The Neural Basis of Love as a Subliminal Prime:
An Event-related Functional Magnetic
Resonance Imaging Study
S. Ortigue1,2, F. Bianchi-Demicheli3, A. F. de C. Hamilton1,
and S. t. Grafton1,2
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
Abstracto
& Throughout the ages, love has been defined as a motivated
and goal-directed mechanism with explicit and implicit mech-
anisms. Recent evidence demonstrated that the explicit repre-
sentation of love recruits subcorticocortical pathways mediating
premio, emotion, and motivation systems. Sin embargo, the neural
basis of the implicit (unconscious) representation of love re-
mains unknown. To assess this question, we combined event-
related functional magnetic resonance imaging (resonancia magnética funcional) con un
behavioral subliminal priming paradigm embedded in a lexical
decision task. en esta tarea, the name of either a beloved partner,
a neutral friend, or a passionate hobby was subliminally pre-
sented before a target stimulus (palabra, nonword, or blank), y
participants were required to decide if the target was a word or
no. Behavioral results showed that subliminal presentation of
either a beloved’s name (love prime) or a passion descriptor
(passion prime) enhanced reaction times in a similar fashion.
Subliminal presentation of a friend’s name (friend prime) did
not show any beneficial effects. Functional results showed that
subliminal priming with a beloved’s name (as opposed to either
a friend’s name or a passion descriptor) specifically recruited
brain areas involved in abstract representations of others and
the self, in addition to motivation circuits shared with other
sources of passion. Más precisamente, love primes recruited the
fusiform and angular gyri. Our findings suggest that love, as a
subliminal prime, involves a specific neural network that sur-
passes a dopaminergic–motivation system. &
INTRODUCCIÓN
Amar, one of the deepest forms of human endeavor, es
defined as a complex state involving chemical, saber-
tive and goal-directed behavioral components (Bianchi-
Demicheli, Grafton, & Ortigue, 2006; Aron et al., 2005;
Pescador, a, & Marrón, 2005; Bartels & Zeki, 2000, 2004;
Buss, 2003; Mashek, a, & Pescador, 2000; Sternberg
& Barnes, 1988; a & a, 1986, 1996; Hatfield &
Walster, 1978). When involved in a passionate and inti-
mate relationship, the person actively strives for the
happiness of the loved person (clark & Mills, 1979).
Passionate love is often coupled with the inability to feel
passion for more than one person at a time (Berscheid
& Meyers, 1996). En este sentido, love has often been in-
tertwined with passion and defined as a motivated, meta-
directed desire (a & a, 1996; Aron et al., 2005;
Ovid, 1986). Por ejemplo, the old adage ‘‘Do what you
love and never have to work another day in your life’’
illustrates the motivation component of love.
As a goal-directed motivator, love has various behav-
ioral effects. Por ejemplo, different explicit effects of love
1Dartmouth College, 2Universidad de California, Santa Bárbara,
3Geneva University Hospital, Suiza
have been reported, such as euphoria, loss of appetite,
hyperactivity, delay of the onset of fatigue, and a de-
creased need for sleep (Aron et al., 2005; Buss, 2003;
Sternberg & Barnes, 1988; Hatfield & Walster, 1978). Él
has also been shown that love has stress-reducing and
health-promoting potentials (Esch & Stefano, 2005).
Recientemente, two functional magnetic resonance imag-
En g (resonancia magnética funcional) studies reinforced the assumption that love
is a goal-directed state that leads to a range of emo-
ciones, rather than a specific emotion (Aron et al., 2005;
Bartels & Zeki, 2000). These studies showed that in-
tense ongoing love, as compared to friendship, recruits
subcorticocortical pathways mediating reward, emotion,
and motivation systems (Aron et al., 2005; Fisher et al.,
2005; Bartels & Zeki, 2000, 2004). Más precisamente, el
blood oxygen level dependent signal (BOLD) specific to
explicit viewing of photographs of a loved partner
revealed activity focally restricted to the mid insula,
anterior cingulate cortex, head of the caudate nucleus,
ventral tegmental area, putamen, bilateral posterior
hippocampus,
left middle
temporal gyrus, and cerebellum (Aron et al., 2005;
Bartels & Zeki, 2000) as well as the right parietal lobe
(only in Aron et al., 2005). críticamente, the central role of
the dopaminergic–motivation system in ongoing love
left inferior frontal gyrus,
D 2007 Instituto de Tecnología de Massachusetts
Revista de neurociencia cognitiva 19:7, páginas. 1218–1230
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
9
mi
7
–
1
pag
2
d
1
F
8
/
1
1
9
9
/
3
6
7
3
/
9
1
6
2
1
oh
8
C
/
norte
1
2
7
0
5
0
6
7
8
1
1
9
0
/
7
j
1
oh
2
C
1
norte
8
.
pag
2
d
0
0
b
7
y
.
gramo
1
tu
9
mi
.
s
7
t
.
oh
1
norte
2
0
1
7
8
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
F
.
.
.
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
has been emphasized by high positive correlations
between the subjective feeling of love (passionate love
escala [PLS] puntuaciones; Hatfield & Sprecher, 1986) and BOLD
responses recorded from the anteromedial caudate
núcleo (Aron et al., 2005). De este modo, there is now some
explicit functional evidence supporting the hypothesis
that love, as a motivator,
is a powerful source of
behavior via dopaminergic circuits (p.ej., Berscheid &
Meyers, 1996; Sternberg & Barnes, 1988; a & a,
1986, 1991).
Sin embargo, it is unknown if the implicit representation
of love involves similar motivation/dopaminergic circuits.
The previous two functional studies of the neural systems
underlying love used only explicit tasks with faces (a
et al., 2005; Bartels & Zeki, 2000) where explicit percep-
tion of familiar stimuli and conscious processing are
potential confounds (due to top-down interference) para
assessing selective unconscious mechanisms (Dehaene,
Changeux, Naccache, Sackur, & Sergent, 2006; Dehaene
& Changeux, 2000; Dehaene, Kerszberg, & Changeux,
1998). De este modo, the true benefit of love and its neural net-
work on information processing without any top-down
conscious interference remains unknown.
implicit cognition. Typically,
Aquí, we specifically assessed this question by com-
bining event-related fMRI and a behavioral lexical deci-
sion task embedded in a subliminal priming paradigm,
which is one of the more reliable ways of observing
unconscious mechanisms if they exist. Subliminal prim-
ing is indeed a powerful experimental manipulation that
provides a unique opportunity to examine the mecha-
in subliminal
nisms of
priming, a brief presentation (approximately less than
50 mseg) of a masked visual stimulus induces a change
(p.ej., facilitation) in the speed or accuracy of the pro-
cessing of a following target stimulus (priming effect;
p.ej., Bianchi-Demicheli et al., 2006; Henson, 2003; Musch
& Klauer, 2003; Dehaene, Naccache, et al., 1998; Eimer
& Schlaghecken, 1998; Murphy & Zajonc, 1993; Fazio,
Sanbonmatsu, Powell, & Kardes, 1986). Facilitation ef-
fects due to subliminal priming may occur not only
when the relation between primes and targets is a
perceptual one, but also when it is a conceptual one
(p.ej., Spruyt, Hermans, De Houwer, & Eelen, 2004;
Musch & Klauer, 2003; Dehaene, Naccache, et al.,
1998). Amar (as opposed to friendship) can be consid-
ered as a goal-directed drive that may induce implicit
facilitation effects on cognitive behavior due to motiva-
tional and emotional mechanisms rather than a pure
selective emotion without any motivational component
(Bianchi-Demicheli et al., 2006; Aron et al., 2005). En
light of this, one might expect our subliminal priming
paradigm used in combination with event-related fMRI
to show that the unconscious processing of a beloved’s
name (as opposed to a control friend’s name) facilitates
the behavioral processing of a subsequent stimulus
through the recruitment of dopamine-rich brain re-
gions (Bianchi-Demicheli et al., 2006; Aron et al., 2005;
Fisher et al., 2005; a & a, 1986, 1996). Sin embargo,
a beloved’s name and a friend’s name do not vary ex-
clusively on the motivation dimension. Many variables
can induce differential brain activities between these
two types of stimuli. One of these variables is the simple
frequency with which someone thinks about their sig-
nificant other, as opposed to their friend. The amount
of time someone spends thinking about a beloved or a
friend or both may indeed play a crucial role on the
salience of each of these stimuli and thus lead to distinct
brain activities between these stimuli. In order to ex-
clude this confounding variable, we also included a novel
type of prime as a control for frequency of thoughts,
eso es, a descriptor for a passionate interest, tailored to
each individual, that induced frequency of thoughts that
were equivalent to the frequency of thoughts about a
beloved. De este modo, in the present study, each experimental
trial was randomly preceded by the subliminal presenta-
tion of one out of three different types of visual stimuli
(primes): the name of the participant’s beloved (love
prime), and two control primes, eso es, a name of the
participant’s friend (friend prime), or the participant’s
pasión (passion prime).
