Semantic Structural Alignment of Neural Representational
Spaces Enables Translation between English
and Chinese Words
Benjamin D. Zinszer1, Andrew J. Anderson1, Olivia Kang2*, Thalia Wheatley2,
and Rajeev D. S. Raizada1
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
Abstracto
■ Two sets of items can share the same underlying conceptual
estructura, while appearing unrelated at a surface level. Humanos
excel at recognizing and using alignments between such under-
lying structures in many domains of cognition, most notably in
analogical reasoning. Here we show that structural alignment
reveals how different people’s neural representations of word
meaning are preserved across different languages, such that
patterns of brain activation can be used to translate words from
one language to another. Groups of Chinese and English speakers
underwent fMRI scanning while reading words in their respective
native languages. Simply by aligning structures representing the
two groups’ neural semantic spaces, we successfully infer all seven
Chinese–English word translations. Beyond language translation,
conceptual structural alignment underlies many aspects of high-
level cognition, and this work opens the door to deriving many
such alignments directly from neural representational content. ■
INTRODUCCIÓN
Two sets of items can differ in their surface appearance
but share the same underlying conceptual structure.
People have a powerful ability to discover such structures
and infer new information by comparing overall the
similarities of known structures with new structures, semejante
as in analogical reasoning (Holyoak & Thagard, 1996),
category learning (Gentner & Namy, 1999), and word
aprendiendo (Gentner & Namy, 2004). This comparison
across structures is commonly referred to as structural
alignment because it is based not on commonalities of
the items themselves but instead on matching the struc-
tural relations between those items. Perhaps the most
important example is seen in language. Different lan-
guages’ lexicons are composed of very different word
formas, but these words describe the same world and thus
shared conceptual structures. Here we test whether
these shared concepts are similarly represented across
brains, regardless of differences in their surface (linguis-
tic) representación. Brain imaging offers the ability to
probe people’s neural activation and (more scientifically
interesante) the representational content that this acti-
vation carries. In behavioral studies of structural align-
mento, correspondences are identified between two sets
of items based on their common relational structure
(Gentner & Herrero, 2012). Here we investigate whether
1University of Rochester, 2Dartmouth College
*oh. Kang is now at the Department of Psychology, Harvard
Universidad.
© 2016 Instituto de Tecnología de Massachusetts
structural alignment can also be applied to neural repre-
sentaciones. This offers a strong test of the degree to which
neuroimaging techniques actually access the intended
conceptual content by asking the following question: Si
we take two sets of neural activity and align their neural
similarity structures, will that alignment reflect an accu-
rate alignment of the conceptual content?
Previously, we have shown that by matching neural
similarities across participants it was possible to perform
neural decoding across participants exposed to several
visual objects (Raizada & Connolly, 2012). Sin embargo, en
that study the different people were presented with stim-
uli that were the same not only conceptually but also per-
ceptually. Aquí, we ask the new question of whether the
process of matching neural similarity structures can yield
alignment at the level of linguistic concepts, even when
the surface appearances of the stimuli (word forms) son
completely different. To address that question, we used
the task of cross-language translation between English
y chino. These two languages differ greatly in sur-
face characteristics: A set of English words and their
Chinese translation equivalents have essentially nothing
in common phonetically or orthographically. We test
whether the process of aligning the neural similarity
structures of Chinese and English speakers is able to
correctly pair English words with their correct Chinese
translations.
Previous work has investigated correlations between
neural similarity structures, a saber, the important and
increasingly popular fMRI approach of representational
Revista de neurociencia cognitiva 28:11, páginas. 1749–1759
doi:10.1162/jocn_a_01000
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
1
2
1
8
/
1
1
7
1
4
/
9
1
1
7
9
4
5
9
1
/
9
1
6
0
7
8
oh
5
C
6
norte
0
_
2
a
/
_
j
0
oh
1
C
0
norte
0
0
_
a
pag
_
d
0
1
b
0
y
0
gramo
0
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
F
t
/
.
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
similarity analysis (RSA; Kriegeskorte, Mur, & Bandettini,
2008). Sin embargo, although RSA assesses the overall de-
gree of match between two sets of neural similarities, él
does not in itself produce an alignment between them. En
otras palabras, RSA does not find pairwise correspon-
dences between items in the structures that it compares.
Outside the domain of neural decoding, algorithms have
been developed to match sets of structural relations
(Turney, 2008; Goldstone & Rogosky, 2002; Laakso &
Cottrell, 2000; Falkenhainer, Forbus, & Gentner, 1989),
and this approach can be used to achieve across-subject
neural decoding (Raizada & Connolly, 2012). Going
beyond RSA’s correlation of overall neural similarity
estructura, structural alignment is used here to reveal the
pairwise correspondences between individual items.
Además, those items dissociate underlying meaning
from surface appearance, as the shared linguistic con-
cepts are represented in languages that are very differ-
ent, both visually and phonetically.
en este estudio, we perform structural alignment between
functional neural responses for speakers of different lan-
calibres. Between these groups, the surface appearances
for stimulus words differ dramatically, but we predict that
underlying conceptual structures are similar enough to
realign individual translation equivalent words. Usando
multivoxel pattern analysis and representational simila-
rity structures, we compare distributed functional brain
activity for separate groups of native speakers of Chinese
e inglés. We ask whether the similarity structures
among the functional brain responses to these word-
elicited concepts in each language are close enough in
structure across languages to perform neural decoding
on group level data and translate words in one language
into the other language.
MÉTODOS
Participantes
On the basis of the sample size of a previous, similar par-
adigm (Mitchell et al., 2008; norte = 9), we aimed to recruit
10 participants per language at Dartmouth College. adi-
tional participants were scheduled in case of failure to
complete the task or meet eligibility requirements (ver
abajo), y por lo tanto 11 English-speaking participants and
12 Chinese-speaking participants were included in the
estudiar. Behavioral data analysis revealed that one Chinese-
speaking participant did not provide complete behavioral
respuestas, and this participant was excluded from the
imaging analyses. De este modo, a total of 11 native speakers of
Inglés (4 M/7 F) y 11 native speakers of Mandarin
Chino (3 M/8 F) were available for the imaging analyses.
These analyses were performed after collection of the
entire sample, with no subsequent changes in sample size
due to exclusion or additional recruitment.
All participants were undergraduate students, graduate
estudiantes, or postdoctoral researchers. Participants self-
reported being native speakers of English or Mandarin
Chino, defined as being born in their native language
environment and speaking that language as their earliest
idioma. The English-speaking participants reported
no knowledge of Chinese language in a verbal screening
before the experiment. The Chinese-speaking partici-
pants were all Chinese–English bilinguals studying or
working in the United States. A separate screening (de-
scribed below) was used to evaluate the Chinese-speaking
participants’ knowledge of the English stimuli.
Materials
We selected seven translation equivalent words in
English and Chinese before the study, meeting four cri-
teria: (1) concrete nouns, (2) monosyllabic in both lan-
calibres, (3) represented by a single Chinese character,
y (4) unlikely for English translations to be known by
the Chinese participants. To ensure that criterion (4) era
met, Chinese participants completed a brief translation
task in which they were asked to write the English trans-
lation for 20 Chinese words. Mean translation accuracy
era 1.56 out of the seven critical stimuli. Although this
result may be surprising at first, this low level of accuracy
confirms that we were successful in our goal of choosing
words that do not often arise in an academic context.
The Chinese-speaking participants were largely graduate
students or postdoctoral researchers from the computer
science department and rarely needed to use these words
en Inglés.
