Seeing and Hearing Meaning: ERP and f MRI Evidence
of Word versus Picture Integration into
a Sentence Context
Roel M. Willems1, Aslı O¨ zyu¨rek1,2, and Peter Hagoort1,2
Abstracto
& Understanding language always occurs within a situational
context and, por lo tanto, often implies combining streams of infor-
mation from different domains and modalities. One such combi-
nation is that of spoken language and visual information, cual
are perceived together in a variety of ways during everyday com-
munication. Here we investigate whether and how words and
pictures differ in terms of their neural correlates when they are
integrated into a previously built-up sentence context. Esto es
assessed in two experiments looking at the time course (measur-
ing event-related potentials, ERPs) and the locus (using func-
tional magnetic resonance imaging, resonancia magnética funcional) of this integration
proceso. We manipulated the ease of semantic integration of
word and/or picture to a previous sentence context to increase
the semantic load of processing. In the ERP study, an increased
semantic load led to an N400 effect which was similar for pictures
and words in terms of latency and amplitude. In the fMRI study,
we found overlapping activations to both picture and word in-
tegration in the left inferior frontal cortex. Specific activations for
the integration of a word were observed in the left superior
temporal cortex. We conclude that despite obvious differences in
representational format, semantic information coming from pic-
tures and words is integrated into a sentence context in similar
ways in the brain. This study adds to the growing insight that the
language system incorporates (semantic) information coming
from linguistic and extralinguistic domains with the same neural
time course and by recruitment of overlapping brain areas. &
INTRODUCCIÓN
Understanding language always occurs within a situa-
tional context, such as knowledge about the person you
are talking to or the location one is in (clark, 1996).
Por lo tanto, understanding language often implies com-
bining streams of information from different modalities.
Por ejemplo, consider a biology teacher describing the
properties of an animal while at the same time show-
ing a slide with a picture of the animal. In such a case,
information do not bear a direct
auditory and visual
physical connection. Eso es, the string of sounds de-
scribing a concept never directly maps onto the visual
appearance of this concept. This raises the question how
verbal (linguistic) and visual (extralinguistic) semantic
information combine to form a coherent interpretation
of a speaker’s message in relation to the overall context.
The current study investigates this question by assessing
integration of semantic information from
the neural
words and pictures embedded within a spoken sentence
contexto. Our main aim was to investigate whether,
despite differences in representational format, semantic
information from pictures and words is integrated into
an overall representation of an utterance in the same
1Radboud University Nijmegen, Nimega, Los países bajos, 2máx.
Planck Institute for Psycholinguistics, Nimega, Los países bajos
way as unimodal semantic information from a word is.
Within the broader context of language comprehension,
we wanted to investigate differences and commonalities
between linguistic and extralinguistic information pro-
cessing during sentence comprehension.
We investigated both the neural time course (measuring
event-related potentials, ERPs) and the locus (using func-
tional magnetic resonance imaging, resonancia magnética funcional) of this multi-
modal
integration process. Combining temporal and
spatial neural information in this way provides a more
complete view of the integration process under study.
Lexical Semantic Information in
a Sentence Context
The seminal ERP studies by Kutas and Hillyard (1980,
1984) showed that words that are semantically anoma-
lous in relation to the preceding sentence context lead
to a more negative deflection in the ERP waveform than
words that are semantically congruent. Por ejemplo, el
sentence-final word of the sentence, ‘‘She spread her
bread with socks,’’ leads to a negative deflection in the
ERP waveform in comparison to the ERP for a congruous
ending as in ‘‘She spread her bread with butter.’’ This
N400 effect occurs between 250 y 500 msec after the
anomalous word, and is usually maximal at central–
posterior electrode sites. N400 effects are also observed
D 2008 Instituto de Tecnología de Massachusetts
Revista de neurociencia cognitiva 20:7, páginas. 1235–1249
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
0
yo
7
mi
–
1
pag
2
d
3
F
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
oh
3
C
5
norte
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
8
mi
.
s
t
2
oh
0
norte
0
8
0
5
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
/
t
.
F
.
oh
norte
1
8
METRO
a
y
2
0
2
1
when a critical word is a possible but unexpected con-
tinuation of a sentence, without being a semantic viola-
ción (Hagoort & Marrón, 1994; Kutas & Hillyard, 1984).
The N400 has become an established ERP component
which is thought to reflect the ease of integration of a
word into a preceding context (see Brown, Hagoort, &
Kutas, 2000; Kutas & Van Petten, 1994 for reviews).
fMRI studies of sentences with semantic anomalies
comparable to those used in N400 studies have reported
increased activation in left inferior frontal and/or left
temporal areas ( Willems, O¨ zyu¨rek, & Hagoort, 2007;
Hagoort, Hald, Bastiaansen, & Petersson, 2004; Friederici,
Ruschemeyer, Hahne, & Fiebach, 2003; Kuperberg et al.,
2000, 2003; Baumgaertner, Weiller, & Buchel, 2002; En
et al., 2000). Based on these findings, it has been claimed
that these areas are involved in semantic integration
because they respond to a higher load of integration
elicited by the difficulty of semantic processing. Reciente
work has indeed shown that increased semantic ambi-
guity without semantic anomalies also leads to increased
activations in left inferior frontal and left temporal regions
(Rodd, davis, & Johnsrude, 2005; Davis et al., 2007).
Extralinguistic Information in
a Sentence Context
The integration of extralinguistic information into a
preceding context has been explored in a variety of ways
in ERP or fMRI studies (p.ej., Hagoort et al., 2004;
Koelsch et al., 2004; Van Petten & Rheinfelder, 1995).
A complete review of how extralinguistic information
inf luences sentence comprehension is beyond the
scope of this article. We therefore restrict our focus on
studies investigating the integration of visual informa-
tion into a preceding (oración) contexto.
Few studies have looked into the integration of
picture information into a sentence context. In an ERP
estudiar, Ganis, Kutas, and Sereno (1996) presented sen-
tences that either ended with a word or a picture that
could be anomalous or not. Similar N400 effects were
found to anomalous words and pictures. Sin embargo, el
scalp distribution for the anomalous pictures was more
frontal than for the anomalous words. Nigam, Hoffman,
and Simons (1992) also found similar N400 effects for
pictures and words, but did not find a difference in scalp
distribución. Sin embargo, this might be due to the limited
number of electrodes that they recorded from, cual
did not cover the frontal part of the brain. Federmeier
and Kutas (2001) found a correlation between the am-
plitude of the N400 effect and the semantic fit of a
picture with respect to the preceding part of a sentence.
De nuevo, there was a frontal scalp distribution for the ef-
efectos. Además, they observed an N300 effect to the
anomalous pictures. Some other ERP studies have inves-
tigated the processing of visual information following a
visual context instead of a language context. West and
Holcomb (2002), por ejemplo, presented a series of pic-
tures forming a simple story. The last picture was either
a congruous or an incongruous ending of the story.
Incongruous pictures elicited increased N300 and N400
efectos, with a maximal distribution over centro-frontal
electrodes. Sitnikova, Kuperberg, and Holcomb (2003)
had congruous or incongruous objects appear in video
clips of real-world events. They observed an N400 effect
for the incongruous objects with a fronto-central maxi-
mum in the scalp distribution. Finalmente, Ganis and Kutas
(2003) had congruent or incongruent objects appear in
still images of real-world events. An increased negativity
strongly resembling the N400 was observed for the in-
congruous as compared to the congruous objects.
Several priming studies report similar findings when
pictures and words are presented outside of a sentence
contexto. Eso es, N300 and N400 effects are reported to
incongruous picture pairs, with a more frontal scalp
distribution than is normally seen for word–word prim-
ing studies (McPherson & Holcomb, 1999; Holcomb &
McPherson, 1994; Barrett & Rugg, 1990).
En resumen, ERP studies manipulating the semantic fit
of pictures in relation to a (oración) context report
similar N400 amplitudes and onset latencies as found for
integration of semantic information conveyed through
a word. Differences are reported, sin embargo, in scalp dis-
tribution which is more frontal for pictures than for
palabras, and in the finding of an earlier separate negativ-
idad, the N300. The latter component has been suggested
to reflect the degree of effort needed to integrate an
object-specific/imagistic representation into a preceding
contexto (p.ej., McPherson & Holcomb, 1999).
From neuroimaging studies, little is known about the
neural localization of sentence-level processing of visual
extralinguistic information. In an earlier study, nosotros
looked at how meaningful co-speech gestures compare
to spoken words when anomalous within a sentence
contexto (Willems et al., 2007). Overlap between lexical
violations and gesture violations was found in the left
inferior frontal cortex. Relevant to the present study is
the considerable literature on the neural correlates of
the semantic representation of visually presented ob-
jects. Such studies mostly find that the ventral temporal
cortex is activated to the perception of a large variety
of objects (Martín & chao, 2001; Schacter & Buckner,
1998). More important for the present study is that many
of these studies also report the inferior frontal cortex to
be sensitive to the repeated presentation of an object
(Martín & chao, 2001; Schacter & Buckner, 1998) or of a
word and an object (p.ej., Lebreton, Desgranges, Landeau,
Baron, & Eustache, 2001). A commonly held view derived
from these and other studies is that ventral temporal
activation is related to semantic knowledge of an ob-
ject, whereas inferior frontal activation is related to
processes of semantic selection or retrieval (Martín &
chao, 2001; Wagner, Pare-Blagoev, clark, & Poldrack,
2001; Thompson-Schill, D'Esposito, Aguirre, & Farah, 1997;
Wagner, Desmond, Demb, guantero, & Gabrieli, 1997). Nosotros
1236
Revista de neurociencia cognitiva
Volumen 20, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
0
yo
7
mi
–
1
pag
2
d
3
F
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
oh
3
C
5
norte
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
8
mi
.
s
t
2
oh
0
norte
0
8
0
5
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
t
.
.
F
/
oh
norte
1
8
METRO
a
y
2
0
2
1
would like to point out that although related to our study,
the priming studies of objects differ in important aspects
from the present study. In the present paradigm, hay
a relatively rich linguistic context to which picture or word
can be integrated. In the repeated presentation of exem-
plars of object categories, this is arguably not the case.