This design allowed us to test whether the neural basis
of the unconscious mental representation of love de-
pends on (a) motivational/emotional components (por
contrasting love primes vs. friend primes and passion
primes vs. friend primes), (b) salience of the stimuli (por
contrasting love primes vs. passion primes), o ambos. Para
instancia, although we cannot exclude that passion for a
person is different from passion for sports, arte, or objects,
we assume that love primes and passion primes, as impli-
cit motivators, might share some common neural mech-
anisms (as opposed to friend primes) that could affect
cognition, as previously predicted (Bianchi-Demicheli
et al., 2006). This neurofunctional assumption is coherent
with the recent Dehaene–Changeux computational
model that suggests the reinforcement (via Hebbian-type
aprendiendo) of conceptual associations by reward-motivated
learning mechanisms (Dehaene & Changeux, 2000;
Gisiger, Dehaene, & Changeux, 2000). According to this
modelo, the internal representation of a reward signal is
coded in n cortical areas of contribution weighted by
‘‘synaptic efficacities’’ w0, w1, . . ., wn. Both reward and
expectation signals converge at the level of the ventral
tegmental area, and learning modifies synaptic weights to
minimize the discrepancies between the two. The output
of this ventral tegmental area adequately reproduces
dopamine release during performance of various tasks
(Gisiger et al., 2000). This Hebbian-based model
illus-
trates the principle of cognitive learning by production
and selection of pre-representation. críticamente, porque
this model also posits that mental representations of
stimuli may be distributed over different brain areas
according to their conceptual structure (Pulvermuller,
1996, 1999), one might expect that our three types of
prime stimuli (love, amigo, and passion) have overlapping
Ortigue et al.
1219
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
9
mi
7
–
1
pag
2
d
1
F
8
/
1
1
9
9
/
3
6
7
3
/
9
1
6
2
1
oh
8
C
/
norte
1
2
7
0
5
0
6
7
8
1
1
9
0
/
7
j
1
oh
2
C
1
norte
8
.
pag
2
d
0
0
b
7
y
.
gramo
1
tu
9
mi
.
s
7
t
.
oh
1
norte
2
0
1
7
8
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
.
.
F
oh
norte
1
8
METRO
a
y
2
0
2
1
and partly distinct neural representations on the basis of
any differences in contextual encoding.
MÉTODOS
Participantes
Thirty-six healthy heterosexual women, aged 20.1 ± 3.2
(mean ± SD) years who were dating, engaged, o
married to someone and who had a favorite passion in
vida (p.ej., ciencia, Deportes, arte) provided written informed
consent to participate in this experiment, which was ap-
proved by the Committee for Protection of Human Sub-
jects. Participants were recruited from the Dartmouth
College experiment scheduling system on the basis of
advertisements indicating that experimenters were seek-
ing individuals who were currently intensively in love. Todo
participants had normal or corrected-to-normal vision,
were not taking antidepressant medication, and had no
chemical dependency and psychiatric or neurological
illnesses. All participants were right-handed (Edimburgo
Handedness Inventory, Oldfield, 1971).
Just prior to the scanning session, one of us (S.O.)
interviewed each participant in a semistructured format
to obtain general personal information (such as date of
birth; handedness) and to establish the range of each
participant’s feelings of passionate love. This semistruc-
tured interview, based on standard studies, provided
insights into the women’s feelings about their beloved,
the duration and the intensity of their love relationship,
and the percentage of time they think about their be-
loved during their waking hours (Pescador, 2004; Hatfield
& Sprecher, 1986). Entonces, each participant completed
one standard self-report questionnaire: the PLS (Hatfield
& Sprecher, 1986; example items: ‘‘For me, (name of
the beloved) is the perfect romantic partner’’; ‘‘I would
rather be with (name of the beloved) than anyone else’’;
‘‘I have an endless appetite for affection from (name
of the beloved)''. The reported duration of ‘‘being in
love’’ with their partner was 1–60 months (15.3 ± 14.5
[mean ± SD]).
De término medio, participants had a PLS score of 7.7 ± 1.22
(Dakota del Sur) out of 9 puntos. The participant’s subjective inten-
sity of love for their partner was 7.36 ± 1.85 (Dakota del Sur) en un
9-point scale.
As part of the same interview, participants were also
asked about their passions in life, and every participant
provided the name of a passionate hobby. As a selec-
tion criterion, only participants who reported think-
ing about their passion 60% of the day were included
in the experiment. A t test conducted between percent
of thoughts dedicated to the partner and percent of
thoughts dedicated to the passion did not reveal any
significant difference, t(35) = 0.299, pag = .766. Porque
this selection criterion was used for all participants,
there was a ceiling effect of this variable. No correlation
could have been done with reaction times (RTs), acumular-
racy, or functional
imaging data. Finalmente, Participantes
were asked to provide the name of a male friend of
similar age, sex and duration of friendship as their part-
ners, with the imposed condition that they did not feel
any emotional, físico, or intellectual attraction for him,
such that this friend was neutral in the love dimension.
The names of the participant’s beloved, pasión, y
friend were used as primes during the fMRI scanning as
detailed below.
Procedimiento
During the scanner session, participants were instructed
to perform a lexical decision task each time that they saw
a visual stimulus flash onto the screen. They were asked
to indicate as rapidly and as accurately as possible whether
or not an English word was presented on that trial. Re-
sponses were made by pressing one of two response but-
tons on a keyboard with fingers of the right hand (‘‘yes’’
response with the index finger to words and ‘‘no’’ re-
sponse with the middle finger to nonwords or blanks).
The visual stimulus on each trial was composed of
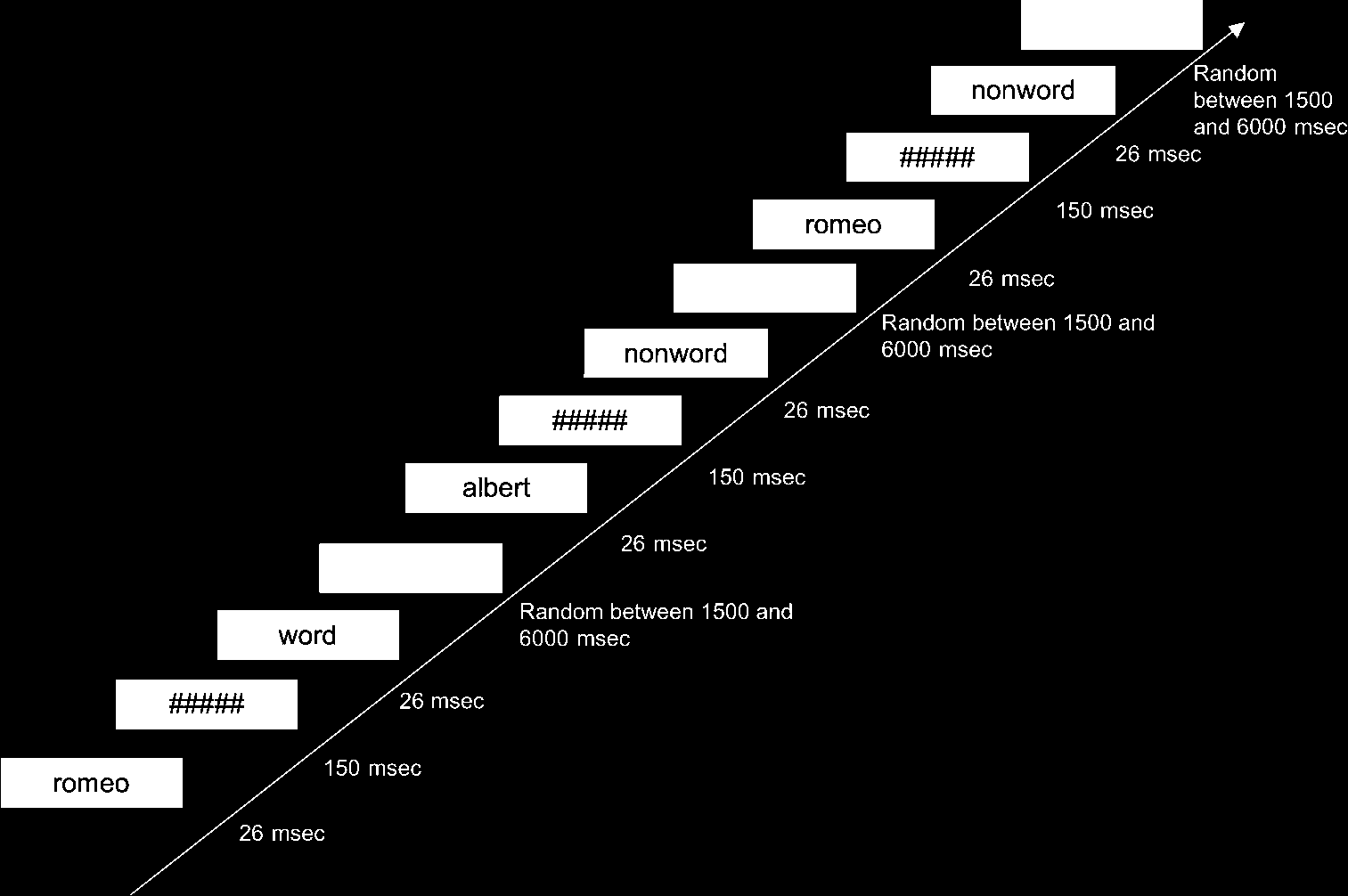
a sequence of three frames (Cifra 1) following a stan-
dard subliminal priming paradigm. Primero, a prime word
was presented for 26 mseg, followed by a mask of
########## symbols for 150 msec and then the
target word for 26 mseg. Stimulus onset asynchrony
(SOA; es decir., the interval between the onset of the prime
and the onset of the target) era 176 mseg. Trials were
separated by an interstimulus interval randomly chosen
entre 1500 y 6000 msec to allow accurate model-
ing of the hemodynamic response. All stimuli were
presented using Cogent 2000 running in Matlab 7.0.1
under Windows XP, which provides control of display
durations and accurate recordings of reaction times.