The seven words that we used are listed in Table 1 con
some common lexical parameters. Frequency data were
obtained from film subtitle frequencies in American
English and Chinese (Cai & Brysbaert, 2010; Brysbaert &
Nuevo, 2009). Concreteness ratings were obtained from
Brysbaert, Warriner, and Kuperman’s (2014) norming
study for English words, and all seven critical stimuli
scored near ceiling (5). To our knowledge, an equivalent
Mesa 1. Seven Critical Word Stimuli in English and Chinese
Inglés
Chino
Word
Frecuencia
Concreteness
Word
Frecuencia
axe
broom
gown
hoof
jaw
mule
raft
1.81
2.13
3.14
0.72
3.37
2.54
1.56
5.00
4.89
4.61
4.89
4.87
5.00
5.00
(fuˇ)
(zho˘u)
(páo)
(tí)
(è)
(luó)
(fá)
2.33
0.03
4.98
2.41
0.63
0.42
0.21
Frequency data are per million words in the SUBTLEX film subtitles
database. Concreteness ratings are on a scale of 1 (most abstract) a
5 (most concrete). See Brysbaert et al. (2014) for details.
1750
Revista de neurociencia cognitiva
Volumen 28, Número 11
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
1
2
1
8
/
1
1
7
1
4
/
9
1
1
7
9
4
5
9
1
/
9
1
6
0
7
8
oh
5
C
6
norte
0
_
2
a
/
_
j
0
oh
1
C
0
norte
0
0
_
a
pag
_
d
0
1
b
0
y
0
gramo
0
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
F
/
t
.
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
database is not yet available in Chinese. The critical stimuli
were presented in three different font faces (Inglés:
Helvetica, American Typewriter, and Times New Roman;
Chino: STFangSong, Kai, and STSong) to reduce the
influence of visual similarity on neural representations
of the stimuli. The functional activity elicited by these
words forms the basis of all the analyses presented here.
Participants completed a semantic relatedness task in-
volving catch trials and 42 filler words interspersed be-
tween the seven critical stimuli to encourage them to
think about word meanings. Filler words were not used in
any of the fMRI analysis and therefore did not conform to
the criteria used to select critical stimuli. Del 42 filler
palabras (translation equivalents in both languages), half
were semantically related to one of the critical stimuli
(p.ej., axe–log) and half were semantically unrelated (p.ej.,
axe–moth) for a total of three related words and three
unrelated words for each critical stimulus. Because seman-
tic relatedness was only a distractor task and not a manip-
ulation in the experiment, the related and unrelated
conditions were based on the experimenters’ judgment,
and participants were not evaluated on their accuracy in
reproducing the same judgments.
Stimuli for this task were presented as black or red text
on a gray background via projector to a screen behind
the MRI scanner. Participants viewed the projected words
through a mirror attached to the scanner’s head coil.
Procedimiento
Experimental procedures were approved by the Dartmouth
Committee for the Protection of Human Subjects. Participe-
pants completed the semantic relatedness task while under-
going fMRI. Words were presented for 2000 mseg, seguido
by a 4000–6000 msec jittered fixation cross. If a catch-
word was presented in red text with a “?" (p.ej., “moth?",
also presented for 2000 mseg), participants responded by
indicating whether the catchword was semantically related
to the word immediately preceding. Catchwords were
always filler words and occurred in approximately one
third of trials to encourage participants to think about
the meanings of each stimulus word. Each functional
run was composed of 45–50 stimulus presentations, acerca de
7 min in duration. Participants completed seven functional
runs for a total of 35 presentations per critical stimulus
palabra.
Scanning Parameters
The study was performed using a Philips Intera Achieva
3-T scanner (Philips Medical Systems, Bothell, Washington) con
a SENSE (SENSEitivity Encoding) 32-channel head coil.
Anatomical images were collected using a high-resolution
3-D MPRAGE sequence (220 slices, 1 mm isotropic voxels,
campo de visión = 240 mm, acquisition matrix = 256 × 256).
Functional images were collected in seven runs using echo-
planar functional images sensitive to BOLD contrast (repe-
tition time = 2000 mseg, echo time = 35 mseg, flip angle =
90°, 3 mm in-plane resolution). During each of the func-
tional runs, 175 sets of axial images (35 slices/volume)
were collected in an interleaved fashion across the whole
cerebro.
Preprocessing and Estimation
Preprocessing and model estimation were all completed
within SPM8 (Penny, Friston, Ashburner, Kiebel, & Nichols,
2011). Functional images across seven runs were regis-
tered to the mean image, smoothed with a 5-mm FWHM
kernel for realignment, realigned, and resliced. Data were
then normalized to the ICBM space template and written
como 3 mm cubic voxels. A general linear model was esti-
mated with separate regressors for each of the seven crit-
ical stimuli and a regressor for the response type (catch
trial or none). Separate parameters were estimated for
each functional run and then averaged in contrasts defined
for each of the critical stimuli.
Multivoxel Pattern Analysis and Neural Similarity
Individual participants’ multivoxel patterns for the critical
stimuli were computed separately in 96 anatomical ROIs
(48 in each hemisphere), as defined by the Harvard-
Oxford Atlas (www.fmrib.ox.ac.uk/fsl/).
Cifra 1 illustrates the procedure for calculating neural
similarity in a single participant. A general linear model
is estimated in SPM for each participant with separate
parameters for each of the seven critical stimulus words.
The estimated multivoxel response pattern for each crit-
ical stimulus (a word) is defined by a contrast map (beta
weights) estimated in the participant level (first-level)
general linear model (Figura 1B). De este modo, for each critical
stimulus, we extract a 1 × n vector of beta values for all
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
1
2
1
8
/
1
1
7
1
4
/
9
1
1
7
9
4
5
9
1
/
9
1
6
0
7
8
oh
5
C
6
norte
0
_
2
a
/
_
j
0
oh
1
C
0
norte
0
0
_
a
pag
_
d
0
1
b
0
y
0
gramo
0
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
t
.
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
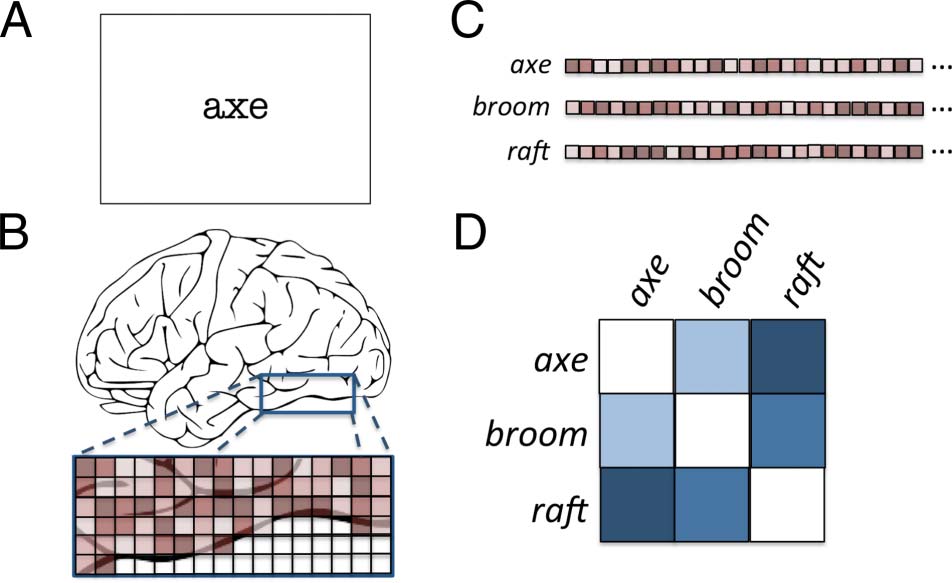
Cifra 1. Procedure for computing a neural similarity matrix. (A)
Stimulus is presented during functional imaging. (B) Individual voxel
responses to stimulus are measured or estimated. (C) Responses for
each stimulus are compared as n-dimensional vectors for n voxels.
