrying out the appropriate ac-
tion for a certain object ( Jax, Buxbaum, & Moll, 2006;
Haaland, Harrington, & Caballero, 2000; Heilman & Rothi,
1993; Heilman, Rothi, & Valenstein, 1982). Además,
sentences with action verbs elicited greater activation
within the left posterior temporal gyrus, in proximity to
the human motion area (MT+; rees, Friston, & Koch,
2000). This brain region is implicated in accessing con-
ceptual information about motion attributes and is also
associated with tool use (Gallivan & Culham, 2015; Brandi,
Wohlschläger, Sorg, & Hermsdörfer, 2014; Ramayya et al.,
2010; Saygin, McCullough, Alac, & Emmory, 2010; Kable,
Kann, wilson, Thompson-Schill, & Chatterjee, 2005). Sev-
eral studies have obtained similar effects of concreteness
within premotor and primary motor cortices (Raposo
et al., 2009; Aziz-Zadeh et al., 2006; Tettamanti et al.,
2005; Pulvermüller, Härle, & Hummel, 2000). En nuestro
estudiar, concrete sentences showed a higher BOLD signal
than abstract sentences within the left precentral gyrus,
at a reduced statistical threshold (corrected p < .09).
Similarly, concrete transitive sentences showed a higher
BOLD signal than abstract transitive sentences within the
left precentral gyrus, at a slightly reduced statistical
threshold (corrected p < .07). Activations were exclu-
sively observed within the left premotor cortex. This re-
sult is in line with previous findings that, during lexical
decisions on manual action verbs, right-handers prefer-
entially activate the left premotor cortex, whereas left-
handers predominantly activate the right premotor
cortex ( Willems, Hagoort, & Casasanto, 2010). In addi-
tion, our activation clusters both fell within the premotor
cortex, anterior to primary motor areas. Such an anterior
shift of activation away from primary sensory and motor
cortices has been observed across a number of studies
investigating the neural correlates of action understand-
ing (Chatterjee, 2010; Willems et al., 2010; Willems,
Hagoort, Toni, Hagoort, & Casasanto, 2009; Wallentin,
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
o
4
c
7
n
0
_
7
a
/
_
j
0
o
0
c
9
n
2
6
_
a
p
_
d
0
0
b
9
y
2
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 5. Areas activated by the CCM–CI, CI-AI, and ACM–AI contrasts. Red-orange colors show greater activation for the CI, CCM and ACM
conditions; blue-cyan colors show greater activation for the ACM, AI, CI, and AI conditions. L = left hemisphere; R = right hemisphere.
van Dam and Desai
701
Table 3. Activations in the Remaining Contrasts
Table 3. (continued )
Volume Max
x
y
z
Structure
Volume Max
x
y
z
Structure
CCM > ACM
3726
2781
459
459
3.86 −49
19
23
L IFG (pars opercularis)
3.46 −61 −52
5
L MTG
3.31 −55 −40
3.15 −4 −34
23
59
L SMG
L paracentral g, s
ACM > CCM
ACM > AI
2889
1377
864
432
AI > ACM
4.63 −22
58
3.68 −40 −49
3.45
7 −67
17
44
L MFG
L AG
0 R oc-temp lingual g
3.07 −55 −37
38
L SMG
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
2052
1674
1647
1350
918
837
CI > AI
864
810
270
AI > CI
1755
1242
891
891
−3.80 −22 −67 −12
L oc-temp fusiform g
−4.54 −4 −67
26
L precuneus
−3.92
52 −58
35 R AG
−3.50 −4 −40
29
L post-dorsal cingul
−4.50
−3.59
61 −10 −15 R MTG
61 −31
0 R MTG
4.06 −46 −76
17
L MTG
3.18 −52 −58
0
L MTG
3.05 −55 −37
29
L SMG
−4.11 −55 −16 −12
L MTG
−4.61
49
13
29 R MFG
−3.49 −52 −37
2
L STS
−3.51
7
40
32 R SFG
3.52
3.28
40
43
17 R MFG
61 −40
23 R SMG
3.63 −34
31
3.55 −34 −49
4.36 −34
3.67 −28
46
43
35
32
20
L MFG
L IPS
L MFG
2
L frontomarginal g, s
CCM > CI
1836
1674
1647
1107
945
837
CI > CCM
1782
1323
999
945
918
−3.98
−3.93
7 −49
8 R post ventral cingul
58 −4 −9 R MTG
−3.95 −55 −1 −9
L MTG
−3.42 −31 −82 −6
L MOG
−4.38
4
49 −6 R rectus g
−4.74 −55
22
20
L IFG (pars opercularis)
−4.67 −55 −16 −3
L STS
−3.85 −49
7 −18
L MTG
−3.88 −4
46
−3.53 −34 −52
41
14
L SFG
L STS
−3.85 −25 −88 −9
L occipital pole
−4.17 −4
19
56
L SFG
−3.43
31 −85
0 R MOG
7020
6264
2943
2862
2781
2673
2160
1647
CA × TI
1026
3.70 −43
28
5
L IFG (pars orbitalis)
The volume of the cluster (μl), peak z score, Talairach coordinates, y
the anatomical structures that the clusters overlap are shown. post = pos-
terior; oc-temp = occipito-temporal; cingul = cingulate. L = left hemi-
sphere; R = right hemisphere; g = gyrus; s = sulcus; ant = anterior.
Ellegaard, Ostergaard, Ostergaard, & Roepstorff, 2005).
Several authors have proposed that such anterior activa-
tions might reflect representations that are abstracted
away from perceptually based representations and sche-
matized over the course of learning, which are neverthe-
less shared between language and perception (ver, p.ej.,
Humphreys, Newling, Jennings, & Gennari, 2013).
Constructions that contained an abstract verb elicited
greater levels of activation within lateral temporal re-
gions, most notably the left middle and anterior STS
and STG, as well as in the posterior cingulate. These re-
sults corroborate earlier findings showing that portions of
the left STS and STG play a role in processing abstract
conceptos ( Wilson-Mendenhall, Simmons, Martín, &
Barsalou, 2013; Desai et al., 2009, 2011; Wang, Conder,
Blitzer, & Shinkareva, 2010; Binder, Westbury, McKiernan,
Possing, & Medler, 2005; Wise et al., 2000). En suma-
mary, the current results of verb concreteness effects are
consistent with a number of previous studies, showing ac-
tivation of higher level sensory–motor regions for action
verb processing. These findings provide evidence for the
embodied framework insofar as they confirm that the
meaning of sentences that refer to action is represented
in a cortical network including areas that play a role in per-
ception and action.