Stimuli appeared in lowercase 43-point Courier New
font, in white on a black background.
Each trial was composed of one of three primes
together with one of 40 palabras, or one of 40 nonwords,
or one of 40 ‘‘blank’’ trials, giving a total of 360 posible
ensayos. Each of the possible trials was then randomly
assigned to one of six blocks by means of a Latin square,
which ensures that each block contained an equal
number of beloved primes, passion primes, and friend
primes, and an equal number of words, nonwords, y
blanks; and words were never presented twice in a
block. Trial order within a block was pseudorandomized
with the constraint of no more than three consecutive
trials with the same target type. Each participant per-
formed six blocks with 60 trials in each block, for a total
de 360 ensayos, which took up to 40 min including breaks
between each block.
Prime Stimuli
Para cada participante, three unique words (3 a 11
characters long) were used as prime stimuli. The first
1220
Revista de neurociencia cognitiva
Volumen 19, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
9
mi
7
–
1
pag
2
d
1
F
8
/
1
1
9
9
/
3
6
7
3
/
9
1
6
2
1
oh
8
C
/
norte
1
2
7
0
5
0
6
7
8
1
1
9
0
/
7
j
1
oh
2
C
1
norte
8
.
pag
2
d
0
0
b
7
y
.
gramo
1
tu
9
mi
.
s
7
t
.
oh
1
norte
2
0
1
7
8
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
F
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Cifra 1. Experimental
diseño. Stimulus sequence.
The visual stimulus on each
trial was composed of a
sequence of three frames. Primero,
a prime word was presented
para 26 mseg, followed by a
mask of ##########
symbols for 150 msec and then
the target word for 26 mseg.
Subjects were not informed
of the presence of the prime.
The type of target letter strings
(emotional words, nonwords,
or blanks) and the type of the
primes (beloved, p.ej., Romeo;
neutral friend, p.ej., Alberto;
and passion, p.ej., piano) eran
randomly presented according
to a Latin square calculated
over the six experimental
bloques. This means a target
stimulus was not presented
twice in the same block in
order to avoid any effects of
familiarization. Además,
the order of experimental
trials was random, con el
constraint of no more than
three consecutive trials with
the same target type.
name (either full first name, such as Robert, or short-
ened first name, such as Bob) of the participant’s
beloved was designated as the ‘‘love prime.’’ No nick-
name was used in the present study. A noun describing
the participant’s passion in life (p.ej., dance, piano,
softball), as supplied by the participant, was designated
as the ‘‘passion prime,’’ and finally, the name of a friend
was designed as the ‘‘friend prime’’ or ‘‘control prime.’’
No difference in name length between love, amigo, y
passion primes was observed, F(2,70) = 2.69; pag = .08.
The presence of the prime was not mentioned to the
subjects. To check that participants were not aware of
the type of prime stimuli, we used an extensive debrief-
ing procedure in which participants were asked increas-
ingly specific questions about the study. This procedure
revealed that all participants reported that they had seen
the flash of the mask. Sin embargo, no participant could
report on the specific emotional or semantic contents of
the flashes.
Target Stimuli
were selected to be high in written word frequency,
and the other half were low in written word frequency
according to the Kucera and Francis linguistic database
(Kucera & Francisco, 1967). Además, a 2 (cid:1) 3 analysis of
variance (ANOVA) performed between low- versus high-
frequency targets and the prime types did not show any
main interaction: F(2,70) = .44, pag = .65 for reaction
veces; F(2,70) = .41, pag = .96 for accuracy. All words
were selected to be high in valence and high in arousal
(Bradley & Lang, 1999). High- (m = 220.95, DE =
144.67) and low-frequency (m = 27.6, DE = 27.13)
emotional words were matched in word length (alto-
frequency words: m = 5.9, DE = 1.48; low-frequency
palabras: m = 6.3, DE = 1.95; pag = .48), valence (alto-
frequency words: m = 7.40, DE = 0.64; low-frequency
palabras: m = 7.55, DE = 0.66; pag = .57), arousal (alto-
frequency words: m = 5.89, DE = 0.44; low-frequency
palabras: m = 6.42, DE = 1.17; pag = .054), abstractness
(high-frequency words: m = 407,38, DE = 123,64; bajo-
frequency words: m = 404.6, DE = 118.84; pag = .95;
Coltheart, 1981).
El 80 target letter-string stimuli (3 a 11 characters
largo) incluido 40 positive emotional English nouns and
a set of 40 pronounceable nonwords (following the
same consonant/vowel structure as words). Emotional
words were selected from the Affective Norms for
English Words (Bradley & Lang, 1999). In order to con-
trol any effect of word frequency, one half of the words
Imagen de resonancia magnética
Imaging was performed with a 3T Phillips MRI scanner
using an eight-channel phased-array head coil. For each
functional run, an echo planar gradient-echo imaging
sequence sensitive to BOLD contrast was used to ac-
quire 30 slices per repetition time (TR) (4 mm thickness,
Ortigue et al.
1221
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
9
mi
7
–
1
pag
2
d
1
F
8
/
1
1
9
9
/
3
6
7
3
/
9
1
6
2
1
oh
8
C
/
norte
1
2
7
0
5
0
6
7
8
1
1
9
0
/
7
j
1
oh
2
C
1
norte
8
.
pag
2
d
0
0
b
7
y
.
gramo
1
tu
9
mi
.
s
7
t
.
oh
1
norte
2
0
1
7
8
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
F
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
0.5 mm gap), with a TR of 1976 mseg, tiempo de eco (EL) de
35 mseg, flip angle of 908, field of view (FOV) de 240 mm,
y 80 (cid:1) 80 matrix. One hundred fifty-seven whole-
brain images were collected in each run. After all the
functional runs, a high-resolution T1-weighted image of
the whole brain was acquired using a spoiled gradient
recalled 3-D sequence (TR = 9.9 mseg; TE = 4.6 mseg;
flip angle = 88, FOV = 240 mm; slice thickness = 1 mm,
matrix = 256 (cid:1) 256).
Functional Image Processing
All analysis was carried out in SPM2 (www.fil.ion.ucl.ac.
uk/spm). Primero, functional images were realigned to cor-
rect for head movement. Entonces, data were coregistered
to the SPM2 EPI template, and normalized to a stan-
dardized Montreal Neurological Institute (MNI) stereo-
taxic space to give images with 2 (cid:1) 2 (cid:1) 2-mm voxels.
A design matrix was fitted for each subject with the
trials in each cell of the three-by-three factorial de-
sign (3 primes (cid:1) 3 target types) modeled by a standard
hemodynamic response function and its temporal de-
rivative. Each trial was modeled as a single event with
zero duration, starting at the onset of the prime stimu-
lus. Rest was not modeled. The design matrix weighted
each raw image according to its overall variability to re-
duce the impact of movement artifacts (Diedrichsen &
Shadmehr, 2005). The design matrix was fit to the data
for each participant individually. After estimation, betas
were smoothed (10-mm full width half maximum) y
taken to the second level for random effect analysis.
Segundo(cid:2)level Data Analysis
Our analysis aimed to identify which brain areas within
the standard ‘‘prime network’’ were preferentially acti-
vated by love and passion regardless of the target type.
To this purpose, we first identified the brain regions in-
volved in the primed lexical decision task. This contrast of
all primes > rest was thresholded at p < .001 uncorrected
and used to generate an inclusive mask of the priming
network in the brain. Within this mask, we calculated con-
trasts for love > amigo, passion > friend, love > passion,
and passion > love, and report results that passed a
threshold of p < .01 and a cluster size of 10 voxels. We
used a masking approach to restrict the likelihood of
false positives rather than a region-of-interest approach,
because the lack of previous studies of subliminal prim-
ing for love and passion means that we cannot make
strong a priori predictions of the locations of our results.
that used a recruitment method similar to ours, we first
calculated correlations between the various scores we
obtained. We tested whether the questionnaire data cor-
related significantly with relationship length or percent
of thoughts (or whether any of these variables corre-
lated with each other); they did not (Table 1). That is,
as was found by Aron et al. (2005), there were no sig-
nificant correlations among PLS scores and length of
being in love (Pearson correlation, r = .15, p = .39).
Because in the present study, scores obtained at the
PLS, 9-point scale, and the percent of daily thoughts
dedicated to the beloved were highly correlated with
each other (Table 1), we calculated a composite mea-
sure by combining the individual measures of these
three scales (averaged as Z scores). There was no cor-
relation between this composite measure and length
of being in love (Pearson correlation, r = .15, p = .38;
Table 1).