To our knowledge, the present study is the first to in-
vestigate the integration of pictures at the sentence level,
using both fMRI and ERPs.
The Present Study
Within the study of the cognition of language, the issue
of how linguistic and extralinguistic information are inte-
grated into a sentence context is reflected in the distinc-
tion between one-step and two-step models of language
comprensión. The implication of two-step models is
eso, primero, the meaning of a sentence is computed and
segundo, the sentence meaning is integrated with extralin-
guistic information such as information about the speak-
er’s identity (p.ej., Lattner & Friederici, 2003; Cutler &
Clifton, 1999). This position is a consequence of Fregean
composicionalidad, which states that the meaning of an
utterance is a function of the meaning of its parts and of
the syntactic rules by which these parts are combined
(see Culicover & Jackendoff, 2006). Because the domain
of syntactic rules is the sentence, the implication of this
idea is that language interpretation takes place in a two-
step fashion. It is important for the present study that the
two-step model at least implies that linguistic computa-
tion should precede the integration of nonlinguistic in-
formation in time (see Hagoort & van Berkum, 2007 para
further discussion). Adherents of a one-step model, en
contrast, take as their starting point the ‘‘immediacy
assumption,’’ that is, the idea that every source of infor-
mation that constrains the interpretation of an utterance
(syntax, prosody, word-level semantics, prior discourse,
world knowledge, knowledge about the speaker, ges-
turas, etc.) poder, in principle, do so immediately (Hagoort
& van Berkum, 2007; Spivey Knowlton & Sedivy, 1995;
Tanenhaus, Spivey Knowlton, Eberhard, & Sedivy, 1995;
Tanenhaus & Trueswell, 1995; Trueswell & Tanenhaus,
1994; Taraban & McClelland, 1990). Summarized, propo-
nents of a two-step model would expect indicators of
semantic integration in the ERP to be manifested earlier
when a word has to be integrated as compared to when a
picture has to be integrated into the previous sentence
contexto.
In terms of cortical areas important for language com-
prehension, a recent neurobiological account of lan-
guage comprehension has argued for the left inferior
frontal cortex to be a general (es decir., not domain-specific)
unification site (Hagoort, 2005a, 2005b). Unification
entails integration of information into a built-up repre-
sentation of the previous sentence context as well as
a selection of appropriate candidates for integration
(Hagoort, 2005a, 2005b). When unification is more dif-
ficult, more resources are needed to integrate linguistic
as well as extralinguistic information, resulting in in-
creased activation levels in the left inferior frontal cortex.
If this is indeed the case, we should observe increased
activation both when a picture and when a word are
harder to integrate. Sin embargo, if this area’s role is re-
stricted to integrating language information, no such in-
crease should be observed when information conveyed
through a picture has to be integrated.
On the basis of previous studies investigating sentence-
integration of co-speech gestures compared to
nivel
palabras, we have argued for linguistic and extralinguistic
information to be integrated in the same way into a
linguistic context (O¨ zyu¨rek, Willems, Kita, & Hagoort,
2007; Willems et al., 2007). In these studies, the seman-
tic fit of a word or of a co-occurring co-speech gesture
to the preceding sentence context was manipulated.
Mismatching spoken words and co-speech gestures elic-
ited N400 effects with similar onset latencies. A pesar de
co-speech gestures and the pictures that we investigate
here are both extralinguistic information, clear differ-
ences exist as well. One important characteristic of co-
speech gestures is that their meaning is not recognized
unambiguously when presented outside of a language
contexto (Krauss, Morrel Samuels, & Colasante, 1991). En
contrast, pictures can stand on their own. Por lo tanto,
the present study is a stronger test for the claim that
semantic integration at the sentence level is not domain
(es decir., idioma) specific. If neural correlates of integra-
tion of pictures and words are similar, it follows that also
information that is not necessarily bound to a language
context is integrated with the same spatio-temporal pro-
file in the brain as linguistic information is.
To address these questions, we presented participants
with spoken sentences in which a critical word was
manipulated to either fit the sentence context or not.
The critical words were accompanied by pictures (es decir.,
line drawings) that could also either match or mismatch
with regard to the previous part of the sentence. Este
manipulation resulted in four conditions (ver tabla 1):
correct condition [Picture (PAG) +, Idioma (l) +];
language mismatch condition (P+L(cid:1)); picture mismatch
condición (PAG(cid:1)L+); double mismatch condition (PAG(cid:1)l(cid:1)).
In the language mismatch, the critical word was harder
to integrate semantically into the preceding sentence
contexto, whereas the co-occurring picture matched the
sentence context. In the picture mismatch condition,
the picture was harder to integrate into the previous
contexto, whereas the critical word matched the spoken
sentence context. In the double mismatch condition,
both the picture and the word were difficult to integrate
into the previous sentence context. Note that in the
language and picture mismatch conditions the critical
word and the overlapping picture locally mismatched
(p.ej., picture CHERRY, word ‘‘flower,’’ and vice-versa),
whereas in the double mismatch condition they lo-
cally matched (p.ej., both ‘‘cherry’’). This manipulation
Willems, O¨ zyu¨rek, and Hagoort
1237
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
0
yo
7
mi
–
1
pag
2
d
3
F
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
oh
3
C
5
norte
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
8
mi
.
s
t
2
oh
0
norte
0
8
0
5
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
.
F
.
t
.
/
oh
norte
1
8
METRO
a
y
2
0
2
1
Mesa 1. An Example of the Stimulus Materials
Dutch: De man gaf zijn vrouw een mooie bloem/kers die avond
Inglés: ‘‘The man gave his wife a nice flower/cherry that
evening’’
Correct condition
P+L+: The man gave his wife a nice flower that evening
Language mismatch
P+L(cid:1): The man gave his wife a nice cherry that evening
Picture mismatch
PAG(cid:1)L+: The man gave his wife a nice flower that evening
Double mismatch
PAG(cid:1)l(cid:1): The man gave his wife a nice cherry that evening
Pictures were displayed time-locked to the onset of the noun (bajo-
lined). Note that the condition coding (P+L+, P+L(cid:1), etc.) refers to
the match/mismatch of either the noun (Idioma: l) or the Picture
(Picture: PAG) to the part of the sentence preceding the word that is
underlined, with a minus sign indicating a mismatch. Eso es, en el
correct condition (P+L+), both the word ‘‘flower’’ as well as the
picture [FLOWER] fit the preceding sentence context. In the language
mismatch condition (P+L(cid:1)), the word ‘‘cherry’’ does fit the preceding
sentence context less well, whereas the picture [FLOWER] does fit.
En cambio, in the picture mismatch condition (PAG(cid:1)L+), the picture
[CHERRY] does not fit the preceding sentence context, mientras que el
word ‘‘f lower’’ does fit. Finalmente, in the double mismatch condition
(PAG(cid:1)l(cid:1)), both the word ‘‘cherry’’ and the picture [CHERRY] do not fit
the preceding sentence context. Mismatching words are indicated in
bold. All stimuli were in Dutch.
enabled us to distinguish integration at the ‘‘local’’ level
of simultaneously occurring word and picture from
integration at the ‘‘global’’ sentence level; eso es, no-
gration into a higher-level representation built-up on the
basis of the preceding context information.
We had three specific hypotheses. Primero, for the ERP
data we hypothesized that manipulating the match of
both picture and word would lead to an N400 effect
comparable in size and onset latency. Además, nosotros
were curious to see if an N300 effect would be apparent
and if so, whether it would be specific to the picture
mismatch condition. Previous ERP studies have mostly
compared the presence of an N300 effect in reaction to
pictures to the absence of an N300 to words indirectly.
Eso es, in most studies, either words or pictures were
presentado. Our design allows for assessing the functional
relevance of the N300 in the sense that if it is sensitive to
semantic load of a picture it should occur in the picture
mismatch condition but not in the language mismatch
condición. Segundo, for the fMRI study, we predict a
stronger involvement of the inferior frontal cortex in both
the picture and word mismatch conditions. En ese caso, este
would be evidence that this region, besides its well-
established role in spoken and written language compre-
hension (p.ej., Vigneau et al., 2006; Bookheimer, 2002),
also takes extralinguistic visual information into account
during language comprehension. Tercero, on the basis of
earlier findings (O¨ zyu¨rek et al., 2007), we hypothesized
our findings to reflect semantic processing at the ‘‘global’’
sentence level but not at the ‘‘local’’ level of the simul-
taneous picture and word. In relation to this, it should
be noted that recent data acquired in our lab show that
without an elaborate context, simultaneously presented
incongruent word and object pairs do not elicit an N400
(O¨ zyu¨rek, Willems, & Hagoort, unpublished results).
En general, our main question regards the similarity or dis-
similarity of integrating linguistic and extralinguistic in-
formation into a sentence context. Differences in neural
indicators of semantic processing would favor an account
in which linguistic information has a preferred status in
sentence integration (Fodor, 1983; Forster, 1979), dónde-
as findings of similar neural correlates would support the
idea that linguistic and extralinguistic information are in-
tegrated with a similar neural time course and by recruit-
ing overlapping cortical areas (Hagoort & van Berkum,
2007). Además, an earlier effect to words than to pic-
tures would be in favor of two-step models of language
comprensión (p.ej., Lattner & Friederici, 2003; Cutler &
Clifton, 1999), whereas similar neural time courses would
favor accounts of immediacy in which a broad range of
information types is immediately incorporated into a
discourse model (Hagoort & van Berkum, 2007; Spivey
Knowlton & Sedivy, 1995; Tanenhaus & Trueswell, 1995;
Tanenhaus et al., 1995; Trueswell & Tanenhaus, 1994;
Taraban & McClelland, 1990).
MÉTODOS
Materials and Procedure
Un total de 328 oraciones (mean duration 3196 mseg, range
2164–4184 msec) were recorded in a sound-attenuated
room at 44.1 kHz, spoken at a normal rate by a native
Dutch female speaker. Half of these sentences differed
in one critical word, which was never in sentence-final
posición. In each sentence, a short context was intro-
duced to which the critical word could fit more or less
easily. Critical words were nouns that corresponded to
the names given by a separate group of participants (norte =
1238
Revista de neurociencia cognitiva
Volumen 20, Número 7
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
F
/
t
t
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
0
yo
7
mi
–
1
pag
2
d
3
F
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
oh
3
C
5
norte
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
8
mi
.
s
t
2
oh
0
norte
0
8
0
5
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
/
.
t
F
.