702
Revista de neurociencia cognitiva
Volumen 28, Número 5
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
oh
4
C
7
norte
0
_
7
a
/
_
j
0
oh
0
C
9
norte
2
6
_
a
pag
_
d
0
0
b
9
y
2
gramo
6
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
i
2
3
mi
s
/
j
t
F
.
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Main Effect of Transitivity
Of central interest was the question of whether con-
structions themselves are associated with meaning, cuando
controlling for verb meaning. In the Caused Motion–
Intransitive contrast, the effects of verbs were eliminated,
as both constructions contained similar action and abstract
verbos. The results show that sentences with caused motion
frames elicited greater levels of activation in the left aIPL,
which overlapped with the activation found because of
action verbs (Cifra 3). As discussed above, this region is
associated with action planning and control. Aunque el
same general region of the left aIPL was activated by both
action verbs and caused motion constructions, some differ-
ences were also observed. Verbs activated a more ventral
aspect of SMG, extending into the parietal operculum.
Parietal operculum is associated with secondary somato-
sensory cortex. Caused Motion constructions, en el otro
mano, activated a somewhat more dorsal section of the
SMG, extending into the anterior IPS (aIPS). The aIPS
is thought to be the human analog of monkey area AIP
(Culham & Valyear, 2006). Both of these dorsal and ventral
regions are commonly coactivated in grasping and tool use
tareas, and it is somewhat difficult to distinguish their role in
relation to control of actions. Sin embargo, the dorsal region,
and especially AIP/aIPS, is associated more strongly with
hand–object interactions, visually guided grasping, y
on-line dynamic control of movements (Reichenbach,
Thielscher, Peer, Bülthoff, & Bresciani, 2014; Rice, Tunik,
& Grafton, 2006; Tunik, Frey, & Grafton, 2005; Culham
et al., 2003; see Tunik, Rice, hamilton, & Grafton, 2007,
para una revisión).
Cifra 6. Areas showing an interaction between Concreteness
(Concrete vs. Abstracto) and Transitivity (Transitive/Caused Motion vs.
Intransitive). Mean percent signal change relative to rest is shown for
the CCM, CI, ACM, and AI conditions, in a sphere of 5-mm radius
around the peak voxel.
The left dorsal AG was also activated for caused motion
constructions relative to intransitive constructions. El
entire AG has been shown to play a role in semantic pro-
cesando (Bonner, Peelle, Cocinar, & Grossman, 2013; Binder
& Desai, 2011; Binder, Desai, Tumbas, & Conant, 2009).
Asimismo, TMS and patient studies have substantiated
the idea that the AG is involved in controlled semantic
Procesando ( Whitney, Kirk, O'Sullivan, Lambon Ralph, &
Jefferies, 2011, 2012; Corbett, Jefferies, & Lambon Ralph,
2009; Jefferies & Lambon Ralph, 2006). More specifically,
the dorsal AG has shown to play a role in action planning
and execution, especially reach-to-grasp actions. In a bi-
focal TMS study, Koch et al. (2010) examined connectiv-
ity between dorsal AG and the primary motor cortex (M1)
as well as between dorsal SMG and M1. They found that,
during early (∼50 msec) preparation of reaching and
grasping movements, AG–M1 connectivity was sensitive
to both type of grasp (whole-hand grasp) y ubicación
of the object in space, whereas SMG–M1 connectivity
was sensitive only to the type of grasp (precision grasp).
Structurally, both dorsal SMG and dorsal AG were con-
nected to M1 and to ventral premotor cortex, a través de
different bundles within the superior longitudinal
fasciculus.
Además, caused motion constructions also activated
the posterior medial aspect of the parietal lobe. Esta re-
gion, dorsal to the parieto-occipital sulcus and near the
posterior edge of the hemisphere, is termed the “parietal
reach region” (PRR) and is found in both monkeys and
humanos ( Yttri, Wang, Liu, & Snyder, 2014; Chang, Papa-
dimitriou, & Snyder, 2009; Filimon, nelson, Huang, &
Sereno, 2009; Connolly, andersen, & Goodale, 2003).
PRR plays a role in planning visually guided reaching
movimientos, possibly by computing the initial response
by taking into account the difference between the initial
hand position and gaze location.
The left MFG and IFS were also activated by both
caused motion constructions and action verbs. A dorsal–
ventral pattern, similar to the one seen in the aIPL, era
also seen in the frontal lobe, whereby the caused mo-
tion constructions activated more dorsal MFG regions,
whereas activation because of action verbs extended
more ventrally to the IFS. These frontal regions have a
wide-ranging role in executive function (p.ej., Duncan &
Owen, 2000) and can be interpreted here as reflecting
executive aspects of action processing.
En resumen, the activation of areas responsible for ac-
tion planning, alcanzando, and grasping, a saber, aIPS/aIPL,
dorsal AG, and PRR, suggests that caused motion con-
structions, regardless of the verb used, are associated
with action semantics, and this conceptual content is
grounded in regions used in action planning and control.
This is consistent with the idea that caused motion con-
structions activate a caused motion schema, cual es
grounded through areas controlling reaching and grasp-
ing actions. We did not obtain an effect of transitivity
within the premotor, primary motor, or supplementary
van Dam and Desai
703
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
oh
4
C
7
norte
0
_
7
a
/
_
j
0
oh
0
C
9
norte
2
6
_
a
pag
_
d
0
0
b
9
y
2
gramo
6
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
i
2
3
mi
s
/
j
/
.
t
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
motor cortex. A potential explanation is that motor reso-
nance elicited by these constructions represents relatively
abstract action plans and does not contain detailed or
specific parameters related to selection and execution of
the actions. In a study of patients with stroke, Desai,
Herter, Riccardi, Rorden, and Fridriksson (2015) presentado
that comprehension accuracy of action-related words
was selectively predicted by detailed parameters (p.ej., ini-
tial direction error or number speed maxima) of reaching
actions performed by the patients. This suggests that such
details are part of simulations at least at a single-word
nivel. Constructions, by their very nature, are general, estafa-
tain slots that can be filled by a variety of words, y son
unlikely to be grounded in specific action parameters.
Bastante, they are grounded in more general, higher-order
action plans.