Our behavioral results showed that both the sublimin-
al presentation of passion (580 ± 19.58 msec [mean ±
SE]) and love (581 ± 19.92 msec [mean ± SE]) induced
beneficial effects (faster reaction times) on the explicit
detection of words, love: F(1,35) = 5.22; p = .03; pas-
sion: F(1,35) = 6.74; p = .01, compared to the sub-
liminal presentation of a control friend’s name (607 ±
23.4 msec [mean ± SE]). No behavioral difference was
found between love and passion ( p = .95). No specific
beneficial effect of prime was found for nonword detec-
tion, F(2,70) = 0.15; p = .87. However, a general main
effect of prime was found independently of the target,
F(2,70) = 4.09; p = .02, indicating that subjects were
faster to respond after a beloved prime (608 ± 19.49
[mean ± SE]) or a passion prime (607 ± 19 [mean ±
Table 1. Pearson’s r Correlation Coefficient between Each
Measure of Love (PLS, 9-Point Scale, and Percent of Thoughts)
and Length of Duration
Pearson’s r
Correlation
Coefficient
9-Point
Intensity
Scale
PLS
Percent of
Thoughts
Length of
Duration Composite
PLS
1
S***
S***
ns
9-Point
.86
1
S***
ns (trend)
intensity
scale
Percent of
thoughts
Length of
duration
.73*** .61***
1
.15
.33
(cid:2).02
ns
1
Composite
–
–
–
–
–
–
.15
1
RESULTS
Behavioral Results
In order to compare our results with two previous
fMRI studies (Aron et al., 2005; Bartels & Zeki, 2000)
Composite averages the individual measures of the three love scales
(as Z scores), that is, PLS, 9-point intensity scale, and percent of
thoughts. PLS = passionate love scale; S = significant; ns = non-
significant, p > .05; trend = trend of significance, .05 < p < .06.
***p < .001.
1222
Journal of Cognitive Neuroscience
Volume 19, Number 7
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
a
e
-
r
p
t
d
i
c
1
l
9
e
7
-
1
p
2
d
1
f
8
/
1
1
9
9
/
3
6
7
3
/
9
1
6
2
1
o
8
c
/
n
1
2
7
0
5
0
6
7
8
1
1
9
0
/
7
j
1
o
2
c
1
n
8
.
p
2
d
0
0
b
7
y
.
g
1
u
9
e
.
s
7
t
.
o
1
n
2
0
1
7
8
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
SE]) than after a friend prime (621 ± 21.41 [mean ±
SE]). A 2 (cid:1) 3 ANOVA involving simple effects of primes
(love, friend, passion) and letter-string types (words,
nonwords) revealed a trend of significant interaction,
F(2,70) = 2.83, p = .06, suggesting that participants tend
to be faster in detecting target words (and not non-
words). This effect occurred especially if the target
words were primed by love/passion (as opposed to
friendship). An overall 3 (cid:1) 3 ANOVA did not show any
interaction between the different types of targets
(words, nonwords, blanks) and the different types of
primes (love, friend, passion), F(4,140) = 1.79; p = .13,
mainly due to absence of main effect of primes for a
blank target, F(2,70) = 0.28, p = .75.
A trend of negative correlation was observed between
PLS scores and reaction times (the more participants
were in love, the faster they were) for target stimuli
primed by a beloved’s name (Pearson correlation, r =
(cid:2).29, p = .04). A similar trend of correlation was found
between PLS scores and reaction times to passion
primes (Pearson correlation, r = (cid:2).29, p = .04). No
similar correlation was found between PLS scores and
reaction times to friend primes (Pearson correlation, r =
(cid:2).25, p = .07). A test evaluating the significance of
differences between these two correlation coefficients
(r = (cid:2).25 and r = .29) was nonsignificant ( p = .86).
In addition, a negative correlation was found between
the percentage of daily thoughts for the partner and
reaction times (the more participants thought about
their beloved every day, the faster they were) for target
stimuli primed by a beloved’s name (Pearson correla-
tion, r = (cid:2).40, p = .008). A similar negative correlation
was also found between percentage of thoughts for the
beloved and reaction times to passion primes (Pearson
correlation, r = (cid:2).38, p = .01) as well as with reaction
times to friend primes (Pearson correlation, r = (cid:2).29,
p = .04). A test evaluating the significance of differences
between the two correlation coefficients related to love
and passion primes (r = (cid:2).40 and r = (cid:2).38) or between
love/passion and friend primes was nonsignificant (love
vs. passion: p = .92; love vs. friend: p = .61; passion vs.
friend: p = .68).
No significant correlation was observed between
length of love relationship and reaction times (Pearson
correlation, for beloved primes: r = .21, p = .11; for
friend primes: r = .20, p = .12; for passion primes: r =
.24, p = .08).
No significant effects were observed for accuracy. An
overall 3 (cid:1) 3 ANOVA did not show any interaction
between the different types of targets and primes,
F(4,140) = .78, p = .54. A 2 (cid:1) 3 ANOVA involving simple
effects of primes (love, friend, passion) and letter-string
types (words, nonwords) did not reveal any significant
interaction, F(2,70) = 0.037, p = .96. There was no
main effect of love, F(1,35) = 0.93, p = .34, or passion,
F(1,35) = 0.68, p = .41, on the detection of target words,
compared to the subliminal presentation of neutral
friend’s name. There was also no specific beneficial effect
of prime on target nonword detection, F(2,70) = 0.06;
p = .94, or target blank detection, F(2,70) = 0.49, p = .61.
No general main effect of prime was found indepen-
dently of the target, F(2,70) = 0.31; p = .73.
Functional Imaging Results
First, our functional results showed that the subliminal
presentation of a beloved’s name (as opposed to a
friend’s name) recruited a cerebral network that is partly
concordant with the cerebral network previously found
for the explicit presentation of love-related stimuli (Aron
et al., 2005; Bartels & Zeki, 2000). More precisely, in
comparison with a control stimulus (love > friend con-
contraste), the subliminal presentation of a beloved’s name
recruited the caudate nucleus, ventral tegmental area,
insula, bilateral fusiform regions, parahippocampal gyri,
angular gyrus, left dorsolateral middle frontal gyrus, izquierda
inferior temporal gyrus, occipital cortex, and cerebel-
lum (Mesa 2). The two previous studies (Aron et al.,
2005; Bartels & Zeki, 2000) that investigated the expli-
cit mechanisms of love using a similar contrast showed
brain area activation restricted to the mid insula, ante-
rior cingulate cortex, head of the caudate nucleus,
ventral tegmental area, putamen, bilateral posterior hip-
pocampus, left inferior frontal gyrus, left middle tem-
poral gyrus, and cerebellum as well as the right parietal
lobe (only in Aron et al., 2005).
Por otro lado, our study demonstrated that the
subliminal presentation of another motivator, como una
passion’s descriptor (as compared to a friend’s name,
es decir., passion > friend contrast), showed greater activa-
tion in the caudate nucleus,
insula, bilateral fusiform
regiones, parahippocampal gyri, right angular gyrus, oc-
cipital cortex, lingual gyrus, and cerebellum (Mesa 2).
Tomados juntos, our functional results thus showed
potentially overlapping brain activation between love
(love > friend contrast) and passion primes (passion >
friend contrast). Both love primes (love > friend con-
contraste) and passion primes (passion > friend contrast) led
to increases in BOLD signal in regions within motivation
and emotion systems, compared with a control friend
prime ( pag < .01; Table 2). Specifically, both love and pas-
sion primes activated the caudate nucleus, insula, bi-
lateral fusiform regions, parahippocampal gyrus, right
angular gyrus, occipital cortex, and cerebellum.
Finally, when we directly compared BOLD responses
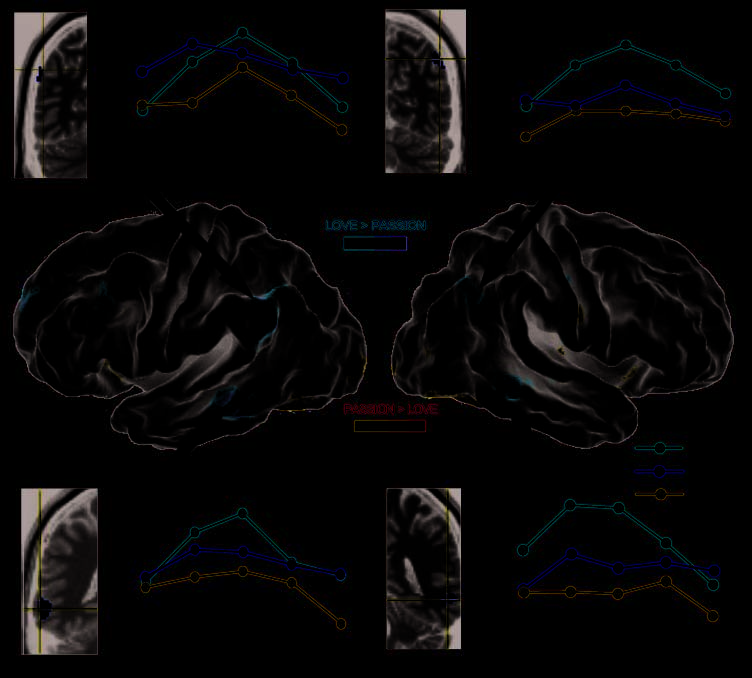
obtained from love and passion primes ( p < .01; Fig-
ure 2; Table 2), we found some differences of brain acti-
vation. Love primes (as opposed to passion primes, love
> passion contrast) revealed significant activation in bi-
lateral angular gyri (Mesa 1; in orange in Figure 2A and
B); and bilateral fusiform regions (Mesa 2; in orange in
Figure 2C and D).