(D) Stimulus representations are correlated to generate the neural
similarity matrix.
Zinszer et al.
1751
n voxels in the ROI (Figura 1C). An individual partici-
pant’s multivoxel patterns for the critical stimuli are
finally abstracted into similarity space. This step is critical
because it eliminates the need for matching up patterns
voxel-by-voxel across participants and instead allows
comparison of the similarity structure to any set of multi-
variate patterns for the same stimuli. The response pat-
terns to each of the seven critical stimuli are pairwise
correlacionado (Pearson’s r), resulting in a 7 × 7 neural sim-
ilarity matrix in which each stimulus is described by the
correlation of its functional response pattern to that of
the other six stimuli (Figure 1D). The correlation values
in each participant’s 7 × 7 similarity matrix are trans-
formed using Fisher’s r-to-z for normalizing correlation
coefficients, and the similarity matrices are averaged
across participants in each group, resulting in a single
7 × 7 group level matrix for English and another for
Chino.
Permutation-based Decoding
To achieve neural translation, a reference matrix (p.ej.,
the English group neural similarity structure) is com-
pared with every possible permutation of stimuli in the
test matrix (p.ej., the Chinese group neural similarity
estructura). If the neural similarity structures are similar
enough between two languages, the permutation of the
test matrix most highly correlated with the reference
matrix will be the correct set of translations. Statistical
significance for the permutation test is determined by ob-
serving all possible permutations for an empirical proba-
bility distribution. The 95th percentile of this distribution
represents a p value of .05. Note that it is important to
distinguish between permutation testing for nonpara-
metrically estimating statistical significance, a common
procedure that has also been used to assess the signifi-
cance of overall correlations between similarity matrices
(p.ej., Kriegeskorte, Mur, Fallar, et al., 2008), Opuesto a
our permutation-based decoding, which establishes
structural alignments between specific pairs of items by
picking as its alignment the permutation of labels that
produces the highest correlation between the permuted
and reference similarity matrices.
RESULTADOS
Participants were prompted to make semantic related-
ness judgments in approximately 30% of stimulus presen-
tations for seven target words and 42 additional filler
palabras. Mean participant response rate was 83% (DE =
20%) with a mean RT of 1398 mseg (DE = 167 mseg).
No measure of response accuracy was performed be-
cause the semantic relatedness judgments are subjective.
Cifra 1 illustrates the procedure for calculating par-
ticipants’ neural similarity matrices based on their unique
patterns of functional activity for word stimuli. Para el
seven word stimuli in each language (Mesa 1), semejanza
matrices were computed and transformed using Fisher’s
r-to-z (the inverse hyperbolic tangent) to normalize the
r distribution, and a group similarity matrix was com-
puted by averaging individual participants’ matrices for
each language. No voxel selection criteria were used in
this or any other analyses in this article, and thus in whole-
brain measures, the entire set of voxels from all parts of
all anatomical regions were used to produce the struc-
tural alignments.
The English and Chinese group neural similarity struc-
tures were also estimated in each anatomical ROI of a
standard and widely used brain atlas (the Harvard-Oxford
atlas, www.fmrib.ox.ac.uk/fsl/ ) and used to attempt neural
decoding of one language using the neural similarity
structures estimated for the other language. Por ejemplo,
the group similarity matrices for English and Chinese
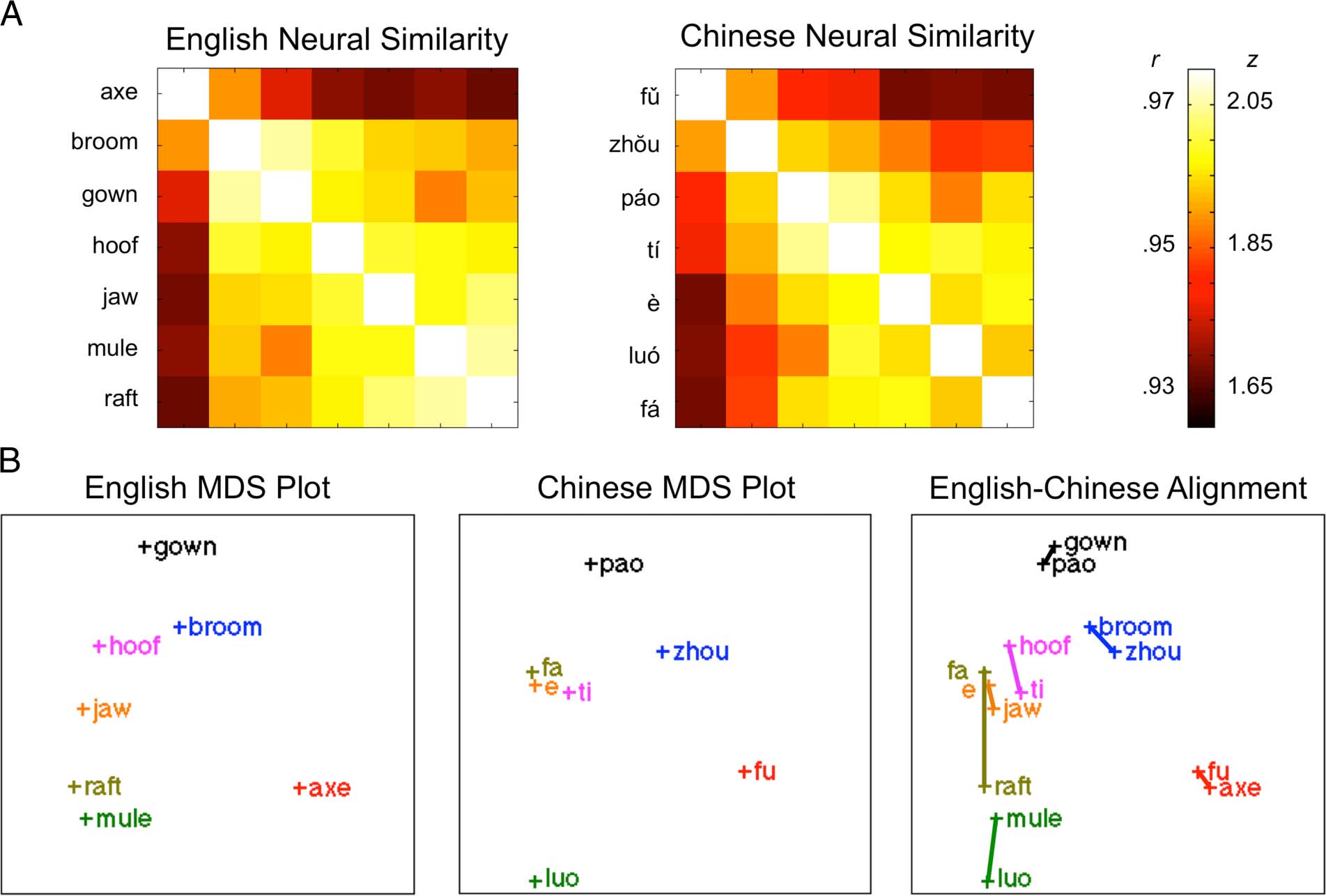
in left postcentral gyrus are illustrated in Figure 2A.
Between-group comparison of these structures provides
a neurally mediated form of translation wherein Chinese
words can be matched to English words based only on
their respective functional brain response patterns, a través de
the neural similarity structures for each language. Estos
seven-by-seven matrices thus provide the sole source of
information for alignment across languages and thus for
the translations.