Sentences with an intransitive frame elicited activations
that are similar to those seen for abstract relative to con-
crete language, both at the word and sentence levels. El
left STS plays a role in processing abstract concepts
( Wilson-Mendenhall et al., 2013; Wang y cols., 2010; Desai
et al., 2009; Sabsevitz, Medler, Seidenberg, & Binder,
2005; Noppeney & Precio, 2004; Mellet, Tzourio, Denis,
& Mazoyer, 1998). The OFC has shown to play a role in
emotion and decision-making and has been argued to
play a role in processing the affective significance of con-
cepts (Rolls & Grabenhorst, 2008; Binder et al., 2005;
Kringelbach, 2005; Bechera, Damasio, & Damasio,
2000). The IFG was activated by intransitives, mostly in-
volving pars orbitalis. An examination of the CCM–CI and
ACM–AI contrasts reveals that this activation was mostly
driven by AIs. This was also the case for most of the ac-
tivation in the left STS/STG. Given that these regions are
commonly activated for abstract relative to concrete con-
cepts, this suggests that AIs were the most abstract of the
condiciones. Abstract words occur in more diverse seman-
tic contexts (Hoffman, Rogers, & Lambon Ralph, 2011),
and their meaning is more dependent on distributional
information and integration with context (Andrews,
Vigliocco, & Vinson, 2009). The left IFG activation can
be interpreted as contributing to deriving meaning
through integration with context. The current results sug-
gest that this applies to intransitive constructions as well,
and their meaning is more abstract and constructed
through integration.
Interaction between Verb and Construction Types
To investigate if the degree to which the main verb elicits
perceptual and motor simulations is modulated by syn-
tactic structure, we calculated an interaction between
the class of the main verb (concrete vs. abstract) y
transitivity (transitive/caused motion vs. intransitive).
Our results show an interaction between these two fac-
tors in the left anterior IFG, involving the pars orbitalis
(spreading slightly into pars triangularis). An examination
of the four conditions indicates that the syntactic frame
modulates activation within the pars orbitalis of the
IFG to a greater extent for sentences with an abstract
as compared with a concrete main verb. Although both
Abstract conditions activated this region relatively more
than the Concrete conditions, the difference between
AI and ACM was greater, giving rise to this interaction.
As the meaning of a concrete noun like “rose” is less
dependent on context, a concrete verb (“throw”) would
strongly guide a specific perceptual/motor simulation
that can be expected to show a weaker modulation by
contexto. The syntactic frame is therefore expected to
have a smaller influence on these verbs. A diferencia de, ab-
stract verbs (“consider”) rely more on the surrounding
frame for their interpretation and should be modu-
lated more. This is precisely what our results show: A
stronger effect of syntactic frame for sentences with an
abstract main verb as compared with a concrete main
verb, within pars orbitalis (BA 47), which is thought to
play a role in controlled semantic retrieval ( Wagner,
Koutstaal, Maril, Schacter, & Buckner, 2000; Gabrieli, Pol-
drack, & Desmond, 1998). In a large-scale meta-analysis
(Binder et al., 2009), BA 47 was found to be sensitive to
semantics, when overall difficulty is controlled (es decir., casos
where conditions with greater executive demands are
compared with relatively easier conditions are eliminat-
ed). Our results suggest that the syntactic frame can
guide the ease with which certain semantic representa-
tions are selected, especially when selection of semantic
information is not easily achieved on the basis of the
main verb alone. Como se ha mencionado más arriba, these findings
are consistent with the proposal by Andrews and col-
ligas (2009) suggesting that concrete and abstract rep-
resentations are learned from different types of statistical
datos. Concrete representations are based on a set of per-
ceived physical properties of an object and the experien-
tial information we acquired while interacting with them.
Abstract representations, por otro lado, rely more
strongly on information of how words are distributed
across different linguistic environments. The results sug-
gest that the left pars orbitalis is especially relevant to in-
tegrating verb and construction-level meaning.
The two-level theory of verb meaning suggests that
such an interface would be provided by the template level
of the verb. This theoretical framework argues for two
separate levels of verb meaning. The composite meaning
of a verb constitutes an association of the root and tem-
plate level of a particular verb (for an overview, see Levin
& Rappaport Hovav, 2005). The uniqueness of every verb
is constituted by the root level of meaning, which is not
visible to grammar and provides subtle distinctions be-
tween each verb in a given class. In contrast to the root
level of verb meaning, the template level is visible to
grammar and provides a schematic representation that
is common to all verbs in a given class (Kemmerer,
2012; Kemmerer & Gonzalez-Castillo, 2010). Several neu-
ropsychological studies have provided evidence that
704
Revista de neurociencia cognitiva
Volumen 28, Número 5
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
oh
4
C
7
norte
0
_
7
a
/
_
j
0
oh
0
C
9
norte
2
6
_
a
pag
_
d
0
0
b
9
y
2
gramo
6
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
i
2
3
mi
s
/
j
F
t
.
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
these two levels of verb meaning are segregated in the
mente. Por ejemplo, studies with brain-damaged partici-
pants have shown a double dissociation in the impair-
ment in the discrimination of features of verb meaning
that are relevant to grammar versus features that are irrel-
evant to grammar (Kemmerer, 2000, 2003; Kemmerer &
Wright, 2002). There is some evidence for the associa-
tion of the left IFG with the schematic motor component
of template-level verb meanings. Por ejemplo, Tettamanti
and colleagues (2005) compared prototypical caused mo-
tion constructions with and without motor components,
such as “I bite an apple” and “I appreciate sincerity” (en
italiano), and found activation in the left pars opercularis
for motor sentences. Sin embargo, both verbs and nouns dif-
fered in these sentences, and it is not clear whether the
differences were due only to verb concreteness. Sobre el
other hand, Desai et al. (2011) performed a similar com-
parison (using simple caused motion sentences such as
“The thief bashed the table” and “The jury criticized the
proposal”) and did not find activation in the left IFG. En
another study, Desai et al. (2009) compared prototypical
transitive sentences (“I throw the ball”) with abstract sen-
tenencias (“I consider the risk”) and again did not find activa-
tion in the left IFG. Además, to control for the effect of
noun manipulability, they compared action sentences with
sentences with visual verbs (“I see the ball”), and no IFG
activation was found in that comparison either. De este modo, el
evidence associating left IFG with simple or prototypical
transitive constructions is mixed. A potentially relevant fac-
tor is that the posterior region of the left IFG (pars oper-
cularis/BA 44) is also involved in overt or covert speech
production and perception. If the action sentences have
longer syllable or phoneme length than the control sen-
tenencias, activation of posterior IFG can be expected. Ambos
Desai et al. (2009) and Desai et al. (2011) ensured that ac-
tion sentences were not longer (in terms of words, letters,
phonemes, or syllables) than the control abstract or visual
oraciones. Tettamanti and colleagues (2005) do not report
syllable or phoneme length of their sentences, and it is not
clear if they were matched. Studies that compare pro-
nounceable stimuli (p.ej., words or sentences) with non-
pronounceable materials (p.ej., false fonts or hash marks)
and report activation of posterior IFG can therefore be
questioned on these grounds. In our study, we did not ob-
serve activation of the pars opercularis. Es posible que
this region was activated equally by all verbs because it is
always involved in processing the argument structure and
is not seen in any contrasts. An alternative possibility is
that template-level meaning is not necessarily attached
to specific verbs, but templates or constructions them-
selves have semantics over and above those of the verb
that is filled in. Several psycholinguistic studies have pro-
vided evidence that syntactic constructions carry meanings
(Kako, 2006; Kaschak & Glenberg, 2000; Pescador, 1994,
1996). Además, a recent study by Allen, Pereira,
Botvinick, and Goldberg (2012) showed different neural
activations for the dative (p.ej., “Sally gave the book to
Joe”) and the ditransitive construction (p.ej., “Sally gave
Joe a book”), which share the same content words. Estos
results provide evidence that different grammatical con-
structions are associated with distinct patterns of neural
activación.