Por otro lado, passion primes (Opuesto a
love primes, passion > love contrast) showed significant
Ortigue et al.
1223
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
9
mi
7
–
1
pag
2
d
1
F
8
/
1
1
9
9
/
3
6
7
3
/
9
1
6
2
1
oh
8
C
/
norte
1
2
7
0
5
0
6
7
8
1
1
9
0
/
7
j
1
oh
2
C
1
norte
8
.
pag
2
d
0
0
b
7
y
.
gramo
1
tu
9
mi
.
s
7
t
.
oh
1
norte
2
0
1
7
8
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
.
.
F
oh
norte
1
8
METRO
a
y
2
0
2
1
Mesa 2. MNI Coordinates of Cerebral Activation Peaks
Regions
Thalamus
Coordinates
Love > Friend
Passion > Friend
Love > Passion
Passion > Love
X
y
z
t
t
Cluster Size
t
Cluster Size
t
Cluster Size
t
Cluster Size
6 (cid:2)8
10 6.14
Caudate nucleus
6
12
0 5.34 4.39
Brainstem/midbrain
6 (cid:2)22 (cid:2)22 4.90 3.21
Insula
(cid:2)26
20 (cid:2)4 4.97 3.75
Occipitotemporal/
fusiform region
36 (cid:2)4
2 3.73 3.72
36 (cid:2)88 (cid:2)18 5.33 2.38
(cid:2)48 (cid:2)50 (cid:2)24 4.65 3.33
64 (cid:2)52
16 4.25
(cid:2)56 (cid:2)70 (cid:2)2 3.47
Parahippocampal gyrus
34 (cid:2)18 (cid:2)28 5.03 4.06
(cid:2)40 (cid:2)8 (cid:2)34 4.91 2.79
Angular gyrus
(cid:2)64 (cid:2)50
22 4.35
Dorsolateral middle
frontal gyrus
66 (cid:2)44
32 4.33 2.45
(cid:2)46
52
26
38
26 4.51 3.09
22 3.78
Superior temporal gyrus
66 (cid:2)14 (cid:2)8 4.18
80
13
2275
291
49
57
405
46
27
112
3.49
193
3.47
3.01
2.62
2.47
2.71
3.15
7
193
17
21
21
100
3.11
2.66
148
113
2.58
17
3.18
52
3.30
191
2.62
46
2.65
2.43
51
68
Inferior temporal gyrus (cid:2)44 (cid:2)26 (cid:2)20 4.39 2.70
43
2.40
21
Occipital cortex
(cid:2)18 (cid:2)96 (cid:2)8 6.00 2.51
138
(cid:2)64 (cid:2)22 (cid:2)10 4.52
Precuneus
18 (cid:2)94 (cid:2)8 5.22
16 (cid:2)76
20 4.32
2.61
3.72
42
744
Lingual gyrus
(cid:2)12 (cid:2)68 (cid:2)2 4.05
2.41
52
Cingulate gyrus
10 (cid:2)40
20 3.97
(cid:2)6 (cid:2)42
8 3.25
Precentral gyrus
64
2
24 3.66
Cerebellum
12 (cid:2)48 (cid:2)38 5.17
2.76
(cid:2)36 (cid:2)78 (cid:2)52 4.04
4.3
142
651
3.48
4.45
518
510
3.21
2.96
19
160
2.63
184
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
9
mi
7
–
1
pag
2
d
1
F
8
/
1
1
9
9
/
3
6
7
3
/
9
1
6
2
1
oh
8
C
/
norte
1
2
7
0
5
0
6
7
8
1
1
9
0
/
7
j
1
oh
2
C
1
norte
8
.
pag
2
d
0
0
b
7
y
.
gramo
1
tu
9
mi
.
s
7
t
.
oh
1
norte
2
0
1
7
8
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
F
.
.
.
.
.
activation in the bilateral temporo-occipital
junction,
left parahippocampal and inferior temporal gyrus, izquierda
cingulate gyrus, right precentral gyrus, and cerebellum
(Mesa 2; in blue in Figure 2).
In order to specifically compare our results with those
obtained previously by Aron et al. (2005), we performed
a between-subjects random effect analysis correlating
BOLD responses and participant’s scores on the PLS
puntuaciones, first focusing on the love > friend contrast, como
the Aron et al. analysis did. This method used previously
by Aron et al. is known to provide strong evidence for
the link of a function with an activated area. In the Aron
et al. estudiar, PLS scores had high positive correlations
with activation in two of the regions that were signifi-
cant for the contrast by itself, the right anteromedial
caudate body (r= .60, pag = .012) and the septum–fornix
región (r= .54, pag = .008). Eso es, the Aron et al. estudiar
participants who self-reported higher levels of love than
others also showed greater activation than others in
this region of the caudate and septum when viewing
their beloved (Aron et al., 2005). (For planned compari-
hijos, Aron et al. applied small volume corrections with
a sphere as a region of interest. The coordinates for the
centers of the following regions of interest were re-
ported: VTA, caudate nucleus, putamen, accumbens/
subcallosal cortex/ventral striatum, amygdala, posterior
oh
norte
1
8
METRO
a
y
2
0
2
1
1224
Revista de neurociencia cognitiva
Volumen 19, Número 7
Cifra 2. BOLD responses
obtained for love compared
to passion (love > passion
in orange; passion > love in
azul), shown on lateral views
of the fiducial left and right
side of the brain. BOLD time
courses, averaged over prime
tipo (rojo, love; azul, pasión;
yellow, amigo) were extracted
for the significant regions
between love and passion
(love > passion) contrasts
and are shown for the angular
gyrus (A, izquierda; B, bien) y
fusiform gyrus (C, izquierda; D,
bien). Brain activities were
mapped using the PALS human
cortical atlas from Caret5
software (VanEssen, 2005).
hippocampus, cingulate, insula, retrosplenial cortex, y
medial and lateral orbitofrontal cortex). As we did for
our previous analyses described above, we performed
this focused ‘‘love > friend contrast’’ between-subjects
random effect analysis within the prime network (as de-
scribed in Table 2). We corrected our correlations for
multiple comparisons and only allowed a significant
threshold at p < .01. Our results showed similar and
also distinct findings as compared to the findings of Aron
et al. Focusing on the love > friend contrast, our find-
ings showed that PLS scores had high positive corre-
lations with activation in some of the cerebral regions
that were significant for the contrast by itself, eso es,
the caudate nucleus (r= .48, pag = .003), right para-
hippocampal gyrus (Correlación de Pearson, r= .46, pag =
.005), angular gyrus (r= .39, pag = .01), insula (izquierda: r=
.41, pag = .01; bien: r= .38, pag = .02), ventral tegmental
área (r= .38, pag = .02), and the left dorsolateral mid-
dle frontal gyrus (r= .40, pag = .01). Eso es, Participantes
who self-reported higher levels of love than others ex-
hibited greater activation than others in these brain
regiones.
A similar method of analysis applied for the passion >
friend contrast (as reported in Table 2) did not reveal
similar significant correlation between BOLD responses
and participant’s scores on the PLS scores.
Focusing on the love > passion contrast (as described
en mesa 2), we found the same method of analysis
showed high positive correlations between PLS scores
and three of the cerebral regions that were significant
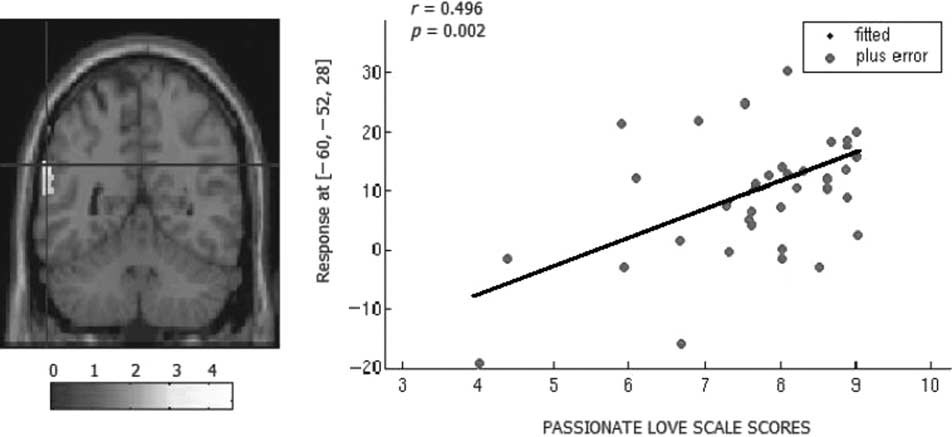
for the contrast by itself, the left angular gyrus (Pearson
correlation, r= .50; pag = .002; Cifra 3, Mesa 3) y el
fusiform region bilaterally (izquierda: r= .46; pag = .005; bien:
r= .47; pag = .003; Mesa 3). Eso es, participants who self-
reported higher levels of love than others also exhibited
greater activation than others in these brain regions.