.
oh
norte
1
8
METRO
a
y
2
0
2
1
32) to a large set of black and white line drawings. Todo
critical words had a picture equivalent with a naming
consistency of 85% or higher. In total there were 26 crit-
ical words with their picture equivalents. All words were
one syllable long and started with a plosive consonant.
Every critical word occurred equally often in a matching
and a mismatching sentence context. The critical word in
the mismatching sentence always had a different onset
consonant than the critical word in the semantically cor-
rect sentence. Sentences were pretested in a cloze prob-
ability test that was given to a separate participant group
(norte = 16). The percentage of participants that gave the
target word as response was taken as a measure of its
cloze probability. En general, the mean cloze probability was
16% for the matching critical words (range 0–69%), y
0% for the semantically anomalous critical words. Nosotros
choose for critical words with low cloze probabilities to
avoid confounding effects of prediction (p.ej., Van Berkum,
Marrón, Zwitserlood, Kooijman, & Hagoort, 2005).
Our manipulation resulted in four conditions (Mesa 1):
(i) correct condition (P+L+); (ii) language mismatch
(P+L(cid:1)); (iii) picture mismatch (PAG(cid:1)L+); (iv) double mis-
match (PAG(cid:1)l(cid:1)). Note that mismatch in these materials is
always defined relative to the preceding sentence context.
Four stimulus lists of 164 trials each were created in
which only one item of every stimulus quartet (as in
Mesa 1) was presented. Sentences were pseudorandom-
ized with the constraint that the same condition occurred
maximally two times in a row. Every list contained an
equal amount of stimuli from the four conditions (41 por
condición). Every target word and picture was repeated,
on average, 6.3 veces (range = 5–8, modus = 6, median =
6 repetitions) in every stimulus list. Pictures were pre-
sented from the onset of the critical word to the end of
la frase.
EXPERIMENT 1: EEG
Participantes
Twenty-four healthy right-handed (Oldfield, 1971) par-
ticipants with Dutch as their mother tongue took part in
the electroencephalogram (EEG) estudiar. None had any
known neurological history or hearing complaints, y
all had normal or corrected-to-normal vision. Eight par-
ticipants’ data had to be discarded because of an ex-
cessive number of blinks and eye movements, leaving
datasets from 16 Participantes (edad media = 22.4 años,
range = 18–34, 11 women). Subjects were paid for
participación. The local ethics committee approved the
study and all participants signed informed consent in
accordance with the Declaration of Helsinki.
Procedimiento
Stimuli were presented using Presentation software
(versión 9.13, www.neuro-bs.com/ ). Pictures had vary-
ing sizes depending upon the object they represented
and were maximally 8 (cid:2) 8 cm, shown at a viewing
distance of 90 cm (58 (cid:2) 58 visual angle). A trial started
con 600 msec blank screen, followed by the spoken
sentence and the picture, 1000 msec blank screen, y
2500 msec with a fixation cross on the screen. Participantes
were instructed to sit still in a comfortable position and to
blink only when a fixation cross was presented. The test
session started with eight trials which contained different
critical words than used in the main part of the experi-
mento. Participants were told to attentively listen to and
watch the stimuli about which they would receive ques-
tions afterward. At the end of the test session, general
questions about the stimuli were asked. All participants
had understood the manipulation in the materials and
could provide examples of stimuli.
Recording and Analysis
The EEG was recorded from 27 electrode sites across
the scalp using an Electrocap with Ag/AgCl electrodes,
each referred to the left mastoid. Electrodes were placed
on standard electrode sites (Fz, FCz, Cz, Pz, Fp2, F3, F4,
F8, F7, FC5, FC1, FC2, FC6, T7, T8, C3, C4, CP5, CP1,
CP2, CP6, P7, P3, P4, P8, O1, O2). Vertical eye move-
ments and blinks were monitored by means of two
electrodes, one placed beneath and one above the left
ojo. Horizontal eye movements were monitored by
means of a left to right bicanthal montage. Activity over
the right mastoid was recorded to determine whether
there were additional contributions of the experimental
variables to the two presumably neutral mastoid sites.
No such differences were observed. The EEG and elec-
trooculogram (EOG) recordings were amplified with
BrainAmp DC amplifiers, using a band-pass filter from
10 sec to 100 Hz. Impedances were kept below 5 k(cid:1) para
all channels. The EEG and EOG signals were recorded
and digitized using Brain Vision Recorder software (ver-
sión 1.03), with a sampling frequency of 500 Hz.
The data were filtered off-line with a 30-Hz low-pass
filter, re-referenced to the mean of the two mastoids,
and segmented from 150 msec before to 1000 mseg después
the critical word. Segments were normalized to the
mean amplitude of a baseline period 150 msec before
the critical word (baseline correction). All segments
were screened for eye movements, electrode drifting,
amplifier blocking, and muscle artifacts. Trials containing
such artifacts were rejected (mean = 8.6%, DE = 5.2%,
range = 0–18%). Rejected trials were equally distributed
across conditions (F < 1). Segments were averaged for
each condition for each participant at each electrode
site. Repeated measures analysis of variance (ANOVA)
was applied to the mean activity in four time windows
(see Results) with factors condition (P+L+, P+L(cid:1),
P(cid:1)L+, P(cid:1)L(cid:1)) and quadrant (left anterior, right anterior,
left posterior, right posterior). Electrodes were assigned
to quadrants as follows: left anterior (F3, F7, FC1, FC5,
C3), right anterior (F4, F8, FC2, FC6, C4), left posterior
Willems, O¨ zyu¨rek, and Hagoort
1239
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
7
e
-
1
p
2
d
3
f
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
o
3
c
5
n
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
0
8
0
5
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
.
f
/
.
o
n
1
8
M
a
y
2
0
2
1
(CP1, CP5, P3, P7, O1), and right posterior (CP2, CP6, P4,
P8, O2). A separate ANOVA was performed for the mid-
line electrodes (Fz, FCz, Cz, Pz). Huynh–Feldt correction
for violation of sphericity assumption was applied when
appropriate (Luck, 2005; Picton et al., 2000; Huynh &
Feldt, 1976). Differences in N400 effect onset latencies
were tested by calculating the time bin (bins of 10 msec)
in which 20% of the total area of the difference waves of
the experimental conditions with the correct condition in
the 200–500 msec latency window was reached (fractional
area latency analysis). Statistical significance of these dif-
ferences was assessed by using the jackknifing procedure
described by Miller, Patterson, and Ulrich (1998).
Results
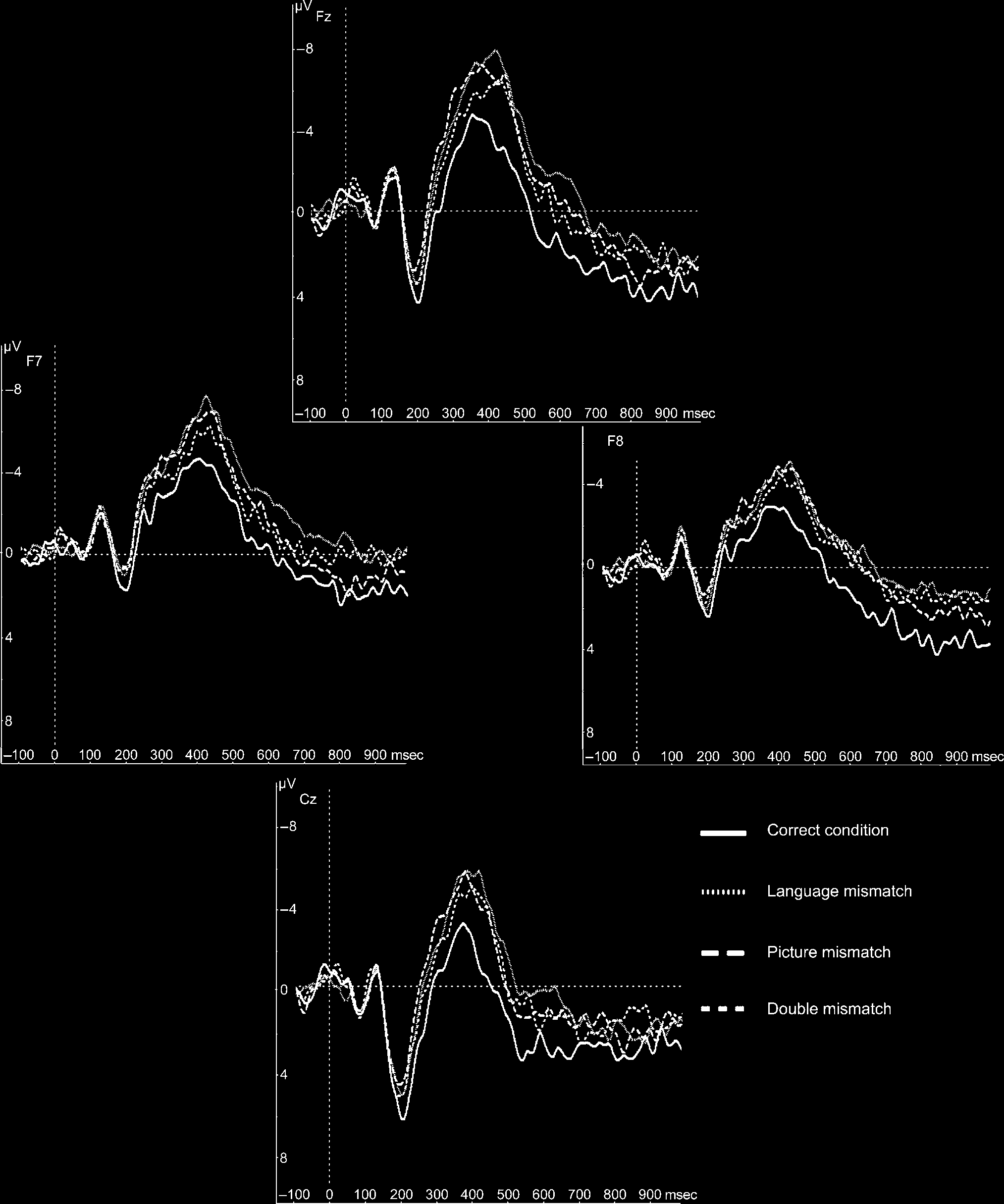
The grand-average waveforms (Figure 1) show clear N1
and P2 components followed by a negativity starting
from 350 msec resembling the N400. The correct con-
dition showed a slightly decreased peak at the N1 and a
stronger positivity at the P2 peak compared to the other
three conditions (Figures 1 and 2A). In the N400 time
window, the three mismatch conditions (P+L(cid:1), P(cid:1)L+,
P(cid:1)L(cid:1)) were more negative than the correct condition.