Finalmente, we note that several areas in various compari-
hijos, such as the left STS and MFG, showed negative ac-
tivations, relative to the baseline of gaps between the
estímulos, for all conditions. Aquí, only the differences be-
tween conditions are relevant, and not the sign, for two
razones. Primero, the sentences were separated only by
7.4 segundo, which is not long enough for the hemodynamic
response to return to baseline or even to become weak.
A “resting” state never occurs during the experiment;
there is only modulation due to reading various sen-
tenencias. De este modo, activations are not against rest but against
a baseline where general processes involved in process-
ing sentences are still active. Segundo, resting is not an
“off” or “neutral” state with respect to semantic process-
ing but is instead a very active period of rich semantic
Procesando. It has been suggested that engagement in ef-
fortful tasks reliably suppresses such task-unrelated se-
mantic processing (Binder et al., 2009). Applying this
logic leads to the prediction that, if the AG plays a role
in semantic processing than it’s expected to be deacti-
vated during nonsemantic tasks, no deactivation is
expected during semantic tasks. The finding that deacti-
vation of the left IPL (particularly the AG), which is part of
the Default Mode Network, is modulated by the semantic
nature of the task, shows that resting state and semantic
networks greatly overlap (Seghier & Precio, 2012; Seghier,
Fagan, & Precio, 2010; but see Humphreys, Hoffman,
Visser, Binney, & Lambon Ralph, 2015).
Conclusión
Our results show that the transitive/caused motion and
intransitive constructions are associated with distinct
patterns of neural activation. The caused motion con-
structions, across both action and abstract verbs, elicited
activation in areas associated with reaching and grasp-
En g. The intransitive constructions were processed simi-
larly to abstract sentences, activating lateral temporal
areas. These results provide evidence that the transitive
construction, by virtue of being associated with a caused
motion schema, elicits activations within sensory–motor
regiones del cerebro. Eso es, the syntactic construction itself
may drive the mental simulation of an event involving
the movement of an object (along a designated path).
Pars orbitalis has an important role in construction and
verb-level meaning, showing an interaction between
verb and construction type. These findings provide evi-
dence for constructionist approaches that claim that
form–meaning correspondences do not exclusively exist
at the level of verbs or lexical items but also at the level
of constructions.
van Dam and Desai
705
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
d
oh
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
oh
4
C
7
norte
0
_
7
a
/
_
j
0
oh
0
C
9
norte
2
6
_
a
pag
_
d
0
0
b
9
y
2
gramo
6
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
i
2
3
mi
s
/
j
t
/
F
.
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Acknowledgment
We thank David Kemmerer and an anonymous reviewer for their
helpful suggestions. All mistakes are ours. This research was
supported by NIH/NIDCD grant R01 DC010783 (R. h. D.).
Reprint requests should be sent to Rutvik H. Desai, 220 Dis-
covery Building, 915 Greene St., University of South Carolina,
Columbia, CAROLINA DEL SUR 29208, o por correo electrónico: rutvik@sc.edu.
Nota
1. On the basis of the data from an experiment that involved
physical application of painful heat at different levels of inten-
sity, Cortejar, Krishnan, and Wagner (2014) suggested using a
voxelwise threshold of p < .001 or lower. The recommendation
of such a stringent threshold is appropriate given that sensory
tasks typically lead to activations that are large in extent. On the
other hand, studies of higher cognitive functions often involve a
comparison of conditions that are closely matched in their sen-
sory properties and differ in more subtle ways. In our experi-
ence, such contrasts show activations that are lower in
magnitude and are much more variable between participants.
To balance false positives and false negatives in group maps,
thresholds in the range of p < .025 to p < .005 are more ap-
propriate in these cases. This is why a large number of cognitive
experiments use voxelwise thresholds in the neighborhood of
p < .01, and similar values are used as defaults in some pack-
ages. Although Woo et al.’s (2014) recommendation remains
valuable in many cases, our choice of threshold is based on
these considerations.
REFERENCES
Agnew, Z. K., van de Koot, H., McGettigan, C., & Scott, S. K.
(2014). Do sentences with unaccusative verbs involve
syntactic movement? Evidence from neuroimaging.
Language, Cognition, and Neuroscience, 29, 1035–1045.
Allen, K., Pereira, F., Botvinick, M., & Goldberg, A. E. (2012).
Distinguishing grammatical constructions with fMRI pattern
analysis. Brain and Language, 123, 174–182.
Andrews, M., Vigliocco, G., & Vinson, D. (2009). Integrating
experiential and distributional data to learn semantic
representations. Psychological Review, 116, 463–498.
Aziz-Zadeh, L., Wilson, S. M., Rizzolatti, G., & Iacoboni, M.
(2006). Congruent embodied representations for visually
presented actions and linguistic phrases describing actions.
Current Biology, 16, 1818–1823.
Barsalou, L. W. (1999). Perceptual symbol systems. Behavioral
and Brain Sciences, 22, 577–660.
Barsalou, L. W. (2008). Grounded cognition. Annual Review of
Psychology, 59, 617–645.
Bechera, A., Damasio, H., & Damasio, A. R. (2000). Emotion,
decision making and the orbitofrontal cortex. Cerebral
Cortex, 10, 295–307.
Ben-Shachar, M., Hendler, T., Kahn, I., Ben-Bashat, D., &
Grodzinsky, Y. (2003). The neural reality of syntactic
transformations: Evidence from functional magnetic
resonance imaging. Psychological Science, 14, 433–440.
Bergen, B. K., & Chang, N. C. (2003). Embodied construction
grammar in simulation-based language understanding. In
J. O. Ostman & M. Fried (Eds.), Construction grammar(s):
Cognitive and cross-language dimensions. Amsterdam:
John Benjamin Publishing Company.