To provide an intuitive visualization of the structural
alignments produced from these similarity matrices, nosotros
performed a classical multidimensional scaling of the sim-
ilarity matrices into 2-D projections. The z-transformed
correlations in the similarity matrices were transformed
into distance matrices using 1/z (where z corresponds
to the transformed correlation coefficient), and overall
structures were plotted for each language. Alignment of
these structures across languages provides some insight
into the translation relationships that would be predicted
based on the left postcentral gyrus. Sin embargo, the MDS
projections depict only the first two dimensions of these
multidimensional data. Correlations between the dis-
tance matrices and their 2-D projections were strong,
indicating that the MDS projections visualized for illus-
tration in Figure 2 account for a large amount of the vari-
ance in the neural representations (Chino: R2 = .66;
Inglés: R2 = .81). Además, the distances in the 2-D
projections were highly correlated between languages,
giving an impression of how well the structures aligned
(R2 = .82) in the third panel of Figure 2B. Fundamentalmente, cómo-
alguna vez, none of these diagnostics nor the MDS solutions as a
whole were used to evaluate translation accuracy. Ellos
are presented strictly as a visualization of the end-results
of the structural alignment analysis. Sin embargo, that struc-
tural alignment analysis itself was performed by permuting
el 7 × 7 similarity matrices, a process that occurred
before and independently of the subsequent 2-D MDS
visualization.
The threshold for statistical significance was computed
by taking the 95th percentile of the full distribution of
1752
Revista de neurociencia cognitiva
Volumen 28, Número 11
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
1
2
1
8
/
1
1
7
1
4
/
9
1
1
7
9
4
5
9
1
/
9
1
6
0
7
8
oh
5
C
6
norte
0
_
2
a
/
_
j
0
oh
1
C
0
norte
0
0
_
a
pag
_
d
0
1
b
0
y
0
gramo
0
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
.
F
t
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
1
2
1
8
/
1
1
7
1
4
/
9
1
1
7
9
4
5
9
1
/
9
1
6
0
7
8
oh
5
C
6
norte
0
_
2
a
/
_
j
0
oh
1
C
0
norte
0
0
_
a
pag
_
d
0
1
b
0
y
0
gramo
0
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
t
.
/
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Cifra 2. (A) Neural similarity matrices (r-to-z transformed) based on all voxels in left postcentral gyrus for each language group. (B) Conceptual
structures represented as MDS projections of neural similarity structures, based on first two dimensions.
accuracy scores for all possible permutations of words in
the test structure. The 95th percentile of the accuracy
distribution for all permutations was 3 de 7 correct trans-
laciones. De este modo, scores above this threshold have less than a
.05 probability of occurring by random selection (Raizada
& Connolly, 2012). Bonferroni correction for multiple
comparisons (96 ROI) results in a significance threshold
de 5 de 7 correct translations. Figure 2B illustrates a strong
visual correspondence between the Chinese and English
similarity structures in left postcentral gyrus.
All ROIs that achieved five or seven correct translations
are outlined in Table 2. En este caso, six correct trans-
lations are impossible because one translation error
would entail swapping two words, resulting in only five
correct translations. Switching the reference and test
matrices yields identical results. Six regions produced 7
de 7 translations, and an additional 11 ROI (listed in
Mesa 2) correctly translated five of the seven word stimuli
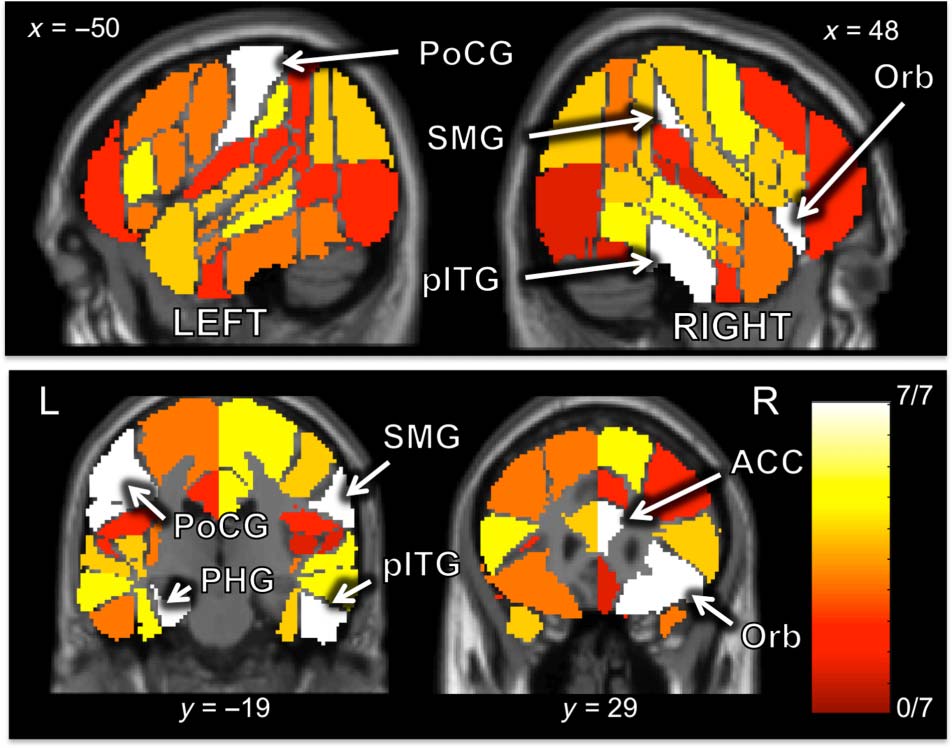
(the corrected threshold value for significance). Cifra 3
(visualized using the xjView toolbox available at alive-
learn.net/xjview) depicts a sample of the cortical regions
tested and highlights all six ROIs with 7 de 7 translations.
Regions with successful neural decoding were largely
located in temporal and parietal regions, although most
regions were not bilaterally equal in decoding accuracy.
Several frontal regions were also included in the set of
successfully decoded ROIs. The majority of regions with
7/7 translations (4 de 6 regiones) and slightly narrower
majority for all regions with 5 de 7 or more translations
(11 de 17 regiones) were right hemisphere regions. El
whole-brain similarity structures yielded 5 de 7 translations.
In one final analysis, we applied a jackknife resampling
procedure to test the reliability of the decoding results.
We excluded participants one at a time from the sample
de todo 22 participants and repeated the between-group
decoding. In the whole-brain data, resampling did not sig-
nificantly change decoding accuracy (mean accuracy =
.73, t(21) = 1.00, pag = .38). For each of the 17 significativo
ROI, we compared the set of 22 resampled means to the
original significance threshold of .71 (5 de 7 translations)
using a single-sample t test. Only the temporooccipital
part of right inferior temporal gyrus (Harv.-Oxf. #15) fell
significantly below this threshold after correction for
multiple comparisons. The mean and standard deviations
for the resampled means are reported in Table 2.
DISCUSIÓN
We have demonstrated that representational similarity
structures for word-elicited concepts are conserved
across speakers of different languages. These similarity
structures are effective for comparing functional brain
Zinszer et al.