The mismatch conditions stayed more negative than the
correct condition until the end of the segment. Conse-
quently, ANOVAs were done on the mean amplitudes in
the latency ranges 170–250 msec, 300–550 msec, and
600–900 msec. Additional tests were performed in the
N300 time window (225–325 msec) given that previous
literature (e.g., McPherson & Holcomb, 1999) reports
picture-specific effects in this time window.
P2 Time Window (170–250 msec)
Statistical analyses in this time window failed to reveal a
significant effect of condition [F(3, 45) = 2.34, p = .09].
There was also no Condition (cid:2) Quadrant interaction (F <
1). However,
in the ANOVA over midline electrodes,
a main effect of condition was found [F(3, 45) = 2.87,
MSE = 9.97, p = .047]. Planned comparisons over the
midline electrodes, of every experimental condition ver-
sus the correct condition, showed this effect to be stron-
gest in the picture mismatch condition [F(1, 15) = 5.82,
MSE = 26.96, p = .029], although there were also mar-
ginally significant effects in the language mismatch versus
correct condition [F(1, 15) = 4.29, MSE = 21.27, p = .056]
and in the double mismatch versus correct condition com-
parisons [F(1, 15) = 4.51, MSE = 10.75, p = .051].
N300 Time Window (225–325 msec)
The morphology of the grand-average waveforms does
not clearly indicate the presence of a separate N300
component. Given previous findings of the N300 for mis-
matching pictures, we tested effects in the 225–325 msec
time window (e.g., McPherson & Holcomb, 1999). A main
effect of condition was observed [F(3, 45) = 3.17, MSE =
42.96, p = .040], but no Condition (cid:2) Quadrant interac-
tion [F(9, 135) = 1.26, MSE = 10.22, p = .293]. Pairwise
comparisons between all conditions were performed and
p values were corrected for the number of tests accord-
ingly. Only the double mismatch versus correct condition
differed significantly from each other [F(1, 15) = 9.82,
MSE = 17.3, p = .042]. Since previous studies found the
distribution of the N300 effect to be frontal, we separately
tested in left and right anterior quadrants. Again, there
was a main effect of condition [left: F(3, 45) = 3.47,
MSE = 22.54, p = .040; right: F(3, 45) = 4.22, MSE =
11.88, p = .010]. Pairwise comparisons revealed only the
double mismatch condition to be significantly different
from the correct condition in the left anterior quadrant
[F(1, 15) = 11.33, MSE = 10.17, p = .024]. The picture
mismatch versus correct condition was marginally signif-
icant in the left anterior quadrant only [F(1, 15) = 8.20,
MSE = 34.96, p = .073]. No other comparisons revealed
significant differences between conditions. Summarized,
although there is a main effect of condition in this time
window, this effect is not specific to the picture and/
or double mismatch conditions, as might have been ex-
pected based upon previous findings.
N400 Time Window (300–550 msec)
Table 2 summarizes the results in this time window.
There was a main effect of condition [F(3, 45) = 11.46,
p < .001], but no Condition (cid:2) Quadrant interaction
[F(9, 135) = 1.56, p = .16]. To explore specific differ-
ences between conditions, pairwise comparisons were
performed. Accordingly, the p values are corrected for
the number of tests performed (see Table 2). Pairwise
comparisons revealed that all mismatch conditions dif-
fered significantly from the correct condition. No other
comparisons showed significant differences between
conditions (Table 2). These effects are spatially smeared
out over the scalp, with a tendency for fronto-central
electrodes to show the greatest effect size (Figure 2B).
To formally test the onset latencies of the N400 effects, a
fractional area latency measure was computed in the
200–500 msec time window (see EEG Methods). The
time point at which 20% of the grand-average difference
waveform was reached was 305 msec for the language
mismatch–correct condition, 285 msec for the picture
mismatch–correct condition, and 295 msec for the dou-
ble mismatch–correct condition. Statistical testing of
these differences (see Miller et al., 1998) revealed that
onset latencies did not differ from each other (all t < 1).
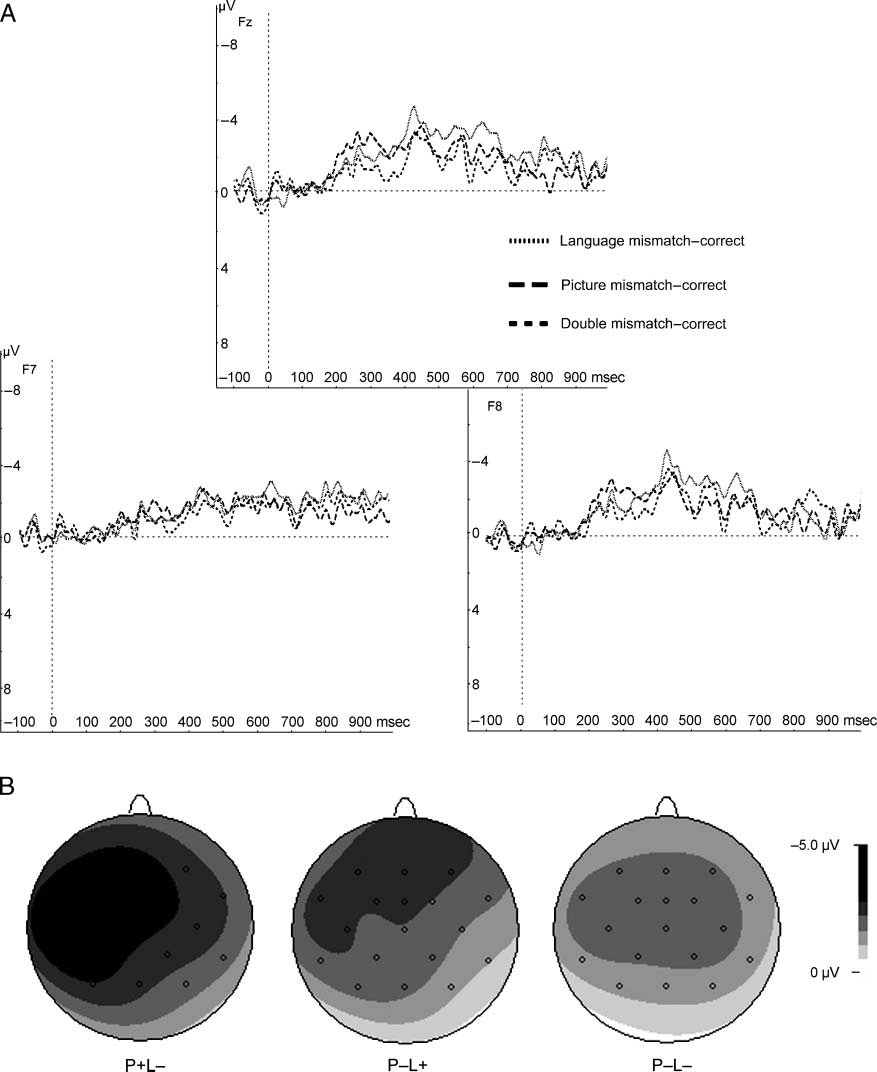
Finally, the scalp distributions of the difference waves
(Figure 2B) were tested in a two-way ANOVA with factors
difference (language mismatch–correct condition, picture
mismatch–correct condition, double mismatch–correct
condition) and quadrant. The lack of an interaction effect
1240
Journal of Cognitive Neuroscience
Volume 20, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
7
e
-
1
p
2
d
3
f
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
o
3
c
5
n
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
0
8
0
5
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
t
.
.
.
/
o
n
1
8
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
7
e
-
1
p
2
d
3
f
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
o
3
c
5
n
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
0
8
0
5
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
/
t
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 1. Grand-average ERPs for the four conditions at electrodes Fz, F7, F8, and Cz. ERPs were time-locked to the onset of the critical word
and picture. Negativity is plotted upward.
Willems, O¨ zyu¨rek, and Hagoort
1241
(F < 1) indicates that the scalp distributions were similar
for the N400 effects elicited by the mismatch conditions.
Late Time Window (600–900 msec)
In the late time window (600–900 msec), there was a
significant main effect of condition [F(3, 45) = 3.71,
MSE = 31.55, p = .018] and a significant Condition (cid:2)
Quadrant interaction [F(9, 135) = 3.62, MSE = 6.26,
p = .003]. Subsequent tests in specific quadrants re-
vealed main effects of condition only in the left ante-
rior [F(3, 45) = 5.82, MSE = 13.91, p = .002] and right
anterior quadrants [F(3, 45) = 5.80, MSE = 11.97, p =
.002]. Again, we tested pairwise comparisons between all
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
7
e
-
1
p
2
d
3
f
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
o
3
c
5
n
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
0
8
0
5
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
/
.
t
.
o
n
1
8
M
a
y
2
0
2
1
Figure 2. (A) Difference waves of the experimental conditions minus the correct condition (language mismatch–correct condition; picture
mismatch–correct condition; double mismatch–correct condition) at electrodes Fz, F7, and F8. Difference waves are time-locked to the onset
of the critical word and picture. Negativity is plotted upward. (B) Spline interpolated isovoltage maps of the mean difference wave 300–550 msec
after the critical word. Displayed are the difference of the language mismatch condition (left), picture mismatch condition (middle), and double
mismatch condition (right) with the correct condition.