Binder, J. R., & Desai, R. H. (2011). The neurobiology
of semantic memory. Trends in Cognitive Sciences,
15, 527–536.
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L. (2009).
Where is the semantic system? A critical review and meta-
analysis of 120 functional neuroimaging studies. Cerebral
Cortex, 19, 2767–2796.
Binder, J. R., Westbury, C. F., McKiernan, K. A., Possing, E. T., &
Medler, D. A. (2005). Distinct brain systems for processing
concrete and abstract words. Journal of Cognitive
Neuroscience, 17, 905–917.
Bolinger, D. (1968). Entailment and the meaning of structures.
Glossa, 2, 199–127.
Bonner, M. F., Peelle, J. E., Cook, P. A., & Grossman, M. (2013).
Heteromodal conceptual processing in the angular gyrus.
Neuroimage, 71, 175–186.
Brandi, M.-L., Wohlschläger, A., Sorg, C., & Hermsdörfer, J.
(2014). The neural correlates of planning and executing
actual tool use. Journal of Neuroscience, 34, 13183–13194.
Brysbaert, M., Warriner, A. B., & Kuperman, V. (2013).
Concreteness ratings for 40 thousand generally known
English word lemmas. Behavior Research Methods, 46,
904–911.
Buccino, G., Binkofski, F., Fink, G. R., Fadiga, L., Fogassi, L.,
Gallese, V., et al. (2001). Action observation activates
premotor and parietal areas in a somatotopic manner:
An fMRI study. European Journal of Neuroscience,
13, 400–404.
Caspers, S., Zilles, K., Laird, A. R., & Eickhoff, S. B. (2010).
ALE meta-analysis of action observation and imitation in
the human brain. Neuroimage, 50, 1148–1167.
Chang, S. W. C., Papadimitriou, C., & Snyder, L. H. (2009).
Using a compound gain field to compute a reach plan.
Neuron, 64, 744–755.
Chatterjee, A. (2010). Disembodying cognition. Language
and Cognition, 2, 79–116.
Chen, E., Widick, P., & Chatterjee, A. (2008). Functional-
anatomical organization of predicate metaphor processing.
Brain and Language, 119, 149–157.
Connolly, J. D., Andersen, R. A., & Goodale, M. A. (2003). fMRI
evidence for a “parietal reach region” in the human brain.
Experimental Brain Research, 153, 140–145.
Corbett, F., Jefferies, E., & Lambon Ralph, M. A. (2009).
Different impairments of semantic cognition in semantic
dementia and semantic aphasia: Evidence from the
non-verbal domain. Brain, 132, 2593–2608.
Cox, R. W. (1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computers
and Biomedical Research, 29, 162–173.
Cox, R. W., & Jesmanowicz, A. (1999). Real-time 3D image
registration of functional MRI. Magnetic Resonance in
Medicine, 42, 1014–1018.
Croft, W. A. (2001). Radical construction grammar: Syntactic
theory in typological perspective. Oxford: Oxford University
Press.
Culham, J. C., Danckert, S. L., DeSouza, J. F. X., Gati, J. S.,
Menon, R. S., & Goodale, M. A. (2003). Visually guided
grasping produces fMRI activation in dorsal but not
ventral stream brain areas. Experimental Brain Research,
153, 180–189.
Culham, J. C., & Valyear, K. F. (2006). Human parietal
cortex in action. Current Opinion in Neurobiology,
16, 205–212.
Desai, R. H., Binder, J. R., Conant, L. L., Mano, Q. R., &
Seidenberg, M. S. (2011). The neural career of
sensory–motor metaphors. Journal of Cognitive
Neuroscience, 23, 2376–2386.
Desai, R. H., Binder, J. R., Conant, L. L., & Seidenberg,
M. S. (2009). Activation of sensory–motor areas
in sentence comprehension. Cerebral Cortex,
20, 468–478.
706
Journal of Cognitive Neuroscience
Volume 28, Number 5
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
o
4
c
7
n
0
_
7
a
/
_
j
0
o
0
c
9
n
2
6
_
a
p
_
d
0
0
b
9
y
2
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Desai, R. H., Conant, L. L., Binder, J. R., Park, H., & Seidenberg,
M. S. (2013). A piece of the action: Modulation of sensory–
motor regions by action idioms and metaphors. Neuroimage,
83, 862–869.
Desai, R. H., Herter, T., Riccardi, N., Rorden, C., & Fridriksson,
Haiman, J. (1985). Natural syntax: Iconicity and erosion.
Cambridge: Cambridge University Press.
Hamilton, A. F. C., & Grafton, S. T. (2006). Goal representation
in human anterior intraparietal sulcus. Journal of
Neuroscience, 26, 1133–1137.
J. (2015). Concepts within reach: Action performance
predicts action language processing in stroke.
Neuropsychologia, 71, 217–224.
Desikan, R. S., Ségonne, F., Fischl, B., Quinn, B. T., Dickerson,
B. C., Blacker, D., et al. (2006). An automated labeling system
for subdividing the human cerebral cortex on MRI scans into
gyral based regions of interest. Neuroimage, 31, 968–980.
Destrieux, C., Fischl, B., Dale, A., & Halgren, E. (2010).
Automatic parcellation of human cortical gyri and sulci using
standard anatomical nomenclature. Neuroimage, 53, 1–15.
Dixon, R. M. W. (2010). Basic linguistic theory. In Grammatical
topics (Vol. 2). Oxford: Oxford University Press.
Duncan, J., & Owen, A. M. (2000). Common regions of the
human frontal lobe recruited by diverse cognitive demands.
Trends in Neurosciences, 23, 475–483.
Filimon, F., Nelson, J. D., Huang, R. S., & Sereno, M. I. (2009).
Multiple parietal reach regions in humans: Cortical
representations for visual and proprioceptive feedback
during online reaching. Journal of Neuroscience, 29,
2961–2971.
Fillmore, C., Kay, P., & O’Connor, C. (1988). Regularity and
idiomaticity in grammatical constructions: The case of let
alone. Language, 64, 501–538.
Fisher, C. (1994). Structure and meaning in the verb lexicon:
Input for a syntax-aided verb learning procedure. Language
and Cognitive Processes, 9, 473–517.
Fisher, C. (1996). Structural limits on verb mapping: The role of
analogy in children’s interpretations of sentences. Cognitive
Psychology, 23, 331–392.
Fisher, M., & Zwaan, R. (2008). Embodied language: A review of
the role of the motor system in language comprehension.
Quarterly Journal of Experimental Psychology, 61, 825–850.
Fodor, J. (1983). The modularity of mind. Cambridge, MA: MIT
Press.