1753
Mesa 2. Regions with Significant Decoding Accuracy
Harvard-Oxford ROI
Anatomical ROI
Resampled Mean (Dakota del Sur)
Regions with 7 de 7 Correct Translations (Accuracy = 1.0)
0.82 (0.22)
0.90 (0.14)
0.84 (0.19)
0.75 (0.29)
0.82 (0.25)
0.96 (0.10)
0.68 (0.15)
0.66 (0.21)
0.66 (0.10)
14
16
18
28
32
33
R inferior temporal gyrus, posterior division
L postcentral gyrus
R supramarginal gyrus, anterior division
R cingulate gyrus, anterior division
R frontal orbital cortex
L parahippocampal gyrus, anterior division
Regions with 5 de 7 Correct Translations (Accuracy = 0.71)
02
04
06
09
11
11
15
17
18
29
37
R superior frontal gyrus
L inferior frontal gyrus, pars triangularis
R precentral gyrus
R superior temporal gyrus, posterior division
0.75 (0.11)
R middle temporal gyrus, posterior division
L middle temporal gyrus, posterior division
0.69 (0.05)
0.73 (0.06)
R inferior temporal gyrus, temporooccipital part
0.62 (0.11)
R superior parietal lobule
L supramarginal gyrus, anterior division
R cingulate gyrus, posterior division
0.69 (0.08)
0.69 (0.14)
0.71 (0.14)
L temporal fusiform cortex, posterior division
0.77 (0.11)
ROIs are numbered according to the Harvard-Oxford anatomical atlas. Translation accuracy was also
resampled using a jackknife ( leave-one-participant-out) procedimiento. Mean and SD of resampled
accuracy scores are reported.
activity across speakers of different languages and enable
cross-language decoding, eso es, neural translation, a través de
structural alignment. We thereby provide strong neuro-
cognitive evidence for an intuitive (but heretofore untested)
afirmar: Although the words we use differ in visual and
auditory form across languages, we conceptualize the
meanings of these words similarly, such that they can be
reliably translated across brains.
This study is distinct from all previous work investigat-
ing the neural decoding of language because we translate
semantic representations between independent groups
for each language. Although previous studies have in-
vestigated neural decoding across languages (Correia,
Jansma, Hausfeld, Kikkert, & Bonte, 2015; Correia
et al., 2014; Buchweitz, Shinkareva, Mason, mitchell, &
Just, 2012), these studies have sidestepped one or more
aspects of the translation challenge by using within-
subject designs, which only allow comparisons within a
single brain. In a similar vein, cross-modal (but within-
idioma) comparison has found significant similarity
for orthographic and auditory elicitations of word mean-
ings (p.ej., Akama, Murphy, Na, Shimizu, & Poesio, 2012)
but also remains limited to within-subject decoding and
Cifra 3. Decoding accuracy projected onto the cortical surface. Ver
Mesa 2 for list of all ROIs with accuracy significantly greater than chance
nivel. ROIs providing 7 de 7 correct translations are labeled as follows:
ACC = anterior cingulate cortex; Orb = frontal orbital cortex;
PHG = parahippocampal gyrus; pITG = posterior inferior temporal
gyrus; PoCG = postcentral gyrus; SMG = supramarginal gyrus.
1754
Revista de neurociencia cognitiva
Volumen 28, Número 11
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
1
2
1
8
/
1
1
7
1
4
/
9
1
1
7
9
4
5
9
1
/
9
1
6
0
7
8
oh
5
C
6
norte
0
_
2
a
/
_
j
0
oh
1
C
0
norte
0
0
_
a
pag
_
d
0
1
b
0
y
0
gramo
0
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
t
.
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
thus makes no generalization about regularities in repre-
sentation across people. One notable exception, Honey,
Thompson, Lerner, and Hasson’s (2012) translation of
narratives between different groups of Russian and
English speaking participants examined only global
semantic processing without addressing word meaning.
The current work is distinct from this previous work be-
cause we translate meaning of individual word-elicited
concepts between independent groups for each language.
Within-subject studies are interesting but reveal only
associative matches between words in each language
(like learning synonyms for a word) or idiosyncratic se-
mantic relationships that are conserved only within an
individual’s brain. Here we use a between-subject design
to demonstrate the generalizability of structural align-
ment across brains, allowing the identification of com-
mon conceptual spaces across speakers and languages.
Our data expand upon dominant models of bilingual
lexical representation and access, which maintain that
bilinguals draw on a single, shared conceptual store for
building lexical semantic connections (Kroll, van Hell,
Tokowicz, & Verde, 2010; Van Hell & De Groot, 1998;
Kroll & Stewart, 1994; De Groot, 1992) by suggesting
that these shared concepts are broadly preserved across
entire groups of speakers of different languages.
The neuroanatomical regions that produce the highest
decoding performance achieve successful translation
because they encode the underlying conceptual struc-
tures evoked by word stimuli while all other aspects of
the word stimuli (p.ej., visual, orthographic, phonetic) dif-
fer between Chinese and English. Previous research in
semantic representation and processing has produced a
broad-reaching network of anatomical regions that inte-
grate perceptual and functional information into con-
cepts and decode auditory or visual word forms to
activate those concepts in language comprehension.
Many of the regions that produced significant cross-
language decoding in this study correspond to areas
widely recognized for the integration of multimodal se-
mantic information, either in traditional semantic pro-
cessing studies or in previous within-language neural
decoding analyses. Here we review these regions and
compare our findings with the existing semantic litera-
tura. We also found a handful of regions that are not typ-
ically reported for involvement in semantic processing,
and we explore these findings in a later section.
Regions Classically Associated with
Semantic Representation
The left fusiform gyrus has been repeatedly implicated in
previous neural decoding studies of semantic represen-
tation (anderson, Bruni, Lopopolo, Poesio, & Baroni,
2015; Chen, Garcea, & Mahon, 2015; Fernandino et al.,
2015; Raizada & Connolly, 2012; Mitchell et al., 2008;
Haxby et al., 2001) and translated five of seven words
accurately in this study. Recent studies have also sug-
gested that the parahippocampal gyri may be a multi-
modal hub for the conjunction of multiple sensory
modalities (Fernandino et al., 2015), which is consistent
with its role in functional representations instead of spe-
cific sensory modalities. The parahippocampal gyri have
also been implicated in specific domains of concept rep-
resentimiento: better for shelter- than tool-specific concepts
(Just, Cherkassky, Aryal, & mitchell, 2010) but also seem
to represent functional information about tools (versus
action/motor information; Chen et al., 2015).
The lateral and ventral temporal cortices are also widely
recognized for integrating multimodal information in word
representación (see reviews by Poeppel, Emmorey, Hickok,
& Pylkkänen, 2012; Precio, 2012; Binder, Desai, Tumbas, &
Conant, 2009). A series of regions proceeding from the
ventral temporooccipital cortex toward the anterior tem-
poral lobe support visual wordform processing, as in this
study’s task. Price’s (2012) account describes the most
posterior regions (the temporooccipital cortex) as per-
forming visual feature extraction whereas more anterior
regiones (the posterior inferior temporal cortex) access
lexical semantics of the identified words. Sin embargo, nosotros
have found that a large segment of posterior regions (ambos
temporooccipital and posterior portions of the inferior
temporal gyrus) in right hemisphere encode sufficient
semantic information to allow cross-language decoding.
This observation is inconsistent with a strict role of visual
feature extraction for the temporooccipital region because
the languages should provide no orthographic or visual
cues to translation. Findings in the left and right middle
temporal gyri are more consistent with conventional
semantic processing models, wherein the middle tempo-
ral gyrus provides cross-modal semantic representation
(Fairhall & Caramazza, 2013) and shows only weak left
lateralization (Poeppel et al., 2012). Finalmente, the left pos-
terior superior temporal gyrus has long been included in
the anatomical definition of Wernicke’s area and asso-
ciated with word comprehension (Poeppel et al., 2012;
although with some debate as to its involvement in audi-
tory processing, auditory word recognition, and seman-
tic access; Precio, 2012; Binder et al., 2009). En este caso,
we find that translation is successful only in the right
superior temporal gyrus (STG). Bilateral involvement
of posterior STG is generally restricted to auditory pro-
cessing functions in the aforementioned reviews, leaving
open the possibility that the visual word forms are elicit-
ing in STG some semantic representation adequate for
achieving translation (perhaps mediated by auditory word
form retrieval).