1242
Journal of Cognitive Neuroscience
Volume 20, Number 7
Table 2. ERP Results in the 300–550 msec Time Window
Source
df
F
MSE
p
ANOVA (4 Conditions (cid:2) 4 Quadrants)
Condition
3, 45
11.46
26.72 < .001***
Condition (cid:2) Quadrant
9, 135
1.56
8.86
.162
Pairwise comparisons
p(corr.)
and double mismatch conditions were not different
from the correct conditions [F(1, 15) = 5.30, MSE =
29.86, p = .216; P(cid:1)L(cid:1) vs. P+L+: F(1, 15) = 8.31, MSE =
27.53, p > .5]. Other comparisons did not reveal signif-
icant differences between conditions. Over the midline
electrodes, there was only a trend for a main effect of
condición [F(3, 45) = 2.45, MSE = 12.41, pag = .076].
42.34
39.18 < .001***
Discussion
P+L(cid:1) vs. P+L+
P(cid:1)L+ vs. P+L+
P(cid:1)L(cid:1) vs. P+L+
P+L(cid:1) vs. P(cid:1)L(cid:1)
P(cid:1)L+ vs. P(cid:1)L(cid:1)
P(cid:1)L+ vs. P+L(cid:1)
1, 15
1, 15
1, 15
1, 15
1, 15
1, 15
11.42
73.08
.025*
15.39
37.05
.008**
4.66
60.93
.285
0.67
37.53
2.38
58.83
ns
ns
Midline (4 Conditions (cid:2) 4 Electrodes)
Condition
3, 45
9.81
10.58 < .001***
Pairwise comparisons
p(corr.)
P+L(cid:1) vs. P+L+
P(cid:1)L+ vs. P+L+
P(cid:1)L(cid:1) vs. P+L+
P+L(cid:1) vs. P(cid:1)L(cid:1)
P(cid:1)L+ vs. P(cid:1)L(cid:1)
P(cid:1)L+ vs. P+L(cid:1)
1, 15
1, 15
1, 15
1, 15
1, 15
1, 15
37.72
13.76 < .001***
10.82
28.82
10.82
19.31
.029*
.030*
2.92
23.76
0.75
13.79
1.50
17.42
ns
ns
ns
Amplitudes of the ERPs were averaged over this time window for every
participant separately and entered into repeated measures ANOVA with
factors condition (four levels) and quadrant (four levels). A separate
ANOVA was performed for the midline electrodes with factors condition
(four levels) and electrode (four levels). Huynh–Feldt correction for
violation of sphericity assumption was applied (Luck, 2005; Picton et al.,
2000; Huynh & Feldt, 1976), but the original degrees of freedom are
reported. The significance levels for the pairwise comparisons were cor-
rected for the number of tests performed by means of Bonferroni cor-
rection. The corrected p levels are reported; effects with p values > .5
are reported as not significant (ns). P+L+ = correct condition; P+L(cid:1) =
language mismatch condition; PAG(cid:1)L+ = picture mismatch condition;
PAG(cid:1)l(cid:1) = double mismatch condition.
*pag < .05.
**p < .01.
***p < .001.
conditions and we report adjusted p values accordingly.
In the left anterior quadrant, the language mismatch dif-
fered significantly from the correct condition [F(1, 15) =
30.74, MSE = 15.10, p < .001]. The double mismatch
condition and the picture mismatch condition did not
differ significantly from the correct condition [F(1, 15) =
4.31, MSE = 40.54, p = .33 and F(1, 15) = 2.43, MSE =
30.47, p > .5, respectivamente]. In the right anterior quad-
rant, a qualitatively similar pattern of results was ob-
served with language mismatch being significantly
different from the correct condition [F(1, 15) = 11.54,
MSE = 32.54, pag = .024], whereas the picture mismatch
D
oh
w
norte
yo
oh
a
d
mi
d
yo
yo
/
/
/
/
j
t
t
F
/
i
t
.
:
/
/
F
r
oh
metro
D
h
oh
t
w
t
norte
pag
oh
:
a
/
d
/
mi
metro
d
i
F
t
r
oh
pag
metro
r
C
h
.
s
pag
i
yo
d
v
i
mi
r
mi
r
C
C
t
.
h
metro
a
i
r
mi
.
d
tu
C
oh
oh
metro
C
/
norte
j
a
oh
r
C
t
i
norte
C
/
mi
a
–
pag
r
d
t
i
2
C
0
yo
7
mi
–
1
pag
2
d
3
F
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
oh
3
C
5
norte
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
oh
pag
C
d
norte
.
b
y
2
0
gramo
0
tu
8
mi
.
s
t
2
oh
0
norte
0
8
0
5
8
.
S
pag
mi
d
pag
F
mi
metro
b
y
b
mi
gramo
r
tu
2
0
mi
2
s
3
t
/
j
F
.
/
.
t
.
oh
norte
1
8
METRO
a
y
2
0
2
1
Highly similar N400 effects were found for all experimen-
tal conditions compared to the correct condition. Com-
paring our ERP findings to earlier studies investigating
semantic processing of pictures, a few differences are
readily apparent. In contrast to some earlier findings
(Federmeier & Kutas, 2001, 2002; Oeste & Holcomb,
2002; McPherson & Holcomb, 1999), we failed to observe
a separate N300 effect, which has been claimed to be
specific to the processing of pictorial stimuli. Sin embargo,
some other studies investigating pictures in a sentence
context have also failed to observe a separate N300 effect
(Ganis et al., 1996; Nigam et al., 1992). Por lo tanto, nosotros
argue for the N300 as not being specific to the processing
of pictures, at least not when presented within a sentence
contexto. The absence of a picture-specific effect in the
ERP waveforms and the similar time course of the N400
suggest that at this level of processing, no differentiation
is made between verbal and visual semantic information.
The double mismatch condition, in which both the
word and the picture did fit the previous context less
Bueno, evoked an N400 similar in latency and amplitude
than the other mismatch conditions,
in which either
word or picture were in discordance with the previous
contexto. Conflicting information coming from the visual
or verbal domain does not add up linearly to increase the
effect size in the double mismatch. Además, the fact
that the double mismatch N400 starts at the same latency
as the other two mismatch conditions speaks in favor of
the ‘‘immediacy assumption’’ which predicts that infor-
mation is used by the language comprehension system as
soon as it is available. En otras palabras, it suggests that
picture and word are not first integrated at a lower level
of processing before being integrated into the sentence
contexto. If this were the case, a delay in the N400 re-
sponse to the language mismatch condition and the pic-
ture mismatch condition would have been expected.
Curiosamente, an earlier, marginally significant difference
between mismatch conditions and the correct condition
could be observed in the time window of the P2 compo-
próximo. This is most parsimoniously explained as a lead-in
effect of the subsequent N400, which was much more
negative for the three mismatch conditions than for the
correct condition.
In line with earlier ERP studies, the scalp distribution
of the N400 effect was more frontal than the centro-
posterior distribution that is normally observed in studies
of spoken or written language. The frontal distribution
Willems, O¨ zyu¨rek, and Hagoort
1243
era, sin embargo, not specific to the picture mismatch
condición. Por lo tanto, although the presence of visual
information might shift the N400 distribution to a more
frontal maximum, the fact that this holds even when the
anomaly is language-internal argues against a picture-
specific integration process that is different from semantic
integration of written or spoken words. Together with
results in an earlier study (O¨ zyu¨rek et al., 2007), it seems
that the mere presence of a visual stimulus (other than a
written word) makes the scalp distribution ‘‘shift’’ to a
more frontal maximum compared to when only linguistic
information is presented.
The stronger negativities to the mismatch conditions
in the late time window resemble the findings in the
N400 time window, although only the language mis-
match condition differed significantly from the correct
condición. Sin embargo, no differences were observed be-
tween the experimental conditions. Por lo tanto, estos
late effects are best explained as a carryover effect of
the strong N400 effects.
EXPERIMENT 2: f MRI
Participantes
Nineteen healthy right-handed (Oldfield, 1971) partícipe-
ipants with Dutch as their mother tongue took part in
the fMRI study. None had any known neurological
history or hearing complaints, and all had normal or
corrected-to-normal vision. Three datasets in the fMRI
study had to be discarded, two because of inattentive
Participantes (see below) and one because of excessive
head motion. Data from the 16 remaining participants
(edad media = 22.3 años, range 20–28, 8 women) eran
entered into the analysis. Participants were paid for
participación. The local ethics committee approved the
study and all participants signed informed consent in
accordance with the Declaration of Helsinki.
Procedimiento
Stimuli were the same as in the EEG experiment. Pic-
tures were projected from outside of the scanner room
onto a screen at the end of the patient table. The screen
was visible through a mirror mounted to the head coil,
at a viewing distance of 80 cm (pictures subtended maxi-
mally 5.78 (cid:2) 5.78 visual angle). Speech was presented
through nonmagnetic headphones (Commander XG,
www.mrivideo.com), which dampened scanner noise.
Intertrial interval was 6, 7, o 8 segundo. During the scanning
session eye movements were recorded using an infrared
IviewX eyetracker (www.smi.de) to formally control par-
ticipant’s vigilance during scanning.
The scanner was switched on during the practice trials
and participants had to indicate whether the volume
should go up or down. No participant asked for the vol-
ume to be increased to the maximally possible level.
Participants were told to attentively listen to and watch
the stimuli about which they would receive questions
afterwards. All participants indicated they were able to
hear and understand the sentences well. At the end of the
scanning session, general questions about the stimuli were
asked. All participants had understood the manipulation
in the materials and could provide examples of stimuli.
Recording and Analysis
MR imaging was performed on a 3-T Siemens Magnetom
Trio scanner (Siemens, Erlangen, Alemania). Approxi-
mately 800 echo-planar whole-brain images were ac-
quired per participant (TR = 2230 mseg; TE = 30 mseg;
flip angle = 808; 32 slices; slice thickness = 4 mm; FOV =
224 mm, voxel resolution = 3.5 (cid:2) 3.5 (cid:2) 4 mm). Ad-
ditionally, a T1-weighted anatomical scan (3-D MP-RAGE,
192 slices, TR = 2300 mseg; TE = 3.93 mseg; FOV =
256 mm; slice thickness = 1 mm) was made. Data were
analyzed using Brainvoyager QX (Brain Innovation, www.
brainvoyager.com). The first five volumes of a session
were discarded to avoid T1 saturation effects. Preprocess-
ing involved rigid-body transformations of all volumes to
the first volume, slice scan time correction, linear trend
removal, and high-pass temporal filtering (cutoff 3 cy-
cles over the time course) and spatial smoothing with a
Gaussian filter kernel of 8 mm full width at half maximum.