Foundas, A. L., Macauley, B. L., Raymer, A. M., Maher, L. M.,
Rothi, L. J., & Heilman, K. M. (1999). Ideomotor apraxia in
Alzheimer disease and left hemisphere stroke: Limb transitive
and intransitive movements. Neuropsychiatry,
Neuropsychology, and Behavioral Neurology, 12, 161–166.
Gabrieli, J. D., Poldrack, R. A., & Desmond, J. E. (1998). The role
of left prefrontal cortex in language and memory.
Proceedings of the National Academy of Sciences, U.S.A., 95,
906–913.
Gallese, V., Fadiga, L., Fogassi, L., & Rizzolatti, G. (1996). Action
recognition in the premotor cortex. Brain, 119, 593–603.
Gallivan, J. P., & Culham, J. C. (2015). Neural coding within
human brain areas involved in actions. Current Opinion in
Neurobiology, 33, 141–149.
Glenberg, A. M. (1997). What memory is for. Behavioral and
Brain Sciences, 20, 1–55.
Goldberg, A. E. (1995). Constructions: A construction
grammar approach to argument structure. Chicago:
University of Chicago Press.
Goldberg, A. E. (2003). Constructions: A new theoretical
approach to language. Trends in Cognitive Sciences, 7,
219–224.
Grosbras, M.-H., Beaton, S., & Eickhoff, S. B. (2012). Brain
regions involved in human movement perception: A
quantitative voxel-based meta-analysis. Human Brain
Mapping, 33, 431–454.
Haaland, K. Y., Harrington, D. I., & Knight, R. T. (2000).
Neural representations of skilled movement. Brain,
123, 2306–2313.
Harnad, S. (1990). The symbol grounding problem. Physica D,
42, 335–346.
Heilman, K. M., & Rothi, L. J. (1993). Apraxia. In K. M. Heilman
& E. Valenstein (Eds.), Clinical neuropsychology (3rd ed.,
pp. 141–163). New York: Oxford University Press.
Heilman, K. M., Rothi, L. J. G., & Valenstein, E. (1982). Two
forms of ideomotor apraxia. Neurology, 32, 342–346.
Hoenig, K., Sim, E.-J., Bochev, V., Herrnberger, B., & Kiefer, M.
(2008). Conceptual flexibility in the human brain: Dynamic
recruitment of semantic maps from visual, motor, and
motion-related areas. Journal of Cognitive Neuroscience,
20, 1799–1814.
Hoffman, P., Rogers, T. T., & Lambon Ralph, M. A. (2011).
Semantic diversity accounts for the “missing” word frequency
effect in stroke aphasia: Insights using a novel method to
quantify contextual variability in meaning. Journal of
Cognitive Neuroscience, 23, 2432–2446.
Humphreys, G., Hoffman, P., Visser, M., Binney, R. J., & Lambon
Ralph, M. A. (2015). Establishing task- and modality-
dependent dissociations between the semantic and default
mode networks. Proceedings of the National Academy of
Sciences, U.S.A., 112, 7857–7862.
Humphreys, G., Newling, K., Jennings, C., & Gennari, S. P.
(2013). Motion and actions in language: Semantic
representations in occipito-temporal cortex. Brain and
Language, 125, 94–105.
Jax, S. A., Buxbaum, L. J., & Moll, A. D. (2006). Deficits in
movement planning and intrinsic coordinate control in
ideomotor apraxia. Journal of Cognitive Neuroscience,
18, 2063–2076.
Jefferies, E., & Lambon Ralph, M. A. (2006). Semantic
impairment in stroke aphasia versus semantic dementia: A
case-series comparison. Brain, 129, 2131–2147.
Kable, J. W., Kann, I. P., Wilson, A., Thompson-Schill, S. L., &
Chatterjee, A. (2005). Conceptual representations of action in
the lateral temporal cortex. Journal of Cognitive
Neuroscience, 17, 1855–1870.
Kako, E. (2006). The semantics of syntactic frames. Language
and Cognitive Processes, 21, 562–575.
Kaschak, M. P., & Glenberg, A. M. (2000). Constructing
meaning: The role of affordances and grammatical
constructions in sentence comprehension. Journal of
Memory and Language, 43, 508–529.
Katz, J. J., & Fodor, J. A. (1963). The structure of a semantic
theory. Language, 39, 170–210.
Kemmerer, D. (2000). Grammatically relevant and
grammatically irrelevant features of verb meaning can be
independently impaired. Aphasiology, 14, 997–1020.
Kemmerer, D. (2003). Why can you hit someone on the arm
but not break someone on the arm? A neuropsychological
investigation of the English body-part possessor ascension
construction. Journal of Neurolinguistics, 16, 13–36.
Kemmerer, D. (2006). Action verbs, argument structure
constructions, and the mirror neuron system. In
M. Arbib (Ed.), Action to language via the mirror
neuron system. Cambridge: Cambridge University
Press.
Kemmerer, D. (2012). The crosslinguistic prevalence of SOV
and SVO word orders reflects the sequential and hierarchical
representation of action in Broca’s area. Language and
Linguistics Compass, 6, 50–66.
Kemmerer, D. (2015). Are the motor features of verb meanings
represented in the precentral motor cortices? Yes, but
van Dam and Desai
707
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
o
4
c
7
n
0
_
7
a
/
_
j
0
o
0
c
9
n
2
6
_
a
p
_
d
0
0
b
9
y
2
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
within the context of a flexible, multilevel architecture for
conceptual knowledge. Psychonomic Bulletin and Review,
22, 1068–1075.
Kemmerer, D., & Gonzalez-Castillo, J. (2010). The two-level
theory of verb meaning: An approach to integrating the
semantics of action with the mirror neuron system. Brain
and Language, 112, 54–76.
Kemmerer, D., & Wright, S. (2002). Selective impairment of
knowledge underlying un-prefixation: Further evidence for
the autonomy of grammatical semantics. Journal of
Neurolinguistics, 15, 403–432.
Kiefer, M., & Pulvermüller, F. (2012). Conceptual representations
in mind and brain: Theoretical developments, current evidence
and future directions. Cortex, 48, 805–825.
Kintsch, W. (2008). Symbol systems and perceptual
representations. In M. De Vega, A. Glenberg, & A. Graesser
(Eds.), Symbols and embodiment (pp. 145–164). Oxford:
Oxford University Press.
Kipper, K., Dang, H. T., & Palmer, M. (2000). Class-based
construction of a verb lexicon. In Proceedings of the Seventh
National Conference on Artificial Intelligence (AAAI-2000),
pp. 691–696.
Koch, G., Cercignani, M., Pecchioli, C., Versace, V., Oliveri, M.,
Caltagirone, C., et al. (2010). In vivo definition of
parieto-motor connections involved in planning of
grasping movements. Neuroimage, 51, 300–312.