Several parietal regions identified in this study have
been emerging in recent literature as playing important
roles in integration of semantic representations across
sensory modalities. Recent studies in neural decoding
have converged upon the supramarginal gyrus as a multi-
modal (or transmodal) integrator of sensory information
(Fernandino et al., 2015). Previous cross-language studies
have found that this region produced stable responses
Zinszer et al.
1755
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
1
2
1
8
/
1
1
7
1
4
/
9
1
1
7
9
4
5
9
1
/
9
1
6
0
7
8
oh
5
C
6
norte
0
_
2
a
/
_
j
0
oh
1
C
0
norte
0
0
_
a
pag
_
d
0
1
b
0
y
0
gramo
0
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
F
t
.
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
across languages in bilinguals (Buchweitz et al., 2012)
and were correlated in Russian and English speakers’
semantic processing (Honey et al., 2012), although these
effects were left lateralized. en este estudio, the bilateral
supramarginal gyrus reached or exceeded the signifi-
cance criterion (5 de 7 translations).
The bilateral (left-dominant) activation of posterior
cingulate gyrus was associated with semantic tasks in
Binder et al.’s (2009) meta-analysis. On the basis of its
involvement in visual, spatial, and emotional processes
in other tasks, Binder and colleagues suggest that the
posterior cingulate interacts with the hippocampus to
process episodic memory and thus is highly correlated
with semantic processing tasks that involve retrieval of
episodic information. This claim is consistent with our
finding that the right posterior cingulate produced accu-
rate translations. Our results also identified the right
anterior cingulate for high decoding accuracy. The ante-
rior cingulate has more typically been described as pro-
viding conflict monitoring for cognitive control (Shenhav,
Straccia, cohen, & Botvinick, 2014; Botvinick, cohen, &
Carretero, 2004), including in the case language conflict in
bilingualism (Verde & Abutalebi, 2013; Abutalebi &
Verde, 2007). The anterior cingulate cortices have not
often been the focus on concept representation research,
but we see some evidence of this modal specificity in that
it is selectively responsive to information about object
forma, while the posterior cingulate decodes across mul-
tiple modalities (Fernandino et al., 2015).
Recent and Novel Findings in
Semantic Representation
Several other anatomical regions where we observed
significant cross-language decoding are not typically
included in neural accounts of semantic representation.
According to the meta-analysis of Binder et al. (2009),
pre- and postcentral gyri and the adjacent superior pari-
etal lobule rarely appear in contrast-based neuroimaging
analiza, perhaps suggesting a lack of overall change in
activation levels across tasks. This absence of significant
spatial contrasts does not rule out the possibility that
multivoxel response patterns still encode important se-
mantic information, and we find that these regions encode
accurate translation information. According to previous
decoding studies, the postcentral gyrus is, En realidad, tuned
for identifying tools (Just et al., 2010), suggesting that it
may represent information related to manipulation of
objects. Relacionado, a region spanning the left postcentral
gyrus and superior parietal lobule are selectively respon-
sive to shape information, and the right precentral gyrus
to manipulation information (Fernandino et al., 2015).
Both of these factors would be highly relevant to the
classification of concrete objects, such as those described
by the seven nouns in our study.
The left inferior frontal gyrus has been widely impli-
cated in language processing tasks, but Binder et al.
(2009) suggested that that only the pars orbitalis of the
inferior frontal gyrus is directly involved in semantic pro-
cessing and that any detected pars triangularis involvement
is attributable to phonological or working memory aspects
of the task. Our findings suggest that the pars triangularis
may also contain semantic information. Previous research
in picture naming has linked pars triangularis with lexical
selection when objects have low name agreement (Kan &
Thompson-Schill, 2004), and this selection may involve
more semantically informed representations than revealed
by contrast analyses, which focus on magnitude of activation.
Left dorsomedial and ventromedial pFC are frequent
correlates of semantic processing, but their direct role
in representation is not clear. According to Binder and
colleagues’ (2009) cuenta, the dorsal and medial sur-
faces of the left superior frontal gyrus may have a role
in retrieval processes, although these effects are primarily
left-lateralized, while our findings were restricted to the
region’s right homologue. Precio (2012) proposes that the
left superior frontal gyrus is principally involved in various
constraints on semantic meaning, either by syntax or by
contexto. The frontal orbital cortex (in the wider ventro-
medial pFC) appears to be associated with affective pro-
cessing and thus likely underlies affective information in
semantic processing (Binder et al., 2009). Poeppel et al.
(2012) more generally attribute ventromedial pFC as
another semantic combinatory region, tasked with repre-
senting syntactically covert meaning. These proposals do
not appear to be mutually exclusive and could be consis-
tent with our current cross-language findings.
A right lateralization effect seems to be suggested by
the relative prevalence of right hemisphere regions
among significant ROIs, but this observation may be ex-
plained in light of the jackknife resampling results. El
stability information provided by the resampling analysis
supports a more balanced account: Although the left
hemisphere regions from the 7 de 7 translation set were
highly stable (mean accuracy ≥ .90 in left parahippo-
campal and post central gyri), right hemisphere regions
with the same group level translation accuracy achieved
markedly lower resampled accuracy and higher variability
(larger standard deviations; ver tabla 2). De este modo, the rela-
tive importance of these regions should be interpreted
with the reliability of their performance across partici-
pants in mind.
Fundamentalmente, the limited number of words used in this
study may simply result in too stark a contrast between
regions with successful and unsuccessful translation per-
rendimiento. Por ejemplo, a region that achieves five of
seven accurate translations has a 50% chance of success-
fully translating the last pair of words (given that only two
permutations of the remaining two words exist). This fact
should limit the importance we assign to whether a re-
gion achieves five versus seven translations, and indeed
the apparent right bias decreases slightly when consider-
ing all significant regions together, as well as the appear-
ance of typical bilateral or left-lateralized regions for
1756
Revista de neurociencia cognitiva
Volumen 28, Número 11
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
1
2
1
8
/
1
1
7
1
4
/
9
1
1
7
9
4
5
9
1
/
9
1
6
0
7
8
oh
5
C
6
norte
0
_
2
a
/
_
j
0
oh
1
C
0
norte
0
0
_
a
pag
_
d
0
1
b
0
y
0
gramo
0
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
/
F
t
.
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
semantic integration (middle temporal gyrus, supra-
marginal gyrus, left fusiform cortex) in the broader 5+/7
set of regions.
Previous neural decoding studies have not specifically
investigated lateralization effects, but one cautious expla-
nation for a right hemisphere advantage might be drawn
from research on lateralization in semantic processing:
Semantic information in the right hemisphere has long
been hypothesized to represent coarser, message level
semantic representations (Beeman, 1993) and more re-
cently been associated with processing semantically dis-
tant or novel associations and semantic context (Vigneau
et al., 2011; Jung-Beeman, 2005) such as in metaphor
comprensión ( Vigneau et al., 2011; Schmidt, DeBuse,
& Seger, 2007). en este estudio, coarser representations
may offer better cross-language symmetry than fine-
grained language-specific or culturally specific information.
Particularly, because our word stimuli were composed of
only seven relatively distant concepts, the coarse repre-
sentations for these concepts could be more consistent
across languages than their left-lateralized, finer-grained
representaciones (such as the exact shape and appearance
of a prototypical broom or raft).