Data were transformed into stereotaxic space (Talairach
& Tournoux, 1988). A whole-brain analysis was performed
in the context of the General Linear Model, con el
conditions as factors of interest and the six parameters
from the motion correction as nuisance factors. Experi-
mental factors were modeled for the duration of each
sentence and convolved with a canonical 2 gamma hemo-
dynamic response function. Parameters were estimated
for every voxel’s time course. Effect sizes were estimated
by constructing contrast (t) maps consisting of differences
between the parameter estimates in every voxel and par-
ticipant separately for contrasts of interest. Después,
contrast maps were taken to a second-level analysis, prueba-
ing for differences from zero in a one-sample t test in a
random effects analysis. Every contrast was tested two-
sided. The multiple comparisons problem was addressed
by thresholding the activation maps at t(15) = 3.9, pag <
.001 at the voxel level and taking the cluster sizes into
account, leading to a correction at an alpha level of p < .05
(Forman et al., 1995). The eyetracking data were used
to control for the vigilance (i.e., wakefulness) of the par-
ticipant. Two datasets had to be discarded because par-
ticipants had their eyes closed in more than 10% of the
trials.
Results
To see effects specific for the language condition, the
language mismatch condition (P+L(cid:1)) was contrasted
against the correct condition (P+L+). The correct con-
1244
Journal of Cognitive Neuroscience
Volume 20, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
7
e
-
1
p
2
d
3
f
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
o
3
c
5
n
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
0
8
0
5
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
t
.
.
f
.
o
n
1
8
M
a
y
2
0
2
1
dition served as a high-level baseline in this way. An
extensive region in the left inferior frontal cortex,
stretching into the premotor cortex, and an area in
the left superior temporal sulcus were found activated
(Table 3; Figure 3A). One area in the right middle fron-
tal sulcus was activated in the reversed contrast (i.e.,
correct condition vs. language mismatch). Second, ef-
fects to the picture condition were assessed, again by
comparing it to the correct condition (P(cid:1)L+ vs. P+L+).
Part of the left inferior frontal sulcus showed significant
activation to this contrast (Table 3; Figure 3B). Finally,
the double mismatch condition compared to correct
condition tested the effect of both picture and word
being in discordance with the sentence context. This
comparison (P(cid:1)L(cid:1) vs. P+L+) led to increased activity in
an extensive part of the inferior frontal cortex stretching
into the premotor cortex, an area in the left superior
temporal sulcus, an area in the left temporo-parietal junc-
tion, and a small area of activation in the right cerebel-
lum (Table 3; Figure 3C).
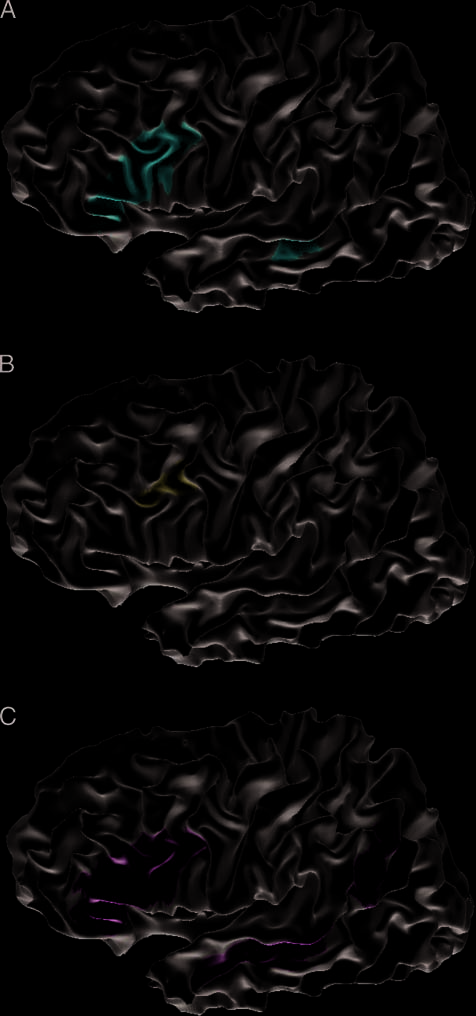
Figure 4A displays the results of a conjunction analysis
(conjunction as in a logical AND; see Nichols, Brett,
Andersson, Wager, & Poline, 2005) testing for overlap
between the comparisons described above (P(cid:1)L+ vs.
P+L+ \ P+L(cid:1) vs. P+L+ \ P(cid:1)L(cid:1) vs. P+L+). One re-
gion in the left inferior frontal cortex (max TAL [(cid:1)40 11
31]) was found activated in this contrast. Compared to
a cytoarchitectonic probability map, 39% of this region
overlapped with BA 44, in contrast to only 3% of the
voxels that were classified as being part of BA 45 (Eickhoff
et al., 2005).
Finally, we tested for differential effects to the local
and global match or mismatch effects. This was done by
comparing the language mismatch and picture mismatch
to the double mismatch condition (P+L(cid:1) vs. P(cid:1)L(cid:1) \
P(cid:1)L+ vs. P(cid:1)L(cid:1)). In this way, all conditions involve a
sentence-level mismatch, but only the language and
picture mismatch conditions had an additional local mis-
match. No areas were found activated in this contrast.
Discussion
We observed increased activation levels in all three
mismatch conditions compared to the correct condition
in the left inferior frontal cortex. This study adds to a
large number of studies showing that the left inferior
frontal cortex is an important node in the speech
comprehension network (for reviews, see Vigneau
et al., 2006; Bookheimer, 2002). We interpret our find-
ings as ref lecting unification processing in the left
inferior frontal cortex. This entails integration of infor-
mation into a built-up representation of the previous
sentence context as well as a selection of appropriate
candidates for integration (Hagoort, 2005a, 2005b). We
show here that integration of extralinguistic information,
such as a visual picture, recruits this area.
The peak of overlapping activation in the inferior fron-
tal cortex was in BA 44, which is at odds with proposals
of a gradient of linguistic subprocesses (such as seman-
tics, syntax, and phonology) within the left inferior fron-
tal cortex ( Vigneau et al., 2006; Bookheimer, 2002).
Semantic processing is centered more ventrally, around
BA 45 and 47 in these proposals (Vigneau et al., 2006;
Bookheimer, 2002). However, as can be seen in Figure 3,
two of the mismatch conditions did activate a more
ventral part of the left inferior frontal cortex (language
Table 3. Results from the fMRI Experiment
Center Coordinates
Contrast
x
Language mismatch versus
(cid:1)45
correct condition
Picture mismatch versus
correct condition
Double mismatch versus
correct condition
Correct condition versus
language mismatch
(cid:1)52
(cid:1)38
(cid:1)43
(cid:1)50
(cid:1)38
17
28
y
15
(cid:1)34
8
17
(cid:1)29
(cid:1)60
(cid:1)38
30
z
24
(cid:1)1
32
Region
t (Max)
Number of Voxels
(1 (cid:2) 1 (cid:2) 1 mm)
Left inferior frontal sulcus/
premotor cortex
Left superior temporal sulcus
Left inferior frontal sulcus
8.25
5.42
6.23
8655
2507
2404
23
Left inferior frontal sulcus/
11.44
17304
premotor cortex
(cid:1)6
14
(cid:1)27
44
Left superior temporal sulcus
11.20
Left temporo-parietal junction
Right cerebellum
Right middle frontal sulcus
5.84
5.46
7.15
9370
3103
906
1965
Regions are presented that were significantly activated in the whole-brain random effects group analysis [t(15) > 3.9, pag < .05, corrected] by con-
trasting each mismatch condition against the correct condition. Displayed are the contrasts, the center coordinates in stereotaxic space (Talairach &
Tournoux, 1988), a description of the region, the t value of the maximally activated voxel, and the number of significant voxels.
Willems, O¨ zyu¨rek, and Hagoort
1245
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
7
e
-
1
p
2
d
3
f
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
o
3
c
5
n
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
0
8
0
5
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
.
/
t
.
o
n
1
8
M
a
y
2
0
2
1
be taken as evidence in favor of or against a gradient of
linguistic processes in the left inferior frontal cortex.
A potential worry is that the activation of the left in-
ferior frontal cortex could be a by-product of using the
mismatch paradigm, bearing little relevance to general
language processing. As is clear from Figure 4B, how-
ever, the correct condition also resulted in an activation
increase in this region. The activation levels of the cor-
rect condition were found to be significantly different
from zero (see Figure 4). We show that the inferior
frontal cortex activation is involved also in the process-
ing of semantically correct sentences and that its ac-
tivation in this study cannot be attributed to the use of
the mismatch paradigm (see also Hasson, Nusbaum, &
Small, 2007; Willems et al., 2007; Hagoort, 2005b; Rodd
et al., 2005).
In relation to the object priming literature cited in
the Introduction, we want to point out that our results
cannot be explained in terms of increased conceptual
priming in the correct condition. That is, word and pic-
ture were conceptually the same in both the correct
condition and the double mismatch condition. How-
increased inferior frontal cortex activation was,
ever,
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
7
e
-
1
p
2
d
3
f
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
o
3
c
5
n
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
0
8
0
5
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
f
.
.
/
Figure 3. Results from the fMRI whole-brain random effects group
analysis [t(15) > 3.9, pag < .05, corrected]. Areas significantly activated
in the (A) language mismatch versus correct condition contrast (red),
(B) picture mismatch versus correct condition contrast (blue), (C)
double mismatch versus correct condition contrast (green). Results
are overlain on a cortical sheet segmented along the gray–white matter
border in stereotaxic (Talairach) space.
mismatch condition and double mismatch condition).
More in general, it should be pointed out that what is
most striking from meta-analyses (such as Vigneau et al.,
2006; Bookheimer, 2002) is the large spread around the
mean center coordinates. Because of this variance across
studies, one single study such as the present one cannot
Figure 4. (A) Part of the left inferior frontal cortex commonly
activated by all mismatch conditions. This area was the result of
a conjunction analysis (Nichols et al., 2005) of each mismatch
condition to the correct condition (P(cid:1)L+ vs. P+L+ \ P+L(cid:1) vs.