Because no studies have investigated the love > passion
contrast previously, we also performed additional ana-
lyses correlating BOLD responses and the different love
measures (9-point subjective intensity scale, por ciento de
thoughts, length of being in love). This approach offers
a broad overview of the present findings, although they
were highly correlated with each other (as shown in
Mesa 1). The 9-point subjective intensity scale had simi-
larly correlated with the same brain regions: la izquierda
angular gyrus (r= .43, pag = .009) and the fusiform region
bilaterally (izquierda: r= .46, pag = .005; bien: r= .44, pag = .007;
Mesa 3). Por otro lado, percent of thoughts only
showed a positive correlation with the right fusiform
región (r= .43, pag = .008) and a trend to a positive cor-
relation with the left angular gyrus (r= .396, pag = .017;
Mesa 3). No correlation was observed between the
Ortigue et al.
1225
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
9
mi
7
–
1
pag
2
d
1
F
8
/
1
1
9
9
/
3
6
7
3
/
9
1
6
2
1
oh
8
C
/
norte
1
2
7
0
5
0
6
7
8
1
1
9
0
/
7
j
1
oh
2
C
1
norte
8
.
pag
2
d
0
0
b
7
y
.
gramo
1
tu
9
mi
.
s
7
t
.
oh
1
norte
2
0
1
7
8
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
F
.
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Cifra 3. Brain activation correlated with the passionate love scale (PLS) scores of participants ( pag < .01 corrected). (Left) Angular gyrus location for the correlation. (Right) Correlation of activity in the left angular gyrus with PLS scores. Participants who scored higher on the PLS scale showed stronger activations in a specific site ((cid:2)60, (cid:2)52, 28; x, y, z [mm] MNI coordinates) of a larger part of the angular gyrus. length of time in love and the regions that were signif- icantly activated for this specific contrast. DISCUSSION Taken together, our results provide the first functional evidence that the subliminal presentation of a beloved’s name (as compared to a friend’s name) activates a spe- cific pattern of brain activity, and these regions are more active for people who are more in love. This network for implicit effects of a beloved’s name is partly concor- dant with the love-related network demonstrated by previous studies that investigated the explicit mecha- nisms of love. Nevertheless, our findings also reveal the involvement of some distinct brain areas. Thus, our results may have important future implications for un- derstanding the unconscious mental representation of love in the brain. Love as an Implicit Motivator First, we aimed to test whether facilitation priming effects could occur for unconscious representation of a beloved’s name using a subliminal priming paradigm. The results are clear-cut. Two main results are important in this respect. First, we found that participants were faster to detect target stimuli when subliminally primed by a beloved’s name, and not when subliminally primed by a friend’s name. This phenomenon occurred espe- cially for word detection. This demonstrates that the subliminal facilitation effect of love occurs at an associa- tive level rather than a perceptual level. Women in love are not inherently faster based on a general emotional arousal but they are faster because of the mental asso- ciations they unconsciously created in their mind in relation to their internal state of being passionately in love. This indicates that subliminal passionate/emotional associations are sufficient to lead to automatic priming Table 3. MNI Coordinates of Correlations between BOLD Responses for Love Minus Passion and Three Different Measures of Love (PLS, 9-Point Scale, and Percent of Thoughts) Coordinates PLS Scores 9-Point Scale Percent of Thoughts D o w n l o a d e d f r o m l l / / / / / j f / t t i t . : / / D h t o w t p n : o / a / d m e i d t f p r o r m c . h s i p l v d e i r r e c c h t . a m i r e . d c u o m o / c j n o a c r t n i c / a e - r p t d i c 1 l 9 e 7 - 1 p 2 d 1 f 8 / 1 1 9 9 / 3 6 7 3 / 9 1 6 2 1 o 8 c / n 1 2 7 0 5 0 6 7 8 1 1 9 0 / 7 j 1 o 2 c 1 n 8 . p 2 d 0 0 b 7 y . g 1 u 9 e . s 7 t . o 1 n 2 0 1 7 8 S . e p p d f e m b b y e r g 2 u 0 e 2 s 3 t / j t f . . . . . o n 1 8 M a y 2 0 2 1 Regions x y z t Love > Passion
pag
Cluster Size
pag
Cluster Size
Occipitotemporal/
fusiform region
(cid:2)48 (cid:2)50 (cid:2)24
4.65
64 (cid:2)52
Angular gyrus
(cid:2)64 (cid:2)50
66 (cid:2)44
16
22
32
4.25
4.35
4.33
3.11
2.66
2.65
2.43
.005
.003
.002
–
37
115
210
–
.005
.007
.009
–
23
24
40
–
pag
–
.008
.017
–
Cluster Size
–
49
8
–
1226
Revista de neurociencia cognitiva
Volumen 19, Número 7
on the basis of associative relatedness, which is defined
as the extent to which the activation of one concept will
call to mind another concept (p.ej., Spruyt et al. 2004).
Además, as expected, we found similar facilitation
effects with a passion prime, in comparison with a friend
prime. This highlights that the observed associative re-
latedness does not only call on emotional associations,
but also on motivation mechanisms. This is coherent
with Hebbian models of cortical language representa-
ción, which suggest that the representations of words
are considered to include not only words’ visual forms,
but also their related memories (p.ej., sounds, smells),
and contextual encoding (Pulvermuller, 1996, 1999).
Within such a Hebbian model, the behavioral priming
effect arises when subliminal love or passion primes ac-
tivate mental representations that are part of that par-
ticular state and, de este modo, implicitly modulates behavior, como
previously suggested for other facilitation priming ef-
efectos (Fergusson & Bargh, 2004; Innes-Ker & Niedenthal,
2002; Dehaene, Naccache, et al., 1998). En otras palabras,
our data suggest that the representation of a beloved’s
name may call for a goal-directed state sustained by
higher order mechanisms rather than a specific emo-
tion without any motivational or reward components
(Bianchi-Demicheli et al., 2006; Aron et al., 2005; a &
a, 1986). These selective facilitation effects also sug-
gest underneath associative mechanisms that may be due
to implicit automatic spreading of activation from both
emotional and motivational associations (p.ej., Spruyt et al.,
2004; Musch & Klauer, 2003; Innes-Ker & Niedenthal,
2002).
Unconscious Mental Representation
of a Loved One
Our fMRI results show that the subliminal presentation
of a beloved’s name (as compared to a friend’s name)
activates some cortical regions that are known to be
also recruited during the conscious presentation of be-
loved’s faces (Aron et al., 2005; Bartels & Zeki, 2000).
De este modo, our results suggest that the subliminal presenta-
tion of love-related stimuli correspond to variations in
neural response that extend beyond a visual memory for
subliminal words. The subliminal presentation of a be-
loved’s name may implicitly activate the structural visual
form of the word and also a more complex representa-
tion of the beloved. In agreement, our fMRI results high-
light that love calls for higher order motivational and
reward mechanisms rather than a specific emotion
per se. Primero, the overlap between the neural network
observed for love primes (love > friend contrast) y
that observed for passion primes (passion > friend con-
contraste) demonstrates that both love and passion primes
recruited emotion/motivation/dopamine systems (a
et al., 2005; Bartels & Zeki, 2000; Gisiger et al., 2000). En
accordance with the Dehaene–Changeux model as well
as the Aron et al. (2005) motivation assumption, estos
results reinforce our behavioral results by demonstrating
dopaminergic-driven facilitation effects of both love and
passion on reaction times. Entonces, the subliminal presen-
tation of a beloved’s name (as opposed to another
motivator such as a passion’s descriptor, love > passion
contrast) also show activation in higher order brain
areas, such as those known to be involved in conceptual
thinking and metaphors (von Bubnoff, 2005). Estos
differences of brain activation contrast with the equiva-
lent behavioral results we obtained for passion and love
primes. Although this result might seem surprising, es
coherent with Hebbian models of cortical
idioma
representación, which suggest that the representations
of words may be segregated throughout cerebral hemi-
spheres by their conceptual structures (Gisiger et al.,
2000; Pulvermuller, 1996, 1999).
Love and the Self
críticamente, love primes recruited bilateral fusiform and an-
gular gyri, two brain regions involved in integration of ab-
stract representations (Arzy, Seeck, Ortigue, Spinelli, &
Blanke, 2006; Jackson, Brunet, Meltzoff, & Decety, 2006;
sajonia & Kanwisher, 2003; Blanke, Ortigue, Landis, &
Seeck, 2002). Although some might attribute our effects
to some bottom-up attention mechanisms because of
the specific salience of the subliminal stimuli (anderson,
2005; Corbetta & Shulman, 2002; anderson & Phelps,
2001), this could not account for all brain activations.
Por ejemplo, even if we agree that arousal mechanisms
may take place when contrasting primes with different
prominencia (such as ‘‘love > friend’’ or ‘‘passion >
friend’’), automatic arousal mechanisms could not ex-
plain the difference of brain activation between two
stimuli with equivalent salience, or at least with equiva-
lent frequency of thoughts on target (such as love
primes and passion primes). En cambio, the implication of
these brain areas calls for higher order mechanisms. Nuestro
assumption is reinforced by the positive correlation
between BOLD responses in these brain areas and the
different measures of love (PLS scores,
intensidad, y
percent of thoughts). Por otro lado, the absence
of correlation between brain activation in these brain
areas and the length of being in love underlines that the
fusiform region and the angular gyrus do not seem di-
rectly ‘‘love time dependent.’’ This is coherent with the
only previous study that assessed this question (a
et al., 2005) and showed changes in several regions as
the relationship changes, but not in the angular gyrus
and the fusiform regions. En cambio, activity related to the
length of the relationship was found in the right insula,
the right anterior and posterior cingulate cortex, y el
right posterior cingulated/retrospenial cortex (a
et al., 2005).