Future Directions
This study is the first to show that neural activation can
be used to structurally align a set of words in speakers of
one language with the translation-equivalent words in
speakers of a very different language. Clearly this prom-
ising result in a new area is just the beginning of a much
broader set of future investigations. en este estudio, nosotros
relied on group level averages of the neural similarity
structures to perform the alignments. This approach re-
duces interparticipant noise in the similarity structures,
but it also limits our ability to generalize about new
participants or new words.
One aspect that could obviously be extended is the
number of words being considered. Our study used only
seven words. Now that cross-language structural align-
ment has been shown to be achievable, the constraints
that led to that small set size could be loosened in future
trabajar. Por ejemplo, each word could be presented a
smaller number of times during the fMRI scanning
(thereby allowing more words to be used without length-
ening the overall scan), and our tight constraints on word
choice (single character in Chinese, monosyllabic in
Inglés, etc.) could also potentially be relaxed. Including
a greater number of words would provide more similarity
data for the representation of each word, and these
additional data could improve individual participant
level translation rather than relying on group-averaged
data to improve the signal-to-noise in the similarity matri-
ces. Además, we have evaluated the overall success
of the structural alignment, but with a greater number of
palabras, comparisons in translation accuracy could be
drawn between words. Translation may be more or less
successful for new sets of words based on their semantic
semejanza, translation equivalence, or other factors that
we simply cannot explore with the current, small stimu-
lus set. Claramente, more research is needed. This study
demonstrates that across-subject neural translation is
posible. Building upon that initial foundation, future
work can explore how well such translation can succeed
across larger numbers and varieties of words.
Además, although the permutation-based decoding
worked well in this study, it has certain computational
limits: It is computationally infeasible to perform the same
exhaustive search for even slightly larger lexicons, como el
number of permutations that must be compared expands
factorially (p.ej., 7 words have 5040 permutations, pero 10
words have over 3.6 million permutations). Sin embargo,
search optimization strategies offer a number of oppor-
tunities to refine the existing algorithm, which would
allow neural translation to scale up to much larger lexicons.
Our comparison of neural similarity structures in native
speakers of Chinese and English yielded a successful
translation between English and Chinese words based
on the functional brain responses of separate groups of
participants using each language. This ability to compare
brain representations of words between speakers of dif-
ferent languages presents a new way of studying transla-
tion asymmetry, such as between abstract nouns for
which experimental evidence indicates translation costs
due to ambiguity ( Van Hell & De Groot, 1998). Neurally
informed translation permits comparison of multiple
translation candidates for their relative fitness to brain
responses elicited by the other language. Además,
language-specific and language-independent elements
of brain representation can be contrasted by examining
translation pairs for correlation to nonlinguistic measures
(p.ej., visual object information) and linguistic measures
(p.ej., word co-occurrence) in a similar fashion to Anderson
et al. (2015).
Human cognition often draws on the ability to find
alignments between structures that differ in their surface
apariencia (such as the relationships between words in
Chinese and English) but share a deeper underlying
estructura (such as the relationships between concepts
underlying words). Structural alignments of this sort sup-
port a broad range of cognitive functions, including ana-
logical reasoning, scientific inference, and language
aprendiendo (Gentner & Herrero, 2012; Gentner & Namy, 1999;
Holyoak & Thagard, 1996). en este estudio, we showed that
such structural alignment of matching concepts could be
performed directly on people’s neural representations. Este
work may therefore open the door to the largely uncharted
domain within cognitive neuroscience of the structural
alignment of neural conceptual spaces.
Expresiones de gratitud
This work was supported in part by NSF award 1228261 (P.I.
Raizada).
Zinszer et al.
1757
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
1
2
1
8
/
1
1
7
1
4
/
9
1
1
7
9
4
5
9
1
/
9
1
6
0
7
8
oh
5
C
6
norte
0
_
2
a
/
_
j
0
oh
1
C
0
norte
0
0
_
a
pag
_
d
0
1
b
0
y
0
gramo
0
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
t
.
/
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Reprint requests should be sent to Benjamin D. Zinszer,
Rochester Center for Brain Imaging, Room 2B201, 430 Elmwood
Ave., Rochester, Nueva York 14627, o por correo electrónico: bzinszer@gmail.com.
representation reflects multimodal abstraction: A
framework for embodied semantics. Corteza cerebral,
26, 2018–2034.
REFERENCIAS
Abutalebi, J., & Verde, D. W.. (2007). Bilingual language
producción: The neurocognition of language representation
and control. Journal of Neurolinguistics, 20, 242–275.
Akama, h., Murphy, B., Na, l., Shimizu, y., & Poesio, METRO. (2012).
Decoding semantics across fMRI sessions with different
stimulus modalities: A practical MVPA study. Fronteras en
Neuroinformatics, 6.
anderson, A. J., Bruni, MI., Lopopolo, A., Poesio, METRO., & Baroni, METRO.
(2015). Reading visually embodied meaning from the brain:
Visually grounded computational models decode visual-object
mental imagery induced by written text. Neuroimagen, 120,
309–322.
Beeman, METRO. (1993). Semantic processing in the right
hemisphere may contribute to drawing inferences from
discourse. Brain and Language, 44, 80–120.
Gentner, D., & Namy, l. (1999). Comparison in the development
of categories. Cognitive Development, 14, 487–513.
Gentner, D., & Namy, l. l. (2004). The role of comparison in
children’s early word learning. In D. GRAMO. Sala & S. R. Waxman
(Editores.), Weaving a lexicon (páginas. 533–568). Cambridge, MAMÁ:
CON prensa.
Gentner, D., & Herrero, l. (2012). Analogical reasoning. In V. S.
Ramachandran (Ed.), Encyclopedia of Human Behavior
(2y ed., páginas. 130–136). Oxford, Reino Unido: Elsevier.
Goldstone, R. l., & Rogosky, B. j. (2002). Using relations within
conceptual systems to translate across conceptual systems.
Cognición, 84, 295–320.
Verde, D. w., & Abutalebi, j. (2013). Language control in
bilinguals: The adaptive control hypothesis. Diario de
Psicología cognitiva, 25, 515–530.
Haxby, j. v., Gobbini, METRO. I., Furey, METRO. l., Ishai, A., Schouten,
j. l., & Pietrini, PAG. (2001). Distributed and overlapping
representations of faces and objects in ventral temporal
corteza. Ciencia, 293, 2425–2430.
Binder, j. r., Desai, R. h., Tumbas, W.. w., & Conant, l. l. (2009).
Holyoak, k. J., & Thagard, PAG. (1996). Mental leaps: Analogy
Where is the semantic system? A critical review and
meta-analysis of 120 functional neuroimaging studies.
Corteza cerebral, 19, 2767–2796.
Botvinick, METRO. METRO., cohen, j. D., & Carretero, C. S. (2004). Conflicto
monitoring and anterior cingulate cortex: An update.
Tendencias en Ciencias Cognitivas, 8, 539–546.
Brysbaert, METRO., & Nuevo, B. (2009). Moving beyond Kucera and
Francisco: A critical evaluation of current word frequency
norms and the introduction of a new and improved word
frequency measure for American English. Behavior Research
Métodos, 41, 977–990.
Brysbaert, METRO., Warriner, A. B., & Kuperman, V. (2014).
Concreteness ratings for 40 thousand generally known
English word lemmas. Behavior Research Methods, 46,
904–911.
Buchweitz, A., Shinkareva, S. v., Mason, r., mitchell, t. METRO., & Just,
METRO. A. (2012). Identifying bilingual semantic neural representations
across languages. Brain and Language, 120, 282–289.