P+L+ \ P(cid:1)L(cid:1) vs. P+L+). (B) Parameter estimates (compared to
implicit baseline) for all conditions from the area in A. Although
this area is more strongly activated in all mismatch conditions
than in the correct condition, the parameter estimate to the
correct condition was significantly different from zero [t(15) = 2.43,
p < .03], indicating that the area is also activated in the correct
condition. Error bars indicate standard error of the mean (SEM).
o
n
1
8
M
a
y
2
0
2
1
1246
Journal of Cognitive Neuroscience
Volume 20, Number 7
nevertheless, observed in the double mismatch condi-
tion. The reason that no priming effects are observed is
probably because picture and word are not presented
after another as is usually done in priming paradigms.
That is, a picture or a word does not form the context
for the other item; rather, the preceding sentence is the
crucial context.
Finally, apart from overlapping areas across condi-
tions, we also found an increase in activation in the left
superior temporal sulcus specific to the language and
double mismatch conditions, but not to the picture mis-
match condition.1 A similar result was obtained in an
earlier study investigating the processing of co-speech
gestures and words (Willems et al., 2007). This suggests
that superior temporal regions might be specifically
involved in verbal semantics.2 Interestingly, no specific
effect was observed for the picture mismatch condition.
Given the role of the ventral temporal cortex in object
representations, this area might have been expected to
be more activated in the picture mismatch condition as
compared to the other conditions. Such effect was not
observed however.
GENERAL DISCUSSION
In this study, we compared the integration of semantic
information conveyed through spoken language (words)
and visual information (pictures) at the sentence level.
Overall, our results provide strong evidence for both pro-
cesses to tax the same neural processes. That is, neural
indicators of semantic integration react the same to both
a higher integration load when information is conveyed
through a word than when it is conveyed through a pic-
ture. A same neural time course is indicated by same on-
set latencies and effect sizes of the N400 effects. The
processing at this level of comprehension does not give
temporal precedence to linguistic information over extra-
linguistic information as indicated by the N400 effects. In
terms of neural locus, part of the left inferior frontal cor-
tex was commonly activated by all mismatch conditions.
A recent neurobiological account of sentence compre-
hension has interpreted increased activation in the left
inferior frontal cortex as being the neural indicator of in-
creased integration load of a word’s meaning into a built-
up (sentence) context (Hagoort, 2005b). Here we provide
evidence for this region not to be domain-specific because
the integration of information presented in a nonlinguistic
modality also taxes this region. This is in line with an ear-
lier study in which we found the left inferior frontal cortex
to be activated more strongly to both spoken words and
co-speech gestures in a sentence context (Willems et al.,
2007). However, there is an important difference between
our previous and the present study. Co-speech gestures
are necessarily bound to a language context; that is, they
do not clearly represent their meaning when presented
on their own (McNeill, 1992; Krauss et al., 1991). Pictures,
on the contrary, are fully meaningful outside of a language
context. In this way, the present study provides more
convincing evidence for the claim that the role of the left
inferior frontal cortex in language comprehension is not
domain-specific. The left inferior frontal cortex plays an
important role in integration and selection operations that
combine linguistic and extralinguistic visual information
into a coherent overall interpretation of an expression.
The current study adds to an understanding of the
language comprehension system as taking several types
of information into account in the same way when
understanding a message (see also Spivey Knowlton &
Sedivy, 1995; Tanenhaus & Trueswell, 1995; Trueswell &
Tanenhaus, 1994; Taraban & McClelland, 1990). That is,
the system does not restrict itself to one source of in-
formation (speech), but seems to use a rich variety of
sources of meaningful information in a qualitatively simi-
lar way when understanding a message. Note that the
visual information in our study was rather simple, con-
sisting of pictures of single objects. In contrast, in the
eye movement literature cited above, visual context often
involves several objects (e.g., Tanenhaus et al., 1995).
For reasons of comparability with earlier studies (e.g.,
O¨ zyu¨rek et al., 2007) as well as for reasons of experimen-
tal control, we restricted ourselves to using pictures of
single objects as stimuli. However, the few ERP studies
that did use a richer visual context seem to suggest that
similar findings would be obtained if the visual stream of
information had been richer (e.g., Ganis & Kutas, 2003;
Sitnikova et al., 2003; West & Holcomb, 2002). This is,
however, an issue that is open for empirical investigation.
Importantly, our results support a theory of language
processing that goes against the classical two-step model
of interpretation (e.g., Lattner & Friederici, 2003; Cutler
& Clifton, 1999). Instead, in line with the immediacy
assumption, all available information is used directly to
co-determine the interpretation of linguistic expres-
sions. Moreover, we show that the role of the inferior
frontal cortex in the language comprehension network
is not restricted to linguistic information. Rather, also an
increased semantic integration load conveyed by a pic-
ture activates this area.
Acknowledgments
This research was supported by a grant from the Netherlands
Organization for Scientific Research (NWO), 051.02.040. Petra
van Alphen is acknowledged for expertly voicing the sentences.
We thank Heidi Koppenhagen and Niels Schiller for providing
the naming consistency information of the line drawings and
Tineke Snijders, Giosue` Baggio and Tessa van Leeuwen, as well
as two anonymous reviewers for helpful comments. We thank
Paul Gaalman for assistance during the scanning sessions and
Miriam Kos for help in EEG data collection.
Reprint requests should be sent to Roel M. Willems, F.C. Donders
Centre for Cognitive Neuroimaging, Radboud University Nijme-
gen, P.O. Box 9101, 6500 HB Nijmegen, The Netherlands, or via
e-mail: roel.willems@fcdonders.ru.nl.
Willems, O¨ zyu¨rek, and Hagoort
1247
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
7
e
-
1
p
2
d
3
f
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
o
3
c
5
n
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
0
8
0
5
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
.
t
.
/
o
n
1
8
M
a
y
2
0
2
1
Notes
Informal visual inspection of the contrast map at a lower sta-
1.
tistical threshold confirmed that in the picture mismatch–correct
condition comparison, the superior temporal cortex was not
activated.
2. This could relate to the debated issue of the semantic system
to be organized in multiple semantic codes or in one common
code (Caramazza, Hillis, Rapp, & Romani, 1990; McCarthy &
Warrington, 1988; Shallice, 1988). Our study was designed to in-
vestigate sentence-level integration of semantic information and
not targeted at revealing the neural representations of words
and pictures. Therefore, we are reluctant to interpret our results
in terms of multiple or modality-specific codes.
REFERENCES
Barrett, S. E., & Rugg, M. D. (1990). Event-related potentials
and the semantic matching of pictures. Brain and
Cognition, 14, 201–212.
Baumgaertner, A., Weiller, C., & Buchel, C. (2002). Event-related
fMRI reveals cortical sites involved in contextual sentence
integration. Neuroimage, 16, 736–745.
Bookheimer, S. (2002). Functional MRI of language: New
approaches to understanding the cortical organization of
semantic processing. Annual Review of Neuroscience, 25,
151–188.
Brown, C. M., Hagoort, P., & Kutas, M. (2000). Postlexical
integration processes in language comprehension: Evidence
from brain-imaging research. In M. S. Gazzaniga (Ed.),
The cognitive neurosciences (2nd ed., pp. 881–895).
Cambridge: MIT Press.
Caramazza, A., Hillis, A. E., Rapp, B. C., & Romani, C. (1990).
The multiple semantics hypothesis: Multiple confusions?
Cognitive Neuropsychology, 7, 161–189.
Clark, H. H. (1996). Using language. New York: Cambridge
University Press.
Culicover, P. W., & Jackendoff, R. (2006). The simpler syntax
hypothesis. Trends in Cognitive Sciences, 10, 413–418.
Cutler, A., & Clifton, C. E. (1999). Comprehending spoken
language: A blueprint of the listener. In C. M. Brown
& P. Hagoort (Eds.), The neurocognition of language.
Oxford: Oxford University Press.
Davis, M. H., Coleman, M. R., Absalom, A. R., Rodd, J. M.,
Johnsrude, I. S., Matta, B. F., et al. (2007). Dissociating
speech perception and comprehension at reduced levels
of awareness. Proceedings of the National Academy of
Sciences, U.S.A., 104, 16032–16037.
Eickhoff, S. B., Stephan, K. E., Mohlberg, H., Grefkes, C.,
Fink, G. R., Amunts, K., et al. (2005). A new SPM toolbox
for combining probabilistic cytoarchitectonic maps and
functional imaging data. Neuroimage, 25, 1325–1335.
Federmeier, K. D., & Kutas, M. (2001). Meaning and modality:
Influences of context, semantic memory organization, and
perceptual predictability on picture processing. Journal
of Experimental Psychology: Learning, Memory, and
Cognition, 27, 202–224.
Federmeier, K. D., & Kutas, M. (2002). Picture the difference:
Electrophysiological investigations of picture processing in the
two cerebral hemispheres. Neuropsychologia, 40, 730–747.
Fodor, J. A. (1983). The modularity of mind. Cambridge:
MIT Press.
Forman, S. D., Cohen, J. D., Fitzgerald, M., Eddy, W. F.,
Mintun, M. A., & Noll, D. C. (1995). Improved assessment
of significant activation in functional magnetic resonance
imaging (fMRI): Use of a cluster-size threshold. Magnetic
Resonance in Medicine, 33, 636–647.
Forster, K. I. (1979). Levels of processing and the structure
of the language processor. In W. E. Cooper & C. T. Walker
(Eds.), Sentence processing: Psycholinguistic essays
presented to Merrill Garrett (pp. 27–85). Hillsdale,
NJ: Erlbaum.
Friederici, A. D., Ruschemeyer, S. A., Hahne, A., & Fiebach,
C. J. (2003). The role of left inferior frontal and superior
temporal cortex in sentence comprehension: Localizing
syntactic and semantic processes. Cerebral Cortex, 13,
170–177.
Ganis, G., & Kutas, M. (2003). An electrophysiological study
of scene effects on object identification. Cognitive Brain
Research, 16, 123–144.
Ganis, G., Kutas, M., & Sereno, M. I. (1996). The search for
‘‘common sense’’: An electrophysiological study of the
comprehension of words and pictures in reading. Journal
of Cognitive Neuroscience, 8, 89–106.