Además, a growing body of evidence demonstrates
that the angular and fusiform gyri not only call for
automatic attentional mechanisms but also higher order
Ortigue et al.
1227
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
9
mi
7
–
1
pag
2
d
1
F
8
/
1
1
9
9
/
3
6
7
3
/
9
1
6
2
1
oh
8
C
/
norte
1
2
7
0
5
0
6
7
8
1
1
9
0
/
7
j
1
oh
2
C
1
norte
8
.
pag
2
d
0
0
b
7
y
.
gramo
1
tu
9
mi
.
s
7
t
.
oh
1
norte
2
0
1
7
8
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
.
F
.
oh
norte
1
8
METRO
a
y
2
0
2
1
procesos, such as episodic retrieval and conceptual
conocimiento (Ashby & O'Brien, 2005; Giesbrecht, Camblin,
& Swaab, 2004). Por ejemplo, love could be considered
as a learned response from episodic memories and
positive associative reinforcements that may be primary
(such as pleasant feeling) and secondary (como
mental association between the beloved’s presence
and good feelings; Brehm, Molinero, Perham, & Molinero,
2002; Herrero & Queller, 2000; Carlston & Herrero, 1996;
Sternberg & Barnes, 1988). More critically, the angular
gyrus is known to be involved in the abstract represen-
tation of the self (Arzy et al., 2006; Blanke et al., 2002).
This lets us speculate that the unconscious representa-
tion of a beloved’s name is among others directed to the
self. This is concordant with previous studies suggesting
that intense lifetime achievements (p.ej., love) are a cen-
tral human motivation to ‘‘expand the self’’ (a &
a, 1986, 1996; Epstein, 1973; Rogers, 1969; Blanco,
1959). Our findings are also in agreement with theories
in social psychology suggesting that the expansion of the
self involves two phases: expansion, then integration of
each new expansion by incorporating aspects (p.ej., nuevo
habilidades, attitudes, resources, behaviors) of another into
the self (a & a, 1986, 1996; Hatfield & Walster,
1978; Epstein, 1973; Rogers, 1969; Bataille, 1962; Blanco,
1959). The angular gyrus is important in (a) integración
of abstract representations of others ( Jackson et al.,
2006), (b) social cognition related to the ability to rea-
son about the contents of mental states, such as de-
sire (sajonia & Kanwisher, 2003), o ambos. Además, el
junction is an association brain area
temporoparietal
considered pivotal in carrying out cross-modal informa-
ción (Bremmer, Schlack, Duhamel, Graf, & Fink, 2001;
Calvert, Campbell, & Brammer, 2000). This is of particu-
lar importance for the concept of love, which is based
on multidimensional components (Sternberg & Barnes,
1988; Hendrick & Hendrick, 1986). Because ‘‘a love
experience will not expand the self if it cannot be inte-
grated’’ (a & a, 1986; James, 1890/1950), this brain
area might be crucial for self-expansion by integrating dif-
ferent love-related sensations and extracting a common,
abstract representation from them.
Possible Limitations and Future Directions
There are some factors that restrict our interpretation
of the data in this study and that could be examined
more closely in the future. Because there are not many
previous studies of the neural basis of love and none
about the neural basis of passion, our study constitutes
a first step and includes some limitations. Primero, ser-
cause no study has previously unraveled the implicit
neural basis of love and passion, we did not feel justi-
fied in using only regions of interest, which might
exclude important and novel results. En particular, el
reliance on a priori regions of interest increases the
likelihood that researchers focus on a narrowly defined
set of brain areas for particular cognitive functions.
Although that approach is appropriate for cognitive
processes such as face recognition where the neural
substrates are well defined, the field of fMRI research
into love and abstract concepts is so novel that we
do not feel a region-of-interest approach is sensible. Por
using a masking approach, we are able to obtain ade-
quate protection against false positives without limiting
our conclusions.
Segundo, in our experiment the participants were only
women. This selection criterion could make the results a
bit different from those obtained in the two previous
studies that used a mix of genders (Aron et al., 2005;
Bartels & Zeki, 2000).
Finalmente, due to the absence of a ‘‘passionate scale’’ for
hobby, as was available for love, no correlation between
intensity of passion and BOLD responses could be done
in the present study. This area would benefit from the
development of standardized questionnaires evaluating
the different types and degrees of passion. This would
be helpful to better understand the neural modulation
of a passion with respect to its intensity.
Our study of subliminal love primes is the first fMRI
investigation of this phenomenon, which is considered
to be both a concept and a motivational factor. Más
work will be needed to distinguish between different
types of passions (for objects or for hobbies) and differ-
ent types of love (for a partner or a child) and to better
understand the conceptual framework underlying love
representations in the brain.
Conclusión
The present study reveals that the neuroscience of love
might provide not only essential clues for understanding
emotion and motivation systems, but also how one’s
unconscious may be enhanced by desire.
Expresiones de gratitud
We thank Prof. METRO. Gazzaniga for his helpful comments on a
previous version of the manuscript. This study was supported
by the Swiss National Foundation for research in Biology and
Medicamento (grant 1223/PASMA 111563/1). The experiment was
carried out using Matlab 7.0.1 and Cogent 2000 developed at the
Laboratory of Neurobiology and the Wellcome Department of
Neurociencia de la imagen, University College London. Los autores
declare that they have no competing financial interests.
Reprint requests should be sent to Scott T. Grafton, Sage Center
for the Study of the Mind and Department of Psychology,
Edificio 251, Room 3837, Universidad de California, Santa Bárbara,
California 93106, o por correo electrónico: grafton@psych.ucsb.edu or ortigue@
psych.ucsb.edu.
REFERENCIAS
anderson, A. k. (2005). Affective influences on the attentional
dynamics supporting awareness. Revista de experimentación
Psychology and Genetics, 134, 258–281.
1228
Revista de neurociencia cognitiva
Volumen 19, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
9
mi
7
–
1
pag
2
d
1
F
8
/
1
1
9
9
/
3
6
7
3
/
9
1
6
2
1
oh
8
C
/
norte
1
2
7
0
5
0
6
7
8
1
1
9
0
/
7
j
1
oh
2
C
1
norte
8
.
pag
2
d
0
0
b
7
y
.
gramo
1
tu
9
mi
.
s
7
t
.
oh
1
norte
2
0
1
7
8
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
.
F
.
oh
norte
1
8
METRO
a
y
2
0
2
1
anderson, A. K., & Phelps, mi. A. (2001). Lesions of the
human amygdala impair enhanced perception of
emotionally salient events. Naturaleza, 411, 305–309.
a, A., & a, mi. norte. (1986). Love and the expansion
of self: Understanding attraction and satisfaction.
Nueva York: Hemisphere.
a, A., & a, mi. norte. (1991). Love and sexuality.
In K. McKinney & S. Sprecher (Editores.), Sexuality in
close relationships (páginas. 25–48). Hillsdale, Nueva Jersey: Erlbaum.
a, A., & a, mi. norte. (1996). Love and expansion of
the self: The state of the model. Personal Relationships,
3, 45–58.
a, A., Pescador, h., Mashek, D. J., Strong, GRAMO., li, h., &
Marrón, l. l. (2005). Reward, motivación, and emotion
systems associated with early-stage intense romantic love.
Revista de neurofisiología, 94, 327–337.
Arzy, S., Seeck, METRO., Ortigue, S., Spinelli, l., & Blanke, oh.
(2006). Induction of an illusory shadow person. Naturaleza,
443, 287.
Ashby, F. GRAMO., & O'Brien, j. B. (2005). Category leaning and
multiple memory systems. Tendencias en Ciencias Cognitivas,
9, 83–89.
Bartels, A., & Zeki, S. (2000). The neural basis of romantic
love. NeuroReport, 11, 3829–3834.
learning in neuronal networks for planning and decision
haciendo. Progress in Brain Research, 126, 217–229.
Dehaene, S., Changeux, j. PAG., Naccache, l., Sackur, J., &
Sergent, C. (2006). Conscious, preconscious, y
subliminal processing: A testable taxonomy. Trends
in Cognitive Sciences.
Dehaene, S., Kerszberg, METRO., & Changeux, j. PAG. (1998).
A neuronal model of a global workspace in effortful
cognitive tasks. Proceedings of the National Academy
of Sciences, EE.UU., 95, 14529–14534.
Dehaene, S., Naccache, l., Le Clec, h. GRAMO., Koechlin, MI.,
Mueller, METRO., Dehaene-Lambertz, GRAMO., et al. (1998).
Imaging unconscious semantic priming. Naturaleza, 395,
597–600.
Diedrichsen, J., & Shadmehr, R. (2005). Detecting and
adjusting for artifacts in fMRI time series data. Neuroimagen,
27, 624–634.