Cai, P., & Brysbaert, METRO. (2010). SUBTLEX-CH: Chinese word
and character frequencies based on film subtitles. Más uno,
5, e10729.
Chen, P., Garcea, F. MI., & Mahon, B. z. (2015). The representation
of object-directed action and function knowledge in the
cerebro humano. Corteza cerebral, 26, 1609–1618.
Correia, J., Formisano, MI., Valente, GRAMO., Hausfeld, l., Jansma, B.,
& Bonte, METRO. (2014). Brain-based translation: fMRI decoding of
spoken words in bilinguals reveals language-independent
semantic representations in anterior temporal lobe.
Revista de neurociencia, 34, 332–338.
Correia, j. METRO., Jansma, B., Hausfeld, l., Kikkert, S., & Bonte, METRO.
(2015). EEG decoding of spoken words in bilingual listeners:
From words to language invariant semantic-conceptual
representaciones. Fronteras en psicología, 6.
De Groot, A. METRO. B. (1992). Bilingual lexical representation: A
closer look at conceptual representations. En R. Frost & l.
katz (Editores.), Orthography, phonology, morfología, y
significado (páginas. 289–412). Ámsterdam: Elsevier.
Fairhall, S. l., & Caramazza, A. (2013). Brain regions that
represent amodal conceptual knowledge. Diario de
Neurociencia, 33, 10552–10558.
Falkenhainer, B., Forbus, k. D., & Gentner, D. (1989). El
structure-mapping engine: Algorithm and examples.
Artificial Intelligence, 41, 1–63.
Fernandino, l., Binder, j. r., Desai, R. h., Pendl, S. l.,
Humphries, C. J., Bruto, W.. l., et al. (2015). Concept
in creative thought. Cambridge, MAMÁ: CON prensa.
Honey, C. J., Thompson, C. r., Lerner, y., & Hasson, Ud..
(2012). Not lost in translation: Neural responses
shared across languages. Revista de neurociencia, 32,
15277–15283.
Jung-Beeman, METRO. (2005). Bilateral brain processes for
comprehending natural language. Tendencias en Cognitivo
Ciencias, 9, 512–518.
Just, METRO. A., Cherkassky, V. l., Aryal, S., & mitchell, t. METRO.
(2010). A neurosemantic theory of concrete noun
representation based on the underlying brain codes. PLoS
Uno, 5, e8622.
Kan, I. PAG., & Thompson-Schill, S. l. (2004). Effect of name
agreement on prefrontal activity during overt and covert
picture naming. Cognitivo, Afectivo & conductual
Neurociencia, 4, 43–57.
Kriegeskorte, NORTE., Mur, METRO., & Bandettini, PAG. (2008).
Representational similarity analysis—Connecting the
branches of systems neuroscience. Frontiers in Systems
Neurociencia, 2.
Kriegeskorte, NORTE., Mur, METRO., Fallar, D. A., Kiani, r., Bodurka, J.,
Esteky, h., et al. (2008). Matching categorical object
representations in inferior temporal cortex of man and
monkey. Neurona, 60, 1126–1141.
Kroll, j. F., & Stewart, mi. (1994). Category interference in
translation and picture naming: Evidence for asymmetric
connections between bilingual memory representations.
Journal of Memory and Language, 33, 149–174.
Kroll, j. F., van Hell, j. GRAMO., Tokowicz, NORTE., & Verde, D. W.. (2010).
The revised hierarchical model: A critical review and
evaluación. Bilingualism, 13, 373–381.
Laakso, A., & Cottrell, GRAMO. (2000). Content and cluster analysis:
Assessing representational similarity in neural systems.
Philosophical Psychology, 13, 47–76.
mitchell, t. METRO., Shinkareva, S. v., Carlson, A., Chang, K.-M.,
Malave, V. l., Mason, R. A., et al. (2008). Predicting human
brain activity associated with the meanings of nouns. Ciencia,
320, 1191–1195.
Penny, W.. D., Friston, k. J., Ashburner, j. T., Kiebel, S. J., &
Nichols, t. mi. (Editores.) (2011). Statistical parametric mapping:
The analysis of functional brain images. Burlington, MAMÁ:
Prensa académica.
Poeppel, D., Emmorey, K., Hickok, GRAMO., & Pylkkänen, l. (2012).
Towards a new neurobiology of language. Diario de
Neurociencia, 32, 14125–14131.
1758
Revista de neurociencia cognitiva
Volumen 28, Número 11
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
1
2
1
8
/
1
1
7
1
4
/
9
1
1
7
9
4
5
9
1
/
9
1
6
0
7
8
oh
5
C
6
norte
0
_
2
a
/
_
j
0
oh
1
C
0
norte
0
0
_
a
pag
_
d
0
1
b
0
y
0
gramo
0
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
.
F
t
/
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Precio, C. j. (2012). A review and synthesis of the first 20 años
of PET and fMRI studies of heard speech, spoken language
and reading. Neuroimagen, 62, 816–847.
Raizada, r., & Connolly, A. (2012). What makes different
people’s representations alike: Neural similarity space solves
the problem of across-subject fMRI decoding. Diario de
Neurociencia Cognitiva, 24, 868–877.
Schmidt, GRAMO. l., DeBuse, C. J., & Seger, C. A. (2007). Right
hemisphere metaphor processing? Characterizing the
lateralization of semantic processes. Brain and Language,
100, 127–141.
Shenhav, A., Straccia, METRO. A., cohen, j. D., & Botvinick, METRO. METRO.
(2014). Anterior cingulate engagement in a foraging context
reflects choice difficulty, not foraging value. Naturaleza
Neurociencia, 17, 1249–1254.
Turney, PAG. (2008). The latent relation mapping engine:
Algorithm and experiments. Journal of Artificial Intelligence
Investigación, 33, 615–655.
Van Hell, j. GRAMO., & De Groot, A. METRO. B. (1998). Conceptual
representation in bilingual memory: Effects of concreteness
and cognate status in word association. Bilingualism:
Language and Cognition, 1, 193–211.
Vigneau, METRO., Beaucousin, v., Hervé, PAG. y., Jobard, GRAMO., Petit, l.,
Crivello, F., et al. (2011). What is right-hemisphere contribution
to phonological, lexico-semantic, and sentence processing?
Insights from a meta-analysis. Neuroimagen, 54, 577–593.
D
oh
w
norte
yo
oh
a
d
mi
d
F
r
oh
metro
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
h
t
t
pag
:
/
D
/
oh
metro
w
i
norte
t
oh
pag
a
r
d
C
mi
.
d
s
F
i
r
oh
yo
metro
v
mi
h
r
C
pag
h
a
d
i
i
r
r
mi
.
C
C
t
.
oh
metro
metro
/
j
mi
oh
d
tu
C
norte
oh
/
C
a
norte
r
a
t
r
i
t
i
C
C
yo
mi
mi
–
pag
–
d
pag
d
2
F
8
/
1
2
1
8
/
1
1
7
1
4
/
9
1
1
7
9
4
5
9
1
/
9
1
6
0
7
8
oh
5
C
6
norte
0
_
2
a
/
_
j
0
oh
1
C
0
norte
0
0
_
a
pag
_
d
0
1
b
0
y
0
gramo
0
tu
.
mi
pag
s
t
d
oh
F
norte
b
0
y
8
S
METRO
mi
I
pag
t
mi
metro
l
i
b
b
mi
r
r
a
2
r
0
2
i
3
mi
s
/
j
.
t
/
F
tu
s
mi
r
oh
norte
1
7
METRO
a
y
2
0
2
1
Zinszer et al.
1759