Korhonen, A., Krymolowski, Y., & Briscoe, T. (2006). A large
subcategorization lexicon for natural language processing
applications. In Proceedings of LREC, pp. 1326–1331.
Kringelbach, M. L. (2005). The human orbitofrontal cortex:
Linking reward to hedonic experience. Nature Reviews
Neuroscience, 6, 691–702.
Kuno, S., & Takami, K. (2004). Functional constraints in
grammar: On the unergative-unaccusative distinction.
Philadelphia: John Benjamins Publishing Co.
Lakoff, G. (1987). Women, fire, and dangerous things: What
categories reveal about the mind. Chicago: University of
Chicago Press.
Langacker, R. W. (1999). Grammar and conceptualizations.
Berlin & New York: Mouton de Gruyter.
Levin, B. (1995). Approaches to lexical semantic representation.
In D. E. Walker, A. Zampolli, & N. Calzolari (Eds.),
Automating the lexicon: Research and practice in a
multilingual environment (pp. 53–91). Oxford: Oxford
University Press.
Levin, B., & Rappaport Hovav, M. (2005). Argument
realization. Cambridge: Cambridge University Press.
Lewis, J. (2006). Cortical networks related to human use of
tools. The Neuroscientist, 12, 211–231.
Lui, F., Buccino, G., Duzzi, D., Benuzzi, F., Crisi, G.,
Baraldi, P., et al. (2008). Neural substrates for observing
and imagining non-object-directed actions. Social
Neuroscience, 3, 261–275.
Luzzatti, C., Raggi, R., Zonca, G., Pistarini, C., Contardi, A., &
Pinna, G. D. (2002). Verb-noun double dissociation in aphasic
lexical impairments: The role of word frequency and
imageability. Brain and Language, 81, 432–444.
Malchukov, A., Haspelmath, M., & Comrie, B. (Eds.) (2010).
Studies in ditransitive constructions: A comparative
handbook. Berlin: De Gruyter.
Mayka, M. A., Corcos, D. M., Leurgans, S. E., & Vaillancourt,
D. E. (2006). Three-dimensional locations and boundaries
of motor and premotor cortices as defined by functional
brain imaging: A meta-analysis. Neuroimage,
31, 1453–1474.
McAllister, T., Bachrach, A., Waters, G., Michaud, J., & Caplan,
D. (2009). Production and comprehension of unaccusatives
in aphasia. Aphasiology, 23, 989–1004.
Mellet, E., Tzourio, N., Denis, M., & Mazoyer, B. (1998). Cortical
anatomy of mental imagery of concrete nouns based on their
dictionary definition. NeuroReport, 9, 803–808.
Meltzer-Asscher, A., Schuchard, J., den Ouden, D.-B., &
Thompson, C. K. (2012). The neural substrates of complex
argument structure representations: Processing “alternating
transitivity” verbs. Language and Cognitive Processes,
28, 1154–1168.
Meteyard, L., Cuadrado, S. R., Bahrami, B., & Vigliocco, G.
(2012). Coming of age: A review of embodiment and
neuroscience of semantics. Cortex, 48, 788–804.
Mozaz, M., Rothi, L. J., Anderson, J. M., Crucian, G. P., &
Heilman, K. M. (2002). Postural knowledge of transitive
pantomimes and intransitive gestures. Journal of the
International Neuropsychological Society, 8, 958–962.
Noppeney, U., & Price, C. J. (2004). Retrieval of abstract
semantics. Neuroimage, 22, 164–170.
Pinker, S. (1989). Learnability and cognition: The acquisition
of argument structure. Cambridge, MA: MIT Press.
Pulvermüller, F. (1999). Words in the brain’s language.
Behavioral and Brain Sciences, 22, 253–336.
Pulvermüller, F., Härle, M., & Hummel, F. (2000).
Neurophysiological distinction of verb categories.
NeuroReport, 11, 2789–2793.
Pylyshyn, Z. W. (1984). Computation and cognition:
Towards a foundation for cognitive science. Cambridge,
MA: MIT Press.
Ramayya, A. G., Glasser, M. F., & Rilling, J. K. (2010).
A DTI investigation of neural substrates supporting
tool use. Cerebral Cortex, 20, 507–516.
Raposo, A., Moss, H. E., Stamatakis, E. A., & Tyler, L. K. (2009).
Modulation of motor and premotor cortices by actions,
action words and action sentences. Neuropsychologia,
47, 388–396.
Rees, G., Friston, K., & Koch, C. (2000). A direct quantitative
relationship between functional properties of human and
macaque V5. Nature Neuroscience, 3, 716–723.
Reichenbach, A., Thielscher, A., Peer, A., Bülthoff, H. H., &
Bresciani, J. P. (2014). A key region in the human parietal
cortex for processing proprioceptive hand feedback during
reaching movements. Neuroimage, 84, 615–625.
Rice, N. J., Tunik, E., & Grafton, S. T. (2006). The anterior
intraparietal sulcus mediates grasp execution independent of
requirement to update: New insights from transcranial
magnetic stimulation. Journal of Neuroscience,
26, 8176–8182.
Rizzolatti, G., Fadiga, L., Gallese, V., & Fogassi, L. (1996).
Premotor cortex and the recognition of motor actions.
Cognitive Brain Research, 3, 131–141.
Roehm, D., Sorace, A., & Bornkessel-Schlesewsky, I. (2012).
Processing flexible form-to-meaning mappings: Evidence for
enriched composition as opposed to indeterminacy.
Language and Cognitive Processes, 28, 1244–1274.
Rolls, E. T., & Grabenhorst, F. (2008). The orbitofrontal cortex
and beyond: From affect to decision-making. Progress in
Neurobiology, 86, 216–244.
Rumiati, R. I., Weiss, P. H., Shallice, T., Ottoboni, G., North, J.,
Zilles, K., et al. (2004). Neural basis of pantomiming the use
of visually presented objects. Neuroimage, 21, 1224–1231.
Saad, Z. S., Glen, D. R., Chen, G., Beauchamp, M. S., Desai, R.,
& Cox, R. W. (2009). A new method for improving
functional-to-structural MRI alignment using local
Pearson correlation. Neuroimage, 44, 839–848.
Sabsevitz, D. S., Medler, D. A., Seidenberg, M., & Binder, J. R.
(2005). Modulation of the semantic system by word
imageability. Neuroimage, 27, 188–200.