Hagoort, P. (2005a). Broca’s complex as the unification
space for language. In A. Cutler (Ed.), Twenty first century
psycholinguistics: Four cornerstones (pp. 157–172).
Mahwah, NJ: Erlbaum.
Hagoort, P. (2005b). On Broca, brain, and binding: A new
framework. Trends in Cognitive Sciences, 9, 416–423.
Hagoort, P., & Brown, C. (1994). Brain responses to lexical
ambiguity resolution and parsing. In L. Frazier, J. Clifton
Charles, & K. Rayner (Eds.), Perspectives in sentence
processing (pp. 45–80). Hillsdale, NJ: Erlbaum.
Hagoort, P., Hald, L., Bastiaansen, M., & Petersson, K. M.
(2004). Integration of word meaning and world knowledge
in language comprehension. Science, 304, 438–441.
Hagoort, P., & van Berkum, J. J. (2007). Beyond the sentence
given. Philosophical Transactions of the Royal Society,
Series B, 362, 801–811.
Hasson, U., Nusbaum, H. C., & Small, S. L. (2007). Brain
networks subserving the extraction of sentence information
and its encoding to memory. Cerebral Cortex, 17,
2899–2913.
Holcomb, P. J., & McPherson, W. B. (1994). Event-related
brain potentials reflect semantic priming in an object
decision task. Brain and Cognition, 24, 259–276.
Huynh, H., & Feldt, L. (1976). Estimation of the box correction
for degrees of freedom from sample data in randomized
block and splitplot designs. Journal of Educational
Statistics, 1, 69–82.
Koelsch, S., Kasper, E., Sammler, D., Schulze, K., Gunter, T.,
& Friederici, A. D. (2004). Music, language and meaning:
Brain signatures of semantic processing. Nature
Neuroscience, 7, 302–307.
Krauss, R. M., Morrel Samuels, P., & Colasante, C. (1991).
Do conversational hand gestures communicate? Journal
of Personality and Social Psychology, 61, 743–754.
Kuperberg, G. R., Holcomb, P. J., Sitnikova, T., Greve, D.,
Dale, A. M., & Caplan, D. (2003). Distinct patterns of
neural modulation during the processing of conceptual
and syntactic anomalies. Journal of Cognitive Neuroscience,
15, 272–293.
Kuperberg, G. R., McGuire, P. K., Bullmore, E. T., Brammer,
M. J., Rabe-Hesketh, S., Wright, I. C., et al. (2000).
Common and distinct neural substrates for pragmatic,
semantic, and syntactic processing of spoken sentences:
An fMRI study. Journal of Cognitive Neuroscience, 12,
321–341.
Kutas, M., & Hillyard, S. A. (1980). Reading senseless
sentences: Brain potentials reflect semantic incongruity.
Science, 207, 203–205.
Kutas, M., & Hillyard, S. A. (1984). Brain potentials during
reading reflect word expectancy and semantic association.
Nature, 307, 161–163.
1248
Journal of Cognitive Neuroscience
Volume 20, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
7
e
-
1
p
2
d
3
f
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
o
3
c
5
n
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
0
8
0
5
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
.
f
/
t
o
n
1
8
M
a
y
2
0
2
1
Kutas, M., & Van Petten, C. K. (1994). Psycholinguistics
electrified: Event-related brain potential investigations.
In M. A. Gernsbacher (Ed.), Handbook of psycholinguistics
(pp. 83–143). San Diego, CA: Academic Press.
Lattner, S., & Friederici, A. D. (2003). Talker’s voice
and gender stereotype in human auditory sentence
processing-evidence from event-related brain potentials.
Neuroscience Letters, 339, 191–194.
Lebreton, K., Desgranges, B., Landeau, B., Baron, J. C., &
Eustache, F. (2001). Visual priming within and across
symbolic format using a tachistoscopic picture identification
task: A PET study. Journal of Cognitive Neuroscience,
13, 670–686.
Luck, S. J. (2005). An introduction to the event-related
potential technique. Cambridge: MIT Press.
Martin, A., & Chao, L. L. (2001). Semantic memory and
the brain: Structure and processes. Current Opinion
in Neurobiology, 11, 194–201.
McCarthy, R. A., & Warrington, E. K. (1988). Evidence for
modality-specific meaning systems in the brain. Nature,
334, 428–430.
McNeill, D. (1992). Hand and mind: What gestures reveal
about thought. Chicago, IL: University of Chicago Press.
McPherson, W., & Holcomb, P. J. (1999). An
electrophysiological investigation of semantic priming with
pictures of real objects. Psychophysiology, 36, 53–65.
Miller, J., Patterson, T., & Ulrich, R. (1998). Jackknife-based
method for measuring LRP onset latency differences.
Psychophysiology, 35, 99–115.
Ni, W., Constable, R. T., Mencl, W. E., Pugh, K. R., Fulbright,
R. K., Shaywitz, S. E., et al. (2000). An event-related
neuroimaging study distinguishing form and content in
sentence processing. Journal of Cognitive Neuroscience,
12, 120–133.
Nichols, T., Brett, M., Andersson, J., Wager, T., & Poline, J. B.
(2005). Valid conjunction inference with the minimum
statistic. Neuroimage, 25, 653–660.
Nigam, A., Hoffman, J. E., & Simons, R. F. (1992). N400 to
semantically anomalous pictures and words. Journal of
Cognitive Neuroscience, 4, 15–22.
Oldfield, R. C. (1971). The assessment and analysis of
handedness: The Edinburgh inventory. Neuropsychologia,
9, 97–113.
O¨ zyu¨rek, A., Willems, R. M., Kita, S., & Hagoort, P. (2007).
On-line integration of semantic information from speech
and gesture: Insights from event-related brain potentials.
Journal of Cognitive Neuroscience, 19, 605–616.
Picton, T. W., Bentin, S., Berg, P., Donchin, E., Hillyard, S. A.,
Johnson, R., Jr., et al. (2000). Guidelines for using human
event-related potentials to study cognition: Recording
standards and publication criteria. Psychophysiology, 37,
127–152.
Rodd, J. M., Davis, M. H., & Johnsrude, I. S. (2005). The
neural mechanisms of speech comprehension: fMRI
studies of semantic ambiguity. Cerebral Cortex, 15,
1261–1269.
Schacter, D. L., & Buckner, R. L. (1998). Priming and the
brain. Neuron, 20, 185–195.
Shallice, T. (1988). Specialisation within the semantic
system. Cognitive Neuropsychology, 5, 133–142.
Sitnikova, T., Kuperberg, G., & Holcomb, P. J. (2003).
Semantic integration in videos of real-world events: An
electrophysiological investigation. Psychophysiology,
40, 160–164.
Spivey Knowlton, M. J., & Sedivy, J. C. (1995). Resolving
attachment ambiguities with multiple constraints.
Cognition, 55, 227–267.
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic
atlas of the human brain. New York: Thieme.
Tanenhaus, M. K., Spivey Knowlton, M. J., Eberhard, K. M.,
& Sedivy, J. C. (1995). Integration of visual and linguistic
information in spoken language comprehension. Science,
268, 1632–1634.
Tanenhaus, M. K., & Trueswell, J. C. (1995). Sentence
comprehension. In J. L. Miller & P. D. Eimas (Eds.),
Speech, language, and communication (pp. 217–262).
San Diego, CA: Academic Press.
Taraban, R., & McClelland, J. L. (1990). Parsing and
comprehension: A multiple-constraint view. In D. A. Balota,
G. B. Flores d’Arcais, & K. Rayner (Eds.), Comprehension
processes in reading (pp. 231–263). Hillsdale, NJ: Erlbaum.
Thompson-Schill, S. L., D’Esposito, M., Aguirre, G. K., & Farah,
M. J. (1997). Role of left inferior prefrontal cortex in retrieval
of semantic knowledge: A reevaluation. Proceedings of the
National Academy of Sciences, U.S.A., 94, 14792–14797.
Trueswell, J. C., & Tanenhaus, M. K. (1994). Toward a
lexicalist framework of constraint-based syntactic ambiguity
resolution. In C. Clifton, Jr. (Ed.), Perspectives on sentence
processing (pp. 155–179). Hillsdale, NJ: Erlbaum.
Van Berkum, J. J. A., Brown, C. M., Zwitserlood, P., Kooijman,
V., & Hagoort, P. (2005). Anticipating upcoming words
in discourse: Evidence from ERPs and reading times.
Journal of Experimental Psychology: Learning, Memory,
and Cognition, 31, 443–467.
Van Petten, C., & Rheinfelder, H. (1995). Conceptual
relationships between spoken words and environmental
sounds: Event-related brain potential measures.
Neuropsychologia, 33, 485–508.
Vigneau, M., Beaucousin, V., Herve, P. Y., Duffau, H.,
Crivello, F., Houde, O., et al. (2006). Meta-analyzing left
hemisphere language areas: Phonology, semantics, and
sentence processing. Neuroimage, 30, 1414–1432.
Wagner, A. D., Desmond, J. E., Demb, J. B., Glover, G. H.,
& Gabrieli, J. D. (1997). Semantic repetition priming for
verbal and pictorial knowledge: A functional MRI study
of left inferior prefrontal cortex. Journal of Cognitive
Neuroscience, 9, 714–726.
Wagner, A. D., Pare-Blagoev, E. J., Clark, J., & Poldrack, R. A.
(2001). Recovering meaning: Left prefrontal cortex guides
controlled semantic retrieval. Neuron, 31, 329–338.
West, W. C., & Holcomb, P. J. (2002). Event-related potentials
during discourse-level semantic integration of complex
pictures. Cognitive Brain Research, 13, 363–375.
Willems, R. M., O¨ zyu¨rek, A., & Hagoort, P. (2007). When
language meets action: The neural integration of gesture
and speech. Cerebral Cortex, 17, 2322–2333.
Willems, O¨ zyu¨rek, and Hagoort
1249
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
7
e
-
1
p
2
d
3
f
5
/
1
2
9
0
3
/
6
7
9
/
2
1
5
2
o
3
c
5
n
/
1
2
0
7
0
5
8
9
5
2
9
0
2
0
8
/
5
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
0
8
0
5
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
.
f
.
.
/
o
n
1
8
M
a
y
2
0
2
1