Eimer, METRO., & Schlaghecken, F. (1998). Effects of masked
stimuli on motor activation: Behavioral and
electrophysiological evidence. Revista de experimentación
Psychology Human Perception and Performance, 24,
1737–1747.
Epstein, S. (1973). The self-concept revisited: Or a theory
of a theory. Psicólogo americano, 28, 404–416.
Bartels, A., & Zeki, S. (2004). The neural correlates of
Esch, T., & Stefano, GRAMO. B. (2005). The neurobiology of love.
maternal and romantic love. Neuroimagen, 21, 1155–1166.
Bataille, GRAMO. (1962). Eroticism (METRO. Dalwood, Trans.). Londres:
Calder.
Berscheid, MI., & Meyers, S. A. (1996). A social categorical
approach to a question about love. Personal Relationships,
3, 19–43.
Bianchi-Demicheli, F., Grafton, S. T., & Ortigue, S. (2006).
The power of love on the human brain. Social
Neurociencia, 1, 90–103.
Blanke, o., Ortigue, S., Landis, T., & Seeck, METRO. (2002).
Stimulating illusory own-body perceptions. Naturaleza, 19,
269–270.
Bradley, METRO. METRO., & Lang, PAG. j. (1999). Affective norms for
English words (ANEW): Estímulos, instruction manual
and affective ratings (Technical report C-1). Gainesville,
Florida: The Center for Research in Psychophysiology,
University of Florida.
Brehm, S. S., Molinero, r., Perham, D., & Molinero, C. S. (2002).
Intimate relationships (3tercera ed.). Nueva York: McGraw-Hill
Humanities.
Bremmer, F., Schlack, A., Duhamel, j. r., Graf, w., & Fink, GRAMO. R.
(2001). Space coding in primate posterior parietal cortex.
Neuroimagen, 14, S46–S51.
Buss, D. METRO. (2003). The evolution of desire. Nueva York:
Libros Básicos.
Neuro Endocrinology Letters, 26, 175–192.
Fazio, R. h., Sanbonmatsu, D. METRO., Powell, METRO. C., & Kardes,
F. R. (1986). On the automatic activation of attitudes.
Journal of Personality and Social Psychology, 50,
229–238.
Pescador, h. (2004). Why we love? Nueva York: Henry Holt.
Pescador, h., a, A., & Marrón, l. l. (2005). Romantic love:
An fMRI study of a neural mechanism for mate choice.
Journal of Comparative Neurology, 493, 58–62.
Giesbrecht, B., Camblin, C. C., & Swaab, t. Y. (2004).
Separable effects of semantic priming and imageability
on word processing in human cortex. Corteza cerebral,
14, 521–529.
Gisiger, T., Dehaene, S., & Changeux, j. PAG. (2000).
Computational models of association cortex. Actual
Opinion in Neurobiology, 10, 250–259.
Hatfield, MI., & Sprecher, S. (1986). Measuring passionate
love in intimate relationships. Journal of Adolescence, 9,
383–410.
Hatfield, MI., & Walster, GRAMO. W.. (1978). A new look at love.
Nueva York: Addison-Welsey.
Hendrick, C., & Hendrick, S. (1986). A theory and method
of love. Journal of Personality and Social Psychology, 50,
392–402.
Henson, R. norte. (2003). Neuroimaging studies of priming.
Calvert, GRAMO. A., Campbell, r., & Brammer, METRO. j. (2000).
Progress in Neurobiology, 70, 53–81.
Evidence from functional magnetic resonance imaging of
crossmodal binding in the human hetero-modal cortex.
Biología actual, 10, 649–657.
Carlston, D. MI., & Herrero, mi. R. (1996). Principles of mental
representación. In E. t. Higgins & A. Kruglanski (Editores.),
Social psychology: Handbook of basic principles
(páginas. 184–210). Nueva York: Guilford Press.
clark, METRO. S., & Mills, j. (1979). Interpersonal attraction
in exchange and communal relationships. Diario de
Personality and Social Psychology, 37, 12–24.
Coltheart, METRO. (1981). The MRC psycholinguistic database.
Revista trimestral de psicología experimental, 33A,
497–505.
Innes-Ker, A., & Niedenthal, PAG. METRO. (2002). Emoción
concepts and emotional states in social judgment and
categorization. Journal of Personality and Social
Psicología, 83, 804–816.
Jackson, PAG. l., Brunet, MI., Meltzoff, A. NORTE., & Decety, j. (2006).
Empathy examined through the neural mechanisms
involved in imagining how I feel versus how you feel
pain. Neuroimagen, 44, 752–761.
James, W.. (1950). The principles of psychology. Nueva York:
Dover. (Trabajo original publicado 1890.)
Kucera, h., & Francisco, W.. norte. (1967). Análisis computacional
of present-day American English. Providencia, Rhode Island: Marrón
Prensa universitaria.
Corbetta, METRO., & Shulman, GRAMO. l. (2002). Control of goal-directed
and stimulus-driven attention in the brain. Nature Review
Neurociencia, 3, 201–216.
Dehaene, S., & Changeux, j. PAG. (2000). Dependiente de la recompensa
Mashek, D., a, A., & Pescador, h. mi. (2000). Identifying,
evoking, and measuring intense feelings of romantic
love. Representative Research in Social Psychology, 24,
48–55.
Ortigue et al.
1229
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
9
mi
7
–
1
pag
2
d
1
F
8
/
1
1
9
9
/
3
6
7
3
/
9
1
6
2
1
oh
8
C
/
norte
1
2
7
0
5
0
6
7
8
1
1
9
0
/
7
j
1
oh
2
C
1
norte
8
.
pag
2
d
0
0
b
7
y
.
gramo
1
tu
9
mi
.
s
7
t
.
oh
1
norte
2
0
1
7
8
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
F
.
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Murphy, S. T., & Zajonc, R. B. (1993). Affect, cognition, y
awareness: Affective priming with optimal and suboptimal
stimulus exposures. Journal of Personality and Social
Psicología, 64, 723–739.
Musch, J., & Klauer, k. C. (2003). The psychology of
evaluación. Nueva York: Erlbaum.
Oldfield, R. C. (1971). The assessment and analysis of
handedness: The Edinburgh inventory. Neuropsicología,
9, 97–113.
Ovid (1986). Metamorphoses (A. D. Melville, Trans.). Nuevo
york: prensa de la Universidad de Oxford.
thinking people. The role of the temporo-parietal junction
in ‘‘theory of mind.’’ Neuroimage, 19, 1835–1842.
Herrero, mi. r., & Queller, S. (2000). Mental representations.
In A. Tesser & norte. Schwarz (Editores.), Blackwell handbook
in social psychology: volumen. 1, Intraindividual processes.
Oxford: Blackwell.
Spruyt, A., Hermans, D., De Houwer, J., & Eelen, PAG. (2004).
Automatic non-associative semantic priming: Episodic
affective priming of naming responses. Acta Psicológica,
116, 39–54.
Sternberg, j. S., & Barnes, METRO. l. (1988). The psychology of
Pulvermuller, F. (1996). Hebb’s concept of cell assemblies
love. nuevo refugio, CT: Prensa de la Universidad de Yale.
and the psychophysiology of word processing.
Psychophysiology, 33, 317–333.
Pulvermuller, F. (1999). Words in the brain’s language.
Ciencias del comportamiento y del cerebro, 22, 253–279; discussion
280–336.
Rogers, C. R. (1969). Freedom to learn. Columbus, OH:
Charles E. Merril.
VanEssen, D. C. (2005). A population-average, landmark-
and surface-based (PALS) atlas of human cerebral cortex.
Neuroimagen, 28, 635–662.
von Bubnoff, A. (2005). Stroke patients shed light on
metaphors [Electronic version]. Naturaleza. Retrieved
Puede 26.
Blanco, R. W.. (1959). Motivation reconsidered: The concept
sajonia, r., & Kanwisher, norte. (2003). People thinking about
of competence. Revisión psicológica, 66, 297–333.
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
D
h
t
oh
w
t
pag
norte
:
oh
/
a
/
d
metro
mi
i
d
t
F
pag
r
oh
r
metro
C
.
h
s
i
pag
yo
v
d
mi
i
r
r
mi
C
C
h
t
.
a
metro
i
r
mi
.
d
C
tu
oh
metro
oh
/
C
j
norte
oh
a
C
r
t
norte
i
C
/
a
mi
–
r
pag
t
d
i
C
1
yo
9
mi
7
–
1
pag
2
d
1
F
8
/
1
1
9
9
/
3
6
7
3
/
9
1
6
2
1
oh
8
C
/
norte
1
2
7
0
5
0
6
7
8
1
1
9
0
/
7
j
1
oh
2
C
1
norte
8
.
pag
2
d
0
0
b
7
y
.
gramo
1
tu
9
mi
.
s
7
t
.
oh
1
norte
2
0
1
7
8
S
.
mi
pag
pag
d
F
mi
metro
b
b
y
mi
r
gramo
2
tu
0
mi
2
s
3
t
/
j
t
.
.
.
.
.
F
oh
norte
1
8
METRO
a
y
2
0
2
1
1230
Revista de neurociencia cognitiva
Volumen 19, Número 7