Sakreida, K., Schubotz, R. I., Wolfensteller, U., & von Cramon,
D. Y. (2005). Motion class dependency in observers’ motor
708
Journal of Cognitive Neuroscience
Volume 28, Number 5
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
o
4
c
7
n
0
_
7
a
/
_
j
0
o
0
c
9
n
2
6
_
a
p
_
d
0
0
b
9
y
2
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
areas revealed by functional magnetic resonance imaging.
Journal of Neuroscience, 25, 1335–1342.
Saygin, A. P., McCullough, S., Alac, M., & Emmory, K. (2010).
Modulation of bold response in motion-sensitive lateral
temporal cortex by real and fictive motion sentences. Journal
of Cognitive Neuroscience, 22, 2480–2490.
Seghier, M. L., Fagan, E., & Price, C. J. (2010). Functional
subdivisions in the left angular gyrus where the semantic
system meets and diverges from the default network.
Journal of Neuroscience, 30, 16809–16817.
Seghier, M. L., & Price, C. J. (2012). Functional heterogeneity
within the default network during semantic processing and
speech production. Frontiers in Psychology, 3, 281.
doi:10.3389/fpsyg.2012.00281.
Shetreet, E., & Friedmann, N. (2012). Stretched, jumped, and
fell: An fMRI investigation of reflexive verbs and other
intransitives. Neuroimage, 60, 1800–1806.
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic
atlas of the human brain. New York: Thieme.
Tettamanti, M., Buccino, G., Saccuman, M. C., Gallese, V.,
Danna, M., Scifo, P., et al. (2005). Listening to action-related
sentences activates fronto-parietal motor circuits. Journal
of Cognitive Neuroscience, 17, 273–281.
Thompson, C. K., Bonakdarpour, B., & Fix, S. F. (2010). Neural
mechanisms of verb argument structure processing in
agrammatic aphasic and healthy age-matched listeners.
Journal of Cognitive Neuroscience, 22, 1–35.
Thompson, C. K., Bonakdarpour, B., Fix, S. F., Blumenfeld, H. K.,
Parrish, T. B., Gitelman, D. R., et al. (2007). Neural
correlates of verb argument structure processing. Journal
of Cognitive Neuroscience, 19, 1753–1767.
Tkach, D., Reimer, J., & Hatsopoulos, N. G. (2007). Congruent
activity during action and action observation in motor cortex.
Journal of Neuroscience, 27, 13241–13250.
Tunik, E., Frey, S. H., & Grafton, S. T. (2005). Virtual lesions of
the anterior intraparietal area disrupt goal-dependent on-line
adjustments of grasp. Nature Neuroscience, 8, 505–511.
Tunik, E., Rice, N. J., Hamilton, A., & Grafton, S. T. (2007).
Beyond grasping: Representation of action in human anterior
intraparietal sulcus. Neuroimage, 36(Suppl. 2), T77–T86.
Van Dam, W. O., Van Dijk, M., Bekkering, H., & Rueschemeyer,
S.-A. (2012). Flexibility in embodied lexical-semantic
representations. Human Brain Mapping, 33, 2322–2333.
Wagner, A. D., Koutstaal, W., Maril, A., Schacter, D. L., &
Buckner, R. L. (2000). Task-specific repetition priming in left
inferior prefrontal cortex. Cerebral Cortex, 10, 1176–1184.
Wallentin, M., Ellegaard, T., Ostergaard, S., Ostergaard, L., &
Roepstorff, A. (2005). Motion verb sentences activate left
posterior middle temporal cortex despite static context.
NeuroReport, 16, 649–652.
Wang, J., Conder, J. A., Blitzer, D. N., & Shinkareva, S. (2010).
Neural representation of abstract and concrete concepts: A
meta-analysis of neuroimaging studies. Human Brain
Mapping, 31, 1459–1468.
Watson, R. T., Fleet, W. S., Rothi, L. J. G., & Heilman, K. M.
(1986). Apraxia and the supplementary motor area. Archives
of Neurology, 43, 787–792.
Whitney, C., Kirk, M., O’Sullivan, J., Lambon Ralph, M. A., &
Jefferies, E. (2011). The neural organization of semantic
control: TMS evidence for a distributed network in left
inferior frontal and posterior middle temporal gyrus.
Cerebral Cortex, 21, 1066–1075.
Whitney, C., Kirk, M., O’Sullivan, J., Lambon Ralph, M. A., &
Jefferies, E. (2012). Executive semantic processing is
underpinned by a large-scale neural network: Revealing
the contribution of let prefrontal, posterior temporal,
and parietal cortex to controlled retrieval and selection
using TMS. Journal of Cognitive Neuroscience,
24, 133–147.
Wierzbicka, A. (1988). The semantics of grammar. The
semantics of English causative constructions, in a
universal-typological perspective. In M. Tomasello (Ed.),
The new psychology of language (Vol. 1, pp. 113–153).
Mahwah, NJ: Lawrence Erlbaum.
Willems, R. M., Hagoort, P., & Casasanto, D. (2010). Body-specific
representations of action verbs: Neural evidence from
right- and left-handers. Psychological Science, 21, 67–74.
Willems, R. M., Hagoort, P., Toni, I., Hagoort, P., & Casasanto,
D. (2009). Neural dissociations between action verb
understanding and motor imagery. Journal of Cognitive
Neuroscience, 22, 2387–2400.
Wilson-Mendenhall, C. D., Simmons, W. K., Martin, A., &
Barsalou, L. W. (2013). Contextual processing of abstract
concepts reveals neural representations of nonlinguistic
semantic content. Journal of Cognitive Neuroscience,
25, 920–935.
Wise, R., Howard, D., Mummery, C. J., Fletcher, P., Leff, A.,
Buchel, C., et al. (2000). Noun imageability and the temporal
lobes. Neuropsychologia, 38, 985–994.
Woo, C.-W., Krishnan, A., & Wagner, T. D. (2014). Cluster-extent
based thresholding in fMRI analyses: Pitfalls and
recommendations. Neuroimage, 91, 412–419.
Yttri, E. A., Wang, C., Liu, Y., & Snyder, L. H. (2014). The
parietal reach region is limb specific and not involved in
eye-hand coordination. Journal of Neurophysiology,
111, 520–532.
Zwaan, R. A., & Kaschak, M. P. (2008). Language in the brain,
body, and world. In M. Robbins & M. Aydede (Eds.),
Cambridge handbook of situated cognition (pp. 368–381).
Cambridge: Cambridge University Press.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
5
2
8
6
/
9
5
3
/
1
6
9
9
5
3
0
/
5
1
7
7
7
8
o
4
c
7
n
0
_
7
a
/
_
j
0
o
0
c
9
n
2
6
_
a
p
_
d
0
0
b
9
y
2
g
6
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
van Dam and Desai
709